
Exploring the Microhabitats of Marsupial Frogs- a Study of the Fo
68
e College of Wooster Libraries Open Works Senior Independent Study eses 2013 Exploring the Microhabitats of Marsupial Frogs: a Study of the Forces Driving Habitat Selection For Flectonotus Fitzgeraldi Within Herbaceous Xanthosoma Jacquinii Populations of Tobago Meredith M. Eyre e College of Wooster Follow this and additional works at: hp://openworks.wooster.edu/independentstudy is Senior Independent Study esis is brought to you by Open Works, a service of e College of Wooster Libraries. It has been accepted for inclusion in Senior Independent Study eses by an authorized administrator of Open Works. For more information, please contact [email protected]. © Copyright 2013 Meredith M. Eyre Recommended Citation Eyre, Meredith M., "Exploring the Microhabitats of Marsupial Frogs: a Study of the Forces Driving Habitat Selection For Flectonotus Fitzgeraldi Within Herbaceous Xanthosoma Jacquinii Populations of Tobago" (2013). Senior Independent Study eses. Paper 4954. hp://openworks.wooster.edu/independentstudy/4954
-
Upload
meredith-eyre -
Category
Documents
-
view
28 -
download
0
Transcript of Exploring the Microhabitats of Marsupial Frogs- a Study of the Fo

The College of Wooster LibrariesOpen Works
Senior Independent Study Theses
2013
Exploring the Microhabitats of Marsupial Frogs: aStudy of the Forces Driving Habitat Selection ForFlectonotus Fitzgeraldi Within HerbaceousXanthosoma Jacquinii Populations of TobagoMeredith M. EyreThe College of Wooster
Follow this and additional works at: http://openworks.wooster.edu/independentstudy
This Senior Independent Study Thesis is brought to you by Open Works, a service of The College of Wooster Libraries. It has been accepted forinclusion in Senior Independent Study Theses by an authorized administrator of Open Works. For more information, please [email protected].
© Copyright 2013 Meredith M. Eyre
Recommended CitationEyre, Meredith M., "Exploring the Microhabitats of Marsupial Frogs: a Study of the Forces Driving Habitat Selection For FlectonotusFitzgeraldi Within Herbaceous Xanthosoma Jacquinii Populations of Tobago" (2013). Senior Independent Study Theses. Paper 4954.http://openworks.wooster.edu/independentstudy/4954

EXPLORING THE MICROHABITATS OF MARSUPIAL FROGS: A STUDY OF THE FORCES DRIVING HABITAT SELECTION FOR
FLECTONOTUS FITZGERALDI WITHIN HERBACEOUS XANTHOSOMA JACQUINII POPULATIONS OF TOBAGO
DEPARTMENT OF BIOLOGY
INDEPENDENT STUDY THESIS
Meredith Milo Eyre
Advisor: Richard Lehtinen and Marilyn Loveless
Submitted in Partial Fulfillment of the Requirement for
Independent Study Thesis in Biology at the
COLLEGE OF WOOSTER 2013

TABLE OF CONTENTS
I. TITLE PAGE
II. TABLE OF CONTENTS 1
III. ABSTRACT 2
IV. INTRODUCTION 3
V. METHODS 16
VI. RESULTS 32
VII. DISCUSSION 37
VIII. ACKNOWLEDGEMENTS 54
IX. LITERATURE CITED 55
X. APPENDIX 61

-‐2-‐
ABSTRACT
This study explored the forces driving habitat selection for a little-known
phytotelm-dwelling tropical marsupial frog, Flectonotus fitzgeraldi (Hylidae), among
phytotelms in the herbaceous Xanthosoma jacquinii (Araceae). Data were collected on
the Caribbean island of Tobago from July 27th to August 3rd, 2012. In order to better
define the frog’s ecological niche, I examined 106 X. jacquinii, of which 11 were
occupied by frogs. Data were collected for environmental variables (canopy cover,
detritus, invertebrate presence, distance to nearest neighboring plants), morphological
characteristics of the plant (height, diameter, length of the longest petiole and leaf,
number of leaf axils), and aquatic variables for the phytotelms within the plants
(dissolved oxygen concentration, temperature, pH, and water depth). A Multiple Logistic
Regression Model found that water depth was the only significant predictor of F.
fitzgeraldi occupancy in X. jacquinii phytotelms (p=0.023). Therefore, the frog species
seems to have a relatively broad ecologic niche based on its ability to withstand a large
range of ecological conditions. This flexibility may allow the frog to occupy other types
of phytotelms including tree holes, bamboo stumps, bromeliads, or phytotelms created by
other plants as long as the microcosm provides an adequate water supply. From a
conservation perspective, this provides hope for the frog’s long term survival despite
possible changes to surrounding environmental or ecological conditions.

-‐3-‐
INTRODUCTION
The field of ecology has developed tremendously in recent years as old theories
are reexamined, new experimental techniques are created, and basic ecological ideas shift
based on new insight. One such concept, the ecological niche, has been redefined many
times over the years. This concept is essential to understand because it is always
intimately linked with at least one of several other major themes in ecology including an
organism’s behavior, morphology, or physiology. In addition, the niche concept provides
insight into how the individual or species functions within its larger community,
specifically with respect to competitor, predator, or prey interactions and resource use.
Therefore, it is a very important concept to define clearly, as it puts other ecological ideas
and theories into a larger context. By fully understanding the niche of a species, one may
ultimately be able to predict distribution or abundance patterns for the species. This
information may be essential to future conservation efforts.
The original description of the niche was focused on understanding the
environmental requirements necessary for the long-term survival and reproduction of an
individual or species (Grinnell 1917). By this theory, the characteristic nature of an
organism’s habitat is the most important factor in determining how an organism or
species fits into the complex ecological web in which it lives. For example, a habitat
comprised mainly of underbrush is essential to the California thrasher because this
structure provides the bird with a place to breed and escape predators in addition to
serving as a source of food. Therefore, characteristics of an individual’s habitat play an
integral role in both the general behavior and long-term survival of the species.

-‐4-‐
Grinnell’s definition of an ecological niche is based on this habitat description. This
niche concept may be used to explain both the behavior of a single individual or
characteristic behavioral patterns of an entire species.
Hutchinson (1957) also developed the niche concept farther by introducing the
idea of a “fundamental” and “realized” niche. A species’ fundamental niche describes
the widest possible range of conditions the individual or species may tolerate (Hutchinson
1957). Historically, biologists have focused considerable time and energy into
determining the factors that contribute to the fundamental niche of an organism.
However, selective pressures may force the species to inhabit a smaller range of
conditions. For example, Robert Paine (1966) studied this phenomenon in the mussel
species Mytilus californianus growing along the shorelines of western North America.
The mussel species grows in a well-defined band in the rocky intertidal zone. This band
of mussels is bordered at its base by a series of predators including the starfish Pisaster
ochraceus. When Pisaster was removed manually, the mussels advanced downward to
inhabit the newly exposed surface relatively quickly. Thus, it can be concluded that even
though M. californianus and P. ochranceus occupy similar ecological niches, the mussels
are forced to only occupy a fraction of their fundamental niche because the starfish will
out-compete any mussels that try to grow into the starfish’s overlapping habitat.
Therefore, the band of intertidal zone that M. californianus occupies represents its
realized niche. Thus, the realized niche is comprised of the smaller range of conditions
the organism is actually able to occupy given the surrounding biotic interactions
(Hutchinson 1957).

-‐5-‐
The distinction between the fundamental and realized niche revolutionized the
niche concept into a more quantifiable measurement based on Hutchinson’s (1957)
mathematical model. This model quantitatively illustrates the smaller realized niche
compared to the full range of conditions the organism could potentially tolerate
comprising the fundamental niche. In this model, each environmental variable is
represented along an axis to illustrate the full range of conditions in which the individual
has the ability to function. For example, this continuum may represent the range of
temperature or resources an individual may withstand. Because each condition is
represented by an axis, the fundamental niche is defined as “a region of an n-dimensional
hypervolume” (Hutchinson 1957). However, interspecific interactions restrict a species
to a fraction of its larger potential. Thus, the realized niche may be depicted
mathematically as a fraction of this larger fundamental niche.
In addition to defining an ecological niche by habitat characteristics and resource
needs, the term has also been used to describe the role of an organism in a community
especially with regard to food consumption (Elton 1927, MacArthur and Levin 1967).
Specifically, Elton (1927) used the term to describe the placement of a species within a
community’s food web. By this definition, Elton observed that arctic foxes and spotted
hyenas occupy very similar niches. The arctic fox eats the eggs of guillemots seasonally,
while relying on the remains of seals killed by polar bears year-round. Similarly, the
spotted hyena relies on the remains of zebras killed by lions year-round, while
supplementing its diet with large numbers of ostrich eggs seasonally. Because the arctic

-‐6-‐
fox and spotted hyena have parallel roles in their community structure, by Elton’s
definition, their ecological niche is essentially the same.
Both Grinnell’s habitat and resource based definition and Elton’s ecological role
based definitions are critical to understanding how an individual or species fits into a
larger community structure and survives long-term in its environment. These definitions
were integrated with the emergence of the competitive exclusion principle, stating that
two competing species must differ in several traits related to their fitness in order to
coexist (Hardin 1960). For example, two species may live within the same habitat
directly in contact with each other, but as long as they have a different role in the food
web, they may coexist. Conversely, the long term survival of two species with identical
ecological roles is not threatened as long as they live in different habitats. This
competitive exclusion principle is based on the assumption that one species will
inevitably out-compete the other species for key resources, eventually forcing it out of the
habitat. However, two species with differing resource requirements and ecological roles
may coexist because each occupies a separate ecological niche within the same
ecosystem.
Because the niche concept is fundamentally integrated into so many aspects of
ecology, it has been used in many ecological papers over the years, often to the point of
confusing its meaning. Chase and Leibold (2003) attempted to redefine the niche concept
with the following definition: “an ecological niche is the joint description of the
environmental conditions that allow a species to satisfy its minimum requirements so that
the birth rate of a local population is equal to or greater than its death rate along with the

-‐7-‐
set of per capita effects of that species on these environmental conditions.” This
definition incorporates both historical perspectives of the ecological niche concept,
essentially combining Grinnell’s resource use definition with Elton’s ecological role
definition. This niche definition may be used in the context of a single organism or an
entire species.
An organism’s mobility or lack thereof plays a crucial role in establishing its
ecological niche. Organisms that are immobile are limited to a single geographic location
and are forced to face the challenges of the environment without moving to avoid harsh
conditions. Thus, plants have developed a wide range of morphological characteristics in
response to the challenges being immobile present. This has resulted in both the
speciation of entire new lineages and phenotypic plasticity of individual plants. In
contrast, mobile organisms have the ability to actively select their habitats and move in
response to environmental stimuli. Individuals that inhabit areas that promote their
survival and reproduction have a clear evolutionary advantage. Furthermore, the
individuals that inhabit less suitable environments will leave fewer offspring (Southwood
1988). Because an organism’s habitat is crucial to its survival and reproduction, it makes
sense for a species to evolve mechanisms that allow its members to perceive and respond
to their environments accordingly. Therefore, evolutionary adaptations that allow for
habitat selection play a crucial role in determining the future success of the species
(Southwood 1988, Holt 1987, Murphy 2003).
Habitat selection, or the concept that individuals are able to choose to occupy the
habitat in which their fitness is greatest, represents another fundamental theme in ecology

-‐8-‐
(Rosenzweig 1991). The term is used to refer to a set of behavioral responses that may
result in the disproportionate use of habitats, ultimately influencing the survival and
fitness of individuals (Jones 2001, Hutto 1985, Block and Brennan 1993). Habitat choice
is demonstrated in, but not limited to, several species of insects, lizards, rodents, and
birds (Rausher 1984, Brown 1998, Hanski and Singer 2001, Odling-Smee et al. 2003).
These studies have shown that organisms move in response to a wide variety of
conditions including resource needs, biotic factors, and environmental variables.
Because of the range of habitats mobile organisms experience, some evolutionists believe
that habitat selection by individuals may channel the direction of adaptive evolution
(Rosenzweig 1987, Holt 1987). Thus, organisms are not simply at the mercy of their
environment, dependent on the limits of their physiology. Instead, mobile organisms
actively play a role in determining their long-term survival by learning how to respond to
environmental cues and develop habitat selection techniques. Previous analyses have
focused primarily on alleles, genotypes, and phenotypes that influence habitat or resource
choice by examining movement patterns for organisms in heterogeneous environments
(Krebs et al. 1978, Stephens and Krebs 1986).
The term “habitat selection” implies that complex behavioral patterns are
understood by biologists on some level (Cherrett 1989). In contrast, “habitat use” simply
refers to the way an organism uses a collection of physical and biological entities in a
habitat (Krausman 1999). Therefore, the distribution pattern of individuals is the end
result of the habitat selection process (Jones 2001, Cherrett 1989). In order to confidently
conclude that habitats have been selected, an important aspect of habitat selection must

-‐9-‐
be met: individuals within the study species must demonstrate the ability to make
decisions about their surroundings. The resulting habitat use may be described and
explained through the analysis of the costs and benefits associated with certain habitat
characteristics. A disproportional use of potential habitats corresponding with these
quality assessments provides evidence for this active habitat selection (Fretwell and
Lucas 1970, Jones 2001). Therefore, understanding patterns in overall habitat quality is
essential in determining if the individual has actively selected its habitat. It is important
to remember that without behavioral or life history information, there is no way to know
if the detected differences in habitat characteristics actually have any direct influence on
the choice of the individual (Jones 2001, Martin 1992, Martin 1998). However, even if
the individual’s resulting fitness is not fully understood, it is still assumed to be adaptive
on some level (Jones 2001, Robertson 1972, Pulliam and Danielson 1991, Martin 1998).
Ultimately, a compromise must be made when assessing the quality of the habitat.
Knowledge of how each environmental factor affects a species must be balanced with
inferred predictions based on observations of similar species (Jones 2001).
Jones’ (2001) study states that habitat selection may result in the disproportionate
use of habitats. Although not the sole evidence for habitat selection, this disproportionate
use of potential habitats based on the quality of habitat may serve as evidence for habitat
selection. Instead of individuals being evenly distributed throughout their entire potential
range, they may be found in larger numbers in a subsection of this habitat. If
characteristics of this smaller habitat prove beneficial to the individual, it may support the
theory that habitats are actively being selected. The ideal free distribution theory further

-‐10-‐
explains this observation (Cherret 1989 pg. 327). This theory is a widely discussed
model that helps predict where animals will be distributed within the environment, based
on the assumption that all the mobile animals are free and able to travel wherever they
like. In addition, it is also assumed that the organisms each have a perfect knowledge of
every habitat within the larger ecosystem. According to this theory, a relatively equal
distribution of species will spread evenly throughout the entire potential range of the
species (Fretwell and Lucas 1970). As the highest quality environments become too
densely populated, their overall quality decreases until their quality matches the
previously unoccupied habitats. At this point, the previously unoccupied habitats become
filled from the overflow of organisms from the more densely populated habitats.
Eventually, a relatively equal distribution of species will result, spread evenly throughout
the habitats of equal quality. Therefore, the disproportional distribution of individuals
throughout the larger range of the species provides evidence that habitat selection has
occurred.
There is a fitness advantage in being able to perceive and respond to certain
environmental cues. However, it is difficult for biologists to recognize and prioritize
which biotic or abiotic conditions contribute most to the quality of a habitat. Examining
the effects of these conditions on an individual’s behavior or physiology is difficult on a
large scale because the individual must respond to both a variety of conditions at once
and a range within each condition (Brown et al. 1996). Therefore, a biologist may spend
years assessing the characteristics of an organism’s habitat, only to find that nothing more
than broad observations may be made because the animal roams a large area of land that

-‐11-‐
encompasses a wide variation in environmental factors. For example, researchers
observed broad patterns that could be used to describe the habitat selected by Asian
elephants. The study found these elephants preferred a dense shrub layer, shorter trees, a
dense canopy, and smaller trunk radii (Limin and Li 2005). However, it would be very
difficult to confidently conclude that tree height alone affects the habitat of the elephants,
as the elephant’s natural range incorporates a large range of tree heights in addition to the
multitude of other factors. Since these organisms move over a large spatial scale, it
would be nearly impossible logistically to quantify and identify all aspects of the
environment that may play a role in habitat choice (Limin and Li 2005). In fact, some
species may require different types of habitats and conditions over the course of their
lifetime. Ultimately, a combination between both biotic and abiotic factors determines
the quality of an organism’s habitat (Heying 2004). Therefore, although each factor may
contribute either positively or negatively to the fitness of the individual, it is often
difficult to specify which factors are most important to an organism’s success.
However, the complex system may be simplified by focusing on a smaller spatial
scale (Lehtinen et al. 2004). Some mobile organisms may spend the majority of their life
living within a self-contained microcosm. Although microcosms may be difficult to
locate in the field, they may be easily and thoroughly observed upon discovery (Lehtinen
et al. 2004). Because of their discrete nature, microcosms serve as model systems for
studying the characteristics of the habitat these mobile organisms inhabit. Each
microcosm may be studied on a very small scale to precisely determine the biotic,
chemical, and physical characteristics affecting the system (Srivastava et al. 2004,

-‐12-‐
Lehtinen et al. 2004). Furthermore, most factors measured will be homogenous
throughout the entire system. Therefore, a more holistic assessment of the habitat that
directly affects the individual is possible. This may provide insight into how mobile
organisms living within microcosms respond to both biotic and abiotic challenges and
utilize the surrounding environment. Microcosms also provide a clear logistical
advantage in that many replicates may be sampled per unit time, thus increasing the
sample size within a study.
Species that inhabit phytotelmata are a prime example of organisms that inhabit
microcosms (Lehtinen et al. 2004, Summers and McKeon 2004). A phytotelm is a body
of water contained entirely within a plant (derived from Greek: phytos=plant, telm=pond)
(Lehtinen et al. 2004). There are many different types of phytotelmata including water-
filled tree holes, bamboo stumps, bromeliad tanks, nut husks, tree buttresses, and leaf
axils (Lehtinen et al. 2004, Silva et al. 2011, Lin and Kam 2008, Chiu and Kam 2006,
Kam et al.1996). The volume of water in these different microcosms may range in size
from several milliliters (Rödel et al. 2004) to tens or hundreds of liters (Schiesari et al.
2003).
Phytotelmata may be used for breeding, feeding, resting, and water-balance for a
variety of species in the tropics (Lehtinen 2002). For example, water-filled axils in screw
pines (Pandanus) were shown to provide microhabitats for 20 different species of reptiles
and amphibians in Madagascar (Lehtinen 2002). In addition, past research has indicated
that there are at least 102 species of phytotelm-breeding frogs, representing at least 2% of
all recognized frog taxa (Lehtinen et al. 2004).

-‐13-‐
A phytotelm presents a unique set of benefits and challenges to the individual
inhabiting it (Lehtinen 2004, Silva et al. 2011, Lehtinen and Carfagno 2011). Although
these microcosms are thought to be relatively safe from the predators and competitors
often present in larger ponds or streams, the unique microhabitat is not necessarily a safe
haven (Caldwell and Araujo 2004, Lehtinen 2004). Invertebrates, reptiles, or other
species of amphibians have been observed acting as predators, competitors, parasites, and
prey for the inhabitants of the phytotelm. In addition to these biotic factors, the small
bodies of water may be prone to desiccation, have very low levels of oxygen, or contain
unpredictable amounts of food (Caldwell and Araujo 2004, Lehtinen 2004). The
inhabitants of the phytotelms must also respond to a range in other abiotic conditions
including temperature, moisture, amount of detritus, and canopy cover (Lehtinen and
Carfagno 2011, Silva et al. 2011). Spatial isolation may affect both the general shape and
size of the plant, in turn affecting the quality of the phytotelm within. Spatial isolation
may also help determine the frog’s ability to locate and inhabit the plant (Silva et al.
2011). In addition, certain morphological characteristics of the plant have been shown to
affect the phytotelm’s ability to hold and retain water and thus influence the quality of the
habitat (Zotz and Thomas 1999, Silva et al. 2011, Lin and Kam 2008). An increased
water volume was also shown to positively influence the occupancy of both Guibemantis
bicalcaratus and Guibemantis punctatus phytotelm-breeding frogs living within the
water-filled axils of the Pandanus plants (Lehtinen and Carfagno 2011). In addition, a
larger plant size may provide larger phytotelms with the ability to hold a greater volume
of water. Therefore, water volume and plant size may also be important factors

-‐14-‐
contributing to the overall quality of the microcosm. Several studies have also analyzed
the effect of water chemistry and other aquatic variables when predicting frog occupancy
of phytotelms (Caldwell and Araujo 2004, Silva et al. 2011). However, results from
these studies varied, especially with respect to levels of dissolved oxygen and pH of the
water within phytotelms. Furthermore, little is known about the effect of water
temperature in terms of the challenges it presents for the frogs.
Evidence supports the hypothesis that some species of phytotelm-breeding frogs
actively select a specific microcosm to occupy based on these biotic and abiotic cues
(Heying 2004, Resetarits 1996). For example, Phrynobatrachus guineensis is highly
mobile and individuals move between water-filled cavities frequently, suggesting that
they have the ability to actively select their habitat on some level (Sandberger et al.
2010). Xenohyla truncate has been observed moving through vegetation at night, as if
actively moving to a more suitable habitat (Silva and Britto-‐Pereira 2006). Similarly,
larvae of both Mantidactylus bicalcaratus and Mantidactylus punctatus have been
observed climbing along leaf surfaces to move to different axils (Lehtinen 2004). Female
Scinax perpusillus have also been observed both dipping their legs or hind body and
diving into and swimming around the small pool as if “testing the water” before selecting
a mate and spawning (Alves-Silva and Silva 2009). Poison dart frogs distribute their
offspring individually between phytotelms they have actively chosen (Summers 1992,
Williams et al. 2007). In most Dendrobates species, this behavior is performed by the
male (Summers 1992). This behavior ensures that the cannibalistic tadpoles do not have
a chance to eat each other, therefore maximizing reproductive success. Habitat selection

-‐15-‐
in phytotelm-breeding frogs seems to be based on a number of biotic and abiotic factors.
However, very little is known regarding the behavior and ecology of most tropical
phytotelm-breeding frogs.
Individuals from one such species, Flectonotus fitzgeraldi (Hemiphractidae), are
commonly known as marsupial frogs. The larvae develop partially in the female’s dorsal
marsupium before they are deposited into the phytotelmata created within the leaf axils of
Xanthosoma jacquinii (Araceae) (Duellman and Gray 1983, Kenny 1969, Murphy 1997
pg. 63-65, RML unpublished observations). Therefore, larvae are already
developmentally advanced and presumably better able to cope with the challenges the
phytotelm presents. This strategy also reduces the amount of time the tadpole has to
remain in the microcosm.
In an attempt to better understand the ecological niche of the phytotelm-breeding
frog F. fitzgeraldi, this study explores the biotic and abiotic conditions contributing to the
overall quality of its habitat. Specifically, this study sought to answer the following
question: To what environmental characteristics do marsupial frogs respond in order to
select habitats that promote optimal fitness? Because very little is known about the
natural history of F. fitzgeraldi, additional insight was noted in this study. Ultimately,
this information may be valuable for future conservation efforts.

-‐16-‐
METHODS
Study area and study system
This study was conducted in the rainforest on the Caribbean island of Tobago
over the course of eight days. Several trials were run the first day to standardize the
measuring technique. These trials were not included in the final analysis. Data analyzed
in this study were collected from July 28 to August 3, 2012 between the hours of 9:00 am
and 6:00 pm. The collection period fell within the rainy season, with temperatures
ranging from 22.6°C-29.1°C and humidity ranging from 85%-100%. Because the air
temperature and humidity were recorded every time a plant was sampled, these ranges
represent conditions within the study sites over the course of the day. Two study sites
were established on the island (see Figure 1). One site was located within the Tobago
Forest Reserve about a 20-minute hike upstream from a bridge on the Roxborough
Parlatuvier Road (11°17.221 N, 60°35.676 W, elevation: 410 m) (see Figure A1). The
second site was located along a tributary of the Bloody Bay River about 30 m upstream
from the coordinates 11°17.967’ N, 60°37.085’ W at an elevation of 56 m (see Figure
A1). Although rarely the dominant species in the rainforest, the understory herb
Xanthosoma jacquinii (Araceae) does occur in relatively high numbers in patches along
streams in both locations.

-‐17-‐
Figure 1. Data for this study were collected on the Caribbean island of Tobago. Tobago is a relatively small island (300 km2) located NE of Trinidad (4,768 km2). Together, the islands form the Republic of Trinidad and Tobago. The points indicate the two sites sampled in this study. The NW point corresponds to the Bloody Bay site and the SE point corresponds to the site in the Forest Reserve. Flectonotus fitzgeraldi was found at both locations. Flectonotus fitzgeraldi tadpoles and adults of both sexes inhabit the phytotelms
created within the leaf axils of X. jacquinii (RML unpublished data, personal
observations) (see Figure 2). Adult individuals have been observed inhabiting
phytotelms in the Xanthosoma plant at both these locations in years past (RML
unpublished data). Kenny (1969) also found F. fitzgeraldi tadpoles in the base of a
Xanthosoma plant. He noted that 26 tadpoles were found together in a single phytotelm
containing approximately 200 mL of water. This is the only historical sighting of F.
fitzgeraldi tadpoles. Adults have also been found in other terrestrial plants, including leaf

-‐18-‐
axils of bromeliads and rolled-up leaves of Heliconia plants (Murphy 1997). The latter
do not retain water so these leaves may only be used for shelter, as reproduction is
limited to phytotelms (although the bracts of Heliconia flowers may serve as viable
phytotelms). Flectonotus fitzgeraldi is probably terrestrial by day and arboreal at night
based on observations made by Duellman and Gray (1983) for F. pygmaeus in
Venezuela. The frogs are crepuscular, and begin calling an hour before and continue
calling an hour after sunset (Murphy 1997, Kenny 1969). At dusk, the frogs were
observed leaping between branches of a bromeliad-laden tree, eventually entering the
bromeliads (Murphy 1997). Murphy notes that the leaps are long and fast, making the
frog’s capture difficult. He also observed adults crossing forest roads on rainy nights.
These observations collectively describe everything previously known about the habitat
of F. fitzgeraldi. On a larger scale, this species occurs on Trinidad, Tobago, and on the
Península de Paria in Venezuela (Murphy 1997).
Figure 2. The relative difference in male and female body shape was used to distinguish between the sexes in the field. This sex determination was based on the assumption that males had a more tapered body than females.

-‐19-‐
Because amplexus and oviposition have not been observed in this species, the
following behaviors are inferred from observations made by Duellman and Maness
(1980) of F. pygmaeus. Immediately after eggs are fertilized, the male is thought to use
his hind limbs to individually push each egg up into the marsupium on the female’s
dorsum where they develop from small fertilized eggs into fairly well-developed
tadpoles. The clutch size for this species is the smallest in the species group, ranging
from 2-6 offspring (Murphy 1997, RML unpublished observations). The developing
embryos form distinct, round protrusions on the female’s dorsum (Murphy 1997,
personal observations). Duellman and Gray (1983) suspect females may be capable of
producing multiple broods per season, but more research is necessary to confirm this
hypothesis. When the tadpoles reach advanced stages of development (stages 39-41 of
Gosner 1960), they are deposited by the female into a phytotelm (Murphy 1997). Water
held within this phytotelm provides a pool just large enough to support the tadpoles as
they metamorphose into adults. Kenny (1969) observed that metamorphosis occurred
five days after leaving the parent. During this period, tadpoles are well supplied with
yolk and do not feed (Kenny 1969). While there are no published studies describing the
feeding behavior and diet for adult F. fitzgeraldi individuals, it may be inferred from
similar species that this frog is a dietary generalist. Therefore, the frog is expected to eat
any non-vegetative matter it can reasonably fit into its mouth.
Flectonotus fitzgeraldi has been observed to inhabit the phytotelms created within
the leaf axils of Xanthosoma jacquinii from the Araceae family (see Figure 3). This plant
has large leaves and thick, fleshy decumbent stems (Simmonds 1950). The plant

-‐20-‐
produces flowers and seeds and also propagates vegetatively (see Figure 4). It produces
grey latex previously thought to be poisonous (Simmonds 1950). The entire plant gives
off a rotten stench (Simmonds 1950, personal observations).
Figure 3. The phytotelms are found at the leaf axils within Xanthosoma jacquinii. This image is drawn from a photo and provides insight into the structure of the plant. To establish a relative size scale, the backpack at the base of the plant is approximately 50 cm tall.
This color distinction is considered the “top of the trunk.” The height of the plant is measured from the ground to this point. The DBH is also measured at this point.

-‐21-‐
a) b) Figure 4. The trunk of Xanthosoma jacquinii. It may either emerge directly from the ground (a) or grow vegetatively (b). In both situations, roots extend from higher on the trunk down into the soil. The height of the plant was measured from the level of the soil.
Very little information is known about this plant, although one study noted it was
found in rocky ravines and along streams on the nearby island of Trinidad (Simmonds
1950). In Trinidad, it has been found throughout the wetter northern parts of the island
growing on metamorphic rock or sandstone. This species has been recorded at altitudes
up to 760 m on this island, although the highest point in Tobago is only 549 m
(Simmonds 1950, Murphy 1997). In Tobago, this plant seems to grow in relatively dense
clusters and seems to be limited to areas of undisturbed deep forest (personal
observations, RML unpublished data). Although Simmonds (1950) describes the plant to
be “locally abundant” in Trinidad, we found that locating these patches in Tobago was
relatively difficult and the ones we found always occurred in undisturbed forested areas.
However, the search strategies used to make this observation were limited to casual
observations while collecting data for this study and others conducted in years past.
Although extensive additional data is needed to support this conclusion, it seems likely
that the plant’s long-term survival is dependent on the preservation of old-growth forests.

-‐22-‐
Plant selection
Five patches of Xanthosoma jacquinii were sampled at each site, each patch
containing ten individuals (see Figure A1). Patches were chosen haphazardly along a
stream. The first plant encountered when hiking up the stream was used as starting point.
From this origin, the nearest nine other individuals were selected based entirely on spatial
distribution. Therefore, the selected plants always occurred further upstream or further
up the bank of the stream in relation to the starting point. Together, these individuals
collectively formed a “patch.” In order to be included, a plant had to have a DBH
≥4.5cm, contain at least two leaf axils, and not exceed 190 cm in height. Preliminary
observations showed that plants smaller than these minimum criteria did not have
phytotelmata large enough to serve as viable microhabitats for F. fitzgeraldi. In addition,
plants exceeding 190 cm in height were not included because they were outside the
observer’s range of visibility. Individuals were not considered a part of the patch if they
were more than three meters away from another Xanthosoma jacquinii already
established to be part of the patch. The distance between any two plants in separate
patches always exceeded 3 meters, and often much more. Plants sampled within these
patches were measured for a number of characteristics (see ‘Habitat Characterization’
below).
In addition to the plants sampled within the established patches, about 10 hours
were spent informally searching plants which had not already been examined for F.
fitzgeraldi. Of this time, approximately 8 hours were spent in the Bloody Bay site and 2
hours were spent in the Forest Reserve site. This searching became crucial to increasing

-‐23-‐
the sample size of occupied plants given the time constraints since relatively few
Xanthosoma in the established patches contained F. fitzgeraldi. During this informal
searching, every plant encountered was examined for frogs. The path taken to find these
plants was random on a small scale but systematically covered a very large area. The
route was dictated by plant densities, as more time was spent in areas more densely
populated with X. jacquinii. At Bloody Bay, a high density of plants often occurred
further up the bank of the stream in addition to the populations already surveyed near the
stream. Therefore, extensive time was spent examining plants high on the surrounding
slopes. At the Forest Reserve, efforts to mimic the sampling technique along the high
slopes were hindered due to topographical differences. Although the slopes along the
streams were not as high, the sampling effort still remained focused on surveying areas
with the largest X. jacquinii density. If F. fitzgeraldi was found during these informal
plant surveys, the occupied plant was measured identically to the formally sampled plants
and the resulting data were included in the final analysis.
Frog surveys
Each plant was surveyed for F. fitzgeraldi immediately upon approach. Leaf axils
were examined closely for any individuals. The surveying continued long enough to be
confident about the occupancy status (frogs present or frogs absent) and varied based on
the relative size of the plant (range: 30 s - 3 min) (see Figure 5). In addition to peering
directly into the top of a phytotelm, a headlamp was held against the base of the petiole to
illuminate the depths of the phytotelm through the petiole (see Figure 6). This technique

-‐24-‐
helped distinguish frogs hiding deep in the base of the phytotelm from debris and other
organic matter. If any organic matter was questionable, an eyedropper was used to prod
the matter for movement. Because of the small size of the phytotelms and structurally
simple nature of the plant, it was easy to survey X. jacquinii plants thoroughly, and
adequate time ensured that few frogs (if any) were missed.
Figure 5. Xanthosoma jacquinii plants may have many leaves. The time required to sample each plant for frogs ranged from 30 s – 3 min. based on the number of leaf axils it contained. This figure shows how the petioles protrude from the main trunk. Although this figure shows flower buds emerging from the central trunk, the vast majority of plants did not have buds, flowers, or fruits at the time the study was conducted. The circled regions indicate the highest and lowest phytotelm. New leaves grow vertically from the top of the main trunk and “uncurl” as they mature. If the petiole was bent downward in a way that positioned the leaf lower than its axil, the phytotelm was destroyed because the water drained from the axil. Therefore, it was not counted in the analysis.

-‐25-‐
Figure 6. A head lamp was held directly against the base of the phytotelm to illuminate its depths more clearly. The arrow indicates the direction of the light.
Habitat characterization
In order to determine the biotic and abiotic factors potentially influencing the
quality of the phytotelms F. fitzgeraldi inhabit, a variety of measurements were taken
(see Table 1).

-‐26-‐
Table 1: The phytotelms were characterized by a number of biotic and abiotic factors. Data were collected based on the following variables. The shaded variables were eliminated from the final multiple logistic regression model because they were highly correlated with at least one other variable. The variables were considered highly correlated if they had a correlation coefficient >0.7 using the Spearman Test. Independent Variables Categorical or
continuous Explanation
Canopy cover Categorical (0-4 scale), 0=100% canopy cover, 4=0% canopy cover Detritus Categorical (0-4 scale), 0=leaf axils completely full of detritus, 4=leaf axils
devoid of detritus Invertebrate density Categorical (0-4 scale), 0=no invertebrates, 4=very high density of invertebrates. Invertebrate presence Categorical 0=no invertebrates, 1=invertebrates present Invertebrate size Categorical 0=no invertebrates, 1=only small invertebrates present (<7mm),
2=only large invertebrates present (>7mm), 3=both small (<7mm) and large (>7mm) invertebrates present
Volume of water Categorical (0-4 scale), 0=no water in phytotelm, 4=phytotelm completely full of water
Depth of water Continuous Measured from the base of the phytotelm to the surface of the water (to the nearest half-centimeter)
Oxygen concentration Continuous Measured by Oakton probe in mgO2/L water
Oxygen concentration Continuous Measured by Oakton probe in % O2 pH Continuous Measured by Oakton probe Temperature of water Continuous Measured by Oakton probe in °C Trunk height Continuous Measured from the ground to the point on the central trunk where a
color change occurs between the green leaf stalks and brown trunk (see Figure 3).
Height of lowest phytotelm
Continuous Measured from the ground to the point on the main stem where the lowest phytotelm protrudes (see Figure 5 and 8). (This is usually only a few centimeters higher than the “trunk height” measurement.)
Height of highest phytotelm
Continuous Measured from the ground to the point on the main stem where the highest phytotelm protrudes (see Figure 5 and 8).
Diameter at breast height (DBH)
Continuous Measured using a DBH tape around the top of the trunk indicated by a color change between the green leaf stalks and brown trunk (the same point on the trunk used to measure trunk height) (see Figure 3). This standard was used because not all the plants were breast height, but all had leaves protruding from a central trunk.
Length of petiole Continuous Measured from the base of the leaf to the connection point between the petiole and the trunk (see Figure 7 and 8). The longest petiole of the plant was measured.
Length of leaf Continuous Measured from the base of the leaf where the petiole connects to the tip of the leaf. The largest leaf on the plant was measured (see Figure 7).
Number of leaf axils Continuous A leaf axil was only counted if it had the ability to hold water (see Figure 5 and 10).
Number of leaves Continuous A leaf was counted if the tip protruded at least 30 cm from the top of the main stem (see Figure 5).
Nearest X. jacquinii neighbor
Continuous Distance from the trunk of the measured X. jacquinii to the trunk of the nearest X. jacquinii neighbor
Nearest neighbor (non-X. jacquinii)
Continuous Distance from the trunk of the measured X. jacquinii to any part of the nearest non-X. jacquinii neighbor
Distance to water Continuous Distance from the trunk of the measured X. jacquinii to the edge of the nearest stream bed

-‐27-‐
Categorical variables included a qualitative estimate of forest canopy cover over
the plant, detritus levels in the leaf axils, invertebrate abundance, and the volume of water
contained within the phytotelm relative to its potential capacity. All of these categorical
variables were measured on a 0-4 scale. In addition, notes were taken on the relative
types of invertebrates and their sizes (larger or smaller than the 7 mm cutoff used by
Caldwell and Araujo (2004) to distinguish between potential competitors or food sources
for the adults). The depth of the water within the phytotelm was also recorded. This
measurement was taken from the base of the phytotelm to the surface of the water,
rounded to the nearest half-centimeter. If the plant was occupied, the specific phytotelm
the F. fitzgeraldi individual inhabited was measured. However, if the plant was
unoccupied, an average of the water level and detritus level in all of the phytotelms
collectively was estimated and recorded.
Water chemistry data was also recorded using a calibrated Oakton 300 series
Dissolved Oxygen/pH/Temperature Meter. Dissolved oxygen was measured in both
mg/L and percent. In addition, pH and water temperature were also recorded. An eye-
dropper was used to extract approximately 25-30 mL of water from the phytotelms and
this water was transferred to the rubber cap normally used to cover the probe of the
Meter. After the probe was inserted into this cap, readings were taken and recorded. If
the plants were occupied, only water from the inhabited phytotelm was measured. If the
plant was unoccupied, the eye-dropper was used to take samples from all of the
phytotelms and this water was all mixed together in the rubber cap before the probe was
inserted.

-‐28-‐
Morphological measurements of the plant included trunk height, highest and
lowest phytotelm heights, longest petiole length, length of largest leaf, number of leaves
and number of leaf axils (see Figures 7 and 8). In addition, several other variables
including distance to the nearest X. jacquinii neighbor, distance to nearest neighboring
plant of any species other than X. jacquinii, and distance to the nearest stream were also
recorded to provide more insight into the surrounding habitat. Collectively, these data
were used to characterize the microhabitat for F. fitzgeraldi.
Figure 7. The length of the petiole and the length of the leaf were measured. For each plant, the longest petiole was measured. Likewise, the largest leaf was also measured. Typically these were correlated so that the longest petiole had the largest leaf.
Figure 8. Phytotelm formed at the base a leaf axil in X. jacquinii. The star indicates the reference point used in the following measurements: height of highest phytotelm, height of lowest phytotelm, and length of the petiole.

-‐29-‐
Selecting variables for the multiple logistic regression model
The Spearman Test for correlation was used to identify any variables that had a
correlation coefficient over 0.7, the cutoff used for this model. Any variables more
highly correlated than 0.7 were no longer considered independent, and thus eliminated
from the model.
Invertebrate density, invertebrate presence or absence, and invertebrate size were
all highly correlated with each other. Ultimately, invertebrate size was chosen to remain
because it represents both an objective measure of invertebrate presence and contains
information indicating the relative size of the invertebrates. Therefore, this variable may
provide insight into the role of the invertebrates, acting as either predators (large
invertebrates >7 mm) or sources of food for the adult F. fitzgeraldi (small invertebrates
<7 mm) (Caldwell and Araujo 2004).
In addition, the relative volume of water in the phytotelm was highly correlated to
the depth of the water in the phytotelm. In theory, these variables represent very different
measurements. The “volume of water” measurement represents the amount of water
within the phytotelm relative to the phytotelms capacity to hold more water. For
example, a phytotelm that was half-filled to its capacity was recorded as a “2” on the 0-4
categorical scale, while a phytotelm filled to its brim with water was recorded as a “4”.
A dry phytotelm was recorded as a “0”, and a phytotelm with water taking up about 25%
or 75% of its total volume was recorded as either a “1” or “3”, respectively. In contrast,
the “depth of water” measurement was a continuous variable representing the actual
depth of the water in the phytotelm, measured from the base of the phytotelm to the

-‐30-‐
surface of the water and estimated to the nearest half-centimeter. However, the “volume
of water” and “depth of water” variables were highly correlated. Ultimately, the “depth
of water” variable was chosen to remain because it represents a more precise, quantitative
estimate of the actual volume of water present in the phytotelms, considering they range
slightly in size.
The dissolved oxygen was recorded in both mg O2/L water and percent O2 within
the water. Because these variables were highly correlated, the variable representing the
measurement in mg O2/L water was chosen because it represents a more quantitative
measurement of the amount of oxygen present in the water.
The trunk height, height of the lowest phytotelm, and height of the highest
phytotelm were also highly correlated. Trunk height was used to represent all of these
variables.
In addition, the number of leaves and the number of leaf axils were highly
correlated. Because some leaves protruded vertically from the top of the plant, they did
not have an axil that could potentially fill with water. Therefore, the variable
representing the number of leaf axils was chosen to remain in the model for further
analysis because it better represents the actual number of potential phytotelms available
for F. fitzgeraldi to inhabit.
After eliminating seven highly correlated variables, fifteen independent variables
remained to be included in the multiple logistic regression model.

-‐31-‐
Preliminary testing
When a multiple logistic regression model was used to analyze the effect of both
the categorical and continuous variables on the single dependent variable (presence or
absence of F. fitzgeraldi), multiple error messages occurred in the SPSS program.
Therefore, a multiple logistic regression model was used to separately analyze the effect
of only the categorical variables (canopy cover, invertebrate size, and detritus) on all 106
plants. This preliminary analysis did not provide any evidence supporting the
significance of these variables (canopy cover: p=0.466, invertebrate size: p=0.638,
detritus: p=0.934). Therefore, the categorical variables were removed from the data set
and the continuous variables were analyzed separately.

-‐32-‐
RESULTS
The ecological niche of F. fitzgeraldi was defined and described using both
categorical and continuous variables. The baseline data provide insight into the range of
conditions the frog faced and the relative frequency that these conditions occurred in this
study (see Table 2 and 3). Although all variables measured during the data collection
period are included in these tables, not all of these variables were included in the final
analysis (see “Selecting Variables for the Multiple Logistic Regression Model” section).
Table 2. Descriptive statistics document the range of conditions that would be encountered by the frog if it were to occupy a phytotelm in the observed plant. Only the continuous variables are included in this table. Because the leaf axils in eight plants were dry, the aquatic variables could not be observed. Note the tight range and low standard deviation for these aquatic variables, in contrast to the relatively wide ranges observed for other variables. Variable n Range Average Standard
Deviation Depth of water 106 0-6 cm 1.7 cm 1.2 cm Oxygen concentration of water 98 90.4%-100% 97.5% 2.4% Oxygen concentration of water 98 7.22 - 8.54 mg O2/L 7.90 mg O2/L 0.29 mg O2/L Temperature of water 98 21.0-28.7°C 26.2°C 2.1°C pH of water 98 4.36-7.30 5.99 .69 Trunk height 106 1-190 cm 55 cm 39 cm Height of lowest phytotelm 106 1-193 cm 63 cm 40 cm Height of highest phytotelm 106 14-200 cm 78 cm 41 cm Diameter at breast height (DBH) 106 4.5-20 cm 7.9 cm 2.44 cm Length of petiole 106 54-197 cm 101 cm 23 cm Length of leaf 106 27-57 cm 41 cm 7 cm Number of leaf axils 106 2-14 6 2.5 Number of leaves 106 3-16 7 2.8 Nearest X. jacquinii neighbor 106 6-260 cm 70 cm 48 cm Nearest neighbor (representing any species other than X. jacquinii)
106 1-120 cm 34 cm 25 cm
Distance to water 106 0-20 m 4.9 m 4.1 m

-‐33-‐
Table 3. Several categorical variables were also used to document the range of conditions that would be encountered by the frog if it were to occupy a phytotelm in the observed plant. Each categorical variable was recorded according to the described scale. For each variable, the number of plants that fit into each category was recorded. Variable n 0 1 2 3 4 Notes Canopy cover
106 52 51 2 0 1 (0-4 scale), 0=100% canopy cover, 4=0% canopy cover
Detritus 106 2 22 31 38 13 (0-4 scale), 0=leaf axils completely full of detritus, 4=leaf axils devoid of detritus
Invertebrate density
106 14 50 17 19 6 (0-4 scale), 0=no invertebrates, 4=very high density of invertebrates.
Invertebrate presence
106 14 92 0=no invertebrates, 1=invertebrates present
Invertebrate size
106 14 54 28 10 0=no invertebrates, 1=only small invertebrates present (<7mm), 2=only large invertebrates present (>7mm), 3=both small (<7mm) and large (>7mm) invertebrates present
Volume of water
106 8 49 32 14 3 (0-4 scale), 0=no water in phytotelm, 4=phytotelm completely full of water
Between both sites, 106 plants were sampled for F. fitzgeraldi. Of these plants,
11 contained frogs (see Table A1). Because the leaf axils in eight of the plants were dry,
water chemistry data could not be taken. After running the multiple logistic regression
model on the categorical variables using all 106 plants, the eight dry plants were removed
from the model to complete further analysis.
A multiple logistic regression model was run using the remaining continuous
variables on the 98 plants containing water in at least one leaf axil (see Table 4). Both
the Omnibus Test of Model Coefficients (p<0.001) and the Hosmer and Lemeshow Test
(p=0.837) indicate that this model was a significant predictor of the presence or absence
of F. fitzgeraldi in X. jacquinii plants.

-‐34-‐
Table 4: This multiple logistic regression model analyzed whether the continuous variables were significant predictors of the frog’s presence or absence. This model found that water depth was the only significant predictor of the frog’s occupancy status. The B value represents the coefficient calculated by the model for each variable (positive coefficients indicate that the likelihood of occupancy increases as the continuous variable increases in value, negative coefficients indicate that the likelihood of occupancy decreases as the continuous variable increases in value). The Wald values provide insight into the significance of the variable in the model. The p-values indicate the statistical significance of each variable. The Odds ratio indicates how the odds of finding a frog changes as the continuous variable increases by one unit. If the Odds ratio is >1, frogs are more likely to be found as the continuous variable increases. If the Odds ratio is <1, the chances of finding a frog decreases as the continuous variable increases. See Table 1 for definitions of variables. Independent variable
B Wald P-value
Odds ratio
Water depth 2.761 5.185 .023* 15.819 Height .005 .120 .729 1.005 Length of petiole .072 3.182 .074 1.075 Length of leaf -.111 .461 .497 .895 Number of axils .315 .823 .364 1.371 DBH -.880 2.449 .118 .415 Neighbor (X. jac.) .015 .536 .464 1.015 Neighbor (other) -.020 .492 .483 .980 Distance to water .002 1.615 .204 1.002 Dissolved O2 (mg/L) 11.970 1.919 .166 158003.132 Water temperature .338 .181 .670 1.402 pH of water .125 .019 .890 1.133

-‐35-‐
Water depth (p=0.023) was the only significant predictor of habitat occupancy (see
Figure 9a). Plants with phytotelms containing a greater volume of water were more likely
to be inhabited by F. fitzgeraldi than plants with phytotelms containing a lesser volume of
water, as measured by the depth of the phytotelm. The length of the plant’s longest petiole
was marginally significant (p=0.074) (see Figure 9b). As the length of the petiole
increased, the odds of finding F. fitzgeraldi in the plant also increased. DBH was the third
highest predictor of the presence or absence of frogs (p=0.118) (see Figure 9c).
Flectonotus fitzgeraldi were more likely to be found in plants with a smaller DBH.
Although dissolved oxygen (measured in mg O2/L) is the fourth highest predictor of frog
occupancy in this model, this variable was not significant (p=0.166) (see Figure 9d). If this
trend was more significant, it would suggest that a higher level of oxygen in the water is
beneficial for the frogs. None of the remaining continuous variables were significant.
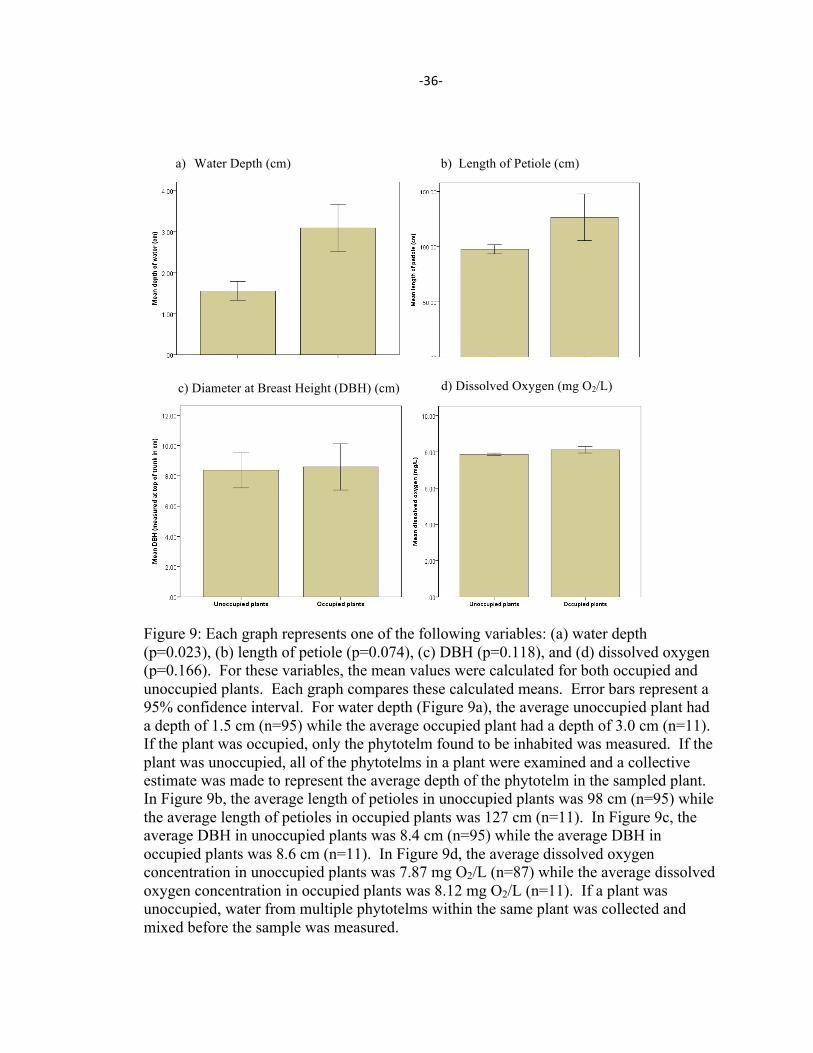
-‐36-‐
Figure 9: Each graph represents one of the following variables: (a) water depth (p=0.023), (b) length of petiole (p=0.074), (c) DBH (p=0.118), and (d) dissolved oxygen (p=0.166). For these variables, the mean values were calculated for both occupied and unoccupied plants. Each graph compares these calculated means. Error bars represent a 95% confidence interval. For water depth (Figure 9a), the average unoccupied plant had a depth of 1.5 cm (n=95) while the average occupied plant had a depth of 3.0 cm (n=11). If the plant was occupied, only the phytotelm found to be inhabited was measured. If the plant was unoccupied, all of the phytotelms in a plant were examined and a collective estimate was made to represent the average depth of the phytotelm in the sampled plant. In Figure 9b, the average length of petioles in unoccupied plants was 98 cm (n=95) while the average length of petioles in occupied plants was 127 cm (n=11). In Figure 9c, the average DBH in unoccupied plants was 8.4 cm (n=95) while the average DBH in occupied plants was 8.6 cm (n=11). In Figure 9d, the average dissolved oxygen concentration in unoccupied plants was 7.87 mg O2/L (n=87) while the average dissolved oxygen concentration in occupied plants was 8.12 mg O2/L (n=11). If a plant was unoccupied, water from multiple phytotelms within the same plant was collected and mixed before the sample was measured.
a) Water Depth (cm) b) Length of Petiole (cm)
c) Diameter at Breast Height (DBH) (cm) d) Dissolved Oxygen (mg O2/L)

-‐37-‐
DISCUSSION
Habitat choice by Flectonotus fitzgeraldi was primarily driven by the availability
of water. It makes sense that the depth of water in the phytotelm was an important factor
influencing the choice of the frog because this water is essential for the development of
the tadpole (Kenny 1969). If tadpoles are deposited into a phytotelm with very little
water, the probability of desiccation is higher (Caldwell and Araujo 2004, Denver 1998,
Rudolf and Rodel 2007). Because tadpole survival is directly dependent on a constant
water supply, habitats that provide a greater assurance that desiccation will not occur are
favored (Caldwell and Araujo 2004). Thus, the ability of the frog to detect this larger
water volume when depositing its offspring should increase the individual’s overall
fitness. Data from this study suggest that water availability serves as the best indicator of
habitat quality, as water depth was the only significant predictor of the occupancy of the
frogs (p=0.023).
In addition to assuring an appropriate habitat for tadpoles to develop, phytotelms
with comparatively more water also seem to provide a microhabitat favored by adults. In
fact, ten of the eleven occupied plants contained adults. Although very little is known
about the natural history or physiological requirements of F. fitzgeraldi, this occupancy
pattern seems to suggest that water-filled leaf axils are also beneficial for adults to
inhabit. This observation is consistent with previous studies which have shown that an
increased water volume positively influences the likelihood of occupancy for other adult
phytotelm-dwelling frogs (Lehtinen and Carfagno 2011, Silva et al. 2011). For example,
Lehtinen and Carfagno (2011) found that the volume of water in the leaf axils of

-‐38-‐
Pandanus plants was one of the best predictors of habitat occupancy for both
Guibemantis bicalcaratus and G. punctatus of Madagascar. In addition, Silvia et al.
(2011) concluded that bromeliad occupancy is an adaptation to water scarcity for adults
of several frog species in Brazil. Data from my study suggests that adult F. fitzgeraldi
also tend to inhabit phytotelms with larger volumes of water. It seems advantageous for
mature females to select phytotelms with more water to inhabit, as tadpole survival
depends on water availability (Kenny 1969). Males may also use water volume as a cue
to select quality habitats in an effort to attract females. If females are attracted to the
quality habitats males are calling from, it may improve the overall fitness for both
individuals. This could simultaneously provide the male with the opportunity to mate
and allow the female to occupy a quality habitat, thus improving the chances of survival
for her future offspring.
In this study, three of eleven plants found to be occupied by F. fitzgeraldi
contained multiple individuals. In every case, all of the individuals occupied the same
leaf axil. Although very little is known about amplexus or other mating behaviors of F.
fitzgeraldi, it seems likely that these behaviors occur within phytotelms. In fact, all of the
phytotelms occupied by multiple adult individuals contained at least one male and one
female. A larger volume of water may provide a larger physical space for these activities
to occur. Therefore, an increased water depth may improve the quality of the
microhabitat for mating purposes and thus serve as an important cue for the frog to
detect.

-‐39-‐
An increased water depth may also provide more protection against certain
environmental conditions, biotic intrusions, or other detrimental outside factors. In
addition to decreasing the chance of desiccation, a larger volume of water may better
resist rapid temperature changes due to fluctuating environmental temperatures (Lin and
Kam 2008, Paradise 2004, Sota et al. 1994). Although the overall environmental
conditions are not likely to vary too drastically in Tobago, the increased water depth may
still reduce the variation the frogs experience, even if only on a small scale. Personal
observation showed frogs were often found wedged tightly between the petiole and the
trunk at the very base of the phytotelm. In fact, many frogs wedged themselves even
deeper upon discovery. These frogs seemed to be seeking maximum protection from the
sudden intrusion. Therefore, a larger volume of water may increase the quality of the
frog’s microhabitat by increasing the spatial barrier between the frog and its environment.
Past research has shown that canopy cover and detritus levels may be correlated
with water volumes. For example, less canopy cover may increase the chances of
capturing new rain while simultaneously decreasing the chances that detritus will fall into
the phytotelm (Lehtinen 2004, Silva et al. 2011, Lehtinen and Carfagno 2011, Lin and
Kam 2008). New rain is beneficial as it helps increase the dissolved oxygen levels
(Caldwell and Araujo 2004). One study found that bromeliads located in the sun had
430% higher dissolved oxygen levels than bromeliads located in the shade (Silva et al.
2011). Silva et al. (2011) suggested that this variation was in part because the bromeliads
in the sun were able to collect fresh, oxygenated rainwater. Studies have found that an
increased level of detritus decreases the level of dissolved oxygen because microbes

-‐40-‐
depend on this oxygen supply to decompose organic matter (Lehtinen 2004, Diaz and
Rosenberg 2008). Considering neither adult F. fitzgeraldi frogs nor tadpoles consume
organic matter, high levels of detritus are presumably not a beneficial characteristic for
the frog (Kenny 1969). The amount of detritus contained in a plant has been shown to
be negatively correlated with a plant’s water-holding capacity (Lehtinen and Carfagno
2011). Lehtinen and Carfagno (2011) suggest that G. punctatus probably uses detritus
load as a cue for water holding capacity, as plants under a relatively dense canopy collect
more detritus and receive less rainfall than plants under a more open canopy (Lehtinen
2004). Therefore, an increased volume of water in the phytotelm may be a direct result
of several other cumulatively important environmental conditions including decreased
canopy cover and decreased levels of detritus. Although these studies suggest both
canopy cover and detritus may be important factors to consider when predicting the
quality of the microhabitat, my study did not support this hypothesis. Neither canopy
cover (p=0.466) nor detritus (p=0.934) were individually significant in predicting the
occupancy of F. fitzgeraldi in X. jacquinii.
Although water availability was the primary driving force for habitat selection in
F. fitzgeraldi, petiole length was also a marginally significant variable (p=0.074).
Although not significant by the 0.05 standard assumed by this study, other pilot studies
have used a 0.1 critical point to figure out where to focus their attention to maximize the
chances of finding significant data in future follow-up research. Flectonotus fitzgeraldi
seemed more likely to occupy Xanthosoma jacquinii with longer petioles, but it is not
likely that they perceived this cue and responded to it directly. Instead, the petiole length

-‐41-‐
may be correlated with the relative age of the leaf, which may in turn affect the quality of
the phytotelm (see Figure 10). Personal observations showed that younger leaves
emerging from the top of the plant had shorter petioles, as they were still growing.
Because these younger leaves often pointed directly upward, the size of the leaf axil was
compromised and thus the phytotelm’s potential to hold water decreased dramatically. In
addition, older leaves located lowest on the trunk also seemed to have shorter petioles
and likely smaller phytotelms as a result, because they emerged when the plant was
smaller. Personal observations showed that phytotelms resulting from older leaves often
appeared stretched out because as the trunk grew wider with age, the depth of the
phytotelm decreased (see Figure 10a). As a result, the phytotelm was not able to hold as
much water. Furthermore, the older leaves often bent downwards with age, ultimately
draining the phytotelm and destroying the microcosm (see Figure 10c). Therefore,
middle-aged leaves with longer petiole lengths seem to be correlated with larger, more
stable phytotelms. However, it is important to note that data measuring multiple petioles
on the same plant were not taken. Therefore, this suggested explanation is only based on
informal observations of the plant and future studies are needed to support the logic of
this argument.

-‐42-‐
A) B) C) Figure 10. Phytotelms created by leaf axils. A) Represents a phytotelm created by a relatively new, young leaf. The petiole is generally thinner, oriented more vertically, and contains a relatively small, though often fairly deep phytotelm. B) Represents a “middle-aged” leaf. Petioles are typically longer and a bit wider at the base than younger leaves. Petioles are also not oriented as vertically as newer petioles, widening the phytotelm. Although this may create an overall larger phytotelm, the more vertical younger leaves may still create a deeper, though smaller phytotelm. C) Represents the leaf axil created by an aging leaf. With age, the petiole bends downward, eventually inhibiting the leaf axil’s ability to hold water. At this point, it is no longer considered a phytotelm, and thus not considered in the analysis.
Alternatively, it is possible that longer petioles are associated with larger
phytotelms. Although this idea cannot be directly supported from my study (considering
the examined leaf axils were never directly measured for size), longer petioles may be
necessarily wider at the base to structurally hold the weight of the leaf (see Figure 7). If
this is the case, the resulting phytotelm will also be larger. Because larger phytotelms
can hold more water, and water depth was found to be the only significant predictor of
frog occupancy, it is possible that longer petioles are associated with higher quality
phytotelms.
Furthermore, the overall shape of the plant is largely dependent on the length of
the petioles, as longer petioles allow the leaves to reach higher and extend farther
horizontally (see Figure 3). Therefore, petiole length may also be associated with overall
plant size, although height and DBH may also be used to represent the size of the plant.

-‐43-‐
A large plant may be easier to locate in the ecosystem because its leaves extend farther
and it may be possible that as the frog moves between plants in the population, long
petioles increase the frog’s likelihood of encountering the plant.
While not significant, it is worth noting that the third highest explanatory variable
in the analysis was diameter at breast height (DBH) (p=0.118). Although minimal, this
variable still has some predictor value and our analysis suggests that frogs may slightly
favor plants with smaller diameters. If smaller diameters are associated with smaller
phytotelms, it may be possible that microhabitats resulting from plants with smaller
diameters are large enough for frogs but too small for larger predators or competitors
including snakes, crabs, or other large organisms. Therefore, plants with smaller
diameters may be beneficial to the frog and thus increase the quality of the plant.
However, the hypothesis that smaller phytotelms are beneficial to the frog directly
contradicts the previously described idea that larger phytotelms are more favored. In
fact, it may be possible that neither DBH nor petiole length is actually directly correlated
to phytotelm size. It may be possible for plants to have both large petioles and small
diameters, as these characteristics are not necessarily mutually exclusive. Furthermore,
the petiole length might be primarily an indicator of phytotelm quality based on the
relative age of the leaf, while DBH might provide more insight into the actual dimensions
of the leaf axil. However, data recording the physical size of the phytotelm was not taken
in this study. Therefore, claims suggesting either petiole length or DBH are correlated
with phytotelm size are not adequately supported by data and only speculative in nature.
Petiole length and DBH were positively correlated with each other based on the

-‐44-‐
Spearman test (correlation coefficient=0.647, p<0.001). Although this suggests that as
petiole length increases, DBH also increases, the Spearman correlation coefficient did not
exceed 0.7, our cutoff for the multiple logistic regression model. Therefore, the variables
were treated independently. When examined individually, the DBH was a greater
predictor of frog occupancy (odds ratio: 0.415) than petiole length (odds ratio: 1.075)
(see Table 4). Ultimately, it is important to remember that neither trend was actually
significant.
The amount of dissolved oxygen in the phytotelm does not significantly predict
frog occupancy in this study (p=0.166). Because frogs have highly permeable skin
essential for cutaneous respiration, it seems possible that dissolved oxygen levels could
affect their physiology (Caldwell and Araujo 2004). Although the relatively large surface
area to volume ratio of water in the phytotelm likely decreases the time needed for the
dissolved oxygen to equilibrate with the environment, dissolved oxygen levels may still
vary greatly. One study found that dissolved oxygen levels varied by 430% within
tropical phytotelmata located in bromeliads in Brazil (Guimaraes-Souza et al. 2006). The
same study suggested that frogs assess the quality of their environment primarily through
cues from water chemistry. In fact, past research has observed frogs presumably
assessing the water chemistry of a small pool of water before selecting a quality habitat
and accompanying mate with which to spawn (Alves-Silva and Silva 2009, Downie et al.
2001). Caldwell and Araujo (2004) hypothesized that oxygen depletion was the reason
for mortality for several Allobates femoralis tadpoles living in phytotelms created by fruit
shells of Bertholletia excelsa, the Brazil nut tree. This study also found that Bufo

-‐45-‐
castaneoticus were revived after a heavy rainfall, and thus their lethargic behavior and
fact that they floated upside down on the water’s surface before the rain could be
attributed to oxygen depletion. These observations suggest that higher levels of dissolved
oxygen in the water of the phytotelms would increase the quality of the microhabitat.
However, data from several other studies have found that water chemistry does not
significantly affect the quality of the phytotelm (Lehtinen 2004, Caldwell and Araujo
2004). Data from our analysis regarding F. fitzgeraldi supports the latter conclusion.
Previous studies have focused on the effect of rainfall on both dissolved oxygen
and amount of detritus in the phytotelms. This research has shown that phytotelmata
containing only rainwater consistently have higher concentrations of dissolved oxygen
than phytotelmata containing mostly detritus (Caldwell and Araujo 2004). These
variables are often related to each other because new rainwater may flush a phytotelm of
some of its accumulated detritus. In addition, an increased detritus often increases the
amount of microbial activity, in turn lowering the concentration of dissolved oxygen
(Lehtinen 2004, Caldwell and Araujo 2004, Ryan and Barry 2011). However, my study
found that dissolved oxygen concentration is not significantly correlated to detritus level
(correlation coefficient: -0.055, p=0.591). Furthermore, dissolved oxygen was also not
correlated with water depth (correlation coefficient: 0.132, p=0.196), contrary to the idea
that rainfall would increase both dissolved oxygen level and water depth simultaneously.
Despite the perceived challenges of dealing with a low dissolved oxygen concentration,
one study found that two phytotelm-breeding frogs in Madagascar, Mantidactylus
bicalcaratus and M. punctatus, actually have higher growth rates in the low oxygen

-‐46-‐
levels (Lehtinen 2004). This is likely due to the fact that detritus served as a food source
for the tadpoles of these species and this food source was more beneficial than the
decreased levels of dissolved oxygen were harmful (Lehtinen 2004). Because F.
fitzgeraldi tadpoles are supplied with a large yolk and do not feed (Kenny 1969), high
levels of detritus were not expected to significantly increase the quality of the phytotelm.
Furthermore, once tadpoles reach a certain stage of development, they possess functional
lungs and can gulp air directly at the surface even if there is no oxygen in the water at all
(Lehtinen 2004). Although a decreased level of dissolved oxygen may decrease the
quality of the habitat for some species of frogs, data from our study concluded that
dissolved oxygen levels were not significant predictors of F. fitzgeraldi occupancy in
phytotelms created by X. jacquinii.
In addition to dissolved oxygen, my study concluded that neither pH nor water
temperature had any notable effect in our model of frog occupancy. Previous studies
have suggested contrasting views on the importance of pH and water temperature in
microcosm selection. A decreased pH increased the mortality rate and delayed
metamorphosis for Rana temporaria (Cummins 1986). Elevated water temperature was
also found to be a significant aquatic factor, accelerating the development of several
species of tadpoles in Virginia, including Rana temporaria (Smith-Gill and Berven 1979,
Laurila and Kujasalo 1999). Although both these studies and additional research state
that cues from the aquatic variables may be important factors contributing to the fitness
of the frog and thus quality of the phytotelm (Guimaraes-Souza et al. 2006, Silva et al.

-‐47-‐
2011), these variables were not shown to be significant in other research (Caldwell and
Araujo 2004, Lehtinen 2004). My study supports the latter conclusion.
However, it is worth noting that the aquatic variables in this study had a relatively
small overall range (see Table 2). Therefore, the observation that frogs did not seem to
distinguish between certain aquatic variables may simply be due to the fact that very little
variation existed in the water temperature and chemistry of the measured phytotelms.
Flectonotus fitzgeraldi in this study seemed to have no choice but to inhabit a relatively
narrow niche when considering aquatic data alone. Therefore, this study found that
aquatic variables are not a valuable dimension on which to describe the ecological niche
of this phytotelm-dwelling species. However, further research may show a greater
variation in the properties of water in phytotelms located in different plant species or over
a larger geographic range in Tobago. Until this variation is found, the ability of F.
fitzgeraldi to tolerate a wide range within the aquatic variables remains unknown.
Invertebrates were recorded for presence, abundance, and relative size. A variety
of ants, crickets, spiders, millipedes, snails, earwigs, and scorpions were found in the
observed plants. Crabs and (although not invertebrates) two snakes (both Imantodes
cenchoa) were also found during informal surveys of X. jacquinii during the collection
period in 2012. My study found that the presence of these invertebrates did not
significantly predict the occupancy of the frog. This contradicts previous research
suggesting that invertebrates may decrease the overall quality of the habitat by acting as
either predators or competitors to the frogs. Caldwell and Araujo (2004) showed that
when potential insect predators were small (<7mm), they were eaten by Dendrobates

-‐48-‐
tadpoles. Although F. fitzgeraldi tadpoles are non-feeding, it may be assumed that the
adults also eat small invertebrates based on observations of related frog species (Elinson
1990). However, if Dendrobates tadpoles were deposited in a water pool with a larger
predaceous invertebrate or larvae already present, the Dendrobates tadpoles were usually
eaten first (Caldwell 1993 and Caldwell and Araujo 2004). This is probably true for F.
fitzgeraldi tadpoles as well. In addition, the invertebrates may act as competitors for the
same resources the phytotelms provide (Caldwell and Araujo 2004, Murphy 2003,
Williams et al. 2007). Despite this research, my data do not show any significant
relationship between invertebrate presence and frog occupancy. Furthermore, neither
invertebrate presence nor size is correlated to any other factors in the analysis.
Other studies suggest that competition between different species of phytotelm-
breeding frogs may be a significant factor to consider when defining the niche of a single
species (Lehtinen and Carfagno 2011). However, as far as we know, F. fitzgeraldi is the
only obligate phytotelm-occupying frog species in Tobago. This study did not consider
the interactions between different species of amphibians, as no other species were found.
It is possible, however, that at least one species of snake (Imantodes cenchoa) is a
predator of F. fitzgeraldi. This hypothesis is based on the fact that two of these snakes
were observed occupying X. jacquinii phytotelms during informal surveys of the sites.
Past research suggests that there may be some relationship between the quality of
the phytotelm and either plant size or overall shape. Although two morphological
variables (petiole length and DBH) were already discussed, the baseline data collected for
this study collectively give a more holistic image of X. jacquinii. Even though Silva et

-‐49-‐
al. (2011) did not examine X. jacquinii specifically, the authors state that a plant’s large
size may be caused by a number of factors that positively influence its growth including
decreased canopy cover, decreased detritus, and increased rainfall (Silva et al. 2011).
Furthermore, sun exposure has been shown to support larger, hardier plants, providing a
larger, more stable phytotelm for the frog to inhabit in both bromeliads in Brazil (Silva et
al. 2011) and bamboo stumps in Taiwan (Lin and Kam 2008). For G. bicalcaratus in
Madagascar, occupation rate increased with increasing plant width. In contrast, the same
study found that G. punctatus were more commonly found in tall plants (Lehtinen and
Carfagno 2011). Some studies have observed the general shape of a plant to be an
important abiotic factor in determining the quality of the plant because shape affects both
the amount of water a leaf axil can hold (Zotz and Thomas 1999) and the quality and
stability of each microhabitat created (Silva et al. 2011). Although there are different
ways to define and describe a plant’s “shape”, Zotz and Thomas (1999) based their
definition for bromeliad shape on three factors: tank capacity, catchment area for
precipitation, and aspects of tank geometry that affect evaporation. Silva et al. (2011)
observed that bromeliad shape was often determined by its position within a larger
population. Individuals located on the edge of a cluster were generally bulkier and
shorter, with lighter green leaves, stronger thorns, more access to sunlight, and less
accumulation of detritus. These bromeliads were observed to be more desirable to frogs
because the general shape allowed for a larger volume of water to be collected between
the leaves, likely reducing desiccation rates. However, these studies of bromeliads
provided little insight into how the morphological measurements of X. jacquinii might

-‐50-‐
affect the quality of the resulting phytotelms simply due to the structural differences
between the plant species.
Because this is the first study to analyze the relationship between the
morphological measurements of X. jacquinii and the resulting characteristics of its
phytotelms, extensive baseline data were taken (see Table 1). However, only
independent variables were used in the Multiple Logistic Regression model. Therefore,
some of these original variables were excluded (see “Selecting Variables for the Multiple
Logistic Regression Model” in Methods section Table 1). Although two of the top three
predictors of F. fitzgeraldi occupancy (including petiole length and DBH) were
morphological characteristics of the plant, no other notable relationships or correlations
were found between any of the other morphological measurements in relation to frog
occupancy.
Although data quantifying several aspects of the plant’s spatial location were
taken, these variables were also not significant predictors of frog occupancy.
Specifically, the distance was measured from the X. jacquinii containing the observed
phytotelm to the nearest other X. jacquinii and nearest plant of any species (other than X.
jacquinii). This was designed to provide insight into the relative density of the X.
jacquinii population in relation to the density of surrounding plant species. However, the
total area the patch covered was not measured, therefore exact densities cannot be
calculated. Personal observations showed that more densely populated X. jacquinii
populations tended to have fewer plants representing different species distributed
amongst the X. jacquinii individuals. Furthermore, X. jacquinii populations that were

-‐51-‐
spread over a wider geographic area tended to have more plants of other species
distributed throughout the range of the observed X. jacquinii patches. Although this trend
seems to be characteristic to the populations of X. jacquinii in this study, neither these
informal observations nor the statistical model suggest these factors are significant when
predicting frog occupancy. In addition, the distance from the measured X. jacquinii to
the nearest stream was also recorded, although this variable was not a significant
predictor of the frog’s occupancy status. This makes sense, as F. fitzgeraldi does not live
directly in the stream during any part of its life cycle, instead relying solely on the
phytotelms for breeding and seeking shelter (Murphy 1997).
Several studies have attempted to explore the relationship between environmental
variables, plant morphology, and the local distribution of frogs (Xavier and Napoli 2011,
Lehtinen and Carfagno 2011, Silva et al. 2011). These studies are based on the
assumption that environmental variables affect plant morphology which in turn affect the
characteristics of the phytotelms. In addition, the studies assume that the frogs will
occupy the phytotelm that maximizes their evolutionary fitness. Similar to the present
study, Silva et al. (2011) sought to explore which biotic or abiotic conditions ultimately
influence the quality of this microhabitat. Morphological characteristics of the plant,
water chemistry, and spatial information were all considered. Silva et al. (2011)
concluded that frogs were more likely to be found under certain conditions that could be
characterized by morphological traits. For example, bromeliads located around the edges
of clusters of multiple plants were shown to be particularly well-inhabited. The authors
rationalized that the overall shape of the edge plants allowed for larger phytotelms in

-‐52-‐
contrast to the relatively crowded bromeliads located in the middle of the patch.
Therefore, occupancy patterns observed on the small scale may be used to provide insight
into occupancy or distribution patterns on a larger scale.
Although it seems safe to assume that factors affecting X. jacquinii distribution
will affect F. fitzgeraldi distribution on some level, the frog distribution cannot be
directly predicted from the distribution of the plants they inhabit (Silva et al. 2011,
Xavier and Napoli 2011). Even though a better understanding of the plant distribution
may provide some insight into the frog geographic distribution, the ranges of both the
plant and the frog species are unlikely to be perfectly correlated. Therefore, extensive
additional data relating to occupancy patterns for F. fitzgeraldi on both the small and
large scale are needed before any real distributional predictions can be made.
My study found that F. fitzgeraldi has a relatively broad ecological niche (see
Table 2 and 3). Because the frogs were found to inhabit phytotelms characterized by a
wide variety of environmental and biotic conditions, it seems safe to conclude that the
species has the ability to withstand a large range of ecological conditions. In fact, my
data showed that water depth was the only significant variable predicting the occupancy
of frogs. Therefore, although habitat selection does seem to occur on some level, the
broad ecological niche of F. fitzgeraldi allows the frog to occupy a wide variety of
phytotelms. As long as the phytotelm provides an adequate water supply, my study
suggests that F. fitzgeraldi is rather flexible in its choice of phytotelm. Therefore, it
might be possible for F. fitzgeraldi to occupy other types of phytotelms including
treeholes, bamboo stumps, bromeliads, or phytotelms created by other plants if human

-‐53-‐
disturbances threaten X. jacquinii. From a conservation perspective, this provides hope
for the frog’s long term survival despite possible changes to surrounding environmental
or ecological conditions.

-‐54-‐
ACKNOWLEDGEMENTS
I gratefully acknowledge the support and guidance I’ve received from a large
number of people throughout the course of this project. First, I’d like to thank Dr.
Lehtinen. To say he’s been a very special mentor over the past three years would be an
incredible understatement. The important lessons I’ve learned and memories I have of
our time together will stay with me for many years to come. I would also like to thank
Dr. Loveless for teaching me more than I imagined I’d learn throughout the whole editing
process. I truly believe her new perspective and helpful insight has proven invaluable in
many significant ways, and I thank her for all of her patience. I am also very thankful for
Dr. Fraga’s insight, and for his wise listening ear. Of course, I am greatly indebted to the
College of Wooster Biology Department for providing me with this amazing opportunity
in the first place, and for the maintinence and use of field equipment. In addition, the
Copeland Fund provided monetary support, making my adventure to Trinidad and
Tobago possible. Finally, I would like to thank my friends and family for their endless
and enthusiastic encouragement throughout the entire project. I will always be grateful
for their support and patience as I truly immersed myself into the “world of IS.” Of
course, the carrel gifts and awesome visits always made me smile, too. I also owe a
special thank you to Matthew Germaine, for both the creation of the most epic carrel-fort
to have ever existed, and for providing me with my very own IS soundtrack. I can
honestly say I now know every song by heart.

-‐55-‐
LITERATURE CITED
Alves-‐Silva, R., Silva, H.R. 2009. Life in Bromeliads: Reproductive Behaviour and the Monophyly of the Scinax perpusillus Species Group (Anura: Hylidae). Journal of Natural History 43(3–4): 205–217.
Block, W.M., Brennan, L.A. 1993. The habitat concept in ornithology: theory and applications. Current ornithology. Vol. 11. Plenum Press, NY pp. 35-91.
Bradford, D.F., Seymour, R.S. 1988. Influence of Environmental PO₂ on Embryonic
Oxygen Consumption, Rate of Development, and Hatching in the Frog Pseudophryne bibroni. Physiological Zoology 61(6): 475–482.
Brown, J. H., Stevens, G.C., Kaufman, D.M. 1996. The Geographic Range: Size, Shape,
Boundaries, and Internal Structure. Annual Review of Ecology and Systematics 27: 597–623.
Brown, J.S. 1998. Game theory and habitat selection. In Game theory and animal
behavior, ed. L.A. Dugatkin and H.K. Reeve. Oxford University Press, Oxford, pp.188-220.
Caldwell, J.P. 1993. Brazil nut fruit capsules as phytotelmata: Interactions among anuran
and insect larvae. Canadian Journal of Zoology 71: 1193-1201. Caldwell, J.P., Araujo, M.C. 2004. Historical and Ecological Factors Influence
Survivorship in Two Clades of Phytotelm-Breeding Frogs (Anura: Bufonidae, Dendrobatidae). Misc. Publ. Mus. Zool., Univ. Mich. 193: 11–21.
Chase, J.M., Leibold, M.A. 2003. Ecological Niches: Linking Classical and
Contemporary Approaches. The University of Chicago Press Ltd., Chicago, 212 pp.
Cherrett, J.M. 1989. Ecological Concepts: The Contribution of Ecology to an
Understanding of the Natural World. Blackwell Scientific Publications, London. Chiu, C., Kam, Y. 2006. Testing the Nest-Homing Abilities of a Phytotelm-breeding
Frog, Chirixalus eiffingeri (Rhacophoridae). Zoological Science 23(6): 501–505. Cummins, C.P. 1986. Effects of aluminum and low pH on growth and development in
Rana temporaria tadpoles. Oecologia 69: 248-252. Denver, R.J. 1998. Hormonal Correlates of Environmentally Induced Metamorphosis in
the Western Spadefoot Toad, Scaphiopus hammondii. General and Comparative Endocrinology 110: 326–336.

-‐56-‐
Diaz, R.J., Rutger, R. 2008. Spreading Dead Zones and Consequences for Marine
Ecosystems. Science 321: 926-929. Downie, J. R., Livingstone, S. R., Cormack, J. R. 2001. Selection of tadpole deposition
sites by male Trinidadian stream frogs, Mannophryne trinitatis (Dendrobatidae): An example of anti-predator behaviour. Herpetological Journal 11(3): 91-100.
Duellman, W.E., Gray, P. 1983. Developmental Biology and Systematics of the Egg-
Brooding Hylid Frogs, Genera Flectonotus and Fritziana. Herpetologica 39(4): 333–359.
Duellman, W.E., Maness, S.J. 1980. The Reproductive Behavior of Some Hylid
Marsupial Frogs. Journal of Herpetology 14(3): 213–222. Elinson, R.P., Del Pino, E.M., Townsend, D.S., Cuesta, F.C., Eichhorn, P. 1990. A
Practical Guide to the Developmental Biology of Terrestrial-Breeding Frogs. Biol. Bull. 179: 163-177.
Elton, C. 1927. Animal ecology. London: Sidgwick and Jackson. Fretwell, S.D., Lucas, H.L. 1970. On territorial behavior and other factors influencing
habitat distribution in birds. Acta Biotheoretica 19: 16-36. Gosner, K.L. 1960. A simplified table for staging anuran embryos and larvae with notes
on identification. Herpetologica: 183-190. Grinnell, Joseph. 1917. The Niche-Relationships of the California Thrasher. The Auk
34(4): 427-433. Guimaraes-Souzai, B.A., Mendes, G.B., Bento, L., Marotta, H., Santoro, A.L., Esteves,
F.A., Pinho, L., Farjalla, V.F., Enrich-Prast, A. 2006. Limnological parameters in the water accumulated in tropical bromeliads. Acta Limnol. Bras. 18(1): 47-53.
Hanski, I., Singer, M.C. 2001. Extinction-colonization and host-plant choice in butterfly
metapopulations. American Naturalist 158: 341-353. Hardin, G. 1960. The Competitive Exclusion Principle. Science 131: 1292-1297. Heying, H. 2004. Reproductive Limitation by Oviposition site in a Treehole Breeding
Madagascan Poison Frog (Mantella laevigata). Misc. Publ. Mus. Zool., Univ. Mich. 193: 23-30.

-‐57-‐
Hutchinson, G. E. 1957. Concluding remarks. Cold Spring Harbor. Symp. Quant. Biol. 22: 415-427.
Hutto, R.L. 1985. Habitat Selection by Nonbreeding, Migratory Land Birds. Habitat
Selection in Birds. Academic Press, Inc. pp. 455-473. Holt, R.D. 1987. Population Dynamics and Evolutionary Processes: The Manifold Roles
of Habitat Selection. Evolutionary Ecology 1(4): 331–347. Jones, J. 2001. Habitat Selection Studies in Avian Ecology: A Critical Review. The Auk
118(2):557-562. Kam, Y., Chuang, Z., Yen, C. 1996. Reproduction, Oviposition-Site Selection, and
Tadpole Oophagy of an Arboreal Nester, Chirixalus eiffingeri (Rhacophoridae), from Taiwan. Journal of Herpetology 30(1): 52–59.
Kenny, J.S. 1969. Amphibia of Tinidad. Studies on the Fauna of Curacao and Other
Caribbean Islands 29(54):1-78. Krausman, P.R. 1999. Some Basic Principles of Habitat Use. Idaho Forest, Wildlife &
Range Exp. Sta. Bull. 70: 85-90. Krebs, J.R., Kacelnick, A., Taylor, P. 1978. Test of optimal sampling by foraging great
tits. Nature 275: 27-31. Laurila, A., Kujasalo, J. 1999. Habitat duration, predation risk and phenotypic plasticity
in common frog (Rana temporaria) tadpoles. Journal of Animal Ecology 68: 1123-1132.
Lehtinen, R.M., Carfagno, G.L.F. 2011. Habitat Selection, the Included Niche, and
Coexistence in Plant-Specialist Frogs from Madagascar. Biotropica 43(1): 58–67. Lehtinen, R.M., Lannoo, M.J., Wassersug, R.J. 2004. Phytotelm-Breeding Anurans:
Past, Present, and Future Research. Misc. Publ. Mus. Zool., Univ. Mich. 193: 1–9. Lehtinen, R.M. 2002. The Use of Screw Pines (Pandanus) by Amphibians and Reptiles
in Madagascar. Herpetological Bulletin 82: 20–25. Lehtinen, R.M. 2004. Tests for Competition, Cannibalism, and Priority Effects in Two
Phytotelm-dwelling Tadpoles from Madagascar. Herpetologica 60(1): 1–13. Limin, F., Li, Z. 2005. Habitat Selection by Asian Elephant (Elephas Maximus) in
Xishuangbanna, Yunnan, China. Acta Theriologica Sinica 25(3): 229 pp.

-‐58-‐
Lin, Y., Kam, Y. 2008. Nest Choice and Breeding Phenology of an Arboreal-Breeding Frog, Kurixalus eiffingeri (Rhacophoridae), in a Bamboo Forest. Zoological Studies 47(2): 129-137.
Macarthur, R., Levins, R. 1967. The Limiting Similarity, Convergence, and Divergence
of Coexisting Species. The American Naturalist 101(921): 377-385. Martin, T.E. 1992. Breeding productivity considerations: what are the appropriate
habitat features for management? J. M. Hagan and D. W. Johnston, ed. Ecology and conservation of Neotropical migrant birds. Smithsonian. Inst. Press, Washington, D.C. pp. 455-473.
Martin, T.E. 1998. Are Microhabitat Preferences of Coexisting Species Under Selection
and Adaptive? Ecology 79(2): 656–670. Murphy, J.C. 1997. Amphibians and Reptiles of Trinidad and Tobago. Krieger Publishing
Company, FL. Murphy, P.J. 2003. Does Reproductive Site Choice in A Neotropical Frog Mirror
Variable Risks Facing Offspring? Ecological Monographs 73(1): 45–67. Odling-Smee, F.J., Laland, K.N., Feldman, M.W. 2003. Niche Construction: The
Neglected Process in Evolution. Monographs in Population Biology, Princeton, NJ, 472 pp.
Paine, R.T. 1966. Food Web Complexity and Species Diversity. The American
Naturalist 100(910): 65-75. Paradise, C.J. 2000. Effects of pH and resources on a processing chain interaction in
simulated treeholes. Journal of Animal Ecology 69: 651-658. Paradise, C.J. 2004. Relationship of water and leaf litter variability to insects inhabiting
treeholes. J.N. AM. Benthol. Soc. 23(4): 793-805. Pulliam, H.R., Danielson, B.J. 1991. Sources, Sinks, and Habitat Selection: A
Landscape Perspective on Population Dynamics. The American Naturalist 137, Supplement: Habitat Selection, pp. S50-S66.
Rausher, M.D. 1984. Tradeoffs in performance on different hosts: evidence from
within- and between-site variation in the beetle Deloyala guttata. Evolution 38: 582-595.
Resetarits, W.J. 1996. Oviposition Site Choice and Life History Evolution. American
Zoologist 36(2): 205–215.

-‐59-‐
Robertson, D.R. 1972. Social control of sex reversal in a coral reef fish. Science 177:
1007-1009. Rödel, M.O., Rudolf, V.H.W., Frohschammer, S., Linsenmair, E. 2004. Life History of
a West African Tree-hole Breeding Frog, Phrynobatrachus guineensis, Guibe, and Lamotte, 1961 (Amphibia: Anura: Petropedetidae). Misc. Publ. Mus. Zool., Univ. Mich. 193: 31-44.
Rosenzweig, M.L. 1987. Habitat selection as a source of biological diversity.
Evolutionary Ecology 1: 315-330. Rosenzweig, M.L. 1991. Habitat Selection and Population Interactions: The Search for
Mechanism. The American Naturalist 137, Supplement: Habitat Selection, pp. S5-S28.
Rudolf, V.H.W., Rodel, M.O. 2007. Phenotypic plasticity and optimal timing of metamorphosis under uncertain time constraints. Evolutionary Ecology 21: 121–
142. Ryan, M.J., Barry, D.S. 2011. Competitive Interactions in Phytotelmata—Breeding
Pools of Two Poison-Dart Frogs (Anura: Dendrobatidae) in Costa Rica. Journal of Herpetology 45(4): 438-443.
Sandberger, L., Feldhaar, H., Lampert, K.P., Lamatsch, D.K., Rödel, M.O. 2010. Small,
Specialised and Highly Mobile? The Tree-hole Breeding Frog, Phrynobatrachus guineensis, Lacks Fine-scale Population Structure. African Journal of Herpetology 59(1): 79–94.
Schiesari, L., Gordo, M., Hödl, W., Lannoo, M.J. 2003. Treeholes as Calling, Breeding,
and Developmental Sites for the Amazonian Canopy Frog, Phrynohyas resinifictrix (Hylidae). Copeia: 263-272.
Silva, H.R., Carvalho, A.L.G., Bittencourt-‐Silva, G.B. 2011. Selecting a Hiding Place:
Anuran Diversity and the Use of Bromeliads in a Threatened Coastal Sand Dune Habitat in Brazil. Biotropica 43(2): 218–227.
Silva, H.R., Britto-‐Pereira, M. C. 2006. How Much Fruit Do Fruit-‐eating Frogs Eat? An
Investigation on the Diet of Xenohyla truncata (Lissamphibia: Anura: Hylidae). Journal of Zoology 270(4): 692–698.
Simmonds, N.W. 1950. Notes on the Biology of the Araceae of Trinidad. Journal of
Ecology 38(2): 277–291.

-‐60-‐
Southwood, T.R.E. 1988. Tactics, Strategies and Templets. Oikos 52(1): 3–18. Srivastava, D.S., Kolasa, J., Bengtsson, J., Gonzalez, A., Lawler, S.P., Miller, T.E.,
Munguia, P., Romanuk, T., Schneider, D.C., Trzcinski, M.K. 2004. Are Natural Microcosms Useful Model Systems for Ecology? Trends in Ecology & Evolution 19(7): 379–384.
Stephens, D.W., Krebs, J.R. 1986. Foraging Theory. Princeton, NJ: Princeton
University Press. Summers, K. 1992. Mating strategies in two species of dart-poison frogs: a comparative
study. Animal Behavior 43: 907-919. Summers, K., McKeon, C.S. 2004. The Evolutionary Ecology of Phytotelmata Use in
Neotropical Poison Frogs. Misc. Publ. Mus. Zool., Univ. Mich. 193: 55-73. Williams, D., MacKay, S.E., Verdonschot, R.C.M., Tacchino, P.J.P. 2007. Natural and
manipulated populations of the treehole mosquito, Ochlerotatus triseriatus, at its northernmost range limit in southern Ontario, Canada. Journal of Vector Ecology 32(2): 328-335.
Xavier, A., Napoli, M. 2011. Contribution of Environmental Variables to Anuran
Community Structure in the Caatinga Domain of Brazil. Phyllomedusa 10(1): 45–64.
Zotz, G., Thomas, V. 1999. How Much Water Is in the Tank? Model Calculations for
Two Epiphytic Bromeliads. Annals of Botany 83(2): 183–192.

-‐61-‐
APPENDIX
a) b) Figure A1. This study was conducted between two sites at both a tributary of the Bloody Bay River (a) and the Tobago Forest Reserve (b). Maps are replicated from simple sketches in field notes. The green ovals represent a sampled patch of X. jacquinii. The numbers labeling each patch correspond to the order in which the patches were sampled, and thus the label of the patch itself. The numbered red dots indicate plants found to be occupied that were not included in a patch. The light green dots indicate relatively high densities of X. jacquinii. Because the maps are not drawn to scale, they provide no more than a rough idea of where the patches are located in relation to each other and to other major landmarks.
= Numbered patch of X. jacquinii = Individual occupied X. jacquinii not included in a patch = Many X. jacquinii located here

-‐62-‐
Table A1: Summary of my field observations for each occupied X. jacquinii. The following notes provide additional insight into the frog’s behavior. Although the model analyzing the significance of the categorical variables was based on an average of all of the leaf axils, informal additional notes were taken describing the characteristics for each inhabited phytotelm. Plant ID Information
F. fitzgeraldi
Notes on frog behavior Extra notes about the individual occupied phytotelm or overall plant
BB3-30 7/30/12 3:20 pm Bloody Bay
2 tadpoles Both still upon first observation. The first was found when it was caught at the tip of the eyedropper being used to collect a water sample. The second was found at the base of the same phytotelm.
Phytotelm: Water depth: 2cm (50% filled), no detritus, no invertebrates, clean fresh-looking middle-aged leaf located 88 cm high (approximately halfway between newest and oldest leaves) Plant: Nothing noteworthy
BBfrog1 7/31/12 4:00 pm Bloody Bay
1 adult Jumped out in a projectile motion toward my face immediately as I pulled the leaf back for inspection. The soft, wet, squishy underside stuck to my hand (which reached up to block my face). Unfortunately, I instinctively shook it off before I realized it was a frog. After a thorough search of the forest floor, it could not be found.
Phytotelm: water depth: 2cm (25% filled), clean fresh-looking leaf axil supporting a full-grown leaf located slightly closer to the newer leaves, small spiders and one large ant crawling between the leaf axils Plant: Growing vegetatively (see Figure 6), located on steep slope

-‐63-‐
BBfrog2 7/31/12 4:30 pm Bloody Bay
1 adult Found wedged deep in the phytotelm, jumped out immediately as leaf was angled back to take a picture.
Phytotelm: Water depth: 3cm (50% filled), no detritus, no invertebrates, clean fresh-looking middle-aged leaf located approximately halfway between newest and oldest leaves Plant: Little fungal vine on outside of trunk
BBfrog3 7/31/12 4:45 pm Bloody Bay
1 adult female carrying 5 fairly developed eggs on her back
When I first saw her, she was sitting just above the water line. As I pulled back the leaf, she sought shelter in the water, wedging herself deeply at the base of the phytotelm. Upon further disturbance (angling leaf for better camera view), she jumped away.
Phytotelm: Water depth: 4cm (50% filled), no detritus although small amount of fine, light-colored sand at the bottom, no invertebrates, clean fresh-looking middle-aged leaf located approximately halfway between newest and oldest leaves Plant: Little fungal vine on outside of trunk, located on a steep slope
-‐64-‐
BBfrog4 8/1/12 4:40 pm Bloody Bay
1 adult Found sitting just above water level. Jumped away as I tried to take a picture.
Phytotelm: Water depth: .5cm (50% filled), no detritus although small amount of fine, light-colored sand at the bottom, no invertebrates, clean fresh-looking middle-aged leaf located slightly closer to the oldest leaves (17 cm from the ground) Plant: Plant growing out from under log on steep slope. Height of the plant was measured from the top of the log because of the extremely steep slope on the other side of the log.
BB5-41 8/2/12 9:30 am Bloody Bay
2 adults (1 male, 1 female)
One large male prominently placed just below the water level with head and forearms clearly visible. One female hiding beneath him wedged deeper in the phytotelm. Each climbed out of the phytotelm before leaping away while I took pictures. The contrast between the body shapes of the frogs when compared side by side allowed me to confidently label the sex of each individual (see Figure 3).
Phytotelm: Water depth: 3cm (50% filled), no detritus, no invertebrates, clean fresh-looking middle-aged leaf located closer to the newest leaves (18 cm from the ground) Plant: patch of white fungus on underside of leaf petiole

-‐65-‐
BB5-43 8/2/12 9:55 am Bloody Bay
1 adult male Male fully submerged in the water of the phytotelm with head and forearms visible.
Phytotelm: Water depth: 3cm (50% filled), no detritus, clean fresh-looking middle-aged leaf located halfway between newest and oldest leaves (at a height of 72 cm) Plant: One large ant crawling between nearby leaf axils
BB5-44 8/2/12 10:15 am Bloody Bay
3 adults (1 male, 1 female, 1 unknown)
Male sitting above water level on the petiole of the leaf. One female submerged below water level with head and forearms visible. One unsexed individual wedged deeply into the base of the phytotelm. This third frog jumped out before I could properly observe its body shape.
Phytotelm: Water depth: 4cm (75% filled), no detritus, one beetle floating on the surface of the water, clean fresh-looking middle-aged leaf located closer to the newest leaves (19 cm from the ground) Plant: Nothing noteworthy
FR5-47 8/3/12 4:30 pm Forest Reserve
1 adult Found submerged in the water with head and forearms clearly visible. Hopped out before it could be sexed.
Phytotelm: Water depth: 3cm (50% filled), no detritus, no invertebrates, clean fresh-looking middle-aged leaf located approximately halfway between newest and oldest leaves Plant: Nothing noteworthy

-‐66-‐
FRfrog1 8/3/12 5:25 pm Forest Reserve
1 adult female with several small but distinct lumps on her back
Seeking shelter in the phytotelm, hopped away before I could take a picture.
Phytotelm: Water depth: 4cm (75% filled), no detritus, no invertebrates, clean fresh-looking middle-aged leaf located approximately halfway between newest and oldest leaves Plant: Nothing noteworthy
FRfrog2 8/3/12 5:30 pm Forest Reserve
1 adult Submerged below water level with head and forearms clearly visible. Hopped out before it could be sexed.
Phytotelm: Water depth: 4cm (50% filled), no detritus, clean fresh-looking middle-aged leaf located approximately halfway between newest and oldest leaves Plant: Spider (>7mm) crawling between leaf axils

















