
Exopolysaccharide Production byViridans Streptococci in ... · EXOPOLYSACCHARIDE PRODUCTION BY...
9
Vol. 43, No. 1 INFECTION AND IMMUNITY, Jan. 1984, p. 359-367 0019-9567/84/010359-09$02.00/0 Copyright © 1984, American Society for Microbiology Exopolysaccharide Production by Viridans Streptococci in Experimental Endocarditis JOHN MILLS,'* LYNN PULLIAM,1t LAWRENCE DALL,lt JOE MARZOUK,1 WALTER WILSON, AND J. WILLIAM COSTERTON3 The Medical Service, San Francisco General Hospital Medical Center and the University of California, San Francisco, California 941101; Section of Infectious Diseases, The Mayo Clinic and Research Foundation, Rochester, Minnesota 559052; and Department of Biology, University of Calgary, Calgary, Alberta, Canada T2N 1N43 Received 6 September 1983/Accepted 13 October 1983 Light and electron microscopy with histochemical staining were used to estimate exopolysaccharide production by strains of viridans streptococci recovered from patients with endocarditis. Six strains were selected for study because they represented a wide range of in vitro polysaccharide production. By light microscopy, there was good agreement between three polysaccharide stains (ruthenium red, periodic acid- Schiff and calcifluor white) in the amount of glycocalyx produced, which ranged from minimal (0 to 1+) to maximal amounts (4+). Two strains selected for minimal (strain 1) and maximal (strain 6) in vitro exopolysaccharide production were studied after we used them to experimentally infect cardiac vegeta- tions. Glycocalyx could be demonstrated surrounding organisms in cardiac vegetations, and the relative amounts produced were similar to those seen in vitro. Vegetations formed by glycocalyx-producing strains were also larger than those formed by glycocalyx-deficient strains. Viridans group streptococci which produce exopolysaccharide in vitro also do so within cardiac vegetations. The relationship of exopolysac- charide production to maintenance of endocardial infection is discussed. Examination of bacteria growing in a wide variety of natural (5), autochthonous (4), and pathogenic (6, 7) environ- ments has revealed that the majority of these cells are found in microcolonies adherent to surfaces. After initial adhesion of free-floating bacteria to these surfaces, the bacteria repli- cate within an enveloping matrix composed of their own exopolysaccharide (glycocalyx) (6, 7). These microcolonies have been shown to attain almost macroscopic dimensions in chronic staphylococcal infections centered on cardiac pace- makers (22) and in chronic Pseudomonas pulmonary infec- tions in cystic fibrosis (16) and much smaller dimensions in acute infections such as intestinal infections caused by enterotoxigenic Escherichia coli (3). The exopolysaccha- rides of the bacterial glycocalyx are chemically varied, but most consist of highly hydrated anionic polymers (30) simi- lar, both chemically and physically, to the majority of commercial ion exchange resins. In addition to its obvious role in adherence, this extracellular matrix has been shown to protect bacteria from opsonization (1) and phagocytosis (29), and it also confers protection from surfactants (12) and certain antibiotics (13). Bacterial endocarditis is of particular interest in this context because the causative agents depend on adhesion to initiate colonization (25, 26, 28). Viridans group streptococci are the most common cause of bacterial endocarditis, and in their natural habitat, the mouth, they are known to produce an exopolysaccharide (glycocalyx) which is predominantly dextran. In experimental endocarditis, production of a su- crose-derived dextran exopolysaccharide has been correlat- ed both to adherence in vitro (25) and infectivity in vivo (8, 9, 25, 26, 28). However, it is not known whether viridans streptococci produce glycocalyx within valvular vegetations * Corresponding author. t Present address: Department of Laboratory Medicine, Veterans Administration Hospital, San Francisco, CA 94121. t Present address: Department of Medicine, University of Mis- souri-Kansas City, Kansas City, MO 64108. 359 (in which no sucrose substrate is available), or whether such production might play a role in sustaining preexisting intra- vascular infection. To answer these questions, we examined a number of strains of viridans group streptococci recovered from patients with endocarditis for their ability to produce an exopolysaccharide glycocalyx in vitro and in cardiac vegeta- tions. (A portion of this material was presented at the 22nd Intersci. Conf. Antimicrob. Agents Chemother., Miami Beach, Fla. abstr. no. 358, 1982.) MATERIALS AND METHODS Test strains. Viridans group streptococci recovered from patients with endocarditis who were hospitalized at San Francisco General Hospital and at the Mayo Clinic Affiliated Hospitals were identified as to species as described by Pulliam et al. (24). Working stock cultures were suspended in Todd-Hewitt broth (Difco Laboratories, Detroit, Mich.) with 15% glycerol and stored frozen at -70°C. For in vitro estimation of glycocalyx production, the strains were grown in Todd-Hewitt broth with added 5% glucose (this medium does not contain sucrose). Animal model of endocarditis. Endocarditis was induced in rabbits according to the procedure originally described by Garrison and Freedman (11) and modified by Durack and Beeson (8). Streptococcal isolates were grown in Todd- Hewitt broth for 7 h to produce an inoculum in the exponen- tial phase of growth. About 8 h after catheterization, 1.0 ml of medium containing the test organism was injected into a marginal ear vein. The inoculum was titrated after injection, and the actual amount given varied between 1 x 108 and 2 x 109 CFU. At intervals of 2 to 48 h after infection, the animals were killed by CO2 inhalation, and the cardiac vegetations were removed and immediately prepared for microscopic examination as described below. Histological techniques. For light microscopy, vegetations were fixed in Zenkers and embedded in Paraplast embedding resin, and 4-,um sections were cut and stained with Mayer's on April 16, 2020 by guest http://iai.asm.org/ Downloaded from
Transcript of Exopolysaccharide Production byViridans Streptococci in ... · EXOPOLYSACCHARIDE PRODUCTION BY...

Vol. 43, No. 1INFECTION AND IMMUNITY, Jan. 1984, p. 359-3670019-9567/84/010359-09$02.00/0Copyright © 1984, American Society for Microbiology
Exopolysaccharide Production by Viridans Streptococci inExperimental Endocarditis
JOHN MILLS,'* LYNN PULLIAM,1t LAWRENCE DALL,lt JOE MARZOUK,1 WALTER WILSON, AND
J. WILLIAM COSTERTON3The Medical Service, San Francisco General Hospital Medical Center and the University of California, San Francisco,California 941101; Section of Infectious Diseases, The Mayo Clinic and Research Foundation, Rochester, Minnesota
559052; and Department of Biology, University of Calgary, Calgary, Alberta, Canada T2N 1N43
Received 6 September 1983/Accepted 13 October 1983
Light and electron microscopy with histochemical staining were used to estimate exopolysaccharideproduction by strains of viridans streptococci recovered from patients with endocarditis. Six strains were
selected for study because they represented a wide range of in vitro polysaccharide production. By lightmicroscopy, there was good agreement between three polysaccharide stains (ruthenium red, periodic acid-Schiff and calcifluor white) in the amount of glycocalyx produced, which ranged from minimal (0 to 1+) tomaximal amounts (4+). Two strains selected for minimal (strain 1) and maximal (strain 6) in vitroexopolysaccharide production were studied after we used them to experimentally infect cardiac vegeta-tions. Glycocalyx could be demonstrated surrounding organisms in cardiac vegetations, and the relativeamounts produced were similar to those seen in vitro. Vegetations formed by glycocalyx-producing strainswere also larger than those formed by glycocalyx-deficient strains. Viridans group streptococci whichproduce exopolysaccharide in vitro also do so within cardiac vegetations. The relationship of exopolysac-charide production to maintenance of endocardial infection is discussed.
Examination of bacteria growing in a wide variety ofnatural (5), autochthonous (4), and pathogenic (6, 7) environ-ments has revealed that the majority of these cells are foundin microcolonies adherent to surfaces. After initial adhesionof free-floating bacteria to these surfaces, the bacteria repli-cate within an enveloping matrix composed of their ownexopolysaccharide (glycocalyx) (6, 7). These microcolonieshave been shown to attain almost macroscopic dimensions inchronic staphylococcal infections centered on cardiac pace-makers (22) and in chronic Pseudomonas pulmonary infec-tions in cystic fibrosis (16) and much smaller dimensions inacute infections such as intestinal infections caused byenterotoxigenic Escherichia coli (3). The exopolysaccha-rides of the bacterial glycocalyx are chemically varied, butmost consist of highly hydrated anionic polymers (30) simi-lar, both chemically and physically, to the majority ofcommercial ion exchange resins. In addition to its obviousrole in adherence, this extracellular matrix has been shownto protect bacteria from opsonization (1) and phagocytosis(29), and it also confers protection from surfactants (12) andcertain antibiotics (13).
Bacterial endocarditis is of particular interest in thiscontext because the causative agents depend on adhesion toinitiate colonization (25, 26, 28). Viridans group streptococciare the most common cause of bacterial endocarditis, and intheir natural habitat, the mouth, they are known to producean exopolysaccharide (glycocalyx) which is predominantlydextran. In experimental endocarditis, production of a su-crose-derived dextran exopolysaccharide has been correlat-ed both to adherence in vitro (25) and infectivity in vivo (8, 9,25, 26, 28). However, it is not known whether viridansstreptococci produce glycocalyx within valvular vegetations
* Corresponding author.t Present address: Department of Laboratory Medicine, Veterans
Administration Hospital, San Francisco, CA 94121.t Present address: Department of Medicine, University of Mis-
souri-Kansas City, Kansas City, MO 64108.
359
(in which no sucrose substrate is available), or whether suchproduction might play a role in sustaining preexisting intra-vascular infection. To answer these questions, we examineda number of strains of viridans group streptococci recoveredfrom patients with endocarditis for their ability to produce anexopolysaccharide glycocalyx in vitro and in cardiac vegeta-tions.(A portion of this material was presented at the 22nd
Intersci. Conf. Antimicrob. Agents Chemother., MiamiBeach, Fla. abstr. no. 358, 1982.)
MATERIALS AND METHODSTest strains. Viridans group streptococci recovered from
patients with endocarditis who were hospitalized at SanFrancisco General Hospital and at the Mayo Clinic AffiliatedHospitals were identified as to species as described byPulliam et al. (24). Working stock cultures were suspendedin Todd-Hewitt broth (Difco Laboratories, Detroit, Mich.)with 15% glycerol and stored frozen at -70°C. For in vitroestimation of glycocalyx production, the strains were grownin Todd-Hewitt broth with added 5% glucose (this mediumdoes not contain sucrose).Animal model of endocarditis. Endocarditis was induced in
rabbits according to the procedure originally described byGarrison and Freedman (11) and modified by Durack andBeeson (8). Streptococcal isolates were grown in Todd-Hewitt broth for 7 h to produce an inoculum in the exponen-tial phase of growth. About 8 h after catheterization, 1.0 mlof medium containing the test organism was injected into amarginal ear vein. The inoculum was titrated after injection,and the actual amount given varied between 1 x 108 and 2 x109 CFU. At intervals of 2 to 48 h after infection, the animalswere killed by CO2 inhalation, and the cardiac vegetationswere removed and immediately prepared for microscopicexamination as described below.
Histological techniques. For light microscopy, vegetationswere fixed in Zenkers and embedded in Paraplast embeddingresin, and 4-,um sections were cut and stained with Mayer's
on April 16, 2020 by guest
http://iai.asm.org/
Dow
nloaded from
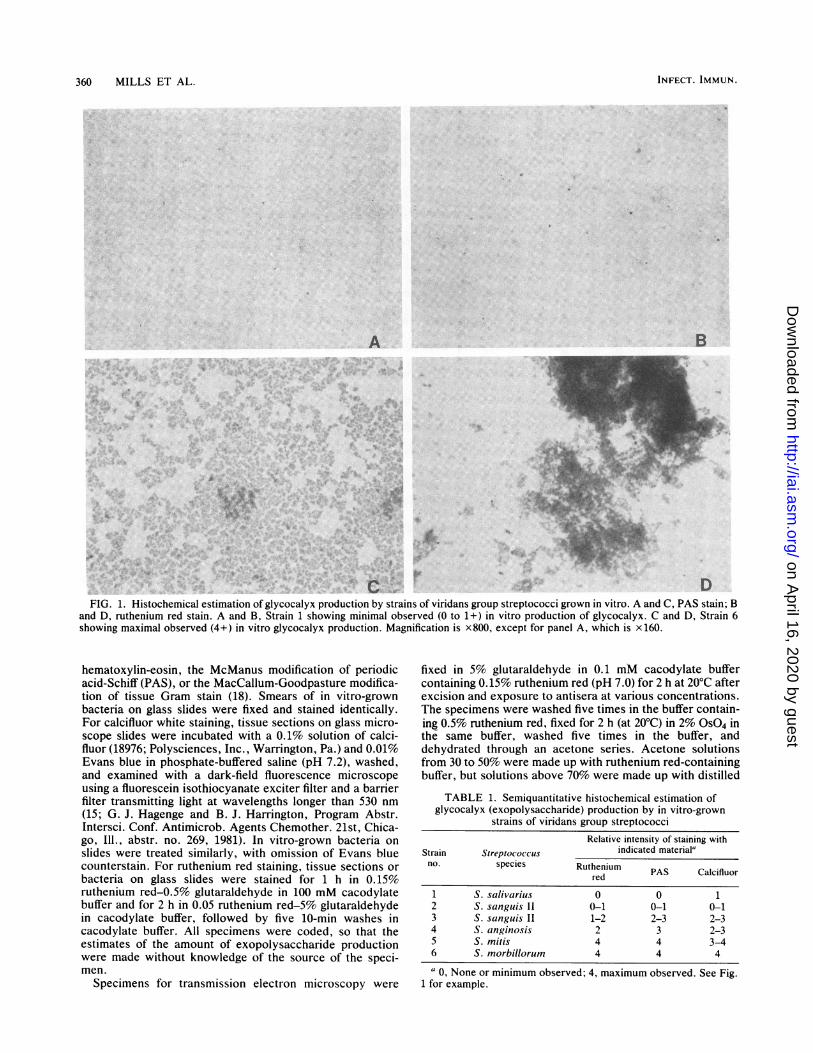
360 MILLS ET AL.
BA
t.
C. D
FIG. 1. Histochemical estimation of glycocalyx production by strains of viridans group streptococci grown in vitro. A and C, PAS stain; Band D, ruthenium red stain. A and B, Strain 1 showing minimal observed (0 to 1+) in vitro production of glycocalyx. C and D, Strain 6showing maximal observed (4+) in vitro glycocalyx production. Magnification is x800, except for panel A, which is x 160.
hematoxylin-eosin, the McManus modification of periodicacid-Schiff (PAS), or the MacCallum-Goodpasture modifica-tion of tissue Gram stain (18). Smears of in vitro-grownbacteria on glass slides were fixed and stained identically.For calcifluor white staining, tissue sections on glass micro-scope slides were incubated with a 0.1% solution of calci-fluor (18976; Polysciences, Inc., Warrington, Pa.) and 0.01%Evans blue in phosphate-buffered saline (pH 7.2), washed,and examined with a dark-field fluorescence microscopeusing a fluorescein isothiocyanate exciter filter and a barrierfilter transmitting light at wavelengths longer than 530 nm(15; G. J. Hagenge and B. J. Harrington, Program Abstr.Intersci. Conf. Antimicrob. Agents Chemother. 21st, Chica-go, Ill., abstr. no. 269, 1981). In vitro-grown bacteria onslides were treated similarly, with omission of Evans bluecounterstain. For ruthenium red staining, tissue sections orbacteria on glass slides were stained for 1 h in 0.15%ruthenium red-0.5% glutaraldehyde in 100 mM cacodylatebuffer and for 2 h in 0.05 ruthenium red-5% glutaraldehydein cacodylate buffer, followed by five 10-min washes incacodylate buffer. All specimens were coded, so that theestimates of the amount of exopolysaccharide productionwere made without knowledge of the source of the speci-men.
Specimens for transmission electron microscopy were
fixed in 5% glutaraldehyde in 0.1 mM cacodylate buffercontaining 0.15% ruthenium red (pH 7.0) for 2 h at 20°C afterexcision and exposure to antisera at various concentrations.The specimens were washed five times in the buffer contain-ing 0.5% ruthenium red, fixed for 2 h (at 20°C) in 2% OS04 inthe same buffer, washed five times in the buffer, anddehydrated through an acetone series. Acetone solutionsfrom 30 to 50% were made up with ruthenium red-containingbuffer, but solutions above 70% were made up with distilled
TABLE 1. Semiquantitative histochemical estimation ofglycocalyx (exopolysaccharide) production by in vitro-grown
strains of viridans group streptococciRelative intensity of staining with
Strain Streptoc oc c us indicated material'no. species Ruthenium PAS Calcifluor
red
1 S. salivarius 0 0 12 S. sanguis 11 0-1 0-1 0-13 S. sanguis II 1-2 2-3 2-34 S. anginosis 2 3 2-35 S. mitis 4 4 3-46 S. morbillorum 4 4 4a 0, None or minimum observed; 4, maximum observed. See Fig.
1 for example.
INFECT. IMMUN.
x1-..
on April 16, 2020 by guest
http://iai.asm.org/
Dow
nloaded from
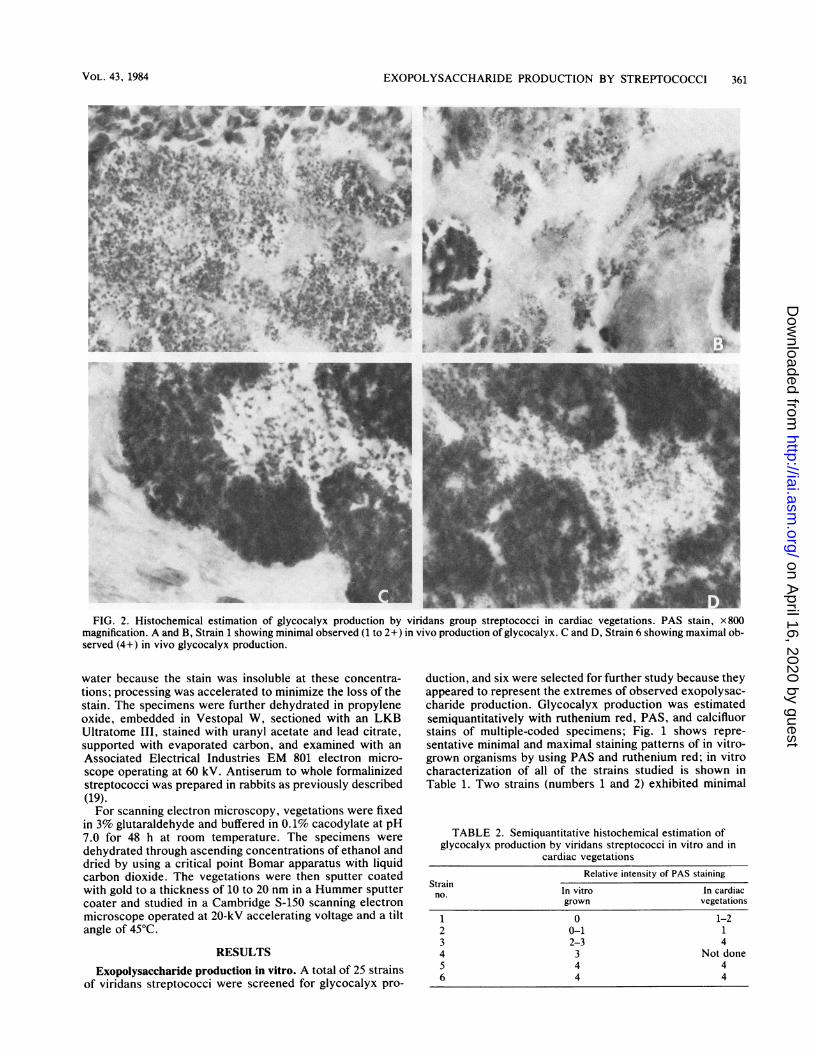
EXOPOLYSACCHARIDE PRODUCTION BY STREPTOCOCCI 361
FIG. 2. Histochemical estimation of glycocalyx production by viridans group streptococci in cardiac vegetations. PAS stain, x 800magnification. A and B, Strain 1 showing minimal observed (1 to 2+) in vivo production of glycocalyx. C and D, Strain 6 showing maximal ob-served (4+) in vivo glycocalyx production.
water because the stain was insoluble at these concentra-tions; processing was accelerated to minimize the loss of thestain. The specimens were further dehydrated in propyleneoxide, embedded in Vestopal W, sectioned with an LKBUltratome III, stained with uranyl acetate and lead citrate,supported with evaporated carbon, and examined with an
Associated Electrical Industries EM 801 electron micro-scope operating at 60 kV. Antiserum to whole formalinizedstreptococci was prepared in rabbits as previously described(19).For scanning electron microscopy, vegetations were fixed
in 3% glutaraldehyde and buffered in 0.1% cacodylate at pH7.0 for 48 h at room temperature. The specimens were
dehydrated through ascending concentrations of ethanol anddried by using a critical point Bomar apparatus with liquidcarbon dioxide. The vegetations were then sputter coatedwith gold to a thickness of 10 to 20 nm in a Hummer sputtercoater and studied in a Cambridge S-150 scanning electronmicroscope operated at 20-kV accelerating voltage and a tiltangle of 45°C.
RESULTS
Exopolysaccharide production in vitro. A total of 25 strainsof viridans streptococci were screened for glycocalyx pro-
duction, and six were selected for further study because theyappeared to represent the extremes of observed exopolysac-charide production. Glycocalyx production was estimatedsemiquantitatively with ruthenium red, PAS, and calcifluorstains of multiple-coded specimens; Fig. 1 shows repre-sentative minimal and maximal staining patterns of in vitro-grown organisms by using PAS and ruthenium red; in vitrocharacterization of all of the strains studied is shown inTable 1. Two strains (numbers 1 and 2) exhibited minimal
TABLE 2. Semiquantitative histochemical estimation ofglycocalyx production by viridans streptococci in vitro and in
cardiac vegetationsRelative intensity of PAS staining
Strainno. In vitro In cardiac
grown vegetations1 0 1-22 0-1 13 2-3 44 3 Not done5 4 46 4 4
VOL. 43, 1984
on April 16, 2020 by guest
http://iai.asm.org/
Dow
nloaded from
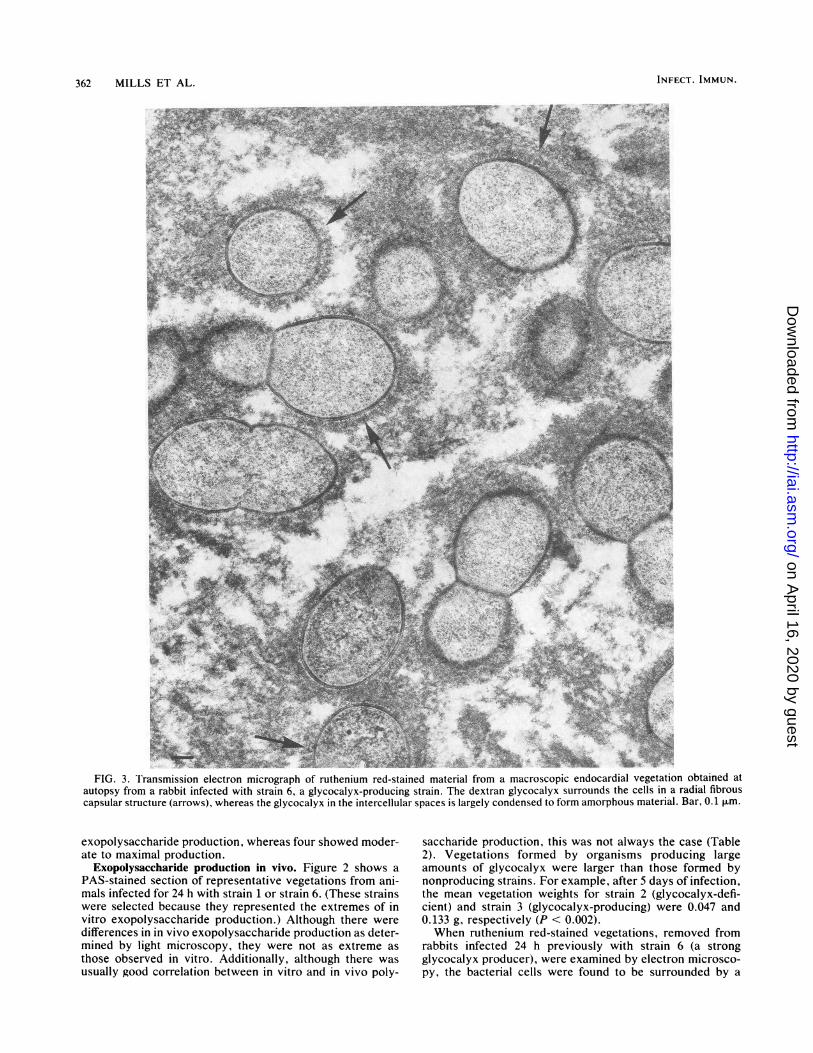
362 MILLS ET AL.
I
.4:.' .FIG. 3. I'ransmission electron micrograph of ruthenium red-stained material from a macroscopic endocardial vegetation obtained at
autopsy from a rabbit infected with strain 6, a glycocalyx-producing strain. The dextran glycocalyx surrounds the cells in a radial fibrouscapsular structure (arrows), whereas the glycocalyx in the intercellular spaces is largely condensed to form amorphous material. Bar, 0.1 ,um.
exopolysaccharide production, whereas four showed moder-ate to maximal production.
Exopolysaccharide production in vivo. Figure 2 shows aPAS-stained section of representative vegetations from ani-mals infected for 24 h with strain 1 or strain 6. (These strainswere selected because they represented the extremes of invitro exopolysaccharide production.) Although there weredifferences in in vivo exopolysaccharide production as deter-mined by light microscopy, they were not as extreme asthose observed in vitro. Additionally, although there wasusually good correlation between in vitro and in vivo poly-
saccharide production, this was not always the case (Table2). Vegetations formed by organisms producing largeamounts of glycocalyx were larger than those formed bynonproducing strains. For example, after 5 days of infection,the mean vegetation weights for strain 2 (glycocalyx-defi-cient) and strain 3 (glycocalyx-producing) were 0.047 and0.133 g, respectively (P < 0.002).When ruthenium red-stained vegetations, removed from
rabbits infected 24 h previously with strain 6 (a strongglycocalyx producer), were examined by electron microsco-py, the bacterial cells were found to be surrounded by a
INFECT. IMMUN.
on April 16, 2020 by guest
http://iai.asm.org/
Dow
nloaded from
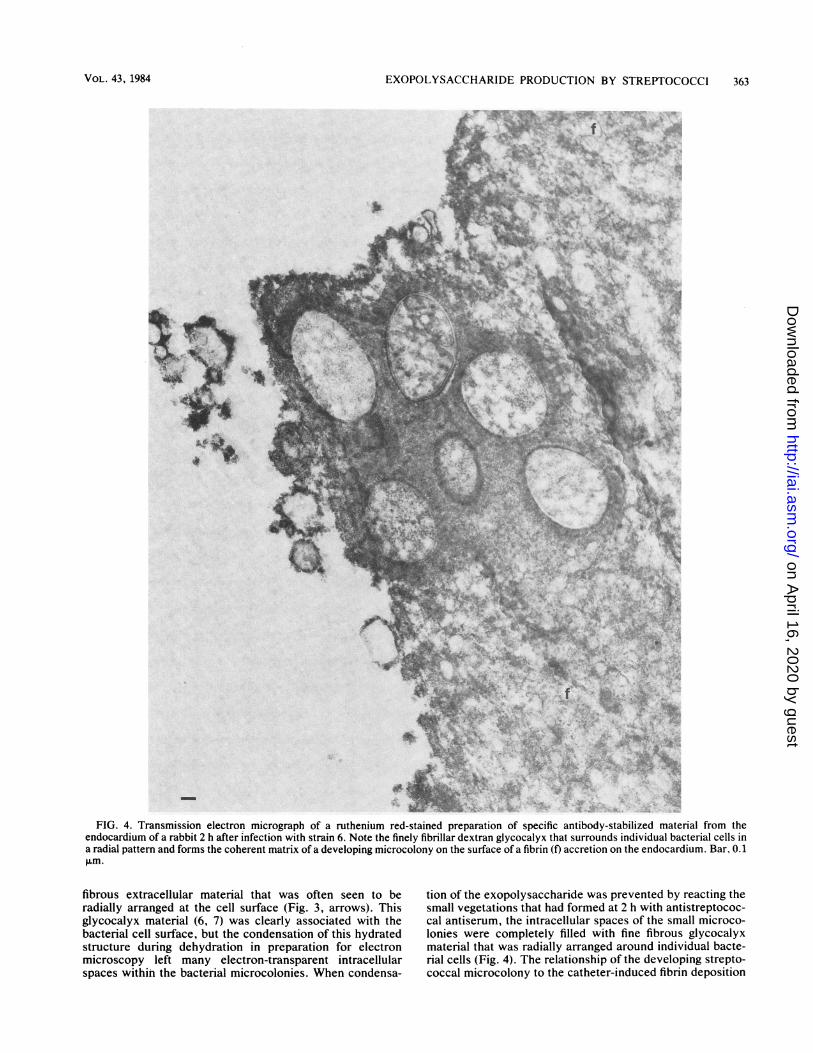
EXOPOLYSACCHARIDE PRODUCTION BY STREPTOCOCCI 363
.4
..
t e:!w
I
'5
>4 ?it.
.4, *, 5.!.
4 t. -.
,>. Ia: ...t
i~~~~~~'
S S ,,. .^114.%. - ,
FIG. 4. Transmission electron micrograph of a ruthenium red-stained preparation of specific antibody-stabilized material from theendocardium of a rabbit 2 h after infection with strain 6. Note the finely fibrillar dextran glycocalyx that surrounds individual bacterial cells ina radial pattern and forms the coherent matrix of a developing microcolony on the surface of a fibrin (f) accretion on the endocardium. Bar, 0.1p.fm.
fibrous extracellular material that was often seen to beradially arranged at the cell surface (Fig. 3, arrows). Thisglycocalyx material (6, 7) was clearly associated with thebacterial cell surface, but the condensation of this hydratedstructure during dehydration in preparation for electronmicroscopy left many electron-transparent intracellularspaces within the bacterial microcolonies. When condensa-
tion of the exopolysaccharide was prevented by reacting thesmall vegetations that had formed at 2 h with antistreptococ-cal antiserum, the intracellular spaces of the small microco-lonies were completely filled with fine fibrous glycocalyxmaterial that was radially arranged around individual bacte-rial cells (Fig. 4). The relationship of the developing strepto-coccal microcolony to the catheter-induced fibrin deposition
VOL. 43, 1984
A ..
.11
on April 16, 2020 by guest
http://iai.asm.org/
Dow
nloaded from
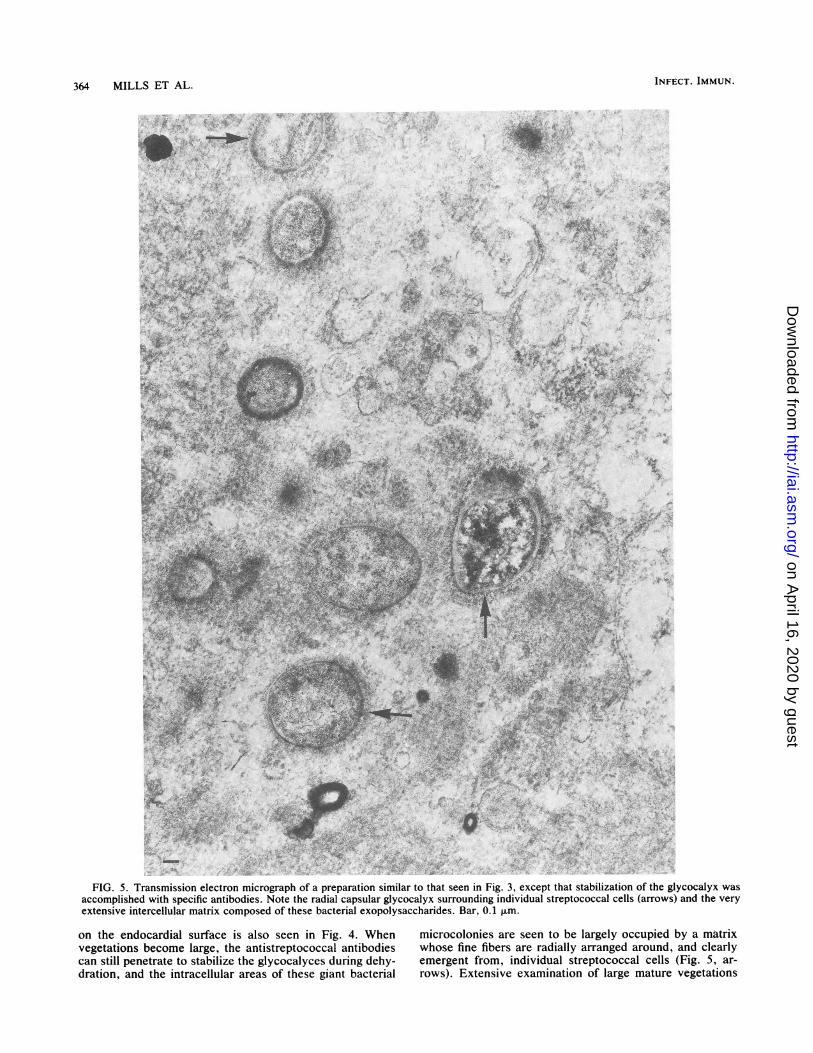
364 MILLS ET AL. INFECT. IMMUN.
....
r
: ~~~~~~~~~~~~~~~~~~~~~~~~~~~V
N~~~~~~~~~~~~~
V;~~~~~
AA
P.-~~~~~~~~~~~~~~
..0:.05
-W : per X i .. s.
-_4:v('./$<,<<g6,i ^@ z fiv ;1 s v'''
/~~~~~~~~~~~~~~~
7~~it,aW / .&siwK; 0 s ; %N-;.56
FIG.5J..Transmission electron micrograph of a preparation similar to that seen in Fig 3 except that stabilization of therg-ycocalyxwa
extensiveineclua mari copoe of ths batra exoplysachrie. Bar, 0.1 p.m.
ontheendocArdialsurface is also seen in Fi'g. 4. When microcolonies are seen to be largely occupied by a matri
*.r .*A ..
FIG. 5. Transmission electron micrograph of a preparation similar to that seen in Fig. 3, except that stabilization of the glycocalyx wasaccomplished with specific antibodies. Note the radial capsular glycocalyx surrounding individual streptococcal cells (arrows) and the veryextensive in'tercellular matrix composed of these bacterial exopolysaccharides. Bar, 0.1 Jim.
on the endocardial surface is also seen in Fig. 4. When microcolonies are seen to be largely occupied by a matrixvegetations become large, the antistreptococcal antibodies whose fine fibers are radially arranged around, and clearlycan still penetrate to stabilize the glycocalyces during dehy- emergent from, individual streptococcal cells (Fig. 5, ar-dration, and the intracellular areas of these giant bacterial rows). Extensive examination of large mature vegetations
on April 16, 2020 by guest
http://iai.asm.org/
Dow
nloaded from
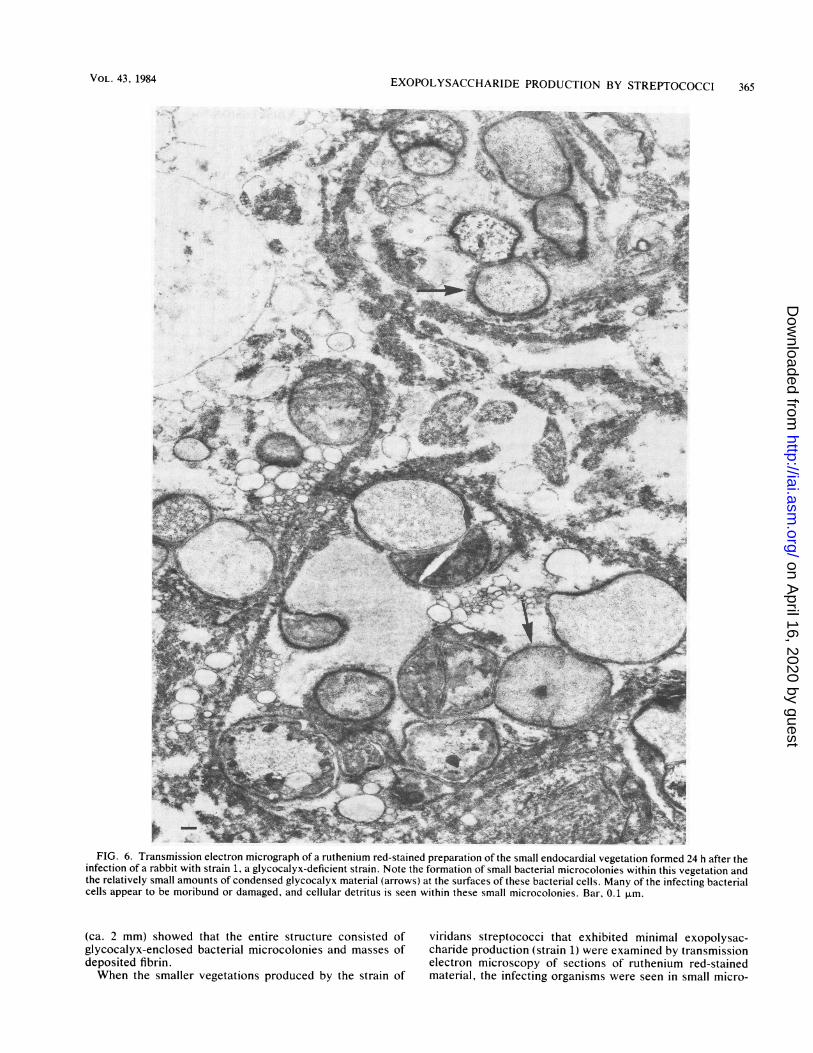
EXOPOLYSACCHARIDE PRODUCTION BY STREPTOCOCCI 365
FIG. 6. Transmission electron micrograph of a ruthenium red-stained preparation of the small endocardial vegetation formed 24 h after theinfection of a rabbit with strain 1, a glycocalyx-deficient strain. Note the formation of small bacterial microcolonies within this vegetation andthe relatively small amounts of condensed glycocalyx material (arrows) at the surfaces of these bacterial cells. Many of the infecting bacterialcells appear to be moribund or damaged, and cellular detritus is seen within these small microcolonies. Bar, 0.1 ,.m.
(ca. 2 mm) showed that the entire structure consisted of viridans streptococci that exhibited minimal exopolysac-glycocalyx-enclosed bacterial microcolonies and masses of charide production (strain 1) were examined by transmissiondeposited fibrin. electron microscopy of sections of ruthenium red-stainedWhen the smaller vegetations produced by the strain of material, the infecting organisms were seen in small micro-
VOL. 43, 1984
on April 16, 2020 by guest
http://iai.asm.org/
Dow
nloaded from
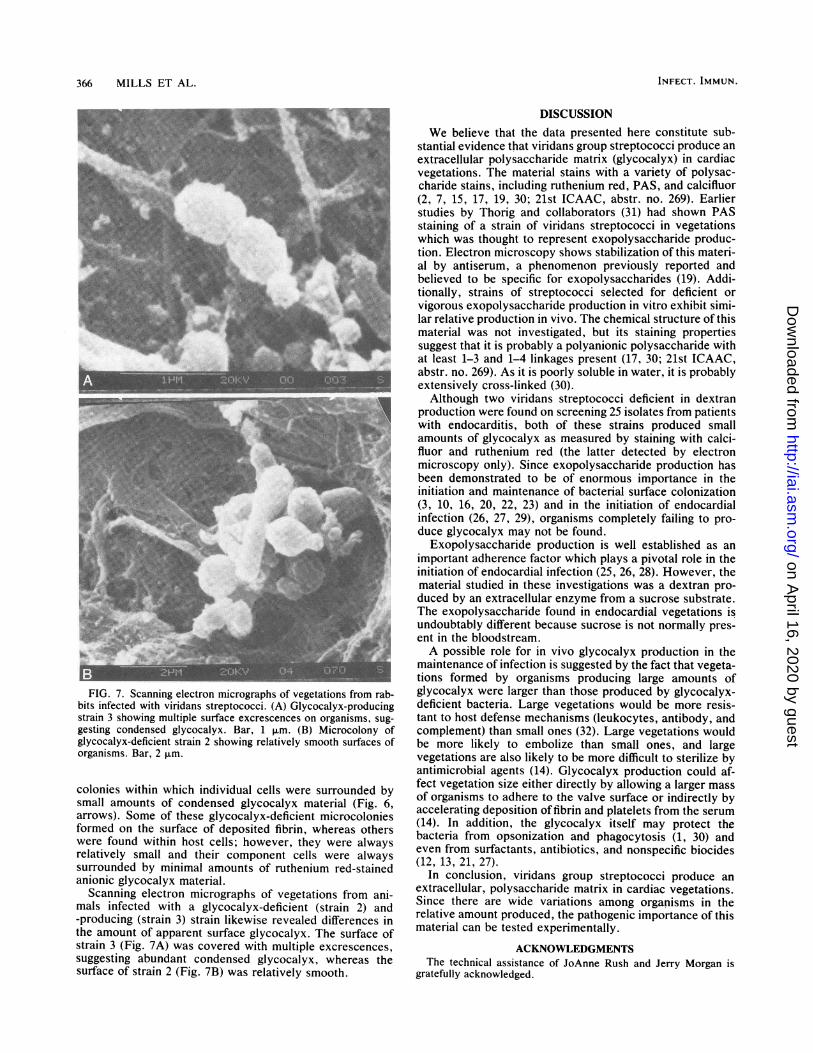
366 MILLS ET AL.
FIG. 7. Scanning electron micrographs of vegetations from rab-bits infected with viridans streptococci. (A) Glycocalyx-producingstrain 3 showing multiple surface excrescences on organisms, sug-gesting condensed glycocalyx. Bar, 1 ,Rm. (B) Microcolony ofglycocalyx-deficient strain 2 showing relatively smooth surfaces oforganisms. Bar, 2 ,.m.
colonies within which individual cells were surrounded bysmall amounts of condensed glycocalyx material (Fig. 6,arrows). Some of these glycocalyx-deficient microcoloniesformed on the surface of deposited fibrin, whereas otherswere found within host cells; however, they were alwaysrelatively small and their component cells were alwayssurrounded by minimal amounts of ruthenium red-stainedanionic glycocalyx material.
Scanning electron micrographs of vegetations from ani-mals infected with a glycocalyx-deficient (strain 2) and-producing (strain 3) strain likewise revealed differences inthe amount of apparent surface glycocalyx. The surface ofstrain 3 (Fig. 7A) was covered with multiple excrescences,suggesting abundant condensed glycocalyx, whereas thesurface of strain 2 (Fig. 7B) was relatively smooth.
DISCUSSIONWe believe that the data presented here constitute sub-
stantial evidence that viridans group streptococci produce an
extracellular polysaccharide matrix (glycocalyx) in cardiacvegetations. The material stains with a variety of polysac-charide stains, including ruthenium red, PAS, and calcifluor(2, 7, 15, 17, 19, 30; 21st ICAAC, abstr. no. 269). Earlierstudies by Thorig and collaborators (31) had shown PASstaining of a strain of viridans streptococci in vegetationswhich was thought to represent exopolysaccharide produc-tion. Electron microscopy shows stabilization of this materi-al by antiserum, a phenomenon previously reported andbelieved to be specific for exopolysaccharides (19). Addi-tionally, strains of streptococci selected for deficient orvigorous exopolysaccharide production in vitro exhibit simi-lar relative production in vivo. The chemical structure of thismaterial was not investigated, but its staining propertiessuggest that it is probably a polyanionic polysaccharide withat least 1-3 and 1-4 linkages present (17, 30; 21st ICAAC,abstr. no. 269). As it is poorly soluble in water, it is probablyextensively cross-linked (30).
Although two viridans streptococci deficient in dextranproduction were found on screening 25 isolates from patientswith endocarditis, both of these strains produced smallamounts of glycocalyx as measured by staining with calci-fluor and ruthenium red (the latter detected by electronmicroscopy only). Since exopolysaccharide production hasbeen demonstrated to be of enormous importance in theinitiation and maintenance of bacterial surface colonization(3, 10, 16, 20, 22, 23) and in the initiation of endocardialinfection (26, 27, 29), organisms completely failing to pro-duce glycocalyx may not be found.
Exopolysaccharide production is well established as animportant adherence factor which plays a pivotal role in theinitiation of endocardial infection (25, 26, 28). However, thematerial studied in these investigations was a dextran pro-duced by an extracellular enzyme from a sucrose substrate.The exopolysaccharide found in endocardial vegetations isundoubtably different because sucrose is not normally pres-ent in the bloodstream.A possible role for in vivo glycocalyx production in the
maintenance of infection is suggested by the fact that vegeta-tions formed by organisms producing large amounts ofglycocalyx were larger than those produced by glycocalyx-deficient bacteria. Large vegetations would be more resis-tant to host defense mechanisms (leukocytes, antibody, andcomplement) than small ones (32). Large vegetations wouldbe more likely to embolize than small ones, and largevegetations are also likely to be more difficult to sterilize byantimicrobial agents (14). Glycocalyx production could af-fect vegetation size either directly by allowing a larger massof organisms to adhere to the valve surface or indirectly byaccelerating deposition of fibrin and platelets from the serum(14). In addition, the glycocalyx itself may protect thebacteria from opsonization and phagocytosis (1, 30) andeven from surfactants, antibiotics, and nonspecific biocides(12, 13, 21, 27).
In conclusion, viridans group streptococci produce anextracellular, polysaccharide matrix in cardiac vegetations.Since there are wide variations among organisms in therelative amount produced, the pathogenic importance of thismaterial can be tested experimentally.
ACKNOWLEDGMENTSThe technical assistance of JoAnne Rush and Jerry Morgan is
gratefully acknowledged.
INFECT. IMMUN.
on April 16, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

EXOPOLYSACCHARIDE PRODUCTION BY STREPTOCOCCI
LITERATURE CITED
1. Baltimore, R. S., and M. Mitchell. 1980. Immunologic investiga-tions of mucoid strains of Pseudomnonas aeruiginosa: compari-son of susceptibility by opsonic antibody in mucoid and nonmu-
coid strains. J. Infect. Dis. 141:238-247.2. Caputy, G. G., and J. W. Costerton. 1982. Morphological
examination of the glycocalyces of Staphylococcus auireu.sstrains Wiley and Smith. Infect. Immun. 36:759-767.
3. Chan, R., S. D. Acres, and J. W. Costerton. 1982. Use of specificantibody to demonstrate glycocalyx, K99 pili, and the spatialrelationship of K99+ enterotoxigenic Esclherichia coli in theileum of colostrum-fed calves. Infect. Immun. 37:1170-1180.
4. Cheng, K. J., R. T. Irvin, and J. W. Costerton. 1981. Autochtho-nous and pathogenic colonization of animal tissues by bacteria.Can. J. Microbiol. 27:461-490.
5. Costerton, J. W., G. G. Geesey, and K. J. Cheng. 1978. Howbacteria stick. Sci. Am. 238:86-95.
6. Costerton, J. W., R. T. Irvin, and K. J. Cheng. 1981. The role ofbacterial cell surface components in pathogenesis. Crit. Rev.Microbiol. 8:303-338.
7. Costerton, J. W., R. T. Irvin, and K. J. Cheng. 1981. Thebacterial glycocalyx in nature and disease. Annu. Rev. Micro-biol. 35:299-324.
8. Durack, D. T., and P. B. Beeson. 1972. Experimental bacterialendocarditis. I. Colonization of a sterile vegetation. Br. J. Exp.Pathol. 53:44-49.
9. Durack, D. T., and P. B. Beeson. 1972. Experimental bacterialendocarditis. II. Survival of bacteria in endocarditis vegeta-tions. Br. J. Exp. Pathol. 53:50-53.
10. Fletcher, M., and G. D. Floodgate. 1973. Electron microscopicdemonstration of an acidic polysaccharide involved in theadhesion of a marine bacterium to solid surfaces. J. Gen.Microbiol. 72:325-334.
11. Garrison, P. K., and L. R. Freedman. 1970. Experiments inendocarditis. Staphylococcal endocarditis in rabbits resultingfrom placement of a polyethylene catheter in the right side of theheart. Yale J. Biol. Med. 42:394-410.
12. Govan, J. R. W. 1975. Mucoid strains of Pseidomnonas aeruigin-osa: the influence of culture medium on the stability of mucousproduction. J. Med. Microbiol. 8:513-522.
13. Govan, J. R. W., and J. A. M. Fyfe. 1978. Mucoid Pseiudomonasaerluginosta and cystic fibrosis: resistance of the mucoid form tocarbenicillin, flucloxacillin, and tobramycin and the isolation ofmucoid variations in vitro. J. Antimicrob. Chemother. 4:233-240.
14. Hook, E. W., III, and M. A. Sande. 1974. Role of the vegetationin experimental Streptococcus viridans endocarditis. Infect.Immun. 10:1433-1438.
15. Hughes, J., and M. E. McCully. 1975. The use of an opticalbrightener in the study of plant structure. Stain Technol.50:319-329.
16. Lam, J., R. Chan, K. Lam, and J. W. Costerton. 1980. Produc-tion of mucoid microcolonies by Pseidoinonas aerluginosa
within infected lungs in cystic fibrosis. Infect. Immun. 28:546-556.
17. Luft, J. H. 1971. Ruthenium red and ruthenium violet. I.Chemistry, purification, methods of use for electron microsco-py, and mechanism of action. Anat. Rec. 171:347-368.
18. Luna, L. G. 1968. Manual of histologic staining methods of theArmed Forces Institute of Pathology. 3rd ed. Blakiston-McGraw Hill, New York.
19. Mackie, E. B., K. N. Brown, J. Lam, and J. W. Costerton. 1979.Morphological stabilization of capsules of group B streptococci,types Ia, lb, II, and III with specific antibody. J. Bacteriol.138:609-617.
20. Marrie, T. J., and J. W. Costerton. 1980. Bacterial adhesion touroepithelial cells: a morphological study. J. Infect. Dis.142:239-246.
21. Marrie, T. J., and J. W. Costerton. 1981. Prolonged survival ofSerratia mnarcescens in chlorhexidine. AppI. Environ. Micro-biol. 42:1093-1102.
22. Marrie, T. J., J. Nelliga, and J. W. Costerton. 1982. A scanningand transmission electron microscopic study of an infectedendocardial pacemaker lead. Circulation 66:1339-1341.
23. McCoy, W. F., J. D. Bryers, J. Robbins, and J. W. Costerton.1981. Observations of fouling biofilm formation. Can. J. Micro-biol. 27:910-917.
24. Pulliam, L., R. K. Porschen, and W. K. Hadley. 1980. Biochemi-cal properties of CO,-dependent streptococci. J. Clin. Micro-biol. 12:27-31.
25. Ramirez-Ronda, C. H. 1978. Adherence of glucan-positive andglucan-negative streptococcal strains to normal and damagedheart valves. J. Clin. Invest. 62:805-814.
26. Ramirez-Ronda, C. H. 1980. Effects of molecular weight ofdextran on the adherence of Streptococcus sanguis to damagedheart valves. Infect. Immun. 29:1-7.
27. Ruseska, I., J. Robbins, E. S. Lashen, and J. W. Costerton. 1982.Biocide testing against corrosion-causing oilfield bacteria helpscontrol plugging. Oil Gas J., p. 253-264.
28. Scheld, W. M., J. A. Vaone, and M. A. Sande. 1978. Bacterialadherence in the pathogenesis of endocarditis. Interaction ofbacterial dextran platelets, and fibrin. J. Clin. Invest. 61:1394-1404.
29. Schwarzmann, S., and J. R. Boring. 1971. Antiphagocytic effectof slime from a mucoid strain of Pseuidomonas aeruginosa.Infect. Immun. 3:762-767.
30. Sutherland, I. W. 1977. Bacterial polysaccharides: their natureand production, p. 27-29. In 1. W. Sutherland (ed.), Surfacecarbohydrates of the prokaryotic cell. Academic Press, Inc.,New York.
31. Thorig, L., J. Thompson, F. Eulderink, J. J. Emeis, and R. VanFurth. 1979. Effects of monocytopenia and anticoaggulation inexperimental Streptococcus sangiis endocarditis. Br. J. Exp.Pathol. 61:108-113.
32. Yersin, B. R., M. P. Glauser, and L. R. Freedman. 1982. Effectof nitrogen mustard on natural history of right-sided streptococ-cal endocarditis in rabbits: role for cellular host defenses.Infect. Immun. 35:320-325.
VOL. 43, 1984 367
on April 16, 2020 by guest
http://iai.asm.org/
Dow
nloaded from


















