Fossils D. Crowley, 2008. Fossils To know how fossils are formed.

Evolving Form and Function:Fossils and DevelopmentProceedings of a symposium honoring Adolf Seilacher for his contributions to paleontology,in celebration of his 80th birthday
Derek E. G. Briggs, Editor
April 1– 2, 2005New Haven, Connecticut
A Special Publication of thePeabody Museum of Natural HistoryYale UniversityNew Haven, Connecticut, U.S.A.
December 2005

Evolving Form and Function: Fossils and DevelopmentProceedings of a symposium honoring Adolf Seilacher
for his contributions to paleontology, in celebration of his 80th birthdayA Special Publication of the Peabody Museum of Natural History, Yale University
Derek E.G. Briggs, Editor
These papers are the proceedings of Evolving Form and Function: Fossils and Development,a symposium held on April 1–2, 2005, at Yale University.
Yale Peabody Museum PublicationsJacques Gauthier, Curatorial Editor-in-Chief
Lawrence F. Gall, Executive Editor
Rosemary Volpe, Publications Editor
Joyce Gherlone, Publications Assistant
Design by Rosemary Volpe • Index by Aardvark Indexing
Cover: Fossil specimen of Scyphocrinites sp., Upper Silurian, Morocco (YPM 202267).Purchased for the Yale Peabody Museum by Dr. Seilacher. Photograph by Jerry Domian.
© 2005 Peabody Museum of Natural History, Yale University. All rights reserved.
Frontispiece: Photograph of Dr. Adolf Seilacher by Wolfgang Gerber.Used with permission. All rights reserved.
In addition to occasional Special Publications, the Yale Peabody Museum publishes the Bulletin of thePeabody Museum of Natural History, Postilla and the Yale University Publications in Anthropology. A com-plete list of titles, along with submission guidelines for contributors, can be obtained from the Yale PeabodyMuseum website or requested from the Publications Office at the address below.
Address correspondence to:
Publications OfficePeabody Museum of Natural HistoryYale University Phone: (203) 432-3786170 Whitney Avenue Fax: (203) 432-5872P.O. Box 208118 E-mail: [email protected] Haven, CT 06520-8118 USA URL: http://www.peabody.yale.edu
Copyright © 2005 Peabody Museum of Natural History, Yale University. All rights reserved.No part of this book, except brief quotations by reviewers, may be used or reproduced in any form ormedia, electronic or mechanical (including photocopying, recording, or by any information storage andretrieval system), without the written permission of the Peabody Museum of Natural History.
Library of Congress Cataloging-in-Publication Data
Evolving form and function : fossils and development : proceedings of a symposium honoring AdolfSeilacher for his contributions to paleontology, in celebration of his 80th birthday : April 1-2, 2005, NewHaven, Connecticut / Derek E.G. Briggs, editor.
p. cm.Includes bibliographical references and index.ISBN-10: 0-912532-72-6 (alk. paper)ISBN-13: 978-0-912532-72-1
1. Evolutionary paleobiology--Congresses. I. Seilacher, Adolf. II. Briggs, D. E. G.QE721.2.E85E97 2005560--dc22
2005034103
ISBN-10: 0-912532-72-6
ISBN-13: 978-0-912532-72-1
Printed in the U.S.A.
This paper meets the requirements of ANSI/NISO Z39.48-1992 (Permanence of Paper).∞

Introduction
Adolf Seilacher’s most important contribution to the science of form and function has been toshow how factors other than adaptation influence the morphology of organisms. (His equally im-portant contributions to our understanding of exceptionally preserved fossil deposits [Lager-stätten] and trace fossils fall mainly outside the subject of this symposium.) Seilacher formalizedhis approach to form in a four-page prospectus, published in Lethaia in 1970, that heralded a re-search program in Germany entitled “Konstruktions–Morphologie,” or constructional mor-phology. The approach was illustrated by a triangle (Figure 1), whose corners represent theinfluence of phylogeny and architecture as well as adaptation. Different terms for these cornershave been used in different descriptions of the approach—historical, morphogenetic and func-tional, for example—but the meanings are the same. It is important to note that, in emphasizinghistorical and morphogenetic factors, Seilacher was in no sense rejecting the importance of adap-tation. The principles of constructional morphology formulated by Seilacher have been applied
© 2005 Derek E. G. Briggs. All rights reserved.
From Evolving Form and Function: Fossils and Development: Proceedings of a symposium honoring Adolf Seilacher for his contributions topaleontology, in celebration of his 80th birthday. D.E.G. Briggs, ed. New Haven: Peabody Museum of Natural History, Yale University.ISBN-10: 0-912532-72-6; ISBN-13: 978-0-912532-72-1. © 2005 Peabody Museum of Natural History, Yale University. All rights reserved.
Seilacher on the Science of Form and FunctionDerek E. G. BriggsDepartment of Geology and Geophysics, Yale UniversityP. O. Box 208109, New Haven, CT 06520-8109 USAE-mail: [email protected]
AbstractAdolf Seilacher’s major contribution to our understanding of the evolution of morphology hasbeen to emphasize that function is an important, but far from complete, explanation of organicform. Recognizing the influence of phylogeny and architecture in addition to adaptation,Seilacher formalized this realization in 1970 as “Konstruktions–Morphologie” (constructionalmorphology). This “triangular” approach was very influential at a time when there was littleinterest in constraints on the evolution of form. In 1990, 20 years on, Seilacher expanded thetriangle to include an environmental dimension; although this cannot be measured directly, itis important conceptually. He renamed the investigation of form “morphodynamics.” Seilacherhighlighted the significant role of fabricational noise, including self-organization, intercala-tional hierarchies and morphogenetic countdowns, in determining morphology. He appliedthe methods of constructional morphology to a range of organisms, from vendobionts to bar-nacles, clams to crinoids. Seilacher’s results are illuminated by his unique explanatory termi-nology and the iconography of his line drawings.
KeywordsAdolf Seilacher, constructional morphology, morphodynamics, bivalve mollusc, Ediacara.

successfully to entire organisms as well as to functional units within them (Schmidt-Kittler andVogel 1991). The approach envisions the organism or feature as one point in a dynamic process(like a single frame from a movie); its morphology provides evidence of the influence of differentfactors during its evolution.
The phylogenetic or historical corner of the triangle (see Figure 1) represents the “bauplan,”or diagnostic morphology, of a major group. This inherited aspect of morphology clearly limitsresponses to natural selection. The accretionary growth of molluscs, for example, incorporates themorphology of the juvenile into the adult, and both must be functionally compatible. Arthropods,on the other hand, grow by molting and the larva may metamorphose into an adult with a com-pletely different lifestyle (such as the free swimming cyprid larva, which becomes a sessile goosebarnacle).
The fabricational or architectural corner has been a particular focus in Seilacher’s research onform. It identifies the influence of available materials and self-organizing processes. These factorsare usually expressed in the growth of the organism. The most familiar example is the computersimulation of the logarithmic spiral in the classic papers by Raup (for example, Raup 1966). Im-portant evidence for the underlying principles are provided by repair structures and fabricationalerrors. The emphasis is on the observation that morphology is a response to factors other thanadaptation.
The functional corner of the triangle acknowledges the insights provided by Rudwick’s(1961) paradigm method for analyzing function. The role of adaptation is often difficult to iden-tify as the form of many structures reflects natural selection for several different purposes (crus-tacean appendages, for example, are often used in feeding, locomotion and respiration). Theparadigm method has been applied in its strictest sense to unusual structures that cannot be in-terpreted through a straightforward comparison (Hickman 1988). In such cases alternative func-
4 Evolving Form and Function: Fossils and Development
Figure 1. The original Konstruktions–Morphologie triangle (after Seilacher 1970).

tions can be identified to provide a basis for paradigms that can be compared with the structureunder consideration.
Rudwick’s (1961, 1964) paradigm method provided a robust procedure for analyzing func-tion in fossils (Gould 1970), but it obscured other important influences on the evolution of form.Gould and Lewontin (1979) criticized the adaptationist programme and emphasized the role ofphylogeny and architecture in determining form (they used the spandrels in the dome of St.Mark’s Cathedral in Venice as an example of a morphological feature without any structural func-tion). Their paper advocated a pluralistic approach to understanding form rather than inter-preting the individual attributes of an organism in isolation. Gould and Lewontin (1979:595)acknowledged the emphasis placed on architectural constraints by Seilacher, “whose work de-serves far more attention than it has received.” They highlighted the example of divaricate patterns(structures that diverge from a point and run obliquely to the growth direction) in the shells ofmolluscs and brachiopods (Figure 3). As Seilacher (1972) pointed out, the divaricate pattern isfunctional only in a small proportion of cases: as burrowing sculpture, mimetic color, and to allowlight to penetrate to endosymbiotic algae. Seilacher used divaricate patterns as a major example informalizing the concept of fabricational noise, that is, morphological features that are the resultof architectural constraints rather than functional adaptation, but can be subject to selection(Seilacher 1973). He showed how these patterns lend themselves to co-option as burrowing sculp-tures. In emphasizing that morphology is a response to factors other than adaptation, Seilacher’sinfluence was profound.
Other authors identified additional factors that influence form: Raup (1972) addedchance and ecophenotypic effects in his review of the analysis of morphology; Hickman(1980) added mechanical, ecological, programmatic, maturational and degenerative factors inher consideration of gastropod radulae. Criteria such as these, however, can be subsumedwithin the major factors that defined Seilacher’s (1970) original triangle (see Thomas 1979).
Seilacher on the Science of Form and Function • Briggs 5
Figure 2. Morphodynamics. The conceptual framework of morphodynamics. After Seilacher 1991, fig. 1.Used with kind permission of Springer Science and Business Media.

The original constructional morphology approach was independent of the postulate of evo-lutionary change, but certainly not incompatible with it (Reif and others 1985). Seilacher (1984)regarded constructional morphology as a method of research rather than a theoretical framework,but acknowledged that accumulated results might lead to the identification of wider patterns.Twenty years after the original formulation Seilacher (1991) expanded the triangle to a tetrahe-dron to include an environmental dimension (Figure 2). At the same time he named the analysisof form “morphodynamics” to emphasize that morphology represents the result of evolution, andtherefore encapsulates evidence for the factors that influenced change in form through time(Seilacher 1991). Seilacher (pers. comm. in 2004) argues, however, that “as environmental para-meters cannot be directly measured in fossil examples, this aspect is important only in a concep-tual sense.”
Since 1970 Seilacher has applied the concept of constructional morphology to a range of ex-amples across the taxonomic spectrum. He regards fossil invertebrate skeletons as “a treasure-house of evolutionary experimentation ready for morphodynamic analysis” (pers. comm. in2004). A major achievement of Seilacher’s approach to the analysis of form has been the identifi-cation of features that transcend specific morphologies and taxa—self-organizational patterns, in-tercalational hierarchies, and terminal and iterative countdowns—and the demonstration thatthey are mainly fabricational noise, and not primarily a response to function.
Self-organization
One of the most striking examples of self-organization is the pneu, a fluid- or gas-filled chamber.No Seilacher course on morphodynamics is complete without the demonstration of a water-filledballoon. Ellipsoids and domes form automatically whenever a tensional membrane encloses adenser or pressurized shape (Seilacher 1991). A water-filled balloon adopts a shape similar to thatof many regular echinoids, and Seilacher argued that tensional forces are translated into an echi-noid test that is “automatically optimized” (1991:257) to distribute compressional forces withinthe rigid wall.
Philippi and Nachtigall (1996) used finite element analysis to show that the shape of an echi-noid is a response to complex factors, particularly the mechanical activity of the ambulacral tubefeet (see Smith 2005). The pneu hypothesis of test shape is doubtless an oversimplification, butthat is precisely the point—it provides an explanation of the origin of the shape. The vaulted
6 Evolving Form and Function: Fossils and Development
Figure 3. Zebra patterns. Left to right: Prints of Seilacher’s index fingers, the left and right view of a zebrahead, and the two valves of a specimen of Lioconcha showing the divaricate pattern in the pigment bands.After Seilacher 1991, fig. 9. Used with kind permission of Springer Science and Business Media.
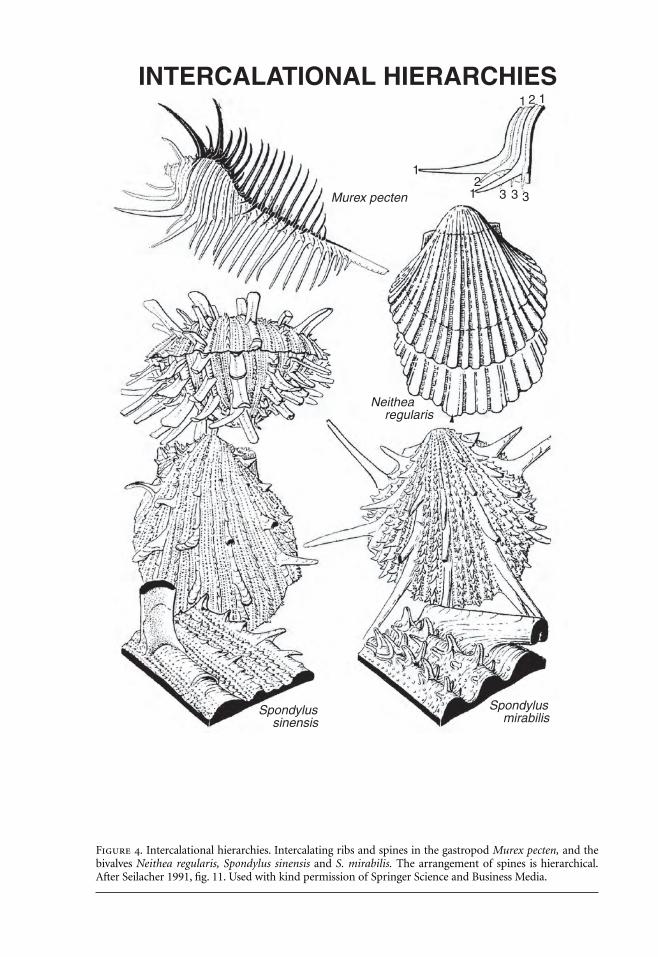
Figure 4. Intercalational hierarchies. Intercalating ribs and spines in the gastropod Murex pecten, and thebivalves Neithea regularis, Spondylus sinensis and S. mirabilis. The arrangement of spines is hierarchical.After Seilacher 1991, fig. 11. Used with kind permission of Springer Science and Business Media.
1
1
domes of echinoid tests, and the configuration of the plates, may reflect the pneu principle eventhough the shape is modifed by differential growth and other factors (Seilacher 1991:258).Seilacher’s (1979) analysis of the constructional morphology of sand dollars showed how the pneushape can be modified for burrowing and sieve feeding with diverse novel features, including al-lometric plate growth and tethering of the upper and lower surfaces with pillars of stereom.Seilacher used the principle of a marginal pneu, likewise modifed by tethering, to explain theshapes of coral polyps (circular in isolation, hexagonal in contact) and the shape of the marginsof bivalves, which grow by accretion (see Seilacher 1991).
Zebra patterns (Figure 3) provide a second example of self-organization. Expressed as linesor spots, in color or sculpture, they are characterized by bilateral asymmetry and occur on manyanimals, not only as the stripes on the eponymous zebra, but in our own fingerprints. Some ofSeilacher’s earliest work (in the 1940s) identified zebra patterns in the ridges on the surface of theteeth of hybodont sharks (see Seilacher 1973). “Normal” zebra patterns occur on nongrowing
8 Evolving Form and Function: Fossils and Development
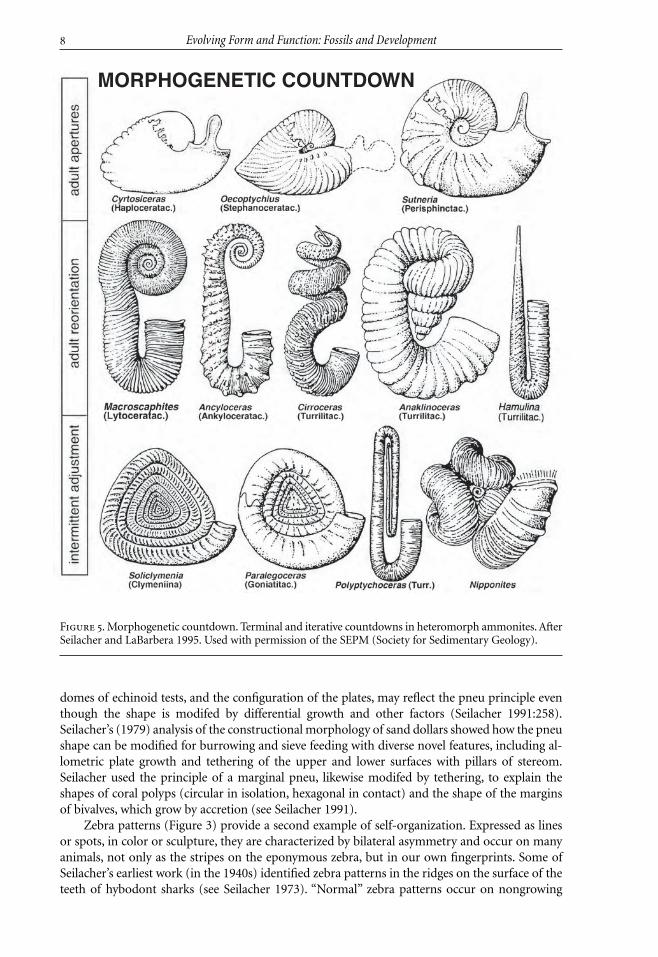
Figure 5. Morphogenetic countdown. Terminal and iterative countdowns in heteromorph ammonites. AfterSeilacher and LaBarbera 1995. Used with permission of the SEPM (Society for Sedimentary Geology).

structures (egg shells, adult cowries) or on expanding skins (fingerprints, mammal skins)(Seilacher and Gunji 1993: 240). The divaricate patterns on bivalves and other shells are the resultof self-organized zebra patterns combined with marginal growth (Seilacher 1991).
Seilacher (1991) considered self-organizing patterns to have begun through a physicalprocess. He envisaged them being brought under genetic control and modified by selection tofulfil a particular function. The evidence for the conflict between self-organization and geneticcontrol is seen in occasional morphological flaws “in which the autonomy of the mechanism ex-presses itself beyond the adaptational straight-jacket” (1991:265). Seilacher has exploited these“mistakes” to identify the nature of the underlying control. He provided an elegant example in apopulation of Strigilla pisiformis from Miami (Seilacher 1984) where the underlying divaricatepattern, which has been modified to provide a burrowing sculpture, is revealed by individuals thathave suffered damage at the shell margin.
Intercalational Hierarchies
Seilacher noted that the cyclic insertion of elements during growth (intercalation) generates a hi-erarchy (Seilacher 1991). This is readily seen in crinoid stems that increase their length by thecyclic introduction of internodals. Similar patterns occur in the ribs of various molluscs and thespines that they bear (Figure 4). Seilacher showed how such intercalation of ribs produces the ex-traordinary arrangement of spines in the gastropod Murex pectin, where rank in the hierarchy de-termines both the size and orientation of the spines. The spines enclose and protect the feedingsnail (Paul 1981). Seilacher (1991) argued that ammonite sutures incorporate elements of an in-tercalational hierarchy in the insertion of tie points where the body membrane was attached to theshell. The shape of the suture, however, was a response to pneu-like bulging between the tie points.
Morphogenetic Countdown
Seilacher used the term “morphogenetic countdown” to describe a change in the pattern ofgrowth in a spiral shell as the adult morphology is reached (Seilacher and Gunji 1993). The con-cept is illustrated by coloration in Nautilus, where color bands disappear on the ventral side of theshell in anticipation of the termination of growth, and the shell becomes countershaded as thefinal aperture forms. Seilacher argued that this change in “terminal” morphology does not implya change in lifestyle, nor a loss of fitness (Seilacher and Gunji 1993).
Seilacher and Gunji (1993) used the concept of morphogenetic countdown to analyze het-eromorphs in gastropods, ammonites and brachiopods. Countdowns are very common in gas-tropods, beginning before the growth of the final aperture and improving fitness only whencomplete. Countdowns extend beyond the self-organization of the logarithmic spiral; inSeilacher’s words they “require a fixed program that must be almost verbally written down in thegenome” (Seilacher and Gunji 1993:245). Countdowns may occur iteratively, a phenomenon par-ticularly well illustrated by muricid gastropods, whose varix spines reflect sequences of morpho-genetic commands superimposed on the underlying program (Seilacher and Gunji 1993; seeFigure 4). Here is a phenomenon that has yet to be explained in terms of evolutionary develop-ment.
The heteromorph ammonites provide a striking example of terminal countdowns (Seilacherand Gunji 1993; Seilacher and LaBarbera 1995; Figure 5). Seilacher noted the tendency for het-eromorphs to turn the margin of the aperture to the horizontal, perhaps to make possible a fil-tering mode of feeding (Seilacher and Gunji 1993). This led to the novel interpretation ofammonites as Cartesian divers (Seilacher and LaBarbera 1995), where the last chamber functionslike an “airbladder” adjusted by muscle action, allowing the animal to move up and down in thewater column as it scans for food. The inferred distribution of lobe and saddle muscles could pro-duce an antagonistic system to lower and raise the last septum (but see Henderson and others2002). Pressurizing the septum with the lobe muscles would result in sinking, while releasing the
Seilacher on the Science of Form and Function • Briggs 9
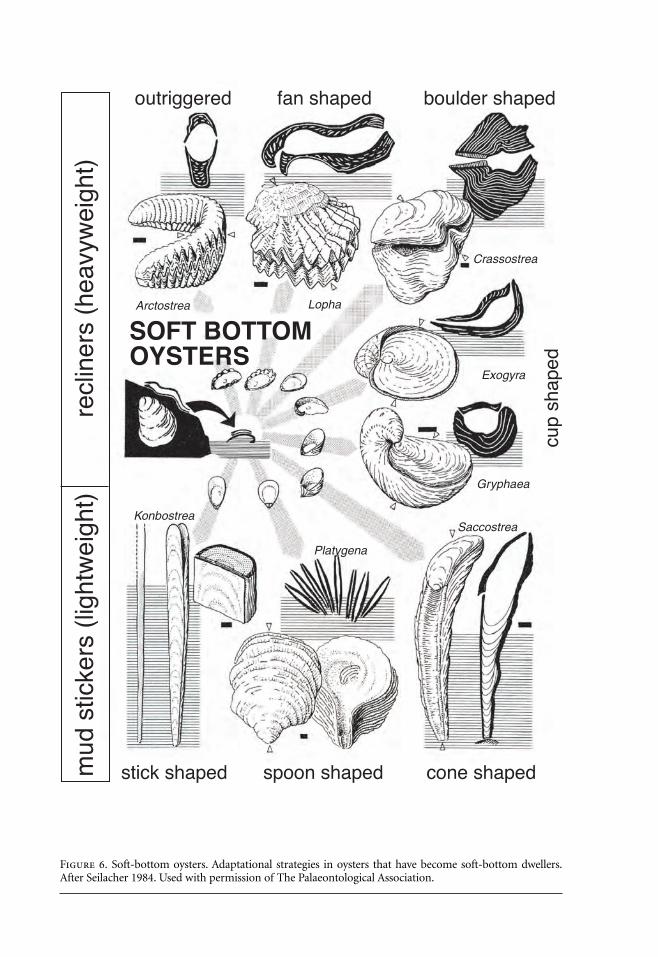
Figure 6. Soft-bottom oysters. Adaptational strategies in oysters that have become soft-bottom dwellers.After Seilacher 1984. Used with permission of The Palaeontological Association.

pressure with the saddle muscles would allow the ammonite to rise in the water column again.The advantage is energy conservation—moving through the water is essentially passive.
This interpretation of heteromorph ammonites is a classic example of the application ofmorphodynamics in place of functional homology (in this case with Nautilus). Seilacher advisesthat we should not be blinkered by comparisons between fossils and their nearest living relatives.The Nautilus model argues for neutral buoyancy and active swimming, difficult to reconcile withsome of the unusual attributes of heteromorph ammonites. Seilacher argued that, while the nau-tiloid septum corresponds to a pneu, the ammonite septum behaves like an elastic membranespread in a frame (Seilacher and LaBarbera 1995). As Seilacher and LaBarbera (1995) pointed out,a difficulty with the Cartesian diver model is that calcification of the membrane in the growingammonite without distortion would seem to require a period during which there was no changein the depth at which the animal was living. The Cartesian diver model may apply only to the laterstages of ammonite evolution.
Constructional Morphology of Bivalve Molluscs
Some of Seilacher’s most compelling applications of constructional morphology have been to bi-valve molluscs. Here he took advantage of the signal provided by convergent morphologies to ex-plore the constraints imposed on the radiation of bivalves in soft-bottom settings (Seilacher 1984,2005b). He considered the function of the elastic ligament that opens the shell, and the role ofburrowing sculptures, particularly those resulting from divaricate patterns. The recolonization ofsoft substrates from sessile attached modes of life is particularly interesting because these bivalveshave lost the foot (which was replaced by the byssus, or cementation for attachment to hard sub-strates) and have no obvious method of anchoring themselves. The colonization of soft substratesoccurred many times and allows a comparison of convergent morphologies. The initial stage in-volves larval attachment to tiny islands of hard-part substrate (shell fragments on the sedimentsurface) and miniaturization of the adult. The major strategy adopted is stabilization of the bodyby shape, weight and size.
In the case of the cemented oysters the growth program had been “derailed” (Seilacher1984:214) to allow them to mimic the substrate, and this may have promoted modification of theshell morphology in derivatives that returned to soft bottoms. They gave rise to a variety of heavy-weight recliners and lightweight mud stickers (Figure 6). Byssate, as opposed to cemented forms,show convergent strategies, but the shells do not become as heavily thickened because the byssusprovides an additional source of stability. Rather than resting on one valve, byssate forms oftenevolved into edgewise recliners. Horn-shaped growth, as in Gryphaea and rudists such as Hippu-rites, allows the shell to be righted passively if it is toppled; erosion creates an upcurrent depres-sion into which the shell rotates.
Seilacher (1998) extended his investigation of bivalves to consider the bizarre rudists of theCretaceous, where the ligament is reduced or absent (he suggested that the shell was opened by aunique system of diductor muscles). The loss of the ligament allowed the two valves of rudists togrow independently: the lower one straight, the upper reduced and anchored as a lid. The pres-ence of diverticles, particularly in the upper valve, indicates that rudists harbored photosymbioticalgae. Rudists, in Seilacher’s view, went beyond “liberation from traditional morphogenetic con-straints” to “deviant lifestyles” (1998:435).
The application of constructional morphology to make comparisons within larger groups,as in Seilacher’s (1984) study of soft-bottom dwelling bivalves and Seilacher and Hauff ’s (2004)on pelagic crinoids, helps to elucidate evolutionary pathways. The approach can also be appliedto a particular clade, as in Thomas’s (1978a, 1978b) analysis of arcoid bivalves, which consid-ered the limits on morphological evolution. Phylogenetic constraints on the evolution of formin arcoid bivalves include: their possession of a spirally coiled calcareous exoskeleton, gills anda muscular foot (as molluscs); their bilateral symmetry, elastic ligament, adductor muscles and
Seilacher on the Science of Form and Function • Briggs 11

Figure 7. Zebra patterns in a micritic crust on a slab of Upper Jurassic limestone from Westerstetten, Ulm,Germany (× 8.75). After Seilacher 1997. Used with permission.

a byssus for attachment (as bivalves); and their shell microarchitecture, with serial ligament andhinge structures (which are specific to arcoids). Fabricational constraints include accretionarygrowth in a spiral, and the simple composition and growth pattern of the ligament. The liga-ment is weak, and is unable to brace the shell firmly against the substrate during burrowing.The simple gill provides an inefficient pump, which may explain why the arcoids did not de-velop siphons. Thus a combination of phylogenetic and fabricational constraints explains whythe radiation of arcoids yielded only generalists, either shallow burrowers or byssate nestlers.
Epibionts
Seilacher pioneered the use of epizoans as important evidence in the interpretation of formand function. He used observations of the standard orientation of a range of epizoan taxa tointerpret the swimming orientation of the ammonite Buchiceras, which they encrusted(Seilacher 1960). This study confirmed that the ammonite lived in the water column, that theshell was not covered by soft parts, and that the adult stage lasted for years. More importantly,Seilacher pointed out that the method could become “a key to the still controversial ecology ofuncoiled and aberrant types” (1960:193). For example, he used traces of barnacle borings inthe guard to confirm the swimming orientation of belemnites (Seilacher 1968). Most recentlyhe showed how some species of barnacle managed to settle on whales (Seilacher 2005a).
Seilacher has also considered the symbiotic relationship between various epizoans and theirhermit crab hosts. Here the morphology of the encruster is controlled by external forces ratherthan morphogenetic information (Seilacher 1991, 2005b). The epizoan grows to maintain its po-sition adjacent to the food supply and to avoid becoming immersed in the substrate on which thecrab lives. Its form is determined by the activities of the hermit crab.
Ediacaran Organisms
One of the most striking examples of Seilacher’s application of the constructional morphologyapproach was to the remarkable Ediacaran fossils of the late Precambrian (see Gehling and others2005), at a time when conventional wisdom held that they all represent metazoans, the ancestorsof modern organisms. Seilacher (1985) was impressed by the anomaly of the widespread occur-rence of a range of soft-bodied organisms preserved (in some cases) in relatively coarse sediment.This taphonomic anomaly has been solved, at least in part, by Gehling’s (1999) death mask hy-pothesis, which explains the preservation of Ediacaran fossils by bacterial precipitation of ironminerals in association with the buried carcasses (Gehling and others 2005). Seilacher explainedthe rarity of this type of preservation in the Phanerozoic as a result of the diversification of bio-turbators (the shift of the seabed from a “matground” to a “mixground”) (Seilacher and Pflüger1994; Seilacher 1999).
More importantly, Seilacher (1985) noted that many of the Ediacaran organisms shared acommon architecture. He deduced that they were constructed as a series of chambers or pneus, astructure akin to quilting, and argued that they could not be interpreted as conventional meta-zoans. “The main function of this provocative hypothesis is to stimulate research in Precambrianpalaeontology and to free it from taxonomic preconceptions” (Seilacher 1985:48). This illustratesone of the major strengths of Seilacher’s approach, his ability to see beyond the organism to itsunderlying organization. In the absence of any phylogenetic constraint, Seilacher (1989) was ableto consider the form of the Ediacaran organisms on purely constructional principles (that is, asarchitecture moderated by functional design). He started with the premise that they “represent anexotic principle of organismic construction” (Seilacher 1989:230). He regarded Dickinsonia, forexample, as a quilted pneu with a thin and flexible outer skin, but with somewhat more rigid in-ternal struts (but see Gehling and others 2005). Seilacher’s “pneu” interpretation suggested thatthe morphology was likely to be a response to a need to maximize surface area for the uptake ofnutrients, and for respiration and excretion. He also considered that these Ediacaran organisms,
Seilacher on the Science of Form and Function • Briggs 13

which he termed Vendobionta (Seilacher 1992), may have harbored bacterial symbionts (Seilacher1989).
Having deduced the architecture of the vendobionts, Seilacher (1989) pointed out thatthere are no living models on which to base an interpretation of function. He recognized thepossibility of radial, unipolar and bipolar growth and, most interestingly, the fractal organiza-tion of some forms. He conceded that his interpretation of the outer skin and internal organi-zation was, of necessity, speculative, but constrained by observations on the nature of theirpreservation. Following research on the specimens found at Mistaken Point in Newfoundland,Seilacher (1992) expanded his analysis of Ediacaran fossils to interpret the various lifestyles ofvendobionts as sediment stickers, flat recliners or erect elevators (once again constraining hisinterpretation of the function of these organisms with observations on the nature of theirpreservation). The interpretation of Ediacaran life developed another novel twist whenSeilacher suggested that many of the vendobionts represent giant protists (Seilacher and others2003), an idea that he had first put forward in 1992 (Seilacher 1992). Seilacher interpreted thoseEdiacaran forms that lived within the microbial mats as xenophyophores (a group of giant ma-rine rhizopodan protists with an agglutinated wall, which today are restricted to abyssaldepths), and considered many of the more familiar vendobionts to be representatives of an ex-tinct class of giant rhizopods (Seilacher and others 2003). This extended interpretation does notalter Seilacher’s analysis of the form of Ediacaran organisms; it simply attempts to make senseof their affinities.
Nonbiological Analogies
Seilacher has a remarkable ability to see pattern and extract lessons from abiotic phenomena suchas balloons, architectural designs, soap bubbles, convection cells in confined fluids and frictionalridges on skis. The experiments that he carried out on candle wax shells with Christian Klug area classic example of this (Seilacher and Klug 1993; Seilacher 1994). Such “shells” are producedwhen molten candle wax flowing into water solidifies. The morphology of the shells is determinedby the temperature of the water: in cold water the shells are narrow and highly convex with promi-nent growth lines; in warmer water they become wider and flatter and the growth lines becomefainter. Seilacher drew a lesson from the similarities to the shells of bivalve molluscs: “regular shellgeometries result from relatively simple sets of parameters and processes” (Seilacher 1994:274),and growth lines are “an essential element of accretionary shell growth” (Seilacher and Gunji1993:239).
Seilacher’s fascination with form led directly to the assembly of his, now internationally trav-elled, exhibition Fossil Art, funded from his 1992 Crafoord Prize. The exhibition presented some34 large slabs with spectacular examples of many of the phenomena that Seilacher has made thesubject of scientific study, from Ediacaran organisms to trace fossils, as well as a small number ofinorganic patterns. These last include sedimentary structures such as mud cracks and ripplemarks, but also a swirling zebra pattern generated by precipitation on a limestone block from theJurassic of Germany (Figure 7). Patterns in nature can be as visually arresting as they are scientif-ically challenging. As Seilacher (1997:60) said, “the limits of ‘art’ are not clearly defined.”
Conclusion
The influence of Seilacher’s research on form and function has been fundamental: several pub-lications have been devoted to constructional morphology (Schmidt-Kittler and Vogel 1991),and the approach has been applied to diverse organisms, stimulating an enormous literature.Seilacher’s ideas are often radical and several, including his interpretations of the Ediacaranbiota, became widely known through the force of his presentations and the power of his per-sonality long before being published in detail (Seilacher 1989). Some encountered resistancefrom reviewers and editors and took some time to appear in the mainstream literature
14 Evolving Form and Function: Fossils and Development

(Seilacher 1994). The long-term influence of Seilacher’s ideas is guaranteed, however, not onlybecause of their scientific content, but also because of his singular explanatory terminology—constructional morphology, morphodynamics, zebra pattern, iterative countdown, sedimentsticker, vendobiont, Fossil Lagerstätten—and his iconic drawing style.
Acknowledgments
I had the privilege of teaching a course on invertebrates with Dolf at Yale in the fall of 2004, whichprovided an opportunity to discuss many of his ideas on the evolution of form. Dolf and EdithSeilacher provided important factual data, including the basis for the Seilacher bibliography (seethe Appendix). Susan Butts prepared the figures based on Seilacher’s originals.
Seilacher on the Science of Form and Function • Briggs 15

Literature Cited
Gehling, J. G. 1999. Microbial mats in terminal Pro-terozoic siliciclastics: Ediacaran death masks.Palaios 14:40–57.
Gehling, J. G., M. L. Droser, S. Jensen and B. N.Runnegar. 2005. Ediacara organisms: relatingform to function. In: D. E. G. Briggs, ed. Evolvingform and function: fossils and development; pro-ceedings of a symposium honoring AdolfSeilacher for his contributions to paleontology, incelebration of his 80th birthday; 2005 Apr 1–2;New Haven. New Haven: Peabody Mus. Nat.Hist., Yale Univ. pp. 43–66.
Gould, S. J. 1970. Evolutionary paleontology and thescience of form. Earth Sci. Rev. 6:77–119.
Gould, S. J. and R. C. Lewontin. 1979. The span-drels of San Marco and the Panglossian para-digm: a critique of the adaptationist programme.Proc. Roy. Soc. London Ser. B 205:581–598.
Henderson, R. A., W. J. Kennedy and W. A.Cobban. 2002. Perspectives of ammonite paleo-biology from shell abnormalities in the genusBaculites. Lethaia 35:215–230.
Hickman, C. S. 1980. Gastropod radulae and the as-sessment of form in evolutionary paleontology.Paleobiology 6:276–294.
—–1988. Analysis of form and function in fossils.Am. Zool. 28:775–793.
Paul, C. R. C. 1981. The function of the spines inMurex (Murex) pecten Lightfoot and relatedspecies (Prosobranchia; Muricidae). J. Conch.London 30:285–294.
Phillipi, U. and W. Nachtigall. 1996. Functionalmorphology of regular echinoid tests (Echino-dermata, Echinoida): a finite element study.Zoomorphology 116:35–50.
Raup, D. M. 1966. Geometric analysis of shell coiling:general problems. J. Paleontol. 40:1178–1190.
—–1972. Approaches to morphologic analysis. In: T.J. M. Schopf, ed. Models in paleobiology. SanFrancisco: Freeman Cooper. pp. 28–44.
Reif, W.-E., R. D. K. Thomas and M. S. Fischer.1985. Constructional morphology: the analysisof constraints in evolution. Acta Biotheor.34:233–248.
Rudwick, M. J. S. 1961. The feeding mechanism ofthe Permian brachiopod Prorichthofenia. Palae-ontology 3:450–471.
—–1964. The inference of function from structure infossils. Brit. J. Philos. Sci. 15:27–40.
Schmidt-Kittler, N. and K. Vogel, eds. 1991.Constructional morphology and evolution.Berlin: Springer-Verlag. 409 pp.
Seilacher, A. 1960. Epizoans as a key to ammonoidecology. J. Paleontol. 34:189–193.
—–1968. Swimming habits of belemnites—recorded
by boring barnacles. Palaeogeogr. Palaeoclimat.Palaeoecol. 4:279–285.
—–1970. Arbeitskonzept zur Konstruktions-Mor-phologie. Lethaia 3:393–396.
—–1972. Divaricate patterns in pelecypod shells.Lethaia 5:325–343.
—–1973. Fabricational noise in adaptive mor-phology. Syst. Zool. 22:451–465.
—–1979. Constructional morphology of sand dol-lars. Paleobiology 5:191–221.
—–1984. Constructional morphology of bivalves:Evolutionary pathways in primary versus sec-ondary soft-bottom dwellers. Palaeontology27:207–237.
—–1985. Discussion of Precambrian metazoans.Philos. Trans. Roy. Soc. London Ser. B 311:47–48.
—–1989. Vendozoa: organismic construction in theProterozoic biosphere. Lethaia 22:229–239.
—–1991. Self-organizing mechanisms in morpho-genesis and evolution. In: N. Schmidt-Kittler andK. Vogel, eds. Constructional morphology andevolution. Berlin: Springer-Verlag. pp. 251–271.
—–1992. Vendobionta and Psammocorallia: lostconstructions of Precambrian evolution. J. Geol.Soc. London 149:607–613.
—–1994. Candle wax shells, morphodynamics andthe Cambrian explosion. Acta Palaeontol. Pol.38:273–280.
—–1997. Fossil art: an exhibition of the GeologischesInstitut Tübingen University, Germany. Tüb-ingen: Geol. Inst., Tübingen Univ. 64 pp.
—–1998. Rudists as bivalvian dinosaurs. In: P. A.Johnston and J. W. Haggart, eds. Bivalves: an eonof evolution. Calgary: Univ. Calgary Pr. pp. 423–436. (Paleobiological studies honoring NormanD. Newell.)
—–1999. Biomat-related lifestyles in the Precam-brian. Palaios 14:86–93.
—–2005a. Whale barnacles: exaptational access to aforbidden paradise. Paleobiology 31(2) Supp:27–35.
—–2005b. Secondary soft-bottom dwellers: conver-gent responses to an evolutionary “mistake.” In:D. E. G. Briggs, ed. Evolving form and function:fossils and development; proceedings of a sym-posium honoring Adolf Seilacher for his contri-butions to paleontology, in celebration of his80th birthday; 2005 Apr 1–2; New Haven. NewHaven: Peabody Museum of Natural History,Yale University. pp. 257–271.
Seilacher, A., D. Grazhdankin and A. Legouta.2003. Ediacaran biota: the dawn of animal life inthe shadow of giant protists. Paleontol. Res. 7:43–54.
Seilacher, A. and Y. P. Gunji. 1993. Morphogeneticcountdown: another view on heteromorph shellsin gastropods and ammonites. Neues. Jahrb.Geol. Paläontol. Abh. 190:73–101.
Evolving Form and Function: Fossils and Development16

Seilacher,A.and R.B.Hauff. 2004.Constructionalmorphology of pelagic crinoids. Palaios 19:3–16.
Seilacher, A. and C. Klug. 1993. Selbst-Organisa-tion bei Kerzenmuscheln. Naturwiss. Rundsch.46:132–134.
Seilacher, A. and M. LaBarbera. 1995. Am-monites as Cartesian divers. Palaios 10:493–506.
Seilacher, A. and F. Pflüger. 1994. From biomatsto benthic agriculture: a biohistoric revolution.In: W. Krumbein, D. M. Paterson and L. J. Stal,eds. Biostabilization of sediments. Oldenburg:Bibl. Infor.-Sys. Univ. Oldenburg. pp. 97–105.
Smith,A.B. 2005.Growth and form in echinoids: theevolutionary interplay of plate accretion versusplate addition. In: D. E. G. Briggs, ed. Evolvingform and function: fossils and development; pro-ceedings of a symposium honoring AdolfSeilacher for his contributions to paleontology, incelebration of his 80th birthday; 2005 Apr 1–2;New Haven. New Haven: Peabody Museum ofNatural History, Yale University. pp. 181–195.
Thomas, R. D. K. 1978a. Shell form and the ecolog-ical range of living and extinct Arcoida. Paleobi-ology 4:181–194.
—–1978b. Limits to opportunism in the evolution ofthe Arcoida (Bivalvia). Philos. Trans. Roy. Soc.London Ser. B 284:335–344.
—–1979. Morphology, constructional. In: R. W. Fair-bridge and D. Jablonski. The encyclopedia of pa-leontology. Stroudsberg: Dowden, Hutchinsonand Ross. pp. 482–487.
Seilacher on the Science of Form and Function • Briggs 17
Appendix: Publications by Adolf Seilacher
1. Seilacher, A. 1943. Elasmobranchierreste ausdem oberen Muschelkalk und dem KeuperWürttembergs. Neues Jahrb. Mineral. Geol.Paläontol. 10(B):256–271, 273–292.
2. Seilacher, A. 1948. Ein weiterer altertümlicherElasmobranchier (Phoebodus keuperinus n. sp.)aus dem Württembergischen Gipskeuper. NeuesJahrb. Mineral. Geol. Paläontol. 1–4(B):24–27.
3. Seilacher, A. 1951. Der Röhrenbau von Laniceconchilega (Polychaeta).Ein Beitrag zur Deutungfossilen Lebensspuren. Senckenbergiana 32:267–280.
4. Seilacher, A. 1953. Studien zur Palichnologie.I. Über die Methoden der Palichnologie. NeuesJahrb. Geol. Paläontol. Abh. 96:421–452.
5. Seilacher, A. 1953. Studien zur Palichnologie.II. Die fossilen Ruhespuren (Cubichnia). NeuesJahrb. Geol. Paläontol. Abh. 98:87–124.
6. Seilacher, A. 1953. Der Brandungssand alsLebensraum in Gegenwart und Vorzeit. Nat.Volk 83:263–273.
7. Seilacher, A. 1954. Die geologische Bedeutungfossiler Lebensspuren. Z. Dtsch. Geol. Ges. 105:214–227.
8. Seilacher, A. 1954. Okologie der TriassischenMuschel Lima lineata (Schloth.) und ihrerEpöken. Neues Jahrb. Geol. Paläontol. Monatsh.4:163–183.
9. Seilacher, A. 1955. Spuren und Lebensweiseder Trilobiten. In: O. H. Schindewolf and A.Seilacher, eds. Beiträge zur Kenntnis des Kam-briums in der Salt Range (Pakistan). Akad. Wiss.Lit. Mainz, Mat.-Nat. Kl.,Abh. 1955(10):342–372.
10. Seilacher, A. 1955. Spuren und Fazies im Un-terkambrium. In: O. H. Schindewolf and A.Seilacher, eds. Beiträge zur Kenntnis des Kam-briums in der Salt Range (Pakistan). Akad. Wiss.Lit.Mainz,Mat.-Nat.Kl.,Abh.1955(10):373–399.
11. Seilacher, A. 1956. Ichnocumulus n. g., eineweitere Ruhespur des schwäbischen Jura. NeuesJahrb. Geol. Paläontol. Monash. 1956:153–159.
12. Seilacher,A.1956. Der Beginn des Kambriumsals biologische Wende. Neues Jahrb. Geol.Paläontol. Abh. 103:155–180.
13. Seilacher, A. 1957. An-aktualistisches Watten-meer? Paläontol. Z. 31:198–206.
14. Seilacher, A. 1958. Zur Ökologischen Charak-teristik von Flysch und Molasse. Eclogae Geol.Helv. 51:1062–1078.
15. Seilacher, A. 1959. Vom Leben der Trilobiten.Naturwissenschaften 46:389–393.
16. Seilacher, A. 1959. Fossilien als Strömungsan-zeiger. Aus der Heimat 67:170–177.
17. Seilacher, A. 1959. Schnecken im Bran-dungssand. Nat. Volk 89:359–366.

18. Seilacher, A. 1960. Epizoans as a key to am-monoid ecology. J. Paleontol. 34:189–193.
19. Seilacher, A. 1960. Strömungsanzeichen imHunsrückschiefer. Notizbl. Hess. Geol. Lan-desamtes Bodenforsch. 88:88–106.
20. Seilacher, A. 1960. Lebensspuren als Leitfos-silien. Geol. Rundsch. 49:41–50.
21. Seilacher,A. 1961. Ein Füllhorn aus dem Hun-srückschiefer. Nat. Volk 91:15–19.
22. Seilacher, A. 1961. Krebse im Brandungssand.Nat. Volk 91:257–264.
23. Seilacher, A. 1961. Holothurien im Hun-srückschiefer (Unter-Devon). Notizbl. Hess.Geol. Landesamtes Bodenforsch. 89:66–72.
24. Seilacher, A. 1962. Paleontological studies onturbidite sedimentation and erosion. J. Geol.70:227–234.
25. Seilacher, A. 1962. Die Sphinctozoa, eineGruppe fossiler Kalkschwämme. Akad. Wiss. Lit.Mainz, Mat.-Nat. Kl., Abh. 1961(10):721–790.
26. Seilacher, A. 1962. Form und Funktion desTrilobiten-Daktylus. Paläontol. Z. (H. SchmidtFestband). pp. 218–227.
27. Seilacher, A. 1963. Lebensspuren und Salin-itäts-Fazies. Fortschr. Geol. Rheinld. Westf. 10:81–94.
28. Seilacher, A. 1963. Kaledonischer Unterbauder Irakiden. Neues Jahrb. Geol. Paläontol.Monash. 10:527–542.
29. Seilacher,A. 1963. Umlagerung und Rolltrans-port von Cephalopoden-Gehäusen. Neues Jahrb.Geol. Paläontol. Monash. 11:593–615.
30. Seilacher, A. 1964. Biogenetic sedimentarystructures. In: J. Imbrie and N. Newell, eds. Ap-proaches to paleoecology. New York: J.Wiley. pp.296–316.
31. Seilacher, A. 1964. Sedimentological classifica-tion and nomenclature of trace fossils. Sedimen-tology 3:253–256.
32. Seilacher,A. and D. Meischner. 1964. Fazies-Analyse im Paläozoikum des Oslo-Gebietes.Geol. Rundsch. 54:596–619.
33. Seilacher, A. 1966. Lobenlibellen und Füll-strukturen bei Ceratiten. Neues Jahrb. Geol.Paläontol. Abh. 125:480–488.
34. Seilacher,A.and C.Hemleben.1966.Beitragezur Sedimentation und Fossilführung des Hun-srückschiefers. 14. Spurenfauna und Bildung-stiefe der Hunsrückschiefer (Unter-Devon).Notizbl. Hess. Geol. Landesamtes Bodenforsch.94:40–53.
35. Seilacher, A. 1967. Tektonischer, sedimentolo-gischer oder biologischer Flysch? Geol. Rundshu.94:40–53.
36. Seilacher, A. 1967. Vorzeitliche Mäander-spuren. In: H. Hediger, ed. Braunschweig: DieStraßen der Tiere. pp. 294–306.
37. Seilacher, A. 1967. Bathymetry of trace fossils.Mar. Geol. 5:413–428.
38. Seilacher, A. 1967. Fossil behavior. Sci. Am.217: 72–80.
39. Seilacher,A.,H.Drozdzewski and R.Haude.1968. Form and function of the stem in apseudoplanctonic crinoid (Seirocrinus). Palae-ontology 11:275–282.
40. Seilacher, A. 1968. Sedimentationsprozesse inAmmonitengehäusen. Akad. Wiss. Lit. Mainz,Mat.-Nat. Kl., Abh. 1967(9):189–203.
41. Seilacher,A. 1968. Swimming habits of belem-nites—recorded by boring barnacles. Palaeo-geogr. Palaeoclimat. Palaeoecol. 4:279–285.
42. Seilacher,A. 1968.Origin and diagenesis of theOriskany Sandstone (Lower Devonian, Ap-palachian) as reflected in its shell fossils. In: G. P.I. T. Müller, ed. Recent developments in car-bonate sedimentology in Central Europe. Hei-delberg: Springer-Verlag. pp. 175–185.
43. Seilacher, A. and T. P. Crimes. 1969. “Euro-pean” species of trilobite burrows in easternNewfoundland. In: M. Kay, ed. North Atlanticgeology and continental drift. Tulsa, OK: Am.Assoc. Pet. Geol. pp. 145–148. (Mem. 12.)
44. Seilacher,A. 1969. Paleoecology of boring bar-nacles. Am. Zool. 9:705–719.
45. Raup, D. and A. Seilacher. 1969. Fossil for-aging behavior: computer simulation. Science166:994–995.
46. Seilacher, A. 1969. Fault-graded beds inter-preted as seismites. Sedimentology 13:155–159.
47. Seilacher, A. 1969. Sedimentary rhythms andtrace fossils in Paleozoic sandstones of Libya. In:Geology, archaeology and prehistory of thesouthwestern Fezzan, Libya. Tripoli: Libya. pp.117–122. (Pet. Explor. Soc. Libya, Ann. Field.Conf., Guidebook 11.)
48. Seilacher, A. 1970. Begriff und Bedeutung derFossil-Lagerstätten. Neues Jahrb. Geol. Paläontol.Monath. 1970:34–39.
49. Seilacher, A. 1970. Cruziana stratigraphy of“non-fossiliferous” Palaeozoic sandstones. In: T.P. Crimes and J. C. Harper, eds. Trace fossils. Liv-erpool: Seel House Pr. pp. 447–476. (Geol. J.Spec. Issue 3.)
50. Seilacher, A. 1970. Arbeitskonzept zur Kon-struktions-Morphologie. Lethaia 3:393–396.
51. Seilacher, A. 1971. Preservational history ofceratite shells. Palaeontology 14:16–21.
52. Goldring, R. and A. Seilacher. 1971. Limulidundertracks and their sedimentologic implica-tions. Neues Jahrb. Geol. Paläontol. Abh.137:422–442.
53. Seilacher, A. and F. Westphal. 1971. “Fossil-Lagerstätten.” In: Sedimentology of parts ofCentral Europe. Frankfurt: Verlag WaldemarKramer. pp. 327–335.
Evolving Form and Function: Fossils and Development18

54. Seilacher, A. 1972. Divaricate patterns in pele-cypod shells. Lethaia 5:325–343.
55. Seilacher, A. 1973. Biostratinomy: the sedi-mentology of biologically standardized particles.In: R. Ginsburg, ed. Evolving concepts in sedi-mentology. Baltimore: Johns Hopkins Univ. Pr.pp. 159–177.
56. Seilacher,A. 1973. Fabricational noise in adap-tive morphology. Syst. Zoo. 22:451–465.
57. Seilacher, A. 1974. Flysch trace fossils: evolu-tion of behavioural diversity in the deep-sea.Neues Jahrb.Geol.Paläontol.Monash.4:233–245.
58. Seilacher, A. 1974. Vorzeitliche Umwelt. DieArbeit des Tübinger Sonderforschungsbereichs53. Tübinger Univ. Zeitung. pp. 54–58.
59. Seilacher, A. 1975. Mechanische Simulationund funktionelle Evolution des Ammoniten-Septums. Paläontol. Z. 49:268–286.
60. Seilacher, A., F. Andalib, G. Dietl and H.Gocht. 1976. Preservational history of com-pressed Jurassic ammonites from southern Ger-many. Neues Jahrb. Geol. Paläontol. Abh.152:307–356.
61. *Seilacher, A. 1976. Echinodermen-Lager-stätten. Zentralbl. Geol. Paläontol. Teil II,1976:227–231.
62. *Seilacher, A. 1976. PhotosymbiontischeMuscheln. Zentralbl. Geol. Paläontol. Teil II,1976:303–305.
63. *Seilacher, A. 1976. Konstruktionsmorpholo-gische Lizenzen des Echinodermen-Skeletts.Zentralbl. Geol. Paläontol. Teil II, 1976:306–311.
64. *Seilacher, A. 1976. Ammoniten-Erhaltung.Zentralbl. Geol. Paläontol. Teil II, 1976:355–362.
65. *Seilacher, A. 1976. Phosphat-Diagenese immarinen Milieu. Zentralbl. Geol. Paläontol. TeilII, 1976: 371–377.
66. *Seilacher, A. 1976. Evolution von Spuren-Vergesellschaftungen. Zentralbl. Geol. Paläontol.Teil II, 1976:396–402.
67. Seilacher, A. 1977. Evolution of trace fossilcommunities. In: A. Hallam, ed. Patterns of evo-lution. Amsterdam: Elsevier. pp. 359–376.
68. Seilacher, A. 1977. Pattern analysis of Paleodic-tyon and related trace fossils. In: T. P. Crimes andJ. C. Harper, eds. Trace fossils 2. Liverpool: SeelHouse Pr. pp. 289–334. (Geol. J. Spec. Issue 9.)
69. Seilacher, A. 1977. Facies problems inCruziana stratigraphy. J. Paleontol. 51:25.
70. Seilacher, A. 1978. Use of trace fossil assem-blages for recognizing depositional environ-ments. In: P. Basan, ed. Trace fossil concepts.Tulsa, OK: Soc. Econ. Paleontol. Mineral. pp.167–180. (Short course 5.)
71. *Seilacher, A. and F. Westphal. 1979. Paleoe-cology, constructions, sedimentology, diagenesisand association of fossils. Neues Jahrb. Geol.Paläontol. Abh. 157:1–2.
72. *Brenner, K. and A. Seilacher. 1979. New as-pects about the origin of the Toarcian PosidoniaShales. Neues Jahrb. Geol. Paläontol.Abh. 157:11–18.
73. *Seilacher, A. and E. Wiesenauer. 1979.Preservational and adaptional history of belem-nites. Neues Jahrb. Geol. Paläontol.Abh. 157:145–149.
74. *Seilacher, A. 1979. Constructional mor-phology and evolutionary ecology of sand dol-lars. Neues Jahrb. Geol. Paläontol. Abh. 157:169–172.
75. *Seilacher, A. 1979. Evolution of trace fossilscommunities in the deep sea. Neues Jahrb. Geol.Paläontol. Abh. 157:251–255.
76. Seilacher, A. 1979. Constructional mor-phology of sand dollars. Paleobiology 5:191–221.
77. Frey, R.W. and A. Seilacher. 1980. Uniformityin marine invertebrate ichnology. Lethaia 13:183–207.
78. Seilacher, A. 1981. Towards an evolutionarystratigraphy. In: Concept and method in paleon-tology: invited papers. Barcelona; 1981 May 5–8.Barcelona: Facultat de Geologia, Univ. Barce-lona. pp. 39–44. (Acta Geol. Hisp. 16.)
79. Seilacher, A. 1981. Konstruktionsmorphologievon Muschelgehäusen. In: W. E. Reif, ed. Funk-tionsmorphologie. Munchen: Paleontol. Gesellsch.,Inst.Paleontol.Hist.Geol.pp.173–186.(Paläontol.Kursbücher 1.)
80. Seilacher,A. 1982.Ammonite shells as habitatsin the Posidonia Shales of Holzmaden—floats orbenthic islands? Neues Jahrb. Geol. Paläontol.Monash. 2:98–114.
81. Seilacher, A. 1982. Posidonia shales (Toarcian,S. Germany)—stagnant basin model revali-dated. In: E. Motanaro Gallitelli, ed. Palaeon-tology, essential of historical geology: proceedingsof the first international meeting on palaeon-tology, essential of historical geology; Venice,Italy; 1981 Jun 2–4. Modena, Italy: S.T.E.M.Mucchi. pp. 279–298.
82. *Seilacher, A. 1982. Erfolgte die Ammoniten-Besiedlung im Holzmadener Posidonienschiefervor oder nach dem Absinken? Neues Jahrb.Geol.Paläontol. Abh. 164:31.
83. *Ghiold, J. and A. Seilacher. 1982. Burrowingstrategies. Neues Jahrb. Geol. Paläontol. Abh.164:221–228.
84. *Chinzei, K., E. Savazzi and A. Seilacher.1982. Adaptational strategies of bivalves living asinfaunal secondary soft bottom dwellers. NeuesJahrb. Geol. Paläontol. Abh. 164:229–244.
85. *Seilacher, A. 1982. “Hammer oysters” as sec-ondary soft bottom dwellers. Neues Jahrb. Geol.Paläontol. Abh. 164:245–250.
86. Seilacher, A. 1982. General remarks about
Seilacher on the Science of Form and Function • Briggs 19

event deposits. In: G. Einsele and A. Seilacher,eds. Cyclic and event stratification. Heidelberg:Springer-Verlag. pp. 161–174.
87. Seilacher,A. 1982.Distinctive features of sandytempestites. In: G. Einsele and A. Seilacher, eds.Cyclic and event stratification. Heidelberg:Springer-Verlag. pp. 333–349.
88. Seilacher, A. 1982. Ammonite shells as habi-tats—floats or benthic islands? In: G. Einsele andA. Seilacher, eds. Cyclic and event stratification.Heidelberg: Springer-Verlag. p. 504.
89. Seilacher, A. 1983. Palökologie—Wechsel-wirkung zwischen geologischen und biologis-chen Prozesser. In: E. Seibold, ed. Forschung inder Bundesrepublik Deutschland. Weinheim:DFG, Verlag Chemie GmbH. pp. 689–696.
90. Seilacher, A. 1983. Upper Paleozoic trace fos-sils from the Gilf-Kebir-Abu-Ras area in south-western Egypt. J. Af. Earth Sci. 1:21–34.
91. Seilacher,A. 1983. Papers on cephalopod pale-obiology and phylogeny. Neues Jahrb. Geol.Paläontol. Abh. 165:327–329.
92. Seilacher,A. 1984. Sedimentary structures ten-tatively attributed to seismic events. Mar. Geol.55:1–12.
93. Seilacher, A. 1984. Storm beds: their signifi-cance in event stratigraphy. In: E. Seibold and J.D. Meulenkamp, eds. Stratigraphy quo vadis?Tulsa, OK: Am. Assoc. Pet. Geol. pp. 49–54.(AAPG Studies in Geol. 16.)
94. Seilacher, A. 1984. Constructional mor-phology of bivalves: evolutionary pathways inprimary versus secondary soft-bottom dwellers.Palaeontology 27:207–237.
95. Seilacher, A. 1984. Late Precambrian and earlyCambrian Metazoa: preservational or real ex-tinctions. In: H. D. Holland and A. F. Trendall,eds. Patterns of change in earth evolution.Dahlem Konferenzen; 1983 May 1–6; Berlin.Heidelberg: Springer-Verlag. pp. 159–168.
96. Seilacher, A. 1985. The Jeram model: eventcondensation in a modern intertidal environ-ment. In: U. Bayer and A. Seilacher, eds. Sedi-mentary and evolutionary cycles. Heidelberg:Springer-Verlag. pp. 336–341. (Lecture Notes inEarth Sci. 1(1).)
97. Seilacher, A., B. A. Matyja and A. Wierz-bowski. 1985. Oyster beds: morphological re-sponse to changing substrate conditions. In: U.Bayer and A. Seilacher, eds. Sedimentary andevolutionary cycles.Heidelberg: Springer-Verlag.pp. 421–435. (Lecture Notes in Earth Sci. 1(1).)
98. Seilacher, A. 1985. Bivalve morphology andfunction. In: D. J. Bottjer, C. S. Hickman and P.D. Ward. Mollusks: notes for a short coursesponsored by the Paleontological Society.Knoxvolle, TN: Univ. TN. Dept. Geol. Sci. pp.88–101. (Studies in Geol. 13.)
99. Seilacher, A., W.-E. Reif and F. Westphal.
1985. Sedimentological, ecological and temporalpatterns of fossil Lagerstätten. Philos. Trans. Roy.Soc. London Ser. B 311:5–23.
100. Seilacher, A. 1985. Discussion of Precambrianmetazoans. Philos. Trans. Roy. Soc. London Ser.B 311:47–48.
101. Seilacher, A. 1985. Trilobite palaeobiology andsubstrate relationships. Trans. Roy. Soc. Edin-burgh 76:231–237.
102. Seilacher, A. and E. Seilacher-Drexler.1986. Sekundäre Weichbodenbewohner unterden Cirripediern. Paläontol. Z. 60:75–92.
103. Alidou, S., J. Lang, J. Bonvalot, E. Roman andA. Seilacher. 1986. Elements de datation enfaveur d'un age Paleozoique et Mesozoique dubassin de Kandi (Nord-Est du Benin, Afrique del'Ouest). J. Af. Earth Sci. 5:339–344.
104. Seilacher,A. 1986. Evolution of behavior as ex-pressed in marine trace fossils. In: M. H. Niteckiand J. A. Kitchell, eds. Evolution of animal be-havior: paleontological and field approaches.New York: Oxford Univ. Pr. pp. 62–87.
105. Seilacher, A., W.-E. Reif, E. Frey, J. Riess andC. Saure. 1987. Bauprinzipien und Evolutionder Skelette von Tieren. Sonderforschungs-bereich Bl/B3, Arcus 1987(1):16–21.
106. Seilacher, A. 1988. Why are nautiloid and am-monite sutures so different? Neues Jahrb. Geol.Paläontol. Abh. 177:41–69.
107. Seilacher, A. and S. Alidou. 1988. Ordovicianand Silurian trace fossils from northern Benin(W. Africa). Neues Jahrb. Geol. Paläontol.7:431–439.
108. Seilacher, A. 1988. Schlangensterne (Aspidura)als Schlüssel zur Entstehungsgeschichte desMuschelkalks. In: H. Hagdorn, ed. NeueForschungen zur Erdgeschichte von Crailsheim.Sonderbände Ges. Nat. Württ. 1:85–98.
109. Seilacher, A. and K. Chinzei. 1988. Selbstbil-dungs-Mechanismen als Prozessoren organis-mischer Entwicklung und Evolution. Leichtbauin Architektur und Natur. Nat. Konstr. 3: 55–63.
110. Seilacher, A. 1989. Vendozoa: organismic con-struction in the Proterozoic biosphere. Lethaia22:229–239.
111. Seilacher, A. 1989. Spirocosmorhaphe, a newgraphoglyptid trace fossil. J. Paleontol. 63:116–117.
112. Seilacher, A. 1990. Aberrations in bivalve evo-lution related to photo- and chemosymbiosis.Hist. Biol. 3:289–311.
113. Seilacher,A. 1990. Paleozoic trace fossils. In: R.Said, ed. The geology of Egypt. Rotterdam:Balkema. pp. 1565–1581.
114. Seilacher,A. 1990. Taphonomy of fossil-Lager-stätten: overview. In: D. E. G. Briggs and P. R.Crowther, eds. Palaeobiology, a synthesis. Ox-ford: Blackwell Sci. Pub. pp. 266–270.
Evolving Form and Function: Fossils and Development20

115. Seilacher, A. 1990. Die Holzmadener Posi-donienschiefer: Entstehung der Fossillagerstätteund eines Erdölmuttergesteins. In: W. K. Wei-dert, ed. Klassische Fundstellen der Paläon-tologie. Korb: Goldschneck-Verlag. pp. 107–131.
116. Seilacher, A. 1990. The sand dollar syndrome:a polyphyletic evolutionary breakthrough. In:M. H. Nitecki, ed. Evolutionary innovations.Chicago: Univ. Chicago Pr. pp. 231–252.
117. Seilacher, A. 1991. Self-organizing mecha-nisms in morphogenesis and evolution. In: N.Schmidt-Kittler and K. Vogel, eds. Construc-tional morphology and evolution. Heidelberg:Springer-Verlag. pp. 251–271.
118. Leszcynski, S. and A. Seilacher. 1991. Ichno-coenoses of a turbidite sole. Ichnos 1:293–303.
119. Alidou, S., J. Lang, J. Bonvalot, E. Roman andA. Seilacher. 1991. Marine influences in the so-called continental sediments of the Paleozoic-Mesozoic Kandi Basin (Northern Benin, WestAfrica). J. Af. Earth Sci. 12:55–65.
120. Seilacher, A. 1991. Precambrian evolutionaryexperiments: Vendozoa and Psammocorallia. In:P. Alberch and G. A. Dover, eds. The referencepoints in evolution. Madrid: Fundacion JuanMarch. pp. 48–53.
121. Seilacher, A. 1991. Morphologic transforma-tion in the wake of behavioral change. In: P. Al-berch and G. A. Dover, eds. The reference pointsin evolution. Madrid: Fundacion Juan March.pp. 80–82.
122. Seilacher, A. 1991. Self-organization: morpho-genetic mechanisms as processors of evolution.Rev. Esp. Paleontol. Extraordin. 5:5–11.
123. Seilacher, A. 1991. Was Fossilien erzählen,oder: zur Taphonomie und Diagenese von Krei-defeuersteinen. Fossilien 4:210–214.
124. Seilacher, A. 1991. Events and their signa-tures—an overview. In: G. Einsele, W. Rickenand A.Seilacher, eds.Cycles and events in stratig-raphy. Heidelberg: Springer-Verlag. pp. 222–226.
125. Seilacher, A. and T. Aigner. 1991. Storm de-position at the bed, facies, and basin scale: the ge-ologic perspective. In: G. Einsele, W. Ricken andA. Seilacher, eds. Cycles and events in stratig-raphy. Heidelberg: Springer-Verlag. pp. 249–267.
126. Brett, C. E. and A. Seilacher. 1991. A tapho-nomic consequence of event sedimentation. In:G. Einsele, W. Ricken and A. Seilacher, eds. Cy-cles and events in stratigraphy. Heidelberg:Springer-Verlag. pp. 283–297.
127. Einsele, G. and A. Seilacher. 1991. Distinc-tion of tempestites and turbites. In: G. Einsele,W. Ricken and A. Seilacher, eds. Cycles andevents in stratigraphy. Heidelberg: Springer-Verlag. pp. 377–382.
128. Pflüger,F.and A.Seilacher. 1991.Flash floodconglomerates. In: G. Einsele, W. Ricken and A.
Seilacher, eds. Cycles and events in stratigraphy.Heidelberg: Springer-Verlag. pp. 383–439.
129. Savrda, C. E., D. J. Bottjer and A. Seilacher.1991. Redox-related benthic events. In: G. Ein-sele, W. Ricken and A. Seilacher, eds. Cycles andevents in stratigraphy. Heidelberg: Springer-Verlag. pp. 524–541.
130. Seilacher, A. 1991. “Medusoid” salt pseudo-morphs. J. Paleontol. 65:330.
131. Seilacher, A. 1991. An updated Cruzianastratigraphy of Gondwanan Paleozoic sand-stones. In: M. J. Salem, O. S. Hammuda and B.A.Eliagoubi, eds. The geology of Libya, Volume 4.Amsterdam: Elsevier. pp. 1565–1581.
132. Seilacher, A. 1992.Vendobionta and Psammo-corallia: lost constructions of Precambrian evo-lution. J. Geol. Soc. London 149:607–613.
133. Seilacher, A. 1992. Whale barnacles: how anevolutionary dream could become true. Mitt.des SFB 230, Nat. Konstr. 8:131–136.
134. Seilacher, A., P. Seilacher and U. Seilacher.1992. Bonebed-Bildungen am heutigen Ama-zonas. Fossilien 4:246–248.
135. Seilacher, A. 1992. Event stratigraphy: dy-namic view of the sedimentary record. In: G.Brown,C.Hawkesworth and C.Wilson,eds.Un-derstanding the earth. Cambridge: CambridgeUniv. Pr. pp. 375–385.
136. Seilacher, A. 1992. How to proceed and suc-ceed in paleontology [feature article]. In: Eu-ropal 1:11–13.
137. Seilacher, A. 1992. Vendobionta: strangest or-ganisms on earth and evolution of trace fossils.In: F. Alvarez, ed. Palaeobiology: preparing forthe twenty-first century. Madrid: Inst. JuanMarch. pp. 57–74. (Centro Reun. Int. Sobre Biol.3.)
138. Seilacher, A. 1992. Self-organization of mor-phologies. In: F. Alvarez, ed. Palaeobiology:preparing for the twenty-first century. Madrid:Inst. Juan March. pp. 89–100. (Centro Reun. Int.Sobre Biol. 3.)
139. Seilacher, A. 1992. Dynamic taphonomy: theprocess-related view of Fossil-Lagerstätten. In: S.Fernandez Lopes ed. Conferencias de la reunionde tafonomia y fosilization. Madrid: Ed. Com-platense. pp. 109–125.
140. Seilacher,A. 1992.Quo vadis, ichnology? In: C.G. Maples and R. R. West, eds. Trace fossils.Knoxville, TN: Paleontol. Soc. pp. 224–238.(Short Courses in Paleontol. 5.)
141. Seilacher, A. 1992. Präkambrisches Leben:Protoplasma in Luftmatratzen. Tübinger Univ.:52: 4–5.
142. Seilacher, A. 1992.Vendobionta als Alternativezu Vielzellern. 34. Phylogenetisches Symposium;1991 Dec 13–15; Hamburg. Mitt. HamburgZool. Mus. Inst. 89:9–20
Seilacher on the Science of Form and Function • Briggs 21

143. Seilacher, A. and F. Pflüger. 1992. Trace fos-sils from the late Proterozoic of North Carolina:early conquest of deep-sea bottoms. In: S.Lidgard and P. R. Crane, eds. Fifth North Amer-ican paleontological convention: abstracts andprogram; 1992 28 Jun–1 Jul; Chicago. Wash-inton, DC: Paleont. Soc. p. 265. (Spec. Pub. 6.)
144. Seilacher, A. 1993. Ammonite aptychi: how totransform a jaw into an operculum. In: P.Dodson and P. Gingerich, eds. Functional mor-phology and evolution. New Haven, CT: KlineGeol. Lab.,Yale Univ. pp. 20–32. (Am. J. Sci. spec.vol. 293A.)
145. Seilacher, A. 1993. Fossil-Lagerstätten imMuschelkalk. In: H. Hagdorn and A. Seilacher,eds. Muschelkalk, Schöntaler Symposium 1991.Sonderbände Ges. Nat. Württ. 2:213–222.
146. Seilacher, A. and C. Klug. 1993. Selbst-Or-ganisation bei Kerzenmuscheln. Naturwiss.Rundsch. 46:132–134.
147. Seilacher, A. 1993. Problems of correlation inthe Nubian sandstone facies. In: U. Thorweiheand H. Schandelmeier, eds. Geoscientific re-search in northeast Africa. Rotterdam: Balkema.pp. 329–333.
148. Skwarko, S. K. and A. Seilacher. 1993. Tracefossils and problematica. In: S. K. Skwarko, ed.Palaeontology of the Permian of Western Aus-tralia. pp. 87, 390–403. (Geol. Surv.W.Aust. Bull.136.)
149. Seilacher, A. and K. Chinzei, eds. 1993.Progress in constructional morphology. NeuesJahrb. Geol. Paläontol. Abh. 190:165–167.
150. Chinzei, K. and A. Seilacher. 1993. Remotebiomineralization I: fill skeletons in vesicularoyster shells. Neues Jahrb. Geol. Paläontol. Abh.190:185–197.
151. Seilacher, A. and K. Chinzei. 1993. Remotebiomineralization II: fill skeletons controllingbuoyancy in shelled cephalopods. Neues Jahrb.Geol. Paläontol. Abh. 190:199–208.
152. Seilacher, A. and Y. P. Gunji. 1993. Morpho-genetic countdown: another view on hetero-morph shells in gastropods and ammonites.Neues Jahrb. Geol. Paläontol. Abh. 190:73–101.
153. Seilacher,A. 1994. Candle wax shells, morpho-dynamics and the Cambrian explosion. ActaPalaeontol. Pol. 38:273–280.
154. Seilacher, A. and E. Seilacher-Drexler.1994. Bivalve trace fossils: a lesson from ac-tuopaleontology. Senckenbergiana Lethaea72:5–15.
155. Buss, L. and A. Seilacher. 1994. The phylumVendobionta: a sister group of the Eumetazoa?Paleobiology 20:1–4.
156. Seilacher, A. 1994. Self-organized structures inthe evolution of early life. In: Natürliche Kon-struktionen, Mitt. des SFB 230, Nat. Konstr.9:303.
157. Seilacher, A. 1994. Early multicellular life: LateProterozoic fossils and the Cambrian explosion.In: S. Bengtson, ed. Early life on earth. New York:Columbia Univ. Pr. pp. 389–400. (Nobel Sympo-sium 84.)
158. Seilacher, A. 1994. How valid is Cruzianastratigraphy? Geol. Rundsch. 83:752–758.
159. Seilacher, A. and F. Pflüger. 1994. From bio-mats to benthic agriculture: a biohistoric revolu-tion. In: W. Krumbein, D. M. Paterson and L. J.Stal, eds. Biostabilization of sediments. Olden-burg: Biblio. Info. Univ. Oldenburg. pp. 97–105.
160. Seilacher,A. 1995. Fossile Kunst: Albumblätterder Erdgeschichte. Korb, Germany: Goldsch-neckverlag. 48 pp.
161. Penalver, E. and A. Seilacher. 1995. Rubielosde Mora: eine untermiozäne Fossil-Lagerstätte.Fossilien 4:211–216.
162. Seilacher, A. and M. LaBarbera 1995. Am-monites as Cartesian divers. Palaios 10:493–506.
163. Seilacher, A. 1995. Black shale models: an ob-servational approach. Europal 8:23–25.
164. Seilacher, A. 1995. Selbstorganisation in derFrühen Evolution des Lebens. Jahresh. Ges.Naturkd. Wüerttemberg 151:73–82.
165. Maeda, H. and A. Seilacher. 1996. Am-monoid taphonomy. In: N. H. Landman, K.Tanabe and R. A. Davis, eds. Ammonoid paleo-biology. New York: Plenum Pr. pp. 543–578.(Topics Geol. 13.)
166. Seilacher, A. 1996. Explosive evolution in thePrecambrian/Cambrian transition. In: E.Molina, ed. Evolucion: aspectos interdiscipli-nares. Zaragoza, Spain: Sem. Interdiscip. Univ.Zaragoza. pp. 15–20. (Cuadernos Interdiscip 6.)
167. Kull, U., W. Maier, W. Nachtigall and A.Seilacher. 1996. Funktions-und Konstruktion-smorphologie. In: K. Teichmann and I. Wilke,eds. Prozess und Form “Natürlicher Konstruk-tionen.” Berlin: Ernst and Sahn Verlag. pp.34–51.
168. Seilacher, A. and R. Goldring. 1996. ClassPsammocorallia (Coelenterata, Vendian-Or-dovician): recognition, systematics and distribu-tion. Geol. Fören. Stockholm För. 118:207–216.
169. Seilacher,A. 1996. Great revolutions in the his-tory of life [in Japanese]. In: R. Morita, ed.Guidebook for the special exhibition of the Nat-ural History Museum and Institute, Chiba, 1996.Chiba, Japan: Syobundo. pp. 2–6.
170. Seilacher, A. 1996. Evolution of burrowing be-havior in Silurian trilobites: ichnosubspecies ofCruziana acacensis. In: M. J. Salem, A. J.Mouzughi and O. S. Hammuda, eds. The ge-ology of Sirt Basin, Volume 1. Amsterdam: Else-vier. pp. 523–530.
171. Gehling, J. G., B. Runnegar and A.Seilacher. 1996. Rasping markings of large
Evolving Form and Function: Fossils and Development22

Metazoan grazers, terminal Neoproterozoic ofAustralia and Cambrian(?) of Saudi Arabia;SEPM meeting abstracts. Tulsa, OK: SEPM (So-ciety for Sedimentary Geology). p. 100.
172. Seilacher, A. 1997. Sandkorallen: Ein aus-gestorbener Lebensformtyp. Fossilien 2:79–84.
173. Fortey, R.A. and A. Seilacher. 1997. The tracefossil Cruziana semiplicata and the trilobite thatmade it. Lethaia 30:105–112.
174. Seilacher,A. 1997. Great revolutions in the his-tory of life. J. Nat. Hist. Mus. Inst. Chiba 4:87–91.
175. Seilacher, A. 1997. Fossil art. J. Nat. Hist. Mus.Inst. Chiba 4:93–95.
176. Seilacher, A. 1997. Fondement écologique etconséquences de l'explosion cambrienne [Theecological base and consequences of the Cam-brian explosion]. Geobios (mem spec.) 21:229–233.
177. Seilacher,A. 1997. Austern und Rudisten—einkonstruktionsmorphologischer Vergleich. Cour.Forsch. Inst. Senckenberg 201:421–432.
178. Seilacher, A. 1997. Warum fossile Fährten oftnur bergauf gehen. Fossilien 6:372–375.
179. Seilacher, A. 1997. Für Eddi Klitzsch. In: H.Schandelmeier and P. Reynolds, eds. Palaeogeo-graphic–palaeotectonic atlas of north-easternAfrica, Arabia, and adjacent areas—Late Neo-proterozoic to Holocene. Rotterdam: Balkema.pp. 15–16.
180. Seilacher, A. 1997. Fossil art: an exhibition ofthe Geologisches Institut Tübingen University,Germany. Drumheller, Alberta, Canada: RoyalTyrell Mus. Palaeontol. 64 pp.
181. Seilacher, A. 1997. The meaning of the Cam-brian explosion. Bull. Natl. Mus. Nat Sci. Peking,China 10:1–9.
182. Seilacher, A. 1998. “Sue”—Das teuerste Fossilaller Zeiten. Fossilien 4:236–238.
183. Seilacher, A. 1998: Rudists as bivalvian di-nosaurs. In: P. A. Johnston and J. Haggart, eds.Bivalves: an eon of evolution. Calgary: Univ. Cal-gary Pr. pp. 423–436. (Paleobiological studieshonoring Norman D. Newell.)
184. Kase, T., P. A. Johnston, A. Seilacher and G.B. Boyce. 1998.Alleged mosasaur marks on LateCretaceous ammonites are limpet (patellogas-tropod) home-scars. Geology 26:947–950.
185. Seilacher, A., P. Bose, P. Pfluger and F.Pflüger. 1998. Triploblastic animals more than1 billion years ago: trace fossil evidence fromIndia. Science 282:80–83.
186. Seilacher, A. 1998. Mosasaurs, limpets or dia-genesis: how Placenticeras shells got punctured.Mitt. Mus. Naturk. Berlin Geowiss. Reihe 1:93–102.
187. Seilacher, A. 1998. Schemas de macroevolu-tion, comment se preparer a l’extinction? [Pat-terns of macroevolution: how to be prepared for
extinction]. C. R. Acad. Sci., Sci. Terre Planet.327:431–440.
188. Seilacher, A. 1999. Oecoptychius Rätsel. Fos-silien 3:131.
189. Seilacher, A. 1999. Pinna. Fossilien 3:138.
190. Seilacher, A. 1999. Biomat-related life styles inthe Precambrian. Palaios 14:86–93.
191. Omarini, R. H., R. I. Sureda, H.-J. Götze, A.Seilacher and F. Pflüger. 1999. Puncoviscanafolded belt in northwestern Argentina: testi-mony of Late Proterozoic Rodinia fragmenta-tion and pre-Gondwana collisional episodes. Int.J. Earth Sci. 88:76–97.
192. Seilacher, A. 1999. Erdgeschichte als Langzeit-Experiment: die großen Revolutionen in der En-twicklung des Lebens. Eclogae Geol. Helv.92:73–79.
193. Keupp, H., M. Röper and A. Seilacher. 1999.Paläobiologische Aspekte von syn vivo-be-siedehen Ammonoideen im Plattenkalk desOber-Kimmeridgiums von Brunn in Ostbayern.Berliner Geowiss. Abh. E 30:121–145.
194. Seilacher-Drexler, E. and A. Seilacher.1999. Undertraces of sea pens and moon snails.Neues Jahrb. Geol. Paläontol. Abh. 214:195–210.
195. Eklöf, J., J. Rydell, J. Fröjmark, M. Johanssonand A. Seilacher. 1999. Orientation of ag-nostid shields in Alum Shale (Upper Cambrian):implications for the depositional environment.Geol. Fören. Stockholm Föerh. 121:301–306.
196. Seilacher, A. 1999. Ediacaran biota. In: R.Singer, ed. Encyclopedia of paleontology.Chicago: Fitzroy Dearborn Pub. pp. 391–397.
197. Seilacher, A. 2000. Fossillagerstätten, Liter-aturbericht. Zentralbl. Geol. Paläontol. Teil II,Heft 5/6, 1999:363–373.
198. Seilacher,A.,A.Meschede,E.W.Bolton andH. Luginsland. 2000. Precambrian “fossil”Ver-miforma is a tectograph. Geology 28:235–238.
199. Seilacher, A. 2000. Ammoniten als KartesischeTaucher? Fossilien 4:231–237.
200. Seilacher, A. and H. Keupp. 2000. Wie sindAmmoniten geschwommen? Fossilien 5:310–313.
201. Seilacher, A. 2000. Leben im Präkambrium.Naturwiss. Rundsch. 629:553–558.
202. Seilacher, A. 2000. Ordovician and Silurianarthrophycid ichnostratigraphy. In: M. A. Solaand D. Worsely, eds. Geological exploration inMurzuq Basin. Amsterdam: Elsevier Sci. pp.237–258.
203. Seilacher, A., C. Marshall, H. C. W. Skinnerand T. Tsuihiji. 2001. A fresh look at sideritic“coprolites.” Paleobiology 27:7–13.
204. Seilacher, A. 2001. Concretion morphologiesreflecting diagenetic and epigenetic pathways.Sediment. Geol. 143:41–57.
Seilacher on the Science of Form and Function • Briggs 23

205. Rydell, J., I. Hammarlund and A. Seilacher.2001. Trace fossils associations in the Mickwitziasandstone (Lower Cambrian): did trilobites re-ally hunt for worms? Geol. Fören. StockholmFöerh. 123:247–250.
206. Omarina, R. H., R. J. Sureda, H. J. Götze, A.Seilacher and F. Pflüger. 2001. Puncovis-cana, folded belt in northwestern Argentina: tes-timony of Late Proterozoic Rodiniafragmentation and pre-Gondwana collisionalepisodes: a reply. Int. J. Earth Sci. 90:894–902.
207. Grazhdankin, D. and Seilacher A. 2002. Un-derground Vendobionta from Namibia. Palae-ontology 45:57–78.
208. Seilacher, A. 2002. Non olet: the strangetaphonomy of coprolites and cololites. In: M. DeRenzi, M. W. P. Alonso, M. Belinchon, E. Pe-nalver, P. Montoya and A. Marques-Aliaga, eds.Current topics on taphonomy and fossilization.Valencia: Ajuntament de Valencia. pp. 233–240.
209. Seilacher, A., S. Lüning, M. A. Martin, E. Kl-itzsch, A. Khja and J. Craig. 2002. Ichnos-tratigraphic correlation of Lower Palaeozoicclastics in the Kufra Basin (SE Libya). Lethaia35:257–262.
210. Seilacher, A. 2003. Vendobionts (Precambrianevolution). In: McGraw-Hill Editorial Staff, ed.McGraw-Hill 2003 yearbook of science andtechnology. New York: McGraw-Hill. pp.458–461.
211. Seilacher, A. 2003. Der Garten von Ediacaraund die Kambrische Explosion. In: W. Hansch,ed. Katastrophen in der Erdgeschichte—Wen-dezeiten des Lebens. Heilbronn: Städ. Mus. pp.70–81. (Museo 19.)
212. Seilacher, A., D. Grazhdankin and A.Legouta. 2003. Ediacaran biota: the dawn ofanimal life in the shadow of giant protists. Pale-ontol. Res. 7:43–54.
213. Seilacher,A. 2003. Anrüchig? Koprolithen undCololithen. Fossilien 4:230–233.
214. Simon, T., H. Hagdorn, M. K. Hagdorn andA. Seilacher. 2003. Swimming trace of a coela-canth fish from the Lower Keuper of south-westGermany. Palaeontology 46:911–926.
215. Seilacher, A., C. Cingolani and R. Varela.2003. Ichnostratigraphic correlation of early Pa-leozoic quartzites in North Africa and centralArgentina. In: M. J. Salem, K. M. Oun and H. M.Dessiq, eds. The geology of northwest Libya.Earth Sci. Soc. Libya 1:275–292.
216. Seilacher, A. and E. Seilacher. 2004. Die Hi-eroglyphen von Moher Cliffs. Fossilien 1:42–45.
217. Seilacher,A. and R. B. Hauff. 2004. Construc-tional morphology of pelagic crinoids. Palaios19:3–16.
218. Seilacher, A. 2004. Constructional “mistakes”in the evolution of blastoid echinoderms. In: T.Heinzeller and J. H. Nebelsick, eds. Echino-
Evolving Form and Function: Fossils and Development24
derms: München. London: Taylor and FrancisGroup. pp. 503–508.
219. Wisshak, M., E. Volohonsky, A. Seilacherand A. Freiwald. 2004. A trace fossil assem-blage from fluvial Old Red deposits (Wood BayFormation; Lower to Middle Devonian) of NW-Spitsbergen, Svalbard. Lethaia 37:149–163.
220. Seilacher, A. 2004. Trittbrettfahrer imMuschelkalkmeer. Fossilien 3:157–160.
221. Seilacher, A. 2004. Principles of ichnostratig-raphy. In: L. A. Buatois and M. G. Mángano, eds.Ichnia 2004. Trelew, Argentina: Mus. Paleontol.Egidio Feruglio. pp. 9–10.
222. Seilacher, A. 2005. Saccocoma: Plankton-Fis-cher im Jurameer. Fossilien 1:48–52.
223. Seilacher, A. 2005. Silurian trace fossils fromAfrica and South America mapping a trans-Gondwanan seaway. Neues Jahrb. Geol.Paläontol. Monash. pp. 129–141.
224. Seilacher, A. 2005. Trace fossils as tourist at-tractions. In: C. Neto de Carvalho, ed. Cruziana’05: paleontological heritage, from discovery torecognition; abstracts from the internationalmeeting on paleontological heritage, geoconser-vation and geotourism; 2005 May 6–7; Idanha-a-Nova, Portugal. Idanha-a-Nova: CentroCultural Raiano. pp. 43–45.
225. Seilacher, A. and MacClintock, C. 2005.Crinoid holdfasts: key to secondary soft-bottomdwelling. Palaios 20:224–240.
226. Seilacher, A. 2005. Pyrit-Sonnen. Globul.11(2003):91–94.
227. Seilacher, A. 2005. Whale barnacles: exapta-tional access to a forbidden paradise. Paleobi-ology 31(2)(suppl.):27–35.
228. Seilacher, A. 2005. Secondary soft-bottomdwellers: convergent responses to an evolu-tionary “mistake.” In: D. E. G. Briggs, ed.Evolving form and function: fossils and develop-ment; proceedings of a symposium honoringAdolf Seilacher for his contributions to paleon-tology, in celebration of his 80th birthday; 2005Apr 1–2; New Haven. New Haven: Peabody Mu-seum of Natural History, Yale University. pp.257–271.
* Reports of Sonderforschungsbereich (SFB) 53, in somecases including results of other members.
![Evolving Hash Functions using Genetic Algorithmsajiips.com.au/papers/V4.1/V4N1.4 - Evolving Hash Functions using... · hash function called "PKP Hash" by Peter.K.Pearson [5] that](https://static.fdocuments.in/doc/165x107/5e3486a76e7276290f0add90/evolving-hash-functions-using-genetic-evolving-hash-functions-using-hash.jpg)