
Experiences from SGC Stockholm - Uppsala...
43
SGC Oxford SGC Toronto SGC Stockholm Experiences from SGC Stockholm Helena Berglund Structural Genomics Consortium Karolinska Institutet, Department of Medical Biochemistry and Biophysics Stockholm, Sweden
Transcript of Experiences from SGC Stockholm - Uppsala...

SGC OxfordSGC Toronto SGC Stockholm
Experiences from SGC Stockholm
Helena BerglundStructural Genomics Consortium
Karolinska Institutet, Department of Medical Biochemistry and Biophysics Stockholm, Sweden

Structural Genomics Consortium
• not-for-profit organisation
• proteins with relevance to human health and disease
• three sites: Toronto, Oxford, Stockholm
• in Stockholm since spring 2005, ~25 people
closing down spring 2011
• Stockholm: > 2 deposited structures / month
• 180 pdb depositions, ~150 different structures
SGC-
Oxford
~65 staff
SGC-
Toronto
~70 staff
SGC-
Stockholm
~25 staff
board of directors
scientific committee
CEO

strategy:
• focus on protein production
• work flow
• biological background, optimisation and trouble shooting, follow up
•cloning, small scale expression screen, large scale culture and purif.
• bioinformatics, x-ray, biophysics expertise and infrastructure
•ELN electronic laboratory notebook (ConturELN)
•LIMS database - BeeHive
• selection of target families Stockholm• suggestions from us, Swedish scientific community, local Swedish scientific
committee
• approval from the SGC scientific committee
management and core scientist
biotechnology
structural biology teams

protein production SGC Stockholm:
•Stockholm main target areas: nucleotide metabolism, domains in apoptosis and receptor
signalling, PARPs, phosphoinositide and lipid signalling, amino acid metabolism, ATPases
• E. coli
• N-terminal His tag
• multi construct approach
• parallel processing of all constructs as far as possible

multi construct approach
• educated guesses of domain boundaries, vary from there
• different protein variants differ in
• expression
• purification yield
• crystallisation properties
• evaluated inGräslund et al. ”The use of systematic N- and C-terminal deletions to promote production and structural studies of recombinant proteins” Protein Expr Purif. 2007 58, 210-221
Savitsky et al. ”High-throughput production of human proteins for crystallization : The SGC experience” J Struct
Biol. 2010 172, 3-13
targeted domain
Start Stop
AB
C
DE

evaluating the multi-construct approach:
a twice as many constructs could be
purified
0
10
20
30
40
50
fraction of targets with at least one construct:
producing highly soluble protein
0
4
8
12
16
20
resulting in well-diffracting crystals
all constructs
shortest &
full-length
all constructs
shortest &
full-length
a four-fold increase in well-diffracting crystals
all constructs (5008)
full-length and
shortest (925)

His tag:
MHHHHHHSSGVDLGTENLYFQ SM...
MGSSHHHHHHSSGLVPRG S....
• often tolerated in crystallization
• His tag removal - comparable with making new constructs
• SGC Stockholm :
~1/3 mounted crystals have the tag removed
-2008 -2011
deposited structures 77 143
tag removed 23 47
His tag making crystal contacts etc 3 ?
thrombin
TEV protease
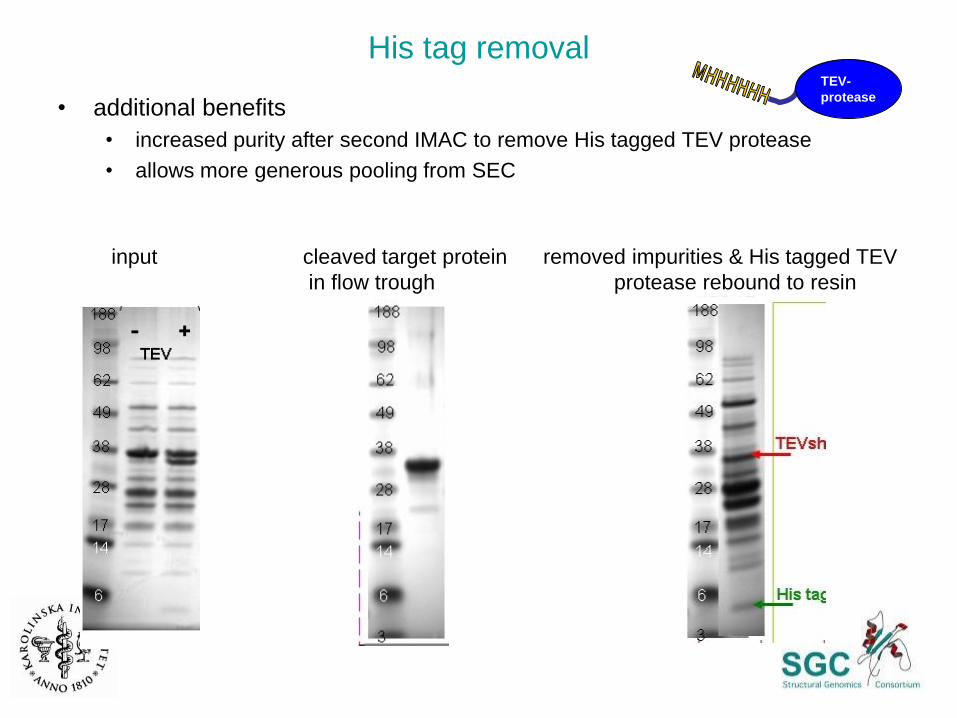
His tag removal
• additional benefits
• increased purity after second IMAC to remove His tagged TEV protease
• allows more generous pooling from SEC
input cleaved target protein removed impurities & His tagged TEV
in flow trough protease rebound to resin
TEV-
protease

moving the His-tag to the C-terminus
• comparable with making new constructs
• between second half of 2008 and 2011 : 16 of ~80 new deposited structures
• initial but not good enough crystals with the N-His construct for most of them, tested new domain borders for a few
....AHHHHHH
non-cleavable C-term His6:
MHHHHHHSSGVDLGTENLYFQ SM....
from cleavable N-term His6:
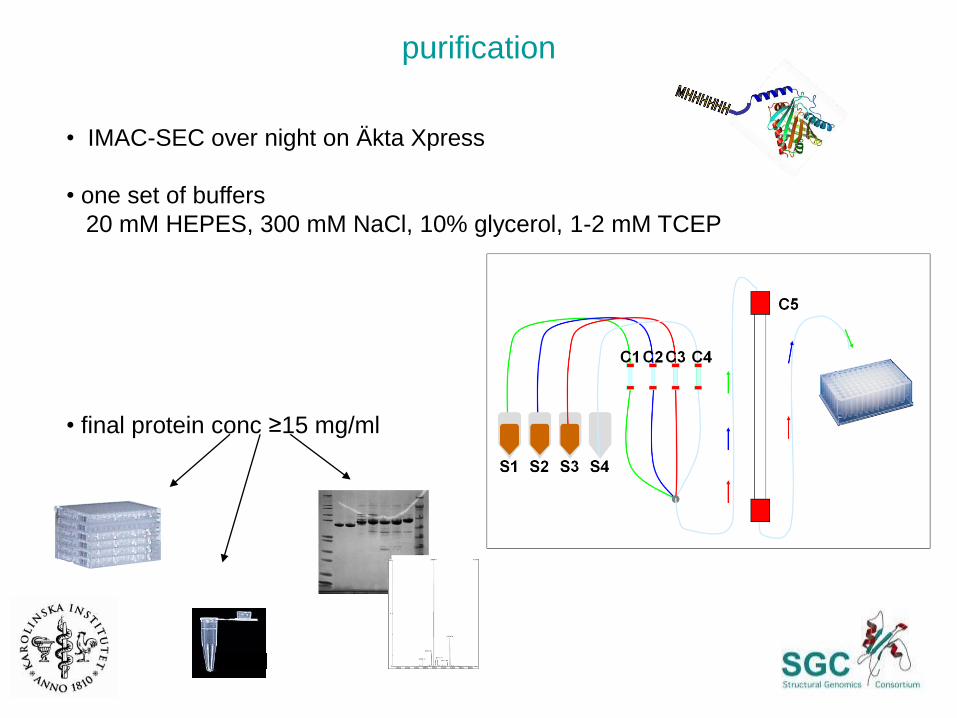
purification
• IMAC-SEC over night on Äkta Xpress
• one set of buffers
20 mM HEPES, 300 mM NaCl, 10% glycerol, 1-2 mM TCEP
• final protein conc ≥15 mg/ml

gel filtration elution profiles as a quality indicator
Gräslund et al. Protein Expr Purif. 2007 Volume 58, Issue 2, April 2008, Pages 210-221
but over truncation!
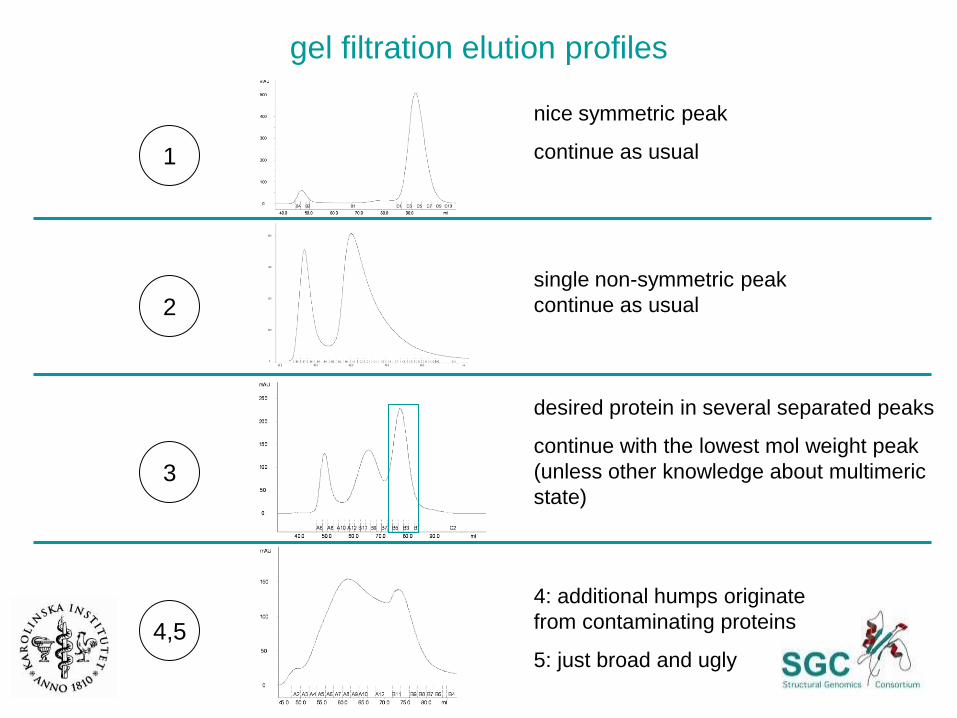
gel filtration elution profiles
2
3
4,5
1
nice symmetric peak
continue as usual
single non-symmetric peak
continue as usual
desired protein in several separated peaks
continue with the lowest mol weight peak
(unless other knowledge about multimeric
state)
4: additional humps originate
from contaminating proteins
5: just broad and ugly

characterization
• SDS-PAGE
• contaminating proteins
• size heterogeneities
• Mass spectrometry
• verify the identity
• chemical homogeneity
• performed at Biotechnology KTH Stockholm
Harry Brumer
21.5
66.3
36.531.0
55.4
14.4
6.0
97.4
FTHFDB GMPR2 ITPAA
ITPKCA PAP39A

mass spectrometrydetected features:
• usually correct mass!
• point mutations
• construct/sample mix-ups
• His tag modification
• α-N-gluconoylation: + 178 Da
Geoghegan et al, Analytical Biochemistry 267 169-184 1999
178 Da
-28 Da
-2 x28 Da
-3 x28 Da
• co-purifying proteins
• E. coli: SlyD, GroEL, Fur, YfbG
• arg-lys substitutions
• differential usage of codons in humans
and E. coli
• E. coli might incorporate a lysine (AAA)
if there is a lack of correct arg-tRNA.
HO-CH2-[CHOH]4-CO-NH-R

mixed projects SGC Stockholm:
0
100
200
300
400
60 70 80 90 100 110 120
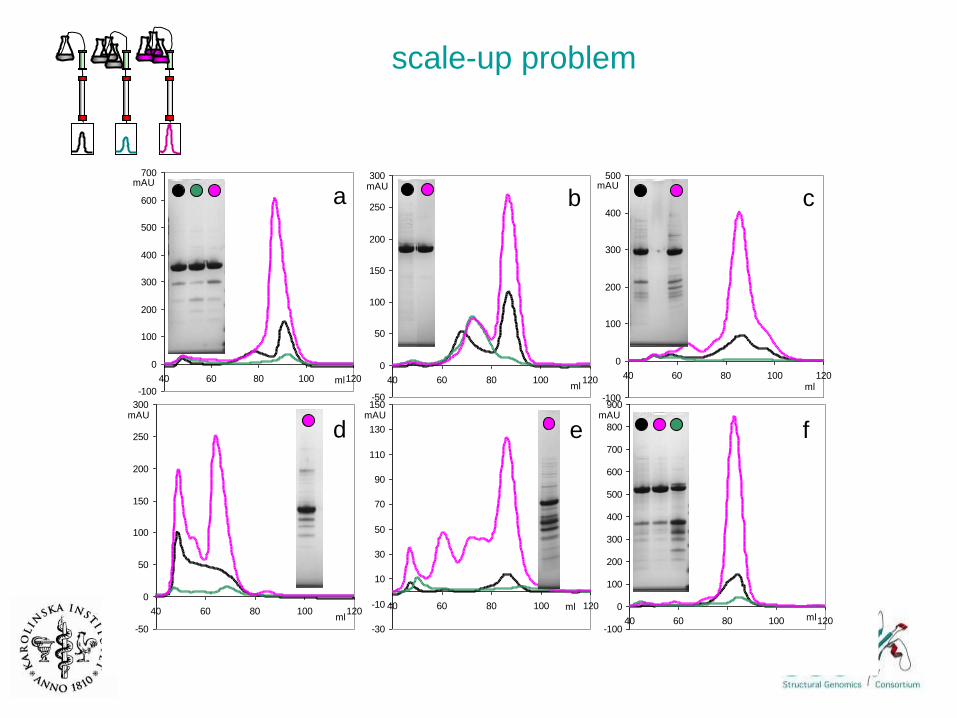
purification scale-up problem
0
100
200
300
400
60 70 80 90 100 110 120
I
M
A
C
S
E
C
ml
mAU
Audur Magnusdottir

model experiment
LYSATEBUFFER
His6-GFP pre-bound to
the top of the columns

scale-up problem
periplasm
removed
BUFFER LYSATE
• not due to :
reducing agent,
pH of lysate,
interferring DNA/RNA pieces,
HEPES buffer
competing E. coli proteins
.....
• work-arounds ?
• E coli lysate induced Ni-leakage
• metal chelators
scale-up problem
-30
-10
10
30
50
70
90
110
130
150
40 60 80 100 120ml
mAU
-100
0
100
200
300
400
500
600
700
40 60 80 100 120ml
mAU
-100
0
100
200
300
400
500
40 60 80 100 120ml
mAU
-50
0
50
100
150
200
250
300
40 60 80 100 120ml
mAU
-50
0
50
100
150
200
250
300
40 60 80 100 120ml
mAU
-100
0
100
200
300
400
500
600
700
800
900
40 60 80 100 120ml
mAU
a b c
d e f

chaperone co-production
Baneyx & Mujacic, Nat Biotech (2004)
co-expression of the target proteins in E.
coli over expressing chaperones to obtain
higher amounts of soluble protein.
TaKaRa plasmid set
pG-KJE8 dnaK+dnaJ+grpE+groEL+groES
pGro7 groES+groEL s
pKJE7 dnaK+dnaJ+grpE x
pG-Tf2 groEL+groES+tig s
pTf16 tig

small scale expression screening
4 3 2 1purified proteins
scoring

0
50
100
150
200
250
-2 -1 0 1 2 3
nu
mb
er
of co
nstr
ucts
chaperone co-expression
difference small scale screening score
score(chaperone strain) - score(standard strain)
>380 constructs tested with different chaperones
48 constructs , 12 different targets side-by-side comparison using pG-
KJE8: dnaK+dnaJ+grpE+groEL+groES
5 structures until now
Martina Nilsson, Martin Hammarström
small set of constructs compared in large scale
positive effect generally lost

E. coli strain arctic express
Arctic Express (DE3) (Stratagene)
express Cpn60/Cpn10 from O.antarctica.
cold-adapted chaperones ( opt at 4-12 ºC )
1wf4.pdb
Oleispira antarctica
74% and 54 % aa identity to the E.coli GroEL/ES,

arctic express
difference small scale screening score
score(arctic express (DE3)) - score(standard strain)
42 constructs , 9 different targets, 4C
no deposited structure yet but 3.1 Å structure for one tricky
protein

PreScission protease TEV protease
GST-LEVLFQG HMHHHHHH-SGVDLGT-ENLYFQ SM.....
fusion proteins:
PreScission protease
Thioredoxin-LEVLFQG HMHHHHHH-SGVDLGT-ENLYFQ SM.....
TEV proteaseMBP-ETVRFQ S HHHHHH-SSGVDLGT-ENLYFQ SM....
TEV protease
constitutively produced, in vivo cleavage
TVMV protease
MHHHHHH-SSG-SHPQFEK-GT-ENLYFQ SM....
TEV protease
STREP-tag
difference small scale screening score
score(standard plasmid) - score(MBP)
still, 1 structure
Martin Hammarström
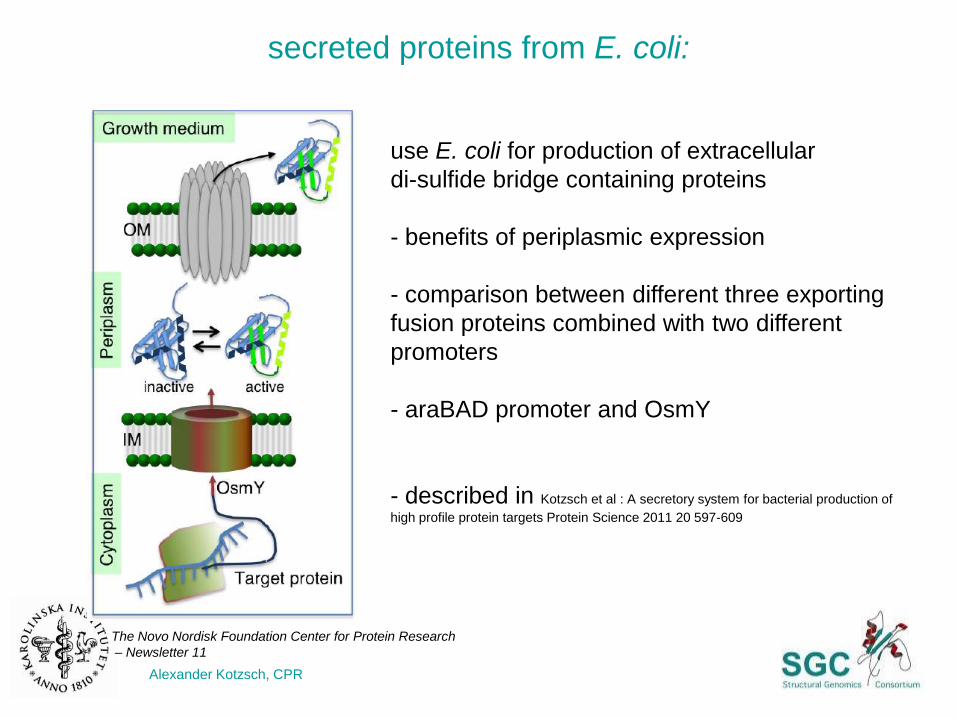
secreted proteins from E. coli:
The Novo Nordisk Foundation Center for Protein Research
– Newsletter 11
use E. coli for production of extracellular
di-sulfide bridge containing proteins
- benefits of periplasmic expression
- comparison between different three exporting
fusion proteins combined with two different
promoters
- araBAD promoter and OsmY
- described in Kotzsch et al : A secretory system for bacterial production of
high profile protein targets Protein Science 2011 20 597-609
Alexander Kotzsch, CPR

secreted proteins from E. coli
proteins in crystallisation
successful protein production
changing host:E. coli insect cells
produced, but
insoluble
all tricks tested
on protein family
members
Ida Johansson, Lionel Trésaugues
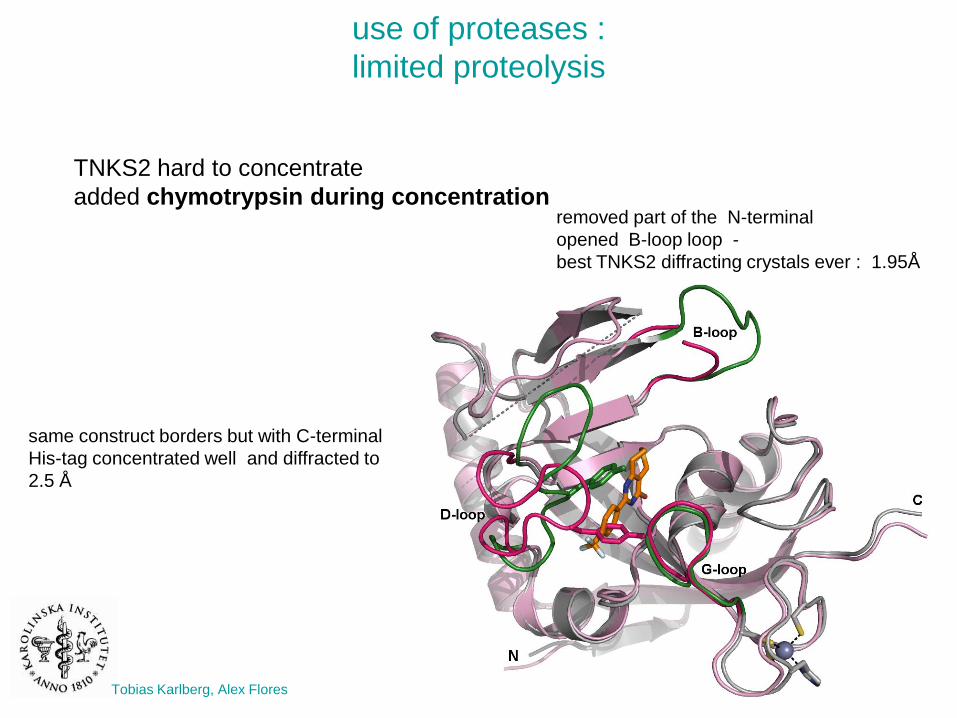
use of proteases :
limited proteolysis
TNKS2 hard to concentrate
added chymotrypsin during concentration
same construct borders but with C-terminal
His-tag concentrated well and diffracted to
2.5 Å
removed part of the N-terminal
opened B-loop loop -
best TNKS2 diffracting crystals ever : 1.95Å
Tobias Karlberg, Alex Flores

in situ proteolysis:
• addition of small amounts of protease ( 1: 200 - 1: 10000) in crystallisation
trypsin R or K, not before P
chymostrypsin F or Y or W , not before P
Glu-C V8 E or D
papain R or K
thermolysin L or F
endoproteinase Lys-C K = hydrophobic aa
Martin Welin
In situ proteolysis for protein crystallization and structure determinationNature Methods 2007 4: 1019 - 1021 work mainly from University of Toronto
In situ proteolysis to generate crystals for structure determination : an update PLoS one 2009 4:e5094
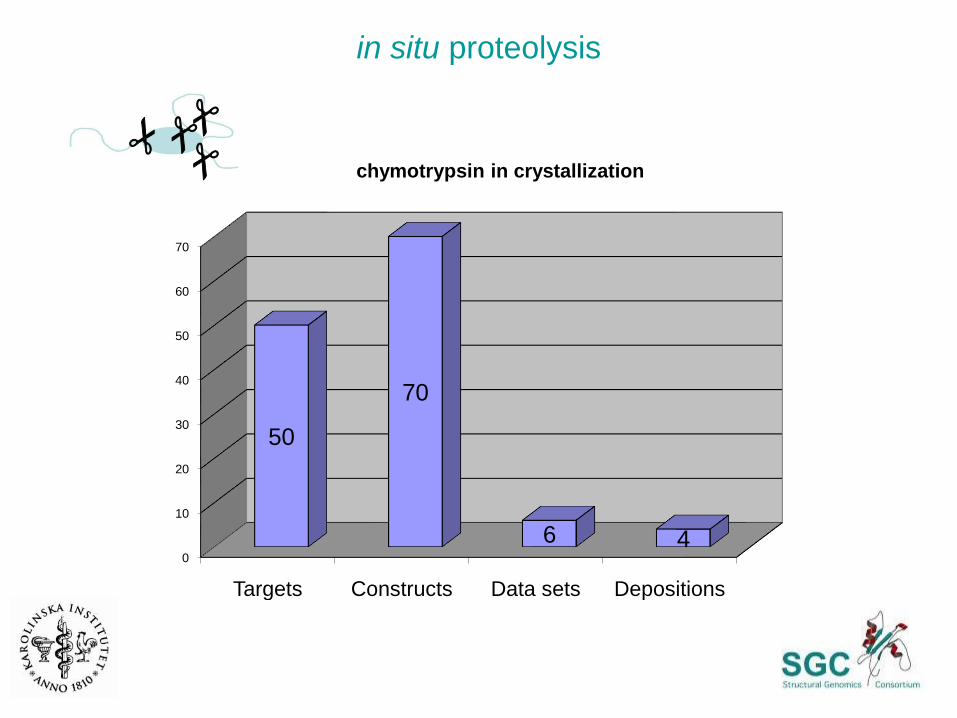
in situ proteolysis
0
10
20
30
40
50
60
70
Targets Constructs Data sets Depositions
50
70
6 4
chymotrypsin in crystallization

in situ proteolysis
SGC Stockholm
• tested on ~90 different targets
• usally one or two proteases
• usually not optimised protase ratio
• 5 structures
•trifunctional human enzyme encoded by the GART-gene
Linker (45) AIRS (475-792) Linker (15)GARS (1-430) GART 808-1010
solved by otherssolved at SGC
insoluble on its own

reductive methylation:
• di-methylation of all accessible lysine side
chains and the N-terminal amino group
• reduces surface disorder
Rayment Methods Enzymol. (1997). 276, 171
NH3+
-NH- -CO-CαH
CβH2
CγH2
CδH2
CεH2
NH
-NH- -CO-CαH
CβH2
CγH2
CδH2
CεH2
+CH3 CH3
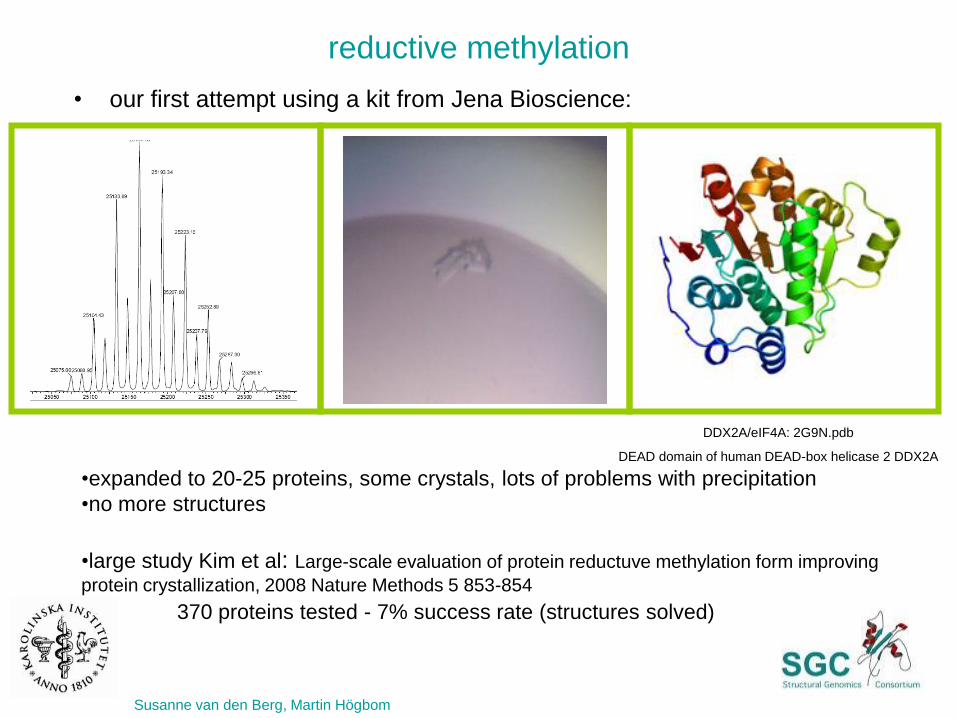
reductive methylation
•expanded to 20-25 proteins, some crystals, lots of problems with precipitation
•no more structures
•large study Kim et al: Large-scale evaluation of protein reductuve methylation form improving
protein crystallization, 2008 Nature Methods 5 853-854
370 proteins tested - 7% success rate (structures solved)
Susanne van den Berg, Martin Högbom
DDX2A/eIF4A: 2G9N.pdb
DEAD domain of human DEAD-box helicase 2 DDX2A
• our first attempt using a kit from Jena Bioscience:

protein stabilisation:
• stabilisation by buffer exchange
• buffer composition, pH, salt,
additives - non specific protein
stabilizing agents, ligands
• ligand induced stabilisation
Flu
ore
scence Inte
nsity →
Temperature →
: SyproOrange fluoresce in low dielectric media (=non-polar environments
such as the interior of an unfolding protein or molten globule)
λex =490 λem =575nm
Pantoliano et al, Journal of Biomolecular Screening,2001;429-440
thermal shift assay: thermofluor

10 0
50 55 60 65 70 75 80 85 90
ΔT
thermal shift assays
• Performed in a RT-PCR instrument:
• 96 samples in ~1.5 hours, different buffers, ligands etc
• ~5-10 µg of protein/well, ~500 µg /96 well plate
• measure fluorescence as a function of temperature in each well
• fit each curve using a sigmoidal function
• extract Tm, transition temperature
• ΔTm -comparison between different conditions
••
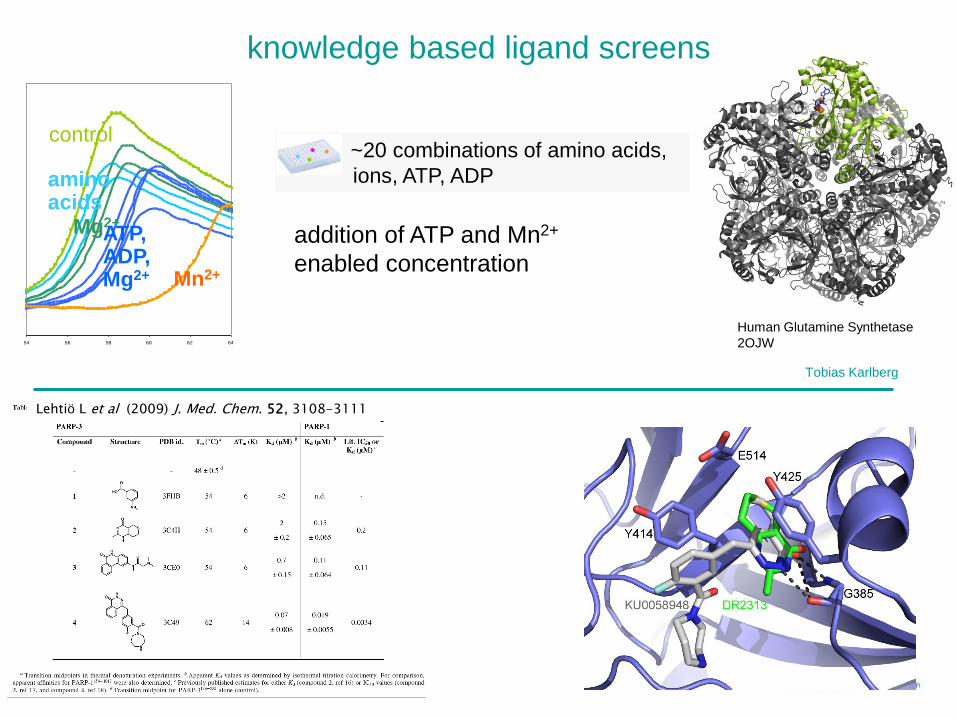
knowledge based ligand screens
54 56 58 60 62 64
control
amino acids
Mg2+ATP,ADP,Mg2+ Mn2+
addition of ATP and Mn2+
enabled concentration
~20 combinations of amino acids,
ions, ATP, ADP•
•••
Tobias Karlberg
Human Glutamine Synthetase
2OJW
Lehtiö L et al (2009) J. Med. Chem. 52, 3108-3111

thermofluor curves of structure determined targetsGMPR2
0
200
400
600
800
1000
1200
20 40 60 80
ITPKC
0
100
200
300
400
500
600
700
20 40 60 80
TULP1
0
500
1000
1500
2000
2500
20 40 60 80
FTHFDB
0
100
200
300
400
500
600
700
800
20 40 60 80
IMPA2
0
200
400
600
800
1000
1200
20 40 60 80
PRPSAP1
0
100
200
300
400
500
600
700
800
900
20 40 60 80
PPP2R4
0
100
200
300
400
500
600
700
20 40 60 80
NUDT3
0
100
200
300
400
500
600
700
800
900
20 40 60 80
DDX25
0
1000
2000
3000
4000
5000
6000
7000
20 40 60 80
DDX2A
0
100
200
300
400
500
600
700
20 40 60 80
PIP5K2C
0
500
1000
1500
2000
2500
20 40 60 80
ITPA
0
50
100
150
200
250
300
350
20 40 60 80
DRP2
0
200
400
600
800
1000
1200
1400
1600
20 40 60 80
CPSF5
0
200
400
600
800
1000
1200
20 40 60 80
FABP1
0
1000
2000
3000
4000
5000
6000
7000
20 40 60 80

alternatives
Guillermo Senisterra
Structural Genomics Consortium
University of Toronto
• only interpret good looking curves
• spin sample hard before thermofluor measurement
• alternative techniques:
Stargazer
for buffers: DLS
analytical SEC

buffer optimisation using DLS
• no crystals in standard buffer PARP13 catalytic domain at 1.1 Å 2x5y.pdb
Ann-Gerd Thorsell, Tobias Karlberg

buffer optimisation using DLS
DLS data at 0.2
mg/ml proteinstandard HEPES buffer pH 7.5 20 mM TRIS, 150 mM NaCl, pH 7.5
TNKS1Amonomers,
99% monodisp
monomers,
99.7% monodisp
PARP10Amonomers and dimers, some oligomers
and LA
monomers
98% monodisp
aggregation with time (1-2 h)
PARP12Aoligomers
98.5% monodisp
multimers
99.4% monodisp
PARP15A dimers and monomers dimer or monomer 97.5% monodisp
PARP13A dimers, tetramers and aggregates >98% monomers,
Natalia Markova

What will happen now?
• Toronto Oxford still running
• for all deposited:
pdb
M&M
reagents
• for many of them:
iSee data packs
• for some additional proteins:
protein production protocols
• publications
• visiting scientist program
what will happen now?
Stockholm site
biotechnology structural biology teamsx-ray

ACKNOWLEDGEMENTS
SGC Stockholm :Aida SehicAlex FloresAlexander KotzschAndreas JohanssonAnnette RoosAnn-Gerd ThorsellAudur MagnusdottirBiljana JovanovicBjörn ForsströmBrendan McManusCamilla PerssonDerek OggElisabeth WahlbergGlareh AskareihHanna WillanderHelena BerglundHerwig Schüler
Ida JohanssonJessica AnderssonJohan NilvebrandtJohan WeigeltJohanna SagemarkJonas UppenbergKarin WalldénKate KouznetsovaKenneth OlesenKerstin MichalkeLari LehtiöLasse DahlgrenLinda SvenssonLionel Trésaugues Lola HermanLouise EgebladLovisa Holmberg-SchiavoneMagdalena Wisniewska
Mailen AnderssonMalin UppstenMarina SiponenMartin HammarströmMartin HällbergMartin HögbomMartin MocheMartin Welin Martina NilssonMonika AbramczukNatalia MarkovaNatalie PerssonPatricia WennerstrandPatrick SchützPer KraulisPetra Nilsson-EhlePetri KursulaPetter NorbergPål Stenmark
Pär NordlundRobert BusamRuairi CollinsStephan KolStina LundgrenSusanne FlodinSusanne GräslundSusanne van den BergTanya KotenyovaTine Kragh-Nielsen Tobias KarlbergTomas Nyman Torun EkbladUlrika ErikssonUlrika IsbergUrzula KosinskaÅsa Kallas
SGC TorontoSGC Oxford
FUNDING PARTNERS
Canadian Institutes for Health Research, Canadian Foundation for Innovation, Genome Canada through the Ontario Genomics Institute, GlaxoSmithKline, Knut and Alice Wallenberg Foundation, Merck & Co., Inc., Novartis Research Foundation, Ontario Innovation Trust, Ontario Ministry for Research and Innovation, Swedish Agency for Innovation Systems, Swedish Foundation for Strategic Research, and Wellcome Trust. www.thesgc.org


















