
Field cage evakluations of Beauveria bassiana and ... Chatriand, Emerald BioAgricultural...
92
February, 2006 Field cage evaluations of Beauveria bassiana and Metarhizium anisopliae in spray and bait formulations for potential use against Mormon cricket in Idaho R. Nelson Foster 1 , Stefan Jaronski 2 , K. Chris Reuter 1 and Lonnie R. Black 1 Organizations 1 USDA-APHIS-PPQ-CPHST-Decision Support and Pest Management Systems Laboratory, Phoenix, Arizona. 2 USDA-ARS-NPARL, Pest Management Unit, Sidney, MT Cooperators Lori Atkins and Donna Hooper, landowner, near Mayfield, Idaho Dave McNeal and Rob McChesney, USDA-APHIS-PPQ, Boise, Idaho David I. B. Vander Hooven, Advanced Organics Inc., Maumee, Ohio Gary Chatriand, Emerald BioAgricultural Corporation, Lancing, Michigan. Tom Correll, Earth BioSciences, New haven, Connecticut Abstract No mortality of immature Mormon crickets was detected in the field for 14 days after treatment with fungal baits or sprays of either Beauveria bassiana Strain GHA or Metarhizium anisopliae Strain F52. Feeding behavior may have adversely affected the success of the bait treatments but confirmation will require additional study. Most importantly, temperatures recorded from the surrogates on the ground in cages indicate that an insufficient number of accumulated hours at temperatures appropriate for fungal development occurred during the study. The data suggest that (32 – 43) and (58 – 75) days of observation would have been required to detect any mortality that could have been attributed to the M. anisopliae and B. bassiana treatments respectively. In field mortality of adult Mormon crickets was not followed, but treated individuals were sent to Sidney, MT for observation. Prevalence of B. bassiana and M. anisopliae infections in these adults, as evidenced by mortality corrected for untreated controls, was 70% and 100% respectively. It is obvious that significantly longer observation times in the field will be required to detect any mortality in Mormon crickets that could be attributable to these fungal treatments. In fact, several weeks may be required after treatment for fungal infections to be expressed as mortality. However, there is some potential for faster fungal growth in Mormon cricket, particularly in the 1
Transcript of Field cage evakluations of Beauveria bassiana and ... Chatriand, Emerald BioAgricultural...

February, 2006
Field cage evaluations of Beauveria bassiana and Metarhizium anisopliae in spray and bait formulations for potential use against Mormon cricket in
Idaho
R. Nelson Foster1, Stefan Jaronski2, K. Chris Reuter1 and Lonnie R. Black1
Organizations
1USDA-APHIS-PPQ-CPHST-Decision Support and Pest Management Systems Laboratory, Phoenix, Arizona. 2USDA-ARS-NPARL, Pest Management Unit, Sidney, MT
Cooperators Lori Atkins and Donna Hooper, landowner, near Mayfield, Idaho Dave McNeal and Rob McChesney, USDA-APHIS-PPQ, Boise, Idaho David I. B. Vander Hooven, Advanced Organics Inc., Maumee, Ohio Gary Chatriand, Emerald BioAgricultural Corporation, Lancing, Michigan. Tom Correll, Earth BioSciences, New haven, Connecticut
Abstract
No mortality of immature Mormon crickets was detected in the field for 14 days after treatment with fungal baits or sprays of either Beauveria bassiana Strain GHA or Metarhizium anisopliae Strain F52. Feeding behavior may have adversely affected the success of the bait treatments but confirmation will require additional study. Most importantly, temperatures recorded from the surrogates on the ground in cages indicate that an insufficient number of accumulated hours at temperatures appropriate for fungal development occurred during the study. The data suggest that (32 – 43) and (58 – 75) days of observation would have been required to detect any mortality that could have been attributed to the M. anisopliae and B. bassiana treatments respectively. In field mortality of adult Mormon crickets was not followed, but treated individuals were sent to Sidney, MT for observation. Prevalence of B. bassiana and M. anisopliae infections in these adults, as evidenced by mortality corrected for untreated controls, was 70% and 100% respectively. It is obvious that significantly longer observation times in the field will be required to detect any mortality in Mormon crickets that could be attributable to these fungal treatments. In fact, several weeks may be required after treatment for fungal infections to be expressed as mortality. However, there is some potential for faster fungal growth in Mormon cricket, particularly in the
1

immature stages. Their aggregating behavior under inclement and nocturnal conditions may conserve and actually promote higher temperatures. The potential value of this aspect of their behavior will require additional study.
Introduction
There are no generally accepted entomopathogen treatments available for use against grasshoppers and Mormon crickets in the United States. Sensitive situations commonly encountered within the western states in potential program areas preclude traditional chemical insecticide treatments. The number of sensitive situations is increasing and in areas of grasshopper or Mormon cricket infestations this presence at best complicates and more often prevents much needed local or area-wide treatments of grasshoppers and/or Mormon crickets on rangeland. In the early 1990’s, extensive work on the development of the fungus Beauveria bassiana GHA by Mycotech Corporation and USDA-APHIS “Methods” (Foster et al. 1991, 1992, 1993, 1996-1999) led to its registration in the US, even though final development for wide acceptance and use was never achieved. While the “stressor concept ” (Foster et al. 1996a; Reuter et al. 1996), where sub lethal doses of insecticides were added to the fungus in an attempt to alter normal behavior of a grasshopper to increase fungal susceptibility, indicated potential promise, the USDA-CPHST efforts in its development were redirected from 1995 until 2002. Interest in B. bassiana has recently been renewed with work conducted by the University of Wyoming which indicated that canola oil as an attractant and diluent may improve results. Collaborative work conducted by the University of Wyoming (Lockwood, 2003) and the USDA- ARS (Jaronski et al. 2002) has indicated that an effective formulation may result by simply applying the spores in canola oil rather than paraffinic oil. While non-replicated, that work indicated substantial differences between B. bassiana treatments applied in canola oil compared to paraffinic oil and untreated control populations. In 2003, a joint APHIS and ARS replicated study (Foster et al. 2003b) conducted to verify the improvement with canola oil compared to a paraffinic oil diluent, showed that no significant increase in grasshopper mortality compared to untreated populations could be attributed to either of the oils studied. Subsequent unpublished laboratory studies conducted by Jaronski, have revealed grasshoppers exhibited greater infection levels and consumption amounts when exposed to paper disks treated with raw canola oil and B. bassiana compared to the refined canola oil and B. bassiana mixture that had been used in both the 2003 and 2004 studies. However, a subsequent joint APHIS and ARS study to evaluate unrefined and refined canola oil mixtures with B. bassiana in field studies revealed no statistical difference in population reductions among the B. bassiana treatments or between those treatments and untreated control populations or populations treated with unrefined canola oil without B. bassiana. (Foster et al. 2004b). The most popular current belief concerning ineffective results of B. bassiana against grasshoppers, in unconfined field populations, supposes that by positioning themselves to increase sun exposure, grasshoppers thermo-regulate and increase body temperatures (Inglis et al. 1996; Inglis et al. 1999). This higher temperature places the insect in a temperature environment much of the day that is non-conducive to fungal growth within the insect. As a
2

result, the mycosis takes so long to grow to a level to cause mortality that detection of population reductions attributable to the fungus, compared to natural population declines is difficult at best. Therefore, lower environmental temperatures may promote more easily detectable mortality that could be attributed to treatment induced fungal infection. Because damaging populations of grasshoppers are generally present in hotter conditions for longer periods during the day than Mormon crickets, and because Mormon crickets did not demonstrate significant “behavioral fever” (thermo-regulation) after infection with B. bassiana (Turnbow 1998), optimal temperature parameters for fungal development within an insect may be more easily encountered with Mormon crickets, particularly with immatures early in the year. Another entomopathogen, Metarhizium anisopliae F52, has recently been registered for control of Coleoptera in horticulture and managed turf, and soft bodied ticks by Earth Biosciences Corp. (Danbury, CT). In laboratory bioassays with immature and adult Mormon crickets, Jaronski (unpublished data) indicated that the F52 strain was highly infectious and virulent, more so than B. bassiana GHA. A novel bait formulation (Taste-E-Bait, a combination of bakery, snack, cereal and confectionary waste) has shown good enhancement of carbaryl efficacy against Mormon cricket (Foster et al. 2003a; Foster et al. 2004a). We hypothesize that such bait could enhance dose transfer of fungal conidia to target Mormon crickets and thus increase efficacy over a ULV spray with a fungus. If so, this result would greatly decrease the cost of fungal pathogens in use against this insect pest. Because non-chemical options are highly desirable and significant work remains to develop fungal pathogens in the U.S. against Orthopteran insects, the following study was conducted to evaluate bait and spray formulations of the fungi B. bassiana and for the first time Metarhizium anisopliae in field cages (redesigned to minimize cage effects) against Mormon crickets and under more acceptable temperatures for fungal development.
Objectives 1. Evaluate B. bassiana GHA and M. anisopliae F52 for potential use against Mormon cricket 2. Compare spray and bait formulations of B. bassiana GHA and Metarhizium
anisopliae F52 against immature and adult Mormon cricket. 3. Determine the accumulated hours of optimal and sub-optimal temperature parameters
necessary for fungal growth to produce mortality under field conditions, using thermal surrogates.
Materials and Methods
The study area was located ca. 18.5 miles south east of the Boise, Idaho airport, near the community of Mayfield on the Lori Atkins property. This location was selected because of the
3

history of Mormon crickets on the property, proximity to current known populations of Mormon crickets, and abundance of historically infested rangeland without livestock. The GHA strain of Beauveria bassiana (Emerald BioAgricultural Corporation, Lansing, Michigan) and the F52 strain of Metarhizium anisopliae (Earth BioSciences, New Haven, Connecticut) were (1) incorporated into untreated Tast - E –Bait (a combination of bakery, snack, cereal, and confectionary food waste produced by Advanced Organics Inc. Maumee, Ohio) graduals and (2) formulated as a liquid spray, initially in Sun Spray Ultra Fine oil. Bait and initial liquid formulations were made at the USDA, ARS, NPARL in Sidney, Montana. Conidia of B. bassiana Strain GHA, as technical grade active ingredient Lot 0304-1, were supplied by Emerald BioAgriculture (now Laverlam International), Butte MT. Conidia of M. anisopliae Strain F52 (Earth Biosciences) were produced by USDA ARS using a biphasic, liquid-solid substrate fermentation process (Bradley et al. 2002). Prior to formulation, conidial viabilities were determined by plating dilute aqueous suspensions of each technical powder onto potato dextrose agar, incubating at 27-28° C. for 16-19 hr, and then examining the conidia with 400X phase contrast microscopy. The Metarhizium conidia required a preliminary step, in which a small quantity of conidia was exposed to 100% relative humidity for 1-2 hr before suspension and plating. A minimum of 400 conidia were examined for germination; a conidium was considered viable (germinated) if it had produced a visible germination peg during the specified incubation time. Viability of the B. bassiana technical powder was 95%, that of M. anisopliae, 85%. The bait formulation consisted of Tast-E-Bait (Advanced Organics Inc., Maumee, Ohio) coated with fungal conidia using a binder of methylated soybean oil. A 1 kg quantity of Tast-E-Bait was treated with a fine spray of Golden Pest Natr’l Oil® (Stoller Manufacturing Inc.) at the rate of 10% v/w, and then mixed in a rotating v-cone blender for 15 minutes. The resulting slightly sticky granules were spread out into a shallow layer and dusted with fungal conidia at a rate equivalent to 1 x 1013 viable conidia / 4 kg (10 lb.) carrier (1.14x1012 conidia/lb formulation, 2.8x106 conidia/bait particle), then mixed in the v-cone blender for 15 minutes. Liquid formulations were prepared by adding sufficient spores of B. bassiana GHA or M. anisopliae F52 to achieve 1x1013 viable conidia /liter of Sun Spray Ultra Fine Oil® (Sun Oil Co.). This concentrate was subsequently diluted in the field 1:4 with canola oil for spraying. Immature Mormon cricket studies. Cages 8.5 in high X 7 in diameter, fashioned from 1/8 inch hardware cloth (Fig. 1) were established on untreated rangeland (Fig. 2) to evaluate the potential of baits and sprays containing the separate fungal pathogens B. bassiana and M. anisopliae against Mormon cricket. Ten cages were established for each of the six treatments including the untreated control and an oil only spray. Each of the treatments including the untreated control, were replicated 4 times. Therefore, the study utilized a total of 240 cages. The appropriate amount of bait was pre-weighed for each cage in the laboratory and placed in the cages prior to cricket introductions. Sprays were applied by using an airbrush (Paasche Type H with 75 regulator) modified with a customized syringe needle for liquid injection to produce droplets which simulate aerially applied sprays (Foster et al. 1996b) to the area to be enclosed by each cage (Figure 3). Drift
4

during spray application was prevented by using portable walls or a containment chamber (Figure 4). All treatments were applied at the rate of 1 X 1013 spores per acre (10 lbs of bait per acre or 0.25 gal (1 L) of oil formulation per acre diluted with canola oil to deliver 1.25 gal (5 L) of spray per acre). The bait application theoretically delivered about 25 particles per cage; the target delivery of the spray was 2.5x105 conidia/cm2 planar surface. Baits were applied on April 29, 2005. Because of the time required to treat each of the cages with sprays, two replications of the spray portion of study were treated on April 30 and May 1, 2005 respectively. Immediately following treatment, each of the cages was stocked with a single third instar stage Mormon cricket. Stocking cages with a single cricket prevented cannibalism, a common occurrence when more than one cricket is confined in a cage. Cricket mortality within cages was monitored daily for 14 days. The study was conducted from April 29 through May 15, 2003. Crickets that died during the study were removed from the cage on the day of death and placed individually for observation in a labeled petri dish. Each dish contained a cotton ball moistened with distilled water to provide very high humidity, to promote fungal outgrowth from the cadaver and sporulation, which was recorded daily. The presence of fungal outgrowth characteristic of Beauveria or Metarhizium was an indication of mycosis. Air temperatures and precipitation were recorded daily for the duration of the study. Mormon cricket thermal surrogates were used to measure the temperature that a cricket would generate on the ground and at canopy level within and outside of a cage. Surrogates consisted of 1.5 ml Eppendorff tubes filled with soy sauce and fitted with a copper-constatin thermocouple wire connected to a data logger recording device (DualogR®, Cole Palmer) (Figures 5 and 6). These thermal surrogates have been shown to reasonably represent the ability of Orthoptera to absorb solar radiant heat as well as indicate typical body temperatures when Orthoptera are unable to thermo-regulate (Lactin and Johnson 1998). The recorder was set to record temperatures in the surrogates every 5 min. for the duration of the field aspect of the study. Adult Mormon crickets studies. Cages were established and sprays were prepared and applied similarly to that described for the immature studies. Thermal surrogates were also used as previously described to measure the temperature that an adult cricket would generate on the ground and at canopy level within and outside of a cage. No baits were studied for efficacy against adults. Again, due to time restraints, spray treatment replications were divided equally over two days, June 2 and 3, 2005. Seventh instar and adult stage Mormon crickets were used in this study. Mortality within cages was monitored daily for 4 days at which time all survivors were transferred via air transport within cooled Styrofoam containers to USDA ARS in Sidney, Montana. Within the shipping container, all crickets within each treatment and replicate were combined in separately labeled ventilated (18 in long x 3.5 in diameter) plastic tube cages containing crumpled newspaper to increase the resting surface area and minimize canabalism. The field aspect of the study was conducted in Idaho from June 2 through June 7. Upon arrival in Sidney, the Mormon crickets from each treatment and replicate were split into two groups. The first group, approximately 20 insects per fungus treatment and 30 for each of the untreated and oil carrier controls, was incubated at a constant 26-28° C. (optimal for fungal mycosis, Fargues et al. 1997; T. Kabaluk unpublished data) and a 16:8 L:D photoperiod in the
5

laboratory. A second group, 10 insects per treatment, chosen at random from those shipped to Sidney, was held outdoors, on vegetated ground, where the Mormon crickets could actively bask and thermo-regulate. In each case individual insects were caged in plastic tubes (8 in. long x 2 in. diameter) with screened caps at each end. All insects were provided a diet of sunflower seed, bird seed and Tetramin® tropical fish food, and cotton-stoppered vials of water. In addition the interior of the cages were sprayed with a water mist each morning to simulate dew. Part of each cage was shaded to provide refuge during the hottest part of each day. A thermal surrogate connected to a data logger was placed in an empty cage together with the outdoor-incubated insects. These insects were incubated in their respective regimes for 14 days. Mortality was determined daily and all cadavers were removed, surface decontaminated by immersion in 0.5% NaOCl for 1 minute, then placed at 100% humidity to allow any mycoses to become expressed. Laboratory bioassays of granules. Concurrent with the field evaluations we also conducted a laboratory bioassay of the granules. Individual Mormon cricket nymphs were confined with 250 mg of treated or untreated bait for 48 hours, then transferred to clean cages and provided seed/fish food diet ad libitum. Fifteen crickets were used for each treatment, but the entire test was not replicated. Crickets were incubated for 14 days. Daily mortality was determined with any cadavers processed to evidence mycosis as described earlier. Statistical analysis. Field data were analyzed using an analysis of variance with the Tukey multiple comparison test to separate means when the ANOVA F was significant. Analyses were performed with SYSTAT for Windows® (SPSS Inc. 1977). Effects of fungal treatments on incubated Mormon Crickets were assessed via Fisher’s Exact test and also in terms of Median Survival Time (Kaplan Meier method) and Cox’s Proportional Hazards technique (Kleinbaum 1996). Latter analyses were performed using Statistix® 8 (Analytical Software Inc. 1998). Mortality in the bioassay and laboratory incubation was manually analyzed with Fisher’s Exact Test.
Results and Discussion
Immature Mormon cricket studies. No significant mortality was detected at 0 -14 days after treatment in the populations treated with bait formulatons of Beauveria bassiana or Metarhizium anisopliae when compared to the untreated population (Fig. 7). No mortality occurred in untreated populations until 3 days after treatment. From then on, mortality in the untreated population increased to 27.5 % at 14 days after treatment. Untreated mortality was slightly higher than expected at 12.5 % from days 5-8 compared to other bait studies with Mormon crickets where untreated population showed mortality of 5% at 7 days after treatment (Foster et al. 2003a, 2004a). However, at the end of field observations, 14 days after treatment, both fungal treated populations and the untreated populations showed an equivalent mean percentage mortality of 27.5 %. This lack in detectable mortality could be explained if crickets were not feeding on the bait. In subsequent small feeding preference studies in the laboratory, Mormon crickets were unexpectedly observed to feed minimally, if at all, on untreated Tast-E-Bait. Further indication of non- feeding by Mormon crickets was seen in laboratory bioassays with Tast-E-Bait where Mormon crickets at 26-28o C
6

showed no significant mortality at 14 days after fungal treatments when compared to untreated populations even when the dose of Beauveria bassiana was increased to 4 times the experimental rate of 1 x 10 13/ acre (Fig. 8). However, these results seems to be at conflict with results of excellent mortality resulting from exposure to Tast-E-Bait containing carbaryl against Mormon cricket in several other studies (Foster et al. 2003a, 2004a). If given equal value and taken together, the studies would seem to suggest that Mormon crickets may have been attracted to the carbaryl or the additive used to incorporate the toxicant into the bait rather than the bait itself. This apparent non feeding behavior alone may explain the lack of mortality. However, only additional testing will fully provide that answer. No significant mortality was detected at 0-14 days after treatment in the populations sprayed with B. bassiana, M. anisopliae, or oil only when compared to untreated populations even though mortalities from populations in M. anisopliae treated and untreated populations resulted in 27.5 % and B. bassiana and oil only treatments resulted in 42.5 and 40 % mortality respectively at 14 days after treatment (Fig. 9). The mortality in untreated populations at 5-7 days after treatment was still about twice (10%) what was expected (5%) based on earlier studies. (Foster et al. 2003a, 2004a). This higher than expected untreated mortality may be explained by sparse vegetation within the cages and therefore a lack of diverse plants needed to insure a sufficient diet of required host plants. Additionally, if this happened, less spray material than expected would have been consumed and therefore a reduced fungal exposure to the cricket would have occurred also helping to explain no detectable mortality. Absence of feeding on the bait could certainly explain the lack of mortality with baits but, probably does not explain the low mortality experienced with the sprays. Topical bioassays of both fungi (Jaronski, unpublished data) revealed LD50s of 1.46x104 and 5.63x103 spores per immature insect for B. bassiana GHA and M. anisopliae F52, respectively (based on Day 10 mortalities). A field spray of 1x1013 conidia/acre should have yielded approx 2.5x105 conidia per cm2 on a planar target. (In comparison, the adult Mormon cricket field trial yielded infection rates (mortality) of 70-100% (Table 1), adjusted for untreated control. In bioassays adults were less susceptible that nymphs to both fungi.) While the field observations appear unsupportive of successful fungal mycosis, data from the surrogates seem to explain the apparent ineffectiveness of both fungal treatments. To understand this, we must first review the optimal temperature parameters under which both fungi can reproduce to a level sufficient to result in mortality. Optimal growth of both fungi occur at 18-30o C (64.4-86.0 o) with some development but less than 50% at intervals of 30-32oC (86.0-89.6
F) and 9-18oC (48.2-64.4o F). No fungal growth occurs above 32o C (89.6o F) or below 9 oC (48.2 o F) (Table 2) (Fargus et al. 1997; Jaronski, unpublished). Based upon USDA ARS bioassays with nymphal and adult Mormon crickets (Jaronski, unpublished), B. bassiana GHA will produce mortality in 10-14 days (240-336 hrs) and Metarhizium anisopliae F52 will grow to produce mortality in grasshoppers in 6–8 days (144-192 hrs) at the LC50 and a constant temperature of 28° C. (83.4° F.). The temperatures measured in the surrogates inside and outside cages on the ground and at the plant canopy demonstrate the possible hours of the day where fungal growth could occur (Figures 10 and 11). Further breakdown of each situation illustrates the actual number of hours
7

each day that could produce optimal fungal growth. (Figures 12-15). When the numbers of optimal fungal growth hours are accumulated for the duration of the study (Table 3) it becomes clear that an insufficient number of hours occurred during the 15 days of the study to produce mortality. Metarhizium anisopliae treatments at the temperatures we experienced during the 15 days of the study, would have required (34.5 – 45 days) and (21 – 28.5 days) for mortality to occur outside of the cages on the ground and at vegetation canopy level respectively. Similarly, it would have required (32-43 days) and (12-15 days) for mortality to occur inside cages on the ground and at vegetation canopy level, respectively. These calculations would appear to indicate that our study should have detected mortality within cages. The quickest estimated mortality should have occurred inside cages at canopy level at 12 – 15 days after treatment. However, it was observed that Mormon crickets during this study spent most of the time at ground level and very little at canopy height. Beauveria bassiana treatments at the temperatures we experienced during the 15 days of the study, would have required (57 – 80 days) and (36 – 50 days) for mortality to occur outside of the cages on the ground and at vegetation canopy level respectively. Similarily, it would have required (57.7 – 75.2 days) and (19.1 – 26.8 days) for mortality to occur inside cages on the ground and at vegetation canopy level, respectively. It is obvious that significantly longer observation times in the field will be required to detect any mortality in Mormon crickets that could be attributable to these fungal treatments. Adult Mormon cricket studies. In an attempt to alleviate the problems associated with detection of mortality during a short period of time in the field that was experienced with immature Mormon crickets, adults were exposed to treatments in the field within cages for 4 days and subsequently sent to Sidney, MT, for field establishment in cages for potentially unlimited observation times. Additionally, some crickets were maintained in the laboratory under constant optimal temperatures for fungal development to determine the actual rate of infection. The indoor incubations showed very high infection rates from the field exposure in Idaho to both fungi (Table 1). Unfortunately many of the crickets that were incubated outdoors died early during the study because of unexpected high temperatures. This situation made these data useless. Nevertheless, we observed that the simulated ULV spray of 1x1013 conidia/acre resulted in infection rates that could have successfully reduced the target populations if body temperatures had allowed. Use of Cox’s Proportional Hazards technique (Table 4) as well as Kaplan Meier Survivorship curves (Figure 16) with the daily mortality data revealed that F52 was slightly more virulent than GHA (Z= 2.12, p =.0344, Relative Risk of death 2.11. There was no difference between mortalities resulting from oil carrier treatments and untreated populations.
Conclusions
No mortality of immature Mormon crickets was detected in the field for 14 days after treatment with fungal baits or sprays. Feeding behavior may have affected the success of the bait
8

treatments but confirmation will require additional study. Most importantly, temperatures recorded from the surrogates on the ground in cages indicate that an insufficient number of accumulated hours at temperatures appropriate for fungal development occurred during the study. The data suggest that (32 – 43) and (58 – 75) days of observation would have been required to detect any mortality that could have been attributed to the Metarhizium anisopliae and Beauveria bassiana treatments respectively. Outdoor studies of adult Mormon crickets treated in Idaho and evaluated in Sidney were inconclusive due to premature death resulting from excessive temperatures that occurred in the Sidney portion of the experiment. Infection and mortality (corrected for control mortality) rates of Mormon crickets collected after field sprays and incubated indoors under ideal temperatures for the fungi were in excess of 70%. It is obvious that significantly longer observation times in the field will be required to detect any mortality in Mormon crickets that could be attributable to these fungal treatments. In fact, several weeks may be required after treatment for fungal infections to be expressed as mortality. However, there is some potential for faster fungal growth in Mormon cricket, particularly in the immature stages. Their aggregating behavior under inclement and nocturnal conditions may conserve and actually promote higher temperatures. The potential value of this aspect of their behavior will require additional study.
Acknowledgements
The authors wish to thank Lori Atkins and Donna Hooper for providing land used in the study, facilities for storage of equipment and supplies, shelter for the researchers during inclement weather, and their continued cooperation, since 2002, toward learning more about Mormon crickets and their management. Thanks are also extended to Dave McNeal and Rob McChesney for their continued assistance in locating and capturing appropriate aged populations of Mormon crickets for testing, and for providing unexpectedly needed miscellaneous equipment and supplies which made our efforts more efficient and to Rob Schlothauer USDA, Sidney, MT for technical assistance in the laboratory assays and incubations.
References Cited
Analytical Software Inc. 1998. Statistix® 8 for Windows. Analytical Software. Tallahassee, Florida.
Bradley, C. A., Wood, P. P., Black, W. E., Kearns, R. D., and Britton, J. 2002. Solid culture substrate including barley. US Patent Application Number 20020006650.
Fargues, J., M.S. Goettel, N. Smits, A. Oudraogo, and M. Rougier. 1997. Effect of temperature on vegetative growth of Beauveria bassiana isolates from different origins. Mycologia 89(3):383-392.
9

Foster, R.N., K.C. Reuter, L. Black, and J. Britton. 1996a. Evaluation of the fungus Beauveria bassiana with selected insecticide stressors for control of unconfined rangeland grasshoppers,1995. Arthropod Management Tests. 21: 280. Foster, R. N., K. Chris Reuter, Lonnie R. Black, Dave McNeal and Rob McChesney. 2004a. Comparison of wheat bran, apple pumice and food waste (combined bakery, snack, cereal, and confectionary) baits containing carbaryl for controlling Mormon crickets: Studies of longevity and primary and secondary mortality. Report. USDA-APHIS-PPQ-CPHST-DS&PMSL, Phoenix, AZ. 12 pp. Foster, R. N., K. C. Reuter, L. Black, E. Kahler, B. Fuller and E. A. Flora. 1996b. Laboratory evaluation of dose and manner of pickup of Spinosad on the migratory grasshopper, 1995. Arthropod Management Tests. (21) p 416. Foster, R. N., K. C. Reuter, C. Bradley, J. Britton, L. Black, S. Drake, L. Leisner, M. Tompkins, B. Fuller, M. Hildreth , E. Kahler, and B. Radsick. 1992. Development of the fungus Beauveria bassiana as a bio-insecticide for grasshoppers on rangeland. In. Cooperative Grasshopper Integrated Pest Management Project 1992 Annual Report. USDA-APHIS, Boise, Idaho. pp 207-215. Foster, R. N., K.C. Reuter, C. A. Bradley and P.P. Wood. 1991. Preliminary investigations on the effect of Beauveria bassiana on several species of rangeland grasshoppers. In. Cooperative Grasshopper Integrated Pest Management Project 1991 Annual Report. USDA-APHIS, Boise Idaho. pp.203-210. Foster, R.N. K. Christian Reuter, Jim Britton, and Cliff Bradley. 1996-1999. Lab Studies and Field Trials with the fungus Beauveria bassiana Against Grasshoppers. In: Cunningham, G.L., and M.W. Sampson tech cords. Grasshopper integrated pest management user handbook. Tech Bull. 1809. Washington, DC: U.S. Dept. of Agriculture, Animal and Plant Health Inspection Service: VII.5-1—VII.5-8.
Foster, R. N., K. Chris Reuter, Dave McNeal and Rob McChesney. 2003a. Comparison of flakey wheat bran and apple pumice baits for controlling Mormon cricket: Evaluating initial and secondary mortality with baits containing 2% and 5% carbaryl. Report. USDA-APHIS-PPQ-CPHST-DS&PMSL, Phoenix, AZ. 10 pp. Foster, R. N., K. C. Reuter, M. T. Tompkins, C. Bradley, J. Britton, N. Underwood, L. R. Black, E. Vigil, G. Daye, B. Radsick, B. Fuller, M. Brinkman and E. Kahler. Development of the fungus Beauveria bassiana as a Bio-insecticide for grasshoppers on rangeland. 1993. In. Cooperative Grasshopper Integrated Pest Management Project 1992 Annual Report. USDA-APHIS, Boise, Idaho. 233-238. Foster, R. N., S. T. Jaronski, K. C. Reuter, B. Helbig, J. Bradley, C. Huddleston, B. Fitzgerald, J. Grace, and B. Radsick. 2003b. Evaluation of the fungus Beauveria bassiana in canola oil and paraffinic oil and identifying the low dose of a bacteria derived toxin, Spinosad, for use against rangeland grasshoppers. Report.USDA-APHIS-PPQ-CPHST-DS&PMSL, Phoenix, AZ. 33 pp.
10

Foster, R. N., S. T. Jaronski, K. C. Reuter, L. Black, B. Helbig, L. Kozel, C. Huddleston and B. Radsick. 2004b. Field evaluation of the fungus Beauveria bassiana in unrefined canola oil and refined soybean oil and evaluation of potential attractants for use against grasshoppers on rangeland. Report. USDA-APHIS-PPQ-CPHST-DS&PMSL, Phoenix, AZ. 13 pp. Inglis, G.D., D.L. Johnson, and M.S. Goettel. 1996. Effects of temperature and thermoregulation on mycosis by Beauveria bassiana in grasshoppers. Biological Control 7(2):131-139 Inglis, G.D.; G.M. Duke, L.M. Kawchuk, and M.S. Goettel. 1999. Influence of oscillating temperatures on the competitive infection and colonization of the migratory grasshopper by Beauveria bassiana and Metarhizium flavoviride. Biological Control 14(2): 111-120 Jaronski, S. T., A. V. Chernysh, S. P. Schell, and J. A. Lockwood. 2002. Enhancement of Beauveria bassiana against grasshoppers by vegetable oil carriers. Poster. Entomological Society of America Annual Meeting. Ft. Lauderdale, Florida. Kleinbaum, D.G. 1996. Survival Analysis, A Self-Learning Text. Springer Verlag, New York NY. Lactin, D. J., and D. L. Johnson. 1998. Environmental, physical, and behavioural determinants of body temperature in grasshopper nymphs (Orthoptera: Acrididae). Can. Entomologist 130(5): 551-577. Lockwood, J. 2003. Canola Oil Proves to be an Effective Attractant-Carrier for Acridicides. In. Advances In Applied Acridology - 2003. The Association for Applied Acridology International. P.16. Reuter, K.C., R.N. Foster, L. Black, and J. Britton. 1996. Laboratory evaluation of Beauveria bassiana with an added chemical stressor, 1995. Arthropod Management Tests. 21:415. SPSS Inc. 1997. SYSTAT for Windows: Statistics, SPSS Inc., Chicago, Illinois.
Turnbow, J.H. 1998. Temperature-sensitive Beauveria bassiana mycoses in the Mormon Cricket, Anabrus simplex. M.S. thesis, Montana State University, Bozeman MT.
11

Figure 1. Hardware cloth cage to minimize cage effects.
Figure 2. Mormon cricket field cages on range vegetation near Boise, Idaho.
12

Figure 3. Using an airbrush system to deliver fungal pathogen sprays.
Figure 4. Containment chamber to eliminate drift during airbrush applications.
13

Figure 5. Mormon cricket surrogates positioned on ground and at plant canopy near cage studies.
Figure 6. Data logger used to record surrogate temperatures.
14

0
10
20
30
40
50
1 2 3 4 5 6 7 8 9 10 11 12 13 14
Days after treatment
Mea
n %
mor
talit
y (4
reps
)
Beauveria baitMetarhizium baitCheck
Figure 7. Beauveria bassiana and Metarhizium anisopliae baits against immature Mormon crickets in field cages – Boise, Idaho (April 29 – May 15, 2005)
Figure 8. Laboratory bioassay of fungal formulations of Tast-E-Bait
15

0
10
20
30
40
50
1 2 3 4 5 6 7 8 9 10 11 12 13 14
Days after treatment
Mea
n %
mor
talit
y (4
reps
)
Beauveria sprayMetarhizium sprayOil only sprayCheck
Figure 9. Beauveria bassiana and Metarhizium anisopliae sprays against immature Mormon crickets in field cages – Boise, Idaho (April 29 – May 15, 2005)
2005 Mormon Cricket Trial #1 Mormon Cricket Surrogate "Body Temperatures"
-10
0
10
20
30
40
50
4/28 4/29 4/30 5/1 5/2 5/3 5/4 5/5 5/6 5/7 5/8 5/9 5/10 5/11 5/12 5/13 5/14 5/15
Date: Time
Tem
pera
ture
(C.)
Inside, On groundInside, in canopy
Figure 10. Surrogate temperatures inside cage on the ground and at canopy level.
16

2005 Mormon Cricket Trial #1 Mormon Cricket "Body Temperatures"
-10
0
10
20
30
40
50
4/28 4/29 4/30 5/1 5/2 5/3 5/4 5/5 5/6 5/7 5/8 5/9 5/10 5/11 5/12 5/13 5/14 5/15
Tem
pera
ture
(C.)
Open groundOpen canopy
Figure 11. Surrogate temperatures outside cage on the ground and at canopy level.
Mormon Cricket "Body Temperatures"Inside Cage, On Ground
16.0014.17 13.00
16.2513.75
16.75 17.25
20.7518.25 17.75
21.0022.50 22.00
14.00 13.25
7.00
6.83
4.33
3.42
4.00
5.75 4.50
3.25
5.003.25
3.001.50 2.00
9.50
3.50
1.003.00
6.674.33
6.25
1.50 2.250.00 0.75
3.00
0.00 0.00 0.00 0.50
7.25
0
5
10
15
20
25
38471 38472 38473 38474 38475 38476 38477 38478 38479 38480 38481 38482 38483 38484 38485
Hou
rs p
er d
ay
>30C18-30C<18C
Figure 12. Accumulated hours of optimal and sub-optimal exposure/day inside cage on the ground.
17

Mormon Cricket "Body Temperatures"Inside Cage, Near top
12.33 12.3311.00
12.3310.83
7.42
0.253.00
5.003.00
12.5013.75 12.75
10.758.50
10.678.67
6.33
7.33
6.9215.08
21.50
21.0018.25
18.00
11.5010.25 11.25
12.75
8.25
1.003.00
6.674.33
6.25
1.50 2.250.00 0.75
3.00
0.00 0.00 0.00 0.50
7.25
0
5
10
15
20
25
38471 38472 38473 38474 38475 38476 38477 38478 38479 38480 38481 38482 38483 38484 38485
Hou
rs p
er d
ay
>30C18-30C<18C
Figure 13. Accumulated hours of optimal and sub-optimal exposure/day inside cage at canopy level.
Mormon Cricket "Body Temperatures"Outside Cage, On Ground
16.3314.67
11.67
15.0013.58
15.6717.00
20.25
17.50 17.7520.25
19.0021.75
13.50 13.25
5.83
5.33
4.00
3.58
6.33 3.75
3.50
4.753.00
3.755.00
2.25
4.502.50
2.42 3.50
7.005.00
6.83
2.003.25
0.251.75
3.25
0.00 0.00 0.00
6.008.25
5.25
0
5
10
15
20
25
38471 38472 38473 38474 38475 38476 38477 38478 38479 38480 38481 38482 38483 38484 38485
Hou
rs p
er d
ay
>30C18-30C<18C
Figure 14. Accumulated hours of optimal and sub-optimal exposure/day outside cage on the ground.
18

Mormon Cricket "Body Temperatures"Outside Cage, 18 cm high
17.33
14.2511.42 11.42
13.42
16.4217.75
21.0018.25 17.50
21.2523.75
21.75
13.00 12.75
6.67
9.58
11.83 11.83
10.58
7.58 5.50
3.00
5.755.75
2.75
0.252.25
11.00
5.75
0.00 0.17 0.75 0.75 0.00 0.00 0.75 0.00 0.00 0.75 0.00 0.00 0.00 0.00
5.50
0
5
10
15
20
25
38471 38472 38473 38474 38475 38476 38477 38478 38479 38480 38481 38482 38483 38484 38485
Hou
rs p
er d
ay
>30C18-30C<18C
Figure 15. Accumulated hours of optimal and sub-optimal exposure/day outside cage at canopy level.
19
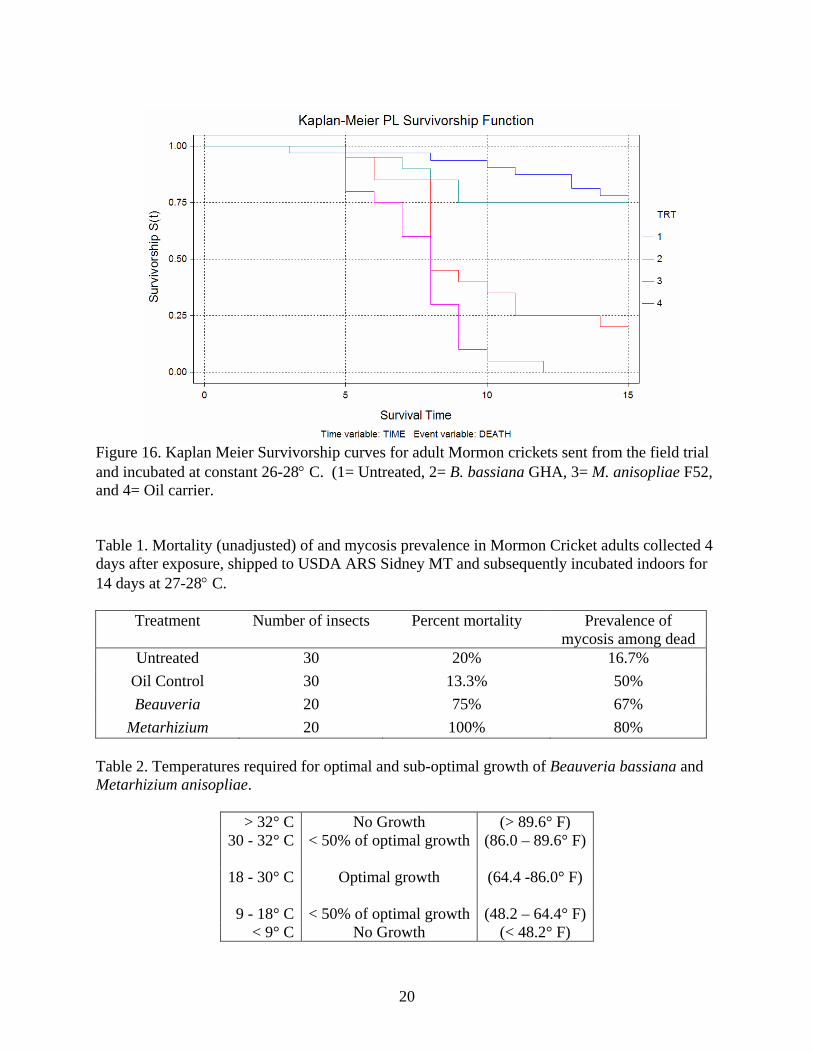
Figure 16. Kaplan Meier Survivorship curves for adult Mormon crickets sent from the field trial and incubated at constant 26-28° C. (1= Untreated, 2= B. bassiana GHA, 3= M. anisopliae F52, and 4= Oil carrier.
Table 1. Mortality (unadjusted) of and mycosis prevalence in Mormon Cricket adults collected 4 days after exposure, shipped to USDA ARS Sidney MT and subsequently incubated indoors for 14 days at 27-28° C.
Treatment Number of insects Percent mortality Prevalence of mycosis among dead
Untreated 30 20% 16.7% Oil Control 30 13.3% 50% Beauveria 20 75% 67%
Metarhizium 20 100% 80% Table 2. Temperatures required for optimal and sub-optimal growth of Beauveria bassiana and Metarhizium anisopliae.
> 32° C No Growth (> 89.6° F) 30 - 32° C < 50% of optimal growth (86.0 – 89.6° F)
18 - 30° C Optimal growth (64.4 -86.0° F)
9 - 18° C < 50% of optimal growth (48.2 – 64.4° F)
< 9° C No Growth (< 48.2° F)
20

Table 3. Accumulated hours of potential fungal development in Mormon cricket surrogates at 18-30° C (duration of field cage study – 15 days) and expected days required for mortality to occur.
Outside cages Inside cages ground canopy ground Canopy
63 100 67 188 Metarhizium anisopliae required growth hours 144 – 192 (6-8 days)
Outside cages Inside cages ground canopy ground Canopy
min – 2.3X min – 1.4X min – 2.1X min – 0.8X (34.5 days needed) (21 days needed) (32 days needed) (12 days needed)
max – 3.0X max – 1.9X max - 2.9X max – 1.0X (45 days needed) (28.5 days needed) (43 days needed) (15 days needed)
Beauveria bassiana required growth hours 240-336 (10-14 days) Outside cages Inside cages
ground canopy ground Canopymin – 3.8X min – 2.4X min – 3.6X min – 1.3X
(57 days needed) (36 days needed) (57.7 days needed) (19.1 days needed) max – 5.3X max – 3.4X max – 5.0X max – 1.8X
(80 days needed) (50 days needed) (75.2 days needed) (26.8 days needed)
Table 4. Cox’s Proportional Hazards analysis based on the survivorship of adult Mormon crickets collected post application and incubated indoors (see Table 1).
Variable Coefficient Std Error Z P Rel. Risk Untreated
vs. Oil 0.05343 0.19526 0.27 0.7843 1.05
Untreated vs.
Beauveria 1.76189 0.46437 3.79 0.0001 5.82
Untreated vs.
Metarhizium 1.52518 0.29794 5.12 0.0000 4.6
Beauveria vs.
Metarhizium 0.74506 0.35219 2.12 0.0344 2.11
21

Oct. 28, 2005
Field evaluations of aerially applied wheat bran and apple pumice baits containing carbaryl for controlling overwintering rangeland grasshopper species in South
Dakota in April
FINAL REPORT
Nelson Foster1, Bruce Helbig2, Chris Reuter1, Amy Mesman2, Bruce Radsick3, Daryl Hill3 , Tim Roland3 and Charles Brown4
Organizations 1USDA-APHIS-PPQ-CPHST, Decision Support and Pest Management Systems Laboratory, Phoenix, Arizona 2USDA-APHIS-PPQ, Pierre, South Dakota 3USDA-APHIS-PPQ, Aircraft and Equipment Operations, Mission, Texas 4USDA-APHIS-PPQ, Riverdale, Maryland
1

Abstract
Populations of the overwintering species Pardalophora haldemani and Eritettix simplex and the early season species Melanoplus confusus demonstrated susceptibility to aerially applied spring treatments of wheat bran or apple pumice baits containing carbaryl. Pardalophora haldemani (99.5% control) and M. confusus (96.9% control) were highly susceptible to wheat bran bait. Eretittex simplex was less susceptible to apple pumice bait (68.5% control) and wheat bran bait (65.9% control). The wheat bran bait treatment resulted in significantly greater mortality on combined species populations (83%) compared to the apple pumice bait (51%). The results indicate that wheat bran and apple pumice carbaryl baits can be significantly effective against overwintering species of rangeland grasshoppers treated in the spring. Comparisons with previous year late season control treatments indicate the early season alternative as the best choice because minimal grasshopper feeding, development, and damage to forage occurs over the winter and significant mortality to the population during the winter is possible. Additionally, a comparison of grasshopper density estimation methods confirmed earlier studies, that demonstrated when the mean grasshopper age nears 3.00 (third instar) and above, the sweep net method results in an underestimate of grasshopper density, compared to the visual ring method.
2

Introduction
Control or suppression of damaging populations of rangeland grasshoppers in the western United States typically occurs during the summer months, particularly June and July. This is when the majority of grasshopper species on western rangelands are in the late instar stages or are young adults. However, a few species are seen very early in the year because they over-winter in the nymphal stage. Populations of these overwintering species, individually or combined, rarely reach levels that cause significant damage, particularly over large areas, and are usually ignored as a potential rangeland problem. However, populations of Pardalophora haldemani, a mostly mixed feeder of grasses with little preference for specific species (Mulkhern et al. 1964; 1969), have reached damaging levels serious enough to require treatments in 1989 and 1990 (Utah), 2002 and 2003 (Nebraska) and in 1990, 2004 and 2005 (South Dakota). It is noteworthy that remarkable numbers of this species also occurred in July of 1932 when nocturnal flights of great magnitude were reported throughout southern South Dakota and northeastern Nebraska (Parker and Connin 1964.) Unquestionably, P. haldemani has accounted for the majority of outbreaks of over-wintering rangeland grasshopper species in the west (Foster et al. 2005). While these occurrences are not unique they are certainly rare but nevertheless require appropriate attention. USDA APHIS sponsored cooperative programs are typically organized toward applying treatments in the summer months after seasonal hires are added for survey and treatment activities. The phenology of P. haldemani places it out of synchrony with the typical rangeland grasshopper activities and the availability of seasonal personnel necessary for conducting control programs. Therefore, treatment timing options are necessary either much earlier or much later than usual. A study of fall treatments showed sprays of carbaryl, diflubenzuron and malathion produced little control while laboratory and field cage studies with solid baits indicated excellent control (Foster et al. 2005). That study also indicated that because of minimal grasshopper feeding, development and damage to forage over the winter, there appeared no economic reason to treat in the fall rather than early in the following spring. The following study was conducted early in the spring to evaluate two solid bait treatments containing carbaryl for control or suppression of populations of Pardalophora haldemani on rangeland in central South Dakota. The study also provided an excellent opportunity to gather operational scale, carbaryl exposure data on personnel loading the baits and pilots applying the baits. The Worker Exposure Study conducted by the Agricultural Handlers Task Force and APHIS is presented by others in a separate report.
3

Objectives Determine the feasibility of early season (April ) treatments against mainly immature stages of overwintering rangeland grasshopper species, particularly Pardalophora haldemani, in central South Dakota. Determine and compare in the field the efficacy of early season (April) aerially applied wheat bran and apple pumice baits containing carbaryl in treatments against overwintering grasshopper species in central South Dakota. Provide an operational program situation to facilitate the collection of data on loader and pilot exposure to carbaryl in activities associated with and during aerial application of bait. This portion of the study was conducted under Good Laboratory Practices and will be available in a separate report. Compare ring delimited visual and sweep net methods of estimating grasshopper densities of overwintering and early hatching species of rangeland grasshoppers.
Methods and Materials
Study Site The study was located in Hughes County, South Dakota near the “North Bend Recreation Area” of the Missouri River on the Crow Creek Indian Reservation and land owned by Charlotte Hyde Eucker and the Chaney-Rush Grazing Association. The centers of the apple pumice bait and wheat bran bait plots were located ca. 1.75 miles north and 8 miles west and ca. 0.75 miles north and 11 miles west of Mac’s Corner, respectively, Figure 1.The general location was selected because of the high density of Pardalophora haldemani, the recent history of problems with this species in the area and the abundance of contiguous P. haldemani infested rangeland. Treatments and Experimental Design The 5% Eco Bran (wheat bran, Peacock Industries Inc.) and Sevin 5% Bait (apple pumice “crumbles”, Wilbur-Ellis) formulations were aerially applied to 320 and 640 acre plots respectively. The baits were applied at 100% coverage to plots each containing 6 evenly distributed predetermined sampling sites to be used for determining grasshopper densities. Six additional sampling sites were also established on untreated rangeland adjacent to each plot for comparison (3 per plot). The 5% EcoBait and Sevin 5% Bait were applied on April 26 and 27, 2005 respectively. Treatments were applied with a Cessna Ag Truck owned by the USDA, Animal and Plant Health Inspection Service (APHIS) and operated by a USDA – APHIS pilot. The aircraft was equipped with a standard commercial Transland Slim-line, 20241 dry material spreader and a differentially corrected guidance and recording system. Ground personnel also provided guidance and ensured acceptable operating parameters during application. All applications occurred from an altitude of 40 to 60 feet. Prior to application the aircraft
4

spreader system was calibrated to deliver bait at the desired rate of 10 lbs/acre. Calibration was accomplished by measuring the amount of material loaded and remaining in the hopper after an application flight of predetermined time, and making adjustments in the gate opening and hopper pressure (vent tube) until the desired output was achieved (Foster and Roland, 1996). The aircraft was operated at 115 mph and calibrated for 45 and 50 feet wide swaths for Ecobait and Sevin Bait, respectively. However, an evaluation of the total amount of material applied and acres treated in the field indicated that the actual applications were 11.16 and 9.29 lbs /acre for Ecobait and Sevin bait respectively. With Ecobait the hopper gate was open 1,540 sec. during 52 passes in which a total of 3,152.5 lbs of bait was dispensed. With Sevin bait, the hopper gate was open 3,420 sec. during 54 passes in which a total of 6,404.5 lbs of bait was dispensed. Humidity and atmospheric pressure may have accounted for the differences in calibration and application rates since the system was calibrated in Mission, Texas but used in South Dakota. While specific wind speeds were not recorded during application it is noted that significant wind occurred during applications on both days. Additionally, even though some precipitation was recorded at the Dakota Lakes Research Farm located ca. 10 miles west of the study area on the days of application, (0.02 and 0.30, April 26 and 27 respectively) and at the Stephan weather station located ca. 12 miles east of the study area (a trace for both April 26 and 27), none occurred during application. The Daily precipitation and mean temperatures as well as averages from 1971 – 2000 recorded at the Stephan weather station for April 24 through May 31 are shown in Figure 2. Grasshopper Density Sampling Generally, grasshopper density and species composition sampling followed protocols established by Foster and Reuter, 1996. Grasshopper populations in treated and untreated plots were counted and sampled 2 to 3 days before treatment (April 24, 2005) and at 19 to 20 days after treatment (May 16, 2005). Untreated control plots were also counted and sampled on any day a treated plot was monitored. Grasshopper densities were determined by counting grasshoppers in (40) 0.1 m2 rings arranged in an approximate 100 yard diameter circle at each of 6 sites in each plot. Rings were separated from adjacent rings by ca. 5 yards. All counts were conducted by the same person (USDA APHIS PPQ, Pierre, South Dakota). The abundance of each species was determined from sweep samples taken, uniformly at each site. With this method, which is employed by federal and state personnel in North Dakota and South Dakota, 40 sweeps, each 180o in length, were conducted with a 15 inch diameter standard sweep net on a 3 foot long handle. Each sweep was performed by dragging the net firmly along the ground with a rapid motion while walking at a moderate pace through the site area. After collection, samples were placed in paper bags, labeled, and cold stored in the laboratory until they could be sorted, identified and counted. The total number of grasshoppers collected in each sample was divided by 10 to arrive at the number of grasshoppers per square yard (Kantack, 1990). This technique resulted from studies conducted in the late 1970’s by Gregory Mulkhern, North Dakota State University (Keith Winks, USDA-APHIS-PPQ retired - personal communication Aug. 25, 2005).
5

Population Maturity Index and Mean Instar Age To quantify the age of the grasshopper populations at each time interval, a population maturity index was developed, Foster et al. 1989. The population maturity index is the ratio of present development to the maximum possible development of a given population. The population maturity index, a ratio of the amount of existing population maturity to the potential population maturity, is described by the equation PMI=6a+5b+4c+3d+2e+f / 6(a+b+c+d+e+f) where a through f are the number of individuals occurring in each of the respective stages of development, adult through first instar stages. For example, if a sample contained 100 individuals and was comprised of 3 adults, 10 fifth instars, 60 fourth instars, 15 third instars, 10 second instars and 2 first instars, the population maturity index would be 6(3) + 5(10) + 4(60) + 3(15) + 2(10) + 1(2) / 6(3+10+60+15+10+2) and 18 + 50 + 240 + 45 + 20 +1 / 6(100) and equals 374 / 600 and equals 0.623, or 62.3 % of total maturity. An index of one would indicate that the population is composed of all adults. To arrive at the mean instar age of a given population, the population maturity index is simply multiplied by 6. In the example, when the population maturity index is 0.623 the mean instar age would be 0.623 x 6 or 3.738 and indicates that the mean age of the population is almost fourth instar, 4.0. Analysis Percentage control data were converted to rank data (Conover and Iman, 1981). An analysis of variance was performed with the Tukey multiple comparison test used to separate means. All analyses were performed with Systat 6.1 For Windows, SPSS Inc. 1997. The three lowest UTC sites were excluded from the analysis due to extremely low numbers.
Results and Discussion Field Evaluation of Baits. Pre-treatment densities from individual sites ranged from 3 to 31 grasshoppers /yd2 in the treated plots and from 2 to 23 grasshoppers /yd2 in the untreated plots. The mean densities were 28, 4 and 10 grasshoppers /yd2 in the Ecobait, Sevin Bait and untreated plots respectively. At the time of treatments the populations were composed of first instars (7%), second instars (1%), third instars (19%), fourth instars (46%), fifth instars (17%) and adults (10%). The total mean instar age was 3.98 (near 4th instar) on April 4, 2005 . At the time of treatment only three different species were found in the study area, Pardalophora haldemani (71%), Eritettix simplex (21 %) and Melanoplus confusus (8%). The relative abundance and associated age of all grasshopper species in pre-treatment and post-treatment samples are shown in Table 1. At 19-20 days after treatment, Ecobait and Sevin Bait treatments resulted in mean total population percentage reductions of 83% and 51%, respectively. During the same time, untreated populations declined 33%, Table 2. Both Ecobait and Sevin bait treatments reduced populations significantly more than occurred in the untreated population.
6

However, Ecobait resulted in significantly greater population reductions compared to Sevin 5% bait. Three species were evaluated individually for susceptibility to the baits. Unfortunately, extremely low numbers of grasshoppers captured in the sweep net samples (Table 1) prevented analyses of any individual species except Eritettix simplex in the apple pumice plot. When sufficient data allowed for analyses, the apple pumice bait demonstrated 68.5% control of E. simplex. With substantially more data, the wheat bran bait demonstrated 65.9% control of E. simplex, 96.9% control of Melanoplus confusus and 99.5% control of Pardalophora haldemani (Table 3). The relative levels of susceptibility to carbaryl bait seen in this study are consistent with those reported by Onsager et al. 1996. Both studies show E. simplex as less susceptible than M. confusus. However, in our study the mortality levels for both species are consistently higher than those reported by Onsager et al. 1996. These higher levels may be attributed to the higher application rate used in this study. Rangeland grasshoppers are usually treated at 1.5 lbs/acre when applied by air and 2.0 lbs/acre when applied by ground. A 10 lb/acre rate, commonly used for controlling Mormon crickets on rangeland was chosen for this study to accommodate the human exposure data that was being collected. While no data exists in the Onsager et al. 1969 study on P. haldemani, it is clear from this study and an earlier late season study (Foster et al. 2005) that P. haldemani is highly susceptible to carbaryl bran and apple pumice baits. Data on P. haldemani from this study is very consistent with the earlier study that showed 5% carbaryl wheat bran baits producing 94% and 96% mortality in laboratory and field cages respectively (Foster at al. 2005). Additionally, in that study, 5% carbaryl apple pumice bait resulted in 90% and 86% mortality to P. haldemani in laboratory and field cages, respectively. Additionally, experience with baits by the senior author has previously indicated Camnula pellucida to probably be the most sensitive species to wheat bran bait an occurrence which appears now to be rivaled by Pardalophora haldemani. Interestingly, both species are bandwinged (Oedipodinae). From April 26 through May 16 the average of daily mean temperatures during this study (43.8o F) were 8.96o F cooler than historic averages (52.8o F). During the first 8-9 days after treatments only two days had maximum temperatures above 50o F (56o F on April 26 and 52o F on May 4). This compares to historical averages of maximum daily temperatures ranging from 62o-65o F during the same dates. These cool temperatures experienced in our study may have affected maximum feeding. Therefore, feeding per given unit time may be greater with resulting mortality occurring sooner in average years. The feeding threshold temperature for this species is not documented. However, it is worthy to note, that several studied non-overwintering species that are susceptible to wheat bran bait, commence feeding activity at ca. 80 o F. (Jech, 1996-1999). This species has most likely evolved to feed at lower temperatures than the non overwintering species. Studies to determine developmental rates and number of instars that occur at different temperatures are currently under way.
7

While solid baits appear a good choice for controlling grasshoppers, they are less economical and more difficult to apply correctly compared to sprays. However, liquid sprays are more susceptible to negative impacts that can be attributed to less than optimal meteorological conditions such as cool temperatures, high winds and rain that can occur in the spring. Comparison of density estimation methods Densities at all mean ages (3.44 – 6.00) were underestimated by the sweep method when compared to the visual method, Table 4. Generally, a higher mean age was directly related to a higher disparity between visual and sweep net estimates of grasshoppers. Estimates with visual methods in our study ranged from 1.7 (28.1 vs. 16.5) to 67.6 (2.0 vs. 0.03) times higher than sweep net estimates. This was not surprising and is consistent with comparative studies of the two methods reported by Foster et al. 1989. In that study, the sweep net method began to underestimate densities when the grasshopper mean age was below 3.00.
Conclusions
The overwintering species, Pardalophora haldemani, and the early hatching species, Melanoplus confusus, are very susceptible to carbaryl wheat bran bait. Another over-wintering species, Eretittex simplex, demonstrated somewhat less susceptibility to both wheat bran and apple pumice baits. The wheat bran bait treatment resulted in significantly greater mortality on combined species populations (83%) compared to the apple pumice bait (51%). The results indicate that wheat bran and apple pumice carbaryl baits can be effective against overwintering species of rangeland grasshoppers treated in the spring. There is no question that Ecobait (carbaryl wheat bran bait) is highly effective in reducing populations of Pardalophora haldemani and in the senior authors view, rivals Camnula pellucida as the species most susceptible to carbaryl bran bait. While Sevin 5% (apple pumice bait) also demonstrated effectiveness, further study should be conducted to determine if apple pumice bait is less effective than wheat bran against older grasshoppers and more commonly treated species. Additionally, the grasshopper density estimation data confirmed earlier studies that demonstrated when the mean grasshopper age nears 3.00 (third instar) and above, the sweep net method results in an underestimate of grasshopper density, compared to the visual ring method. Solid baits are only one treatment option and when early spring treatments are planned, liquid sprays should also be considered. While liquid sprays may be economically advantageous compared to baits, they are also more susceptible to negative impacts attributable to less than optimal early spring meteorological conditions that may occur. Because of the potential for difficulties associated with liquid sprays and spring
8

meteorological conditions, solid baits should be seriously considered when unusually early season treatments are necessary.
Acknowledgements
The authors thank the Crow Creek Indian Tribe, Charlotte Hyde Eucket and the Chaney-Rush Grazing Association for allowing the study to be conducted on their land. Additionally, we thank the USDA APHIS PPQ Western Region for funding the study separately and in addition to normal funding mechanisms and schedules. Thanks are also extended to Dennis Todey, South Dakota Extension / State Climatologist for providing recent and historical weather data. Special thanks are extended to Charlie Brown for organizing the human exposure study and in loading the aircraft. Additionally, thanks are extended to Tracey Horner, Roeland Ellison, and Larry Law for loading the aircraft and to Richard Honneycutt, Dan Hill, Brian DeGeare, Kissette Collier, and Lisa Mosser for conducting the exposure sampling, which will be reported separately.
9

References Cited
Conover, W.J. and R.L. Iman. 1981. Rank Transformations as a Bridge Between Parametic and Nonparametric Statistics. The American Statistician. 35: (3) 124-129. Foster, R.N., W.J. Cushing, M. Smith, T. Battaglia and S. Battaglia. 1989. Standardization of rangeland grasshopper survey methodology: a comparison of visual, ring and sweep net techniques in estimating grasshopper populations on rangeland. In: USDA-APHIS-PPQ Grasshopper Integrated Pest Management Project. Annual Report (Fiscal Year 1989). pp.165-172.
Foster, R.N. and K.C. Reuter. 1996-1999. Evaluation of rangeland grasshopper controls: A general protocol for efficacy studies of insecticides applied from the air. In: Cunningham, G.L., and M.W. Sampson tech cords. Grasshopper integrated pest management user handbook. Tech Bull. 1809. Washington, DC: U.S. Dept. of Agriculture, Animal and Plant Health Inspection Service: II.2-1—II.2-7.
Foster, R.N., B. Helbig, C. Reuter, L. Black, A. Mesman, and B. Radsick. 2005. Controlling the overwintering rangeland grasshopper, Pardalophora haldemani, in South Dakota in September: Field evaluations of aerially applied sprays with cage evaluations of three types of solid baits. Report. USDA, APHIS, PPQ, DS&PMSL, Phoenix, Arizona. 23pp. Foster, R.N. and T.J. Roland. 1996-1999. Equipment Modification, Swath Width Determination, and Calibration for Aerial Application of Bran Bait With Single-Engine Fixed-Wing Aircraft. In: Cunningham, G.L., and M.W. Sampson tech cords. Grasshopper integrated pest management user handbook. Tech Bull. 1809. Washington, DC: U.S. Dept. of Agriculture, Animal and Plant Health Inspection Service: II.18-1—II.18-11. Jech, Larry. 1996. What, when and where do grasshoppers eat. In: Cunningham, G.L., and M.W. Sampson tech cords. Grasshopper integrated pest management user handbook. Tech Bull. 1809. Washington, DC: U.S. Dept. of Agriculture, Animal and Plant Health Inspection Service. II.13-1 – II.13.2.
Kantack, Benjamin H. 1990. Grasshopper Control Economics Thresholds. Cooperative Extension Service, South Dakota State University, US Department of Agriculture, Brookings , South Dakota. FS 859. Mulkhern, G. B., D. R. Toczek and M. A. Brusvan. 1964. Biology and ecology of North Dakota grasshoppers II. Food habits and preferences of grasshoppers associated with the sand hills prairie. Researsch Report No. 11. North Dakota Agricultural Experiment Station, Fargo, North Dakota. 59 pp.
10

11
Mulkhern, G. B., K. E. Pruess, H. Knutson, A. F. Hagen, J. B. Campbell and J. D. Lambley. 1969. Food habits and preferences of grasslands grasshoppers of the North Central Great Plains. Bulletin No. 481. Agricultural Experimental Station, North Dakota State University, Fargo, North Dakota 58102. 32 pp. Onsager, J.A., R.N. Foster and L. Jech. 1996. Bait Acceptance by Different Grasshopper Species and Instars. In. Cunningham, G.L., and M.W. Sampson tech cords. Grasshopper integrated pest management user handbook. Tech Bull. 1809. Washington, DC: U.S. Dept. of Agriculture, Animal and Plant Health Inspection Service. II.12-1 – II.12-5.
Parker, J. R., and R. V. Connin. 1964. Grasshoppers: Their habits and damage. U.S. Dept. of Agriculture . Info. Bull. 287: 1-28. SPSS Inc. 1997. SYSTAT for Windows: Statistics, SPSS Inc., Chicago, Illinois.

Table 1. Grasshopper species composition in the study area on the Crow Creek Indian Reservation near Pierre, South Dakota – 2005.
Pre-treatment (04/24/2005) Post-treatment (05/16/2005) Instars Instars Treatment/Species 1 2 3 4 5 Adults Total % 1 2 3 4 5 Adults Total % Apple pumice Aeropedellus clavatus 0 0 0 0 0 0 0 0 1 0 0 0 0 0 1 50 Eritettix simplex 0 0 0 0 0 6 6 100 0 0 0 0 0 1 1 50
Total 0 0 0 0 0 6 6 1 0 0 0 0 1 2 % 0 0 0 0 0 100 50 0 0 0 0 50
Wheat Bran Eritettix simplex 0 0 0 3 120 115 238 24 0 0 0 0 7 36 43 96 Melanoplus confusus 51 5 0 0 0 0 56 6 0 0 0 0 0 0 0 0 Pardalophora haldemani 0 0 190 447 61 0 698 70 0 0 0 0 2 0 2 4
Total 51 5 190 450 181 115 992 0 0 0 0 9 36 45 % 5 1 19 45 18 12 0 0 0 0 20 80
Untreated Eritettix simplex 0 0 0 1 9 7 17 7 0 0 0 0 0 9 9 7 Melanoplus confusus 41 4 0 0 0 0 45 19 1 9 14 2 0 0 26 19 Pardalophora haldemani 0 0 40 116 15 0 171 73 0 0 0 16 85 1 102 74
Total 41 4 40 117 24 7 233 1 9 14 18 85 10 137 % 18 2 17 50 10 3 1 7 10 13 62 7
12

Table 2. Field efficacy of selected carbaryl baits against predominantly overwintering grasshoppers in April and May, 2005 near Pierre, South Dakota. Mean pre- & post-treatment grasshopper density & percentage reduction Treatment pre-treatment mean % reduction 1post-treatment
4/24 5/16 19 – 20 DAT Eco Bait 5% 28.1 4.6 83 a Sevin 5% 4.4 2.0 51 b Untreated 9.9 8.1 33 c 1 Rank transformations of the data were evaluated by analysis of variance with a Tukey multiple comparison method. Means in a column followed by the same letter are not significantly different (P ≤ 0.05). Table 3. Total percentage control (based on the sweep net method) of selected species treated with carbaryl baits in April and May, 2005 near Pierre, South Dakota. Treatment Species Percentage Control1
Apple pumice Eritettex simplex 68.5 Wheat bran Eritettix simplex 65.9 Melanoplus confusus 96.9 Pardalophora haldemani 99.5 1Corrected for untreated check mortality.
13

Table 4 Comparison of grasshopper density and (percentage reduction) with visual and sweep net methods.1
Pre-count 4/24/05 Post-count 5/16/05 Treatment Visual Sweep Mean Age Visual Sweep Mean Age Eco Bait 28.1 16.5 4.05 4.6 (84) 0.75 (95) 5.74 Sevin Bait 4.4 0.1 6.00 2.0 (77) 0.03 (70) 6.00 Untreated 9.9 3.9 3.44 8.1 (18) 2.30 (41) 4.51 1 Density recorded as grasshoppers/yd2.
Figure 1. The study area, east-southeast of Pierre, South Dakota, is shown in the above map. Green or red diamond symbols represent sites in the wheat bran or apple pumice carbaryl bait plots, respectively and a circle with an x inside represents the untreated sites for each treatment plot.
14

0
10
20
30
40
50
60
70
80
4/24 4/27 4/30 5/3 5/6 5/9 5/12 5/15 5/18 5/21 5/24 5/27 5/30
2005
Tem
pera
ture
o F
00.20.40.60.811.21.41.61.8
Prec
ipita
tion
(inch
es)
Daily mean °F Historic daily mean °F Daily precipitation Historic daily mean precipitation
Figure 2. Daily means and amounts and historical (1971-2000) daily means for temperature and precipitation observed at the Stephan climatological recording station in Hyde County, South Dakota. (Arrows indicate treatment dates)
15

Sept. 14, 2005
Controlling the overwintering rangeland grasshopper, Pardalophora haldemani, in South Dakota in
September: Field evaluations of aerially applied sprays with cage evaluations of three types of solid baits.
Nelson Foster1, Bruce Helbig2, Chris Reuter1, Lonnie Black1, Amy Mesman2 and Bruce Radsick3
Organizations 1USDA-APHIS-PPQ-CPHST, Decision Support and Pest Management Systems Laboratory, Phoenix, Arizona 2USDA-APHIS-PPQ, Pierre, South Dakota 3USDA-APHIS-PPQ, Aircraft and Equipment Operations, Mission Texas
1

Controlling the overwintering rangeland grasshopper, Pardalophora haldemani, in South Dakota in September: Field evaluations of aerially
applied sprays with cage evaluations of three types of solid baits
Nelson Foster1, Bruce Helbig2, Chris Reuter1, Lonnie Black1, Amy Mesman2 and Bruce Radsick3
Adult female and second instar Pardalophora haldemani (Scudder)
Organizations
1USDA-APHIS-PPQ-CPHST, Decision Support and Pest Management Systems Laboratory, Phoenix, Arizona 2USDA-APHIS-PPQ, Pierre, South Dakota 3USDA-APHIS-PPQ, Aircraft and Equipment Operations, Mission Texas
Cooperators Todd Cowan and Dick Carmody, ranchers, Duane D. Big Eagle, Sr., Chairman, Crow Creek Sioux Tribe Indian Reservation Elton Hawkwing, Acting Superintendent, and Tony Willman, Range Conservationists, Bureau of Indian Affairs, Crow Creek Agency, Fort Thompson, South Dakota.
2

Abstract
Treatments of carbaryl, diflubenzuron and malathion sprays aerially applied in September were evaluated on rangeland for controlling the overwintering rangeland grasshopper species Pardalophora haldemani in central South Dakota. Additionally, evaluations of field and laboratory cage studies of wheat bran, apple pumice and food waste baits containing carbaryl were conducted. Among sprays, carbaryl clearly performed better than the others. At two weeks after treatment carbaryl, malathion, diflubenzuron and untreated plots showed mean percentage reductions of 64, 56, 0 and 28, respectively. However, grasshopper densities declined naturally and were nearly the same in treated and untreated plots the following spring. At 189 days after treatment carbaryl, malathion, diflubenzuron and untreated populations showed accumulated grasshopper feeding days per square yard of 1920, 4590, 4815 and 3822, respectively, with higher values indicating greater damage. Compared to untreated populations, malathion produced only a short term reduction, carbaryl produced both a short and long term reduction and diflubenzuron produced no significant reductions at any time. While significant, the reductions attributed to carbaryl and malathion were not high enough to be considered effective or economical. Additionally, even though substantial reductions had occurred naturally, grasshopper densities remained sufficiently high in the spring to cause substantial damage. Baits produced quicker and much more substantial mortality than sprays and are clearly the choice if fall treatments are required. At 7 days after treatment, mortality in field cages attributed to baits without molasses ranged from 74% to 90 %, while untreated populations declined by only 2%. Results of both laboratory and field cage studies of bait comparisons in September on second instar P. haldemani demonstrated no advantage with higher concentrations of carbaryl, 5% vs. 2%, for any of the three different types of bait. Additionally no advantage was seen with higher application rates, 10 lb vs 2 lbs /acre, in field cage studies. Substantial reductions in chemical, carrier and application costs will result from the lower carbaryl concentration and application rate uses. Such reductions are also environmentally and biologically attractive. The data suggest that spring treatments of this overwintering species would most likely be more economical and should be considered in lieu of fall treatments.
3

Introduction
Control and or suppression of damaging populations of rangeland grasshoppers in the western United States typically occurs during the summer months, particularly June and July. This is when the majority of grasshopper species on western rangelands are in the late instar stages or are young adults. However, a few species are seen very early in the year because they over-winter in the nymphal stage, compared to most species that over-winter in the egg stage. These early occurring species are usually the band-winged (Oedipodinae), Arphia conspersa, Chortophaga viridifasciata, Pardalophora apiculata, P. haldemani, Xanthippus corallipes and X. montanus and the slant-faced (Gomphocerinae) Eritettix simplex, Psoloessa delacatula and P. texana. Populations of these species, individually or combined, rarely reach levels sufficient to cause significant damage, particularly over large areas and are usually ignored as a potential rangeland problem. However, populations of Pardalophora haldemani, a mostly mixed feeder of grasses with little preference for specific species (Mulkhern et al. 1964; 1969), reached damaging levels in 2002 in several areas of Nebraska and required treatment that year conducted by private landowners on ca. 50,000 acres. In 2003 the problem became so severe in Nebraska that 10 separate cooperative private, state and federal control programs sponsored by the USDA APHIS PPQ were conducted in Buffalo, Custer, Dawson, and Sherman Counties to protect 291,622 acres. (Steve Johnson, USDA-APHIS-PPQ Lincoln, Nebraska – personal communication, 5/16/05). Although over lesser acreages, populations of this species also reached troubling levels in central South Dakota in 2004 and 2005 where 3,540 acres and 7,460 acres respectively were treated in APHIS sponsored programs. Additionally, ca. 8,000 acres in southwestern South Dakota were treated in May, 2005 by private landowners to control this species. Populations of Pardalophora haldemani had previously reached problem levels in South Dakota in 1990 when a total of 5,120 acres in central South Dakota were treated for control in two APHIS sponsored programs (May 23 – 1,600 acres treated with carbaryl and Sept 12 - 3,520 acres treated with malathion) with mixed successes. The May treatment was ineffective, yielding little, if any, mortality. However, it must be noted that in post program interviews it was learned that the carbaryl used was 8 years old, had settled out in the containers and was not remixed sufficiently to insure an adequate level of active ingredient (AI) applied in the final sprayed formulation. While the September treatment resulted in some mortality it was viewed as minimally acceptable at best and generally uneconomical by the program director. Mixed populations of Pardalophora haldemani and Xanthippus corallipes at densities up to 20 /yd2 in Millard, Beaver and Tooele counties of western Utah required at least two APHIS sponsored cooperative programs in each of both 1989 and 1990 to control damage (Greg Abbott, USDA-APHIS-PPQ, Richfield, Utah – personal communication 5/16/05). It is noteworthy that previous outbreaks of P. haldemani have also been recorded. Remarkable numbers of this species also occurred in July of 1932 when nocturnal flights of great magnitude were reported throughout southern South Dakota and northeastern Nebraska (Parker and Connin 1964.)
4

Other reports of populations of over-wintering species requiring treatment exist. In the late 1970’s a very large over-wintering species (perhaps Xanthippus) was treated for control in Utah (Tom Crowe, USDA-APHIS-PPQ retired – personal communication, 5/11/05). Pfadt (1994) reported outbreaks of Xanthippus corallipes at 18 to 20/yd2 damaging rangeland grasses, and cultivated crested wheatgrass in 1989 and alfalfa fields in 1990 in western Utah. In the late 1950’s a large grasshopper species near Casa Grande, Arizona was treated early in the year to protect cotton (Bruce Thornly, USDA-APHIS-PPQ retired – personal communication, 5/11/05). Unquestionably, P. haldemani has accounted for the majority of outbreaks of overwintering rangeland grasshopper species in the west. While these occurrences are not unique they are certainly rare but nevertheless require appropriate attention. USDA APHIS sponsored cooperative programs are typically organized toward applying treatments in the summer months after seasonal hires are added for survey and treatment activities. The phenology of the overwintering species and in particular Pardalophora haldemani places it out of synchrony with the typical rangeland grasshopper activities and the availability seasonal personnel necessary for control programs. Therefore, treatment timing options are necessary either much earlier or much later than usual. Early or late season treatment activities each pose somewhat unusual if not unique, meteorologically related grasshopper developmental and behavioral problems. All of the chemicals (carbaryl, diflubenzuron and malathion) used in APHIS approved treatments require ingestion to achieve optimal results (Pfadt et al. 1970 and Lloyd et al. 1974). Early or late season treatment programs potentially place the grasshopper in a significantly cooler environ that may cause reduced activity, including feeding and thus subsequent longer instar stadia development. Reduced activity impacts on the pickup (ingestion) of sufficient amounts of active ingredient to produce acceptable mortality within the available active residual window for the particular treatment. These same parameters also extend molting intervals which may have a substantial impact on the success of diflubenzuron, an insect growth regulator. The following study was conducted to evaluate potential treatment options exercised late in the season (Sept. 16-24) for control or suppression of populations of over-wintering species of grasshoppers, particularly Pardalophora haldemani, on rangeland in central South Dakota. These populations are the progeny of a problem population that existed earlier in the year.
Objectives Determine the feasibility of late season (September) treatments against primarily first and second instar stage of overwintering rangeland grasshopper species, particularly Pardalophora haldemani, in central South Dakota. Determine and compare in the field the potential for using traditional aerial sprays of carbaryl, diflubenzuron and malathion against first and second instar stage overwintering grasshopper species, particularly Pardalophora haldemani, in late September.
5

Evaluate and compare wheat bran, apple pumice and food waste baits containing carbaryl against first and second instar stage Pardalophora haldemani in laboratory and field cages. Compare two doses of carbaryl in wheat bran, apple pumice and food waste baits applied at selected rates against first and second instar stage Pardalophora haldemani in laboratory and field cages.
Methods and Materials
Field Study Site The center of the study was located in Hughes County, South Dakota, 3 miles west and one-half mile north of the “North Bend Recreation Area” of the Missouri River on the Crow Creek Indian Reservation and land owned by Todd Cowen. The general location was selected because of the high density of Pardalophora haldemani, the recent history of problems with this species in the area and the abundance of contiguous P. haldemani infested rangeland. Treatments and Experimental Design Field evaluations- The treatments studied were: (1) The Sevin XLR Plus formulation of carbaryl applied at 0.50 lbs AI/ac (16 fluid ounces). The treatment was applied at a 1:1 ratio with water buffered to < 7 ph with LI 700 surfactant, penetrant, acidifier, (Loveland Industries Inc. Greeley. CO) at a total volume of 32 oz/acre, (2) The Dimilin 2L formulation of diflubenzuron applied at 0.015625 lb AI/ac = 7.1 grams AI /ac (1 fluid ounce). The treatment was applied specifically in 31 total fluid ounces/ac containing one fluid ounce of Dimilin 2L, 10 fluid ounces of CornbeltR Premium Crop Oil Concentrate and 20 fluid ounces of water / acre and (3) The Fyfanon ULV formulation of malathion applied at 0.61 lb AI/ac (8 fluid ounces). Each treatment was applied at 100% coverage to a single 320 acre plot containing 6 evenly distributed predetermined sampling sites. Severe inclement weather prevented all treatments from being applied on consecutive days. The malathion, diflubenzuron and carbaryl were applied on Sept. 16, 17 and 24, 2004 respectively. Treatments were applied with a Cessna Ag Truck owned by the USDA, Animal and Plant Health Inspection Service (APHIS) and equipped with winglets (DBA- Ag Tips: Clack Oberholtzer, Alberta, Canada). Winglets are added to spray aircraft to reduce the production of fine droplets and to improve handling characteristics. The aircraft was operated by a USDA – APHIS pilot. The aircraft was equipped with a standard commercial spraying system and differentially corrected guidance and recording system. Ground personnel also provided guidance and ensured acceptable operating parameters during application. All applications occurred from an altitude of 40 to 60 feet. Prior to application the aircraft spray system was calibrated to operate under parameters which resulted in delivery of spray within 1% of the desired rate per acre. Calibration was accomplished by collecting and measuring the amount of material sprayed through each nozzle for a predetermined
6

amount of time, and making adjustments in pressure until the desired output was achieved. The aircraft was calibrated for a 100 foot wide swath for all treatments. The specific mix, total volume applied per acre, number of nozzles, nozzle screen size, nozzle tip size, boom pressure, aircraft speed, and swath width used for each of the 3 different treatments is summarized in Table 1. Winds during application ranged from 0 to 11 mph and averaged 4.3 mph. Other meteorological conditions recorded during application are summarized in Table 2. The precipitation recorded by a temporary weather station established in the immediate area of the study from Sept. 14, through Oct. 25, 2004 is shown in Figure 1. The daily precipitation and mean temperatures as well as averages from 1971-2000 recorded at the Stephan weather station located ca. 12 miles east of the study area are shown in Figure 2. Cage Evaluations- The Eco Bran (wheat bran, Peacock Industries Inc.), Sevin 2% and 5% Bait (apple pumice “crumbles”, Wilbur-Ellis) and Tast-E-Bait (food waste, Advanced Organic Inc.) formulations of bait containing 2% and 5% carbaryl were applied to rangeland in cages. Additionally, treatments consisting of fine particles of 2% and 5% Eco Bran obtained by sifting material through an 18 mesh sieve were also applied. All formulations and concentrations were applied at the rate of 10 lbs/acre. An additional treatment of 2% Eco Bait was applied at 2 lb/acre. Ten cages (two gallon bottomless buckets modified with screen sides and top) (Fig. 3) were established on rangeland for each treatment. Ten additional cages were established on untreated vegetation as controls for comparison. This design, including the untreated control, therefore used a total of 110 cages. The appropriate amount of bait was pre-weighed in the laboratory and introduced into each cage separately in the field. Each cage was stocked with five, second instar stage Pardalophora haldemani grasshoppers collected from a nearby untreated area. Grasshoppers were introduced into cages immediately after bait was applied. After exposure to bait treatments, grasshoppers in all cages, including untreated cages, were monitored daily for 7 days for mortality, (Fig. 4.). The study occurred from Sept. 17–24, 2004. A study of the identical treatments was similarly conducted (Sept. 18-24, 2004) using 4.5 inch diameter x 3 inch high squat paper cups with screen tops as cages (Fig. 3) located in the USDA, APHIS, PPQ laboratory in Pierre, South Dakota. Data for both types of studies were analyzed as five replications per treatment with 10 observations per replicate. Field Sampling Methods Two methods, sweep net and ring delimited counts, of estimating grasshopper densities were employed. Both methods are equivalent in estimating densities at young instar ages (Foster and Reuter, 1996).The same person from the Pierre South Dakota, USDA, APHIS, PPQ office conducted all of the grasshopper density estimates using one or the other of the two methods at each of 6 evenly distributed predetermined sampling sites within each plot and at 6 additional untreated sites evenly distributed in untreated rangeland located adjacent to and surrounding the treated plots. The sweep method was
7

employed when less time was available to make density estimates. Grasshopper populations in treated and untreated plots were counted and sampled at 0 to 3 days before treatment and as weather and schedules permitted at 3, 8, 14, 21, days after treatment in 2004 and 193 and 197 days after treatment in 2005 (malathion); 7, 15, 21, 28 and 38 days after treatment in 2004 and 192 and 196 days after treatment in 2005 (diflubenzuron); and 8, 14, 21 and 31 days after treatment in 2004 and 185 and 189 days after treatment in 2005 (carbaryl). Pretreatment counts were conducted on Sept. 14 and Sept 24, 2004. Post treatment counts for one or all treatments were conducted on Sept. 19, 24, 30, Oct 2, 7, 8, 15, 25, of 2004 and on March 28 and April 1 of 2005. Untreated control plots were counted and sampled on any day a treated plot was monitored.
The majority of the data was collected using the sweep net method employed by federal and state personnel in North Dakota and South Dakota. With this method, 40 sweeps, each 180o in length, were conducted with a 15 inch diameter standard sweep net on a 3 foot long handle. Each sweep was performed by dragging the net firmly along the ground with a rapid motion while walking at a moderate pace through the site area. After collection samples were placed in paper bags, labeled and cold stored in the laboratory until they could be sorted, identified and counted. The total number of grasshoppers collected in each sample was divided by 10 to arrive at the number of grasshoppers per square yard This technique resulted from studies conducted in the late 1970’s by Gregory Mulkhern, North Dakota State University (Keith Winks, USDA-APHIS-PPQ retired - personal communication Aug. 25, 2005) and (Kantack, 1990). Alternately, grasshopper density evaluations generally followed protocols established by Foster and Reuter, 1996. Grasshopper densities were determined by counting grasshoppers in (40) 0.1 m2 rings arranged in an approximate 100 yard diameter circle at each sampling site. Rings at each site were separated from adjacent rings by ca, 5 yards. The four square meter count was converted to grasshoppers per square yard.
Analysis Field densities were recorded as grasshoppers /yd2. Post treatment data were further expressed as percent mortality based on pretreatment counts in the same plot. Percentage mortality data were converted to rank data (Conover and Iman, 1981) for analysis. Because post treatment evaluation intervals were not always equal, the data were grouped into intervals of 3 days after treatment (DAT), 7-8 DAT, 14-15 DAT, 21 DAT, 28-31 DAT, 398 DAT and 189-197 DAT for convenience and analyses. An analysis of variance was performed on ranked data with the Tukey Multiple Comparison Test used to separate means. Additional analyses were conducted because great differences in pretreatment densities between sites within the same plot and between plots confounded traditional analysis and comparisons. To alleviate this problem a theoretical population, equal to the pretreatment density mean of all plots, including the untreated check, (66 grasshoppers /yd2) was generated as a starting point for each site. These density values were then reduced by the actual percentage reduction values determined at each site at each visitation. Theoretical grasshopper density curves were then generated and graphed through time for each site. The area under the curve, termed grasshopper feeding days (Hewitt and Onsager 1982)
8

was then calculated for each site for a given length of time. When the feeding days were totaled for a given interval they were termed accumulated grasshopper feeding days. Grasshopper feeding days were considered an index of damage with the highest accumulated feeding days an indication of the greatest damage. The accumulated grasshopper feeding days values calculated for each site were then analyzed as described earlier at 1, 2 and 3 weeks post treatment in the fall and at 189 days post treatment after two complete sets of data had been collected from each plot the following spring. Field and laboratory cage data were recorded as percentage mortality, converted to rank data, and analyzed using a one-way analysis of variance with The Tukey Multiple Comparison Test to separate means. All analyses were performed with Systat 6.1 for Windows
Results and Discussion
Field Evaluation of Sprays. Pretreatment densities from individual sites ranged from 14 to 231 grasshoppers /yd2 in the treated plots and from 1 to 117 grasshoppers /yd2 in the untreated plots. The mean densities were 120, 80 and 20 grasshoppers /yd2 in the malathion, carbaryl and diflubenzuron plots respectively and were 43 grasshoppers/yd2 in the untreated sites. At the time of treatments the populations were composed predominately of first instars (13 and 32%), second instars (48 and 61%), third instars (13 and 8%) and fourth instars (5 and 6%). The total mean instar age was 1.96 on Sept. 14, 2004 and 2.22 on Sept. 24, 2004. The two most dominate species were Pardalophora haldemani 86% and 90 % on Sept. 14, 2004 and Sept. 24, 2004 respectively and Eritettix simplex 13 % and 9 % on Sept. 14, 2004 and Sept. 24, 2004 respectively. Sixteen different species were found in the study area at the time of treatment. The relative abundance and associated age of all species in pretreatment samples for both dates are shown in Table 3. Abundance and age data from post treatment samples will be available in the future upon completion of all sample processing and identification. Generally, populations in all four plots (treated and untreated) started at substantially different levels. The mean densities for the entire study are shown in Fig 5. In the season of treatment it was evident that populations demonstrated sharper initial declines in both malathion and carbaryl treated plots. Diflubenzuron and untreated populations declined similarly in the season of treatment. By mid to late Oct. any differences between population levels was indiscernible due to the onset of winter. In the spring of 2005 populations in all plots showed substantial declines. Mean percentage grasshopper reductions in treated and untreated plots are shown in Table 4. At 3 days after treatment the malathion treated population demonstrated a substantial 58 percent reduction while the untreated population demonstrated 37 percent reduction. At 7-8 days after treatment carbaryl and malathion treated populations were reduced significantly more than diflubenzuron treated populations. In fact, populations
9

had actually increased in all of the diflubenzuron sites and four of the untreated sites, an indication of additional hatch. At 14-15 days after treatment, Carbaryl treated populations were reduced significantly more than diflubenzuron treated populations but were statistically equivalent to malathion treated populations. At 21 days after treatment all three treatments demonstrated statistically equivalent reductions. At 28 to 38 days after treatment (Oct.15-25) populations in all plots including the untreated control, demonstrated reductions in excess of 84 percent. This was not unexpected. Many non overwintering species were disappearing from old age and exposure and overwintering species were difficult to see or capture due to the onset of winter and colder temperatures. Data (ring delimited) was collected the following spring on April 1, 2005, when mean populations were 14.1, 11.75, 12.6 and 12.8 grasshoppers / yd2 in the malathion, diflubenzuron, carbaryl and untreated plots respectively. The population reductions were statistically equivalent in malathion and carbaryl treated plots but, greater than reductions in the diflubenzuron plot. Untreated populations revealed an 88 % mean reduction, Table 4. Compared to ring delimited data, sweep net collected data which was collected 4 days earlier on March 28, 2005 showed somewhat different mean populations of 24.9, 5.95, 15.25 and 13.7 grasshoppers / yd2 in untreated plots respectively. These differences were inexplicable since both techniques provide equivalent estimates with young grasshoppers but show underestimation of population levels with the sweep net technique when grasshoppers are older (Foster et al. 1989). The great differences in pre-treatment densities within the same plot and between plots confounded comparisons and analyses with the field collected data. In an additional evaluation, considered more robust by the authors, the percentage reduction values observed in each treatment were applied to a theoretical equivalent pretreatment density for all plots. Density curves through time were generated using the field observed reduction values (Figure 7) and the area under the curves, termed accumulated grasshopper feeding days (AGFD), were totaled for analyses. Grasshopper feeding days (GFDs) are an indication of feeding and therefore forage damage with higher numbers indicating the greatest damage. Modeled data at one week after treatment indicated there was no significant difference in AGFD between untreated populations and those treated with diflubenzuron, Table 5. This was not unexpected. Little if any molting was expected within the interval. Cool temperatures had not only reduced grasshopper activity and the feeding required to pick-up the toxicants, but more importantly had reduced the speed of grasshopper development required for molting to occur. Diflubenzuron requires molting to the next development stage before any mortality and reduction in population would occur. There was no significant difference in GFDs between malathion and carbaryl treated plots but both plots demonstrated significantly lower GFDs compared to diflubenzuron and untreated plots, The reduced GFD indicated some level of control had occurred in the malathion and carbaryl plots. Some or all of this mortality may be attributed to contact activity with the insecticides. Even though most of the mortality that results from these treatments is due to feeding and ingestion of the toxicant, which could have been suppressed in this study, some of the mortality associated with these treatments is a result of direct contact.
10

Analyses at two and three weeks after treatment showed similar results to that seen at one week after treatment. Again this was not unexpected as temperature dependant development was effected by the cool temperatures, Fig 2. However, from late October, 2004 through Late April, 2005, average temperatures in the study were higher than average years. From Oct. 15, 2004 through April 29, 2005 daily mean temperatures averaged 32.1o F in this study compared to the historic average daily mean of 26.6o F (Fig. 2).This suggests that in most years grasshopper development would have been even slower than seen in this study. When viewed from treatment day (Sept. 16 – 24, 2004) to early spring (April 1, 2005), no significant differences were detected in GFD accumulated for 185 and 189 days between sites treated with Fyfanon ULV and Dimilin or between those sites and the untreated sites, Table 5. However, there were significantly lower AGFD in the plot treated with carbaryl compared to the other plots. Compared to the untreated plot, AGFD increased 20% and 26% in the Fyfanon ULV and Dimilin treated plots respectively. The AGFD in the Sevin treated plots declined 50% compared to the untreated plot/population. Some hatch was still occurring after treatment. This recruitment in the population was evidenced by increasing percentages of first instars found in the sweep samples. Some of the population increases in treated plots could have been offset as long as any treatment residues were active. Residual activity is longest with diflubenzuron, followed by carbaryl and the shortest with malathion. Not withstanding, some level of mortality in malathion and particularly carbaryl plots occurred. GFD were increasing (between 7 and 21 days) with the least increase in accumulated GFD occurring in the carbaryl treated plot, Table 5. Carbaryl treatments appeared to have greater effect than malathion as evidenced by the slowest growing GFD and an increasing total percentage reduction, Table 5. Total percentage reduction in malathion sites remained relatively stable at 61, 45 and 56 % reduction for 1, 2 and 3 weeks after treatment compared to respective reductions in carbaryl sites of 58, 64 and 89 %. This later additional mortality seen in carbaryl sites was not unexpected since carbaryl has much longer residual activity than malathion. Through the spray study, malathion produced only a short term reduction, carbaryl produced both a short and long term reduction and Dimilin produced neither a short or long term reduction while the untreated populations declined substantially on their own. Lab Cage Evaluation of Baits. At three days after treatment, all treatments resulted in significantly higher mortality than occurred in untreated populations. All of the 5% concentrations of bait produced statistically equivalent mortality. All of the 2% concentrations of bait produced statistically equivalent mortality at the 10 lb application rate. The 5% concentration showed no advantage over the 2% concentration in any of the standard formulations of Eco Bait, Crumbles or Tast E Bait at the 10 lb application rate. However, with fine particles of Ecobait, significantly higher mortality resulted from the 5% concentration when compared to 2%. The addition of molasses to the 2% Tast E Bait produced no
11

significant increase in mortality compared to the same bait without molasses. The 2 lb/ac application rate of 2% Ecobait resulted in significantly lower mortality than all of the other treatments. At 6 days after treatment all baits applied at 10 lbs/acre produced equivalent mortality that was significantly higher than in untreated populations. Untreated populations declined only 2% during the 6 days of the study. Results of analyses conducted at other post treatment intervals are shown in Table 6. Field Cage Evaluation of Baits At three days after treatment all baits resulted in significantly higher mortality than occurred in the untreated populations, Table 7. Results in field cages were similar to those from laboratory cages with three exceptions. (1) The addition of molasses to 2% Tast E Bait resulted in significantly lower mortality compared to the same bait without molasses. It was obvious in the field that molasses resulted in reduced performance of the bait. In the presence of available native foods, the addition of molasses may have rendered the bait less appealing and therefore produced less consumption and mortality compared to the laboratory. In the laboratory study no alternative native food was available. (2) There was no significant difference between mortalities resulting from fine particles of 2% and 5% Ecobait. However, fine particles may be more easily found in the laboratory than the field. Compared to larger particles, fine particles of bait may contain higher concentration of carbaryl that could explain higher levels of mortality in the laboratory. (3) There was no statistical advantage resulting from the 10 lb/acre application rates of any of the baits compared to the 2 lb application rate of 2% Ecobait. All baits produced equivalent mortality except 2% Tast E Bait and molasses. However, in the laboratory the 2 lb application rate resulted in significantly less mortality than 10 lb application rates. The difference between field and laboratory mortality resulting from the 2 lb application rate is unexplainable. However, it is noteworthy that 4 of the 5 Eco-bait formulations resulted in numerically higher mortality in the field compared to the lab. In field cages at 2, 3 and 7 days after treatment, all baits produced equivalent mortality that was significantly higher than in untreated populations. Untreated populations declined only 2% during the 7 days of the study. Results of analyses conducted at other post treatment intervals are shown in Table 7. Feeding activity that is directly related to temperature was considered usual for this time of year because the mean of the daily average temperatures (60.4o F) during the study (Sept. 17-24, 2004) were similar to the mean of daily averages (58.4oF) for 1971-2000, (Fig.2). In both laboratory and field studies, no real advantages were seen with the higher concentrations of carbaryl. In the field there was clearly no advantage with a higher application rate. Compared to 5 % concentrations of carbaryl, the 2 % formulations reduce the cost of carbaryl required by 60 %. Reducing the application rate from 10 to 2 lbs / not only reduces the cost of carrier used in the bait by 80 %, but results in less physical material required per acre which translates into a higher acre capacity in the hoppers of spray aircraft, and yields reductions in application costs. While these reductions are very substantial in terms of chemical, carrier and application costs savings, such reductions are also highly desired relative to environmental and non-target concerns.
12

Conclusions
Grasshopper feeding and development as well as loss of forage were minimal because of cool weather during this study. Cool temperatures affected the consumption of sprayed vegetation and the developmental progression from one instar to the next and therefore the mortality expected from all liquid sprays. For the first three weeks after treatment temperatures were similar to historical averages for the area. However, from mid Oct, 2004 through late April, 2005 temperatures during our study averaged 5.5o F higher than usual. In years with temperatures closer to average (colder), feeding and development would most likely be even slower, thus further impacting mortality expected from sprays. When sprays were applied in September, carbaryl clearly performed better than the other treatments. However, grasshopper densities declined naturally and were nearly the same in treated and untreated plots the following spring. Compared to untreated populations, malathion produced only a short term reduction, carbaryl produced both a short and long term reduction and diflubenzuron produced neither a short or long tern reduction. While significant, these reductions were not high enough to be considered effective or economical. Additionally, even though substantial reductions had occurred naturally, grasshopper densities remained sufficiently high to cause substantial damage. Unquestionably, baits produced quicker and much more substantial mortality than sprays and are clearly the choice if fall treatments are required. However, the question remains, regarding the value of such treatments. During the period from fall to spring, little, grasshopper development and forage destruction is occurring, and in normal years grasshopper development would be, if anything, even slower than seen in this study. Our data indicate there is little advantage for treating in the fall rather that waiting until the following spring. If late season treatments are anticipated, several issues/areas should be considered. (1) Weather conditions that impact application. However, if early spring is the option, it may be no better than late fall for optimal application parameters. (2) The potential for substantial natural mortality to occur over the winter. (3) Minimal grasshopper development as well as minimal loss of forage between late fall and next spring (4) Seasonal meteorological conditions following treatment through the required time frame for optimal results of the selected treatments. The major component of mortality of these treatments is attributed to ingestion. Grasshopper feeding is related to temperature as is development. Sufficiently high temperatures must exist to stimulate feeding on vegetation containing the toxicants and also to promote development to the stage that molting will occur which results in mortality when the insect growth regulator, diflubenzuron is used. (5) Economics. Sprays are generally less expensive than solid baits. Traditional sprays of carbaryl and diflubenzuron are similar in costs but more expensive than the traditional spray of malathion. Conversely, if a Reduced Agent Area Treatment strategy (RAATs) is used, malathion is more expensive than carbaryl or diflubenzuron (Foster et al. 2000, 2004). If solid baits are selected as the treatment, the costs of both materials and application must be considered. Material costs are less a
13

consideration when low concentration and rate is used (2 lb/ac of 2%), but in any case, application costs are substantially higher with baits compared to sprays because of the bulk density and the capacity in terms of “acres of material” that the hopper of an aircraft can accommodate. Our data suggest spring treatments of this overwintering species would most likely be more economical and should be considered in lieu of fall treatments. Treatments in South Dakota between late April and late May should allow time for any additional natural mortality to occur and would still present a window for treating immature grasshoppers before any potential wide scale damage, movement or egg laying could occur. Additionally, any of the available treatment options could be considered during this time period.
Acknowledgements
The authors thank the Crow Creek Indian Tribe and Todd Cowen for allowing the study to be conducted on their land. Additionally, we thank the USDA APHIS PPQ Western Region for funding the study separately and in addition to normal funding mechanisms and schedules. Special thanks are extended to retired APHIS employees Dick Jackson, Bruce Thornley, Tom Crowe, Dan Kail and Keith Winks and current APHIS employees Bruce Helbig, Steve Johnson and Greg Abbott for providing historical information on outbreaks of overwintering species of grasshoppers. Thanks are also extended to Dennis Todey, South Dakota Extension / State Climatologist for providing recent and historical weather data.
References Cited Conover, W.J. and R.L. Iman. 1981. Rank Transformations as a Bridge Between Parametic and Nonparametric Statistics. The American Statistician. 35: (3) 124-129. Foster, R.N. and K.C. Reuter. 1996-1999. Evaluation of rangeland grasshopper controls: A general protocol for efficacy studies of insecticides applied from the air. In: Cunningham, G.L., and M.W. Sampson tech cords. Grasshopper integrated pest management user handbook. Tech Bull. 1809. Washington, DC: U.S. Dept. of Agriculture, Animal and Plant Health Inspection Service: II.2-1—II.2-7.
Foster, R.N., K. Chris Reuter, K. Fridley, D. Kurtenbach, R. Flakus, R. Bohls, B. Radsick, J.B. Helbig, A. Wagner and L.J. Jech. 2000,2004. Field and economic evaluations of operational scale Reduced Agent and Reduced Area Treatments (RAATs) for management of grasshoppers on South Dakota rangeland, 1997-1999. Report, 40 pp. 2000. Also in: Branson, D.H. and Bethany Redlin (eds). 2004. Grasshoppers: Their Biology, Identification and Management. 2th edition. CD-ROM. US Department of Agriculture, Agricultural research Service.
14

Foster, R.N., W.J. Cushing, M. Smith, T. Battaglia and S. Battaglia. 1989. Standardization of rangeland grasshopper survey methodology: a comparison of visual, ring and sweep net techniques in estimating grasshopper populations on rangeland. In: USDA-APHIS-PPQ Grasshopper Integrated Pest Management Project. Annual Report (Fiscal Year 1989). pp 165-172.
Hewitt, G B. and J. A. Onsager. 1982. A Method for Forcasting Potential Losses from Grasshopper Feeding on Northern Mixed Prairie Forages. Journal of Range Management. 35: (1) 53-57.
Kantack, Benjamin H. 1990. Grasshopper Control Economics Thresholds. Cooperative Extension Service, South Dakota State University, US Department of Agriculture, Brookings , South Dakota. FS 859. Lloyd, J. E., R. E. Pfadt and M. Ali. 1974. Manner of Pickup of ULV Carbaryl by Aulolcara elliotti from Aerially Sprayed Rangeland. Jour. Econ. Entomol. 67:596-597.
Mulkhern, G. B., D. R. Toczek and M. A. Brusvan. 1964. Biology and ecology of North Dakota grasshoppers II. Food habits and preferences of grasshoppers associated with the sand hills prairie. Researsch Report No. 11. North Dakota Agricultural Experiment Station, Fargo, North Dakota. 59 pp.
Mulkhern, G. B., K. E. Pruess, H. Knutson, A. F. Hagen, J. B. Campbell and J. D. Lambley. 1969. Food habits and preferences of grasslands grasshoppers of the North Central Great Plains. Bulletin No. 481. Agricultural Experimental Station, North Dakota State University, Fargo, North Dakota 58102. 32 pp.
Parker, J. R., and R. V. Connin. 1964. Grasshoppers: Their habits and damage. U.S. Dept. of Agriculture . Info. Bull. 287: 1-28.
Pfadt, R. E. 2002. Field Guide to Common Western Grasshoppers. Third Ed. Redshanked Grasshopper Xnathippus corallipes (Haldeman).Wyoming Agricultural Experiment Station. Bulletin 912. University of Wyoming Pfadt, R. E., J. E. Lloyd, M. Ali, and G. Sharafi. 1970. Manner of pickup of ULV malathion by grasshoppers from aerially sprayed rangeland. Journal of Economic Entomology. 63:1210-1214. SPSS Inc. 1997. SYSTAT for Windows: Statistics, SPSS Inc., Chicago, Illinois.
15

Table 1. Summary of treatments and calibration parameters near Pierre, SD 2004. Nozzle Treatment Diluent Total Nozzle Tip Screen Pressure Aircraft Swath (lb AI/ac) fl oz/ac fl oz/ac No. Size Size psi Speed Width carbaryl 16 H2O 32 18 8003 50 40 120 100 (0.50) diflubenzuron 10 oil + 31 18 8003 50 40 125 100 (0.015625) 20 H2O malathion none 8 8 8002 50 40 120 100 (0.61) Table 2. Meteorological conditions recorded during aerial application of treatments in the grasshopper study near Pierre, S. D. 2004. Temperatures °F Time AM Ground Air Wind (mph) Treatment Date Start Stop Start Stop Start Stop Pilot Start Stop malathion 9/16 8:17 8:56 51 53 54 55 50 6-8 SE 7-11SE diflubenzuron 9/17 7:00 7:40 49 55 58 57 57 0-3 NE 2-6 NE carbaryl 9/24 7:17 8:00 38 40 43 51 49 2-3 NE 1-2 NE
16
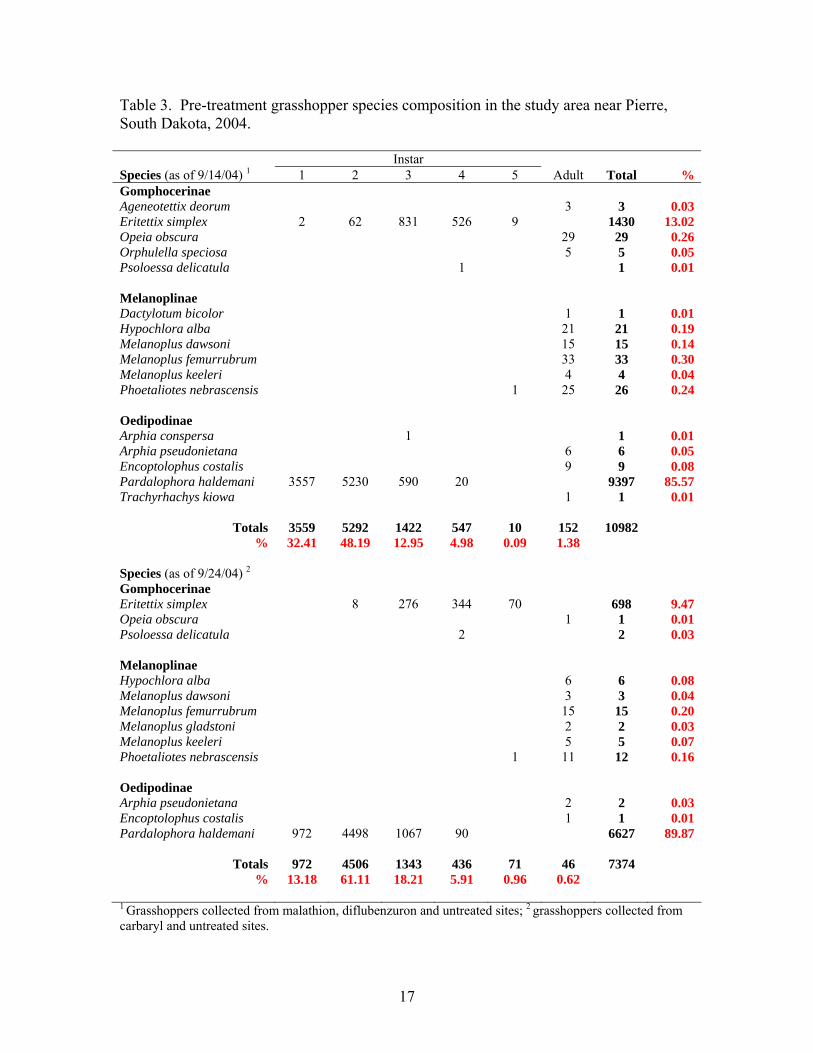
Table 3. Pre-treatment grasshopper species composition in the study area near Pierre, South Dakota, 2004. Instar Species (as of 9/14/04) 1 1 2 3 4 5 Adult Total % Gomphocerinae Ageneotettix deorum 3 3 0.03 Eritettix simplex 2 62 831 526 9 1430 13.02 Opeia obscura 29 29 0.26 Orphulella speciosa 5 5 0.05 Psoloessa delicatula 1 1 0.01 Melanoplinae Dactylotum bicolor 1 1 0.01 Hypochlora alba 21 21 0.19 Melanoplus dawsoni 15 15 0.14 Melanoplus femurrubrum 33 33 0.30 Melanoplus keeleri 4 4 0.04 Phoetaliotes nebrascensis 1 25 26 0.24 Oedipodinae Arphia conspersa 1 1 0.01 Arphia pseudonietana 6 6 0.05 Encoptolophus costalis 9 9 0.08 Pardalophora haldemani 3557 5230 590 20 9397 85.57 Trachyrhachys kiowa 1 1 0.01
Totals 3559 5292 1422 547 10 152 10982 % 32.41 48.19 12.95 4.98 0.09 1.38
Species (as of 9/24/04) 2 Gomphocerinae Eritettix simplex 8 276 344 70 698 9.47 Opeia obscura 1 1 0.01 Psoloessa delicatula 2 2 0.03 Melanoplinae Hypochlora alba 6 6 0.08 Melanoplus dawsoni 3 3 0.04 Melanoplus femurrubrum 15 15 0.20 Melanoplus gladstoni 2 2 0.03 Melanoplus keeleri 5 5 0.07 Phoetaliotes nebrascensis 1 11 12 0.16 Oedipodinae Arphia pseudonietana 2 2 0.03 Encoptolophus costalis 1 1 0.01 Pardalophora haldemani 972 4498 1067 90 6627 89.87
Totals 972 4506 1343 436 71 46 7374
% 13.18 61.11 18.21 5.91 0.96 0.62
1 Grasshoppers collected from malathion, diflubenzuron and untreated sites; 2 grasshoppers collected from carbaryl and untreated sites.
17

Table 4. Mean percentage reduction of grasshoppers in treated and untreated plots near Pierre, South Dakota – 2004, 2005. % reduction – DAT 1
Treatment 3d 7-8d 14-15d 21d 28-31d 38d 189-197d malathion 58 61 a 45 ab 56 a 95 a diflubenzuron -87 b 0 b 33 a 85 a 84 78 b carbaryl 58 a 64 a 89 a 97 a 94 a untreated 37 16 28 52 91 97 88 1 A one-way analysis of variance with a Tukey multiple comparison procedure determined statistical differences (P ≤ 0.05). Means in a column followed by the same letter are not significantly different. Table 5. Mean modeled accumulated grasshopper feeding days and (total grasshopper percentage reduction). Mean grasshopper feeding days (total reduction) 1
Treatment 7-8 d 14-15 d 21 d 185 d 189 d Fyfanon ULV 475 a (61) 676 A (45) 912 a (56) 4756 b (79) 4590 b (94) Dimilin 2L 960 b (-87) 1720 B (0) 2039 b (33) 4519 b (72) 4815 b (77) Sevin XLR 473 a (58) 631 A (64) 740 a (89) 1886 a (81) 1920 a (93) Untreated 927 b (16) 1289 B (28) 1645 b (52) 3757 b (66) 3822 b (84) 1 Rank transformations of grasshopper feeding days data were evaluated by analysis of variance with a Tukey multiple comparison method. Means in a column followed by the same letter are not significantly different (P ≤ 0.05).
18

Table 6. Efficacy of selected carbaryl baits against second instar Pardalophora haldemani grasshoppers in laboratory cages – Pierre, SD 2004. Mean % mortality 1
Treatment 1d 2d 3d 4d 5d 6d Crumbles 5% 64 ab 76 a 82 ab 88 ab 90 ab 90 a Crumbles 2% 46 bc 66 ab 70 b 74 b 76 b 80 a Tast-E-Bait 5% 74 ab 80 a 82 ab 92 ab 94 ab 94 a Tast-E-Bait 2% 66 ab 78 a 82 ab 86 ab 92 ab 94 a Tast-E-Bait 2% mol 52 abc 62 ab 68 b 76 b 78 ab 80 a Eco-bait 5% fine 76 a 86 a 94 a 98 a 98 a 98 a Eco-bait 2% fine 52 abc 60 ab 68 b 80 ab 82 ab 82 a Eco-bait 5% 70 ab 70 ab 76 ab 78 b 88 ab 94 a Eco-bait 2% 62 ab 80 a 84 ab 86 ab 88 ab 88 a Eco-bait 2%/2lb 30 c 40 b 42 c 44 c 48 c 54 b Untreated 0 d 0 c 0 d 0 d 0 d 2 c 1 A one-way analysis of variance with a Tukey multiple comparison procedure determined statistical differences (P ≤ 0.05). Means in a column followed by the same letter are not significantly different.
19

Table 7. Efficacy of selected carbaryl baits against second instar Pardalophora haldemani grasshoppers in field cages – Pierre, SD 2004. Mean % mortality Treatment 1d 2d 3d 7d Crumbles 5% 50 b 76 a 80 a 86 a Crumbles 2% 50 b 70 ab 74 ab 80 ab Tast-E-Bait 5% 60 ab 82 a 84 a 88 a Tast-E-Bait 2% 66 ab 78 a 88 a 88 a Tast-E-Bait 2% mol 22 c 36 b 52 b 56 b Eco-bait 5% fine 66 ab 66 ab 74 ab 74 ab Eco-bait 2% fine 60 ab 74 a 82 a 86 a Eco-bait 5% 76 a 94 a 96 a 96 a Eco-bait 2% 66 ab 84 a 90 a 90 a Eco-bait 2%/2lb 58 ab 74 a 86 a 90 a Untreated 0 c 0 c 2 c 2 c 1 A one-way analysis of variance with a Tukey multiple comparison procedure determined statistical differences (P ≤ 0.05). Means in a column followed by the same letter are not significantly different.
20

0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
9/14 9/17 9/20 9/23 9/26 9/29 10/2 10/5 10/8 10/11 10/14 10/17 10/20 10/23 10/26
2004
Prec
ipita
tion
(inch
es)*
x x x x x x x x x x x x x x x x
*Bar values represent total precipitation recorded since the previous reading (X).
Figure 1. Specific amounts of precipitation recorded in the study area.
-20-10
0102030405060708090
9/1 9/21 10/11 10/31 11/20 12/10 12/30 1/19 2/8 2/28 3/20 4/9 4/29
2004/2005
Tem
pera
ture
o F
00.10.20.30.40.50.60.70.80.91
Prec
ipita
tion
(inch
es)
Daily mean °F Historic daily mean °F Daily precipitation Historic daily mean precipitation
Figure 2. Daily means and historical daily means for temperature and precipitation observed at the Stephan climatological recording station in Hyde County, South Dakota.
21

Figure 3. Field and laboratory cages used to evaluate selected baits for efficacy against Pardalophora haldemani.
Figure 4. Field cages established for evaluating mortality of Pardalophora haldemani.
22
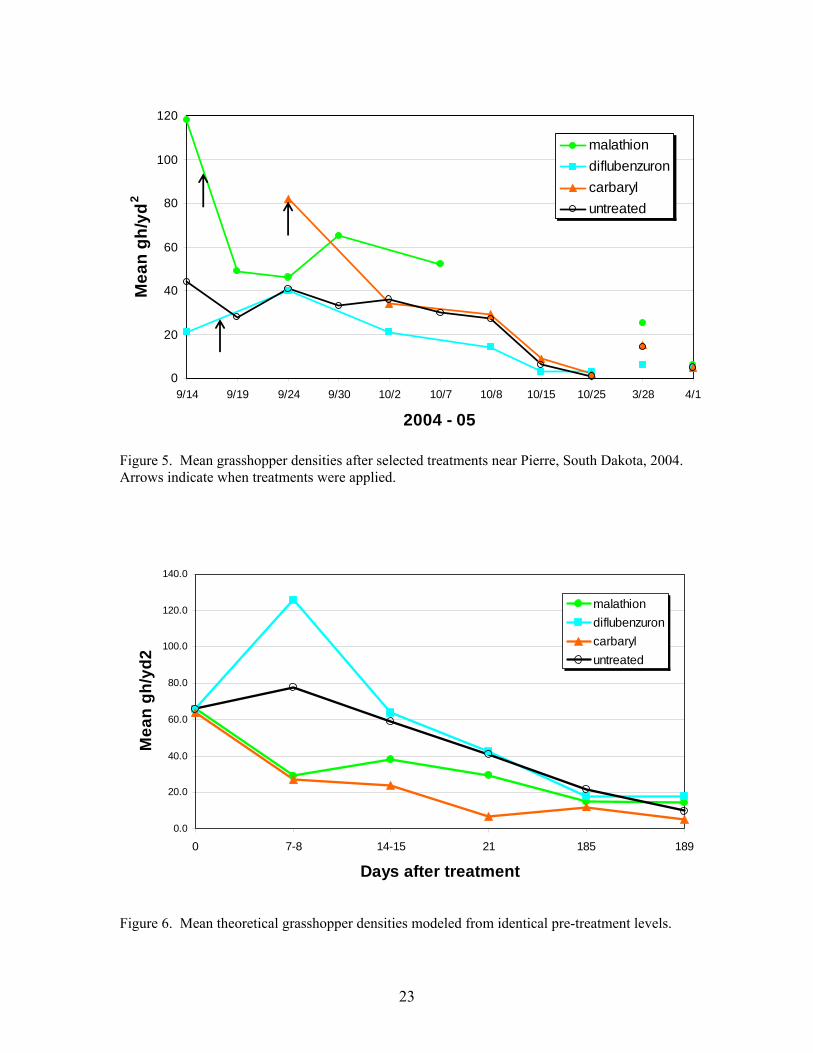
0
20
40
60
80
100
120
9/14 9/19 9/24 9/30 10/2 10/7 10/8 10/15 10/25 3/28 4/1
2004 - 05
Mea
n gh
/yd2
malathiondiflubenzuroncarbaryluntreated
Figure 5. Mean grasshopper densities after selected treatments near Pierre, South Dakota, 2004. Arrows indicate when treatments were applied.
0.0
20.0
40.0
60.0
80.0
100.0
120.0
140.0
0 7-8 14-15 21 185 189
Days after treatment
Mea
n gh
/yd2
malathiondiflubenzuroncarbaryluntreated
Figure 6. Mean theoretical grasshopper densities modeled from identical pre-treatment levels.
23

Jan. 2006
Initial evaluations of apple pumice and food waste baits against rangeland grasshopper species not susceptible to the wheat bran bait standard
R. Nelson Foster, K. Chris Reuter, and Lonnie R. Black
Organizations
USDA/APHIS/PPQ/CPHST/Decision Support and Pest Management Laboratory, Phoenix, Arizona
Cooperators
Bruce Helbig, USDA/APHIS/PPQ, Pierre, South Dakota Mark Tubbs, Edgemont, South Dakota
Neil Wagner, Peacock Industries, Hague, Saskatchewan, Canada Gary Saxton, Wilbur-Ellis, Fresno, California
David I. B. Vander Hooven, Advanced Organics Inc., Maumee Ohio
Abstract
A study conducted on southwestern South Dakota rangeland with apple pumice and food waste baits against grasshopper species categorized to be nonsusceptible to the standard wheat bran bait demonstrated minimal if any success against separate caged populations of Amphitornus coloradus (Thomas), Cordillacris occipitalis (Thomas) and Trachyrhachys kiowa (Thomas). Treatments of the three baits on C. occipitalis showed no consistent significant mortality resulted with any of the three baits when compared to untreated populations. Treatment with the food waste bait produced no significant mortality on A. Coloradus when compared with untreated populations. Treatment with food waste bait produced significantly higher mortality on T. kiowa than occurred in untreated populations, 38% and 8% mortality respectively. However, even though one species demonstrated higher mortality than in controls, all three species would still be categorized as nonsusceptible to the alternative baits. This is consistent with responses expected with wheat bran baits.
1

Introduction
The level of acceptance by different rangeland grasshopper species to wheat bran bait containing carbaryl is well documented (Onsager et al. 1980 a, b; Quinn et al. 1989, 2000; Jech et al. 1993; Foster et al 1998, 1999). Some species are categorized as sensitive where control is expected to average about 70% with worst-case and best-case scenarios about 55% and 85% respectively. Some species are vulnerable where control is expected to average about 42% with worst-case and best-case scenarios about 12% and 72% respectively. Some species are categorized as non-susceptible where control is expected to average about 15% with worst-case and best-case scenarios about 0% and 30% respectively (Onsager et al. 1996-1999). The Mormon cricket, a long-horned grasshopper, is extremely susceptible to wheat bran bait containing carbaryl (Foster et al. 1979). During the last three years studies with alternative baits based on apple pumice and food waste (combined bakery, snack, cereal and confectionary waste) have shown activity equal to the standard wheat bran bait against Mormon cricket. (Foster et al. 2003, 2004). These alternative baits may provide an opportunity to expand the usefulness of baits against grasshoppers. Species that are moderately or nonsusceptible to wheat bran bait may be more susceptible to alternative baits. However, little information exists on the level of susceptibility of rangeland grasshopper species to these baits. The following study was conducted to develop data on the level of susceptibility that could be expected when alternative apple pumice or food waste based carbaryl baits are used against rangeland grasshoppers categorized as non-susceptible to the current wheat bran based bait standard.
Materials and Methods
The study was conducted in Fall River County of southwestern South Dakota ca. 12 miles west and 9 miles north of the town of Edgemont on the Mark Tubbs ranch during the period of June 30 through July 6, 2005. The location was selected because of the diversity in grasshopper species, density of grasshoppers, history of grasshoppers in the area, and availability of a rangeland pasture without livestock for the period of time required in the study. Three species of rangeland grasshoppers categorized as non-susceptible to carbaryl wheat bran bait, were selected for separate cage studies against the alternative baits. The baits used in the studies were Sevin Bait (Wilbur-Ellis), Tast-E-Bait (Advanced Organics) and the standard, Eco Bran (Peacock Industries), formulations each containing 2% carbaryl. Cages were two gallon bottomless buckets modified with screen sides and top Figs 1-3. All baits were applied within cages at the rate of 10 lbs/ acre. The appropriate amount of bait was pre-weighed in the laboratory and introduced into each cage separately in the field. Grasshoppers were introduced into cages after bait was applied. In the first study Cordillacris occipitalis (Fig. 4), the most prevalent nonsusceptible species in the area, was exposed to the Eco Bran, Sevin Bait and Tast-E-Bait. Ten cages
2

were established for each formulation. Ten additional cages were established on nearby untreated vegetation as controls for comparison. Each cage was stocked with five, mostly adults and a few fifth instar stage Cordillacris occipitalis grasshoppers collected from an adjacent untreated area. Both treated and untreated cages were monitored daily for 6 days after exposure to treatments occurred In the second study Amphitornus coloradus (Fig. 5) was exposed to Tast-E-Bait in 10 cages. Ten additional cages were established on nearby untreated vegetation as controls for comparison. Each cage was stocked with five, mostly fifth instar and a few adult stage Amphitornus coloradus grasshoppers collected from an adjacent untreated area. The rarity of this species precluded direct comparison with Eco Bran and Sevin Bait. Both treated and untreated cages were monitored daily for 6 days after exposure to treatments occurred In the third study Trachyrhachys kiowa (Fig. 6) was exposed to Tast-E-Bait in 10 cages. Ten additional cages were established on nearby untreated vegetation as controls for comparison. Each cage was stocked with five, mostly fifth instar and a few adult stage Trachyrhachys kiowa grasshoppers collected from an adjacent untreated area. The rarity of this species also precluded direct comparison with Eco Bran and Sevin Bait. Both treated and untreated cages were monitored daily for 3 days after exposure to treatments occurred.
Results and Discussion
Cordillacris occipitalis feeds on a high variety of grasses (where it is not highly selective) and some sedges. It rarely consumes forbes or ground litter, although, it is known to have been attracted to a discarded apple core (Pfadt, 2002).Our study with C. occipitalis, showed there was no significant difference in mortality resulting from any of the three baits and the untreated populations, except at 5 days after treatment, where apple pumice bait produced significantly more mortality than occurred with the other baits (Table 1; Fig. 7). This generally nonsusceptible response is consistent with substantial data evaluated and reported by Onsager et al. 1996-1999. Our data suggest that this species should be considered as not susceptible to apple pumice and food waste based baits. Amphitornus coloradus feeds almost exclusively on grasses and sedges and rarely feeds on the ground (Pfadt, 2002). He also reports that field tests indicate only 4% of the population will feed on wheat bran. In our study with A. coloradus, the food waste bait did not produce mortality significantly greater than occurred in untreated populations. (Table 2; Fig. 8).The untreated populations in this study demonstrated much higher mortality than the untreated populations in any of the other studies, most likely indicating that the small cage area limited or precluded availability of critical and or preferred host food sources in the field. Our data suggest that this species should be considered nonsusceptible to food waste bait.
3

Trachyrhachys kiowa feeds exclusively on grasses and sedges with a great preference for blue gramma grass. Nymphs have never been observed eating plant litter and adults infrequently. Lab and field evidence indicate it does not feed on bran bait (Pfadt, 2002). In our study with T. kiowa, the food waste bait produced mortality at 2 and 3 days after treatment that was significantly (highly) different than occurred in the untreated populations (Table 3: Fig. 9). This suggests some potential for alternative baits on this species. However, the level of mortality even though highly significant, is at the upper range for the nonsusceptible category of species set by Onsager et al.1996-1999. Our data suggest that this species should be considered nonsusceptible to food waste bait. After adjustment for natural mortality at 3-6 days after treatment, the average mortality of the three species we studied (13.2 %) is consistent with the average (15 %) reported for the entire nonsusceptible category of rangeland grasshoppers reported by Onsager et al. 1996-1999.
Other studies on food waste bait against two grasshopper species highly susceptible to wheat bran bait, Ageneotettix deorum and Aulocara elliotti (Foster et al. 2003, 2004) indicate high levels of susceptibility that are consistent with the category for species sensitive to wheat bran reported by Onsager et al. The results of our study are not completely unexpected. All three species were categorized as nonsusceptible to wheat bran baits. The behavioral response that renders wheat bran unattractive apparently also renders the tested alternatives unattractive. The data does suggest that the categorization for wheat bran bait may hold true for species treated with apple pumice or food waste baits. However, additional study with species categorized as moderately susceptible (vulnerable) or highly susceptible (sensitive) to wheat bran bait should be conducted to insure more confidence in extrapolating wheat bran susceptibility to apple pumice and food waste baits.
Conclusions
C occipitalis showed no significant mortality response to any of the three baits. A. coloradus showed no significant mortality response to the food waste bait. Even though T. kiowa showed a significant mortality response to the food waste bait, the level was not sufficient to categorize the species above nonsusceptible. These studies suggest that susceptibility levels of grasshopper species to apple pumice and food waste baits may be extrapolated from previous wheat bran bait results. However, to insure equal acceptance of all three baits by individual species some additional testing of these alternative baits is still warranted on species categorized as moderately and highly susceptible to wheat bran bait.
4

Acknowledgements
The authors thank Neil Wagner (Peacock Industries), Gary Saxton (Wilbur-Ellis), and David I. B. Vander Hooven (Advanced Organic Inc.) for providing the material for testing. Thanks are extended to Mark Tubbs for providing the rangeland for the study and his continued cooperation since 1989.
References
Foster, R. N., C. H. Billingsley, R. T. Staten and D. J. Hamilton 1979. Field cage tests for concentrations of carbaryl in a bait and its application rates for control of Mormon Cricket. Jour. Econ. Entomol. 72: 295-297.
Foster, R. Nelson, K. Chris Reuter, Bruce Helbig and John Bradley. 2003. Field cage evaluation of Tast-E-Bait against the rangeland grasshopper Ageneotettix deorum. Report. USDA / APHIS / PPQ / CPHST / Decision Support and Pest Management Systems Laboratory. 4pp. Foster, R. N., K. C. Reuter, D. McNeal and R. McChesney, 2003. Report, Comparison of wheat bran and apple pumice baits for controlling Mormon cricket: Evaluating initial and secondary mortality with baits containing 2% and 5% carbaryl. Nov. 2003. 10 pp. Foster, R. Nelson, K. Chris Reuter, Lonnie Black, Bruce Helbig, and Lane Kozel. 2004. Field cage comparison of wheat bran, apple pumice and food waste baits containing carbaryl for controlling the rangeland grasshoppers, Ageneotettix deorum (Scudder) and Aulocara elliotti (Thomas). Report. USDA / APHIS / PPQ / Decision Support and Pest Management Systems Laboratory. 9 pp. Foster, R. N., K. C. Reuter, L. R. Black, D. McNeal, and R. McChesney. 2004.Report Comparison of wheat bran, apple pumice and a combined (bakery, snack, cereal and confectionary) food waste baits containing carbaryl for controlling Mormon crickets: Studies of bait longevity and primary and secondary mortality. USDA-APHIS-PPQ-CPHST-DS&PMSL. ??? pp. OCT 2004 Foster, R. N., M. A. Quinn, K. C. Reuter, D. Colletto, R. Houston, M. J. Puclik, A. Scott, and B. Radsick. 1999.Comparison of single and multiple applications of an insecticide bait for controlling grasshoppers (Orthoptera: Acrididae) on mixed-grass prairie. Jour. Kansas Entomol. Soc. 72: 181-189. Foster, R. N., M. A. Quinn, L. L. Herbaugh, D. D. Walgenbach, R. L. Bohls, K. C. Reuter, and T. J. Roland. 1998. Effect of multiple concentrations and rates of an insecticidal bait on grasshoppers (Orthoptera: Acrididae) of mixedgrass prairie. Jour. Kansas Entomol. Soc. 71: 1-10.
5

Jech, L. E., R. N. Foster, D. Coletto, D. D. Walgenbach, T. J. Roland, G. D. Rodriguez, R. Bohls, R. D. Houston, W. K. Meeks, R. L. Queener, C. L. Jackson, J. L. Dines, M. K. Puclik, and A. K. Scott. 1993. Field evaluations of diflubenzuron and carbaryl bran baits against grasshopper (Orthoptera: Acrididae) populations in South Dakota. Jour. Econ. Entomol. 86: 557-565. Onsager, J. A., J. E. Henry, and R. N. Foster. 1980. A model for predicting efficacy of carbaryl bait for control of rangeland grasshoppers. Jour. Econ. Entomol. 73: 726-729. Onsager, J. A., J.E. Henry, R. N. Foster, and R. T. Staten. 1980. Acceptance of wheat bran bait by species of rangeland grasshoppers. Jour. Econ. Entomol. 73: 548-551. Onsager, Jerome A., R. Nelson Foster and Larry Jech. 1996-1999. Bait Acceptance by Different Grasshopper Species and Instars. In: Cunningham, G.L., and M.W. Sampson tech cords. Grasshopper integrated pest management user handbook. Tech Bull. 1809. Washington, DC: U.S. Dept. of Agriculture, Animal and Plant Health Inspection Service: II.12-1—II.12-5. Pfadt, Robert E. 2002. A Field Guide To Common Western Grasshoppers. 3rd ed. Wyoming Agricultural Experiment Station. University of Wyoming. Bull. 912. 288 pp. Quinn, M. A., R. L. Kepner, D. D. Walgenbach, R. A. Bohls, and P. D. Pooler: R. N. Foster, K. C. Reuter, and J. L. Swain. 1989. Immediate and 2nd-year effects of insecticide spray and bait treatments on populations of rangeland grasshoppers. Canadian Entomologist. 121: 589-602. Quinn, M. A., R. N. Foster, W. J. Cushing, D. C. Hirsch, K. Winks, and K. C. Reuter. 2000. The North Dakota grasshopper integrated pest management demonstration project. U.S. Dept. Agri. APHIS. Tech. Bull. 1891. 124pp.
6

Figure 1. Bucket cages used for evaluating baits for rangeland grasshoppers.
Figure 2. Evaluating selected grasshopper baits in cage studies near Edgemont, South Dakota, 2005.
7

Figure 3. Monitoring bucket cages used in evaluating grasshopper mortality in South Dakota
Figure 4. Cordillacris occipitalis Adult male Photo courtesy of Robert E. Pfadt
Figure 5. Amphitornus coloradus Adult male Photo courtesy of Robert E. Pfadt
Figure 6. Trachyrhachys kiowa Adult male Photo courtesy of Robert E. Pfadt
8

0
5
10
15
20
25
30
35
1 2 3 4 5 6
Days after treatment
Mea
n %
mor
talit
y
Tast-E-Bait 2% Crumbles 2% Wheat bran 2% Untreated
Figure 7. Mortality of Cordillacris occipitalis treated with selected carbaryl baits in field cages near Edgemont, South Dakota, 2005.
05
1015202530354045
1 2 3 4 5 6
Days after treatment
Mea
n %
mor
talit
y
Tast-E-Bait 2% Untreated
Figure 8. Mortality of Amphitornus coloradus treated with a food waste bait in field cages near Edgemont, South Dakota, 2005.
9
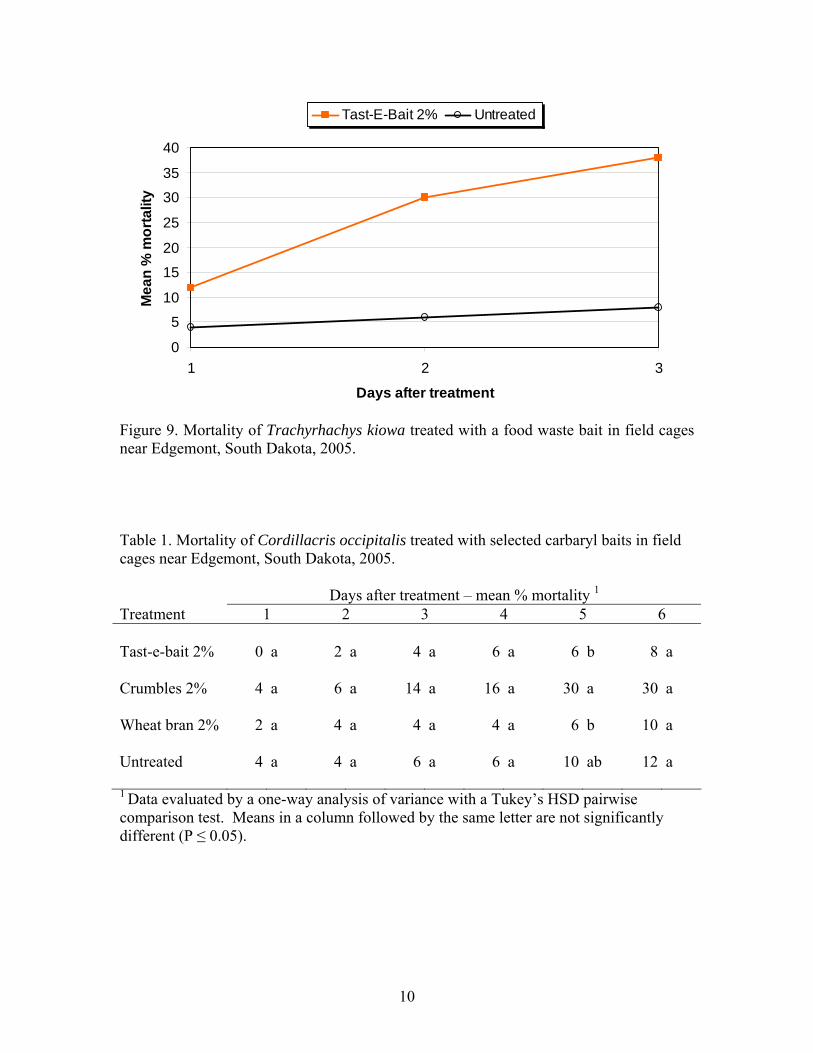
0
5
10
15
20
25
30
35
40
1 2 3
Days after treatment
Mea
n %
mor
talit
y
Tast-E-Bait 2% Untreated
Figure 9. Mortality of Trachyrhachys kiowa treated with a food waste bait in field cages near Edgemont, South Dakota, 2005. Table 1. Mortality of Cordillacris occipitalis treated with selected carbaryl baits in field cages near Edgemont, South Dakota, 2005. Days after treatment – mean % mortality 1
Treatment 1 2 3 4 5 6 Tast-e-bait 2% 0 a 2 a 4 a 6 a 6 b 8 a Crumbles 2% 4 a 6 a 14 a 16 a 30 a 30 a Wheat bran 2% 2 a 4 a 4 a 4 a 6 b 10 a Untreated 4 a 4 a 6 a 6 a 10 ab 12 a 1 Data evaluated by a one-way analysis of variance with a Tukey’s HSD pairwise comparison test. Means in a column followed by the same letter are not significantly different (P ≤ 0.05).
10
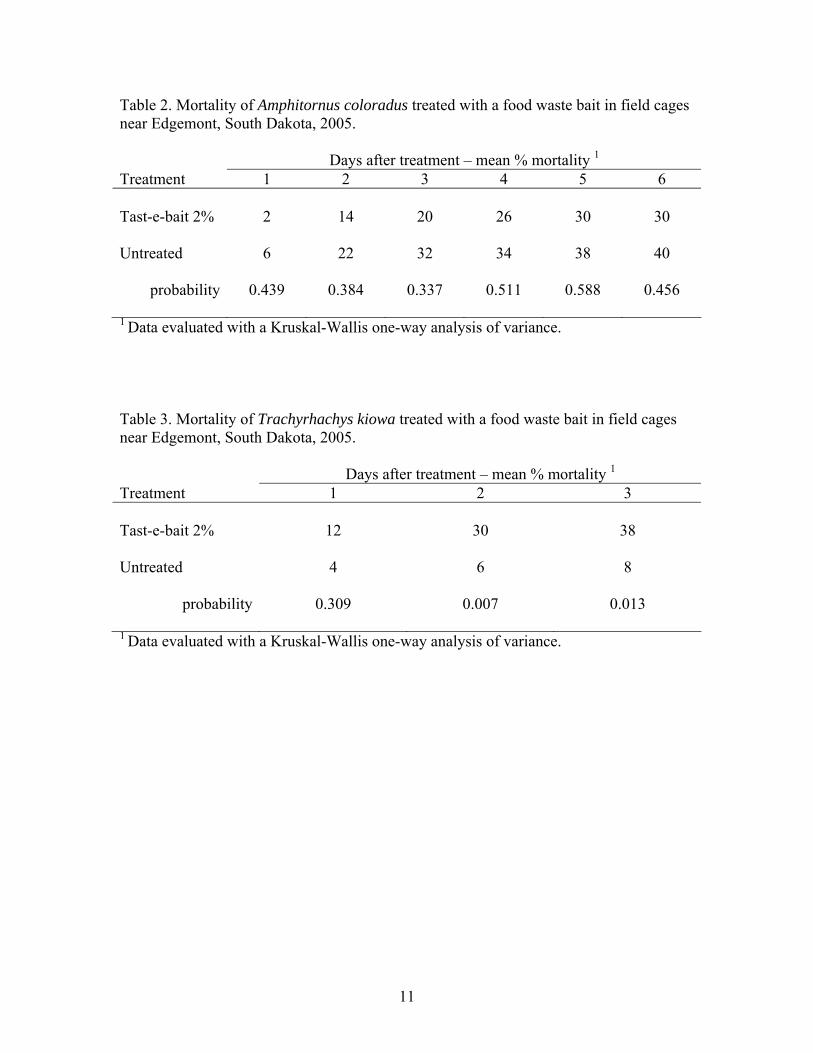
11
Table 2. Mortality of Amphitornus coloradus treated with a food waste bait in field cages near Edgemont, South Dakota, 2005. Days after treatment – mean % mortality 1
Treatment 1 2 3 4 5 6 Tast-e-bait 2% 2 14 20 26 30 30 Untreated 6 22 32 34 38 40
probability 0.439 0.384 0.337 0.511 0.588 0.456
1 Data evaluated with a Kruskal-Wallis one-way analysis of variance. Table 3. Mortality of Trachyrhachys kiowa treated with a food waste bait in field cages near Edgemont, South Dakota, 2005. Days after treatment – mean % mortality 1
Treatment 1 2 3 Tast-e-bait 2% 12 30 38 Untreated 4 6 8
probability 0.309 0.007 0.013 1 Data evaluated with a Kruskal-Wallis one-way analysis of variance.

March, 2006
Re-examining the secondary mortality of Mormon crickets resulting from consumption of carbaryl bait induced cadavers
R. Nelson Foster, K. Chris Reuter and Lonnie R. Black
Organization
USDA-APHIS-PPQ-CPHST-Decision Support and Pest Management Systems Laboratory, Phoenix, Arizona.
Cooperators Dave McNeal and Rob McChesney USDA,APHIS,PPQ, Boise, Idaho Lori Atkins and Donna Hooper, landowner, near Mayfield, Idaho Neil Wagner, Peacock Industries, Hague, SK, Canada Joe Millinax, Wilbur-Ellis, Fresno, California David I. B. Vander Hooven, Advanced Organics Inc., Maumee, Ohio
1

Abstract No secondary mortality occurred when Mormon crickets fed on cadavers resulting from wheat bran baits containing 2% and 5% carbaryl. Both concentrations in apple pumice and food waste baits produced statistically significant secondary mortality. There was no significant advantage with the higher dose for either bait. While secondary mortality may be advantageous in some cases, justification for the higher concentration appears unwarranted. Thoroughly washing the bait induced cadavers (Mormon cricket cadavers resulting from mortality produced by poison baits) before presentation to unexposed Mormon crickets was required to accurately assess the level of secondary mortality.
Introduction
Treatments for controlling Mormon crickets have historically relied on ground applied wheat bran or steamrolled oat baits containing carbaryl. Even though diflubenzuron sprays have recently proved highly effective (Foster et al. 2002) and become extremely popular, baits applied by ground or air remain an important tool for controlling Mormon crickets. Flaky wheat bran bait (2% carbaryl) has been used in many states, but steam-rolled wheat and/or apple pumice bait (5% carbaryl) have gained recent popularity in some states. Apple pumice bait was used for the first time in APHIS sponsored Mormon cricket programs in Idaho in 2002. Another bait, based on food waste products (a combination of bakery, snack, cereal and confectionary waste) became available for evaluation in 2003. Mortality from solid baits is popularly thought to be attributed to both primary and secondary pickup of the toxicant by Mormon crickets. However, unpublished data has shown that the older typical carbaryl wheat bran bait treatments did not demonstrate secondary kill properties (Foster et al. 1979, 1996). More recently, little evidence of secondary mortality was seen with the newer baits when immature Mormon crickets consumed carbaryl bait induced cadavers (Mormon cricket cadavers resulting from mortality produced by poison baits) (Foster et al. 2003). In that study, an indication (although not consistently statistically significant within the test) of increased secondary mortality was seen with the higher concentration of carbaryl in the apple pumice bait against immature Mormon crickets in single cadaver presentation studies at 8 days after treatment. An additional study, with multiple cadaver presentations, indicated a significant statistical increase in secondary mortality only after fresh cadavers, produced with apple pumice bait, were provided to test Mormon crickets successively for 6 days. However, in secondary mortality studies with adults, Foster et al. (2004), showed apple pumice bait (“Crumbles” 5%) and food waste bait (Tast-E-Bait 2% and 5%) resulted in substantially significant secondary mortality to Mormon crickets while wheat bran bait (Eco Bran) resulted in no significant secondary mortality. While those results were very compelling, based on subsequent un-replicated studies, the degree of differences that resulted from the treatments were considered inaccurate as a result of inconsistent procedures used to insure no adherence of bait to the cadaver.
2

The development of additional secondary mortality data on Mormon cricket will help to clarify the activity levels and potential of the different baits and to standardize the use of carbaryl concentrations in those baits. Additionally, new information may lead to a new bait option or standard for Mormon cricket control. Where secondary kill is not considered an advantage, lower use rates of carbaryl could have a reduced impact on the environment and should reduce the cost of the formulated bait. Where higher rates may be needed, an overall increased effectiveness in a control effort may result. Secondary mortality may be important in situations where extremely large bands of migrating Mormon crickets overwhelm the treatment and consume all of the bait. The following study was conducted to re-examine and compare the effectiveness of wheat bran, apple pumice and food waste baits containing 2% and 5% carbaryl for secondary mortality against adult Mormon crickets in Idaho and to confirm the suspicions concluded in the 2004 study on secondary mortality (Foster et al. 2004).
Objectives Re-examine in field cages, the level of secondary mortality that can be attributed to apple pumice bait (crumbled), wheat bran bait and a food waste bait (combined bakery, snack, cereal and confectionary waste) each containing carbaryl, when adult Mormon crickets are exposed to potential meals of single bait-induced cadavers (Mormon cricket cadavers resulting from mortality produced by poison baits). More specifically to: (1) Compare bait types for secondary mortality. (2) Compare 2% and 5% concentrations for secondary mortality.
Materials and Methods The study area was located ca. 18.5 miles southeast of the Boise, Idaho airport, near the community of Mayfield on rangeland provided by Lori Atkins. This location was selected because of the history of Mormon crickets in the area, proximity to current known populations of Mormon crickets, and availability of historically infested rangeland without livestock. Small cages (two gallon bottomless buckets modified with screen sides and top) were established on untreated, improved rangeland to determine the secondary mortality of Mormon crickets resulting from consumption of poisoned cadavers (Figures 1 and 2). The cadavers (freshly killed crickets) were produced by confining single unexposed crickets to 10 lbs/acre of 2% and 5% carbaryl in apple pumice (Sevin 2% and 5% Bait “Crumbles”, Wilbur Ellis), wheat bran (Eco Bran, Peacock Industries Inc.), and food waste (Tast-E-Bait, Advanced Organics Inc.) baits in 4.5 inch diameter x 3 inch high squat paper cups with screen tops in a temporary laboratory established for this purpose (Fig. 3). All crickets were 7th instar stage or adult. Cadavers were washed in running water before placement in cages to prevent fine particles of bait from adhering to the
3

cadavers. All treatments were replicated 4 times with 10 cages per replicate utilizing a total of 280 squat cups and 280 bucket cages. An artificially killed (frozen overnight) cricket was included in the cages designated as untreated controls. Cricket mortality within cages was monitored daily for 7 days. The study was conducted from June 1 - 7, 2005. Analysis. Analysis of all cage study data relied on a one-way analysis of variance and an all pair- wise multiple comparison procedure (Tukey Test). Analyses were conducted on both unranked and ranked data (SPSS Inc., 1997).
Results
No significant secondary mortality resulted from cadavers produced with the 2% and 5% concentrations of carbaryl in wheat bran bait when compared to untreated cadavers at any of the intervals studied (Fig. 4, Table 1). Also the 2% concentration in the food waste bait produced no significant secondary mortality compared to untreated cadavers at one day after treatment. With that exception, both concentrations in the apple pumice and food waste baits resulted in significantly higher secondary mortality than produced by untreated cadavers at all intervals examined. These levels of mortality were also significantly higher than that produced by either concentration in wheat bran except for 2% in apple pumice at one day after exposure that was equivalent to 5% wheat bran and 2% in food waste bait that was equivalent to both concentrations in wheat bran at one day after exposure and equivalent to the 5% concentration in wheat bran at 4 days after exposure. There was no significant difference in secondary mortality resulting from cadavers produced with the 5% concentration and the 2% concentration for any of the three baits even though all treatments performed in numerical dose rank order. The two concentrations in apple pumice and food waste baits resulted in equivalent secondary mortality at all three intervals examined except for 2% in food waste that was significantly lower than 5% in apple pumice at 7 days after exposure. Within each replicate of each treatment 70 to 100 % of bait induced cadavers had been fed upon at one day after exposure (80 to 100 % in untreated controls). By 4 days after exposure the mean percentage of cadavers that had been fed upon by each treatment ranged from 92.5 to 97.5 % (95% controls). The percentage of cadavers that had been fed upon, for all of the studied intervals, is shown in Table 2.
Discussion
In this study, it is clear that irregardless of carbaryl concentration, wheat bran baits produced no secondary mortality while both apple pumice and food waste baits produced secondary mortality. These differences may reflect a higher level of toxicant in cadavers which fed on particles of greater mass compared to wheat bran. Particles of apple pumice and food waste baits have greater mass than those of wheat bran. Particles of greater
4

mass, containing more toxicant, may have allowed for transfer of a higher level of carbaryl to the secondary consumer, especially if a single “bite” (initial feeding) of these types of particles provides substantially more than a lethal dose. It is reasonable to expect that as soon as the cricket consumes a dose sufficient to cause sickness, that feeding will discontinue, and in the case with small particles, less total bait will be consumed. Therefore, less toxicant will be transferred to be picked up by the cannibalizing cricket. Because of the small particle mass, more wheat bran bait may remain on the ground for other cricket consumers compared to the other baits. Conversely, the small particle size may decrease the overall effectiveness of the treatment effort if particles are undetected by the crickets. It must be noted that in most cases it has been observed that after the initial “bite” (feeding) little more bait is consumed. Although an exception was observed when a single cricket consumed the entire amount of wheat bran bait applied at the rate of 10 lbs/ acre within a squat cup container and then became sick and incapacitated in 7 minutes. The lack of secondary mortality attributed to wheat bran is consistent with the results from unpublished secondary kill studies conducted in Nevada in 1995 (Foster et al. 1996). The secondary mortality attributed to apple pumice and food waste baits is consistent with results from earlier studies (Foster et al. 2004), even though the degree of differences in those results were questioned by the authors. It is also clear from that study, and this one, that cadavers must be thoroughly washed to insure no particles of bait may be adhering to the cadavers before being used in secondary mortality/kill studies.
Secondary kill may not be desirable because of the potential for reducing populations of other non-target arthropods that may feed on cadavers of Mormon crickets, although this has not been demonstrated. However, there may be cases where secondary mortality is desirable, in order to increase the effectiveness of the treatment actions. Additional mortality provided through secondary action on cannibalizing Mormon crickets may actually reduce if not eliminate the need for retreating the same area again, a frequent occurrence. This could reduce the overall pesticide load on both the immediate and general area. As long as bait is present on the ground there should be no need for secondary mortality. Studies show that the level of control attributed to secondary mortality, when it occurs, is always substantially less than initial mortality. More primary mortality is seen with these baits than secondary mortality. However, if all of the applied bait has been consumed, then mortality attributed to consumption of bait induced cadavers would represent additional mortality than would be expected without bait on the ground, and would represent a benefit to the control effort. Secondary mortality could also be beneficial if crickets move beyond or across a strip where bait was applied. However, this activity would not substantially expand the active area as observations have indicated that once bait is ingested, sickness and subsequent spasms with incapacitation occurs as soon as 7 minutes, followed by eventual death within hours. Once sick, the cricket is incapable of significant ground movement.
5

When discussing the concentration of carbaryl in bait it is important to note that no differences in primary mortality were seen between 2 and 5 % concentrations in wheat bran based baits in 1977 and 1978 on nymphs or adults (Foster et al. 1979) or in 1995 on nymphs (Foster et al. 1996). However, those studies were conducted with bakers bran, which contained large particles of wheat bran, compared to the currently used bran. It is noteworthy that, changes in the wheat bran bait have occurred over the years as a result of several changes in producers. Most notable is the presence of finer particles of feed grade wheat bran compared to the large or course flaky bakers grade wheat bran used in 1950’s and 1970’s. Unpublished data collected during the earliest study (Foster et al. 1979), revealed no secondary mortality resulted from wheat bran bait. In 2003 no difference was seen between the 2 and 5 % concentrations in wheat bran and apple pumice in primary mortality with nymphs but some evidence on adults indicated secondary kill with 5 % apple pumice bait (Foster et al. 2003). In the 2004 study with adults, no statistical difference was seen between 2 and 5 % concentrations in apple pumice and food waste baits on primary kill but some evidence was seen with wheat bran that indicated lower mortality with the lower concentration (Foster et al. 2004). The 2% wheat bran resulted in lower mortality and took 9 to 10 days after treatment to attain statistically similar mortality. Those differences were considered possibly a result of finer particle composition in the wheat bran bait noted during the studies. Other tests conducted during that study indicated that coarser particles of bran produced higher mortality than fine particles. However, the lower mortality associated with fine particles may simply be a function of particle discovery by the insect. While wheat bran produced no secondary mortality, both apple pumice and food waste baits produced secondary mortality and showed some advantage (although that data was questioned by the authors) with the higher concentration. These later two studies used wheat bran baits consisting of substantially more fine particles compared to the coarser particles of bakers bran used in the earlier studies.
To summarize four earlier studies and this one (Table 3): Wheat bran baits produced no secondary mortality and showed no advantage with the higher concentration and the older coarser bait on nymphs or adults. With the more recent baits containing finer particles, no advantage against immatures was seen with the higher concentration in primary mortality and no secondary mortality was produced. However against adults, while no secondary mortality was produced, an advantage of the higher concentration was seen in primary mortality. In contrast, both apple pumice and food waste baits produced significant secondary mortality but the increased concentration provided no advantage in producing either primary or secondary mortality.
If secondary mortality is desired, then it is clear that apple pumice and food waste baits are superior to the current wheat bran baits. However, because there is no statistical significant advantage in terms of mortality, the use of higher concentrations of carbaryl (5% vs 2%) in these baits appears unjustified.
6

Conclusions
No significant secondary mortality resulted from 2% and 5% carbaryl induced cadavers produced with wheat bran bait. Secondary mortality resulting from apple pumice and food waste bait induced cadavers was significantly higher than mortality from consumption of normal (non-bait induced) cadavers. However, there was no significant difference in secondary mortalities resulting from 2% and 5% concentrations of carbaryl in both apple pumice and food waste based baits. While secondary mortality may be advantageous in some cases, there is no justification for the higher concentration. Further studies should be conducted to refine the knowledge regarding the impact of wheat bran particle sizes on primary and secondary mortality.
Acknowledgements
The authors wish to thank Lori Atkins and Donna Hooper for providing land used in the study, facilities for storage of equipment and supplies, shelter for the researchers during inclement weather, and their continued cooperation, since 2002, toward learning more about Mormon crickets and their management. Thanks are also extended to Dave McNeal and Rob McChesney for their continued assistance in locating and capturing appropriate aged populations of Mormon crickets for testing, and for providing unexpectedly needed miscellaneous equipment and supplies which made our efforts more efficient. Thanks are also extended to Neal Wagner, Peacock Industries for providing wheat bran bait, to Gary Saxton, Wilber Ellis for providing the apple pumice bait, and to David I. B. Vander Hooven, Advanced Organics Inc. for providing the food waste bait.
References Cited
Foster, R.N. C.H. Billingsley, R.T. Staten and D.J. Hamilton 1979. Field cage tests for concentrations of carbaryl in a bait and its application rates for control of Mormon Cricket. Jour. Econ. Entomol. 72: 295-297. Foster, R. N., K. C. Reuter, G. Abbott, S. Prestwich and C. Gentry. Dec. 2002. Report. Field evaluation of aerially applied Dimilin and field cage evaluation of selected doses of Dimilin and Fipronil baits against Mormon cricket, Anabrus simplex Haldeman, on rangeland in Utah. USDA-APHIS-PPQ-CPHST-DS&PMSL.
Foster, R. N., K. Chris Reuter, Lonnie R. Black, Dave McNeal and Rob McChesney. 2004. Comparison of wheat bran, apple pumice and food waste (combined bakery, snack, cereal, and confectionary) baits containing carbaryl for controlling Mormon crickets: Studies of longevity and primary and secondary mortality. Report. USDA-APHIS-PPQ-CPHST-DS&PMSL, Phoenix, AZ. 12 pp.
7

Foster, R. N., K. Chris Reuter, Dave McNeal and Rob McChesney. 2003. Comparison of flakey wheat bran and apple pumice baits for controlling Mormon cricket: Evaluating initial and secondary mortality with baits containing 2% and 5% carbaryl. Report. USDA-APHIS-PPQ-CPHST-DS&PMSL, Phoenix, AZ. 10 pp. Foster, R. N., K. C. Reuter, G. Nash and D. Kail. 1996. Carbaryl bran bait: Secondary kill in Mormon crickets. Oral presentation, printed abstract. Proceedings of the National Grasshopper management Board Annual Meeting. Jan. 9 - 11, Rapid City, SD. SPSS Inc. 1997. SYSTAT for Windows: Statistics, SPSS Inc., Chicago, Illinois.
Figure 1. Close-up view of field bucket cage used to confine Mormon crickets.
8

Figure 2. View of field bucket cage setup used in Mormon cricket bait studies.
Figure 3. Squat cups (16 oz) used to confine single Mormon crickets treated with 10 lbs of bait per acre to produce fresh cadavers.
9

01020304050607080
1 4 7
Days after treatment
Mea
n %
mor
talit
yUntreated Wheat bran 2% Wheat bran 5% Apple pumice 2%Apple pumice 5% Food waste 2% Food waste 5%
c
bc
ab
a
abc
a
c c
bc
a
a
ab
a
d d
cd
abc
a
bc
ab
c
Figure 4. Secondary mortality of 7th instar and adult Mormon crickets exposed to single carbaryl bait induced cadavers in field cages near Boise, Idaho – 2005. Table 1. Secondary mortality of 7th instar and adult Mormon crickets exposed to single carbaryl bait induced cadavers in field cages near Boise, Idaho – 2005. Days after treatment – mean % mortality 1
Treatment 1 2 3 4 5 6 7 Eco-bait 2% 5.0 bc 5.0 cd 5.0 c 5.0 c 5.0 c 5.0 d 5.0 d Eco-bait 5% 12.5 bc 12.5 bcd 12.5 bc 12.5 bc 12.5 bc 17.5 cd 17.5 cd Crumbles 2% 30.0 ab 35.0 ab 40.0 a 42.5 a 47.5 a 47.5 ab 47.5 ab Crumbles 5% 40.0 a 47.5 a 52.5 a 52.5 a 55.0 a 60.0 a 67.5 a Tast-e-bait 2% 20.0 abc 25.0 abc 32.5 ab 32.5 ab 35.0 ab 35.0 bc 37.5 bc Tast-e-bait 5% 42.5 a 42.5 a 47.5 a 52.5 a 55.0 a 55.0 ab 55.0 ab Untreated 0.0 c 0.0 d 2.5 c 2.5 c 5.0 c 5.0 d 5.0 d
10

Table 2. Percentage of bait induced cadavers showing evidence of being fed on by unexposed Mormon crickets. Days after exposure - % cadavers fed on Treatment rep 1 2 3 4 5 6 7 Eco-bait 2% 1 80 80 80 100 100 100 100 2 80 90 90 100 100 100 100 3 90 90 90 100 100 100 100 4 80 80 90 90 90 90 100
mean 82.5 85 87.5 97.5 97.5 97.5 100 Eco-bait 5% 1 80 90 90 100 100 100 100 2 80 80 80 100 100 100 100 3 80 80 100 100 100 100 100 4 80 90 90 90 100 100 100
mean 80 85 90 97.5 100 100 100 Crumbles 2% 1 70 70 80 90 90 90 100 2 80 100 100 100 100 100 100 3 80 80 90 100 100 100 100 4 90 90 100 100 100 100 100
mean 80 85 92.5 97.5 97.5 97.5 100 Crumbles 5% 1 100 100 100 100 100 100 100 2 80 80 80 80 90 100 100 3 90 90 90 90 90 90 90 4 90 100 100 100 100 100 100
mean 90 92.5 92.5 92.5 95 97.5 97.5 Tast-e-bait 2% 1 80 80 80 80 100 100 100 2 80 80 80 90 100 100 100 3 80 100 100 100 100 100 100 4 100 100 100 100 100 100 100
mean 85 90 90 92.5 100 100 100
Tast-e-bait 5% 1 100 100 100 100 100 100 100 2 70 80 90 90 90 90 90 3 90 90 100 100 100 100 100 4 80 90 90 90 90 90 90
mean 85 90 95 95 95 95 95
Untreated 1 80 90 90 90 90 90 100 2 80 100 100 100 100 100 100 3 100 100 100 100 100 100 100 4 80 80 90 90 90 90 90
mean 85 92.5 95 95 95 95 97.5
11

Table 3. Summary of bait activity against Mormon cricket in field cage studies, 1976 – 2006. Primary Mortality Year Secondary Mortality
Bait Immature Adult Immature Adult Type 2% vs. 5% 2% vs. 5% 2% vs. 5% 2% vs. 5%
1976-1977 wheat bran = = 0 1995 wheat bran = 0 2003 wheat bran = 0 apple pumice = ≠ 2004 wheat bran ≠ 0 apple pumice = ≠? food waste = ≠? 2005 wheat bran 0 apple pumice = food waste = = mortality resulting from 2% and 5% carbaryl in bait statistically equal ≠ 5% carbaryl in bait resulted in statistically higher mortality than 2% carbaryl in bait ≠? data questioned 0 no significant mortality
12

13

February, 2006
Field cage comparison of a food waste bait containing three doses of Spinosad for activity against the grasshopper, Aulocara elliotti (Thomas) on rangeland
R. Nelson Foster, K. Chris Reuter, Lonnie Black
Organizations
USDA/APHIS/PPQ/CPHST/DS&PMSL, Phoenix, AZ.
Cooperators
David I. B. Vander Hooven, Advanced Organic Inc., Maumee. Ohio. Dow AgroSciences, Indianapolis, Indiana
Abstract
Doses of 2.65, 0.66 and 0.17 g AI/ac of Spinosad in a food waste bait (combined bakery, snack, cereal and confectionary food waste) applied at 10 lbs/ac against a carbaryl wheat bran bait susceptible grasshopper species, Aulocara elliotti, on southwestern South Dakota rangeland resulted in mortality of 60, 49 and 12 % respectively at 7 days after treatment. These doses were 1/16 of the doses used in previous aerial spray treatment studies with Spinosad. Results are extremely encouraging and suggest further work should be conducted on higher doses of Spinosad in bait formulations with wheat bran, apple pumice and food waste carriers.
Introduction
Grasshopper control /management/suppression programs sponsored by the USDA APHIS to protect rangelands rely on both sprays and solid bait. Several toxicant options, carbaryl, diflubenzuron or malathion, exist when sprays are selected. However, when solid bait is selected as the treatment of choice, carbaryl is the only toxicant choice available even though several carriers, wheat bran, apple pumice and food waste based baits are available. While some attempts to develop baits with alternative toxicants have occurred, little advancement in this area has been achieved. Malathion on corn cob grits showed little effectiveness (Foster et al. 1989) and early development of chlorpyrifos bait and diflubenzuron bait showed no advantage over the existing carbaryl bait formulations (Jech et al. 1989; 1993). However, solid baits are deemed highly desirable when
1

opposition to aerial spray exists or in some sensitive areas where use of lower levels of toxicant are advantageous. Spinosad, a bacteria derived toxin produced by Saccharopolyspora spinosa an aerobic, non-antibiotic actinomycete (Dow AgroSciences), has demonstrated some potential for development as an alternative spray. It is desirable because of it safety to humans, fish, birds, other non-target organisms and registration for organic crops. Results from two aerially applied spray studies conducted in South Dakota indicated that doses in the range of 0.75 to 1.5 fluid oz/acre (10.6 - 21.3 g AI/ac) could produce acceptable reduction in grasshoppers while leaving little residual activity at 3 days after treatment (Foster et al. 2002 and Foster et al. 2003). Since mortality results primarily from ingestion (Foster et al 1996), that activity suggests potential if developed also as a solid bait. Baits containing carbaryl have been used for many years to suppress grasshopper or Mormon cricket populations on western rangelands. Currently, carbaryl is used in wheat bran and apple pumice based baits at both 2% and 5% concentrations against Mormon crickets, Anabrus simplex Haldeman, and is commonly applied at a rate of 10 lbs/acre with ground or aerial application. Rangeland grasshoppers are currently treated with 2% carbaryl wheat bran bait applied at 2 lbs/acre by ground and 1.5 lbs/acre by air. While little work has been conducted with baits on Mormon cricket since 1977 (Foster et al. 1979), extensive work has been conducted with wheat bran bait containing carbaryl on numerous species of grasshoppers (Onsager et al. 1980 a, b; Onsager et al. 1996-99; Quinn et al. 1996; Jech et al. 1996; Reuter et al. 1996; Foster et al. 1998; Foster et al. 1999).The recent increase in grasshopper and particularly Mormon cricket populations in the western states has renewed interest in developing new bait options for controlling these rangeland pests. An apple pumice bait has recently received considerable attention for use against Mormon cricket (Foster et al. 2003). Additionally, a new product, Tast-E-Bait, (a combined bakery, snack, cereal and confectionary food waste based bait produced by Advanced Organic Inc.) has recently become available for consideration. It has been recently evaluated substantially against both immature and adult Mormon crickets (Foster et al. 2004 a). Effective concentrations of carbaryl in bran bait exist at 1/16 of the spray rate, 0.03 lb AI/ac and 0.50 lb AI/ac respectively. A commensurate reduction from spray rate to bait rate with Spinosad would improve the economics of that toxicant and may provide an additional option when solid baits are needed to combat grasshoppers on rangeland. Initial studies with a food waste carrier and an apple pumice carrier against two wheat bran bait susceptible species of grasshoppers (Ageneotettix deorum and Aulocara elliottti) indicated similar results with carbaryl irregardless of the carrier (Foster et al. 2004 b). Therefore any or all three carriers may be acceptable for future grasshopper bait development. The following study was designed to initially evaluate and compare in field cages, 3 doses of spinosad in a food waste bait carrier for activity against a common rangeland grasshopper, Aulocara elliotti, known to be susceptible to carbaryl in wheat bran bait (Onsager et al. 1996-1999) as well as in apple pumice and food waste based baits (Foster et al 2004 b).
2

Methods and Materials
The study was conducted in Fall River County of southwestern South Dakota ca. 12 miles west and 9 miles north of the town of Edgemont on the Mark Tubbs ranch during the period of June 29 through July 6, 2005. The location was selected because of the diversity in grasshopper species, density of grasshoppers, history of grasshoppers in the area, closeness to other studies occurring in the area and availability of a rangeland pasture without livestock for the period of time required in the study.
The Tracer formulation of Spinosad, produced by DowAgroScience (containing 4 lbs / gal of AI Spinosad) was used to formulate three different doses of Spinosad on the food waste bait carrier, Tast-E-Bait, (a combined bakery, snack, cereal and confectionary food waste based bait produced by Advanced Organic Inc.) Doses were arrived at by using 1/16 of the liquid doses previously used or proposed in earlier rangeland studies (3, 0.75 and 0.1875 fl oz of Spinosad/acre). This reduction was based on the reduction from 0.50 lb AI/ac of carbaryl used in spray formulations (16 fluid ozs of Sevin XLR Plus / ac) to 0.03 lb AI/ac of carbaryl used in bait formulations (1.5 lbs/ac of 2% carbaryl in wheat bran bait). The three doses were calculated as follows: 1. .06 x 3 fluid ounces = .06 x 88.71 ml = 5.3 ml in 1.5 to 2.0 lbs of bait /acre 2. .06 x 0.75 fluid ounces = .06 x 22.17 ml = 1.33 ml in 1.5 to 2 lbs of bait / acre 3. .06 x 0.1875 fluid ounces = .06 x 5.54 ml = 0.3326 ml in 1.5 to 2.0 lbs of bait / acre Continuing: (ml in one pound of bait – one half of above) 1. 2.65 ml up to 10 ml with water and applied to one pound of bait 2. 0.66 ml up to 10 ml with water and applied to one pound of bait 3. 0.17 ml up to 10 ml with water and applied to one pound of bait Continuing: (3785 ml contains 4 lbs AI) 1. 0.002899 lb AI/lb bait x 453.6 = 1.315 grams AI/lb bait x 10 = 13.15 gm AI/ac 2. 0.000697 lb AI/lb bait x 453.6 = 0.316 grams AI/lb bait x 10 = 3.16 gm AI/ac 3. 0.000184 lb AI/lb bait x 453.6 = 0.083 grams AI/lb bait x 10 = 0.83 gm AI/ac Each 10 ml solution was applied by using a DeVilbiss 15 spray atomizer supplied with continuous 5 psi of air from a small electric pump. Each formulation was sprayed for ca. 1 to 1.5 minutes into a 5 gallon bucket containing one pound of bait which was rotating on a small 5 gallon size electric powered mixer. After all material was sprayed the mixer was allowed to tumble the formulation for 10 additional minutes. Upon completion of mixing, all materials were stored at room temperature and out of any light until they were applied in the study. All formulations and concentrations were applied at the rate of 10 lbs/acre which resulted is Spinosad doses of 13.15 g AI/ac, 3.16 g AI/ac and 0.83 g AI/ac. Ten cages (two gallon bottomless buckets modified with screen sides and top) (Fig. 1) were established on
3

rangeland for each treatment. Ten additional cages were established on untreated vegetation as controls for comparison. This design including the untreated control, therefore used a total of 40 cages and 200 grasshoppers (Fig. 2). The appropriate amount of bait was pre-weighed in the laboratory and introduced into each cage separately in the field. Each cage was stocked with five, mostly adults with a few fifth instar stage, Aulocara elliotti grasshoppers (Fig. 3) collected from an adjacent untreated area. This species was selected because of abundance and susceptibility in earlier studies to wheat bran, apple pumice and food waste baits containing carbaryl. Grasshoppers were introduced into cages after bait was applied. After exposure to bait treatments, grasshoppers in all cages including untreated cages were monitored daily for 7 days for mortality. Data were analyzed using an analysis of variance with the Tukey multiple comparison test to separate means. Analyses were performed with SYSTAT for Windows (SPSS Inc. 1977).
Results and Discussion
No mortality occurred in untreated populations until 4 days after treatment (Fig. 1). From then on mortality increased only to 10% at 7 days after treatment. All treatments performed in dose rank order. At 2 days after treatment, there was no significant difference in mortality resulting from any of the treatments and the untreated population. From 2 days after treatment and through the end of the study at 7 days there was no significant difference in mortality resulting from the lowest dose of Spinosad (0.17 g AI/ac) and what occurred in the untreated population. From 2 days after treatment and throughout the study, the two highest doses of Spinosad (2.65 and 0.66 g AI/ac) performed equally well and both resulted in significantly greater mortality than occurred with the lowest dose and in the untreated population. All treatments demonstrated slight but steady increases in mortality during the study. However, mortality attributed to the highest two doses appeared to increase faster than the lowest dose or the untreated population. Mortality from the 2.65 g AI/ac, 0.66 g AI/ac and 0.17 g AI/ac doses in bait at 7 days after treatment were 60 %, 49 % and 12 % respectively and compare with mortality of 97 %, 78 %, 76 % and 63 % from spray doses of 42.54 g AI/ac, 21.27 g AI/ac, 10.64 g AI/ac and 5.32 g AI/ac respectively (Foster et al. 2002, 2003). While it was anticipated that a 1/16 reduction in AI dose from spray to bait would yield a highly acceptable level of mortality, that did not occur. However, the results are very encouraging. Levels of mortality at the two highest bait doses were substantial and indicate that an increase in bait dose may provide acceptable mortality and yet remain at a sufficiently low level of AI (below 5.32 g AI/ac) to become economical. A summary of spray and bait dose responses is seen in Table 1.
4

These levels of mortality compare with levels ranging from 94 % to 100 % with 2 and 5 % carbaryl in wheat bran bait applied at 10 lbs/ac, 90.8 g AI/ac and 227 g AI/ac respectively. This study indicates significant activity with an extremely low dose of Spinosad against a wheat bran carbaryl bait susceptible species. The results suggest further study with higher doses of Spinosad in wheat bran, apple pumice and or food waste based baits is warranted. Slightly higher doses should produce acceptable mortality and may prove more economical than current available sprays containing Spinosad.
Conclusions
Extremely low levels of Spinosad on food waste baits produced substantial mortality and are very encouraging. The experimental treatments with bait produced results in dose rank order. Our data suggests that increases in Spinosad doses in bait should produce acceptable levels of mortality which may be more economical than current available Spinosad sprays being tested against grasshoppers on rangeland. Additional studies conducted at higher doses of Spinosad with wheat bran, apple pumice and food waste based baits is warranted.
Acknowledgements
Special thanks are extended to Mark Tubbs for providing the rangeland for the study and for his continued interest and cooperation since 1989 in developing information relative to improving grasshopper management on rangeland.
References Cited
Foster, R. Nelson, K. Chris Reuter, Bruce Helbig, Cheryl Huddleston and Nathan Bradley. 2002. Initial Evaluations of Malathion and Spinosad Formulations for Control of Grasshoppers on Rangelands in South Dakota. Report. USDAS,APHIS,PPQ, CPHST, DS&PMSL, Phoenix, Arizona. 17 pp. Foster, R. N., K. C. Reuter, and L. Black. 1996. Laboratory evaluation of dose and manner of pickup of Spinosad on the migratory grasshopper 1995. Arthropod Management Tests: 1996 (21) p 416. Foster, R. Nelson, K. Chris Reuter, Lonnie Black, Bruce Helbig, and Lane Kozel. 2004. Field cage comparisons of wheat bran, apple pumice and food waste baits containing carbaryl for controlling the rangeland grasshoppers Ageneotettix deorum (Scudder) and Aulocara elliotti (Thomas).
5

Foster, R.N., L.E. Jech, D.D. Walgenbach, R. Bohls, D. Colletto, J.A. Henserson, W. Meeks, R. Queener, T.J. Roland and J. Burgess. 1989. Initial evaluation of malathion corn cob grit bait for control or suppression of grasshoppers on rangeland near Camp Crook, South Dakota, 1988. In: USDA-APHIS-PPQ, Grasshopper Integrated Pest Management Project (Fiscal Year 1988). pp 217-222. USDA-APHIS-PPQ, Boise, Idaho. Foster, R. N., M. A. Quinn, K. C. Reuter, D. Colletto, R. Houston, M. J. Puclik, A. Scott, and B. Radsick. 1999.Comparison of single and multiple applications of an insecticide bait for controlling grasshoppers (Orthoptera: Acrididae) on mixed-grass prairie. Jour. Kansas Entomol. Soc. 72: 181-189. Foster, R. N., M. A. Quinn, L. L. Herbaugh, D. D. Walgenbach, R. L. Bohls, K. C. Reuter, and T. J. Roland. 1998. Effect of multiple concentrations and rates of an insecticidal bait on grasshoppers (Orthoptera: Acrididae) of mixedgrass prairie. Jour. Kansas Entomol. Soc. 71: 1-10. Foster, Nelson, Stephan Jaronski, Chris Reuter, Bruce Helbig, John Bradley, Cheryl Huddleston, Bryan Fitzgerald, Julie Grace and Bruce Radsick. 2003. Evaluation of the fungus Beauveria bassiana in canola oil and paraffinic oil and identifying the low dose of a bacteria derived toxin, spinosad, for use against rangeland grasshoppers. Report. USDAS,APHIS,PPQ, CPHST, DS&PMSL, Phoenix, Arizona. 33 pp. Jech, L. E., D. D. Walgenbach, R. N. Foster, R. A. Bohls, D. Colletto, J. Burgess,W. Meeks, R. Queener.1989. Field trials of chlorpyrifos and carbaryl baits for control of grasshoppers on rangeland near Buffalo, South Dakota. In: USDA-APHIS-PPQ Grasshopper Integrated Project. Annual Report (Fsical Year 1988). pp.206-217. USDA-APHIS-PPQ, Boise, Idaho. Jech, L. E., R. N. Foster, D. Coletto, D. D. Walgenbach, T. J. Roland, G. D. Rodriguez, R. Bohls, R. D. Houston, W. K. Meeks, R. L. Queener, C. L. Jackson, J. L. Dines, M. K. Puclik, and A. K. Scott. 1993. Field evaluations of diflubenzuron and carbaryl bran baits against grasshopper (Orthoptera: Acrididae) populations in South Dakota. Jour. Econ. Entomol. 86: 557-565. Onsager, Jerome A., R. Nelson Foster and Larry Jech. 1996-1999. Bait Acceptance by Different Grasshopper Species and Instars. In: Cunningham, G.L., and M.W. Sampson tech cords. Grasshopper integrated pest management user handbook. Tech Bull. 1809. Washington, DC: U.S. Dept. of Agriculture, Animal and Plant Health Inspection Service: II.12-1—II.12-5. SPSS Inc. 1997. SYSTAT for Windows: Statistics, SPSS Inc., Chicago Illinois.
6

Figure 1. Bucket cage used for evaluating baits for rangeland grasshoppers.
Figure 2. Evaluating selected doses of Spinosad bait against Aulocara elliotti.
7

Figure 3. Aulocara elliotti, adult male. A common rangeland grasshopper species that is susceptible to wheat bran, apple pumice and food waste based baits containing carbaryl.
0
10
20
30
40
50
60
70
2 3 4 5 6 7
Days after treatment
Mea
n %
mor
talit
y
Spinosad 2.65 Spinosad 0.66 Spinosad 0.17 Untreated
a
aaa
aa
bb
a
a
b
b b
b
a
a
a
a
b
b
b
b
a
a
Figure 4. Mean mortality of Aulocara elliotti treated with selected doses of Spinosad (g AI/ac) incorporated into a food waste bait in field cages – Edgemont, SD – 2005.
8
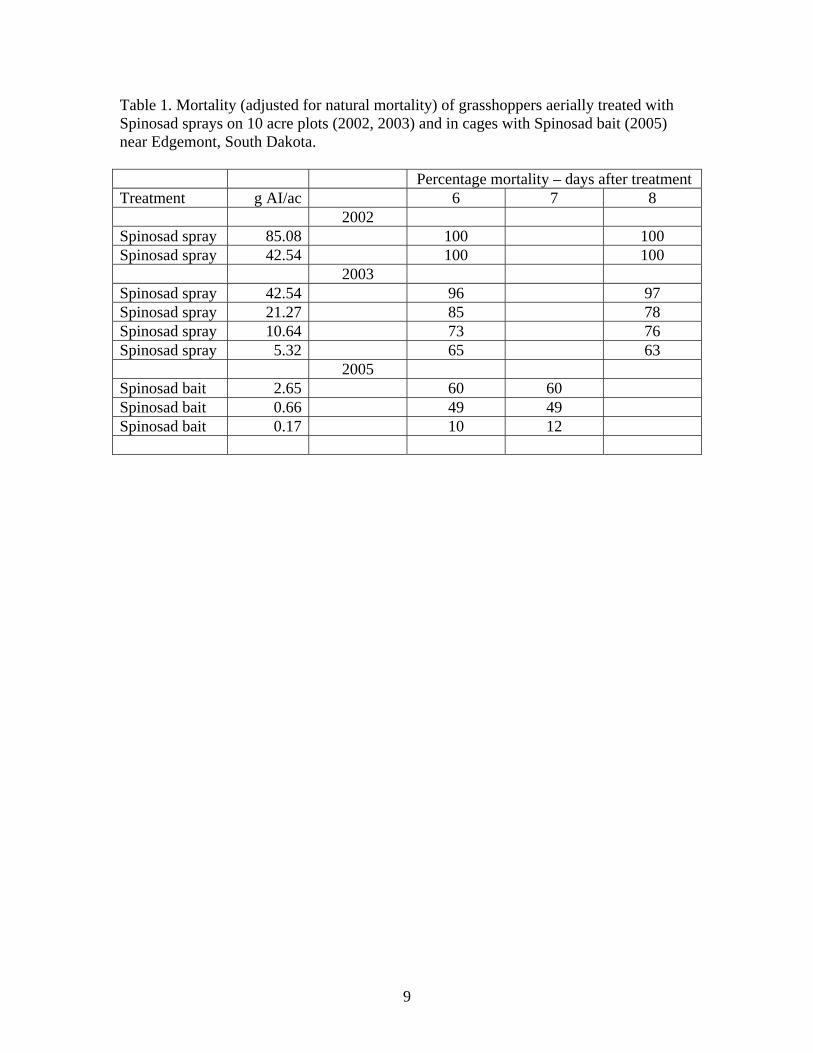
9
Table 1. Mortality (adjusted for natural mortality) of grasshoppers aerially treated with Spinosad sprays on 10 acre plots (2002, 2003) and in cages with Spinosad bait (2005) near Edgemont, South Dakota. Percentage mortality – days after treatment Treatment g AI/ac 6 7 8 2002 Spinosad spray 85.08 100 100 Spinosad spray 42.54 100 100 2003 Spinosad spray 42.54 96 97 Spinosad spray 21.27 85 78 Spinosad spray 10.64 73 76 Spinosad spray 5.32 65 63 2005 Spinosad bait 2.65 60 60 Spinosad bait 0.66 49 49 Spinosad bait 0.17 10 12


















