EZ-Root-VIS: A Software Pipeline for the Rapid Analysis ... · (Armengaud et al., 2009). For...
36
1 1 EZ-Root-VIS: A Software Pipeline for the Rapid Analysis and Visual Reconstruction of 2 Root System Architecture 3 4 Zaigham Shahzad 1 , Fabian Kellermeier 1 , Emily M. Armstrong 1 , Simon Rogers 2 , Guillaume 5 Lobet 3,4 , Anna Amtmann 1* and Adrian Hills 1* 6 7 1 Institute of Molecular, Cell and Systems Biology, University of Glasgow, G12 8QQ, UK 8 2 School of Computing Science, University of Glasgow, G12 8QQ, UK 9 3 Agrosphäre (IBG-3), Forschungszentrum Jülich GmbH, Jülich, 52428, Germany 10 4 Earth and Life Institute, Université catholique de Louvain, Louvain-la-Neuve, 1348, 11 Belgium 12 13 *Corresponding Authors: 14 Anna Amtmann [email protected], Adrian Hills [email protected] 15 16 Short title: Reconstructing root architecture. 17 18 One-sentence summary: 19 EZ-Root-VIS is a free software package that enables the rapid quantification and visual 20 reconstruction of averaged root system architectures for investigating environment-genotype 21 interactions. 22 23 Author contributions: 24 The EZ-Root-VIS pipeline was conceived by F.K. and A.A., and developed by A.H with 25 input from S.R. and G.L. Root growth rate assays were carried out by F.K. GWAS and 26 genotype-environment interaction studies were designed and performed by Z.S. E.M.A. 27 helped with the development of the software pipeline and tested the most recent versions 28 using experimental data. AA managed the project. Z.S., A.A., and A.H., wrote the paper. 29 30 Funding sources: 31 The work was funded by the BBSRC (BB/N018508/1) and by the Gatsby Charitable Trust 32 (Sainsbury studentship to FK). 33 Plant Physiology Preview. Published on June 12, 2018, as DOI:10.1104/pp.18.00217 Copyright 2018 by the American Society of Plant Biologists www.plantphysiol.org on June 6, 2020 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.
Transcript of EZ-Root-VIS: A Software Pipeline for the Rapid Analysis ... · (Armengaud et al., 2009). For...

1
1
EZ-Root-VIS: A Software Pipeline for the Rapid Analysis and Visual Reconstruction of 2
Root System Architecture 3
4
Zaigham Shahzad1, Fabian Kellermeier
1, Emily M. Armstrong
1, Simon Rogers
2, Guillaume 5
Lobet3,4
, Anna Amtmann1*
and Adrian Hills1*
6
7
1 Institute of Molecular, Cell and Systems Biology, University of Glasgow, G12 8QQ, UK 8
2 School of Computing Science, University of Glasgow, G12 8QQ, UK 9
3 Agrosphäre (IBG-3), Forschungszentrum Jülich GmbH, Jülich, 52428, Germany 10
4 Earth and Life Institute, Université catholique de Louvain, Louvain-la-Neuve, 1348, 11
Belgium 12
13
*Corresponding Authors: 14
Anna Amtmann [email protected], Adrian Hills [email protected] 15
16
Short title: Reconstructing root architecture. 17
18
One-sentence summary: 19
EZ-Root-VIS is a free software package that enables the rapid quantification and visual 20
reconstruction of averaged root system architectures for investigating environment-genotype 21
interactions. 22
23
Author contributions: 24
The EZ-Root-VIS pipeline was conceived by F.K. and A.A., and developed by A.H with 25
input from S.R. and G.L. Root growth rate assays were carried out by F.K. GWAS and 26
genotype-environment interaction studies were designed and performed by Z.S. E.M.A. 27
helped with the development of the software pipeline and tested the most recent versions 28
using experimental data. AA managed the project. Z.S., A.A., and A.H., wrote the paper. 29
30
Funding sources: 31
The work was funded by the BBSRC (BB/N018508/1) and by the Gatsby Charitable Trust 32
(Sainsbury studentship to FK). 33
Plant Physiology Preview. Published on June 12, 2018, as DOI:10.1104/pp.18.00217
Copyright 2018 by the American Society of Plant Biologists
www.plantphysiol.orgon June 6, 2020 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

2
34
www.plantphysiol.orgon June 6, 2020 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

3
ABSTRACT 35
If we want to understand how the environment has shaped the appearance and behavior of 36
living creatures we need to compare groups of individuals that differ in genetic make-up and 37
environment experience. For complex phenotypic features, such as body posture or facial 38
expression in humans, comparison is not straightforward because some of the contributing 39
factors cannot easily be quantified or averaged across individuals. Therefore, computational 40
methods are used to reconstruct representative prototypes using a range of algorithms for 41
filling in missing information and calculating means. The same problem applies to the root 42
system architecture (RSA) of plants. Several computer programs are available for extracting 43
numerical data from root images but they usually do not offer customized data analysis or 44
visual reconstruction of RSA. We have developed Root-VIS, a free software tool, which 45
facilitates the determination of means and variance of many different RSA features across 46
user-selected sets of root images. Furthermore, Root-VIS offers several options to generate 47
visual reconstructions of root systems from the averaged data to enable screening and 48
modeling. We have confirmed the suitability of Root-VIS, combined with a new version of 49
EZ-Rhizo, for the rapid characterization of genotype-environment interactions and gene 50
discovery through genome-wide association studies in Arabidopsis (Arabidopsis thaliana). 51
52
INTRODUCTION 53
Roots play a critical role in soil water and nutrient uptake and thus in plant productivity. Root 54
system architecture (RSA) is determined by the relative growth of different parts of the root 55
system as well as frequency of branching and branch angles. RSA is a key determinant of 56
root function and is highly plastic, being controlled by genotype–environment interactions 57
(Yu et al., 2016; Shahzad and Amtmann, 2017; Morris et al. 2017). Optimization of RSA is 58
thus imperative for the improvement of plant resilience, particularly to soil-borne stresses 59
such as soil drought, flooding, nutrient deficiencies, and biotic factors (Julkowska and 60
Testerink, 2015; Lynch, 2015; Rogers and Benfey 2015). Correlation of root morphology 61
with DNA sequence information allows the identification of the genetic loci that underpin 62
root features, for example through mutant screens, quantitative trait locus (QTL) analysis or 63
genome-wide association studies (GWAS). New sequencing technology has vastly increased 64
the amount of available genetic information, and better phenotyping assays are now required 65
to meet the pace of these advancements. Depending on the research question, different 66
www.plantphysiol.orgon June 6, 2020 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

4
phenotyping approaches need to be taken, because there is a trade-off between (a) reflecting 67
realistic environments, (b) controlling environmental factors, (c) obtaining precise 68
information on individual parts of the root systems, and (d) throughput. 69
Several platforms to obtain root images from plants grown in soil (field, pots or rhizotrons) 70
are available, together with software to capture and model the overall size and shape of the 71
complex root systems (Lobet et al., 2011; Bucksch et al., 2014; Kalogiros et al., 2016). These 72
studies are complemented by analyses of roots developing on synthetic surfaces, which offer 73
more precise control and manipulation of the root environment. Open source software such as 74
BRAT (Slovak et al., 2014; Satbhai et al., 2017) and RootTrace (Naeem et al., 2011) have 75
been developed to precisely track the early main root growth from scanned images of 76
Arabidopsis thaliana seedlings growing on plates. An intermediate analysis space is occupied 77
by software such as EZ-Rhizo (Armengaud et al., 2009), archiDART (Delory et al. 2016, 78
2018), RootScape (Ristova et al., 2013), and the commercial WinRhizo (Arsenault et al., 79
1995). They have facilitated the quantitative assessment of more advanced root systems of 80
Arabidopsis and other dicot plants, including information on lateral root features such as 81
position, length, density and angle. This information has generated knowledge on branching 82
patterns, rather than just primary root growth or overall shape, and has enabled large-scale 83
phenotyping of Arabidopsis roots to investigate natural variation and responses to multiple 84
environmental cues (Gruber et al., 2013; Kellermeier et al., 2013, 2014; Julkowska et al., 85
2017). 86
With a good range of root image analysis software in place, the next challenge is how to 87
process the acquired numerical data in order to extract biologically meaningful and 88
statistically valid results. Averaging across replicates is easy for some simple RSA traits, 89
such as main root (MR) length and lateral root (LR) number, but it is less straightforward for 90
other traits. For example, how should we calculate the mean LR length if the number of LRs 91
differs between replicate plants? We can either average the length of all laterals in each plant 92
before averaging across plants, or we can average individual LRs across replicates. The latter 93
can be done either by the order of appearance (1st, 2
nd LR etc.) or by their position on the 94
main root (e.g. all LRs within a particular sector of the main root). There is no one ‘correct’ 95
solution to this problem; rather, each of the approaches is more or less suited to answer a 96
specific question. For example, the first average provides a measure of overall growth 97
investment into lateral roots, the second provides information on how individual LRs develop, 98
and the last approach best reflects the root shape. It is therefore important that RSA analysis 99
www.plantphysiol.orgon June 6, 2020 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

5
software, in addition to enabling data acquisition, also offers different options for extracting 100
mean root traits and their variance, which can then be used as a numerical input for genetic 101
studies or developmental models. 102
Another challenge is how to visually represent averaged root systems in a manner that clearly 103
and faithfully reflects differences between genotypes or environmental conditions. Current 104
root image analysis software packages rarely include options to visually reconstruct single-105
plant or mean RSAs from the measured data. In publications, the individual averaged RSA 106
traits are usually represented in multiple bar graphs. A picture of one ‘representative’ root 107
system is often included, but considering the variability and complexity of RSA it is unlikely 108
that one individual plant can indeed be representative in all RSA traits. The reader is left with 109
the difficult task of deducing the mean RSA from bar graphs of all individual traits. 110
To address these problems we have developed a new application, Root-VIS, which enables 111
calculation and visual reconstruction of mean RSAs from measured data. We exemplify the 112
possibilities offered by Root-VIS using data acquired with a greatly enhanced version of the 113
previously published EZ-Rhizo program. The combined EZ-Root-VIS pipeline provides an 114
easy procedure to capture RSA features of many individual plants and to visualize averaged 115
RSAs for different genotypes under various environments or at different time points. Root-116
VIS offers a range of choices for how to calculate means and how to visually represent the 117
average root phenotypes. For example, it allows users to calculate means of LR-related traits 118
on a per-plant or a per-branch basis. It provides possibilities to redraw average root 119
architectures from the extracted data or to regenerate root shapes in the form of flag plots, as 120
well as growth-investment or LR length profiles. We have validated the utility of the EZ-121
Root-VIS pipeline for studying root growth dynamics, quantitative genetics, and genotype-122
environment interactions. 123
124
RESULTS 125
Description of the EZ-Root-VIS pipeline 126
The software is a ‘standalone’ implementation, compatible with all Microsoft Windows 127
operating systems from Windows XP (SP-3) onwards, including Vista, Windows 7, 8/8.1 and 128
10 (in ‘Classic Desktop’ mode). Separate versions are available for 32-bit and 64-bit 129
platforms. The installation package includes Root-VIS and the latest version of the EZ-Rhizo 130
program, which has a number of additions and improvements over the original 2009 release 131
www.plantphysiol.orgon June 6, 2020 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

6
(Armengaud et al., 2009). For example, EZ-Rhizo now accepts input images in any of the 132
standard formats: Windows Bitmap (BMP) files, Graphics Interchange Format (GIF), Joint 133
Photographic Experts Group compressed images (JPEG), Portable Network Graphics (PNG) 134
or Tagged Image File Format (TIFF) files, and at any resolution. The software presents a 135
typical, Windows-style graphical user interface (GUI) (Supplemental Figure 1A) which will 136
be very familiar to users of the Microsoft®
Visual Studio development environment. 137
Extensive (and context-sensitive) on-line help is included with the software (Supplemental 138
Figure 1B). The software also provides numerous, user-selectable options for controlling the 139
content and style of displays, and the parameters used in the reconstruction and averaging 140
algorithms. These are adjustable via a “Settings” property sheet (Supplemental Figure 1C). 141
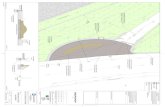
Figure 1A summarizes the workflow of RSA analysis with the EZ-Root-VIS pipeline. Images 142
of plants grown on agar plates in 2D are usually acquired with flatbed scanners (Figure 1B), 143
but any image capturing device that generates sufficient contrast and resolution is suitable. 144
First, EZ-Rhizo is used to process the images and extract numerical data of RSA features of 145
individual roots (Figure 1C). After image cropping, conversion into black and white and 146
background noise filtering, roots are pixelated, skeletonized, edited and detected as described 147
before (Armengaud et al., 2009). In the new version of EZ-Rhizo, we have included an option 148
to re-define the main root (the original version assumed the longest root path formed the MR 149
and this is also the default setting). The numerical results of the analysis are saved in a user-150
selectable database, which is now in XML format, allowing easy sharing of datasets between 151
the acquisition and analysis modules. Root-VIS then ‘reads’ the database generated by EZ-152
Rhizo and displays the files in a database window (left panel in Supplemental Figure 1A). 153
Prior to averaging RSA data from replicate roots, Root-VIS offers the possibility to draw a 154
visual reconstruction of any individual root in the dataset (Figure 1D), which allows the user 155
to verify the data that were captured by the image acquisition software. 156
At the heart of the Root-VIS software lies the concept of the ‘collection analysis’, in which 157
data extracted from the database are grouped according to the metadata parameters entered 158
during image analysis, including genotype, media type and plant age. Analyses can be carried 159
out with zero, one or two of these parameters being variants. The database structure displayed 160
in the analysis window makes it easy to remove or pool individual samples or groups of 161
samples from the analysis (Supplemental Figure 1B). The averaged root for each group can 162
then be displayed visually using a ‘reconstruction’ algorithm described in more detail in the 163
Methods section. Two types of averaged root systems can be generated: “absolute” and 164
www.plantphysiol.orgon June 6, 2020 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

7
“binned” (Figure 1E-F). In the ‘absolute’ mode, lateral roots are averaged according to their 165
order of appearance; in the ‘binned’ mode, the branched zone is divided into sectors (the 166
number of which is equal to the mean number of LRs), and the lengths of LRs in each sector 167
(‘bin’) are averaged. The program also provides an option to visualize the variation between 168
individual replicate roots in ‘alpha blends’ (Figure 1G). Additional visualization options are 169
‘flag plots’ (Figure 1H), which represent the overall density and shape of the root system, and 170
LR profiles (Figure 1I), which represent the size of LRs in different parts of the root system. 171
Flag plots are based on LR angle and LR density as well as the lengths of branched and un-172
branched zones of the MR as depicted in Supplemental Figure 2. For the LR profiles, the MR 173
length is divided into a user-defined number of sections, and for each section LR length is 174
displayed as a rectangle of appropriate size. Each of the display modes has a number of sub-175
options, which the user can select in the ‘Settings’ windows. For example, LR profiles can be 176
chosen to show either the sum length of all LRs (LR size, LRS) or the mean LR length (LRL) 177
in each MR sector. The former provides a measure of total growth investment into a certain 178
part of the root, whereas the latter reflects the local root system width. The different displays 179
are exemplified below using experimental data. We also generated an optional plug-in 180
module, XL-Orate, which allows users to create formatted tables of averaged trait data, 181
including standard errors and number of replicates using Microsoft® Excel (Figure 1J, 182
Supplemental Dataset 1), and to rapidly generate bar charts for any data generated with EZ-183
Rhizo and Root-VIS. The obtained datasets can now be used in a range of downstream 184
studies including growth models, quantitative genetics and breeding. 185
186
EZ-Root-VIS captures root growth dynamics 187
To validate the ability of the EZ-Root-VIS pipeline to capture root system growth, we 188
measured RSA of Arabidopsis Col-0 plants every two days over a period of 4 to 14 days after 189
germination (DAG). Quantitative data for 16 root traits (see Abbreviations) were obtained 190
using EZ-Rhizo. A 3D bar chart generated for three root traits (MRL, LRL, LRD-BZ) (Figure 191
2A) exemplifies the difficulty of visually representing the complexity of RSA changes in bar 192
graphs; several additional graphs would be required to represent the overall RSA. In contrast, 193
the visual root system reconstructions implemented in Root-VIS accommodate all the root 194
traits and represent averaged RSA (either ‘absolute’ or ‘binned’) as a single image (Figure 2B, 195
C). In this example, the display incorporated bends into the MR path to account for the 196
measured difference between MR path length and MR vector length (straightness). The 197
www.plantphysiol.orgon June 6, 2020 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

8
program accommodates any surplus of path length over vector length across equally sized 198
bends, which are placed at LR emergence sites in the branched zone and at equal distances 199
along the apical zone (see Methods). However, the user should keep in mind that EZ-Rhizo 200
does not determine the number, size and position of bends, and the displayed bending pattern 201
could look very different from the original images. To avoid over-interpretation of the 202
displayed bending pattern it might be safer to display either path or vector length as a straight 203
line. Therefore, in the ‘Settings’ property sheet, the user can toggle between displaying bends 204
(in the MR and/or LR), or not. 205
Figure 2D shows the superposition of all re-constructed individual roots as alpha blends, 206
reflecting the variation across replicates. A similar darkness of the blends indicated that the 207
variation of RSA between individual plants did not substantially increase with plant age. The 208
flag plots (Figure 2E) show how the root system increases in depth, width and density over 209
time. For this figure, flag plots were produced based on absolute values of MR and LR 210
lengths and angles, and on the slope of the linear regression of average LR lengths at the 211
given LR positions (see Supplemental Figure 2 and Methods). The shade of the flag fill color 212
indicates LR density with darker shades representing higher density. Finally, LRS profiles 213
(Figure 2F) allow the reader to quickly evaluate the growth investment into LRs in different 214
parts of the root. In this case, the MR was divided into four equal sections (quartiles). The 215
profiles show a sharp increase of mean LRS from 8 DAG onwards, especially in the two 216
basal quartiles of the main root. The data underpinning Figure 2, extracted using XL-Orate 217
(Supplemental Dataset 2), provide a convenient input for root growth models. 218
219
EZ-Root-VIS reveals genotype by environment interactions 220
To demonstrate the effectiveness of the pipeline in capturing genotype by environment 221
interactions, RSA data were acquired with EZ-Rhizo from replicate plants of nine 222
Arabidopsis accessions, grown in four nutrient media: control, low phosphate (P, 20 µM), 223
low potassium (K, 10 µM), or combined low PK media (Figure 3, Supplemental Dataset 3; 224
for growth media see Methods). The visual reconstructions (Figure 3) generated by Root-VIS 225
facilitate the rapid identification of interesting responses and RSA ideotypes, while Excel 226
generated bar graphs (Figure 4) depict means and standard errors of the individual traits. As 227
reported before (Chevalier et al., 2003; Reymond et al., 2006; Svistoonoff et al., 2007), the 228
Shahdara accession showed a strong inhibition of MR growth in response to P limitation. An 229
www.plantphysiol.orgon June 6, 2020 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

9
even stronger reduction of MR length was found for T1080 despite similar MR length on 230
control media. In most other accessions, e.g. Aitba-2 and Cerv-1, MR growth was not 231
sensitive to P availability, and in the Ber accession, MR growth was even stimulated by 232
decreasing the P supply (Figure 3A, Figure 4A). By contrast, in Ber, Aitba-2 and Cerv-1, low 233
P resulted in a reduction in the LR system size (LRS; measured as total lateral root path 234
length) (Figure 4B), particularly in the basal (upper) quartile of the main root (LRS (0.25), 235
Figure 3A, Figure 4D). Unlike low P, low K decreased MR length in all accessions tested, 236
albeit to varying extents, and caused a decrease of LRS in most accessions (Figure 3A). 237
Exceptions to the latter were Gie-0 and N13, which already had very small LRS in control 238
conditions. 239
The effects of combined P and K limitation were striking and in many cases could not have 240
been predicted from individual deficiency effects. In accessions experiencing MR inhibition 241
under low P (e.g. Shahdara, T1080), combined PK limitation had an additive inhibitory effect, 242
but in several other accessions (Gie-0, Ber, N13, Aitba-2) we recorded a complete or partial 243
reversal of the low K-induced MR inhibition by decreasing P (Figure 4A). In Cerv-1 and 244
Dog-4, MR inhibition by low K was maintained in low PK and dominated over MR 245
insensitivity to low P. In contrast to the genotype-dependent response of MR to single and 246
double deficiency, the LRS of all genotypes was smaller under combined PK limitation than 247
under single deficiencies (Figure 4B). LR density in the branched zone (LRD-BZ) was in 248
most accessions stable across treatments (Figure 4C) with the exception of Ber, N13 and 249
Aitba-2, which showed an increase of LRD-BZ under low K. To separate the effects of LR 250
number and LR length on total LRS, LRL profiles and flag plots were also plotted 251
(Supplemental Figure 3). Figure 4E shows the extracted data from these profiles as bar 252
graphs. 253
The re-constructed average RSAs (Figure 3B) pinpointed additional differences. For example, 254
MR growth was more or less vertical depending on genotype and environment. In control 255
medium, the MR angle (MRA) differed between accessions (Figure 4D), although the 256
direction was always the same (‘positive’, see Methods for sign definition). Under low P, 257
MRA remained similar to control in most accessions, but markedly increased in Ber, N13 and 258
Aitba-2, and decreased in T1080. Under low K, MRAs decreased in all accessions apart from 259
N13. In most accessions the low-K MRA phenotype was reverted back to control or to low-P 260
phenotypes in low PK, with the notable exception of Shahdara. 261
www.plantphysiol.orgon June 6, 2020 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

10
In summary, the EZ-Root-VIS pipeline enabled the analysis of complex responses of various 262
root architectural traits to either single or combined nutrient limitations. The averaged visual 263
representations allow the fast identification of nutrient interactions and of accessions that 264
show particularly interesting RSA responses. It is remarkable that the relatively small number 265
of accessions and nutrients tested here already encompass a diverse range of combinatorial 266
phenotypic outputs. 267
268
EZ-Root-VIS enables large scale RSA genetic studies 269
To validate the utility of our pipeline for large-scale quantitative genetic studies, we 270
measured RSA of 147 Arabidopsis accessions with EZ-Rhizo, and determined means of 16 271
root traits with Root-VIS for downstream analyses (Supplemental Dataset 4). Five plants per 272
accession were grown on standard medium, and it took one person 13 hours to analyze all 273
root images obtained at one time point (12 DAG) using the EZ-Root-VIS pipeline. 274
Ten of the analyzed root traits exhibited a normal distribution (Supplemental Figure 4), but 275
several traits showed a positively skewed trait distribution, including for example LR size and 276
number. Correlation analysis revealed a high degree of inter-dependence of the RSA traits 277
(Figure 5A). Only main root angle (MRA) and lateral root angle (LRA) were relatively 278
independent of other RSA traits, although they showed correlation with the size of MR and 279
LRs, respectively. Surprisingly, MR length was not correlated with the LR density over the 280
MR (LRD-MR) but was negatively correlated with LR density over the branched zone (LRD-281
BZ). This observation stresses the importance of determining LR density in the branched 282
zone rather than relating the number of LRs to the entire main root. Interestingly, LR density 283
was also negatively correlated with the length of the basal zone (BsZL). In fact, BsZL was 284
negatively correlated with most LR traits, suggesting that the amount of main root growth 285
before emergence of the first LR is an important determinant of other RSA features. 286
Hierarchical clustering was performed to classify the accessions into groups sharing similar 287
RSA based on 15 RSA architecture traits (LR angle was excluded from this analysis due to 288
some negative values). As shown in Figure 5B, the accessions were assigned into eight 289
clusters (Supplemental Dataset 4). Four clusters contained only one accession (Da(1)-12, 290
Lp2-2, N13 and Kn-0), while the largest cluster contained 73 accessions. Average LRS 291
profiles of the central genotype of each cluster (Figure 5C) indicated different growth 292
investment into MR and LR. Averaged RSA reconstructions revealed differences in MRA. 293
www.plantphysiol.orgon June 6, 2020 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

11
For example, the Sanna-2 and Mt-0 groups exhibited more positively skewed MRs than other 294
groups. Thus, when applied to natural variation studies, Root-VIS quickly identifies root 295
traits that are critical determinants of the overall RSA. 296
To test if the data extracted with EZ-Root-VIS can facilitate genetic mapping studies, we 297
performed GWA mapping for 16 root traits using an accelerated mixed model (AMM) 298
implemented in the GWAPP web application (Seren et al., 2012). In total, seven unique 299
locus-specific single nucleotide polymorphisms (SNPs) were found to be associated with 300
various root traits at 5% FDR (Supplemental Dataset 5). Out of these, one (SNP4 6374071) 301
was associated with TRS, LRS and LRS (0.25) (Figure 6A), three were associated with basal 302
zone length (BsZL) and one was associated with lateral root density (LRD-MR). 303
Forty-seven additional SNPs were identified to be associated with root traits with an adjusted 304
threshold of -log10(P) > 5.0 (Supplemental Dataset 5). One of them resembled SNP4 6374071 305
insofar as it showed association with LRS (0.25), LRS and TRS, suggesting a strong 306
contribution of the lateral root size in the basal quartile to the overall root size. We named the 307
two loci Lateral Root Size Locus (LRSL) and General Regulator Factor 1 (GRF1) (Figure 308
6A). LRS designates a 5 kb region that harbors four candidate genes (Supplemental Figure 309
5B). The GRF1 locus was named after the only gene, AT4G09000 (GRF1), present in a 5 kb 310
window around the strongest associated SNPs for this region (Supplemental Figure 5A). 311
Interestingly, GRF1 has been reported to be downregulated during LR emergence (Voß et al., 312
2015), and the expression of one of the candidates in LRSL, AT4G10270, is strongly induced 313
around the time of LR emergence and in emerged LRs (Supplemental Figure 5C-D). We 314
therefore investigated root phenotypes of Col-0 T-DNA insertion mutants for these two genes. 315
Indeed, the LRS profiles showed that knockout mutants of GRF1 or AT4G10270 exhibited 316
reductions in LRS (0.25), LRS and TRS compared to wild-type plants (Figure 6B), and the 317
extracted data confirmed that the differences were statistically significant (Figure 6C). 318
In addition, we characterized the root phenotypes of T-DNA insertion lines of candidate 319
genes underlying 18 other associations (4 significant, 14 between -log10(p) > 5.0 and 320
significant threshold) in the Col-0 or Col-3 backgrounds (Supplemental Dataset 5). In total, 321
for 14 out of the 21 associations, the mutants showed significant phenotypic differences from 322
wild type in root traits for which associations were initially identified, including MR angle, 323
MR length, LRD-BZ, LR number, BsZL, and apical ZL (AZL) (Supplemental Figure 6, 324
Supplemental Dataset 5). Some genes, such as ALF5 and AXR5, were found to contribute to 325
more RSA traits than initially highlighted by the GWAS, suggesting a more general role in 326
www.plantphysiol.orgon June 6, 2020 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

12
root growth and development. Data for all RSA traits in all lines are provided in 327
Supplemental Dataset 6. These results confirm that the EZ-Root-VIS pipeline is a useful tool 328
for fast gene discovery by high-throughput RSA phenotyping. 329
330
DISCUSSION 331
Root-VIS is novel for its ability to average RSA traits using several different algorithms and 332
to visually reconstruct the averaged RSA in several manners. During the development of the 333
software, great effort has been made to create an ergonomically efficient user interface, 334
which is a critical factor in determining the software’s ease of use. Both EZ-Rhizo and Root-335
VIS present a Windows-style graphical user interface (GUI) that will be easy to handle for 336
new users. Other features include numerous keyboard accelerators and the option to skip 337
parameter-setting dialogues using the ‘shift’ key, and an “Undo” command to avoid having to 338
restart processing following inadvertent errors. Such features may, at first, seem trivial but, 339
when considering that a typical experiment may involve processing several hundred images 340
per day, they can make an enormous difference to the overall user experience and greatly 341
reduce problems arising from ‘software fatigue’. 342
We have validated our pipeline to capture fairly complex RSA of 12-day-old Arabidopsis 343
plants in multiple contexts, including time series experiments, large scale natural variation 344
and genotype–environment interaction studies. We provide the options to extract means 345
based on order (‘absolute’) or local position of the lateral (‘binned’). It was noted that 346
depending on how much LR number varies between individual plants, the different averaging 347
methods can lead to about 10% variation in means determined for LR-related traits. Such 348
variation can be crucial when handling a large number of genotypes since the phenotypic 349
spectrum across accessions is continuous with relatively small differences between the 350
individual accessions. We do not consider one of the averaging methods to be superior to the 351
other; instead we provide the user with both options, allowing them to consider which 352
approach is particularly suitable to answer their questions. 353
We have exemplified the utility of the EZ-Root-VIS pipeline to represent the plastic 354
responses of various Arabidopsis accessions to environmental cues, with a particular focus on 355
LRS profiles. Indeed, this analysis helped us to identify extreme and interesting genotypes in 356
terms of their sensitivity to P or K limitation and combined deficiency. Under the low-P 357
conditions applied here none of the Arabidopsis accessions showed preferential growth 358
www.plantphysiol.orgon June 6, 2020 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

13
investment into upper (basal) lateral roots despite the widespread view that low-P adapted 359
RSA ideotypes preferentially explore residual P in the topsoil (Lynch and Brown, 2001). This 360
suggests that the accessions have not evolved in a P-limited environment, or that they only 361
extend laterals if they perceive an actual P gradient. Our pipeline also revealed that 362
interactive effect of K and P limitation modulate RSA in a genotype-specific manner that 363
could not have been predicted from measuring RSA under single deficiency or in one 364
accession only. 365
We have further validated the usefulness of the EZ-Root-VIS pipeline for facilitating GWAS 366
on RSA. A total of 56 loci controlling various root traits could be mapped at a cut-off 367
threshold of –log10(P) > 5.0. We adjusted the threshold to this level for two reasons: first, 368
SNPs in genes encoding known regulators affecting root traits such as IAA1/AXR5 and 369
ALF5 were associated with LRD-BZ and MRL with a –log10(P) > 5.0 and < 5% FDR 370
significance threshold, respectively. Second, some associations which are not significant at 5% 371
FDR, yet above –log10(P) > 5.0, have been shown to be false negatives, because multiple 372
testing correction methods are overly conservative (Kalladan et al., 2017; Meijón et al., 2013; 373
Müller et al., 2016). The vast majority of the loci identified here did not contain any 374
previously known RSA regulators. Indeed, the follow-up characterization of Col-0 knockout 375
mutants pointed to several genes with functions in RSA development, including AT4G09000 376
(GRF1), AT4G10270, AT4G10570 (UBP9), AT4G01550 (NTM2), AT3G29300, 377
AT5G51560 and AT3G04260 (PTAC3). We also found natural variation in genes that had 378
previously been shown to affect root traits such as AT4G14560 (IAA1), AT5G22300 (NIT4) 379
and AT3G23560 (ALF5) (Supplemental Datasets 5 and 6). These findings provide an 380
excellent starting point for understanding the genetics of previously unresolved traits such as 381
LRS (0.25) and MR angle, and the role of RSA in plant adaptation to their natural habitats. 382
Studying these traits can also provide insights into fundamental biological processes. For 383
example, MR angle studies are expected to advance our understanding of plagiogravitropism, 384
cell wall integrity (van der Does et al., 2017) and voltage-sensing (Kellermeier et al., 2014), 385
as Arabidopsis mutants impaired in cellulose biosynthesis or membrane depolarization show 386
altered MR angle. 387
Approaches such as GWAS become highly time and cost-effective if data for multiple traits 388
can be obtained from the same experiment. Using the EZ-Root-VIS pipeline, it is currently 389
possible to generate data for 20 root traits (more can be added) over a mapping population 390
comprising about 150 accessions in the course of a few days. While the throughput is lower 391
www.plantphysiol.orgon June 6, 2020 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

14
than for some other software packages (e.g. BRAT, Slovak et al., 2014) the RSAs quantified 392
by EZ-Root-VIS can be more complex and hence more traits can be extracted. Importantly, 393
the visual reconstructions of averaged root systems replace the need to rely on ‘representative’ 394
example pictures and enable fast integration of the measured individual RSA traits into 395
meaningful root system shapes, which can be considered a breakthrough in the field of plant 396
root biology. In publications, provision of averaged root system images rather than 397
representative images is critical for interpretation and follow-up studies and it may also 398
improve the reproducibility of results between various labs. Visual root shape reconstruction 399
can also aid breeding efforts aimed at improving plant resilience to soil-borne stresses, 400
facilitating the selection of desired root ideotypes from large phenotypic datasets. 401
EZ-Rhizo and Root-VIS have been developed over several years, and the versions released 402
with this paper have already been beta-tested by volunteer researchers. The software 403
therefore represents a community effort, and we will continue to make improvements in 404
response to suggestions and requests from the academic community. 405
406
MATERIAL AND METHODS 407
Software development 408
The EZ-Rhizo and Root-VIS applications, and their various plug-in modules (collectively 409
packaged as The Rhizo-II Root Biometrics Suite), were developed on, and for, the Microsoft®
410
Windows®
operating systems, in the C++ language, using Microsoft®
Visual Studio®
411
2010/2017. The graphical user interface (GUI) was implemented using objects derived from 412
the Microsoft Foundation Class (MFC) library and interaction with the XML database is built 413
on code from the CMarkup (V-11.5) C++ XML Parser (www.firstobject.com). The software 414
package can be downloaded from http://www.psrg.org.uk/download/Rhizo-64.msi (for 64-bit 415
platforms) or http://www.psrg.org.uk/download/Rhizo-32.msi (for 32-bit platforms); it is free 416
of charge but we ask users to register their installation. 417
The XL-Orate plug-in is an ‘application extension module’ (dynamic link library), which is 418
activated via the “Plug-Ins” command in the “File” menu. It was also written in C++ and 419
developed using the same tools and coding strategy as for the main programs. It requires 420
installation of the Microsoft Excel software on the user’s computer and interacts with Excel 421
directly (without user involvement) through programmed automation. 422
www.plantphysiol.orgon June 6, 2020 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

15
Root-VIS was developed to be used in conjunction with EZ-Rhizo. Nevertheless, in principle, 423
any dataset can be analysed with Root-VIS so long as it is presented in the correct format and 424
contains all the data required for the visual reconstructions. To facilitate data input from other 425
sources Root-VIS is already compatible with RSML (Lobet et al., 2015), and procedures for 426
enabling Root-VIS to read those data files are currently under development. 427
Visual reconstructions 428
The Root-VIS program uses a subtle, smart algorithm to reconstruct visually representative 429
forms, (optionally) adding ‘bends’ to roots based on the available information (the length to 430
vector ratio, and the number, position and orientation of lateral roots). We have implemented 431
five types of average visual reconstructions of root systems in Root-VIS, named Absolute 432
Averages, Binned Averages, Alpha Blends, Flag Plots, and LR Profiles. The information 433
available from the database, for a main root, comprises its path length (the total length of the 434
root, from base to apex), its vector length (the straight-line distance between base and apex), 435
the angle of its vector from vertical (positive angles being clockwise when looking at the root 436
from the back of the plate) and the number of lateral roots. Lengths are expressed in 437
centimeters and angles are in degrees. For a lateral root, the information also includes its 438
position (the distance along its parent root from its parent’s base). 439
Adding bending patterns 440
Roots are usually not straight and therefore any visualization has to accommodate the 441
difference between path length and vector length. However, the precise bending patterns are 442
currently not quantified by EZ-Rhizo. We have implemented different visualization options 443
in Root-VIS consisting of either straight lines representing path or vector length, or added 444
bends. For roots that have no sub-root, bends can be added in a pre-decided number, equally 445
spaced along the vector’s axis, with the displacement of each point from the axis being 446
calculated by the application of Pythagoras’ Theorem. However, for roots with one or more 447
sub-roots, a correlation has been demonstrated between the bending of a root and the 448
emergence of a lateral root and that lateral roots will preferentially form at bends and follow 449
an alternating directional pattern (Laskowski et al., 2008). We therefore decided to place 450
bends along the vector, not at equal intervals, but rather at points corresponding to the 451
position of each lateral root, with the direction of the off-axis displacement being determined 452
by the sign of the branch angle. If two consecutive lateral roots branch in the same direction, 453
Root-VIS adds an extra bend point between them, being displaced in the direction opposite to 454
www.plantphysiol.orgon June 6, 2020 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

16
that of the two real sub-roots. Finally, we need to allow for a degree of bending in the apical 455
part of the root: for this, we distribute the total amount of added ‘bend’ between the branched 456
and apical zones according to the relative lengths of these two root segments, and allow the 457
user to specify how many (equally-spaced) bends to add in the apical zone. Clearly, the 458
described procedure does not recreate the original bending pattern but applies existing 459
evidence and user’s choices to fill in missing information. While the displayed bending 460
pattern will not be faithful, the approach allowed us to correctly display both path and vector 461
length in one picture. 462
Averaged RSA; absolute and binned modes 463
The Root-VIS program provides two different methods for calculating the averaged RSA of a 464
given dataset, which we call “absolute averaging” and “binned averaging.” For absolute 465
averages, first, the mean parameters (length, vector and angle) for the main root are 466
determined by dividing the totals for all plants by the number of samples; for the angle 467
parameter, averages are formed using the absolute values (i.e. not including the sign) and the 468
modal sign is then given to the calculated mean. Next, the median number of lateral roots is 469
calculated. Then, for each lateral root, in order of appearance, their means are calculated (in 470
the same way as for the main root, with the addition of the position parameter). If a plant in 471
the dataset has less than the median number of lateral roots, zeros are given as the four 472
parameter values for any ‘missing’ roots, and the divisor for the parameter totals for those 473
lateral roots is reduced by one. Since LRs are numbered from base to apex in EZ-Rhizo any 474
‘missing’ lateral roots will always be at the tip of the parent root. 475
For the binned averaging algorithm, first, each main root within the dataset is split into three 476
sections: the basal zone, the branched zone and the apical zone as described (Armengaud et 477
al., 2009). The path and vector lengths are then established for each of these zones and 478
accumulated across all of the roots, and the means calculated. By adding each mean, the total 479
mean path length and mean vector length of the root are determined. Next, the mean angle 480
and median number of laterals are calculated as for absolute averages. However, to calculate 481
the mean parameters for each lateral root, we first split the branched zone into a number of 482
sections – or bins – equal to the mean number of lateral roots. The lateral roots for each 483
sample in the dataset are allocated to one of these bins according to their position. The mean 484
values for LR length, vector and angle are then calculated for each bin (as before) and for 485
visualization they are positioned at the midpoint of the bin section. This averaging algorithm 486
takes into account that the order of appearance does not necessarily reflect where a LR is 487
www.plantphysiol.orgon June 6, 2020 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

17
placed; for example, the 4th
lateral root in one plant could be located near the top of the main 488
root whereas in another it could be positioned in the middle. In the visual reconstruction the 489
representative average LR is placed at the halfway point of the respective parent root segment. 490
Alpha blends 491
The RSA reconstructions do not include an indication of the amount of variance between 492
samples in each dataset (although standard errors are included in the numerical output). The 493
alpha blend option allows the reconstructions of the individual roots to be overlaid in a semi-494
transparent fashion, thus indicating the degree of variance across the dataset. By default, 495
when creating alpha blends, the vector angles of the main roots are normalised to the average 496
vector angle for the samples included. Additional user-selectable options also allow for the 497
MR path lengths and/or vector lengths to be normalised. 498
Flag plots 499
The various components of a flag plot are indicated in Supplemental Figure 2; the flagpole is 500
divided into three sections, the lengths of which are proportional to the lengths of each of the 501
root zones (basal, branched and apical); the angles between the upper and lower edges of the 502
flag and the pole represent the mean insertion angles of the LRs (upper edge) and the slope of 503
the linear regression between the mean length of the LRs and their position on the main 504
root. Alternatively, the length of the upper edge can represent the average length of the LRs, 505
in which case the angle between the lower edge and the flagpole will not represent any 506
directly measured RSA data. The density of the flag’s fill color represents the average lateral 507
root density – the value of which is normalized (i.e. scaled to a value between 0 and 1) within 508
the given set of plots. 509
The software provides a number of options to control how to draw flag plots, including the 510
selections described above, selections for root path or vector data, and choice of fill color. 511
LR profiles 512
The fact that EZ-Rhizo determines the position of each LR allows simple visualization of 513
local differences in LR growth. To construct the LR profiles, the MR is divided into a user-514
defined number of sectors (2-10) and the LR data within each sector is represented as a 515
rectangle over the sector. The width of the rectangle can be chosen to represent either the 516
total LR size (added lengths of al LRs in the sector, LRS profile) or the mean length of the 517
LRs in the sector (LRL profile). 518
www.plantphysiol.orgon June 6, 2020 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

18
Plant material, growth conditions, and root image processing 519
Arabidopsis (Arabidopsis thaliana) accessions from the RegMap panel (Horton et al., 2012), 520
and homozygous T-DNA insertion mutant lines for candidate genes and their corresponding 521
wild types, Col-0 or Col-3, were obtained from the Nottingham Arabidopsis Stock Centre. 522
Seeds were surface sterilized in absolute ethanol for 1 min and then washed for 5 min with a 523
solution containing 2.8% (w/v) sodium hypochlorite and 0.1% (v/v) Tween®
-20 followed by 524
five washes with sterile distilled water. Seeds were incubated at 4°C for 5-7 days. The 525
sterilized and stratified seeds were sown on vertical plates with minimal media (Kellermeier 526
et al., 2014) containing 1% (w/v) agar (Formedium), 0.5% (w/v) sucrose and pH 5.6 (0.2 M 527
MES/Tris). For nutrient limitation experiments concentrations of K and P were lowered to 10 528
µM and 20 µM, respectively, and the osmotic potential of these media was adjusted as 529
described before (Kellermeier et al., 2014). Plants were a grown in a growth chamber at 60% 530
relative humidity and 20°C, with 9 h of light (120 µE m-2
s-1
). Images of each plate were 531
acquired using a flatbed scanner at 200 dpi as previously described (Kellermeier and 532
Amtmann, 2013). The RSA of each plant was quantified from the images using the EZ-Rhizo 533
as described before (Armengaud et al., 2009), facilitated by some new software functions 534
described in the Help files of the latest version of EZ-Rhizo package released with this 535
publication. 536
Statistical tests 537
Averaged root trait data obtained from Root-VIS using the XL-Orate plugin were used for 538
statistical analyses unless otherwise stated. Trait frequency distribution and Shapiro-Wilk test 539
of uniformity, correlation and agglomerative hierarchical clustering analyses were performed 540
using XLSTAT software. Genome-wide association mapping was conducted using GWAPP 541
web interface (https://gwas.gmi.oeaw.ac.at/) with 250K SNP data and the accelerated mixed-542
model (AMM) algorithm method (Seren et al., 2012). Statistically significant differences 543
between wild-type and mutant plants was assessed using a Student’s t test (*: p < 0.05) using 544
Microsoft®
Excel. 545
Accession numbers 546
Sequence data from this article can be found in the Arabidopsis Genome Initiative or 547
GenBank/EMBL databases under the following accession numbers: AT4G09000 (GRF1), 548
AT4G10270 (in locus LRS), AT4G10570 (UBP9), AT4G01550 (NTM2), AT3G29300 (in 549
www.plantphysiol.orgon June 6, 2020 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

19
locus LRDB), AT5G51560 (in locus LRK-LRN), AT3G04260 (PTAC3), AT4G14560 (IAA1), 550
AT5G22300 (NIT4), AT3G23560 (ALF5). See also Supplemental Data Sets 5 and 6. 551
Supplemental Data Files 552
The following materials are available in the online version of this article. 553
Supplemental Data Set 1: Numerical data for average RSA traits, number of replicate plants 554
and standard error underpinning the roots shown in Figure 1 E, F. 555
Supplemental Data Set 2: Numerical data for average RSA traits, number of replicate plants 556
and standard error underpinning the roots shown in Figure 2. 557
Supplemental Data Set 3: Means and SE of RSA traits of different accessions in four 558
nutrient conditions (data underpinning Figures 3, 4 and Supplemental Figure 3) 559
Supplemental Data Set 4: Means of RSA traits of 147 Arabidopsis accessions under control 560
conditions and hierarchical clustering of accessions based on root traits 561
Supplemental Data Set 5: Description of the GWAS data set. 562
Supplemental Data Set 6: Means and standard errors of root system architecture traits of 563
mutants of candidate genes identified through GWAS (data underpinning Figure 6 and 564
Supplemental Figure 6). 565
Supplemental Figures: Supplemental Figures 1-6 with legends, including: 566
Supplemental Figure 1. Screenshots of the Root-VIS software 567
Supplemental Figure 2. Visual explanation of the data incorporated into a Root-VIS flag 568
plot. 569
Supplemental Figure 3. Root-VIS representations of average RSAs of nine Arabidopsis 570
accessions in four different nutrient conditions. 571
Supplemental Figure 4. Frequency distribution of RS traits in Arabidopsis accessions. 572
Supplemental Figure 5. Genes and transcript profiles for two RSA associations. 573
Supplemental Figure 6. Candidate gene validation 574
Abbreviations 575
MRL Main root path length
MRV Main root vector length
MRA Main root angle
BsZL Basal zone length (from hypocotyl to the first lateral root)
BZL Branched zone length
AZL Apical zone length
TRS Total root system
LRS Total lateral root path length
LRS (0.25) Total lateral root path length in the top quartile of main root
www.plantphysiol.orgon June 6, 2020 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

20
LRL Average lateral root path length
LRL (0.25) Average lateral root path length in the upper quartile of main root
LRN Number of lateral roots
LRD-MR Lateral root density over the entire main root path (LRN/MRL)
LRD-BZ Lateral root density in the branched zone of the main root (LRN/BZL)
LRA Lateral root angle
OAD Overall depth
576
ACKNOWLEDGEMENTS 577
We are grateful to Geraldine Goldie who carried out preliminary consultation and 578
programming as part of a student project at the University of Glasgow. We are also grateful 579
to Mike Blatt for his support in the development of this software. The work was funded by 580
the BBSRC (BB/N018508/1) and by the Gatsby Charitable Trust (Sainsbury studentship to 581
FK). 582
583
584
www.plantphysiol.orgon June 6, 2020 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

21
FIGURE LEGENDS 585
Figure 1. Overview of EZ-Root-VIS pipeline 586
(A) EZ-Rhizo and Root-VIS software packages provide a convenient analysis pipeline 587
transforming root images into a numerical and statistical data output describing root system 588
architecture. 589
(B) A suitable image of Arabidopsis plants growing on a vertical agar plate can be acquired 590
with a flatbed scanner at 200 dpi. 591
(C) Skeletonized roots obtained after image processing with EZ-Rhizo provide the basis for 592
quantification of RSA features by EZ-Rhizo. The obtained data is saved in a searchable 593
database. 594
(D) Root-VIS re-constructs the individual roots using the data extracted by EZ-Rhizo. 595
(E-J) Root-VIS generates visual reconstructions of root system architecture from a user-596
defined set of individual roots (replicates), including absolute (E) and binned (F) average 597
RSA, alpha blends (G), flag plots (H) and LR profiles (I). The examples shown are based on 598
data from the five individual Arabidopsis plants shown in B. Roots images were taken at 12 599
DAG. 600
(J) For subsequent generation of graphs, statistical analyses and modelling the numerical data 601
obtained with Root-VIS is saved and can be displayed in Excel using the XL-Orate plug-in. 602
The screenshot shows an example Excel output from a large dataset. Data for each RSA 603
parameter is provided in a separate datasheet. MRL: Main root path length, LRN: lateral root 604
number, MRA: Main root angle, MRV: Main root vector length. 605
606
Figure 2. Visual representation of RSA development over time 607
(A) Change of selected RSA features over time displayed in a conventional bar graph. Plotted 608
are plant age in days after germination (x-axis), mean lateral root density over the branched 609
zone (LRD-BZ, y-axis), mean main root length (MRL, z-axis), and mean lateral root length 610
(LRL, color) of Arabidopsis Col-0 plants growing on control media. Numbers of roots 611
analyzed at each time point are given in (B). 612
(B-F) Root-VIS offers several options to visualize the RSA of many replicate plants; 613
including absolute (B) and binned (C) average RSA reconstructions, superimposition of 614
normalized RSAs in alpha blends (D) as well as shape reconstructions in the form of flag 615
plots (E) and LRS profiles (F). All Root-VIS reconstructions shown were based on EZ-Rhizo 616
data obtained from Arabidopsis Col-0 plants grown in control conditions. Plant age (in days, 617
d) and numbers of replicate roots (n) are given in (B). 618
www.plantphysiol.orgon June 6, 2020 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

22
Figure 3. Environment- and genotype-dependent growth investment into main root and 619
lateral roots 620
Lateral root size (LRS) profiles (A) and average RSA reconstruction (B) of Arabidopsis 621
accessions grown in control, low K, low P, and combined low P and low K (low PK) media. 622
Vertical lines represent the mean main root path length. The main root path was divided into 623
four equally sized sections and lengths of laterals were added within each section. The width 624
of each square represents the mean LRS of 3-6 replicate plants. 625
Figure 4. RSA traits of Arabidopsis accessions grown in different nutrient conditions 626
(A-E) Means and standard errors of selected RSA features of 12-day-old Arabidopsis 627
accessions grown in control (black bars), low K (blue bars), low P (red bars), or low PK 628
(green bars) media. The data was generated by EZ-Rhizo and means across replicates (n = 3-629
6 plants) were calculated with Root-VIS for (A) main root length (MRL), (B) total lateral root 630
size (LRS), (C) lateral root density over the branched zone (LRD-BZ), and (D) main root 631
angle (MRA). (E) Means and standard errors of lateral root length (LRL) within four sections 632
of the main root. Data were extracted from LRL profiles generated by Root-VIS and 633
transferred to Excel using the XL-Orate plug-in. 634
Figure 5. Use of the EZ-Root-VIS pipeline for large-scale natural variation studies 635
(A) Heatmap of pairwise correlations (Pearson correlation coefficient) of 16 RSA traits in 636
147 Arabidopsis accessions. Plants were grown in control conditions. The correlations 637
between different root features were calculated using means of 5 plants for each genotype. 638
Stars indicate significance at P < 0.05 (Student’s t test). 639
(B) Hierarchical clustering of Arabidopsis accessions by genotype. Calculation of similarity 640
was based on 15 mean RSA traits calculated by Root-VIS. See Supplemental Data Set 5 for 641
the class assignment of the individual accessions. 642
(C) Visual reconstructions of RSA and LRS profiles of the central accession of each cluster. 643
Accession names are colored according to (B). 644
Figure 6. Genome-wide association studies for root system architecture traits 645
(A) Manhattan plots for GWA mapping of three RSA traits; lateral root size over the basal 646
quartile of the main root (LRS (0.25)), total lateral root size (LRS), and total root size (TRS). 647
The horizontal dotted line corresponds to a 5% FDR threshold. Light blue ticks labeled 648
‘GRF1’ and ‘LRSL’ indicate the location of the most significant associations. 649
(B) LRS profiles of Arabidopsis Col-0 wild type and mutant lines. Plants were grown on 650
control media in three independent experiments. The total number of replicate roots 651
contributing to each LRS profile is given in brackets. 652
www.plantphysiol.orgon June 6, 2020 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

23
(C) Means and standard errors (n as shown in B) of some RSA traits in wild type Col-0 and 653
mutant lines. Significant differences to wild type are indicated by asterisks (P <0.05, 654
Student’s t test). 655
656
www.plantphysiol.orgon June 6, 2020 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

24
REFERENCES 657
Armengaud, P., Zambaux, K., Hills, A., Sulpice, R., Pattison, R.J., Blatt, M.R., and 658
Amtmann, A. (2009). EZ-Rhizo: Integrated software for the fast and accurate 659
measurement of root system architecture. Plant J 57: 945–956. 660
Arsenault, J., Pouleru, S., Messier, C., and Guay, R. (1995). WinRHlZOTM
, a Root-661
measuring system with a unique overlap correction method. HortScience 30: 906. 662
Bucksch, A., Burridge, J., York, L.M., Das, A., Nord, E., Weitz, J.S., and Lynch, J.P. 663
(2014). Image-based high-throughput field phenotyping of crop roots. Plant Physiol 166: 664
470–486. 665
Chevalier, F., Pata, M., Nacry, P., Doumas, P., and Rossignol, M. (2003). Effects of 666
phosphate availability on the root system architecture: large-scale analysis of the natural 667
variation between Arabidopsis accessions. Plant Cell Environ 26: 1839–1850. 668
Delory, B.M., Li, M., Topp, C.N., Lobet, G. (2018) archiDART v3.0: A new data analysis 669
pipeline allowing the topological analysis of plant root systems. F1000Res. 2018;7. 670
Delory, B.M., Baudson, C., Brostaux, Y., Lobet, G., du Jardin, P., Pagès, L., et al. (2016). 671
archiDART: an R package for the automated computation of plant root architectural 672
traits. Plant Soil 398: 351–365. 673
Gruber, B.D., Giehl, R.F.H., Friedel, S., and von Wirén, N. (2013). Plasticity of the 674
Arabidopsis root system under nutrient deficiencies. Plant Physiol 163: 161–79. 675
Horton, M.W. et al. (2012). Genome-wide patterns of genetic variation in worldwide 676
Arabidopsis thaliana accessions from the RegMap panel. Nat Genet 44: 212–216. 677
Julkowska, M.M. and Testerink, C.(2015). Tuning plant signaling and growth to survive 678
salt. Trends Plant Sci 20: 586–594. 679
Julkowska, M., Koevoets, I.T., Mol, S., Hoefsloot, H.C., Feron, R., Tester, M., 680
Keurentjes, J.J.B., Korte, A., Haring, M.A., de Boer, G.-J., and Testerink, C. (2017). 681
Genetic Components of Root Architecture Remodeling in Response to Salt Stress. Plant 682
Cell doi:10.1105/tpc.16.00680. 683
Kalladan, R., Lasky, J.R., Chang, T.Z., Sharma, S., Juenger, T.E., and Verslues, P.E. 684
(2017). Natural variation identifies genes affecting drought- induced abscisic acid 685
accumulation in Arabidopsis thaliana. PNAS 114: 11536-11541. 686
www.plantphysiol.orgon June 6, 2020 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

25
Kalogiros, D.I., Adu, M.O., White, P.J., Broadley, M.R., Draye, X., Ptashnyk, M., 687
Bengough, A.G., and Dupuy, L.X. (2016). Analysis of root growth from a phenotyping 688
data set using a density-based model. J Exp Bot 67: 1045–1058. 689
Kellermeier, F. and Amtmann, A. (2013). Phenotyping jasmonate regulation of root growth. 690
Methods Mol Biol 1011: 25–32. 691
Kellermeier, F., Armengaud, P., Seditas, T.J., Danku, J., Salt, D.E., and Amtmann, A. 692
(2014). Analysis of the root system architecture of Arabidopsis provides a quantitative 693
readout of crosstalk between nutritional signals. Plant Cell 26: 1480–1496. 694
Kellermeier, F., Chardon, F., and Amtmann, A. (2013). Natural variation of Arabidopsis 695
root architecture reveals complementing adaptive strategies to potassium starvation. 696
Plant Physiol 161: 1421–1432. 697
Laskowski, M., Grieneisen, V.A., Hofhuis, H., Ten Hove, C.A., Hogeweg, P., Marée, 698
A.F.M., and Scheres, B. (2008). Root system architecture from coupling cell shape to 699
auxin transport. PLoS Biol 6: 2721–2735. 700
Lobet, G., Pagès, L., and Draye, X. (2011). A Novel image-analysis toolbox enabling 701
quantitative analysis of root system architecture. Plant Physiol 157: 29–39. 702
Lobet G., Pound M.P., Diener J. et al. (2015) Root System Markup Language: toward an 703
unified root architecture description language. Plant Physiol 167:617–627. 704
Lynch, J.P. (2015) Root phenes that reduce the metabolic costs of soil exploration: 705
opportunities for 21st century agriculture. Plant Cell Environ 38:1775–1784. 706
Lynch, J.P. and Brown, K.M. (2001). Topsoil foraging - An architectural adaptation of 707
plants to low phosphorus availability. Plant Soil 237: 225–237. 708
Meijón, M., Satbhai, S.B., Tsuchimatsu, T., and Busch, W. (2013). Genome-wide 709
association study using cellular traits identifies a new regulator of root development in 710
Arabidopsis. Nat Genet 46: 77–81. 711
Morris, E.C., Griffiths, M., Golebiowska, A. , Mairhofer, S., Burr-Hersey, J. , Goh, T. , 712
von Wangenheim, D., Atkinson, B. , Sturrock, C. J. , Lynch, J.P. , Vissenberg, K., 713
Ritz, K. , Wells, D. M. , Mooney, S. J. , Bennett, M. J. (2017) Shaping 3D root system 714
architecture. Curr Biol 27: 919-930 715
Müller, L.M., Lindner, H., Pires, N.D., Gagliardini, V., and Grossniklaus, U. (2016). A 716
www.plantphysiol.orgon June 6, 2020 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

26
subunit of the oligosaccharyltransferase complex is required for interspecific 717
gametophyte recognition in Arabidopsis. Nat Commun 7: 10826 doi: 718
10.1038/ncomms10826. 719
Naeem, A., French, A.P., Wells, D.M., and Pridmore, T.P. (2011). High-throughput 720
feature counting and measurement of roots. Bioinformatics 27: 1337–1338. 721
Reymond, M., Svistoonoff, S., Loudet, O., Nussaume, L., and Desnos, T. (2006). 722
Identification of QTL controlling root growth response to phosphate starvation in 723
Arabidopsis thaliana. Plant Cell Environ 29: 115–125. 724
Ristova, D., Rosas, U., Krouk, G., Ruffel, S., Birnbaum, K.D., and Corruzi, G. (2013). 725
RootScape: landmark-based system for rapid screening of root architecture in 726
Arabidopsis. Plant Physiol 161: 1086–1096. 727
Rogers E.D., Benfey P.N. (2015) Regulation of plant root system architecture: implications 728
for crop advancement. Curr Opin Biotechnol 32: 93–98. 729
Satbhai, S.B., Setzer, C., Freynschlag, F., Slovak, R., Kerdaffrec, E., and Busch, W. 730
(2017). Natural allelic variation of FRO2 modulates Arabidopsis root growth under iron 731
deficiency. Nat Commun 8: 15603 doi: 10.1038/ncomms15603. 732
Seren, U., Vilhjalmsson, B.J., Horton, M.W., Meng, D., Forai, P., Huang, Y.S., Long, Q., 733
Segura, V., and Nordborg, M. (2012). GWAPP: a web application for genome-wide 734
association mapping in Arabidopsis. Plant Cell 24: 4793–4805. 735
Shahzad, Z. and Amtmann, A. (2017). Food for thought : how nutrients regulate root 736
system architecture. Curr Opin Plant Biol 39: 80–87. 737
Slovak, R., Su, X., Shimotani, K., Shiina, T., and Busch, W. (2014). A scalable open-738
source pipeline for large-scale root phenotyping of Arabidopsis. Plant Cell 26: 2390–739
2403. 740
Svistoonoff, S., Creff, A., Reymond, M., Sigoillot-Claude, C., Ricaud, L., Blanchet, A., 741
Nussaume, L., and Desnos, T. (2007). Root tip contact with low-phosphate media 742
reprograms plant root architecture. Nat Genet 39: 792–796. 743
Van der Does, D., Boutrot, F., Engelsdorf, T., Rhodes, J., Mckenna, F., Vernhettes, S., et 744
al. (2017). The Arabidopsis leucine-rich repeat receptor kinase MIK2 / LRR-KISS 745
connects cell wall integrity sensing, root growth and response to abiotic and biotic 746
stresses. PLoS Genet 13:e1006832. 747
www.plantphysiol.orgon June 6, 2020 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

27
Voß, U. et al. (2015). The circadian clock rephases during lateral root organ initiation in 748
Arabidopsis thaliana. Nat Commun 6: 7641 doi: 10.1038/ncomms8641. 749
Yu, P. , Gutjahr, C., Li, C.J. , Hochholdinger, F. (2016) Genetic control of lateral root 750
formation in cereals. Trends Plant Sci 21: 951-961 751
752
753
www.plantphysiol.orgon June 6, 2020 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

1
Figure 1. Overview of EZ-Root-VIS pipeline
(A) EZ-Rhizo and Root-VIS software packages provide a convenient analysis pipeline transforming root
images into a numerical and statistical data output describing root system architecture.
(B) A suitable image of Arabidopsis plants growing on a vertical agar plate can be acquired with a flatbed
scanner at 200 dpi.
(C) Skeletonized roots obtained after image processing with EZ-Rhizo provide the basis for quantification of
RSA features by EZ-Rhizo. The obtained data is saved in a searchable database.
(D) Root-VIS re-constructs the individual roots using the data extracted by EZ-Rhizo.
(E-J) Root-VIS generates visual reconstructions of root system architecture from a user-defined set of
individual roots (replicates), including absolute (E) and binned (F) average RSA, alpha blends (G), flag plots
(H) and LR profiles (I). The examples shown are based on data from the five individual Arabidopsis
plants shown in B. Roots images were taken at 12days after germination.. (J) For subsequent generation of graphs, statistical analyses and modelling the numerical data obtained with
Root-VIS is saved and can be displayed in Excel using the XL-Orate plug-in. Note that the screenshot shown
was extracted from a much larger dataset than the example shown in A-E. Data for each RSA parameter is
provided in a separate datasheet. MRL: Main root path length, LRN: lateral root number, MRA: Main root
angle, MRV: Main root vector length.
Image acquisition
Visual reconstruction
Create analysis
Export image/analysis
Root-VIS
Import image
Detect roots
Redefine main root
Save analysis
EZ-Rhizo
Image treatment
Downstream studies
(modelling, quantitative
genetics, physiology, breeding)
Image file .jpg, .bmp, …
Database XML
Data file .txt, .xlsx
A B
C
D
E F G H I
J
www.plantphysiol.orgon June 6, 2020 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

1
Figure 2. Visual representation of RSA development over time
(A) Change of selected RSA features over time displayed in a conventional bar graph. Plotted are plant age in days after
germination (x-axis), mean lateral root density over the branched zone (LRD-BZ, y-axis), mean main root length (MRL,
z-axis), and mean lateral root length (LRL, color) of A. thaliana Col-0 plants growing on control media. Numbers of
roots analyzed at each time point are given in (B).
(B-F) Root-VIS offers several options to visualize the RSA of many replicate plants; including absolute (B) and binned
(C) average RSA reconstructions, superimposition of normalized RSAs in alpha blends (D) as well as shape
reconstructions in the form of flag plots (E) and LRS profiles (F). All Root-VIS reconstructions shown were based on
EZ-Rhizo data obtained from A. thaliana Col-0 plants growing in control conditions. Plant age (in days, d) and numbers
of replicate roots (n) are given in (B).
4 d
(n=6)
Ab
s. A
vg.
B 6 d
(n=27)
8 d
(n=58)
10 d
(n=61)
12 d
(n=63)
14 d
(n=52)
Bin
. A
vg.
C
α-B
len
dF
lag P
lot
D
E
LR
S p
rofile
F
LRD-BZ
LRL (cm)
A
0.78
0.46
0.37
0.20
0.11
0.06
12
34
5
7
6
8
www.plantphysiol.orgon June 6, 2020 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

1
Figure 3. Environment- and genotype-dependent growth investment into main root and
lateral roots
Lateral root size (LRS) profiles (A) and average RSA reconstruction (B) of Arabidopsis
accessions grown in control, low K, low P, and combined low P and low K (low PK) media.
Vertical lines represent the mean main root path length. The main root path was divided into
four equally-sized sections and lengths of laterals were added within each section. The width
of each square represents the mean LRS of 3-6 replicate plants.
Control
Low K
Low P
Low PK
Shahdara T1080 Cerv-1 Dog-4 Wa-1 Gie-0 Ber N13 Aitba-2
Control
Low K
Low P
Low PK
B
A
www.plantphysiol.orgon June 6, 2020 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

1
Figure 4. RSA traits of Arabidopsis accessions grown in different nutrient conditions
(A-E) Means and standard errors of selected RSA features of 12-d old Arabidopsis accessions
grown in control (black bars), low K (blue bars), low P (red bars), or low PK (green bars)
media. The data was generated by EZ-Rhizo and means across replicates (n = 3-6 plants)
were calculated with Root-VIS for (A) main root length (MRL), (B) total lateral root size
(LRS), (C) lateral root density over the branched zone (LRD-BZ), (D) main root angle
(MRA). (E) Means and standard errors of lateral root length (LRL) within four sections of the
main root. Data were extracted from LRL profiles generated by Root-VIS and transferred to
Excel using the XL-Orate plug-in.
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
1.8
2
Contr
ol
Low
K
Low
P
Low
PK
Contr
ol
Low
K
Low
P
Low
PK
Contr
ol
Low
K
Low
P
Low
PK
Contr
ol
Low
K
Low
P
Low
PK
Contr
ol
Low
K
Low
P
Low
PK
Contr
ol
Low
K
Low
P
Low
PK
Contr
ol
Low
K
Low
P
Low
PK
Contr
ol
Low
K
Low
P
Low
PK
Contr
ol
Low
K
Low
P
Low
PK
Shahdara T1080 Cerv-1 Dog-4 Wa-1 Gie-0 Ber N13 Aitba-2
LR
L (
cm
)
E
0
2
4
6
8
10
02468
101214
LR
S (
cm
)
MR
L (
cm
)
0
5
10
15
20
LR
D-B
Z (
cm
-1)
A
C
B
D
-5
0
5
10
15
20
25
30
35
40
MR
A (
o)
Section 1Section 2Section 3
C K P PK
www.plantphysiol.orgon June 6, 2020 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

1
Figure 5. Use of the EZ-Root-VIS pipeline for large-scale natural variation studies
(A) Heatmap of pairwise correlations (Pearson correlation coefficient) of 16 RSA traits in
147 A. thaliana accessions. Plants were grown in control conditions. The correlations
between different root features were calculated using means of 5 plants for each genotype.
Stars indicate significance at P < 0.05 (Student’s t test).
(B) Hierarchical clustering of A. thaliana accessions by genotype. Calculation of similarity
was based on 15 mean RSA traits calculated by Root-VIS. See Supplemental Data Set 5 for
the class assignment of the individual accessions.
(C) Visual reconstructions of RSA and LRS profiles of the central accession of each cluster.
Accessions names are colored according to (B).
B
C
Da(
1)-
12
Lp2
-2K
ult
ure
n-1
Ct-
1Ta
-0V
an-0
WA
RH
od
Rsc
h-4
St-0
Ove
-0R
ak-2
Eden
-2W
l-0
Tom
egap
-2C
ol-
0Ei
-2D
o-0
Bay
-0Jl
-3N
13
HR
-5Se
i-0
Ty-0
Ste-
3U
ll2-5
Kn
-0Si
-0Lm
-2K
ro-0
Kel
ster
bac
h-4
Lillo
-1C
an-0
Ha-
0Sa
p-0
Li-7
Ag-
0Se
-0Li
aru
mH
s-0
Bo
r-1
Nz1
Sq-8
Mc-
0Lu
nd
Ws-
0U
trec
ht
Nw
-2G
el-1
Rev
-2B
g-2
Bsc
h-0
Bla
-1C
nt-
1P
la-0
Ro
u-0
Zu-1
Bo
ot-
1K
on
dar
aC
hat
-1Es
t-0
Mn
z-0
Ler-
1N
p-0
Ull2
-3Ts
u-0
Bu
r-0
Am
el-1
San
na-
2JE
AYo
-0A
a-0
Cvi
-0B
u-0
Ga-
0C
IBC
5Ed
i-0
An
-1W
a-1
Ts-1
Ste-
0Ta
mm
-2A
lc-0
Nc-
1T1
08
0R
mx-
A1
80
Hh
-0G
e-0
Ben
k-1
Ho
v4-1
In-0
Ba-
1G
ie-0
Je-0
Hey
-1P
og-
0G
ot-
7C
om
-1A
lst-
1G
r-1
Tin
g-1
Hn
-0A
ng-
0Jm
-1H
i-0
C2
4K
l-5
Ca-
0M
h-0
Ms-
0P
u2
-23
Or-
0R
a-0
NFA
-8R
ub
ezh
no
e-1
Pro
-0Lo
m1
-1Li
sse
Old
-1Ts
cha-
1R
en-1
Pn
a-1
7B
or-
4C
IBC
-17
Du
kG
y-0
Shah
dar
aFe
i-0
HSm
Kel
ster
bac
h -
2M
z-0
No
k-3
Hau
-0N
o-0
Lip
-0U
od
-7O
b-1
RR
S-7
Ho
vdal
a-2
Es-0
Mt-
0N
d-1
Dra
3-1
Ap
p1
-16
Oy-
0T1
11
0T1
13
0
0.68
0.73
0.78
0.83
0.88
0.93
0.98
Sim
ilarity
MRL MRV MRA BsZL BZL AZL TRS LRS LRS (0.25) LRL TQLRL LRA LRN LRD-MRLRD-BZ OAD
1 0.99* 0.20* 0.09 0.80* 0.83* 0.81* 0.56* 0.58* 0.31* 0.38* 0.11 0.62* -0.11 -0.53* 0.96* MRL
1 0.19* 0.10 0.78* 0.85* 0.80* 0.55* 0.57* 0.30* 0.38* 0.10 0.59* -0.14 -0.52* 0.97* MRV
1 0.19* 0.08 0.22* 0.02 -0.08 -0.09 -0.13 -0.08 0.09 0.00 -0.19* -0.12 -0.01 MRA
1 -0.08 0.05 -0.17* -0.31* -0.35* -0.20* -0.16 0.04 -0.34* -0.53* -0.25* 0.06 BsZL
1 0.37* 0.85* 0.73* 0.65* 0.38* 0.49* 0.10 0.81* 0.30* -0.54* 0.77* BZL
1 0.54* 0.27* 0.38* 0.17* 0.18* 0.07 0.29* -0.37* -0.30* 0.81* AZL
1 0.93* 0.89* 0.68* 0.70* 0.19* 0.84* 0.32* -0.28* 0.80* TRS
1 0.92* 0.79* 0.77* 0.21* 0.84* 0.53* -0.08 0.57* LRS
1 0.76* 0.78* 0.18* 0.75* 0.41* -0.07 0.59* LRS (0.25)
1 0.85* 0.14 0.38* 0.22* -0.12 0.32* LRL
1 0.13 0.48* 0.28* -0.16* 0.40* TQLRL
1 0.17* 0.10 0.09 0.08 LRA
1 0.69 -0.03 0.60* LRN
1 0.41* -0.10 LRD-MR
1
-
0.50* LRD-BZ
1 OAD
A
Col-0Lp2-2 Ste-3 Sanna-2 Mt-0Da(1)-12 N-13 Kn-0
www.plantphysiol.orgon June 6, 2020 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

1
Figure 6. Genome-wide association studies for root system architecture traits
(A) Manhattan plots for GWA mapping of three RSA traits; lateral root size over the basal
quartile of the main root (LRS (0.25)), total lateral root size (LRS), and total root size (TRS).
The horizontal dotted line corresponds to a 5% FDR threshold. Light blue ticks labelled
‘GRF1’ and ‘LRSL’ indicate the location of the most significant associations.
(B) LRS profiles of A. thaliana Col-0 wildtype and mutant lines. Plants were grown on
control media in three independent experiments. The total number of replicate roots
contributing to each LRS profile is given in brackets.
(C) Means and standard errors (n as shown in B) of some RSA traits in wildtype Col-0 and
mutant lines. Significant differences to wildtype are indicated by asterisks (P <0.05,
Student’s t test).
LRS (0.25)
LRS
TRS
GRF1LRSL
A
B CCol-0
(14)
at4g10270
(22)
grf1-1
(18)
Tra
it v
alu
e /cm
0
2
4
6
8
10
12
LRS (0.25) LRS TRS MRL
Col-0
grf1-1
****
**
*
at4g10270
www.plantphysiol.orgon June 6, 2020 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

Parsed CitationsArmengaud, P., Zambaux, K., Hills, A., Sulpice, R., Pattison, R.J., Blatt, M.R., and Amtmann, A. (2009). EZ-Rhizo: Integrated software forthe fast and accurate measurement of root system architecture. Plant J 57: 945–956.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Arsenault, J., Pouleru, S., Messier, C., and Guay, R. (1995). WinRHlZOTM, a Root-measuring system with a unique overlap correctionmethod. HortScience 30: 906.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Bucksch, A., Burridge, J., York, L.M., Das, A., Nord, E., Weitz, J.S., and Lynch, J.P. (2014). Image-based high-throughput fieldphenotyping of crop roots. Plant Physiol 166: 470–486.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Chevalier, F., Pata, M., Nacry, P., Doumas, P., and Rossignol, M. (2003). Effects of phosphate availability on the root systemarchitecture: large-scale analysis of the natural variation between Arabidopsis accessions. Plant Cell Environ 26: 1839–1850.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Delory, B.M., Li, M., Topp, C.N., Lobet, G. (2018) archiDART v3.0: A new data analysis pipeline allowing the topological analysis of plantroot systems. F1000Res. 2018;7.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Delory, B.M., Baudson, C., Brostaux, Y., Lobet, G., du Jardin, P., Pagès, L., et al. (2016). archiDART: an R package for the automatedcomputation of plant root architectural traits. Plant Soil 398: 351–365.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Gruber, B.D., Giehl, R.F.H., Friedel, S., and von Wirén, N. (2013). Plasticity of the Arabidopsis root system under nutrient deficiencies.Plant Physiol 163: 161–79.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Horton, M.W. et al. (2012). Genome-wide patterns of genetic variation in worldwide Arabidopsis thaliana accessions from the RegMappanel. Nat Genet 44: 212–216.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Julkowska, M.M. and Testerink, C.(2015). Tuning plant signaling and growth to survive salt. Trends Plant Sci 20: 586–594.Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Julkowska, M., Koevoets, I.T., Mol, S., Hoefsloot, H.C., Feron, R., Tester, M., Keurentjes, J.J.B., Korte, A., Haring, M.A., de Boer, G.-J.,and Testerink, C. (2017). Genetic Components of Root Architecture Remodeling in Response to Salt Stress. Plant Celldoi:10.1105/tpc.16.00680.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Kalladan, R., Lasky, J.R., Chang, T.Z., Sharma, S., Juenger, T.E., and Verslues, P.E. (2017). Natural variation identifies genes affectingdrought- induced abscisic acid accumulation in Arabidopsis thaliana. PNAS 114: 11536-11541.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Kalogiros, D.I., Adu, M.O., White, P.J., Broadley, M.R., Draye, X., Ptashnyk, M., Bengough, A.G., and Dupuy, L.X. (2016). Analysis of rootgrowth from a phenotyping data set using a density-based model. J Exp Bot 67: 1045–1058.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Kellermeier, F. and Amtmann, A. (2013). Phenotyping jasmonate regulation of root growth. Methods Mol Biol 1011: 25–32.Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Kellermeier, F., Armengaud, P., Seditas, T.J., Danku, J., Salt, D.E., and Amtmann, A. (2014). Analysis of the root system architecture ofArabidopsis provides a quantitative readout of crosstalk between nutritional signals. Plant Cell 26: 1480–1496.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Kellermeier, F., Chardon, F., and Amtmann, A. (2013). Natural variation of Arabidopsis root architecture reveals complementingadaptive strategies to potassium starvation. Plant Physiol 161: 1421–1432.
Pubmed: Author and Title www.plantphysiol.orgon June 6, 2020 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

Google Scholar: Author Only Title Only Author and Title
Laskowski, M., Grieneisen, V.A., Hofhuis, H., Ten Hove, C.A., Hogeweg, P., Marée, A.F.M., and Scheres, B. (2008). Root systemarchitecture from coupling cell shape to auxin transport. PLoS Biol 6: 2721–2735.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Lobet, G., Pagès, L., and Draye, X. (2011). A Novel image-analysis toolbox enabling quantitative analysis of root system architecture.Plant Physiol 157: 29–39.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Lobet G., Pound M.P., Diener J. et al. (2015) Root System Markup Language: toward an unified root architecture description language.Plant Physiol 167:617–627.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Lynch, J.P. (2015) Root phenes that reduce the metabolic costs of soil exploration: opportunities for 21st century agriculture. Plant CellEnviron 38:1775–1784.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Lynch, J.P. and Brown, K.M. (2001). Topsoil foraging - An architectural adaptation of plants to low phosphorus availability. Plant Soil237: 225–237.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Meijón, M., Satbhai, S.B., Tsuchimatsu, T., and Busch, W. (2013). Genome-wide association study using cellular traits identifies a newregulator of root development in Arabidopsis. Nat Genet 46: 77–81.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Morris, E.C., Griffiths, M., Golebiowska, A. , Mairhofer, S., Burr-Hersey, J. , Goh, T. , von Wangenheim, D., Atkinson, B. , Sturrock, C. J. ,Lynch, J.P. , Vissenberg, K., Ritz, K. , Wells, D. M. , Mooney, S. J. , Bennett, M. J. (2017) Shaping 3D root system architecture. Curr Biol27: 919-930
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Müller, L.M., Lindner, H., Pires, N.D., Gagliardini, V., and Grossniklaus, U. (2016). A subunit of the oligosaccharyltransferase complex isrequired for interspecific gametophyte recognition in Arabidopsis. Nat Commun 7: 10826 doi: 10.1038/ncomms10826.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Naeem, A., French, A.P., Wells, D.M., and Pridmore, T.P. (2011). High-throughput feature counting and measurement of roots.Bioinformatics 27: 1337–1338.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Reymond, M., Svistoonoff, S., Loudet, O., Nussaume, L., and Desnos, T. (2006). Identification of QTL controlling root growth responseto phosphate starvation in Arabidopsis thaliana. Plant Cell Environ 29: 115–125.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Ristova, D., Rosas, U., Krouk, G., Ruffel, S., Birnbaum, K.D., and Corruzi, G. (2013). RootScape: landmark-based system for rapidscreening of root architecture in Arabidopsis. Plant Physiol 161: 1086–1096.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Rogers E.D., Benfey P.N. (2015) Regulation of plant root system architecture: implications for crop advancement. Curr Opin Biotechnol32: 93–98.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Satbhai, S.B., Setzer, C., Freynschlag, F., Slovak, R., Kerdaffrec, E., and Busch, W. (2017). Natural allelic variation of FRO2 modulatesArabidopsis root growth under iron deficiency. Nat Commun 8: 15603 doi: 10.1038/ncomms15603.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Seren, U., Vilhjalmsson, B.J., Horton, M.W., Meng, D., Forai, P., Huang, Y.S., Long, Q., Segura, V., and Nordborg, M. (2012). GWAPP: aweb application for genome-wide association mapping in Arabidopsis. Plant Cell 24: 4793–4805.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Shahzad, Z. and Amtmann, A. (2017). Food for thought : how nutrients regulate root system architecture. Curr Opin Plant Biol 39: 80–87.Pubmed: Author and Title www.plantphysiol.orgon June 6, 2020 - Published by Downloaded from
Copyright © 2018 American Society of Plant Biologists. All rights reserved.

Google Scholar: Author Only Title Only Author and Title
Slovak, R., Su, X., Shimotani, K., Shiina, T., and Busch, W. (2014). A scalable open-source pipeline for large-scale root phenotyping ofArabidopsis. Plant Cell 26: 2390–2403.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Svistoonoff, S., Creff, A., Reymond, M., Sigoillot-Claude, C., Ricaud, L., Blanchet, A., Nussaume, L., and Desnos, T. (2007). Root tipcontact with low-phosphate media reprograms plant root architecture. Nat Genet 39: 792–796.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Van der Does, D., Boutrot, F., Engelsdorf, T., Rhodes, J., Mckenna, F., Vernhettes, S., et al. (2017). The Arabidopsis leucine-rich repeatreceptor kinase MIK2 / LRR-KISS connects cell wall integrity sensing, root growth and response to abiotic and biotic stresses. PLoSGenet 13:e1006832.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Voß, U. et al. (2015). The circadian clock rephases during lateral root organ initiation in Arabidopsis thaliana. Nat Commun 6: 7641 doi:10.1038/ncomms8641.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Yu, P. , Gutjahr, C., Li, C.J. , Hochholdinger, F. (2016) Genetic control of lateral root formation in cereals. Trends Plant Sci 21: 951-961Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
www.plantphysiol.orgon June 6, 2020 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.