
Exposure of fH-crystallin to hydroxyl radicals enhances the ...
6
Biochem. J. (1993) 295, 399-404 (Printed in Great Britain) 399 Exposure of fH-crystallin to hydroxyl radicals enhances the transglutaminase-susceptibility of its existing amine-donor and amine-acceptor sites Patricia J. T. A. GROENEN,* Milfred SECCIA,t Ronald H. P. H. SMULDERS,* Enrico GRAVELA,t Kevin H. CHEESEMAN,t Hans BLOEMENDAL* and Wilfried W. DE JONG*§ *Department of Biochemistry, University of Nijmegen, Nijmegen, The Netherlands, tDeparment of Biology and Biochemistry, Brunel University, Uxbridge UB8 3PH, U.K., and tDipartimento di Scienze Cliniche e Biologiche, Universita di Torino, Torino, Italy /H-crystallin was exposed to radiolytically generated hydroxyl radicals at defined radical concentrations, and its capacity to act as an amine-acceptor substrate and as an amine-donor substrate for transglutaminase were investigated. [14C]Methylamine was used as a probe for labelling amine-acceptor sites; a novel biotinylated hexapeptide was used to label amine-donor sites. The results demonstrate that both primary amine incorporation and hexapeptide incorporation by transglutaminase are con- siderably increased after oxidative attack on the crystallin. The identity of the labelled subunits was established, and it is shown INTRODUCTION Oxidative stress is a major threat to the ocular lens in vivo. This stress originates both from oxidants present in the fluids sur- rounding the lens and from oxidants generated within the lens itself (Spector, 1985). Oxidative modifications have been shown to accumulate in the long-lived lens proteins, and this damage has been considered as one of the major factors contributing to aging and cataractogenesis (Augusteyn, 1981; Harding, 1991). Oxidation-induced. modifications of crystallins may lead directly to cross-linking of crystallins (McDermott et al., 1988; Zigler et al., 1989) and to a shift to more acidic crystallins (Garland et al., 1986), which are found in human lenses upon aging and in senile cataract. It is known that oxidative damage to proteins makes them more susceptible for proteolysis (Davies, 1987); this mech- anism also operates on the eye lens crystallins (Carmichael and Hipkiss, 1989). The discovery of transglutaminase activity in the lens of various mammalian species and of significant amounts of y- glutamyl-e-lysine isopeptide bonds in cataractous lenses led to the suggestion that transglutaminase-mediated changes may be important in the aging of the lens and in cataractogenesis (Lorand et al., 1981). Transglutaminases are a group of Ca2+- dependent enzymes involved in the formation of e-(y- glutamyl)lysine isopeptide cross-links in polypeptides and in the conjugation of amines to certain glutamine residues in proteins. The latter glutamine sites are referred to as 'amine-acceptor sites' and the lysine residues involved in polypeptide cross- linking are known as 'amine-donor' or (in the present paper) 'hexapeptide-incorporation' sites. Recently, it was demonstrated that exposure of crystallins to oxidizing free radicals also enhances their susceptibility to transglutaminase, as measured by that, in both cases, this increased incorporation is not due to the production of new substrates, but that the existing incorporation sites become more susceptible. Moreover, using the newly developed probe, we could identify, for the first time, the major crystallin subunits active as amine-donor substrates (both before and after treatment) to be fB 1-, flA3- and flA4-crystallin. These data support the proposal that oxidative stress and trans- glutaminase activity may be jointly involved in the changes found in lens crystallins with age and in the development of cataract. primary amine incorporation (Brossa et al., 1990; Seccia et al., 1991). The enhanced susceptibility of crystallins to transglutaminase after free-radical treatment could be due either to a generalized degradation of proteins, exposing additional and new substrate sites, or to structural changes that make the pre-existing substrate sites more accessible. To distinguish between these two possibil- ities, we studied the influence of hydroxyl-radical (OH') modifi- cation on the transglutaminase substrate capacity of fiH- crystallin using probes for both amine-acceptor and amine- donor activity. Using two-dimensional gel electrophoresis to identify the labelled polypeptides, we demonstrate that oxidative stress does not lead to formation of new substrates, but it increases the susceptibility of the existing transglutaminase substrates. In the bovine eye lens, three fi-crystallin subunits (fiB2-, ,fB3- and flA3-crystallin) have previously been shown to act selectively as amine-acceptor substrates for transglutaminase (Berbers et al., 1984a). In the present study we have now also identified the amine-donor substrates for transglutaminase using a biotinylated amine-acceptor peptide modelled on the sequence around the amine-acceptor glutamine residue in bovine flA3-crystallin (Groenen et al., 1992) as a probe. MATERIALS AND METHODS Exposure of crystallins to free radicals fH-Crystalhn was isolated from the water-soluble fraction of calf lens cortex by gel-permeation chomatography over Ultragel AcA34 (Pharmacia LKB) as previously described (Groenen et al., 1992). It was dissolved in water at pH 7.0 at a concentration Abbreviation used: OH', hydroxyl radical. § To whom correspondence should be sent. Biochem. J. (1 993) 295, 399-404 (Printed in Great Britain) 399
Transcript of Exposure of fH-crystallin to hydroxyl radicals enhances the ...

Biochem. J. (1993) 295, 399-404 (Printed in Great Britain) 399
Exposure of fH-crystallin to hydroxyl radicals enhances thetransglutaminase-susceptibility of its existing amine-donorand amine-acceptor sitesPatricia J. T. A. GROENEN,* Milfred SECCIA,t Ronald H. P. H. SMULDERS,* Enrico GRAVELA,t Kevin H. CHEESEMAN,tHans BLOEMENDAL* and Wilfried W. DE JONG*§*Department of Biochemistry, University of Nijmegen, Nijmegen, The Netherlands, tDeparment of Biology and Biochemistry, Brunel University, Uxbridge UB8 3PH, U.K.,and tDipartimento di Scienze Cliniche e Biologiche, Universita di Torino, Torino, Italy
/H-crystallin was exposed to radiolytically generated hydroxylradicals at defined radical concentrations, and its capacity to actas an amine-acceptor substrate and as an amine-donor substratefor transglutaminase were investigated. [14C]Methylamine was
used as a probe for labelling amine-acceptor sites; a novelbiotinylated hexapeptide was used to label amine-donor sites.The results demonstrate that both primary amine incorporationand hexapeptide incorporation by transglutaminase are con-
siderably increased after oxidative attack on the crystallin. Theidentity of the labelled subunits was established, and it is shown
INTRODUCTIONOxidative stress is a major threat to the ocular lens in vivo. Thisstress originates both from oxidants present in the fluids sur-
rounding the lens and from oxidants generated within the lensitself (Spector, 1985). Oxidative modifications have been shownto accumulate in the long-lived lens proteins, and this damagehas been considered as one of the major factors contributing toaging and cataractogenesis (Augusteyn, 1981; Harding, 1991).Oxidation-induced. modifications of crystallins may lead directlyto cross-linking of crystallins (McDermott et al., 1988; Zigler etal., 1989) and to a shift to more acidic crystallins (Garland et al.,1986), which are found in human lenses upon aging and in senilecataract. It is known that oxidative damage to proteins makesthem more susceptible for proteolysis (Davies, 1987); this mech-anism also operates on the eye lens crystallins (Carmichael andHipkiss, 1989).The discovery of transglutaminase activity in the lens of
various mammalian species and of significant amounts of y-
glutamyl-e-lysine isopeptide bonds in cataractous lenses led tothe suggestion that transglutaminase-mediated changes may beimportant in the aging of the lens and in cataractogenesis(Lorand et al., 1981). Transglutaminases are a group of Ca2+-dependent enzymes involved in the formation of e-(y-glutamyl)lysine isopeptide cross-links in polypeptides and in theconjugation of amines to certain glutamine residues in proteins.The latter glutamine sites are referred to as 'amine-acceptorsites' and the lysine residues involved in polypeptide cross-
linking are known as 'amine-donor' or (in the present paper)'hexapeptide-incorporation' sites. Recently, it was demonstratedthat exposure of crystallins to oxidizing free radicals alsoenhances their susceptibility to transglutaminase, as measured by
that, in both cases, this increased incorporation is not due to theproduction of new substrates, but that the existing incorporationsites become more susceptible. Moreover, using the newlydeveloped probe, we could identify, for the first time, the majorcrystallin subunits active as amine-donor substrates (both beforeand after treatment) to be fB 1-, flA3- and flA4-crystallin. Thesedata support the proposal that oxidative stress and trans-glutaminase activity may be jointly involved in the changesfound in lens crystallins with age and in the development ofcataract.
primary amine incorporation (Brossa et al., 1990; Seccia et al.,1991).The enhanced susceptibility of crystallins to transglutaminase
after free-radical treatment could be due either to a generalizeddegradation of proteins, exposing additional and new substratesites, or to structural changes that make the pre-existing substratesites more accessible. To distinguish between these two possibil-ities, we studied the influence of hydroxyl-radical (OH') modifi-cation on the transglutaminase substrate capacity of fiH-crystallin using probes for both amine-acceptor and amine-donor activity. Using two-dimensional gel electrophoresis toidentify the labelled polypeptides, we demonstrate that oxidativestress does not lead to formation of new substrates, but itincreases the susceptibility of the existing transglutaminasesubstrates.
In the bovine eye lens, three fi-crystallin subunits (fiB2-, ,fB3-and flA3-crystallin) have previously been shown to act selectivelyas amine-acceptor substrates for transglutaminase (Berbers etal., 1984a). In the present study we have now also identified theamine-donor substrates for transglutaminase using a biotinylatedamine-acceptor peptide modelled on the sequence around theamine-acceptor glutamine residue in bovine flA3-crystallin(Groenen et al., 1992) as a probe.
MATERIALS AND METHODS
Exposure of crystallins to free radicals
fH-Crystalhn was isolated from the water-soluble fraction ofcalf lens cortex by gel-permeation chomatography over UltragelAcA34 (Pharmacia LKB) as previously described (Groenen etal., 1992). It was dissolved in water at pH 7.0 at a concentration
Abbreviation used: OH', hydroxyl radical.§ To whom correspondence should be sent.
Biochem. J. (1 993) 295, 399-404 (Printed in Great Britain) 399
400 P. J. T. A. Groenen and others
CL
~i ,B1nlb ,Z 5f3B3
:.:.::.
CL
l3B1a__lb
(ii)
.... W .
§....;....e.-. CL
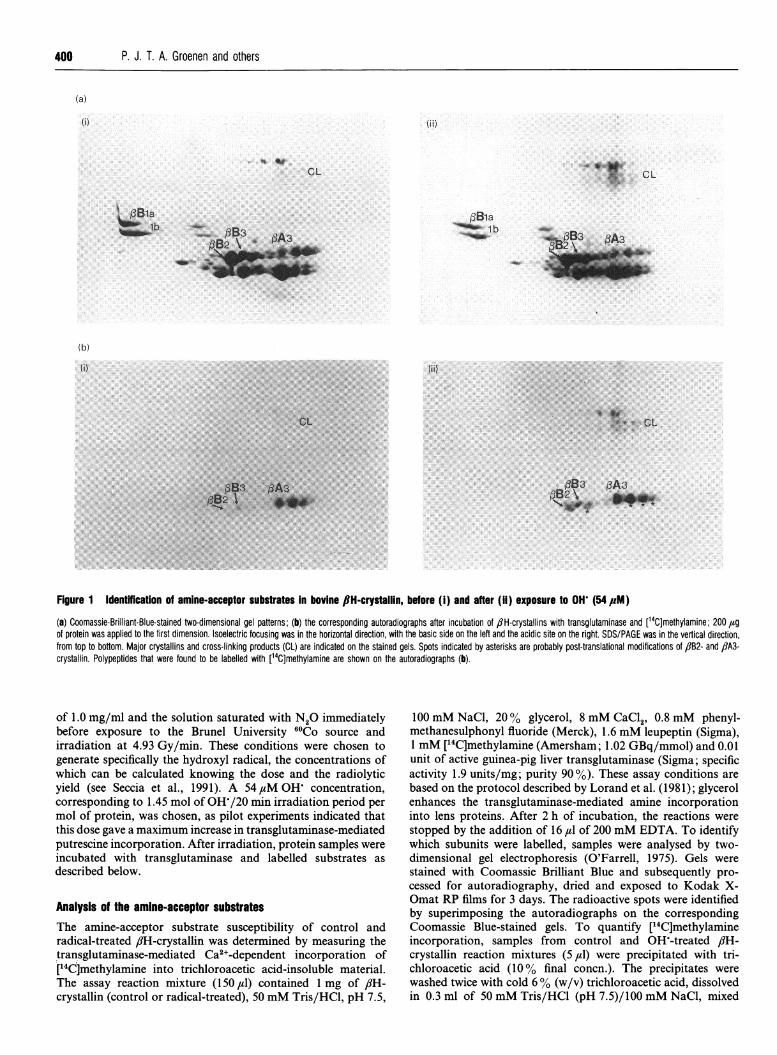
Figure 1 Identffication of amine-acceptor substrates In bovine 8H-crystallin, before (i) and after (ii) exposure to OH (54,pM)
(a) Coomassie-Brilliant-Blue-stained two-dimensional gel patterns; (b) the corresponding autoradiographs after incubation of flH-crystallins with transglutaminase and [14C]methylamine; 200 ,ugof protein was applied to the first dimension. Isoelectric focusing was in the horizontal direction, with the basic side on the left and the acidic site on the right. SDS/PAGE was in the vertical direction,from top to bottom. Major crystallins and cross-linking products (CL) are indicated on the stained gels. Spots indicated by asterisks are probably post-translational modifications of flB2- and flA3-crystallin. Polypeptides that were found to be labelled with [14C]methylamine are shown on the autoradiographs (b).
of 1.0 mg/ml and the solution saturated with N20 immediatelybefore exposure to the Brunel University 60Co source andirradiation at 4.93 Gy/min. These conditions were chosen togenerate specifically the hydroxyl radical, the concentrations ofwhich can be calculated knowing the dose and the radiolyticyield (see Seccia et al., 1991). A 54 ,tM OH concentration,corresponding to 1.45 mol of OH-/20 min irradiation period permol of protein, was chosen, as pilot experiments indicated thatthis dose gave a maximum increase in transglutaminase-mediatedputrescine incorporation. After irradiation, protein samples wereincubated with transglutaminase and labelled substrates asdescribed below.
Analysis of the amine-acceptor substratesThe amine-acceptor substrate susceptibility of control andradical-treated /?H-crystallin was determined by measuring thetransglutaminase-mediated Ca2+-dependent incorporation of[14C]methylamine into trichloroacetic acid-insoluble material.The assay reaction mixture (150 ul) contained 1 mg of /?H-crystallin (control or radical-treated), 50 mM Tris/HCl, pH 7.5,
100 mM NaCl, 20% glycerol, 8 mM CaCl2, 0.8 mM phenyl-methanesulphonyl fluoride (Merck), 1.6 mM leupeptin (Sigma),1 mM [14C]methylamine (Amersham; 1.02 GBq/mmol) and 0.01unit of active guinea-pig liver transglutaminase (Sigma; specificactivity 1.9 units/mg; purity 90 %). These assay conditions arebased on the protocol described by Lorand et al. (1981); glycerolenhances the transglutaminase-mediated amine incorporationinto lens proteins. After 2 h of incubation, the reactions werestopped by the addition of 16 ,u of 200 mM EDTA. To identifywhich subunits were labelled, samples were analysed by two-dimensional gel electrophoresis (O'Farrell, 1975). Gels werestained with Coomassie Brilliant Blue and subsequently pro-cessed for autoradiography, dried and exposed to Kodak X-Omat RP films for 3 days. The radioactive spots were identifiedby superimposing the autoradiographs on the correspondingCoomassie Blue-stained gels. To quantify [14C]methylamineincorporation, samples from control and OH-treated /JH-crystallin reaction mixtures (5 ,u) were precipitated with tri-chloroacetic acid (10% final concn.). The precipitates werewashed twice with cold 6% (w/v) trichloroacetic acid, dissolvedin 0.3 ml of 50 mM Tris/HCl (pH 7.5)/100 mM NaCl, mixed
(a)
(i) (ii)
(b)
(i)
aB2 \,4wiiiiv::>

Exposure of /?H-crystallin to hydroxyl radicals 401
with 4 ml of Aqua Luma Plus (Lumac) and counted in aBeckman Model 2800 liquid-scintillation counter. The increasein [14C]methylamine incorporation in the radical-treated /JH-crystallin above control activity was determined as:
14C incorporation (radical-treated 8lH) - 14C incorporation (
of the amine-acceptor probe increases the molecular mass andlowers the pl of the substrate crystallins. Nevertheless, theidentity ofthe labelled subunits can be deduced from the positions
14C incorporation (control flH)The reported values are the means of triplicate trichloroaceticacid precipitations of flH-crystallin reaction mixtures.
Analysis of the amino-donor substrates (hexapeptideincorporation)Control and radical-treated /JH-crystallins were incubated withactive transglutaminase (Sigma) and a biotinylated amine-acceptor hexapeptide (H-Thr-Val-Gln-Gln-Glu-Leu-OH), asdescribed by Groenen et al. (1992). Samples were analysed bySDS/PAGE (Laemmli, 1970) and two-dimensional gel electro-phoresis (O'Farrell, 1975), followed by staining with CoomassieBrilliant Blue or-streptavidin blot analysis (Groenen etal,- 1992).Quantification of the hexapeptide incorporation was performedin duplicate, scanning the one-dimensional pattern ofstreptavidinblots, loaded with serial dilutions of control and radical-treated,8H-crystallin reaction mixtures, with an LKB-Bromma Ultro-scan XL Enhanced Laser Densitometer.
RESULTSAnalysis of amine-acceptor substratesThe amine-acceptor substrates in control and OW-treated /1H-crystallin were identified by selective transglutaminase-mediatedincorporation of [14C]methylamine. Methylamine is the smallestpossible substrate for transglutaminase and has the advantage ofnot producing noticeable changes in molecular mass and chargeof the proteins in which it is incorporated. The stained electro-phoretic pattern and corresponding autoradiograph of control,/H-crystallin incubated with [14C]methylamine and trans-glutaminase (Figure 1) show that three ,-chains (/3B2-, flB3- and/3A3-crystallin) have been labelled, in agreement with the data ofBerbers et al. (1984a), although flB2-crystallin is only weaklyrepresented. Some modification products of flA3-crystallin arestrongly labelled, and some cross-linked products can also bedetected. After hydroxyl radical treatment of flH-crystallin andincubation with [14C]methylamine and transglutaminase, theintensity of the same crystallins (flB2-, flB3- and flA3-crystallin),including the crosslinking products, is increased; no other fi-crystallin chains become labelled (Figure 1). The substratecapacity of control and radical-treated flH-crystallin was de-termined by measuring the [14C]methylamine incorporation ofthe reaction mixtures. In OW-treated flH-crystallin, an increaseof 77+10 % above control activity was observed. No incor-poration of [14C]methylamine was found in the absence oftransglutaminase or in the absence of Ca2+, either in OH'-treated flH-crystallin or in control flH-crystallin (results notshown).
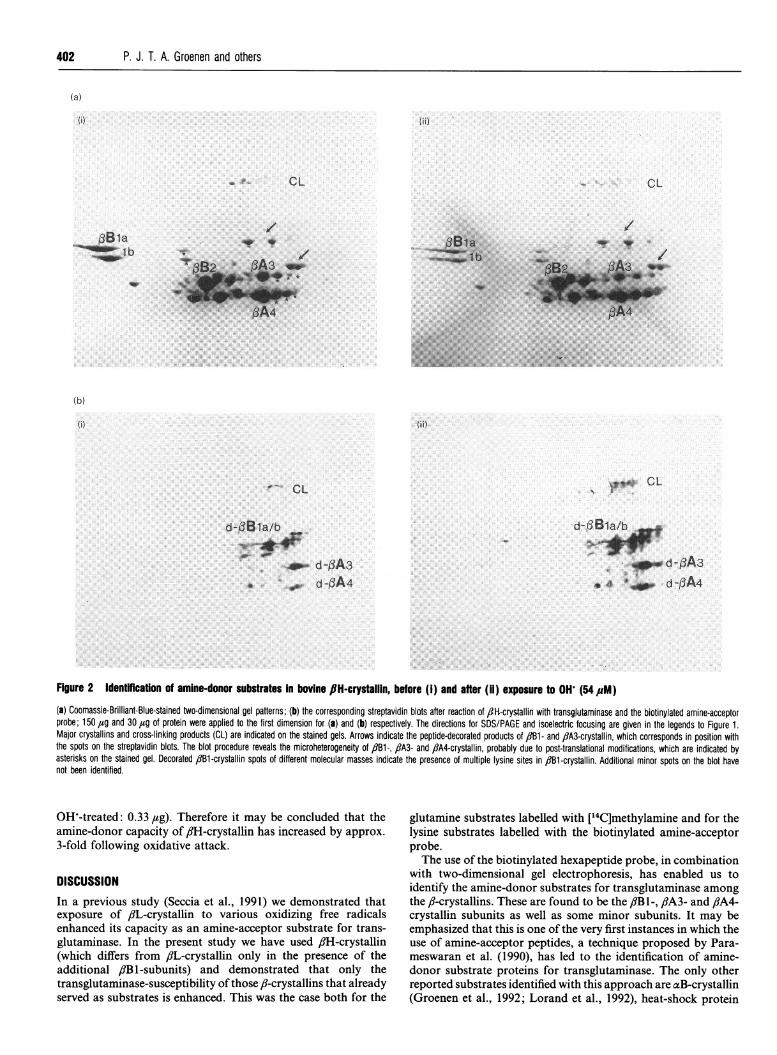
Analysis of amino-donor substratesControl and OHW-treated ,fH-crystallin samples were incubatedwith the biotinylated amine-acceptor probe and analysed by two-dimensional gel electrophoresis. The positions of the subunits inthis separation system are well established from our previouswork (Berbers et al., 1984b). The stained electrophoretic patternand corresponding steptavidin blot are rather complex, as binding
of the peptide-decorated products relative to the unmodifiedsubunits and, in the case of fiBl-crystallin, from the use of aspecific antiserum. The positions of the streptavidin-positivespots demonstrate that 8Bi-, flA3- and flA4-crystallins are thesubunits that serve as amine-donor substrates for trans-glutaminase. The involvement of fiBl-crystallin was confirmedby the positive response of a set of these spots to antiserumspecific for this protein. It appears that both the primary geneproduct fB Ila, its naturally-occurring truncated form /JBlb, andalso their differently charged forms, all react with the hexapeptideprobe. The pattern of the hexapeptide-decorated fiB 1-crystallinsubunits on the streptavidin blot (Figure 2) indicates that itcontains multiple lysine substrate sites for transglutaminase; thisis in accordance with the known presence of four lysine residuesin the N-terminal extension (Mulders et al., 1987).
Antisera against f8A3- or flA4-crystallin were not available,but purification of these subunits and incubation with the probeand transglutaminase confirmed their ability to act as amine-donor substrates (data not shown). The relative intensities of thehexapeptide-decorated subunits on the streptavidin blot [Figure2(ii)] indicate that fiBI- and fiA3-crystallins are better amine-donor substrates for transglutaminase than fiA4-crystallin. Thebinding of the amine-acceptor probe to the substrate proteinsseems to cause a much greater shift in fiB la/b-crystallin than inthe other polypeptides. This may be due to a discontinuous pH-gradient on the basic side during isoelectrofocussing. Moreover,fiB la/b-crystallins have a long N-terminal extension, mainlycomposed of repeating Pro-Ala sequences, which are supposedto induce extraordinary conformational properties that influencethe electrophoretic mobility (Berbers et al., 1983). Decoration ofthe lysine substrate(s) in this sequence may therefore lead to amore drastic change in the apparent pI of the proteins thanexpected.The finding of fiA3-crystallin as amine-donor substrate for
transglutaminase is particularly interesting, because evidencewas provided earlier that fiA3-crystallin can function as amine-acceptor substrate as well (Berbers et al., 1984a). The presence ofmultiple substrate sites within certain crystallins, such as in fiA3-and in fiBI-crystallin, probably explains the labelling of thecross-linking products that is found upon incubation with[14C]methylamine (Figure 1) and with the biotinylated amine-acceptor probe (Figure 2).Comparing the electrophoretic patterns (both stained gels and
corresponding streptavidin blots) of control and OH treatedfiH-crystallin samples reveals that the polypeptides that arelabelled are essentially the same, but the intensity of thestreptavidin-positive spots is increased in the oxidized sample(Figure 2). Spontaneous, transglutaminase-independent incor-poration of the biotinylated hexapeptide in control or radical-treated 8iH-crystallin does not occur (results not shown). Toquantify the increase in lysine-substrate capacity, serial dilutionsof control and radical-treated fH-crystallin reaction mixtureswere analysed on a one-dimensional SDS/PAGE gel, and theintensities of the bands on the streptavidin blot were measuredby laser densitometry. In Figure 3, lane 2a (fiH-crystallin: 1.0 ,ag)is seen to have about the same intensity as lane Sb (,#H-crystallin;
402 P. J. T. A. Groenen and others
(a)
{i)
,3B1at_ lb
(b)
(i)
w- CL
(ii)
9*4* CLN
d-f3Bla/b.1'.
dp-_ d-OA30 aor d-fA4
d-3B1a/b
d-f3A3.S :r#
0. 4 .,_ d-gA4
Figure 2 Identfflcatfon of amine-donor substrates in bovine 8H-crystallin, before (i) and after (ii) exposure to OH (54M#M)(a) Coomassie-Brilliant-Blue-stained two-dimensional gel patterns; (b) the corresponding streptavidin blots after reaction of ,-H-crystallin with transgjutaminase and the biotinylated amine-acceptorprobe; 150 tg and 30 ,ug of protein were applied to the first dimension for (a) and (b) respectively. The directions for SDS/PAGE and isoelectric focusing are given in the legends to Figure 1.Major crystallins and cross-linking products (CL) are indicated on the stained gels. Arrows indicate the peptide-decorated products of 1B1- and fiA3-crystallin, which corresponds in position withthe spots on the streptavidin blots. The blot procedure reveals the microheterogeneity of f1B1-, ,8A3- and fiA4-crystallin, probably due to post-translational modifications, which are indicated byasterisks on the stained gel. Decorated 1B1-crystallin spots of different molecular masses indicate the presence of multiple lysine sites in 13Bl-crystallin. Additional minor spots on the blot havenot been identified.
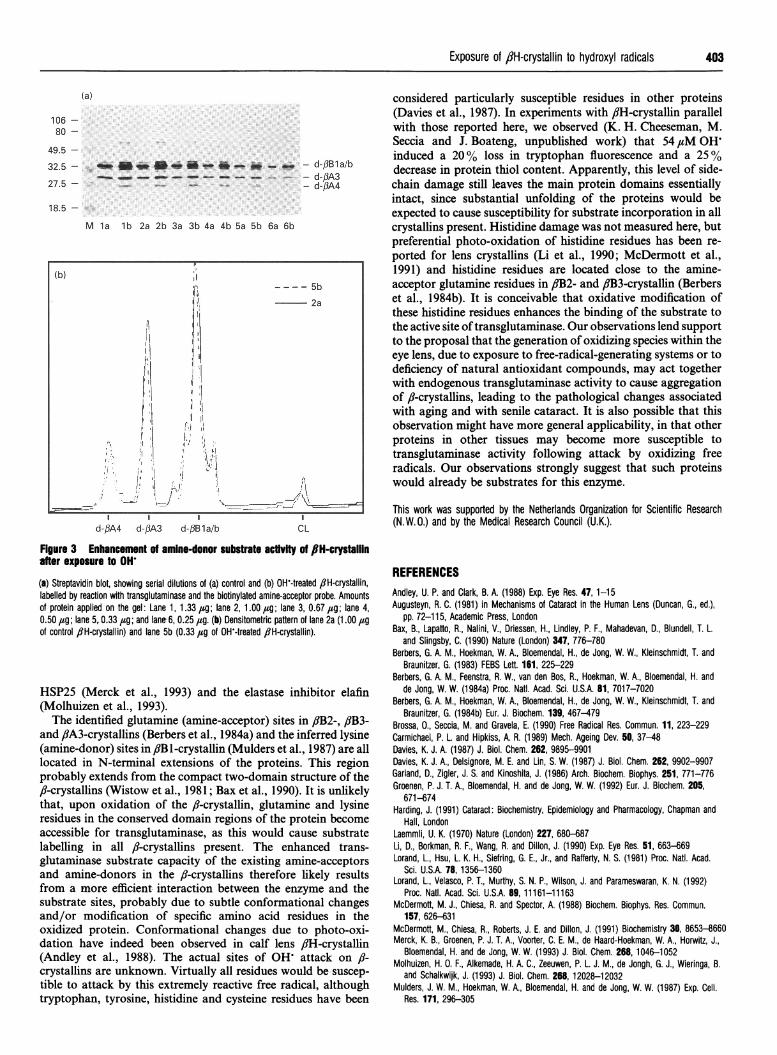
OH'-treated: 0.33,tg). Therefore it may be concluded that theamine-donor capacity of flH-crystallin has increased by approx.3-fold following oxidative attack.
DISCUSSIONIn a previous study (Seccia et al., 1991) we demonstrated thatexposure of /L-crystallin to various oxidizing free radicalsenhanced its capacity as an amine-acceptor substrate for trans-glutaminase. In the present study we have used flH-crystallin(which differs from /L-crystallin only in the presence of theadditional fiB l-subunits) and demonstrated that only thetransglutaminase-susceptibility ofthose fi-crystallins that alreadyserved as substrates is enhanced. This was the case both for the
glutamine substrates labelled with [14C]methylamine and for thelysine substrates labelled with the biotinylated amine-acceptorprobe.The use of the biotinylated hexapeptide probe, in combination
with two-dimensional gel electrophoresis, has enabled us toidentify the amine-donor substrates for transglutaminase amongthe ,-crystallins. These are found to be the ,fB 1-, /3A3- and ,8A4-crystallin subunits as well as some minor subunits. It may beemphasized that this is one of the very first instances in which theuse of amine-acceptor peptides, a technique proposed by Para-meswaran et al. (1990), has led to the identification of amine-donor substrate proteins for transglutaminase. The only otherreported substrates identified with this approach are aB-crystallin(Groenen et al., 1992; Lorand et al., 1992), heat-shock protein
(ii)
CL
/w
N**
f3Bia.:_.Wf:l b
CL
/
VI.W_..
Exposure of flH-crystallin to hydroxyl radicals 403
(a)
106 -80 -
49.5 -
32.5- -d-fBla/b- _ _ _ _ --- _ -- .~ - d-PA327.5 .-. - d-PA4
18.5-
M la lb 2a 2b 3a 3b 4a 4b 5a 5b 6a 6b
I Id-,BA4 d-PA3 d-13B 1 a/b CL
Figure 3 Enhancement of amine-donor substrate activity of flH-crystaillinafter exposure to OH'
(a) Streptavidin blot, showing serial dilutions of (a) control and (b) OH-treated 8lH-crystallin,labelled by reaction with transglutaminase and the biotinylated amine-acceptor probe. Amountsof protein applied on the gel: Lane 1,1.33 /sg; lane 2,1.00 #g; lane 3, 0.67 ,ug; lane 4,0.50 ,ug; lane 5, 0.33 ,ug; and lane 6, 0.25 aug. (b) Densitometric pattern of lane 2a (1.00 ,lgof control 8lH-crystallin) and lane 5b (0.33 j#g of OHW-treated flH-crystallin).
HSP25 (Merck et al., 1993) and the elastase inhibitor elafin(Molhuizen et al., 1993).The identified glutamine (amine-acceptor) sites in flB2-, flB3-
and 81A3-crystallins (Berbers et al., 1984a) and the inferred lysine(amine-donor) sites in /JBI-crystallin (Mulders et al., 1987) are alllocated in N-terminal extensions of the proteins. This regionprobably extends from the compact two-domain structure of the,8-crystallins (Wistow et al., 1981; Bax et al., 1990). It is unlikelythat, upon oxidation of the ,-crystallin, glutamine and lysineresidues in the conserved domain regions of the protein becomeaccessible for transglutaminase, as this would cause substratelabelling in all f8-crystallins present. The enhanced trans-glutaminase substrate capacity of the existing amine-acceptorsand amine-donors in the /?-crystallins therefore likely resultsfrom a more efficient interaction between the enzyme and thesubstrate sites, probably due to subtle conformational changesand/or modification of specific amino acid residues in theoxidized protein. Conformational changes due to photo-oxi-dation have indeed been observed in calf lens 8iH-crystallin(Andley et al., 1988). The actual sites of OH' attack on ,?-crystallins are unknown. Virtually all residues would be suscep-tible to attack by this extremely reactive free radical, althoughtryptophan, tyrosine, histidine and cysteine residues have been
considered particularly susceptible residues in other proteins(Davies et al., 1987). In experiments with /3H-crystallin parallelwith those reported here, we observed (K. H. Cheeseman, M.Seccia and J. Boateng, unpublished work) that 54 ,uM OHinduced a 20% loss in tryptophan fluorescence and a 25%decrease in protein thiol content. Apparently, this level of side-chain damage still leaves the main protein domains essentiallyintact, since substantial unfolding of the proteins would beexpected to cause susceptibility for substrate incorporation in allcrystallins present. Histidine damage was not measured here, butpreferential photo-oxidation of histidine residues has been re-ported for lens crystallins (Li et al., 1990; McDermott et al.,1991) and histidine residues are located close to the amine-acceptor glutamine residues in ,8B2- and /.B3-crystallin (Berberset al., 1984b). It is conceivable that oxidative modification ofthese histidine residues enhances the binding of the substrate tothe active site oftransglutaminase. Our observations lend supportto the proposal that the generation of oxidizing species within theeye lens, due to exposure to free-radical-generating systems or todeficiency of natural antioxidant compounds, may act togetherwith endogenous transglutaminase activity to cause aggregationof /J-crystallins, leading to the pathological changes associatedwith aging and with senile cataract. It is also possible that thisobservation might have more general applicability, in that otherproteins in other tissues may become more susceptible totransglutaminase activity following attack by oxidizing freeradicals. Our observations strongly suggest that such proteinswould already be substrates for this enzyme.
This work was supported by the Netherlands Organization for Scientific Research(N.W.O.) and by the Medical Research Council (U.K.).
REFERENCESAndley, U. P. and Clark, B. A. (1988) Exp. Eye Res. 47, 1-15Augusteyn, R. C. (1981) in Mechanisms of Cataract in the Human Lens (Duncan, G., ed.),
pp. 72-115, Academic Press, LondonBax, B., Lapatto, R., Nalini, V., Driessen, H., Lindley, P. F., Mahadevan, D., Blundell, T. L.
and Slingsby, C. (1990) Nature (London) 347, 776-780Berbers, G. A. M., Hoekman, W. A., Bloemendal, H., de Jong, W. W., Kleinschmidt, T. and
Braunitzer, G. (1983) FEBS Left. 161, 225-229Berbers, G. A. M., Feenstra, R. W., van den Bos, R., Hoekman, W. A., Bloemendal, H. and
de Jong, W. W. (1984a) Proc. Natl. Acad. Sci. U.S.A. 81, 7017-7020Berbers, G. A. M., Hoekman, W. A., Bloemendal, H., de Jong, W. W., Kleinschmidt, T. and
Braunitzer, G. (1984b) Eur. J. Biochem. 139, 467-479Brossa, O., Seccia, M. and Gravela, E. (1990) Free Radical Res. Commun. 11, 223-229Carmichael, P. L. and Hipkiss, A. R. (1989) Mech. Ageing Dev. 50, 37-48Davies, K. J. A. (1987) J. Biol. Chem. 262, 9895-9901Davies, K. J. A., Delsignore, M. E. and Lin, S. W. (1987) J. Biol. Chem. 262, 9902-9907Garland, D., Zigler, J. S. and Kinoshita, J. (1986) Arch. Biochem. Biophys. 251, 771-776Groenen, P. J. T. A., Bloemendal, H. and de Jong, W. W. (1992) Eur. J. Biochem. 205,
671-674Harding, J. (1991) Cataract: Biochemistry, Epidemiology and Pharmacology, Chapman and
Hall, LondonLaemmli, U. K. (1970) Nature (London) 227, 680-687Li, D., Borkman, R. F., Wang, R. and Dillon, J. (1990) Exp. Eye Res. 51, 663-669Lorand, L., Hsu, L. K. H., Siefring, G. E., Jr., and Rafferty, N. S. (1981) Proc. Natl. Acad.
Sci. U.S.A. 78, 1356-1360Lorand, L., Velasco, P. T., Murthy, S. N. P., Wilson, J. and Parameswaran, K. N. (1992)
Proc. Natl. Acad. Sci. U.S.A. 89, 11161-11163McDermott, M. J., Chiesa, R. and Spector, A. (1988) Biochem. Biophys. Res. Commun.
157, 626-631McDermott, M., Chiesa, R., Roberts, J. E. and Dillon, J. (1991) Biochemistry 30, 8653-8660Merck, K. B., Groenen, P. J. T. A., Voorter, C. E. M., de Haard-Hoekman, W. A., Horwitz, J.,
Bloemendal, H. and de Jong, W. W. (1993) J. Biol. Chem. 268, 1046-1052Molhuizen, H. 0. F., Alkemade, H. A. C., Zeeuwen, P. L. J. M., de Jongh, G. J., Wieringa, B.
and Schalkwijk, J. (1993) J. Biol. Chem. 268, 12028-12032Mulders, J. W. M., Hoekman, W. A., Bloemendal, H. and de Jong, W. W. (1987) Exp. Cell.
Res. 171, 296-305
fbI II5b
2a
iii
I.'
I, ,I
s I
(b)
r.
.1
11!I
---,/!

P. J. T. A. Groenen and others
O'Farrell, P. H. (1975) J. Biol. Chem. 250, 4007-4021Parameswaran, K. N., Velasco, P. T., Wilson, J. and Lorand, L. (1990) Proc. Nati. Acad.
Sci. U.S.A. 87, 8472-8475Seccia, M., Brossa, O., Gravela, E., Slater, T. F. and Cheeseman, K. V. (1991) Biochem. J.
274, 869-873
Received 22 December 1992/21 May 1993; accepted 4 June 1993
Spector, A. (1985) in The Ocular Lens (Maisel, H., ed.), pp. 405-438, Marcel Dekker, NewYork
Wistow, G., Slingsby, C., Blundell, T., Driessen, H., de Jong, W. W. and Bloemendal, H.(1981) FEBS Lett. 133, 9-16
Zigler, J. S., Jr., Huang, 0. and Du, X. (1989) Free Radical Biol. Med. 7, 499-505
404







![Geochemical Transactions BioMed Central...ies. Hydroxyl radicals have been directly implicated in lung diseases related to exposures to asbestos [6], silica [7,8], and other airborne](https://static.fdocuments.in/doc/165x107/613013dc1ecc51586943dd22/geochemical-transactions-biomed-central-ies-hydroxyl-radicals-have-been-directly.jpg)


![In Vivo Cell Wall Loosening by Hydroxyl Radicals …...In Vivo Cell Wall Loosening by Hydroxyl Radicals during Cress Seed Germination and Elongation Growth1[W][OA] Kerstin Mu¨ller,](https://static.fdocuments.in/doc/165x107/5fa270bcf76a9306b149591c/in-vivo-cell-wall-loosening-by-hydroxyl-radicals-in-vivo-cell-wall-loosening.jpg)







