
Evolutionary and biogeographical history of an ancient …boyer/Site/Publications_files/Giribet et...
39
Evolutionary and biogeographical history of an ancient and global group of arachnids (Arachnida: Opiliones: Cyphophthalmi) with a new taxonomic arrangement GONZALO GIRIBET 1 *, PRASHANT P. SHARMA 1 , LIGIA R. BENAVIDES 2 , SARAH L. BOYER 3 , RONALD M. CLOUSE 1,4 , BENJAMIN L. DE BIVORT 1,5 , DIMITAR DIMITROV 6 , GISELE Y. KAWAUCHI 1 , JEROME MURIENNE 1,7 and PETER J. SCHWENDINGER 8 1 Museum of Comparative Zoology, Department of Organismic and Evolutionary Biology, Harvard University, 26 Oxford Street, Cambridge, MA 02138, USA 2 Department of Biological Sciences, The George Washington University, 2023 G Street NW, Washington, DC 20052, USA 3 Biology Department, Macalester College, 1600 Grand Avenue, Saint Paul, MN 55105, USA 4 Division of Invertebrate Zoology, American Museum of Natural History, Central Park West at 79th Street, New York, NY 10024, USA 5 Rowland Institute at Harvard, Harvard University, 100 Edwin Land Boulevard, Cambridge, MA 02142, USA 6 Center for Macroecology, Evolution and Climate, Zoological Museum, University of Copenhagen, Department of Entomology, Universitetsparken 15, DK-2100, Copenhagen, Denmark 7 UMR 7205 CNRS, Département Systématique et Évolution, CP 50, Muséum national d’Histoire naturelle, 45, rue Buffon, 75005 Paris, France 8 Muséum d’histoire naturelle, Département des Arthropodes et d’Entomologie I, 1, Route de Malagnou, Case Postale 6434, CH-1211 Genève 6, Switzerland Received 27 May 2011; revised 8 July 2011; accepted for publication 8 July 2011We investigate the phylogeny, biogeography, time of origin and diversification, ancestral area reconstruction and large-scale distributional patterns of an ancient group of arachnids, the harvestman suborder Cyphophthalmi. Analysis of molecular and morphological data allow us to propose a new classification system for the group; Pettalidae constitutes the infraorder Scopulophthalmi new clade, sister group to all other families, which are divided into the infraorders Sternophthalmi new clade and Boreophthalmi new clade. Sternophthalmi includes the families Troglosironidae, Ogoveidae, and Neogoveidae; Boreophthalmi includes Stylocellidae and Sironidae, the latter family of questionable monophyly. The internal resolution of each family is discussed and traced back to its geological time origin, as well as to its original landmass, using methods for estimating divergence times and ancestral area reconstruction. The origin of Cyphophthalmi can be traced back to the Carboniferous, whereas the diversification time of most families ranges between the Carboniferous and the Jurassic, with the exception of Troglosironidae, whose current diversity originates in the Cretaceous/Tertiary. Ancestral area reconstruction is ambiguous in most cases. Sternophthalmi is traced back to an ancestral land mass that contained New Caledonia and West Africa in the Permian, whereas the ancestral landmass for Neogoveidae included the south-eastern USA and West Africa, dating back to the Triassic. For Pettalidae, most results include South Africa, or a combination of South Africa with the Australian plate of New Zealand or Sri Lanka, as the most likely ancestral landmass, back in the Jurassic. Stylocellidae is reconstructed to the Thai-Malay Penisula during the Jurassic. Combination of the molecular and morphological data results in a hypothesis for all the cyphophthalmid genera, although the limited data available for some taxa represented only in the morphological partition negatively affects the phylogenetic reconstruction by decreasing nodal support in most clades. However, it resolves *Corresponding author. E-mail: [email protected] Biological Journal of the Linnean Society, 2011, ••, ••–••. With 10 figures © 2011 The Linnean Society of London, Biological Journal of the Linnean Society, 2011, ••, ••–•• 1
Transcript of Evolutionary and biogeographical history of an ancient …boyer/Site/Publications_files/Giribet et...

Evolutionary and biogeographical history of an ancientand global group of arachnids (Arachnida: Opiliones:Cyphophthalmi) with a new taxonomic arrangement
GONZALO GIRIBET1*, PRASHANT P. SHARMA1, LIGIA R. BENAVIDES2,SARAH L. BOYER3, RONALD M. CLOUSE1,4, BENJAMIN L. DE BIVORT1,5,DIMITAR DIMITROV6, GISELE Y. KAWAUCHI1, JEROME MURIENNE1,7 andPETER J. SCHWENDINGER8
1Museum of Comparative Zoology, Department of Organismic and Evolutionary Biology, HarvardUniversity, 26 Oxford Street, Cambridge, MA 02138, USA2Department of Biological Sciences, The George Washington University, 2023 G Street NW,Washington, DC 20052, USA3Biology Department, Macalester College, 1600 Grand Avenue, Saint Paul, MN 55105, USA4Division of Invertebrate Zoology, American Museum of Natural History, Central Park West at 79thStreet, New York, NY 10024, USA5Rowland Institute at Harvard, Harvard University, 100 Edwin Land Boulevard, Cambridge, MA02142, USA6Center for Macroecology, Evolution and Climate, Zoological Museum, University of Copenhagen,Department of Entomology, Universitetsparken 15, DK-2100, Copenhagen, Denmark7UMR 7205 CNRS, Département Systématique et Évolution, CP 50, Muséum national d’Histoirenaturelle, 45, rue Buffon, 75005 Paris, France8Muséum d’histoire naturelle, Département des Arthropodes et d’Entomologie I, 1, Route deMalagnou, Case Postale 6434, CH-1211 Genève 6, Switzerland
Received 27 May 2011; revised 8 July 2011; accepted for publication 8 July 2011bij_1774 1..39
We investigate the phylogeny, biogeography, time of origin and diversification, ancestral area reconstruction andlarge-scale distributional patterns of an ancient group of arachnids, the harvestman suborder Cyphophthalmi.Analysis of molecular and morphological data allow us to propose a new classification system for the group;Pettalidae constitutes the infraorder Scopulophthalmi new clade, sister group to all other families, which aredivided into the infraorders Sternophthalmi new clade and Boreophthalmi new clade. Sternophthalmi includesthe families Troglosironidae, Ogoveidae, and Neogoveidae; Boreophthalmi includes Stylocellidae and Sironidae,the latter family of questionable monophyly. The internal resolution of each family is discussed and traced backto its geological time origin, as well as to its original landmass, using methods for estimating divergence timesand ancestral area reconstruction. The origin of Cyphophthalmi can be traced back to the Carboniferous, whereasthe diversification time of most families ranges between the Carboniferous and the Jurassic, with the exceptionof Troglosironidae, whose current diversity originates in the Cretaceous/Tertiary. Ancestral area reconstruction isambiguous in most cases. Sternophthalmi is traced back to an ancestral land mass that contained New Caledoniaand West Africa in the Permian, whereas the ancestral landmass for Neogoveidae included the south-easternUSA and West Africa, dating back to the Triassic. For Pettalidae, most results include South Africa, or acombination of South Africa with the Australian plate of New Zealand or Sri Lanka, as the most likely ancestrallandmass, back in the Jurassic. Stylocellidae is reconstructed to the Thai-Malay Penisula during the Jurassic.Combination of the molecular and morphological data results in a hypothesis for all the cyphophthalmid genera,although the limited data available for some taxa represented only in the morphological partition negativelyaffects the phylogenetic reconstruction by decreasing nodal support in most clades. However, it resolves
*Corresponding author. E-mail: [email protected]
Biological Journal of the Linnean Society, 2011, ••, ••–••. With 10 figures
© 2011 The Linnean Society of London, Biological Journal of the Linnean Society, 2011, ••, ••–•• 1

the position of many monotypic genera not available for molecular analysis, such as Iberosiro, Odontosiro,Speleosiro, Managotria or Marwe, although it does not place Shearogovea or Ankaratra within any existingfamily. The biogeographical data show a strong correlation between relatedness and formerly adjacentlandmasses, and oceanic dispersal does not need to be postulated to explain disjunct distributions, especiallywhen considering the time of divergence. The data also allow testing of the hypotheses of the supposed totalsubmersion of New Zealand and New Caledonia, clearly falsifying submersion of the former, although the datacannot reject the latter. © 2011 The Linnean Society of London, Biological Journal of the Linnean Society,2011, ••, ••–••.
ADDITIONAL KEYWORDS: biogeography – distribution modelling – Gondwana – Laurasia – MAXENT –New Caledonia – New Zealand – Pangea.
INTRODUCTION
The harvestman suborder Cyphophthalmi (Fig. 1)constitutes an ancient lineage of arachnids and wasprobably one of the earliest inhabitants of terrestrialecosystems. Currently distributed on all continentallandmasses (with the exception of Antarctica) and onmost large islands of continental origin, the group isconsidered to have been in close association to theselandmasses since its origins (Juberthie & Massoud,1976; Boyer et al., 2007b). The fact that deep geneticdivergences in cytochrome c oxidase subunit I (COI)have been reported within one species (Boyer, Baker& Giribet, 2007a), and are suspected for many others(R. Clouse & P. Sharma, unpubl. data), corroboratesthe observations that individuals may live a long time(Juberthie, 1960b) and do not disperse far during thecourse of life history. These, together with the oldhistory of the group [a Burmese amber specimenprobably belonging to Stylocellidae is known from theEarly Cretaceous (Poinar, 2008) and the origins of thegroup has been estimated to have taken place duringthe Devonian or Carboniferous using moleculardating techniques (Giribet et al., 2010)] have resultedin a broad use of Cyphophthalmi for biogeographicalinferences and zoogeographical discussions (Rambla,1974; Juberthie & Massoud, 1976; Boyer, Karaman &
Giribet, 2005; Boyer & Giribet, 2007; Clouse &Giribet, 2007; Giribet & Kury, 2007; Boyer et al.,2007b; Boyer & Giribet, 2009; Clouse, de Bivort &Giribet, 2009; Karaman, 2009; Murienne & Giribet,2009; Sharma & Giribet, 2009a; Clouse & Giribet,2010; de Bivort & Giribet, 2010; Murienne, Karaman& Giribet, 2010b; Clouse et al., 2011). These includesome recent and more general debates on the totalsubmersion of large fragment islands such as NewCaledonia and New Zealand (Sharma & Giribet,2009a; Giribet & Boyer, 2010; Giribet et al., 2010).
However, to use a system for biogeographical infer-ences, a sound systematic hypothesis of the group isrequired. The taxonomy of Cyphophthalmi has ben-efitted from the contributions of many studies, espe-cially the synthetic work of Hansen & Sørensen(1904), who produced the first and still best mono-graph on the group, and established the first classi-fication system of the suborder Cyphophthalmi withone family, Sironidae, and two subfamilies, Stylocel-lini (including the genera Stylocellus Westwood, 1874,Ogovia Hansen & Sørensen, 1904, which was pre-occupied and became Ogovea Roewer, 1923, and Miop-salis Thorell, 1890) and Sironini (including PettalusThorell, 1876, Purcellia Hansen & Sørensen, 1904,Siro Latreille, 1796, and Parasiro Hansen &Sørensen, 1904). Another major contributor was
�Figure 1. Habitus. A, Karripurcellia harveyi (Pettalidae) from Warren National Park, Western Australia, July 2004. B,Pettalus thwaitesi (Pettalidae) from Peradeniya Botanical Gardens, Central Province, Sri Lanka, October 2007. C, Rakaiapauli (Pettalidae) from Kelcey’s bush, near Waimate, North Island, New Zealand, February 2008. D, Aoraki longitarsa(Pettalidae) from Governor’s bush, Mt Cook, South Island, New Zealand, January 2006. E, male Pettalus thwaitesi(Pettalidae) from Peradeniya Botanical Gardens, Central Province, Sri Lanka, June 2004. F, Ogovea cameroonensis(Ogoveidae) from Ototomo Forest, Central province, Cameroon, June 2009. G, Parogovia sp. (Neogoveidae) from Mt.Koupé, South-West Province, Cameroon, June 2009. H, two species of Parogovia from Campo Reserve, Littoral Province,Cameroon, June 2009; upper left, adult specimen of Parogovia n. sp.; lower right, juvenile specimen of Parogovia cf.sironoides. I, juvenile specimen of Paramiopsalis ramulosus (Sironidae) from P.N. Peneda Gerés, Portugal, May 2008. J,Paramiopsalis ramulosus (Sironidae) from P.N. Peneda Gerés, Portugal, May 2008. K, Parasiro minor (Sironidae) fromMonte Rasu, Sardinia, Italy, March 2008. L, Suzukielus sauteri (Sironidae) from Mt. Takao, Tokyo Prefecture, Honshu,Japan, April 2005. M, juvenile specimen of Leptopsalis sp. (Stylocellidae) from Bantimurung-Bulusaraung N.P., SulawesiSelatan, Indonesia, June 2006. N, female Leptopsalis sp. (Stylocellidae) from Bantimurung-Bulusaraung N.P., SulawesiSelatan, Indonesia, June 2006.
2 G. GIRIBET ET AL.
© 2011 The Linnean Society of London, Biological Journal of the Linnean Society, 2011, ••, ••–••

AA
C D E
F G
I J K
L M N
H
B
EVOLUTIONARY AND BIOGEOGRAPHICAL HISTORY OF ARACHNIDS 3
© 2011 The Linnean Society of London, Biological Journal of the Linnean Society, 2011, ••, ••–••

Juberthie, who described and monographed manygenera (e.g. Juberthie, 1956, 1958, 1960a, 1961, 1962,1969, 1970a, b; Juberthie & Muñoz-Cuevas, 1970;Juberthie, 1979) in addition to his contributions tothe biology of the group; the regional work of Forster(1948, 1952) in New Zealand, that of Lawrence (1931,1933, 1939, 1963) in South Africa, that of Rambla(Rambla & Fontarnau, 1984, 1986; Rambla, 1991,1994) in the Iberian Peninsula and southeast Asia, tomention just a few of them. More recently, Shear hascontributed with descriptions of numerous species inalmost all cyphophthalmid families (Shear, 1977,1979a, b, 1985, 1993a, b, c; Shear & Gruber, 1996). Healso proposed the bases of modern cyphophthalmidsystematics in a seminal first cladistic analysis of thegroup (Shear, 1980), with five families, three of whichwere new, and two infraorders, equivalent to Hansenand Sørensen’s subfamilies (Table 1). A sixth family,Troglosironidae, was also proposed a few years later(Shear, 1993b).
Two decades after Shear’s classification systemappeared, Giribet (2000) compiled all the cyphoph-thalmid literature to date, recognizing 113 species in26 genera. A subsequent analysis including represen-tatives of most genera and based on a numericalcladistic analysis of 32 morphological characters(Giribet & Boyer, 2002) recognized most of the fami-lies erected by Shear (1980) but also challenged someof his systematic propositions because the root ofthe tree, based on a limited molecular data set alsopublished in the same study, was placed betweenstylocellids and the rest (rendering Tropicophthalmiparaphyletic) or between pettalids and the rest (ren-dering Temperophthalmi paraphyletic). Shear (1993b)had also proposed the new family Troglosironidaeas sister to (Pettalidae + Sironidae) and this resultwas refuted by Giribet & Boyer (2002), who found itnested within an unresolved Neogoveidae. After someminor familial reassignments – Huitaca Shear, 1979was removed from Ogoveidae (Giribet & Prieto, 2003)
and subsequently included in Neogoveidae (Giribet,2007b); Fangensis Rambla, 1994 was transferred fromSironidae to Stylocellidae (Schwendinger & Giribet,2005); Metasiro was transferred from Sironidae toNeogoveidae (Giribet, 2007b); Meghalaya Giribet,Sharma & Bastawade, 2007 was included in Stylocel-lidae (Clouse et al., 2009); and Shearogovea mexasca(Shear, 1977) was excluded from Neogoveidae(Benavides & Giribet, 2007; Giribet, 2011) – the fami-lies are currently considered to be stable.
Recent phylogenetic analyses based on nucleotidesequence data have resolved the relationship amongsome of these families, providing strong support fora relationship of Troglosironidae and Neogoveidae(Boyer et al., 2007b; Boyer & Giribet, 2009; Sharma& Giribet, 2009a; Giribet et al., 2010), a result alsoobtained in a recent analysis of morphometric char-acters (de Bivort, Clouse & Giribet, 2010). Monophylyof Pettalidae is well supported both by discrete andcontinuous morphological characters (Giribet &Boyer, 2002; Giribet, 2003a; Boyer & Giribet, 2007; deBivort et al., 2010; de Bivort & Giribet, 2010), as wellas a diversity of molecular analyses (Boyer & Giribet,2007, 2009; Boyer et al., 2007b; Giribet et al., 2010).Stylocellidae is also well supported based on morphol-ogy (Giribet & Boyer, 2002; Clouse et al., 2009) andmolecules (Schwendinger & Giribet, 2005; Clouse &Giribet, 2007; Boyer et al., 2007b; Clouse et al., 2009;Clouse & Giribet, 2010; Giribet et al., 2010). However,monophyly of Sironidae, especially the membership ofthe Mediterranean genus Parasiro and the JapaneseSuzukielus Juberthie, 1970, remains controversial,both based on morphology (Giribet & Boyer, 2002; deBivort & Giribet, 2004; de Bivort et al., 2010), as wellas on molecular analyses (Boyer et al., 2005; Boyer &Giribet, 2007; Giribet et al., 2010).
In addition to the uncertainty about the monophylyof Sironidae, which we approach here by includingan expanded taxon sampling in problematic generapreviously represented by a single species (Parasiro),we include a much larger diversity of Neogoveidae,both from the Neotropics (29 species versus six usedin Boyer et al., 2007b; including data on the newBrazilian genus Canga DaSilva, Pinto-da-Rocha &Giribet, 2010) and from the Afrotropics (12 terminalsversus seven used in Boyer et al., 2007b). Most impor-tantly, we include the first molecular data on thefamily Ogoveidae, from specimens collected in Cam-eroon in 2009. In total, we provide novel sequencedata for 34 species (of a total of 162 molecular ter-minals), include 27 genera and a family previouslyunsampled, and include new landmasses (Mindanao,the eastern Neotropics, the westernmost distributionof the Afrotropics) not considered in previous phylo-genetic analyses. The present study also providesthe first total evidence analysis of molecules and
Table 1. Classification system of Shear (1980, 1993)
Suborder CyphophthalmiInfraorder Tropicophthalmi Shear, 1980
Superfamily Stylocelloidea Hansen & Sørensen, 1904Family Stylocellidae Hansen & Sørensen, 1904
Superfamily Ogoveoidea Shear, 1980Family Ogoveidae Shear, 1980Family Neogoveidae Shear, 1980
Infraorder Temperophthalmi Shear, 1980Superfamily Sironoidea Simon, 1879
Family Sironidae Simon, 1879Family Pettalidae Shear, 1980Family Troglosironidae Shear, 1993
4 G. GIRIBET ET AL.
© 2011 The Linnean Society of London, Biological Journal of the Linnean Society, 2011, ••, ••–••

morphology for the whole suborder Cyphophthalmiand new data on the timing of diversification andcladogenesis of the group, aiming to revisit interest-ing biogeographical topics. Finally, we provide anestimate of the ancestral area for each lineage andpresent the first habitat suitability and distributionalpatterns analysis for this dispersal-limited, yetglobally-distributed group of arthropods. Studyingmacroecological patterns in Cyphophthalmi is compli-cated as a result of the scarce occurrence data formost species. Thus, species-level assessment of large-scale distributional patterns and their primary eco-logical and evolutionary drivers is difficult. Recently,theoretical and practical arguments for the utility ofmodelling distributional patterns or even ecologicalniche characteristics above the species level havebeen proposed (Heino & Soininen, 2007; Hadly,Spaeth & Li, 2009; Diniz, De Marco & Hawkins,2010). Despite some obvious limitations (Diniz et al.,2010), this approach may be very useful to evaluatepatterns in groups with limited distributional datasuch as insects and other arthropods includingCyphophthalmi.
MATERIAL AND METHODSSPECIMENS
Most specimens used in the molecular part ofthis study (Fig. 2) were collected by one or moreof the authors through direct sifting of leaf litterand transferred to approximately 95% ethanol for
molecular and morphological study. Museum speci-mens have also been used for the morphologicalstudies. A detailed discussion of the specimen col-lecting effort is provided in the Supporting informa-tion (Appendix S1).
The study includes the first molecular data for thefamily Ogoveidae (Ogovea cameroonensis Giribet &Prieto, 2003) and includes additional sampling withinall other families, building upon previous studies onthe phylogenies of Pettalidae (Boyer & Giribet, 2007;Boyer et al., 2007b). Stylocellidae (Clouse et al., 2009;Clouse & Giribet, 2010; Clouse et al., 2011), Troglo-sironidae (Sharma & Giribet, 2009a), and Sironidae(Boyer et al., 2005; Giribet & Shear, 2010; Murienneet al., 2010b). However, data for Neogoveidae wererestricted to a single previous study (Boyer et al.,2007b), and the family was poorly sampled. For theAfrican diversity, we are now able to add data onanother described species, Parogovia gabonica (Juber-thie, 1969), from near its type locality (IpassaReserve, Makokou, Gabon). We also add new data ontwo new species from Mount Koupé and the Camporeserve, in Cameroon, and an additional specimen ofParogovia cf. sironoides also from the Campo reservein Cameroon. The third African species, Parogoviapabsgarnoni Legg, 1990, is known only from its typelocality in Sierra Leone, resulting in a large biogeo-graphical gap from the known distribution of speciesin the Gulf of Guinea, and differs morphologicallyfrom the other species in the genus in many keycharacters, showing very different spermatopositor
Figure 2. Distribution map of the sampled specimens for the molecular study (Table 2). Pettalidae are represented inred, Troglosironidae in purple, Ogoveidae in cyan, Neogoveidae in green, Sironidae in orange, and Stylocellidae in navyblue.
EVOLUTIONARY AND BIOGEOGRAPHICAL HISTORY OF ARACHNIDS 5
© 2011 The Linnean Society of London, Biological Journal of the Linnean Society, 2011, ••, ••–••

from the species in the Gulf of Guinea (Legg & Pabs-Garnon, 1989; Legg, 1990). A possible close relative tothis species is included here, based on data from afemale collected in Ivory Coast. The South Americansampling has also been enriched considerably. Themonotypic genus Huitaca is now represented by datafrom seven species all from Colombia, including thenominal Huitaca ventralis Shear, 1979. Metagovea isnow represented by 13 specimens in 12 putative newspecies, including one from Guyana. These includethe Colombian species identified as Neogovea in ourprevious studies, although they appear to be relatedto Metagovea. A large biogeographical gap in ourprevious studies was the easternmost distribution ofthe genus Neogovea (type species Neogovea immsiHinton, 1938, from Amapá, Brazil). We now includeputative representatives of this clade based on speci-mens collected in French Guiana and Guyana, includ-ing specimens from the recently described N. virginieJocqué & Jocqué, 2011. With the exception of onespecimen from Guyana clustering with Metagovea,these specimens group with the unidentified juvenilefrom Venezuela used in Boyer et al. (2007b) and witha species from the ‘Tepuis’ in Colombia, which wereassign here to the genus Brasilogovea Martens,1969, previously considered a synonym of Neogovea(Shear, 1980; Giribet, 2000). Finally, we were able toinclude data on the monotypic genus Canga based ona specimen from its type locality (DaSilva et al.,2010). However, large areas of the Neotropics withknown specimens of Neogoveidae remain unsampledin our molecular phylogeny, and they should beincluded in future studies (Benavides & Giribet, 2007:fig. 1).
MOLECULAR DATA
DNA extraction, amplification, and sequencing wereperformed as described in several of our previousstudies on molecular systematics of Cyphophthalmiusing the same markers (Schwendinger & Giribet,2005; Boyer et al., 2007b; Boyer & Giribet, 2009;Sharma & Giribet, 2009a; Clouse & Giribet, 2010).We used the five markers as in these previous studies,including the nuclear ribosomal 18S and 28S rRNA,the nuclear protein-encoding histone H3, the mito-chondrial ribosomal 16S rRNA, and the mitochondrialprotein-encoding COI genes. For outgroups, we usedseven noncyphophthalmid Opiliones from the subor-ders Eupnoi, Dyspnoi, and Laniatores (Table 2). Pub-lished sequence data from these studies and the noveldata presented here are deposited in GenBank andare shown in Table 2.
All sequence files for each gene were prepared withMacGDE (Linton, 2005). 18S rRNA sequence datawere divided into six fragments and it was available
for 170 terminals. The 28S rRNA fragment wasdivided into ten regions and was available for 169terminals. 16S rRNA was divided into eight frag-ments and was available for 127 terminals. All theribosomal genes were submitted to direct optimiza-tion or to multiple sequence alignment for homologyassignment. The 143 COI sequences, unlike in manyother organisms, show clade-specific considerablesequence length variation, and hence the gene wasdivided into seven fragments and analyzed underdynamic homology (Wheeler, 2005) or submitted tomultiple sequence alignment. The histone H3 datawere available for 108 terminals and were treated asprealigned in all analyses because they show nolength variation.
MORPHOLOGICAL DATA MATRIX
A morphological matrix of 62 characters was compiledfor 161 taxa based in part on our previous studies(Giribet & Boyer, 2002; Giribet, 2003a; de Bivort &Giribet, 2004; Boyer & Giribet, 2007; de Bivort &Giribet, 2010), direct observation of specimens,mostly through scanning electron microscopy, andcomplemented by some new literature sources(Karaman, 2009). All 19 multistate characters wereunordered. Spermatogenesis in Cyphophthalmi is apromising source of phylogenetic characters, asrecently outlined by Alberti, Giribet & Gutjahr (2009;see also Juberthie & Manier, 1976; Juberthie, Manier& Boissin, 1976; Juberthie & Manier, 1978; Alberti,1995, 2005), although taxon sampling is still sparseand these characters were not considered in this dataset (G. Alberti & G. Giribet, unpubl. data). We did nothave access to specimens of a few species that wereincluded in the data matrix based entirely on litera-ture sources. These have missing data for severalcharacters, especially those observed through scan-ning electron microscopy, such as the prosomalsternal characters. These species include Ankaratrafranzi Shear & Gruber, 1996, Manangotria taolanaroShear & Gruber, 1996, and Odontosiro lusitanicusJuberthie, 1961. Similarly, several CyphophthalmusJoseph, 1868 species, included in our molecularmatrix, were not scored for several morphologicalcharacters because males were never available forexamination, and their published descriptions do notinclude scanning electron micrographs of the relevantcharacters and their descriptions and illustrations areinadequate for scoring those features. Finally, a fewspecies scored in the matrix are not known for onegender, and therefore the corresponding scorings aremissing. The total number of missing cells was thus1798 (17% of cells). In the present study, we were notable to use morphometrics, as we have done in pre-vious studies (Clouse, 2010; de Bivort et al., 2010; de
6 G. GIRIBET ET AL.
© 2011 The Linnean Society of London, Biological Journal of the Linnean Society, 2011, ••, ••–••

Tab
le2.
Spe
cim
enan
dco
llec
tion
data
wit
hG
enB
ank
acce
ssio
nn
um
bers
Vou
cher
Loc
alit
yC
oord
inat
es18
SrR
NA
28S
rRN
A16
SrR
NA
CO
IH
3
FA
MIL
YP
ET
TA
LID
AE
Aor
aki
calc
arob
tusa
wes
tlan
dic
aD
NA
1011
29N
ewZ
eala
nd
-41.
7838
7,17
2.36
903
EU
6736
26D
Q51
8038
DQ
5180
70E
U67
3667
EU
6737
03A
orak
icr
ypta
DN
A10
1289
New
Zea
lan
d-3
7.53
404,
175.
7414
0D
Q51
8000
DQ
5180
43D
Q51
8068
DQ
5181
20D
Q51
8156
Aor
aki
den
ticu
lata
den
ticu
lata
DN
A10
0955
New
Zea
lan
d-4
2.09
283,
171.
3409
6E
U67
3618
EU
6736
54E
U67
3584
DQ
9923
09E
U67
3698
Aor
aki
den
ticu
lata
maj
orD
NA
1009
59N
ewZ
eala
nd
-43.
0340
8,17
1.76
464
EU
6736
20E
U67
3656
EU
6735
85D
Q99
2203
EU
6737
00A
orak
igr
anu
losa
DN
A10
1841
New
Zea
lan
d-3
9.93
478,
175.
6404
6D
Q51
7999
DQ
5180
39D
Q51
8071
––
Aor
aki
hea
lyi
DN
A10
0940
New
Zea
lan
d-4
1.08
673,
174.
1382
6D
Q51
8002
DQ
5180
42D
Q51
8067
DQ
5181
22D
Q51
8160
Aor
aki
iner
ma
DN
A10
0966
New
Zea
lan
d-3
8.79
686,
177.
1247
2E
U67
3622
EU
6736
58–
–E
U67
3702
Aor
aki
lon
gita
rsa
DN
A10
1806
New
Zea
lan
d-4
3.73
666,
170.
0922
2E
U67
3613
EU
6736
52–
DQ
9923
13E
U67
3695
Aor
aki
cf.
tum
idat
aD
OC
094
New
Zea
lan
d-3
9.66
7,17
5.63
7*E
U67
3614
––
DQ
9923
18–
Aor
aki
sp.
DN
A10
1126
New
Zea
lan
d-4
1.08
708,
174.
1370
9E
U67
3624
EU
6736
59–
DQ
9923
19–
Au
stro
purc
elli
aar
ctic
osa
DN
A10
0951
Qu
een
slan
d,A
ust
rali
a-1
6.16
610,
145.
4156
0D
Q51
7984
DQ
5180
23–
DQ
5181
11D
Q51
8147
Au
stro
purc
elli
ad
avie
sae
DN
A10
0947
Qu
een
slan
d,A
ust
rali
a-1
7.24
560,
145.
6420
7D
Q51
7985
DQ
5180
24–
DQ
5181
12D
Q51
8148
Au
stro
purc
elli
afo
rste
riD
NA
1009
45Q
uee
nsl
and,
Au
stra
lia
-16.
0615
1,14
5.46
217
DQ
5179
83D
Q51
8022
DQ
5180
64D
Q51
8110
DQ
5181
46A
ust
ropu
rcel
lia
scop
aria
DN
A10
0946
Qu
een
slan
d,A
ust
rali
a-1
6.59
458,
145.
2792
7D
Q51
7982
DQ
5180
21D
Q51
8065
DQ
5181
08E
U67
3678
Ch
ileo
gove
aoe
dip
us
DN
A10
0413
Ch
ile
-41.
5083
3,-7
2.61
666
DQ
1337
21D
Q13
3733
DQ
5180
55D
Q13
3745
–C
hil
eogo
vea
sp.
DN
A10
0490
Ch
ile
-39.
6666
6,-7
3.28
333*
DQ
1337
22D
Q13
3734
DQ
5180
54D
Q13
3746
EU
6736
72K
arri
purc
elli
ah
arve
yiD
NA
1013
03W
este
rnA
ust
rali
a-3
4.49
500,
115.
9752
7D
Q51
7980
DQ
5180
19D
Q51
8062
DQ
5181
06D
Q51
8143
Neo
purc
elli
asa
lmon
iD
NA
1009
39N
ewZ
eala
nd
-44.
1078
0,16
9.35
527
DQ
5179
98E
U67
3650
DQ
5180
66D
Q82
5638
EU
6736
94P
arap
urc
elli
am
onti
cola
DN
A10
0386
Sou
thA
fric
a-2
9.05
347,
29.3
8516
DQ
5189
73D
Q51
8009
–D
Q51
8098
DQ
5181
35P
arap
urc
elli
asi
lvic
ola
DN
A10
0385
Sou
thA
fric
a-2
8.74
421,
31.1
3763
AY
6394
94D
Q51
8008
DQ
5180
53A
Y63
9582
DQ
5181
36P
etta
lus
thw
aite
siD
NA
1012
23S
riL
anka
7.27
251,
80.5
9333
EU
6735
92E
U67
3633
EU
6735
69E
U67
3666
EU
6736
77P
etta
lus
sp.
DN
A10
1282
Sri
Lan
ka7.
3848
3,80
.816
96D
Q82
5537
EU
6736
32–
DQ
8256
36E
U67
3676
Pet
talu
ssp
.D
NA
1012
83S
riL
anka
7.38
483,
80.8
1696
DQ
5179
74D
Q51
8016
DQ
5180
56D
Q51
8100
DQ
5181
37P
etta
lus
sp.
DN
A10
1285
Sri
Lan
ka6.
9251
7,80
.819
49D
Q51
7976
DQ
5180
17D
Q51
8058
DQ
5181
02D
Q51
8139
Pet
talu
ssp
.D
NA
1012
86S
riL
anka
6.92
517,
80.8
1949
DQ
5179
77D
Q51
8013
DQ
5180
59D
Q51
8103
DQ
5181
40P
etta
lus
sp.
DN
A10
1287
Sri
Lan
ka6.
8235
9,80
.849
96D
Q51
7978
DQ
5180
14D
Q51
8060
DQ
5181
04D
Q51
8141
Pet
talu
ssp
.D
NA
1012
88S
riL
anka
6.55
551,
80.3
7030
DQ
5179
79D
Q51
8015
DQ
5180
61D
Q51
8105
DQ
5181
42P
urc
elli
ail
lust
ran
sD
NA
1003
87S
outh
Afr
ica
-33.
9829
4,18
.424
64E
U67
3589
EU
6736
29D
Q51
8052
EU
6736
65E
U67
3673
Rak
aia
anti
pod
ian
aD
NA
1009
57N
ewZ
eala
nd
-43.
2514
5,17
1.36
761
DQ
5179
88D
Q51
8031
DQ
5180
72D
Q51
8115
DQ
5181
51R
akai
ad
orot
hea
DN
A10
0943
New
Zea
lan
d-4
1.28
163,
174.
9096
1D
Q51
7990
DQ
5180
33D
Q51
8077
DQ
9923
31–
Rak
aia
flor
ensi
sD
NA
1012
95N
ewZ
eala
nd
-40.
8325
8,17
2.96
896
DQ
5179
86D
Q51
8025
DQ
5180
83D
Q51
8113
DQ
5181
49R
akai
ali
nd
sayi
DN
A10
1128
New
Zea
lan
d-4
6.89
327,
168.
1039
8D
Q51
7995
DQ
5180
27D
Q51
8081
DQ
5181
18D
Q51
8154
Rak
aia
mac
raD
NA
1018
08N
ewZ
eala
nd
-45.
9200
0,17
0.02
805
EU
6735
96E
U67
3636
EU
6735
71E
U67
3668
–R
akai
am
agn
aau
stra
lis
DN
A10
0962
New
Zea
lan
d-4
2.33
225,
172.
1712
6E
U67
3601
EU
6736
40E
U67
3575
DQ
9923
33E
U67
3684
Rak
aia
med
iaD
NA
1012
92N
ewZ
eala
nd
-39.
9347
8,17
5.64
046
DQ
5179
96D
Q51
8030
DQ
5180
74D
Q51
8125
DQ
5181
57R
akai
am
inu
tiss
ima
DN
A10
1291
New
Zea
lan
d-3
9.41
641,
175.
2185
8D
Q51
7987
DQ
5180
26D
Q51
8082
DQ
5181
14D
Q51
8150
Rak
aia
pau
liD
NA
1009
68N
ewZ
eala
nd
-44.
7007
3,17
0.96
557
DQ
5179
92D
Q51
8032
DQ
5180
73E
U67
3670
DQ
5181
61R
akai
aso
lita
ria
DN
A10
1294
New
Zea
lan
d-4
1.46
852,
175.
4488
5D
Q51
7997
DQ
5180
29D
Q51
8075
DQ
5181
19D
Q51
8155
Rak
aia
sore
nse
ni
sore
nse
ni
DN
A10
0969
New
Zea
lan
d-4
6.10
967,
167.
6903
4D
Q51
7993
DQ
5180
36D
Q51
8079
DQ
5181
16D
Q51
8153
Rak
aia
sore
nse
ni
dig
itat
aD
NA
1009
70N
ewZ
eala
nd
-46.
5817
7,16
9.20
901
DQ
5179
89D
Q51
8035
DQ
5180
78D
Q51
8123
DQ
5181
62R
akai
ast
ewar
tien
sis
DN
A10
0944
New
Zea
lan
d-4
6.89
327,
168.
1039
8D
Q51
7994
DQ
5180
28D
Q51
8080
DQ
5181
17–
Rak
aia
un
iloc
aD
NA
1018
12N
ewZ
eala
nd
-41.
2208
3,17
3.43
944
EU
6735
99E
U67
3638
–E
U67
3671
–R
akai
asp
.D
NA
1012
97N
ewZ
eala
nd
-40.
9759
5,17
5.11
747
EU
6736
08E
U67
3647
EU
6735
79D
Q99
2344
EU
6736
91R
akai
asp
.D
NA
1018
07N
ewZ
eala
nd
-45.
9016
6,16
9.46
277
EU
6736
06E
U67
3645
––
EU
6736
89R
akai
asp
.D
NA
1009
58N
ewZ
eala
nd
-43.
8086
9,17
3.02
144
EU
6735
97E
U67
3637
EU
6735
73D
Q99
2349
EU
6736
81R
akai
asp
.D
NA
1012
93N
ewZ
eala
nd
-40.
8518
1,17
4.93
233
EU
6736
10E
U67
3649
EU
6735
81D
Q99
2322
EU
6736
93R
akai
asp
.D
NA
1009
54N
ewZ
eala
nd
-41.
1575
7,17
5.02
168
EU
6736
03E
U67
3642
EU
6735
76D
Q99
2348
EU
6736
86
EVOLUTIONARY AND BIOGEOGRAPHICAL HISTORY OF ARACHNIDS 7
© 2011 The Linnean Society of London, Biological Journal of the Linnean Society, 2011, ••, ••–••

Tab
le2.
Con
tin
ued
Vou
cher
Loc
alit
yC
oord
inat
es18
SrR
NA
28S
rRN
A16
SrR
NA
CO
IH
3
FA
MIL
YS
IRO
NID
AE
Cyp
hop
hth
alm
us
corf
uan
us
DN
A10
2111
Gre
ece
39.6
1056
,20
.339
44F
J946
390
FJ9
4641
5F
J946
364
FJ9
4643
8–
Cyp
hop
hth
alm
us
du
rico
riu
sD
NA
1004
87S
love
nia
46.0
1667
,14
.666
67A
Y63
9461
DQ
5131
20A
Y63
9526
AY
6395
56–
Cyp
hop
hth
alm
us
ere
DN
A10
0499
Ser
bia
43.8
3333
,20
.05
AY
6394
62D
Q82
5593
AY
6395
27A
Y63
9557
AY
6394
44C
yph
oph
thal
mu
scf
.gj
orgj
evic
iD
NA
1004
98M
aced
onia
41.9
6667
,21
.35
AY
6394
64D
Q82
5587
AY
6395
29A
Y63
9559
–C
yph
oph
thal
mu
sgo
rdan
iD
NA
1004
95M
onte
neg
ro42
.45,
19.2
6667
AY
6394
67D
Q82
5592
AY
6395
32–
AY
6394
46C
yph
oph
thal
mu
sh
lava
ciD
NA
1020
99C
roat
ia43
.413
44,
16.9
1244
FJ9
4638
4F
J946
409
FJ9
4635
8F
J946
433
–C
yph
oph
thal
mu
sm
arko
iD
NA
1004
97M
aced
onia
41.4
1667
,22
.266
67A
Y63
9469
AY
6395
04A
Y63
9534
AY
6395
61A
Y63
9447
Cyp
hop
hth
alm
us
mar
ten
siD
NA
1004
94M
onte
neg
ro42
.4,
18.7
6667
AY
6394
71D
Q82
5589
AY
6395
36A
Y63
9563
AY
6394
49C
yph
oph
thal
mu
sm
inu
tus
DN
A10
0493
Mon
ten
egro
42.6
5,18
.666
67A
Y63
9473
DQ
8255
91A
Y63
9537
AY
6395
65A
Y63
9450
Cyp
hop
hth
alm
us
ogn
jen
ovic
iD
NA
1010
39B
osn
ia&
Her
zego
vin
a43
.016
67,
18.5
1667
AY
6394
75D
Q82
5594
–A
Y63
9567
AY
6394
51
Cyp
hop
hth
alm
us
rum
ijae
DN
A10
0492
Mon
ten
egro
42.1
6667
,19
.333
33A
Y63
9477
DQ
8255
88A
Y63
9539
AY
6395
69A
Y63
9453
Cyp
hop
hth
alm
us
serb
icu
sD
NA
1020
98S
erbi
a43
.279
17,
22.0
6389
FJ9
4638
3F
J946
408
FJ9
4635
7F
J946
432
–C
yph
oph
thal
mu
ste
yrov
skyi
DN
A10
0910
Mon
ten
egro
42.2
3333
,19
.066
67A
Y63
9482
DQ
5131
18A
Y63
9544
AY
6395
71A
Y63
9454
Cyp
hop
hth
alm
us
treb
inja
nu
sD
NA
1010
38B
osn
ia&
Her
zego
vin
a42
.233
33,
19.1
6667
AY
6394
83D
Q51
3119
–A
Y63
9572
–
Cyp
hop
hth
alm
us
zeta
eD
NA
1009
07M
onte
neg
ro42
.933
33,
18.5
AY
6394
85A
Y63
9515
AY
6395
46A
Y63
9574
AY
6394
56P
aram
iops
alis
edu
ard
oiD
NA
1018
78S
pain
43.4
1718
,-8
.063
56E
U63
8284
EU
6382
87E
U63
8281
EU
6382
88JF
7864
15P
aram
iops
alis
ram
ulo
sus
DN
A10
0459
Spa
in42
.315
80,
-8.4
8697
AY
6394
89D
Q51
3121
AY
6395
50D
Q82
5641
–P
aram
iops
alis
ram
ulo
sus
DN
A10
3538
Por
tuga
l41
.569
44,
-8.1
4027
JF93
4956
JF93
4990
JF93
5023
JF78
6389
–P
aram
iops
alis
sp.
DN
A10
4624
Spa
in43
.319
68,
-6.8
7282
JF93
4957
JF93
4991
JF93
5024
JF78
6390
JF78
6416
Par
asir
oco
iffa
iti
DN
A10
1383
Spa
in42
.152
51,
1.93
039
AY
9188
72D
Q51
3122
AY
9188
77D
Q82
5642
AY
9188
82P
aras
iro
min
orD
NA
1035
35S
ardi
nia
,It
aly
40.4
3047
,9.
0094
8JF
9349
58JF
9349
92JF
9350
25JF
7863
91–
Sir
oac
aroi
des
DN
A10
0488
Ore
gon
,U
SA
44.5
833,
-123
.516
6*A
Y63
9490
DQ
5131
28A
Y63
9551
DQ
8256
43–
Sir
obo
yera
eD
NA
1016
14W
ash
ingt
on,
US
A46
.992
21,
-121
.846
41D
Q51
3139
DQ
5131
25–
DQ
5131
12–
Sir
oca
lave
ras
DN
A10
1623
Cal
ifor
nia
,U
SA
38.2
7744
,-1
20.3
0543
DQ
5131
46D
Q51
3133
*–
––
Sir
oex
ilis
DN
A10
0489
Mar
ylan
d,U
SA
39.4
833,
-79.
4333
AY
6394
91D
Q82
5585
–A
Y63
9579
–S
iro
cf.
kam
iake
nsi
sD
NA
1016
11Id
aho,
US
A47
.746
46,
-116
.702
07D
Q51
3147
DQ
5131
34*
–D
Q51
3115
–S
iro
kam
iake
nsi
sD
NA
1016
13Id
aho,
US
A46
.867
77,
-117
.157
77JF
9349
59JF
9349
93–
––
Sir
oru
ben
sD
NA
1004
57F
ran
ce44
.083
38,
3.58
140
AY
4288
18D
Q82
5584
–D
Q51
3111
–S
iro
shas
taD
NA
1016
22C
alif
orn
ia,
US
A41
.063
67,
-122
.360
45D
Q51
3149
DQ
5131
36*
––
–S
iro
vall
eoru
mD
NA
1004
61It
aly
45.9
833,
9.86
66*
AY
6394
92D
Q51
3123
AY
6395
52A
Y63
9580
AY
6394
57.1
Su
zuki
elu
ssa
ute
riD
NA
1015
43Ja
pan
35.6
3440
,13
9.24
122
DQ
5131
38D
Q51
3116
DQ
5180
86D
Q51
3108
DQ
5181
66S
uzu
kiel
us
sau
teri
DN
A10
1550
Japa
n34
.833
33,
138.
9316
6D
Q82
5541
DQ
8255
83D
Q82
5615
DQ
8256
40D
Q82
5520
FA
MIL
YO
GO
VE
IDA
EO
gove
aca
mer
oon
ensi
sD
NA
1046
17C
amer
oon
3.64
621,
11.2
9079
JF93
4960
JF93
4994
JF93
5026
JF78
6392
JF78
6417
FA
MIL
YT
RO
GL
OS
IRO
NID
AE
Trog
losi
roae
llen
iD
NA
1003
45N
ewC
aled
onia
-21.
1833
,16
5.30
59A
Y63
9497
DQ
8255
80A
Y63
9555
AY
6395
84D
Q51
8164
Trog
losi
roju
bert
hie
iD
NA
1003
44N
ewC
aled
onia
-22.
0500
,16
6.46
67D
Q82
5540
EU
8871
21E
U88
7077
EU
8870
47–
Trog
losi
rolo
ngi
foss
aD
NA
1008
67N
ewC
aled
onia
-22.
3527
,16
6.97
36D
Q51
8089
DQ
8255
82D
Q51
8084
DQ
8256
39D
Q51
8165
Trog
losi
rom
onte
ith
iD
NA
1015
80N
ewC
aled
onia
-21.
6000
,16
5.71
67E
U88
7101
EU
8871
16E
U88
7074
EU
8870
43–
Trog
losi
ron
inqu
aD
NA
1005
77N
ewC
aled
onia
-21.
7500
,16
6.15
00D
Q51
8088
DQ
8255
81D
Q51
8055
DQ
5181
28–
Trog
losi
rou
rban
us
DN
A10
1710
New
Cal
edon
ia-2
2.19
45,
166.
5017
EU
8871
02E
U88
7119
EU
8870
73E
U88
7040
JF78
6340
Trog
losi
row
ilso
ni
DN
A10
2324
New
Cal
edon
ia-2
2.17
70,
166.
5106
EU
8871
07E
U88
7125
EU
8870
75E
U88
7061
–
8 G. GIRIBET ET AL.
© 2011 The Linnean Society of London, Biological Journal of the Linnean Society, 2011, ••, ••–••

Tab
le2.
Con
tin
ued
FA
MIL
YS
TY
LO
CE
LL
IDA
EF
ange
nsi
sin
sula
nu
sD
NA
1003
88,
DN
A10
1063
Th
aila
nd
7.88
5,98
.436
94G
Q48
8337
DQ
8255
51–
GQ
4881
81–
Fan
gen
sis
spel
aeu
sD
NA
1006
69T
hai
lan
d14
.299
72,
98.9
8306
DQ
1337
12D
Q82
5554
GQ
4881
95A
Y63
9583
AY
6394
60L
epto
psal
isly
dek
keri
DN
A10
1064
New
Gu
inea
,In
don
esia
-2.7
1667
,13
4.5*
DQ
1337
17G
Q48
8439
–G
Q48
8153
–
Lep
tops
alis
nov
agu
inea
DN
A10
1510
New
Gu
inea
,In
don
esia
-0.8
333,
134.
0333
*G
Q48
8322
GQ
4884
51G
Q48
8230
––
Lep
tops
alis
sp.
DN
A10
1514
Bor
neo
,M
alay
sia
1.76
667,
110.
3166
7G
Q48
8317
GQ
4884
35D
Q82
5611
GQ
4881
78G
Q48
8114
Lep
tops
alis
sp.
DN
A10
1932
Java
,In
don
esia
-6.7
9833
,10
7.01
583
GQ
4882
64G
Q48
8382
GQ
4882
06G
Q48
8144
GQ
4881
21L
epto
psal
issp
.D
NA
1019
44,
DN
A10
1945
Java
,In
don
esia
-6.7
5694
,10
6.52
333
GQ
4882
67G
Q48
8385
GQ
4882
26G
Q48
8146
GQ
4881
23
Lep
tops
alis
sp.
DN
A10
1093
,D
NA
1004
96T
hai
lan
d6.
67,
101.
15*
GQ
4882
83G
Q48
8402
GQ
4882
21G
Q48
8137
–
Lep
tops
alis
sp.
DN
A10
1483
Mal
aysi
a4.
3969
4,10
2.43
056
DQ
8255
32G
Q48
8454
DQ
8256
10G
Q48
8182
GQ
4881
28L
epto
psal
issp
.D
NA
1014
89M
alay
sia
3.71
639,
101.
7386
1D
Q51
8095
GQ
4884
56D
Q51
8087
GQ
4881
84–
Lep
tops
alis
sp.
DN
A10
1930
Java
,In
don
esia
-6.7
4,10
7.01
278
GQ
4882
62G
Q48
8380
GQ
4882
04–
GQ
4881
19L
epto
psal
issp
.D
NA
1019
37S
ula
wes
i,In
don
esia
1.49
028,
125.
1527
8G
Q48
8298
GQ
4884
22G
Q48
8211
GQ
4881
67G
Q48
8132
Lep
tops
alis
sp.
DN
A10
1938
Su
law
esi,
Indo
nes
ia-5
.042
5,11
9.73
556
GQ
4882
99G
Q48
8359
GQ
4882
12G
Q48
8168
GQ
4881
33L
epto
psal
issp
.D
NA
1020
32S
um
atra
,In
don
esia
-0.1
0583
,10
0.66
389
GQ
4883
08G
Q48
8361
GQ
4881
88G
Q48
8171
GQ
4881
34L
epto
psal
issp
.D
NA
1020
33,
DN
A10
2048
Su
mat
ra,
Indo
nes
ia0.
3461
1,10
0.06
917
GQ
4883
07G
Q48
8428
–G
Q48
8170
–
Lep
tops
alis
sp.
DN
A10
2039
Su
mat
ra,
Indo
nes
ia-0
.945
83,
100.
5436
1G
Q48
8250
GQ
4884
33G
Q48
8190
GQ
4881
75–
Lep
tops
alis
sp.
DN
A10
2042
Su
mat
ra,
Indo
nes
ia3.
2211
1,98
.497
22G
Q48
8314
GQ
4884
34G
Q48
8213
GQ
4881
76G
Q48
8136
Lep
tops
alis
sp.
DN
A10
2061
Su
mat
ra,
Indo
nes
ia-0
.477
22,
100.
3538
9G
Q48
8312
GQ
4884
32–
GQ
4881
74G
Q48
8135
Lep
tops
alis
sp.
DN
A10
3250
Th
aila
nd
9.76
667,
98.4
1389
GQ
4882
78G
Q48
8394
GQ
4881
92G
Q48
8155
–M
egh
alay
asp
.D
NA
1010
94,
DN
A10
1500
Th
aila
nd
7.88
528,
98.4
3722
DQ
8255
34G
Q48
8352
–G
Q48
8158
GQ
4881
27
Meg
hal
aya
sp.
DN
A10
1494
,D
NA
1015
06,
DN
A10
1765
Th
aila
nd
9.91
806,
98.9
4278
DQ
8255
30G
Q48
8398
–D
Q82
5632
–
Meg
hal
aya
sp.
DN
A10
1767
Th
aila
nd
6.97
,10
0.10
*G
Q48
8276
GQ
4883
90–
GQ
4881
54–
Meg
hal
aya
sp.
DN
A10
2051
Indi
a25
.507
78,
90.2
3167
GQ
4882
61G
Q48
8379
––
GQ
4881
18M
egh
alay
asp
.D
NA
1032
42,
DN
A10
3243
,D
NA
1032
44
Ch
ina
27.6
8833
,98
.277
78G
Q48
8233
GQ
4883
77G
Q48
8219
–G
Q48
8117
Meg
hal
aya
sp.
DN
A10
3251
Th
aila
nd
9.76
667,
98.4
1389
GQ
4882
80G
Q48
8396
GQ
4881
96–
–M
egh
alay
asp
.D
NA
1032
65T
hai
lan
d14
.25,
101.
98*
GQ
4882
39G
Q48
8392
––
–M
iops
alis
sp.
DN
A10
1519
Bor
neo
,In
don
esia
0.64
,11
7.09
*D
Q82
5526
GQ
4884
36G
Q48
8228
GQ
4881
80G
Q48
8115
Mio
psal
issp
.D
NA
1032
59B
orn
eo,
Mal
aysi
a5.
81,
116.
24*
GQ
4882
60G
Q48
8375
GQ
4881
93G
Q48
8142
GQ
4881
16M
iops
alis
sp.
DN
A10
1468
,D
NA
1019
50B
orn
eo,
Indo
nes
ia0.
64,
117.
09*
GQ
4883
28G
Q48
8444
––
–
Mio
psal
issp
.D
NA
1015
17B
orn
eo,
Indo
nes
ia1.
0666
7,11
7.83
333*
DQ
8255
27.1
DQ
8255
64–
GQ
4881
79D
Q82
5508
.1M
iops
alis
sp.
DN
A10
2032
,D
NA
1020
53,
DN
A10
2058
Su
mat
ra,
Indo
nes
ia-0
.105
83,
100.
6638
9G
Q48
8305
GQ
4884
25–
––
Mio
psal
issp
.D
NA
1032
49B
orn
eo,
Mal
aysi
a6.
01,
116.
53*
GQ
4882
52G
Q48
8367
GQ
4881
94G
Q48
8139
–M
iops
alis
sp.
DN
A10
3254
Bor
neo
,M
alay
sia
1.75
833,
110.
3297
2G
Q48
8257
GQ
4883
71–
––
Mio
psal
issp
.D
NA
1049
81M
inda
nao
,P
hil
ippi
nes
6.48
,12
5.09
*H
Q59
3868
HQ
5938
69–
HQ
5938
70H
Q59
3871
FA
MIL
YN
EO
GO
VE
IDA
EB
rasi
logo
vea
sp.
DN
A10
1665
Col
ombi
a0.
1797
2,-7
2.62
333
JF93
4963
JF93
5011
JF93
5028
JF78
6414
JF78
6430
‘Bra
silo
gove
a’sp
.To
boga
nD
NA
1008
69V
enez
uel
a5.
6500
0,-6
7.63
333
DQ
8255
45D
Q82
5600
DQ
8256
17–
–
EVOLUTIONARY AND BIOGEOGRAPHICAL HISTORY OF ARACHNIDS 9
© 2011 The Linnean Society of London, Biological Journal of the Linnean Society, 2011, ••, ••–••

Tab
le2.
Con
tin
ued
Vou
cher
Loc
alit
yC
oord
inat
es18
SrR
NA
28S
rRN
A16
SrR
NA
CO
IH
3
Can
gare
nat
aeD
NA
1056
80B
razi
l-6
.410
55,
-50.
3231
9JF
9349
64JF
9349
97JF
9350
29JF
7863
95JF
7864
20H
uit
aca
ven
tral
isD
NA
1016
74C
olom
bia
7.41
667,
-72.
4333
3JF
9349
80JF
9350
14–
JF78
6399
–H
uit
aca
sp.
DN
A10
1683
Col
ombi
a3.
5583
3,-7
6.58
278
JF93
4979
JF93
5016
–JF
7863
96JF
7864
21H
uit
aca
sp.
DN
A10
1407
Col
ombi
a5.
7795
6,-7
3.45
377
DQ
5180
90D
Q82
5596
DQ
5180
50D
Q51
8129
DQ
5181
67H
uit
aca
sp.
DN
A10
1681
Col
ombi
a5.
0995
6,-7
5.40
594
JF93
4977
JF93
5015
JF93
5032
JF78
6397
JF78
6422
Hu
itac
asp
.D
NA
1021
50C
olom
bia
3.55
833,
-76.
5827
8JF
9349
81JF
9350
12JF
9350
33–
JF78
6423
Hu
itac
asp
.D
NA
1046
46C
olom
bia
3.55
833,
-76.
5827
8JF
9349
82JF
9350
17JF
9350
30–
–H
uit
aca
sp.
DN
A10
1671
Col
ombi
a7.
4166
7,-7
2.43
333
JF93
4978
JF93
5013
JF93
5031
JF78
6398
JF78
6424
Met
agov
easp
.D
NA
1016
80C
olom
bia
5.09
542,
-75.
3907
5JF
9349
72JF
9349
99JF
9350
36JF
7864
00–
Met
agov
easp
.D
NA
1046
47C
olom
bia
3.55
833,
-76.
5827
8JF
9349
88JF
9350
03JF
9350
37–
–M
etag
ovea
sp.
DN
A10
4648
Col
ombi
a3.
5583
3,-7
6.58
278
JF93
4989
JF93
5004
JF93
5038
––
Met
agov
easp
.D
NA
1014
08C
olom
bia
-4.0
4495
,-6
9.98
979
DQ
8255
43D
Q82
5598
DQ
8256
18–
DQ
8255
14M
etag
ovea
sp.
DN
A10
1410
Col
ombi
a1.
2850
0,-7
8.07
367
DQ
5180
91D
Q82
5597
JF93
5034
GQ
9128
60D
Q51
8168
Met
agov
easp
.D
NA
1016
54C
olom
bia
1.25
,-7
8.25
*JF
9349
70JF
9350
00JF
9350
35JF
7864
01JF
7864
25M
etag
ovea
sp.
DN
A10
2151
Col
ombi
a5.
4858
3,-7
6.01
667
JF93
4971
JF93
5001
–JF
7864
02–
Met
agov
easp
.D
NA
1016
85C
olom
bia
3.56
889,
-76.
5886
1JF
9349
73JF
9349
98–
JF78
6403
–M
etag
ovea
sp.
DN
A10
1670
Col
ombi
a1.
6163
9,-7
6.10
417
JF93
4984
–5JF
9350
02–
––
Met
agov
easp
.D
NA
1014
09C
olom
bia
1.28
500,
-78.
0736
7D
Q82
5544
DQ
8255
99D
Q82
5619
DQ
8256
46JF
7864
26M
etag
ovea
sp.
DN
A10
1642
Col
ombi
a3.
5583
3,-7
6.58
278
JF93
4986
JF93
5006
JF93
5042
––
Met
agov
easp
.D
NA
1016
86C
olom
bia
3.56
889,
-76.
5886
1JF
9349
87JF
9350
05–
JF78
6404
–M
etag
ovea
sp.
DN
A10
5826
Gu
yan
a1.
3365
5,-5
8.96
510
JF93
4983
JF93
5010
JF93
5041
JF78
6408
–M
etas
iro
amer
ican
us
DN
A10
1532
Flo
rida
,U
SA
30.5
6489
,-8
4.95
163
DQ
8255
42D
Q82
5595
DQ
8256
16D
Q82
5645
DQ
8255
13M
etas
iro
amer
ican
us
DN
A10
5644
Sou
thC
arol
ina,
US
A35
.062
31,
-82.
795
JF93
4961
JF93
4995
–JF
7863
93JF
7864
18M
etas
iro
amer
ican
us
DN
A10
5645
Sou
thC
arol
ina,
US
A32
.189
23,
-81.
08JF
9349
62JF
9349
96JF
9350
27JF
7863
94JF
7864
19N
eogo
vea
virg
inie
DN
A10
4823
Fre
nch
Gu
ian
a4.
1951
1,-5
2.14
936
JF93
4974
JF93
5007
–JF
7864
05–
Neo
gove
avi
rgin
ieD
NA
1058
24F
ren
chG
uia
na
4.08
813,
-52.
6752
0JF
9349
75JF
9350
08JF
9350
39JF
7864
06–
Neo
gove
asp
.D
NA
1058
25G
uya
na
1.38
803,
-58.
9463
2JF
9349
76JF
9350
09JF
9350
40JF
7864
07–
Par
ogov
iaga
bon
ica
DN
A10
4620
Gab
on0.
5044
8,12
.795
25JF
9349
69JF
9350
19JF
9350
47JF
7864
11–
Par
ogov
iasi
ron
oid
esD
NA
1010
59B
ioko
,E
quat
oria
lG
uin
ea3.
7257
0,8.
8382
8D
Q51
8092
DQ
8256
06D
Q51
8051
DQ
5181
31D
Q51
8169
Par
ogov
iasi
ron
oid
esD
NA
1010
61B
ioko
,E
quat
oria
lG
uin
ea3.
7028
4,8.
8752
0D
Q82
5550
DQ
8256
07JF
9350
43D
Q82
5650
DQ
8255
19
Par
ogov
iacf
.si
ron
oid
esD
NA
1004
62E
quat
oria
lG
uin
ea2.
1830
5,9.
8030
5A
Y63
9493
DQ
8256
03–
–A
Y63
9459
Par
ogov
iacf
.si
ron
oid
esD
NA
1010
53E
quat
oria
lG
uin
ea1.
6581
5,10
.311
43D
Q82
5548
DQ
8256
04D
Q82
5623
–D
Q82
5517
Par
ogov
iacf
.si
ron
oid
esD
NA
1010
56E
quat
oria
lG
uin
ea1.
4485
8,9.
7808
6D
Q82
5549
DQ
8256
05D
Q82
5624
DQ
8256
50D
Q82
5518
Par
ogov
iacf
.si
ron
oid
esD
NA
1046
19C
amer
oon
2.74
108,
9.88
181
JF93
4967
JF93
5022
JF93
5045
JF78
6409
JF78
6428
Par
ogov
iasp
.D
NA
1010
52E
quat
oria
lG
uin
ea1.
6581
5,10
.311
43D
Q82
5546
DQ
8256
01D
Q82
5620
DQ
8256
48D
Q82
5515
Par
ogov
iasp
.D
NA
1010
57E
quat
oria
lG
uin
ea2.
1311
9,9.
8718
7D
Q82
5547
DQ
8256
02D
Q82
5621
–D
Q82
5516
Par
ogov
iasp
.D
NA
1046
15C
amer
oon
4.80
084,
9.66
326
JF93
4966
JF93
5021
JF93
5044
JF78
6410
JF78
6427
Par
ogov
iasp
.D
NA
1046
18C
amer
oon
2.74
108,
9.88
181
JF93
4968
JF93
5020
JF93
5046
JF78
6412
JF78
6429
Par
ogov
iasp
.D
NA
1056
71Iv
ory
Coa
st5.
8333
3,-7
.350
00*
JF93
4965
JF93
5018
JF93
5048
JF78
6413
–
OU
TG
RO
UP
SP
roto
loph
us
sin
gula
ris
DN
A10
1033
Cal
ifor
nia
,U
SA
EF
0280
95E
F02
8096
EF
1085
81E
F10
8586
EF
1085
92M
egal
opsa
lis
sp.
DN
A10
0783
SI,
New
Zea
lan
dE
F10
8573
EF
1085
76E
F10
8582
EF
1085
87E
F10
8593
Hes
pero
nem
asto
ma
mod
estu
mD
NA
1003
12O
rego
n,
US
AA
F12
4942
EF
1085
77E
F10
8583
EF
1085
88E
F10
8594
Den
dro
lasm
apa
rvu
lum
DN
A10
0318
Japa
nE
F10
8574
EF
1085
78E
F10
8584
EF
1085
89–
Equ
itiu
sd
oria
eD
NA
1006
07A
ust
rali
aU
3700
3E
F10
8579
–E
F10
8590
EF
1085
95S
and
okan
mal
ayan
us
DN
A10
0321
Mal
aysi
aE
F10
8575
EF
1085
80E
F10
8585
EF
1085
91E
F10
8596
Ast
eris
ksin
dica
teap
prox
imat
eco
ordi
nat
es.
10 G. GIRIBET ET AL.
© 2011 The Linnean Society of London, Biological Journal of the Linnean Society, 2011, ••, ••–••

Bivort & Giribet, 2010), as a result of the largernumber of specimens based on literature sources andthe lack of scanning electron micrographs of severalspecies.
When selecting the morphological terminals, weattempted to maximize overlapping with the molecu-lar matrix and also attempted to include the typespecies of each genus, with a few exceptions. Allmonotypic genera were also included, irrespective ofwhether molecular data were available or not. Mono-typic genera not represented by molecular data areAnkaratra Shear & Gruber, 1996, Iberosiro de Bivort& Giribet, 2004, Manangotria Shear & Gruber, 1996,Marwe Shear, 1985, Odontosiro Juberthie, 1961, Spe-leosiro Lawrence, 1931, and Stylocellus. Similarly,Shearogovea mexasca, now not considered as amember of Neogoveidae or Neogovea (Benavides &Giribet, 2007; Giribet, 2011), is not represented bymolecular data but was included in the combinedanalysis.
When a species was represented by multiplemolecular terminals, the morphological data matrixwas replicated so all molecular terminals were repre-sented by the same morphological codings. Thisapplies to the three populations of Metasiro america-nus (Davis, 1933), the two specimens of Parogoviasironoides Hansen, 1921 and four specimens of P. cf.sironoides, the two specimens of Metagovea sp.(DNA104648 and DNA104647), two specimens ofNeogovea virginie, and two specimens of Suzukielussauteri (Roewer, 1916).
The annotated morphological data matrix has beendeposited in Morphobank (morphobank.org) withaccession number P199 (http://morphobank.org/permalink/?P199).
PHYLOGENETIC ANALYSIS: DYNAMIC HOMOLOGY
UNDER PARSIMONY
Parsimony analysis under direct optimization(Wheeler, 1996) used the software POY, version 4.1.2(Varón, Sy Vinh & Wheeler, 2010) on six processors on
a Quad-Core Intel Xeon 3 GHz Mac Pro or on 40processors in the Odyssey cluster at Harvard Univer-sity FAS Research computing facility. Timed searches(multiple Wagner trees followed by SPR + TBR +ratchet and tree fusing) of 6–12 h each were run forthe combined analyses of all molecules under sixanalytical parameter sets (see below). Two additionalrounds of sensitivity analysis tree fusing (SATF)(Giribet, 2007a), taking all input trees from the pre-vious round of analyses, were conducted for the com-bined analysis of molecules under the multipleparameter sets evaluated. These were also 6-h timedsearches, and the results of these were plotted tocheck for stability in the results. Once a parameterset stabilized and the optimal result was found mul-tiple times, we stopped that inquiry but continuedwith additional rounds of searches for those param-eter sets that continued improving or that found theoptimal solution only once. The results of these analy-ses are shown in Table 3.
Because a broad parameter space has already beenexplored in detail in earlier studies (Boyer et al.,2007b), we restricted the dynamic homology analysesto six parameter sets, named 111, 121, 211, 221, 3221,and 3211. Parameter set 3221 (indel opening cost = 3;indel extension cost = 1; transversions = transitions= 2) has been favoured in many analyses and hasbeen justified philosophically as the best way of ana-lyzing data under direct optimization (De Laet, 2010).In addition, we explored a parameter set, named3211, where transversions and transitions receive dif-ferent costs (indel opening cost = 3; indel extensioncost = 1; transversion cost = 2; transition cost = 1),extending the idea of mixed-parameter sets ofSharma et al. (2011). Four other parameter sets 111,121, 211, and 221, optimal in the analyses of Boyeret al. (2007b) and aiming to limit the differencebetween indel costs and transformation costs (Spagna& Álvarez-Padilla, 2008), were explored. To calculatethe WILD (Wheeler, 1995; Sharma et al., 2011) eachindividual partition, or the combination of the twonuclear ribosomal RNA partitions, were run with a
Table 3. Search strategy and tree length stabilization after subsequent rounds of sensitivity analysis tree fusing (TFN)for each parameter set
TF4 TF5 TF6 TF7 TF8 TF9
111 27101 27074 27074 – – –121 41849 41773 41773 – – –211 28975 28944 28940 28940 – –221 45211 45179 45131 45118 45115 451153221 55982 55744 55729 55729 – –3211 43121 42874 42854 42854 – –
111 and 121 stabilized after five rounds of tree fusing; 221 stabilized after eight rounds of tree fusing
EVOLUTIONARY AND BIOGEOGRAPHICAL HISTORY OF ARACHNIDS 11
© 2011 The Linnean Society of London, Biological Journal of the Linnean Society, 2011, ••, ••–••
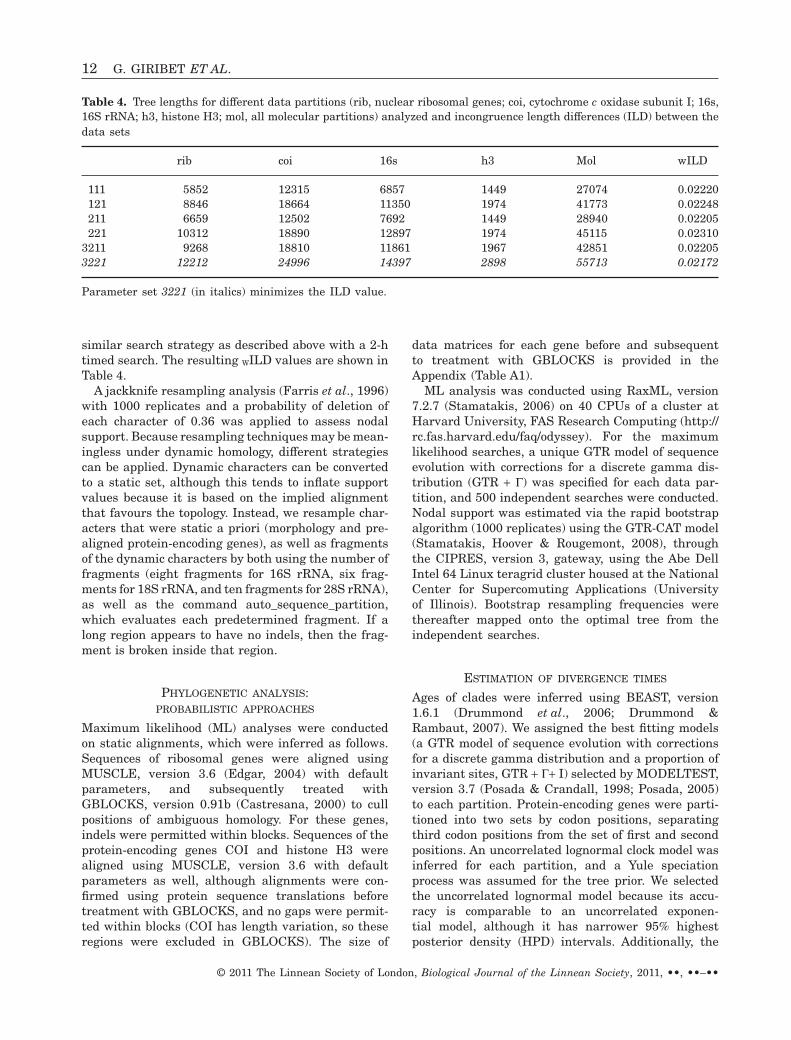
similar search strategy as described above with a 2-htimed search. The resulting WILD values are shown inTable 4.
A jackknife resampling analysis (Farris et al., 1996)with 1000 replicates and a probability of deletion ofeach character of 0.36 was applied to assess nodalsupport. Because resampling techniques may be mean-ingless under dynamic homology, different strategiescan be applied. Dynamic characters can be convertedto a static set, although this tends to inflate supportvalues because it is based on the implied alignmentthat favours the topology. Instead, we resample char-acters that were static a priori (morphology and pre-aligned protein-encoding genes), as well as fragmentsof the dynamic characters by both using the number offragments (eight fragments for 16S rRNA, six frag-ments for 18S rRNA, and ten fragments for 28S rRNA),as well as the command auto_sequence_partition,which evaluates each predetermined fragment. If along region appears to have no indels, then the frag-ment is broken inside that region.
PHYLOGENETIC ANALYSIS:PROBABILISTIC APPROACHES
Maximum likelihood (ML) analyses were conductedon static alignments, which were inferred as follows.Sequences of ribosomal genes were aligned usingMUSCLE, version 3.6 (Edgar, 2004) with defaultparameters, and subsequently treated withGBLOCKS, version 0.91b (Castresana, 2000) to cullpositions of ambiguous homology. For these genes,indels were permitted within blocks. Sequences of theprotein-encoding genes COI and histone H3 werealigned using MUSCLE, version 3.6 with defaultparameters as well, although alignments were con-firmed using protein sequence translations beforetreatment with GBLOCKS, and no gaps were permit-ted within blocks (COI has length variation, so theseregions were excluded in GBLOCKS). The size of
data matrices for each gene before and subsequentto treatment with GBLOCKS is provided in theAppendix (Table A1).
ML analysis was conducted using RaxML, version7.2.7 (Stamatakis, 2006) on 40 CPUs of a cluster atHarvard University, FAS Research Computing (http://rc.fas.harvard.edu/faq/odyssey). For the maximumlikelihood searches, a unique GTR model of sequenceevolution with corrections for a discrete gamma dis-tribution (GTR + G) was specified for each data par-tition, and 500 independent searches were conducted.Nodal support was estimated via the rapid bootstrapalgorithm (1000 replicates) using the GTR-CAT model(Stamatakis, Hoover & Rougemont, 2008), throughthe CIPRES, version 3, gateway, using the Abe DellIntel 64 Linux teragrid cluster housed at the NationalCenter for Supercomuting Applications (Universityof Illinois). Bootstrap resampling frequencies werethereafter mapped onto the optimal tree from theindependent searches.
ESTIMATION OF DIVERGENCE TIMES
Ages of clades were inferred using BEAST, version1.6.1 (Drummond et al., 2006; Drummond &Rambaut, 2007). We assigned the best fitting models(a GTR model of sequence evolution with correctionsfor a discrete gamma distribution and a proportion ofinvariant sites, GTR + G+ I) selected by MODELTEST,version 3.7 (Posada & Crandall, 1998; Posada, 2005)to each partition. Protein-encoding genes were parti-tioned into two sets by codon positions, separatingthird codon positions from the set of first and secondpositions. An uncorrelated lognormal clock model wasinferred for each partition, and a Yule speciationprocess was assumed for the tree prior. We selectedthe uncorrelated lognormal model because its accu-racy is comparable to an uncorrelated exponen-tial model, although it has narrower 95% highestposterior density (HPD) intervals. Additionally, the
Table 4. Tree lengths for different data partitions (rib, nuclear ribosomal genes; coi, cytochrome c oxidase subunit I; 16s,16S rRNA; h3, histone H3; mol, all molecular partitions) analyzed and incongruence length differences (ILD) between thedata sets
rib coi 16s h3 Mol wILD
111 5852 12315 6857 1449 27074 0.02220121 8846 18664 11350 1974 41773 0.02248211 6659 12502 7692 1449 28940 0.02205221 10312 18890 12897 1974 45115 0.02310
3211 9268 18810 11861 1967 42851 0.022053221 12212 24996 14397 2898 55713 0.02172
Parameter set 3221 (in italics) minimizes the ILD value.
12 G. GIRIBET ET AL.
© 2011 The Linnean Society of London, Biological Journal of the Linnean Society, 2011, ••, ••–••

variance of the uncorrelated lognormal model canbetter accommodate data that are already clock-like(Drummond et al., 2006). Priors were sequentiallyoptimized in a series of iterative test runs (data notshown). Markov chains were run for 50 000 000 gen-erations, sampling every 1000 generations. Conver-gence diagnostics were assessed using TRACER,version 1.5 (Rambaut & Drummond, 2007).
Fossil taxa were used to calibrate divergence times.We constrained the age of Eupnoi to 410 Mya usingthe Devonian harvestman Eophalangium sheari[Dunlop et al. 2004 [Dunlop et al., 2003; 2004 (for2003)]; a normal distribution with a standard devia-tion of 5 Myr was applied to this node to account foruncertainty in estimation of fossil age. Dyspnoi wereconstrained using a normal distribution with a meanof 300 Mya and a standard deviation of 10 Myr, on thebasis of the Carboniferous fossils Eotrogulus fayoliThevenin, 1901 and Nemastomoides elaveris Thev-enin, 1901 (Dunlop, 2007).
We explored constraining the family Stylocellidaeusing the Early Cretaceous Burmese amber fossilPalaeosiro burmanicum Poinar, 2008 (Poinar, 2008)1.We used a gamma distribution with shape param-eters (a, b) = (8, 14), and an offset of 105 Myr for thediversification of Stylocellidae; such a prior distribu-tion establishes a floor in the age of stylocellids(105 Mya), at the same time enabling estimates ofdiversification as early as the Late Permian, in accor-dance with previous estimates (Boyer et al., 2007b;Clouse & Giribet, 2010). However, because the inclu-sion of this last constraint did not affect the ageestimate of Stylocellidae, we ultimately did notinclude it in the analysis.
ANCESTRAL AREA RECONSTRUCTION
Likelihood analysis of ancestral area reconstructionwas conducted using the software LAGRANGE (Reeet al., 2005; Ree & Smith, 2008). We divided the datedtree from BEAST analysis into three parts for ana-lytical tractability: (1) the Pettalidae subtree; (2) the(Troglosironidae + Ogoveidae + Neogoveidae) subtree;and (3) the (Sironidae + Stylocellidae) subtree. Foreach subtree, we implemented stratified dispersalconstraint matrices for multiple spans of time for therelevant areas inhabited by the constituent taxa ofeach subtree. Geological events used to delimit the
time spans are sensu Sanmartín & Ronquist (2004)and Hall (2002). The maximum number of areas inancestral ranges was held at two (this conventionreflects empirical observations of Cyphophthalmispecies, the majority of which are narrowly distrib-uted endemics), and dispersal constraints were set to1.0 (if landmasses were connected), 0.1 (if landmasseswere disjunct) or 0 (if landmasses did not exist). Areasand geological intervals for each subtree are indicatedin the Appendix Table A2 (the Python scripts specify-ing dispersal constraint matrices are available uponrequest from the authors).
HABITAT SUITABILITY AND DISTRIBUTION MODELLING
To generate predictions of habitat suitability andpotential lineage distributions, habitat suita-bility models (HSMs) of the major lineages ofCyphophthalmi were reconstructed using the19 bioclimatic variables of Hijmans et al. (2005:http://www.worldclim.org/). These variables provide asummary of the monthly temperature and precipita-tion worldwide. These variables are well documentedand are widely used in studies relaying on niche anddistribution modelling (Evans et al., 2009; Smith &Donoghue, 2010). By contrast to the raw temperatureand precipitation data, they do provide biologicallyrelevant information. We have used all 19 variablesat 10 arc minutes resolution. In addition, analyseswith a subset of the environmental variables repre-senting only the most important variables were per-formed (thus reducing the dimensionality of theanalyses and the risk of over fitting) and the resultsobtained were compared. To evaluate the variablessignificance, we used jackknife (as implemented inMAXENT).
HSMs were built using the maximum entropyalgorithm implemented in MAXENT, version 3.3.3a(Phillips, Anderson & Schapire, 2006; Phillips &Dudik, 2008). Maximum entropy has shown a highperformance score in comparison with other methods(Araujo & Rahbek, 2006) and also allows workingwith fewer data points (Pearson et al., 2007). Thetotal number of unique localities with occurrenceobservations used for the modelling of habitat suit-ability was: Pettalidae, N = 107; Sironidae, N = 60;Stylocellidae, N = 127; and Sternophthalmi, N = 90.To evaluate the performance of the model, cross-validation as implemented in MAXENT (ten repli-cates) was used in all runs.
To test the sensitivity of the results to the model-ling algorithm, we have run the same set of analysisusing the simpler BIOCLIM (Nix, 1986) profilemethod as implemented in openModeller, version1.1.0 (de Souza Muñoz et al., 2011). Climatic enve-lopes’ extent and distribution were modelled world-
1Palaeosiro burmanicum Poinar, 2008 was placed withinSironidae in the original description based on the lack of asternal apophysis with gland pores and dentition on the tarsalclaw of leg II, although these only rule out placement withinTroglosironidae and Neogoveidae. Moreover, the shape andposition of the ozophores, the presence of eyes, the carina ofthe anal plate (similar to some Fangensis Schwendinger &Giribet, 2005), and the collecting locality of the fossil, are allconsistent with an early diverging lineage of Stylocellidae.
EVOLUTIONARY AND BIOGEOGRAPHICAL HISTORY OF ARACHNIDS 13
© 2011 The Linnean Society of London, Biological Journal of the Linnean Society, 2011, ••, ••–••

wide to compare the actual linage distributions withthe distribution of potentially suitable climates.
The software package ENMTools, version 1.3(Warren, Glor & Turelli, 2010) was used to accessclimatic envelopes’ differentiation. ENMTools imple-ments the I, Schoener’s D and relative rank metrics tocompare models predictions (Schoener, 1968; Warren,Glor & Turelli, 2008). The three indices measuresimilarity of predicted habitat suitability distribu-tions and range from 0, indicating no overlap, to 1,indicating complete overlap. In addition, habitat suit-ability score differences were evaluated by comparingthe similarity indices (models overlap) for the modelsbuilt from the actual occurrences of the two species toa null distribution generated by nonparametric resa-mpling. Comparisons were performed using the nicheidentity test (Warren et al., 2008) implemented inENMTools.
RESULTS
Analysis of the combined molecular data matrixunder selected parameter sets for direct optimizationresulted in topologies that agree on several basicaspects of cyphophthalmid phylogeny, includingmonophyly of the suborder, a sister group relationshipof Pettalidae to all other families, and a clade con-taining all members of the families Troglosironidae,Ogoveidae, and Neogoveidae. All parameter setsalso resulted in very similar WILD numbers, with3221 being slightly favoured above all others(WILD = 0.02172; the worst parameter set being 121,with WILD = 0.02248). Stabilization of parameter set3221 occurred after nine rounds of tree fusing. Theoptimal tree, along with the Navajo rugs (Giribet,2003b) for the familial monophyly and relationships,is presented in Figure 3. A clade containing the fami-lies Sironidae and Stylocellidae is also found undermost analytical conditions (Fig. 3).
Monophyly of Pettalidae, Troglosironidae, Stylocel-lidae, and Ogoveoidea (= Ogoveidae + Neogoveidae) issupported under every analyzed parameter set, asare many internal clades within the familiesStylocellidae, Sironidae, and Neogoveidae. However,Sironidae is not monophyletic under any parameterset when combining all data (Sironidae is monophyl-etic when nuclear ribosomal genes are analyzedalone). In this case, a clade containing the generaSiro, Paramiopsalis, and Cyphophthalmus is stableto parameter variation, although Parasiro and
Suzukielus often appear at the base of Stylocellidae,or as sister to a clade including the familiesStylocellidae, Troglosironidae, Ogoveidae, andNeogoveidae (parameter set 3211). The North Ameri-can Siro and the European Siro form reciprocallymonophyletic groups and this clade is sister to Para-miopsalis + Cyphophthalmus. In the case ofNeogoveidae, most parameter sets find Metasiro to bethe sister genus to all other neogoveids but, underparameter sets 111 and 211, Ogovea appears as sisterto the African genus Parogovia, both forming thesister clade to Metasiro. These are the only param-eter sets that find monophyly of the South Americanneogoveids, with Canga as sister genus to all otherSouth American genera. All other parameter setsinstead support monophyly of Neogoveidae, Metasiroas the sister genus to all other species, the Braziliangenus Canga as sister to the African genusParogovia, and the stable relationship of ((Brasil-ogovea, Neogovea) (Huitaca, Metagovea)). Relation-ships of Stylocellidae are well resolved, as:(Fangensis (Meghalaya (Miopsalis, Leptopsalis))).Although all pettalid genera are supported, theirrelationships remain unstable to parameter setvariation, and stable are only the sister group rela-tionships of Purcellia to Chileogovea Roewer, 1961and of Karripurcellia Giribet, 2003 to Pettalus. Twogenera appear as candidate sister groups to all otherpettalids, the South African genus ParapurcelliaRosas Costa, 1950 or the north-eastern Australianendemic Austropurcellia Juberthie, 1988. Jackknifesupport values for the pettalid generic relationshipsare, for the most part, below 50%.
The maximum likelihood analysis resulted in a treetopology with lnL = -103 563.078879. The likelihoodtree topology (Fig. 4) is largely comparable to resultsfrom parsimony analyses but notably recovers amonophyletic Sironidae (i.e. including the generaParasiro and Suzukielus), albeit with low nodalsupport (BS = 44%). As in the direct optimizationoptimal tree, Parapurcellia is sister to all other pet-talid genera, and Purcellia + Chileogovea form a sup-ported clade (66% bootstrap support; BS), whereasKarripurcellia and Pettalus form a clade without sig-nificant nodal support. No other generic relationshipsfind high support. Troglosironidae is sister to Ogoveo-idea (84% BS), and the structure of Neogoveidae isalmost identical to that of the optimal direct optimi-zation tree, with the exception that Brasilogovea ishere monophyletic (the sequences for one terminal
Figure 3. Phylogenetic tree based on the parsimony direct optimization analysis of molecular data under parameter set3221 (55 713 weighted steps). Clade colours correspond to those in Fig. 2. Navajo rugs indicate monophyly (black) ornon-monophyly (white) of a given node under the parameter set specified in the legend. Numbers above nodes indicatejackknife support values.
�
14 G. GIRIBET ET AL.
© 2011 The Linnean Society of London, Biological Journal of the Linnean Society, 2011, ••, ••–••
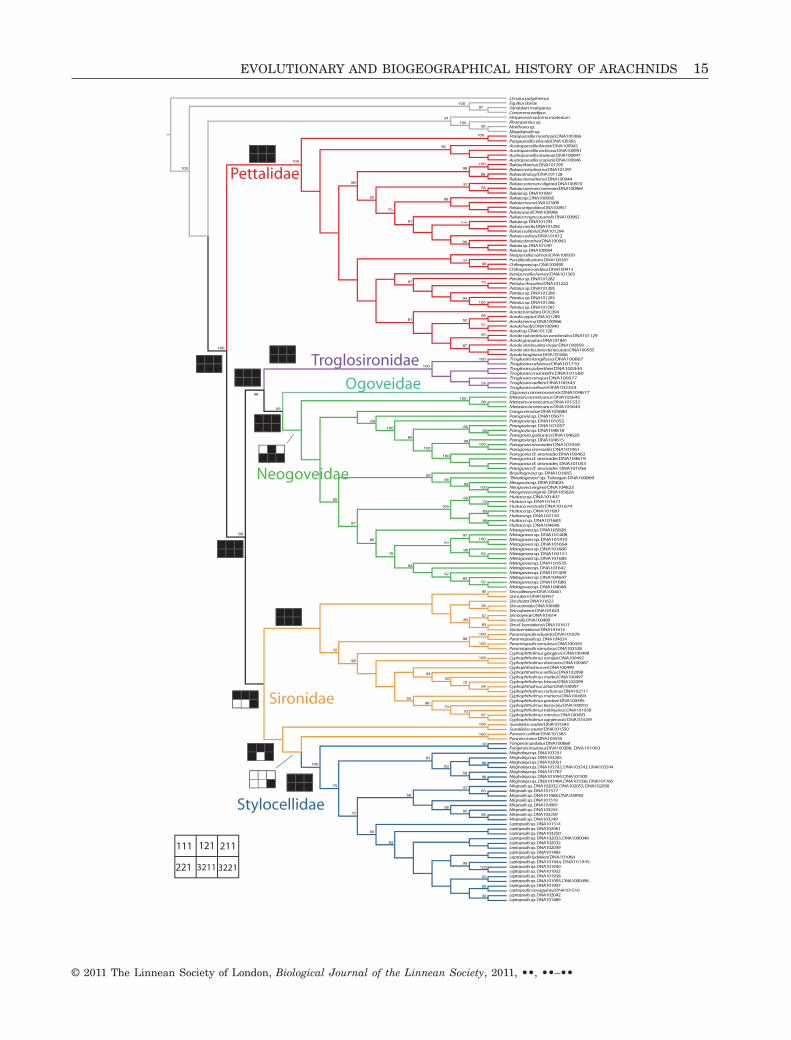
DNA101383
Metagovea sp. DNA101680
Rakaia sp. DNA101807
Rakaia macra DNA101808
Pettalus sp. DNA101286
Rakaia sp. DNA101297
Metagovea sp. DNA101408
Cyphophthalmus ognjenovici DNA101039
Leptopsalis sp. DNA101514
Cyphophthalmus duricorius DNA100487
Cyphophthalmus markoi DNA100497
Austropurcellia scoparia DNA100946
Aoraki sp. DNA101126
Parogovia sp. DNA101057
Metagovea sp. DNA101410
Rakaia stewartiensis DNA100944
Pettalus sp. DNA101282
Miopsalis sp. DNA101519
Miopsalis sp. DNA101517
DNA101295
Rakaia dorothea DNA100943
Cyphophthalmus trebinjanus DNA101038
Meghalaya sp. DNA101767
Pettalus sp. DNA101288
Pettalus sp. DNA101287
Rakaia sp. DNA100954
Aoraki tumidata DOC094
Siro shasta DNA101622
Parapurcellia monticola DNA100386
Meghalaya sp. DNA102051
Conomma oedipus
Marthana sp.
Pettalus sp. DNA101285
Austropurcellia forsteri DNA100945
Aoraki denticulata major DNA100959
Leptopsalis sp. DNA101483
Cyphophthalmus zetae DNA100907
Troglosiro longifossa DNA100867
Leptopsalis sp. DNA103250
Huitaca ventralis DNA101674
Rakaia minutissima DNA101291
Parogovia cf. sironoides DNA104619
Miopsalis sp. DNA102032, DNA102053, DNA102058
Parogovia gabonica DNA104620
Siro rubens DNA100457
Hesperonemastoma modestum
Huitaca sp. DNA101683
Siro calaveras DNA101623
Leptopsalis sp. DNA101489
Rakaia uniloca DNA101812
Suzukielus sauteri DNA101550
Meghalaya sp. DNA103242, DNA103243, DNA103244
Miopsalis sp. DNA103259
Aoraki crypta DNA101289
Metagovea sp. DNA104648
Metasiro americanus DNA101532
Neogovea virginie DNA104823
Pettalus sp. DNA101283
Cyphophthalmus teyrovskyi DNA100910
Metasiro americanus DNA105644
Brasilogovea sp. DNA101665
Leptopsalis sp. DNA101930
Troglosiro ninqua DNA100577
“Brasilogovea” sp. Tobogan DNA100869
Fangensis insulanus DNA100388, DNA101063
Parasiro minor DNA103535
Parogovia sp. DNA104615
Huitaca sp. DNA101681
Leptopsalis sp. DNA102039
Neogovea sp. DNA105825
Metagovea sp. DNA101654
Leptopsalis sp. DNA101938
Huitaca sp. DNA101407
Austropurcellia arcticosa DNA100951
Cyphophthalmus gordani DNA100495
Austropurcellia daviesae DNA100947
Siro boyerae DNA101614
Karripurcellia harveyi DNA101303
Rakaia sorenseni digitata DNA100970
Huitaca sp. DNA104646
Miopsalis sp. DNA103249
Cyphophthalmus rumijae DNA100492
Rakaia media DNA101292
Siro valleorum DNA100461
Troglosiro juberthiei DNA100344
Pettalus thwaitesi DNA101223
Troglosiro aelleni DNA100345
Sandokan malayanus
Miopsalis sp. DNA101468, DNA100950
Meghalaya sp. DNA103251
Troglosiro wilsoni DNA102324
Cyphophthalmus gjorgjevici DNA100498
Aoraki calcarobtusa westlandica DNA101129
Neogovea virginie DNA105824
Leptopsalis sp. DNA101093, DNA1000496
Siro kamiakensis DNA101613
Parogovia cf. sironoides DNA101056
Metagovea sp. DNA101642Metagovea sp. DNA101409
Siro cf. kamiakensis DNA101611
Rakaia antipodiana DNA100957
Paramiopsalis ramulosus DNA103538
Miopsalis sp. DNA103254
Rakaia pauli DNA100968
Parogovia sironoides DNA101059
Paramiopsalis sp. DNA104624
Metagovea sp. DNA101670
Rakaia lindsayi DNA101128
Purcellia illustrans DNA100387
Metagovea sp. DNA105826
Cyphophthalmus minutus DNA100493
Leptopsalis lydekkeri DNA101064
Cyphophthalmus serbicus DNA102098
Leptopsalis sp. DNA102042
Huitaca sp. DNA102150
Siro acaroides DNA100488
Leptopsalis sp. DNA102032
Metagovea sp. DNA101685
Parogovia cf. sironoides DNA101053
Leptopsalis sp. DNA101944, DNA101945
Aoraki longitarsa DNA101806
Parogovia sironoides DNA101061
Paramiopsalis eduardoi DNA101878
Rhampsinitus sp.
Meghalaya sp. DNA101094, DNA101500
Rakaia sp. DNA100958
Parogovia sp. DNA104618
Aoraki granulosa DNA101841
Chileogovea sp. DNA100490
Metagovea sp. DNA104647
Neopurcellia salmoni DNA100939
Canga renatae DNA105680
Limulus polyphemus
Paramiopsalis ramulosus DNA100459
Aoraki healyi DNA100940
Siro exilis DNA100489
Metagovea sp. DNA101686
Leptopsalis sp. DNA101932
Metasiro americanus DNA105645
Cyphophthalmus ere DNA100499
Parogovia cf. sironoides DNA100462
Fangensis spelaeus DNA100669
Meghalaya sp. DNA103265
Parogovia sp. DNA101052
Rakaia sorenseni sorenseni DNA100969
Rakaia magna australis DNA100962
Suzukielus sauteri DNA101543
Chileogovea oedipus DNA100413
Troglosiro monteithi DNA101580
Ogovea cameroonensis DNA104617
Miopsalis sp. DNA104981
Leptopsalis sp. DNA102033, DNA1000048
Rakaia solitaria DNA101294
Metagovea sp. DNA102151
Cyphophthalmus hlavaci DNA102099
Cyphophthalmus martensi DNA100494
Leptopsalis novaguinea DNA101510
Megalopsalis sp.
Parogovia sp. DNA105671
Aoraki denticulata denticulata DNA100955
Huitaca sp. DNA101671
Rakaia sp. DNA101293
Equitius doriae
Troglosiro urbanus DNA101710
Meghalaya sp. DNA101494, DNA101506, DNA101765
Leptopsalis sp. DNA102061
Cyphophthalmus corfuanus DNA102111
Leptopsalis sp. DNA101937
Aoraki inerma DNA100966
Parapurcellia silvicola DNA100385
93
98
92
97
87
98
82
100
100
100
55
52
83
57
60
80
97
98
100
94
92
76
83
64
97
99
65
81
100
73
82
52
99
98
99
73
58
88
100
88
96
59
51
100
100
92
80
64
52
56
94
70
83
56
99
100
81
72
96
81
56
69
57
87
88
72
72
64
52
100
69
99
52
58
75
99
67
83
99
100
73
85
54
100
98
68
87
59
100
69
91
55
98
83
100
91
70
99
97
32
100
100
100
100
99
99
82
Troglosironidae
Ogoveidae
Pettalidae100
Neogoveidae
Sironidae
Stylocellidae
100
100
100
100
99
99
100
100
100
111 121 211
221 3211 3221
EVOLUTIONARY AND BIOGEOGRAPHICAL HISTORY OF ARACHNIDS 15
© 2011 The Linnean Society of London, Biological Journal of the Linnean Society, 2011, ••, ••–••

Pettalus sp. DNA101282
Pettalus sp. DNA101287Pettalus sp. DNA101286
Pettalus sp. DNA101283Pettalus sp. DNA101288
Pettalus sp. DNA101285Pettalus thwaitesi DNA101223
Aoraki sp. DNA101126Aoraki healyi DNA100940
Aoraki inerma DNA100966Aoraki crypta DNA101289
Aoraki calcarobtusa westlandica DNA101129 Aoraki granulosa DNA101841
Aoraki tumidata DOC094Aoraki denticulata major DNA100959
Aoraki denticulata denticulata DNA100955Aoraki longitarsa DNA101806
Austropurcellia daviesae DNA100947Austropurcellia scoparia DNA100946
Austropurcellia arcticosa DNA100951Austropurcellia forsteri DNA100945
Parapurcellia monticola DNA100386Parapurcellia silvicola DNA100385
Purcellia illustrans DNA100387
Chileogovea sp. DNA100490Chileogovea oedipus DNA100413
Karripurcellia harveyi DNA101303
Neopurcellia salmoni DNA100939
Rakaia sp. DNA101293
Rakaia media DNA101292Rakaia solitaria DNA101294
Rakaia magna australis DNA100962
Rakaia sp. DNA100954Rakaia sp. DNA101297Rakaia dorothea DNA100943
Rakaia uniloca DNA101812
Rakaia pauli DNA100968Rakaia antipodiana DNA100957
Rakaia macra DNA101808Rakaia sp. DNA100958
Rakaia sp. DNA101807Rakaia sorenseni sorenseni DNA100969
Rakaia sorenseni digitata DNA100970
Rakaia stewartiensis DNA100944Rakaia lindsayi DNA101128
Rakaia minutissima DNA101291DNA101295
Troglosiro wilsoni DNA102324
Troglosiro aelleni DNA100345Troglosiro ninqua DNA100577
Troglosiro monteithi DNA101580
Troglosiro juberthiei DNA100344
Troglosiro urbanus DNA101710Troglosiro longifossa DNA100867
Ogovea cameroonensis DNA104617
Parogovia sp. DNA105671
Metasiro americanus DNA105645
Metasiro americanus DNA101532Metasiro americanus DNA105644
Canga renatae DNA105680
Parogovia sp. DNA101057Parogovia sp. DNA101052
Parogovia sp. DNA104618Parogovia gabonica DNA104620
Parogovia sp. DNA104615Parogovia sironoides DNA101059
Parogovia sironoides DNA101061
Parogovia cf. sironoides DNA101053
Parogovia cf. sironoides DNA100462Parogovia cf. sironoides DNA101056
Parogovia cf. sironoides DNA104619
Neogovea virginie DNA104823Neogovea virginie DNA105824
Neogovea sp. DNA105825
“Brasilogovea” sp. Tobogan DNA100869Brasilogovea sp. DNA101665
Neogovea sp. DNA101686
Metagovea sp. DNA104648Metagovea sp. DNA104647
Neogovea sp. DNA101409
Metagovea sp. DNA101642Metagovea sp. DNA101670
Neogovea sp. DNA105826
Metagovea sp. Valle DNA101685Metagovea sp. DNA102151m
Metagovea sp. DNA101680
Metagovea sp. DNA101654Metagovea sp. DNA101410
Metagovea sp. DNA101408
Huitaca ventralis DNA101674Huitaca sp. DNA101407
Huitaca sp. DNA101671
Huitaca sp. DNA101683Huitaca sp. DNA104646
Huitaca sp. DNA102150Huitaca sp. DNA101681
Suzukielus sauteri DNA101550Suzukielus sauteri DNA101543
Parasiro minor DNA103535 DNA101383
Cyphophthalmus ognjenovici DNA101039Cyphophthalmus minutus DNA100493
Cyphophthalmus trebinjanus DNA101038Cyphophthalmus teyrovskyi DNA100910
Cyphophthalmus gordani DNA100495Cyphophthalmus martensi DNA100494
Cyphophthalmus zetae DNA100907
Cyphophthalmus corfuanus DNA102111Cyphophthalmus hlavaci DNA102099
Cyphophthalmus markoi DNA100497
Cyphophthalmus serbicus DNA102098
Cyphophthalmus ere DNA100499
Cyphophthalmus rumijae DNA100492Cyphophthalmus duricorius DNA100487
Cyphophthalmus gjorgjevici DNA100498
Paramiopsalis ramulosus DNA103538Paramiopsalis ramulosus DNA100459
Paramiopsalis sp. DNA104624Paramiopsalis eduardoi DNA101878
Siro shasta DNA101622Siro kamiakensis DNA101613
Siro cf. kamiakensis DNA101611
Siro exilis DNA100489Siro boyerae DNA101614
Siro calaveras DNA101623Siro acaroides DNA100488
Siro rubens DNA100457Siro valleorum DNA100461
Leptopsalis sp. DNA101932Leptopsalis sp. DNA101930
Leptopsalis sp. DNA101944, DNA101945
Leptopsalis lydekkeri DNA101064
Leptopsalis novaguinea DNA101510Leptopsalis sp. DNA101937
Leptopsalis sp. DNA102042
Leptopsalis sp. DNA102039
Leptopsalis sp. DNA102032
Leptopsalis sp. DNA102061Leptopsalis sp. DNA103250
Leptopsalis sp. DNA102033, DNA102048
Leptopsalis sp. DNA101093, DNA101496Leptopsalis sp. DNA101938
Leptopsalis sp. DNA101489Leptopsalis sp. DNA101483
Leptopsalis sp. DNA101514
Miopsalis sp. DNA101517
Miopsalis sp. DNA101468, DNA101950Miopsalis sp. DNA102032, DNA102053, DNA102058
Miopsalis sp. DNA101519
Miopsalis sp. DNA103249 Miopsalis sp. DNA103259
Miopsalis sp. DNA103254Stylocellidae gen. sp. DNA104981
Meghalaya sp. DNA101094, DNA101500Meghalaya sp. DNA101494, DNA101506, DNA101765
Meghalaya sp. DNA101767
Meghalaya sp. DNA103265
Meghalaya sp. DNA102051Meghalaya sp. DNA103242, DNA103243, DNA103244
Meghalaya sp. DNA103251Fangensis insulanus DNA100388
Fangensis spelaeus DNA100669
Equitius doriae DNA100607
Megalopsalis sp. DNA100783Marthana sp. DNA100613
Sandokan malayanus DNA100321Conomma oedipus DNA101051
Hesperonemastoma modestum DNA100312
Rhampsinitus sp. DNA100710
* ** *
85
*
91
72
0.1
**
66
97
**
*
*9985
60
85
98
68
50 67
90
55 94
*
97
* **
*
9870
*69
65 *86
*
*83
53** 92
84
81
8395
*99
*99 58
*98
88 92
76
** ***
*
**66
*90
75
99
** 93
* 94
* 6277
**99
97 71
69
* 9774
99
*
*99
8098
*87
*
*
7161
58
9550
*
**
* *
9896
53
92
92* *
*80 89
97
**
99
87
70
Pettalidae
Trogosironidae
Sironidae
Neogoveidae
Stylocellidae
Ogoveidae
I.O. Sternophthalmi
I.O. Boreophthalmi
I.O. Scopulophthalmi
Figure 4. Single most likely tree (lnL = -103 563.078879) for the combined molecular data set aligned using MUSCLEand subsequently trimmed with GBLOCKS and analyzed in RAxML under GTR + G. Clade colours correspond to thosein Fig. 2. Bootstrap support values are represented above each node; asterisks indicate 100% bootstrap value.
16 G. GIRIBET ET AL.
© 2011 The Linnean Society of London, Biological Journal of the Linnean Society, 2011, ••, ••–••

are based on a juvenile specimen, so the assignmentto this genus is tentative). Within Sironidae, Parasirois sister to all other genera, followed by Suzukielus,although bootstrap support for these basal nodes islow, as is the clade including the remaining sironidgenera. In this case, there is also reciprocal mono-phyly of the European and North American Siro, andthese form the sister group of Paramiopsalis +Cyphophthalmus. Structure of the genera within Sty-locellidae matches that of the direct optimizationanalyses.
The run of BEAST reached stationarity after10 000 000 generations; 20 000 000 generations werediscarded as burn-in. The tree topology recovered byBEAST (Fig. 5) is almost identical to that of theparsimony analysis under the parameter set 3221,with Parasiro and Suzukielus forming a paraphyleticgrade at the base of Stylocellidae, although posteriorprobabilities for the corresponding nodes are low(0.788 and 0.880, respectively). In all other aspects, italso resembles the topology of the maximum likeli-hood analysis, especially in the internal relationshipsamong the pettalid genera.
The diversification of Cyphophthalmi is estimatedat approximately 332 Mya (95% HPD: 297–362 Mya).Diversification times for the described familiesof Cyphophthalmi are estimated as: Neogoveidae,236 Mya (95% HPD: 208–266 Mya); Pettalidae,183 Mya (95% HPD: 148–218 Mya); Sironidae(excluding Parasiro and Suzukielus), 278 Mya (95%HPD: 243–311 Mya); Stylocellidae, 167 Mya (95%HPD: 140–195 Mya); and Troglosironidae, 57 Mya(95% HPD: 40–73 Mya). Ogoveidae, represented by asingle exemplar, diverged from Neogoveidae 261 Mya(95% HPD: 231–292 Mya), and Troglosironidaediverged from Ogoveoidea 279 Mya (95% HPD: 248–311 Mya). These results largely corroborate previousestimates of divergence times (Boyer et al., 2007b;Giribet et al., 2010), with the exception of Stylocel-lidae, the diversification of which is recovered asyounger than previously reported (Clouse & Giribet,2010). Although some species represented by multipleterminals are young (e.g. Suzukielus sauteri,Neogovea virginie, Parogovia sironoides), Metasiroamericanus is an old species, perhaps reflecting theexistence of cryptic species along its range.
All probabilistic approaches recognize a clade oftrans-Tasman Cyphophthalmi (the Australian andNew Zealand genera), although none of these land-masses or their constituent terranes appears mono-phyletic (Fig. 6). The ancestral area reconstruction ofthis clade is ambiguous, with the highest probabilityfor an origin in the Australian plate of New Zealand.The ancestral area of the family shows more ambigu-ity, the three most likely scenarios being a mixedSouth Africa/New Zealand Australian plate
(P = 0.291), South African (P = 0.207) or mixed SouthAfrica/Sri Lankan (P = 0.194). The ancestral areareconstruction of the clade including the three fami-lies with sternal opisthosomal gland openings (Fig. 7)is mostly West African/New Caledonian (P = 0.787),with the ogoveoid families being most likely WestAfrican (P = 0.662) or mixed between West Africa andthe south-eastern USA (P = 0.206), the latter beingonce connected to West Africa. A South American(Amazonian) origin of the family Neogoveidaereceives little support.
COMBINED ANALYSIS OF MOLECULES
AND MORPHOLOGY
The position of morphology-only taxa was unstable inthe first rounds of analyses, which (for example) didnot group the two Ogovea species, one represented bymorphology only, whereas the other one was repre-sented by molecules and morphology, despite beingalmost identical for the morphological data matrix.This appears to be a problem of the Wagner addition,as designed in most phylogenetic software, and wasresolved by fusing a jackknife tree and a first treeobtained during a normal search, as described above.The resulting trees of each subsequent analysis werethen fused to the previous pool of trees until results(topology and tree length) stabilized. The combinedanalysis of molecules and morphology in POYrequired eight rounds of tree fusing until stabilizingin a tree length of 56 984 weighted steps and findingthree trees differing only in the position of some of themorphology-only taxa (Fig. 9).
The overall topology is very similar to those of theanalyses with molecular data only, with a few excep-tions, and lowered jackknife support values. Pettal-idae is monophyletic (63%), and includes bothSpeleosiro and Manangotria from the morphology-only taxa. Speleosiro appears as sister to Purcelliaand Managotria is sister to Karripurcellia, althoughthese relationships, as with most other intergenericpettalid relationships, receive low support. Ankaratradoes not appear within Pettalidae and, instead,is basal to the clade containing Sironidae andStylocellidae.
Troglosironidae appears as sister to Ogoveoidea,although this tree differs from all previous trees inthat Neogoveidae is paraphyletic with respect toOgovea, which is sister to Parogovia, constituting anAfrican clade, sister to all the American species, withCanga as the sister group to Metasiro, and thisclade being sister to the remaining neogoveids [56%jackknife frequency (JF)]. The type species andmorphology-only species of Neogovea and Brasil-ogovea appear in a clade, although there is littlecorrespondence between the complete taxa and those
EVOLUTIONARY AND BIOGEOGRAPHICAL HISTORY OF ARACHNIDS 17
© 2011 The Linnean Society of London, Biological Journal of the Linnean Society, 2011, ••, ••–••

Pettalidae
Troglosironidae
Sironidae
Neogoveidae
Stylocellidae
Ogoveidae
Pettalus sp. DNA101282
Pettalus sp. DNA101287Pettalus sp. DNA101286Pettalus sp. DNA101283Pettalus sp. DNA101288 Pettalus sp. DNA101285Pettalus thwaitesi DNA101223
Aoraki sp. DNA101126Aoraki healyi DNA100940Aoraki inerma DNA100966Aoraki crypta DNA101289Aoraki calcarobtusa westlandica DNA101129 Aoraki granulosa DNA101841 Aoraki tumidata DOC094
Aoraki denticulata major DNA100959 Aoraki denticulata denticulata DNA100955
Aoraki longitarsa DNA101806
Austropurcellia daviesae DNA100947Austropurcellia scoparia DNA100946
Austropurcellia arcticosa DNA100951Austropurcellia forsteri DNA100945
Parapurcellia monticola DNA100386Parapurcellia silvicola DNA100385Purcellia illustrans DNA100387
Chileogovea sp. DNA100490Chileogovea oedipus DNA100413
Karripurcellia harveyi DNA101303Neopurcellia salmoni DNA100939
Rakaia sp. DNA101293Rakaia media DNA101292Rakaia solitaria DNA101294
Rakaia magna australis DNA100962
Rakaia sp. DNA100954Rakaia sp. DNA101297
Rakaia dorothea DNA100943
Rakaia uniloca DNA101812
Rakaia pauli DNA100968
Rakaia antipodiana DNA100957
Rakaia macra DNA101808Rakaia sp. DNA100958
Rakaia sp. DNA101807Rakaia sorenseni sorenseni DNA100969
Rakaia sorenseni digitata DNA100970Rakaia stewartiensis DNA100944Rakaia lindsayi DNA101128
Rakaia minutissima DNA101291Rakaia florensis DNA101295
Troglosiro wilsoni DNA102324
Troglosiro aelleni DNA100345Troglosiro ninqua DNA100577
Troglosiro monteithi DNA101580 Troglosiro juberthiei DNA100344 Troglosiro urbanus DNA101710Troglosiro longifossa DNA100867 Ogovea cameroonensis DNA104617
Parogovia sp. DNA105671
Metasiro americanus DNA105645Metasiro americanus DNA101532Metasiro americanus DNA105644Canga renatae DNA105680
Parogovia prietoi DNA101057Parogovia sp. DNA101052
Parogovia sp. Campo2 DNA104618Parogovia gabonica DNA104620Parogovia sp. Koupe DNA104615
Parogovia sironoides DNA101059Parogovia sironoides DNA101061
Parogovia cf. sironoides DNA101053Parogovia cf. sironoides DNA100462
Parogovia cf. sironoides DNA101056Parogovia cf. sironoides DNA104619
Metagovea sp. Valle DNA101686
Metagovea sp. DNA104648Metagovea sp. DNA104647Metagovea sp. Nambi DNA101409
Metagovea sp. Pasto DNA101642Metagovea sp. Huila DNA101670
Metagovea sp. Guyana DNA105826
Metagovea sp. Valle DNA101685Metagovea sp. Tatama DNA102151m
Metagovea sp. Caldas DNA101680
Metagovea sp. Planada DNA101654Metagovea sp. Nambi DNA101410
Metagovea sp. Leticia DNA101408
Huitaca ventralis DNA101674Huitaca boyacaensis DNA101407
Huitaca tama DNA101671
Huitaca bitaco DNA101683Huitaca sharkeyi DNA104646Huitaca depressa DNA102150Huitaca caldas DNA101681
Neogovea virginie DNA104823Neogovea virginie DNA105824
Neogovea sp. Guyana DNA105825Brasilogovea sp. Tobogan DNA100869Brasilogovea chiribiqueta DNA101665
Suzukielus sauteri DNA101550Suzukielus sauteri DNA101543
Parasiro minor DNA103535Parasiro coiffaiti DNA101383
Cyphophthalmus ognjenovici DNA101039Cyphophthalmus minutus DNA100493Cyphophthalmus trebinjanus DNA101038Cyphophthalmus teyrovskyi DNA100910Cyphophthalmus gordani DNA100495Cyphophthalmus martensi DNA100494
Cyphophthalmus zetae DNA100907
Cyphophthalmus corfuanus DNA102111Cyphophthalmus hlavaci DNA102099
Cyphophthalmus markoi DNA100497
Cyphophthalmus serbicus DNA102098
Cyphophthalmus ere DNA100499
Cyphophthalmus rumijae DNA100492Cyphophthalmus duricorius DNA100487
Cyphophthalmus gjorgjevici DNA100498
Paramiopsalis ramulosus DNA103538Paramiopsalis ramulosus DNA100459Paramiopsalis sp. DNA104624Paramiopsalis eduardoi DNA101878
Siro shasta DNA101622Siro kamiakensis DNA101613Siro cf. kamiakensis DNA101611Siro exilis DNA100489Siro boyerae DNA101614
Siro calaveras DNA101623Siro acaroides DNA100488
Siro rubens DNA100457Siro valleorum DNA100461
Leptopsalis sp. DNA101932Leptopsalis sp. DNA101930
Leptopsalis sp. DNA101944, DNA101945Leptopsalis lydekkeri DNA101064
Leptopsalis novaguinea DNA101510Leptopsalis sp. DNA101937
Leptopsalis sp. DNA102042Leptopsalis sp. DNA102039Leptopsalis sp. DNA102032
Leptopsalis sp. DNA102061Leptopsalis sp. DNA103250
Leptopsalis sp. DNA102033, DNA102048
Leptopsalis sp. DNA101093, DNA100496Leptopsalis sp. DNA101938
Leptopsalis sp. DNA101489Leptopsalis sp. DNA101483
Leptopsalis sp. DNA101514
Miopsalis sp. DNA101517
Miopsalis sp. DNA101468, DNA101950
Miopsalis sp. DNA102032, DNA102053, DNA102058Miopsalis sp. DNA101519
Miopsalis sp. DNA103249 Miopsalis sp. DNA103259
Miopsalis sp. DNA103254Miopsalis sp. DNA104981
Meghalaya sp. DNA101094, DNA101500Meghalaya sp. DNA101494, DNA101506, DNA101765Meghalaya sp. DNA101767
Meghalaya sp. DNA103265Meghalaya sp. DNA102051Meghalaya sp. DNA103242, DNA103243, DNA103244
Meghalaya sp. DNA103251
Fangensis insulanus DNA100388, DNA101063Fangensis spelaeus DNA100669
65
CARBONIFEROUS PERMIAN TRIASSIC JURASSIC CRETACEOUS TERTIARY
144206248290354
*
*
*
*
**
*
0.983
0.929
0.792
0.871 *
Figure 5. Evolutionary time-tree of Cyphophthalmi inferred from BEAST analysis of all molecular data. Clade colourscorrespond to those in Fig. 2. Coloured bars indicate 95% highest posterior density (HPD) intervals for nodes of interest.Number on nodes indicate posterior probabilities; asterisks indicate posterior probability of 1.00.
18 G. GIRIBET ET AL.
© 2011 The Linnean Society of London, Biological Journal of the Linnean Society, 2011, ••, ••–••

represented by morphology only (i.e. Neogovea andBrasilogovea are not reciprocally monophyletic).
Ankaratra and Shearogovea form a grade at thebase of the Sironidae – Stylocellidae clade, withSironidae paraphyletic, as in the prior POY andBEAST analyses. Marwe and Iberosiro form a cladewith Paramiopsalis, and Odontosiro forms a cladewith Parasiro. Stylocellidae is monophyletic (63% JF),including the morphology-only species Stylocellussumatranus Westwood, 1874, Meghalaya annandaleiGiribet, Sharma & Bastawade, 2007, Miopsalis puli-caria Thorell, 1890, and Leptopsalis beccarii Thorell,1882–1883. Stylocellus sumatranus, the type speciesof Stylocellus, appears nested within the molecularMeghalaya clade; Meghalaya annandalei, the typespecies of Meghalaya, appears unresolved at the baseof the molecular Leptopsalis clade; Miopsalis puli-caria and Leptopsalis beccarii, the type species oftheir respective genera, appear nested deep within
the clade Leptopsalis. Although the stylocellid resultsmake little sense, this may be a result of the lack ofdiscrete characters useful for resolving their phyloge-netic relationships (see below).
A NEW CLASSIFICATION SYSTEM FOR
CYPHOPHTHALMI
Based on the results reported above, we providea new classification system for Cyphophthalmi,introducing three new infraorders: Scopulophthalminew clade, Sternophthalmi new clade, and Boreo-phthalmi new clade (Table 6). Scopulophthalmiis diagnosed as Pettalidae, and the name refers tothe presence of a scopula in the anal region ofthe male in many pettalid species. Sternophthalmiincludes the families Troglosironidae, Ogoveidae, andNeogoveidae, with its etymology referring to the pres-ence of an exocrine gland opening in the opisthosomal
Pettalus sp. DNA101282
Pettalus sp. DNA101287Pettalus sp. DNA101286Pettalus sp. DNA101283Pettalus sp. DNA101288 Pettalus sp. DNA101285Pettalus thwaitesi DNA101223
Aoraki sp. DNA101126Aoraki healyi DNA100940Aoraki inerma DNA100966Aoraki crypta DNA101289Aoraki calcarobtusa westlandica DNA101129 Aoraki granulosa DNA101841 Aoraki tumidata DOC094
Aoraki denticulata major DNA100959 Aoraki denticulata denticulata DNA100955
Aoraki longitarsa DNA101806
Austropurcellia daviesae DNA100947Austropurcellia scoparia DNA100946
Austropurcellia arcticosa DNA100951Austropurcellia forsteri DNA100945
Parapurcellia monticola DNA100386Parapurcellia silvicola DNA100385Purcellia illustrans DNA100387
Chileogovea sp. DNA100490Chileogovea oedipus DNA100413
Karripurcellia harveyi DNA101303Neopurcellia salmoni DNA100939
Rakaia sp. DNA101293Rakaia media DNA101292Rakaia solitaria DNA101294
Rakaia magna australis DNA100962
Rakaia sp. DNA100954Rakaia sp. DNA101297
Rakaia dorothea DNA100943
Rakaia uniloca DNA101812
Rakaia pauli DNA100968
Rakaia antipodiana DNA100957
Rakaia macra DNA101808Rakaia sp. DNA100958
Rakaia sp. DNA101807Rakaia sorenseni sorenseni DNA100969
Rakaia sorenseni digitata DNA100970Rakaia stewartiensis DNA100944Rakaia lindsayi DNA101128
Rakaia minutissima DNA101291Rakaia florensis DNA101295
65
JURASSIC CRETACEOUS TERTIARY
144206
pr = 0.938
pr = 0.898
pr = 0.050
pr = 0.609
pr = 0.154pr = 0.054
pr = 0.871
pr = 0.053
pr = 0.909
pr = 0.056
pr = 0.679pr = 0.140pr = 0.106
pr = 0.601pr = 0.151pr = 0.078
pr = 0.565
pr = 0.310pr = 0.108
pr = 0.720
pr = 0.152pr = 0.119
South Africa
Chile
Eastern Australia
Western Australia
New Zealand, Australian plate
New Zealand, Pacific plate
Sri Lanka
pr = 0.536pr = 0.168pr = 0.143
pr = 0.637pr = 0.187
pr = 0.443pr = 0.167pr = 0.137pr = 0.051
pr = 0.355
pr = 0.236pr = 0.112pr = 0.061
pr = 0.291
pr = 0.194pr = 0.207
pr = 0.098pr = 0.088
Figure 6. Ancestral range reconstructions for Pettalidae inferred by Lagrange analysis, using stratified models. Colouredsquares at terminals indicate ranges occupied by sampled species. Coloured squares on nodes indicate ranges recon-structed for hypothetical ancestors. Numbers on nodes indicate relative probability of ranges reconstructed.
EVOLUTIONARY AND BIOGEOGRAPHICAL HISTORY OF ARACHNIDS 19
© 2011 The Linnean Society of London, Biological Journal of the Linnean Society, 2011, ••, ••–••

sternal region of males in all troglosironids, allogoveids, and most neogoveids, as opposed to theother three families where the opisthosomal exocrineglands, when present, open in the posterior tergites.We maintain Shear’s superfamily Ogoveoidea, andrestrict Sironoidea to the family Sironidae and Stylo-celloidea to the family Stylocellidae, although we donot introduce other superfamilies because they wouldeach contain a single family. Boreophthalmi includesthe families Stylocellidae and Sironidae, which sub-sequent to Hansen & Sørensen’s (1904), had beenconsidered the representatives of the two maincyphophthalmid clades (Shear, 1980). The term refersto the mostly northern hemisphere distribution ofthese two families, although the origin of Stylocel-lidae can be probably traced to northern Gondwana(Clouse & Giribet, 2010). Sternophthalmi is sistergroup to Boreophthalmi.
The following taxa are thus abandoned as a resultof being non-monophyletic according to our phyloge-netic results: Infraorder Tropicophthalmi Shear, 1980and Infraorder Temperophthalmi Shear, 1980.Shear’s infraorders do not reflect the phylogeneticrelationships obtained here, as suggested in previousstudies (Giribet & Boyer, 2002; Boyer et al., 2007b;Giribet et al., 2010).
DISTRIBUTION MODELLING AND HABITAT
SUITABILITY OVERLAP
Habitat suitability models predicted by both theMAXENT and BIOCLIM methods were highly con-gruent and therefore we present only results fromMAXENT (Fig. 10) because it was found to outper-form other modelling algorithms (Elith et al., 2006).Model predictions were significantly distinct fromrandom and area under the curve (AUC) values werehigh or moderately high (in the range 0.84–0.99) inall runs independently of the modelling algorithm andthe set of variables used to build the model. For theanalyses with a reduced number of variables, we keptall variables that had jackknife regularized traininggain greater than one. As expected, when correlationamong variables is present, using a lower number ofvariables does not change significantly the AUCvalues but reduces over-fitting; hence, the resultingmodels find broader areas with suitable conditions.These are, however, congruent with results frommodels built with all BIOCLIM variables and differ-ences are generally associated with areas wherehabitat suitability is low (Fig. 10).
The variables with highest average relative contri-bution to the MAXENT habitat suitability model for
Troglosiro wilsoni DNA102324
Troglosiro aelleni DNA100345Troglosiro ninqua DNA100577
Troglosiro monteithi DNA101580 Troglosiro juberthiei DNA100344 Troglosiro urbanus DNA101710Troglosiro longifossa DNA100867 Ogovea cameroonensis DNA104617
Parogovia sp. DNA105671
Metasiro americanus DNA105645Metasiro americanus DNA101532Metasiro americanus DNA105644Canga renatae DNA105680
Parogovia sp. DNA101057Parogovia sp. DNA101052
Parogovia sp. DNA104618Parogovia gabonica DNA104620Parogovia sp. DNA104615
Parogovia sironoides DNA101059Parogovia sironoides DNA101061
Parogovia cf. sironoides DNA101053Parogovia cf. sironoides DNA100462
Parogovia cf. sironoides DNA101056Parogovia cf. sironoides DNA104619
Metagovea sp. DNA101686
Metagovea sp. DNA104648Metagovea sp. DNA104647Metagovea sp. DNA101409
Metagovea sp. DNA101642Metagovea sp. DNA101670
Metagovea sp. DNA105826
Metagovea sp. DNA101685Metagovea sp. DNA102151
Metagovea sp. DNA101680
Metagovea sp. DNA101654Metagovea sp. DNA101410
Metagovea sp. DNA101408
Huitaca ventralis DNA101674Huitaca sp. DNA101407
Huitaca sp. DNA101671
Huitaca sp. DNA101683Huitaca sp. DNA104646Huitaca sp. DNA102150Huitaca sp. DNA101681
Neogovea virginie DNA104823Neogovea virginie DNA105824
Neogovea sp. DNA105825“Brasilogovea” sp. Tobogan DNA100869Brasilogovea sp. DNA101665
65
PERMIAN TRIASSIC JURASSIC CRETACEOUS TERTIARY
144206248290
New Caledonia
West Africa
SE USA
Amazonia
pr = 0.993pr = 0.969
pr = 1.000
pr = 1.000
pr = 1.000
pr = 1.000
pr = 1.000
pr = 0.832pr = 0.152
pr = 0.986
pr = 0.696pr = 0.148pr = 0.088
pr = 0.206pr = 0.662
pr = 0.077
pr = 0.996
pr = 0.787pr = 0.109pr = 0.058
Figure 7. Ancestral range reconstructions for Sternophthalmi (Troglosironidae, Ogoveidae, Neogoveidae) inferred byLagrange analysis, using stratified models. Coloured squares at terminals indicate ranges occupied by sampled species.Coloured squares on nodes indicate ranges reconstructed for hypothetical ancestors. Numbers on nodes indicate relativeprobability of ranges reconstructed.
20 G. GIRIBET ET AL.
© 2011 The Linnean Society of London, Biological Journal of the Linnean Society, 2011, ••, ••–••

Pettalidae were isothermality (33.8%), precipitationof the driest month (22.8%) and annual mean tem-perature (9.2%). Jackknife tests of variable impor-tance indicate that temperature seasonality had thehighest gain in isolation. Mean diurnal temperaturerange decreased the gain the most when omitted,suggesting that it contained the most information not
present in the other variables. For Sironidae, theprecipitation of the coldest quarter (40.0%), meantemperature of the coldest quarter (16.3%), andannual mean temperature were the variables withhighest contribution. Mean temperature of thecoldest quarter had the highest gain in isolation, andannual precipitation decreased the gain the most
Suzukielus sauteri DNA101550Suzukielus sauteri DNA101543
Parasiro minor DNA103535Parasiro coiffaiti DNA101383
Cyphophthalmus ognjenovici DNA101039Cyphophthalmus minutus DNA100493Cyphophthalmus trebinjanus DNA101038Cyphophthalmus teyrovskyi DNA100910Cyphophthalmus gordani DNA100495Cyphophthalmus martensi DNA100494
Cyphophthalmus zetae DNA100907
Cyphophthalmus corfuanus DNA102111Cyphophthalmus hlavaci DNA102099
Cyphophthalmus markoi DNA100497
Cyphophthalmus serbicus DNA102098
Cyphophthalmus ere DNA100499
Cyphophthalmus rumijae DNA100492Cyphophthalmus duricorius DNA100487
Cyphophthalmus gjorgjevici DNA100498
Paramiopsalis ramulosus DNA103538Paramiopsalis ramulosus DNA100459Paramiopsalis sp. DNA104624Paramiopsalis eduardoi DNA101878
Siro shasta DNA101622Siro kamiakensis DNA101613Siro cf. kamiakensis DNA101611Siro exilis DNA100489Siro boyerae DNA101614
Siro calaveras DNA101623Siro acaroides DNA100488
Siro rubens DNA100457Siro valleorum DNA100461
Leptopsalis sp. DNA101932Leptopsalis sp. DNA101930
Leptopsalis sp. DNA101944, DNA101945Leptopsalis lydekkeri DNA101064
Leptopsalis novaguinea DNA101510Leptopsalis sp. DNA101937
Leptopsalis sp. DNA102042Leptopsalis sp. DNA102039Leptopsalis sp. DNA102032
Leptopsalis sp. DNA102061Leptopsalis sp. DNA103250
Leptopsalis sp. DNA102033, DNA102048
Leptopsalis sp. DNA101093, DNA101496Leptopsalis sp. DNA101938
Leptopsalis sp. DNA101489Leptopsalis sp. DNA101483
Leptopsalis sp. DNA101514
Miopsalis sp. DNA101517
Miopsalis sp. DNA101468, DNA101950
Miopsalis sp. DNA102032, DNA102053, DNA102058Miopsalis sp. DNA101519
Miopsalis sp. DNA103249 Miopsalis sp. DNA103259
Miopsalis sp. DNA103254Miopsalis sp. DNA104981
Meghalaya sp. DNA101094 500Meghalaya sp. DNA101494, DNA101506, DNA101765Meghalaya sp. DNA101767
Meghalaya sp. DNA103265Meghalaya sp. DNA102051Meghalaya sp. DNA103242, DNA103243, DNA103244
Meghalaya sp. DNA103251
Fangensis insulanus DNA100388Fangensis spelaeus DNA100669
65
CARBONIFEROUS PERMIAN TRIASSIC JURASSIC CRETACEOUS TERTIARY
144206248290354
Thai-Malay Peninsula
Eastern Himalayas
Borneo/Philippine plate
Indo-Malay Archipelago
North America
Western Europe
Mediterranean plate
Balkan Peninsula
Iberian Peninsula
Japan
pr = 0.940
pr = 0.971
pr = 0.943
pr = 0.154
pr = 0.767
pr = 0.619
pr = 0.193pr = 0.069
pr = 0.736
pr = 0.081
pr = 0.330pr = 0.222pr = 0.153
pr = 0.736
pr = 0.609 pr = 0.980
pr = 0.974
pr = 0.963
pr = 0.736
pr = 0.962
pr = 0.996
pr = 0.981
pr = 0.975
pr = 0.941
pr = 0.139pr = 0.722
pr = 0.727pr = 0.074
pr = 0.354pr = 0.178pr = 0.125
Figure 8. Ancestral range reconstructions for Boreophthalmi (Sironidae, Stylocellidae) inferred by Lagrange analysis,using stratified models. Coloured squares at terminals indicate ranges occupied by sampled species. Coloured squares onnodes indicate ranges reconstructed for hypothetical ancestors. Numbers on nodes indicate relative probability of rangesreconstructed.
EVOLUTIONARY AND BIOGEOGRAPHICAL HISTORY OF ARACHNIDS 21
© 2011 The Linnean Society of London, Biological Journal of the Linnean Society, 2011, ••, ••–••

Leptopsalis beccariiLeptopsalis sp. DNA101483Miopsalis pulicariaLeptopsalis lydekkeri DNA101064Leptopsalis sp. DNA101930Leptopsalis sp. DNA101932Leptopsalis sp. DNA101944, DNA101945Leptopsalis novaguinea DNA101510Leptopsalis sp. DNA101937Leptopsalis sp. DNA101489Leptopsalis sp. DNA102042Leptopsalis sp. DNA101093, DNA101496Leptopsalis sp. DNA101938Leptopsalis sp. DNA102032Leptopsalis sp. DNA102039Leptopsalis sp. DNA102061Leptopsalis sp. DNA103250Leptopsalis sp. DNA102033, DNA102048Meghalaya annandaleiLeptopsalis sp. DNA101514Miopsalis sp. DNA103259Miopsalis sp. DNA103249Miopsalis sp. DNA103254Miopsalis sp. DNA104981Miopsalis sp. DNA101517Stylocellus sumatranusMiopsalis sp. DNA101468, DNA101950Miopsalis sp. DNA102032, DNA102053, DNA102058Miopsalis sp. DNA101519Meghalaya sp. DNA101094, DNA101500Meghalaya sp. DNA101494, DNA101506, DNA101765Meghalaya sp. DNA101767Meghalaya sp. DNA102051Meghalaya sp. DNA103242, DNA103243, DNA103244Meghalaya sp. DNA103265Meghalaya sp. DNA103251Fangensis insulanus DNA100388Fangensis spelaeus DNA100669Suzukielus sauteri DNA101550Suzukielus sauteri DNA101543
DNA101383Parasiro minor DNA103535Odontosiro lusitanicusCyphophthalmus minutus DNA100493Cyphophthalmus ognjenovici DNA101039Cyphophthalmus trebinjanus DNA101038Cyphophthalmus teyrovskyi DNA100910Cyphophthalmus gordani DNA100495Cyphophthalmus martensi DNA100494Cyphophthalmus corfuanus DNA102111Cyphophthalmus zetae DNA100907Cyphophthalmus hlavaci DNA102099Cyphophthalmus markoi DNA100497Cyphophthalmus serbicus DNA102098Cyphophthalmus ere DNA100499Cyphophthalmus duricorius DNA100487Cyphophthalmus rumijae DNA100492Cyphophthalmus gjorgjevici DNA100498Paramiopsalis ramulosus DNA103538Paramiopsalis ramulosus DNA100459Paramiopsalis eduardoi DNA101878Paramiopsalis sp. DNA104624Iberosiro distylosMarwe coarctataSiro boyerae DNA101614Siro exilis DNA100489Siro cf. kamiakensis DNA101611Siro kamiakensis DNA101613Siro calaveras DNA101623Siro shasta DNA101622Siro acaroides DNA100488Siro rubens DNA100457Siro valleorum DNA100461Shearogovea mexascaAnkaratra franziMetagovea sp. DNA104648Metagovea sp. DNA104647Metagovea sp. DNA101686Metagovea sp. DNA101670Metagovea sp. DNA101409Metagovea sp. DNA101642Metagovea sp. DNA105826Metagovea sp. DNA101680Metagovea sp. DNA102151Metagovea sp. DNA101685Metagovea sp. DNA101410Metagovea sp. DNA101654Metagovea sp. DNA101408Huitaca sp. DNA101681Huitaca sp. DNA102150Huitaca sp. DNA101683Huitaca sp. DNA104646Huitaca ventralis DNA101674Huitaca sp. DNA101671Huitaca sp. DNA101407Neogovea sp. DNA105825Brasilogovea microphagaNeogovea kamakusaNeogovea virginie DNA104823Neogovea virginie DNA105824“Brasilogovea” sp. Tobogan DNA100869Brasilogovea sp. DNA101665Neogovea immsiNeogovea kartaboMetasiro americanus DNA105644Metasiro americanus DNA101532Metasiro americanus DNA105645Canga renatae DNA105680Parogovia cf. sironoides DNA104619Parogovia cf. sironoides DNA100462Parogovia cf. sironoides DNA101056Parogovia cf. sironoides DNA101053Parogovia sironoides DNA101061Parogovia sironoides DNA101059Parogovia sp. DNA104615Parogovia sp. DNA104618Parogovia gabonica DNA104620Parogovia sp. DNA101052Parogovia sp. DNA101057Parogovia pabsgarnoniParogovia sp. DNA105671Ogovea cameroonensis DNA104617Ogovea nasutaTroglosiro longifossa DNA100867Troglosiro urbanus DNA101710Troglosiro juberthiei DNA100344Troglosiro monteithi DNA101580Troglosiro wilsoni DNA102324Troglosiro ninqua DNA100577Troglosiro aelleni DNA100345Rakaia sp. DNA101293Rakaia solitaria DNA101294Rakaia media DNA101292Rakaia magna australis DNA100962Rakaia sp. DNA101297Rakaia sp. DNA100954Rakaia dorothea DNA100943Rakaia uniloca DNA101812Rakaia antipodiana DNA100957Rakaia pauli DNA100968Rakaia macra DNA101808Rakaia sp. DNA100958Rakaia sorenseni digitata DNA100970Rakaia sp. DNA101807Rakaia sorenseni sorenseni DNA100969
DNA101295Rakaia minutissima DNA101291Rakaia lindsayi DNA101128Rakaia stewartiensis DNA100944Aoraki calcarobtusa westlandica DNA101129Aoraki granulosa DNA101841Aoraki tumidata DOC094Aoraki sp. DNA101126Aoraki denticulata denticulata DNA100955Aoraki longitarsa DNA101806Aoraki denticulata major DNA100959Aoraki crypta DNA101289Aoraki inerma DNA100966Aoraki healyi DNA100940Austropurcellia daviesae DNA100947Austropurcellia scoparia DNA100946Austropurcellia arcticosa DNA100951Austropurcellia forsteri DNA100945Parapurcellia monticola DNA100386Parapurcellia silvicola DNA100385Neopurcellia salmoni DNA100939Chileogovea oedipus DNA100413Chileogovea sp. DNA100490Karripurcellia harveyi DNA101303Manangotria taolanaroPurcellia illustrans DNA100387Speleosiro argasiformisPettalus sp. DNA101285Pettalus sp. DNA101282Pettalus thwaitesi DNA101223Pettalus sp. DNA101288Pettalus sp. DNA101286Pettalus sp. DNA101287Pettalus sp. DNA101283
61
67
98
96
64
10098
6293
8383
87
7487
97
8571
91
5467
83
8396
88
83 99
5094
9568
67
5093
6453
99
56
86
80
85 67
6788
93
56
90 98
87
77
74
63
72
50
95
95
7887
8198
6867
5796
9764
6364
100
100
100
8954
10093
83
87
9051
8367
85 9285
100
10098
9852
6799
10074
Troglosironidae
Ogoveidae
Pettalidae
Neogoveidae
Sironidae
Stylocellidae
imla
hthp
olup
ocS .
O.Ii
mlaht
hpon
retS
.O.I
iml a
h thp
oero
B .O.I
Figure 9. Combined analysis of morphology and molecules. Strict consensus of three optimal trees based on theparsimony direct optimization analysis under parameter set 3221 (56 984 weighted steps). Clade colours correspond tothose in Fig. 2; taxa in red are represented by morphology only. Numbers above nodes indicate jackknife support values.
22 G. GIRIBET ET AL.
© 2011 The Linnean Society of London, Biological Journal of the Linnean Society, 2011, ••, ••–••

when omitted. The variables with highest contribu-tion for Stylocellidae were temperature seasonality(42.9%), annual precipitation (26.3%), and precipita-tion of the warmest quarter (11.8%). Annual pre-cipitation had the highest gain in isolation, andprecipitation of the warmest quarter decreased thegain the most when omitted. For the clade Sternoph-thalmi, the variables with the highest contributionwere annual precipitation (33.8%), precipitation ofthe driest quarter (24.1%), and isothermality (13.9%).Temperature annual range had the highest gainin isolation and also reduced gain the most whenomitted.
Results of the identity test for the MAXENT modelsbased on all variables are shown in Table 5. Resultsfrom the analysis using the Bioclim algorithm arecongruent (not shown). The identity test shows that,when considering relative ranks, the calculatedhabitat suitability scores for most of the groups aresignificantly distinct except for Stylocellidae versusSternophthalmi, the two tropical clades. Habitat suit-ability identity cannot be rejected either for Pettal-idae versus Stylocellidae at the 0.01% significancelevel. The higher values for I and D in those two casesalso show that there is significant overlap of thesuitable habitat of these clades. Pettalidae versusSternophthalmi shows also high values of I and D butidentity tests reject the null hypothesis of habitatsuitability identity.
DISCUSSION
The present data, of worldwide scope, and spanningthe geological scale from the Palaeozoic to the
present, allow us to study a group of soil arthropodsto a level of detail rarely seen in biogeographicaland phylogenetic studies. Taxonomic representationin the molecular data includes species from all non-monotypic genera and several monotypic genera; allgenera are represented in the morphological dataset. Geographical coverage includes all known worldregions where Cyphophthalmi have been reported,with the exception of Mexico (a few specimens fromtwo caves; Shear, 1977, 1980), Madagascar (fourspecimens known in total for two species; Shear &Gruber, 1996), Kenya (five specimens known from asingle cave; Shear, 1985), and the Philippine islandof Palawan (a single adult specimen known; Shear,1993c).
Our phylogenetic results provide the basis for anew classification of the suborder Cyphophthalmi.The results also set the geological time framework forthe origin and diversification of each family and theevolution of the niche preference in selected familiesor suprafamilial clades. This allows testing specificbiogeographical hypotheses, such as the supposedtotal submersion of New Caledonia (Murienne et al.,2005) or New Zealand (Goldberg, Trewick & Paterson,2008), or the reconstruction of the ancestral areas ofeach cyphophthalmid lineage.
DIRECT OPTIMIZATION ANALYSIS
To a lesser degree than for static homology, dynamichomology searches are difficult to evaluate in terms ofoptimality. In the present study, we used a strategy ofSATF with multiple rounds of analyses to decidewhen to stop the searches, and saw that searches of6–12 h run on a desktop computer stabilized after fiveto ten rounds, depending on the parameter set. Thestability of the results is used here as a criterion for
Table 5. Habitat suitability overlap statistics based onthe MAXENT analysis with all bioclim variables
Clade Relative rank I D
Pettalidae versusSironidae
0.727** 0.392 0.187
Pettalidae versusStylocellidae
0.826* (P = 0.015) 0.525 0.234
Pettalidae versusSternophthalmi
0.832** 0.638 0.329
Sironidae versusStylocellidae
0.666** 0.126 0.038
Sironidae versusSternophthalmi
0.768** 0.292 0.114
Stylocellidae versusSternophthalmi
0.836 (P = 0.133) 0.819 0.554
Relative rank significance calculated using ENMtoolsidentity test results. *0.01 < P < 0.05, **P < 0.01. I, the Istatistic (Warren et al., 2008); D, Schoener’s D (Schoener,1968).
Table 6. Classification system for Cyphophthalmi, usingestablished family and superfamily names
Suborder CyphophthalmiInfraorder Scopulophthalmi new clade
Family Pettalidae Shear, 1980Infraorder Sternophthalmi new clade
Family Troglosironidae Shear, 1993Superfamily Ogoveoidea Shear, 1980
Family Ogoveidae Shear, 1980Family Neogoveidae Shear, 1980
Infraorder Boreophthalmi new cladeSuperfamily Stylocelloidea Hansen & Sørensen,
1904 new compositionFamily Stylocellidae Hansen & Sørensen, 1904
Superfamily Sironoidea Simon, 1879 newcomposition
Family Sironidae Simon, 1879
EVOLUTIONARY AND BIOGEOGRAPHICAL HISTORY OF ARACHNIDS 23
© 2011 The Linnean Society of London, Biological Journal of the Linnean Society, 2011, ••, ••–••

reporting results, in the same fashion that drivensearches and similar techniques have been applied tothe computational problem of tree searching (Giribet,2007a; Goloboff, Farris & Nixon, 2008).
In previous studies of cyphophthalmid and harvest-men data, analyses based on direct optimization haveyielded results sometimes differing from those ofanalyses based on static homology (Boyer et al.,2007b; Giribet et al., 2010). However, this is not thecase in the present study, where taxon sampling andgeographical representation have been thoroughlyoptimized. One major difference remains, the mono-phyly of Sironidae (see below), although some ofthe static homology analyses (Fig. 5) are congruentwith the direct optimization tree (Fig. 3), whereas themaximum likelihood tree (Fig. 4) differs from theBayesian estimate (Fig. 5). Another major difference(but, again, among analyses, and not necessarily theresult of differences between dynamic and staticnotions of homology) is the internal resolution of thepettalid genera (see below).
SYSTEMATICS
The monophyly of Cyphophthalmi has been well sup-ported in all morphological analyses (Giribet & Boyer,2002), as well as from the earliest molecular analysesusing just a few sequences in the families Sironidaeand Stylocellidae (Giribet et al., 1999; Shultz &Regier, 2001; Giribet et al., 2002), a few representa-tives of the suborder (Giribet & Boyer, 2002) or, morerecently, in several much denser analyses (Boyeret al., 2007b; Giribet et al., 2010). Our new data addcorroboration to this well-delimited taxon, with thefamilial inter-relationships and their internal struc-ture being the real focus of the study, although, in thecombined analysis including taxa with morphologicaldata only, support for the monophyly of Cyphoph-thalmi decreases to 61%, probably as a result of someeffects of the missing data (see below). Among these,Pettalidae, Stylocellidae, Troglosironidae, andNeogoveidae are monophyletic in most of our analyses(but see discussion on Neogoveidae), Ogoveidae isrepresented by a single specimen in the molecularanalyses, and Sironidae remains contentious, espe-cially with respect to the placement of the two generaSuzukielus and Parasiro.
One of the outstanding issues in cyphophthalmidphylogenetics has been the placement of the root,which was suggested to occur: (1) between Stylocel-lidae and the remaining families; (2) between Pettal-idae and the remaining families; or (3) between aclade containing Suzukielus and Pettalidae and theremaining families (Giribet & Boyer, 2002), based onthe molecular rooting of a morphological tree, becausemost cyphophthalmid morphological characters are
inapplicable or meaningless outside the suborder.Subsequent analyses found alternative resolutionsplacing the root between Pettalidae and the rest orbetween Stylocellidae and the rest (Boyer et al.,2007b), depending on the analysis and optimalitycriterion employed. Different studies have assumedeither one of these alternative rootings until a recentbroader Opiliones study found the root between Pet-talidae and the rest (Sternophthalmi + Boreoph-thalmi), this time without distinction amongoptimality criteria or method of analysis (Giribetet al., 2010). This latter result is further corroboratedin the present study. This position of Pettalidae assister group to all other cyphophthalmid families fal-sifies the two infraorders introduced by Shear (1980),which should be abandoned, and allows for a muchclearer reconstruction of the cyphophthalmid ances-tor, which must have had laterally positioned simpleocelli (Alberti, Lipke & Giribet, 2008), a lamelliformadenostyle in the male fourth tarsus, and opisthoso-mal exocrine glands opening in the anal region in themale.
Internal resolution of Pettalidae does not differconsiderably from the source studies of this pettaliddata set (Boyer & Giribet, 2007, 2009) and, as inthese studies, South Africa, New Zealand and Aus-tralia are not monophyletic. Diversification of thefamily started 183 Mya, and therefore paralogy ofsome of its landmasses is easily explained by clado-genesis prior to the split of Gondwana into itscurrent continents. Nonetheless, relationships withinPettalidae remain unstable or poorly supported andimportant African diversity is missing from themolecular sampling, both from South Africa (deBivort & Giribet, 2010) and Madagascar (Shear &Gruber, 1996), although the combined analyses withmorphology place Speleosiro as sister group to Pur-cellia (64% JF) (Giribet, 2003a; de Bivort et al., 2010;de Bivort & Giribet, 2010), and Manangotria assister group to Karripurcellia, although with lownodal support.
Results within Pettalidae are congruent amongmethods of analysis in the monophyly of each genus,although their relationships remain contentious. Atrans-Tasman clade is found, albeit with low support,in the probabilistic analyses but not in the directoptimization analysis. Similarly, the most-basal posi-tion of Parapurcellia is not universally accepted.Other relationships discussed above are poorly sup-ported, with the exception of a Chileogovea + Purcel-lia clade. Whether the deficient sampling in SouthAfrica (whose genera appear to have influence at thebase of the tree) or perhaps lineage extinction duringthe cooling of Antarctica are responsible for the lackof resolution in the pettalid relationships, remainsuntested.
24 G. GIRIBET ET AL.
© 2011 The Linnean Society of London, Biological Journal of the Linnean Society, 2011, ••, ••–••

The present study introduces the first geneticdata for the monogeneric family Ogoveidae, whichclearly forms part of the previously establishedTroglosironidae–Neogoveidae clade (Boyer et al.,2007b; Sharma & Giribet, 2009a), now namedSternophthalmi. Ogoveidae forms a clade withNeogoveidae in all analyses, corroborating Shear’ssuperfamily Ogoveoidea, although not his infraorderTropicophthalmi, because Stylocellidae are unrelatedto Ogoveoidea. Ogoveoidea is thus a Pantropical cladeof probable African origin, although its original diver-sification dates back to 261 Mya. Some analyses (twosuboptimal parameter sets under direct optimization)place Ogoveidae as ingroup Neogoveidae, althoughmost analyses support monophyly of Neogoveidae.This is also found in the combined analysis withmorphology, where Ogovea and Parogovia form aclade of African Ogoveoidea, although support for thisclade is low. The latter clade is sister to a clade ofAmerican neogoveids. However, because of the uniquemorphology of ogoveids (Juberthie, 1969; Giribet &Prieto, 2003), and monophyly of Neogoveidae in mostanalyses, the family Ogoveidae is maintained as valid(after rediagnosis from Giribet & Prieto, 2003). Shear(1980) included the genus Huitaca in this family,although, earlier, he had postulated a sister grouprelationship of Huitaca and Metagovea (Shear,1979a), as shown in our analyses.
Neogoveidae began its own diversification soonafter (236 Mya), long before the opening of the Atlan-tic Ocean, as illustrated by the amphi-Atlantic claderelating the Eastern Brazilian genus Canga with theAfrican Parogovia (specimens from Cameroon, Gabon,and Equatorial Guinea), or the sister group relation-ships of the North American genus Metasiro to theAmazonian/West African clade. The specimen fromIvory Coast, probably related to P. pabsgarnoni, con-stitutes a new genus that will be described elsewhere.Other than Canga, the South American species forma well supported clade that we currently assign tofour genera: Brasilogovea, which we resurrect here,includes species from Amazonia and the ‘Tepuis’region of Colombia; Neogovea, represented by twospecies from Guyana and French Guiana; Huitaca,still endemic to Colombia, including a large number ofundescribed species; and Metagovea, including notonly most specimens from the Andean region, but alsosome Amazon specimens from Leticia and a specimenfrom Guyana, with the latter being sister to all otherMetagovea and possibly constituting another newgenus (L. Benavides & G. Giribet, unpubl. data). Thisspecies is clearly unrelated to the genus Neogovea,occurring in this part of the Neotropics, and it ischaracterized by a conspicuous opisthosomal mid-dorsal longitudinal sulcus; an adenostyle ending in abrush of setae and located at the base or towards the
centre of the dorsal side of tarsus IV; absence ofopisthosomal exocrine glands; and a spermatopositorcomplex with a crown-shaped structure at the tip,with additional perpendicular projections (L. Bena-vides & G. Giribet, unpubl. data). Our Metagoveaclade includes specimens that we previously placed inthe genera Metagovea and Neogovea (Boyer et al.,2007b; Giribet et al., 2010) because they differ consid-erably in their anatomy. Further subdivision ofMetagovea may be warranted, although not untilspecimens from the Manaus area (Brazil) are avail-able for molecular study. Nonetheless, relationshipsamong the four genera are well established, withBrasilogovea + Neogovea being the sister group to aclade including Huitaca and Metagovea, and thelatter genus generally divided among small species,or ‘typical’ Metagovea and larger species more similarto Neogovea. The large sampling within the super-family, including all the currently recognized genera,and new data for many mostly undescribed speciesand genera not represented in previous studies,allows us to provide a more comprehensive under-standing of this Pantropical group. The addition ofmorphological data of the types of the generaNeogovea and Brasilogovea did not fully resolve thisclade, although this is considered to be a result of thepoor preservation of these specimens (missing theventral opisthosomal region) that does not allowexamination of key characters such as the sternum orthe opisthosomal exocrine glands.
The sister group of Ogoveoidea is without doubt theNew Caledonia endemic genus Troglosiro, and bothseparated approximately 279 Mya at a time whenNew Caledonia was geographically located at theeastern margin of Gondwana. Diversification of Tro-glosironidae is, however, much more recent (57 Mya)and the error associated with this date does not allowfor an unambiguous interpretation of the postulatedtotal submersion of the island (Grandcolas et al.,2008; Murienne et al., 2008; Murienne, 2009). Theanalyses recognize a group with sternal opisthosomaldepressions associated with the sternal exocrineglands of the males, sensu Sharma & Giribet (2009a).
Stylocellidae have gone from being the most poorly-known group to arguably the most stable and bestunderstood phylogenetically. The results of thepresent study corroborate those from the recentstudies mostly by R. Clouse (Clouse & Giribet, 2007;Clouse et al., 2009; Clouse, 2010; Clouse & Giribet,2010; Clouse et al., 2011), from which all the dataincluded here were derived; see also Schwendinger &Giribet (2005). Fangensis, a clade with its origins inthe terrane that today constitutes the Thai-Malaypeninsula, is sister to all other stylocellids, whichdiversified towards the north in the genus Meghalayaand towards the south in the genera Miopsalis and
EVOLUTIONARY AND BIOGEOGRAPHICAL HISTORY OF ARACHNIDS 25
© 2011 The Linnean Society of London, Biological Journal of the Linnean Society, 2011, ••, ••–••

Leptopsalis, the former mostly found on Borneo,although giving rise to species in the Philippines andSumatra, and the latter radiating rapidly asSumatra, Java, and Sulawesi became accessible, andalso extending to New Guinea. The family diversified167 Mya and includes the only reported cases ofpossible transoceanic dispersal in Cyphophthalmi(Clouse & Giribet, 2007; see also Clouse et al., 2011).Morphological analysis of discrete character data alsosupport earlier studies excluding the nominal genusStylocellus from any of the four genera adopted here,with Stylocellus remaining monotypic (Clouse et al.,2009). However, the addition of the type species of thegenera Leptopsalis, Meghalaya, Miopsalis, and Stylo-cellus, not available for molecular analysis, does notresult in a well-resolved taxonomy. The problem,however, lies in the nature of the data because thetype of Stylocellus, an old, pinned, deformed speci-men, is difficult to position based on discrete-characters only (Clouse et al., 2009), whereas the typeof Miopsalis is a female and thus misses most discretecharacters coded for other specimens, and was notavailable to be included in the morphometric analysesof Clouse et al. (2009). Leptopsalis is, however,well placed within its supposed molecular clade,although the type of Meghalaya does not find goodsupport.
Sironidae monophyly has been disputed in previousanalyses based on morphology and molecules, wheretwo genera, Parasiro and Suzukielus, often do notcluster with the remaining sironids (in the generaSiro, Cyphophthalmus, Paramiopsalis, and Iberosiro)(Giribet & Boyer, 2002; de Bivort & Giribet, 2004;Boyer et al., 2007b; Giribet et al., 2010). Odontosiro,never included in a molecular analysis, is sister toParasiro outside of the typical sironids. The KenyanMarwe has been placed within sironids in some analy-ses based on morphological data (de Bivort & Giribet,2004), as also shown here, and appears related toParamiopsalis and Iberosiro. Sironids found theirmonophyly, however, in a recent morphological analy-sis of continuous characters (de Bivort et al., 2010)and in the maximum likelihood analysis of Giribetet al. (2010), as does the maximum likelihood analysisof the present data set, albeit with bootstrap supportbelow 50%. Monophyly of Sironidae is also foundin the analysis of the nuclear ribosomal data underdirect optimization, suggesting that the non-monophyly of the family may be an artefact intro-duced most probably by their unusual COI evolution(Boyer et al., 2005). However, the membership inSironidae of the genera Iberosiro, Odontosiro, or evenMarwe remains untested with molecular data andfuture sampling effort in the north-western IberianPeninsula and in Kenya should focus on these highlycontroversial genera.
MORPHOLOGICAL DATA
It is well known that combined analyses of moleculesand morphology are fundamental for understandingthe systematics of groups that include many taxa forwhich molecular data cannot be obtained (Eernisse &Kluge, 1993; Nixon & Carpenter, 1996). A typicalexample of the latter case is provided by fossil taxa(Giribet, 2010; Murienne, Edgecombe & Giribet,2010a; Pyron, 2011). When dealing with such taxa,missing data can become a concern, although it hasbeen shown, both with simulation and with empiricalresults, that missing data in and of themselves arenot always problematic; instead, it is informationcontent in the data at hand what really matters(Wiens, 2003; Goloboff et al., 2009; Hejnol et al., 2009;Wiens, 2009).
The fossil record of Cyphophthalmi is scarce(Dunlop & Giribet, 2003; Poinar, 2008; Dunlop &Mitov, 2011), and it does not add important diversitythat can be coded into an explicit data matrix.However, the problem of missing data is of greatimportance in Cyphophthalmi as a result of thegroup’s almost global but highly localized distribu-tion, making collecting an arduous task. For thesereasons, many species are known only from oldmuseum material, from a single male, or even fromonly females or juveniles, making it impossible toinclude them in the molecular matrix or presentingconsiderable amounts of missing data in our morpho-logical matrix. To mention just a few examples, thetype species of the genera Miopsalis and Ogovea areknown only from female individuals (Thorell, 1890–1891; Hansen & Sørensen, 1904); the second speciesin the genus Pettalus, Pettalus brevicauda Pocock1897, was based on a juvenile specimen (Giribet,2008); Stylocellus sumatranus, currently the onlyspecies in the genus, is based on a deformed specimenin very poor condition (Clouse et al., 2009); andseveral monotypic genera have never been examinedunder a scanning electron microscope: Ankaratra,Manangotria, Marwe, and Odontosiro.
When trying to maximize the diversity representedin the present study, we included all currently recog-nized genera in our morphological matrix, althoughsome of the species representing these genera aremissing important characters. Despite this problem,we followed earlier recommendations into a combinedanalysis in POY under the optimal parameter set andsubmitted it to a jackknife analysis. The resultingtree was not too different from that of an early analy-sis of all cyphophthalmid genera (Giribet & Boyer,2002) with respect to the lack of resolution for manyclades, which is otherwise well supported by themolecular data sets. Notable results are the place-ment of Managotria taolanaro within Pettalidae (63%
26 G. GIRIBET ET AL.
© 2011 The Linnean Society of London, Biological Journal of the Linnean Society, 2011, ••, ••–••

jackknife support), the monophyly of Stylocellidae(including the types of the genera Leptopsalis, Megha-laya, Miopsalis, and Stylocellus, despite the lack ofmolecular data; 63% jackknife support) or the mono-phyly of the Neotropical Neogoveidae (exceptingCanga), including the types and morphology-onlyspecies of Neogovea [N. immsi, Neogovea kartabo(Davis, 1937)], Neogovea kamakusa Shear, 1977), andBrasilogovea microphaga Martens, 1969 (56% jack-knife support). The inclusion of Parogovia pabsgar-noni affects the monophyly of the African neogoveids,as Parogovia sp. DNA105671 does not form a cladewith the other Parogovia (54% jackknife support).However, the instability of species such asShearogovea mexasca and Ankaratra franzi affectsthe monophyly of groups that are otherwise robustto molecular analysis such as Sternophthalmi +Boreophthalmi, Sternophthalmi, Ogoveoidea, Neogo-veidae or Boreophthalmi.
The current results combining morphology withmolecules lowered overall support for the tree. This isan unfortunate result because simulations haveshown that the accuracy in the phylogenetic place-ment of fossils often improves or stays the same whenusing molecular data and that only in a few casesaccuracy was significantly decreased (Wiens, 2009).The problem here may be related to the low numbersof discrete morphological characters available forthese Opiliones, often limited to variation amonggroups of species; hence, the recent use of continuouscharacters in some analyses of the group (Clouseet al., 2009; de Bivort et al., 2010; de Bivort & Giribet,2010). Some characters show low levels of homoplasy,greatly structuring the data (some of these characterswere the basis for older classification systems) butmany of our wild taxa show ‘unexpected’ states inthese characters. Character 7 is notable in thisrespect. The coxae of the walking legs of Cyphoph-thalmi show different degrees of fusion, with coxae IIIand IV of each side always fused and coxae I remain-ing moveable. Coxa II can be free (state 0) or fused tocoxae III and IV; among the former are most membersof the families Pettalidae, Troglosironidae, andSironidae (except for the genera Paramiopsalis andIberosiro); among the latter are the members of thefamilies Ogoveidae, Stylocellidae, and Neogoveidae(except for Canga and Metasiro). It is therefore notunexpected that this character defined the majorgroups Sironoidea and Stylocelloidea sensu Hansen &Sørensen (1904), and that a genus such as Metasirowas considered a member of Sironidae in previousstudies, nor that these genera are among the mostunstable ones when morphology is used. Thepresence/absence of eyes (character 1), ozophore type(character 2), and spiracle shape (character 49) arealso characters with relatively low levels of
homoplasy, which have played an important role incyphophthalmid systematics, and, again, it is notunexpeced that the taxa that present odd characterstates become unstable.
BIOGEOGRAPHICAL PATTERNS IN CONTINENTS
Cyphophthalmi have been shown to present a highcorrelation of their systematic position and landmassaffinity (Juberthie & Massoud, 1976; Shear, 1980;Giribet, 2000), to show strong genetic structure acrossshort distances (Boyer et al., 2007a), and have beenused as models to study vicariance biogeography(Giribet, 2003a; Boyer et al., 2005; Boyer & Giribet,2007; Giribet & Kury, 2007; Boyer et al., 2007b; Boyer& Giribet, 2009; Clouse et al., 2009; Sharma &Giribet, 2009a; Clouse, 2010; Clouse & Giribet, 2010;de Bivort & Giribet, 2010; Murienne et al., 2010b;Clouse et al., 2011). This study corroborates earlierfindings that suggest a temperate Gondwanan clade(Pettalidae; Fig. 6), a Pantropical clade (Sternoph-thalmi; Fig. 7), one clade originating in the Thai-Malay Peninsula (Stylocellidae; Fig. 8), and two ormore Laurasian clades whose ancestral area is diffi-cult to reconstruct with high probability but thatincludes the Iberian Peninsula, North America, andWestern Europe (Sironidae; Fig. 8). The origin of allthese clades is ancient, preceding the fragmentationof Pangea and therefore suggesting that many clado-genetic events were older than the vicariant eventsthat followed. This has important biogeographicalimplications with respect to using vicariant events ascalibration points because the mismatch between thetwo events could be very large (Kodandaramaiah,2011).
Most biogeographical patterns observed, in conjunc-tion with a well-dated phylogenetic hypothesis and areconstruction of the ancestral landmasses for eachclade, allow a thorough explanation of each clade. Theancestral area reconstruction of the family Pettalidae(Fig. 6) involves several cladogenetic events at thegenus-level because each genus is currently recog-nized to be restricted to a single landmass or toadjacent terranes (Boyer & Giribet, 2007). Althoughresolution among the genera finds low support, mostanalyses suggest the South African genus Parapur-cellia to be the sister group to all other genera, andplace the other South African genus, Purcellia, in thatclade, lending support to South Africa as one of thepossible centres of origin of the family. A relationshipbetween South Africa (Purcellia) and South America(Chileogovea) is found in most analyses, as is alsofound in the members of the peripatopsid Ony-chophora, with similar distribution and habitatrequirements as pettalids (Allwood et al., 2010). It isalso notable that the two Australian genera Austro-
EVOLUTIONARY AND BIOGEOGRAPHICAL HISTORY OF ARACHNIDS 27
© 2011 The Linnean Society of London, Biological Journal of the Linnean Society, 2011, ••, ••–••

purcellia (Queensland) and Karripurcellia (WesternAustralia) never form a clade, supporting earlierviews about using microareas in biogeographicalstudies of small soil organisms (Giribet & Edgecombe,2006). A relationship of Sri Lanka–Australia–NewZealand is found in several analyses.
The biogeographical patterns of Sternophthalmi(Fig. 7) are easily reconstructed, with two ancestrallineages occurring in the Neotropics (Canga is sisterto the African Parogovia clade), two ancestral lin-eages in Africa (Ogovea and Parogovia), and onelineage in North America (Metasiro), which separatedfrom the remaining neogoveids during the Triassic.Although older analyses suggested a relationship ofMetasiro to Parogovia, this was based on analyses
without several neogoveid lineages and withoutogoveids, and the current results are very stable. Thesister group relationship of Ogoveoidea to the NewCaledonian endemic genus Troglosiro has been foundin previous studies and it is discussed in more detailbelow. No paralogy is needed in this tree when con-sidering the timing of the diversification events,as the separation of the Neotropics from the Afrotro-pics is dated at 95 Mya (Raven & Axelrod, 1972;Sanmartín, 2002).
Stylocellid biogeography and their ancestral areashave been discussed recently (Clouse & Giribet, 2010)and our results corroborate this earlier analysis.The Thai-Malay Peninsula is reconstructed as theancestral terrane for the family with subsequent
A
B
C
D
Figure 10. MAXENT models of habitat suitability. Left column with all bioclim variables, right column with variableswith jackknife regularized training gain greater than one. Warmer (red-yellow) colours represent more suitable habitats.Maps in miniature represent actual presence observations. A, Pettalidae; B, Sironidae; C, Stylocellidae; D, Sternophthalmi(Troglosironidae + Ogoveidae + Neogoveidae).
28 G. GIRIBET ET AL.
© 2011 The Linnean Society of London, Biological Journal of the Linnean Society, 2011, ••, ••–••

expansions to the Eastern Himalayas during theCretaceous/Tertiary boundary, and radiations into theBorneo/Philippine plate and into the Indo-MalayArchipelago during the Cretaceous. Several lineagesmay have returned to the Thai-Malay Peninsula ormoved between islands around the Cretaceous/Tertiary boundary during a period in which south-east Asia was subjected to drastic changes and theIndo-Malay Archipelago variously connected (Hall,2002; Ali & Aitchison, 2008).
Reconstruction of the biogeographical history ofSironidae remains hindered by the instability of therelationships of Suzukielus and Parasiro; the formerendemic to Japan and the latter found in the Iberianand Mediterranean plates. Parasiro has its origins inthe Jurassic/Cretaceous of the Iberian Peninsula,where cyphophthalmids are so far restricted to areaswith Paleozoic rocks (Murienne & Giribet, 2009). Themain sironid clade includes three genera found inNorth America (Siro), Western Europe (Siro), theIberian Peninsula (Paramiopsalis), and the Balkanregion and Eastern Europe (Cyphophthalmus). Siroshows reciprocal monophyly of the two landmasses,the lineages separating during the Triassic, a resultnot supported in a recent analysis of the North Ameri-can diversity (Giribet & Shear, 2010). The sister-group relationship of the Iberian/Balkan clade hasbeen discussed thoroughly in recent studies (Boyeret al., 2005; Murienne et al., 2010b), which have alsoillustrated a correlation between an evolutionaryexplosion and the coming into contact of ancestrallandmasses in the Mediterranean region (Murienneet al., 2010b). From a geological point of view, theBalkan Peninsula, supporting the explosive evolutionof Cyphophthalmus, includes the margin of bothEurasia (the Moesian microplate) and Gondwana (theAdria microplate), as well as remnants of the Tethysand related marginal seas (made up of oceanic crust)(Karamata, 2006). The Adria microplate is the largestlithospheric fragment in the Central Mediterraneanregion. It was connected to Iberia in the west and tonorth-west Africa in the south (Wortmann et al.,2001) until the Middle–Late Triassic episodes ofrifting and breakup (Channell, D’Argenio & Horváth,1979; Pamic, Gusic & Jelaska, 1998), around thecladogenesis time for the split between Paramiopsalisand Cyphophthalmus. For most of the time, the Adriamicroplate was in a shallow-water environment(Scheibner & Speijer, 2008) in which the SouthernTethyan Megaplatform formed before disintegratinginto several carbonate platforms in the Early Jurassic(Vlahovic et al., 2005), before the diversification ofCyphophthalmus during the Jurassic/Cretaceous,when cycles of land submergence and emergence havebeen recorded in some carbonate platforms (Vlahovicet al., 2005; Márton et al., 2008). The ancestral area of
the family is however difficult to infer, perhaps,amongst other factors, as a result of the large numberof terranes that existed around the Tethys.
BIOGEOGRAPHY IN CONTINENTAL ISLANDS: THE
CASES OF NEW CALEDONIA AND NEW ZEALAND
Cyphophthalmi are present in most islands of conti-nental origin (fragment islands sensu Gillespie &Roderick, 2002), including Sri Lanka (Pocock, 1897;Sharma & Giribet, 2006; Giribet, 2008; Sharma,Karunarathna & Giribet, 2009), Chiloé (Roewer,1961; Juberthie & Muñoz-Cuevas, 1970; Shear,1993a), Corsica and Sardinia (Simon, 1872; Juber-thie, 1958), Honshu (Roewer, 1916; Juberthie, 1970b;Suzuki & Ohrui, 1972; Giribet, Tsurusaki & Boyer,2006), and the Indo-Malay archipelago (Westwood,1874; Thorell, 1882–1883; Pocock, 1897; Hansen &Sørensen, 1904; Shear, 1979b; Rambla, 1991; Shear,1993c; Giribet, 2002; Schwendinger & Giribet, 2005;Clouse & Giribet, 2007; Clouse et al., 2009; Clouse &Giribet, 2010), and, in all these cases, their presencein these islands is best explained as a result of vicari-ance. Similarly, New Caledonia and New Zealandhost a considerable diversity of Cyphophthalmi,although their presence in these islands as a resultof one or more vicariant evens has been recentlydisputed.
New Caledonia currently has 13 described speciesin the genus Troglosiro, the only genus in the familyTroglosironidae (Juberthie, 1979; Shear, 1993b;Sharma & Giribet, 2005, 2009a; Sharma & Giribet,2009b) considered to be endemic to the Grande Terreand unambiguously recovered as the sister group tothe Equatorial Ogoveoidea from Equatorial WestAfrica and the Equatorial Neotropical belt. Geologicaldata on the origins of the New Caledonian biodiver-sity argue in favour of a series of submersions duringthe Palaeocene and Eocene (Paris, Andreieff &Coudray, 1979; Aitchison et al., 1998; Pelletier, 2006),which has been used to support a total submersion ofthe island, re-emerging 37 Mya (Murienne et al.,2005; Grandcolas et al., 2008; Murienne et al., 2008).Indeed, molecular dating analyses of several NewCaledonian clades has supported diversification pro-cesses post-dating the critical date of 37 Mya (Muri-enne et al., 2005; Page et al., 2005; Murienne et al.,2008; Espeland & Johanson, 2010; Murienne, Edge-combe & Giribet, 2011), which has led some studies tosuggest that the entirety of the New Caledonian ter-restrial biota must have arrived to the islands viadispersal and that no trace of ancient vicariance isleft. One notable exception may be the family Troglo-sironidae, whose diversification has been dated at28–49 Mya by Boyer et al. (2007b) and 52–102 Mya byGiribet et al. (2010), although these studies used few
EVOLUTIONARY AND BIOGEOGRAPHICAL HISTORY OF ARACHNIDS 29
© 2011 The Linnean Society of London, Biological Journal of the Linnean Society, 2011, ••, ••–••

troglosironid samples. Refined analyses here suggesta Late Cretaceous–Early Tertiary diversificationof the family (57 Mya), predating the supposedre-emergence of New Caledonia, although the errorassociated with this date does not allow unambiguousdistinction of the hypothesis owing to the temporalproximity of the re-emergence of the island (37 Mya)and the floor of the diversification age estimate (95%HPD: 40–73 Mya). The ancestral area reconstructionfor the split between Troglosironidae and Ogoveoideais supported as a contiguous landmass containingWest Africa and New Caledonia, deep in the Permian,indicating that the range of the clade was muchbroader than it currently is (Figs 5, 7), and thatmassive extinctions may have occurred during theperiod comprised between 279–57 Mya. However,relict taxa (and Troglosironidae certainly is such anexample) and the problem of extinction, especially inthe absence of a fossil record, are mysteries that aredifficult to address in biogeography (Crisp, Trewick &Cook, 2011). This is indeed a unique case, whereTroglosironidae constitute a special lineage in thisrespect.
Another possibility is a trans-Pacific dispersal,again during the 279–57 Mya period, a phenomenonalso observed in at least two other opilionid lineages(e.g. the families Zalmoxidae and Samoidae; Sharma& Giribet, 2011). However, dispersals in the Cenozoicare possible to reconstruct unambiguously in Zal-moxidae and Samoidae insofar as lineages in one partof the Pacific form a grade with respect to a clade inanother part of the Pacific. In both these cases, Neo-tropical lineages form the paraphyletic grade withrespect to Pacific island lineages, rendering the ances-tral area reconstruction for the origin of these radia-tions as Neotropical. By contrast, Troglosironidae andthe clade (Ogoveidae + Neogoveidae) form reciprocallymonophyletic groups that diverged 279 Mya, which isinconsistent with recent dispersal. Moreover, thePermian origin of Troglosironidae also suggests thatany putative dispersal event had to have occurredsometime between 279–57 Mya, a hypothesis that isdifficult to test. As stated previously, we submit thatthe biogeographical history of Troglosironidae isinherently difficult to reconstruct as a result of therelictual nature of this lineage (Sharma & Giribet,2009a).
A similar case has been proposed for New Zealand,which includes 29 species in three pettalid genera(Aoraki, Neopurcellia, and Rakaia) (Forster, 1948,1952; Boyer & Giribet, 2003, 2007) found in twogeological terranes (the Australian plate and thePacific plate) (Boyer & Giribet, 2009). New Zealand’sgeology and biota reflect a dynamic history of ancientGondwanan origin, long-term isolation from othercontinental landmasses, marine inundating during
the Oligocene, glaciation during the Pleistocene, andevolutionary radiations that have produced a spec-tacular proportion of endemic species (Gibbs, 2006).Studies have focused on New Zealand’s biogeographywith particular vigour over the past two decadesbecause molecular systematics has provided newtools with which to approach evolutionary questions.Molecular systematists have addressed topics suchas the number and location of Pleistocene refugia(Marske et al., 2009; Buckley, Marske & Attanayake,2010), the Alpine Fault Hypothesis (Heads & Craw,2004), and, most contentiously, a vicariance versusdispersal-based origin of New Zealand’s terrestrialbiota (Trewick, Paterson & Campbell, 2007; Phillipset al., 2010). Although studies have long recognizedthat land area was drastically reduced (i.e. to lessthan 15% of its current size) during the marineincursions of the Oligocene (Cooper & Cooper, 1995),more recently Waters & Craw (2006) have suggestedthat there is no strong evidence for continuouslyemergent land throughout the period (Landis et al.,2008). Trewick et al. (2007) and Wallis & Trewick(2009) asserted that the preponderance of biogeo-graphical evidence favours a scenario of completesubmergence during the Oligocene, and some studieshave gone further, suggesting that the entire terres-trial biota arrived via dispersal during the last 22Myr (Landis et al., 2008), and that it is thereforemore like that of an oceanic archipelago than a con-tinent (Goldberg et al., 2008). Few have questionedthis new trend in New Zealand biogeography (Knappet al., 2007; Edgecombe & Giribet, 2008; Boyer &Giribet, 2009; Allwood et al., 2010; Giribet & Boyer,2010).
The evolutionary history of the New Zealandcyphophthalmid genera (Aoraki, Rakaia, and Neopur-cellia) has long been of interest as part of the evalu-ation of a hypothesis proposing total submersion ofNew Zealand in the Oligocene (Waters & Craw, 2006;Trewick et al., 2007; Goldberg et al., 2008; Landiset al., 2008; Wallis & Trewick, 2009; Giribet & Boyer,2010; Phillips et al., 2010). The persistence of theselineages through the Oligocene bottleneck was con-sidered to represent evidence of incomplete submer-sion of this landmass (Boyer & Giribet, 2007, 2009),although this hypothesis was not previously accom-panied by molecular dating. Consequently, the agesof diversification of these lineages have been opento interpretation as very young (e.g. approximately5 Mya; Goldberg et al., 2008). Moreover, Crisp et al.(2011) suggested that an important criterion for evi-dence of vicariance events is diversification time coin-cident with the timing of the geological event thatprecipitated the vicariance; in this case, the rifting ofZealandia from the Australian plate approximately85 Mya, although cladogenesis could be expected to be
30 G. GIRIBET ET AL.
© 2011 The Linnean Society of London, Biological Journal of the Linnean Society, 2011, ••, ••–••

much older than the vicariant event in taxa with lowvagility and small distribution ranges.
The present study, utilizing a robust methodologyfor simultaneous estimation of tree topology and cladedivergence times (sensu Crisp et al., 2011), and cali-brated using fossil taxa exclusively (Kodandara-maiah, 2011), obtains the following diversificationtimes for the New Zealand endemic genera Rakaiaand Aoraki: 91 Mya (95% HPD: 72–108 Mya) and90 Mya (95% HPD: 75–108 Mya), respectively. Thesediversification age estimates coincide with the riftingof Zealandia in the Late Cretaceous. We present evi-dence, therefore, based upon tree topology and cladedivergence times, of the persistence of multiple lin-eages through the Oligocene. In addition, the presentstudy reconstructs the origin of the genus Aoraki tothe Australian plate during the Cretaceous and thatof the genus Rakaia to a composite terrane in thePacific and Australian plate also in the middle of theCretaceous, although, later on, our study clearlyassigns a clade to each terrane (Figs 5, 6). Unfortu-nately, the generic relationships are highly unstableacross methods and parameter sets and furtherspeculation about the relationships of the Australianand New Zealand genera awaits further data. Wesubmit that these data falsify the hypothesis of com-plete submersion of New Zealand during the Oli-gocene Drowning. Recent and forthcoming studies ofother invertebrate lineages (Allwood et al., 2010;Giribet & Boyer, 2010; Murienne et al., 2010a;Marshall, 2011) are anticipated to corroborate thisconclusion.
HABITAT SUITABILITY MODELS
One common characteristic of all models that wepresent here is the larger suitable habitat than thearea actually occupied by the four clades of interest.This constitutes further evidence for the old cladogen-esis and low dispersal abilities of Cyphophthalmibecause many areas of suitable habitat have neverbeen in contact with a landmass occupied by the cladeof interest. This pattern also corroborates the hypoth-esis that tectonic movements and vicariance eventshave defined distributions and driven diversificationin this group of soil arthropods. Mysteries remainbecause certain temperate clades have migrated towarmer climates (e.g. Pettalus in Sri Lanka orAustropurcellia in Queensland, Australia), whereasothers may not have been able to adapt to changingclimates. This suggests that, in several lineages, pro-cesses of niche evolution might have taken place.However, the lack of detailed occurrence observationsfor many species does not currently allow studyingthe niche evolution in Cyphophthalmi in greaterdetail.
CONCLUDING REMARKS
Cyphophthalmi constitute an ancient lineage of Opil-iones distributed in temperate to tropical rainforestsworldwide but restricted for the most part to conti-nents and islands of continental origin, representingan ideal group of organisms for studying vicariancebiogeography. Both phylogenetic patterns derivedfrom molecular and morphological data and moleculardating using Opiliones fossils as calibration pointscorroborate the old age of the group and of its con-stituent clades. Ancestral area reconstruction furthercorroborates our biogeographical predictions byrequiring only minimal switches between land-masses, most of them through contiguous land, there-fore showing that the actual distribution is muchmore restricted than the potential distributiondefined by the modelled habitat suitability for thedifferent familial/suprafamilial clades. The data alsopermit tests of more general biogeographical hypoth-eses, such as the total submersion of New Caledoniaand New Zealand and, at least in the former case,contradict a scenario of complete inundation. Thepresent study provides refinement not only ofthe phylogenetic relationships and taxonomy of thegroup, but also its evolutionary and biogeographicalhistory.
ACKNOWLEDGEMENTS
This work is the result of more than a decade ofintense research on Cyphophthalmi and many indi-viduals and institutions have contributed to it. Firstand foremost, this work has been possible in largepart as a result of grants from the National ScienceFoundation (DEB-0236871 to G.G., DEB-0508789 toS.B. and G.G., and DEB-0205982 to M. Sharkey) andto numerous Putnam Expeditions Grants from theMuseum of Comparative Zoology to collect in Austra-lia, Cameroon, Gabon, Indonesia, New Zealand,South Africa, and Sri Lanka. J.M. was supported by aMarie Curie International Outgoing Fellowship(grant 221099) within the 7th European CommunityFramework Program. Many colleagues have assistedus in the field, and they are acknowledged in ourprevious papers. We want to emphasize, however, thehelp of Carlos Prieto for work in Equatorial Guinea,Indika Karunarathna for work in Sri Lanka, CahyoRahmadi for work in Indonesia, Nobuo Tsurusaki forwork in Japan, Phil Sirvid and Ricardo Palma forwork in New Zealand, Hervé Jourdan for work inNew Caledonia, Nono Legrand for work in Cameroon,and many other colleagues who provided samples,especially Louis Deharverg for south-east Asiansamples, Mike Sharkey for his Colombian samples,Rudy Jocqué for samples from Ivory Coast and
EVOLUTIONARY AND BIOGEOGRAPHICAL HISTORY OF ARACHNIDS 31
© 2011 The Linnean Society of London, Biological Journal of the Linnean Society, 2011, ••, ••–••

Guyana, Eduardo Mateos for Spanish samples, andRicardo Pinto-da-Rocha for Brazilian samples. GregEdgecombe, Salvador Carranza, and Michele Nishigu-chi accompanied us on many collecting trips. We arealso indebted to many museums and curators fortheir long-term loans that made this research pos-sible, especially Lorenzo Prendini and Norman Plat-nick from the American Museum of Natural History,New York, NY; Charles Griswold from the CaliforniaAcademy of Sciences, San Francisco, CA; Petra Sier-wald from the Field Museum, Chicago, IL; Jan Bec-caloni from The Natural History Museum, London;Arturo Muñoz Cuevas from the Muséum nationald’Histoire naturelle, Paris; Phil Sirvid from Te PapaTongarewa, Wellington; Jonathan Coddington fromthe US National Museum (Smithsonian Institution),Washington, DC; Nikolaj Scharff from the ZoologicalMuseum, Natural History Museum of Denmark,Copenhagen; and Jason Dunlop from the Zoologis-chen Museums, Berlin. Finally, Laura Leibenspergerhas assisted with specimen loans during all theseyears. The editor John A. Allen, an anonymousreviewer, and Gustavo Hormiga provided commentsthat helped to improve upon an earlier version of thismanuscript.
REFERENCES
Aitchison JC, Ireland TR, Clarke GL, Cluzel D, DavisAM, Meffre S. 1998. Regional implifications of U/PbSHRIMP age constraints on the tectonic evolution of NewCaledonia. Tectonophysics 299: 333–343.
Alberti G. 1995. Comparative spermatology of Chelicerata:review and perspective. In: Jamieson BGM, Ausió J, JustineJ-L, eds. Advances in spermatozoal phylogeny and tax-onomy. Mémoires du Muséum National d’Histoire Naturelle166: 203–230.
Alberti G. 2005. Double spermatogenesis in Chelicerata.Journal of Morphology 266: 281–297.
Alberti G, Giribet G, Gutjahr M. 2009. Ultrastructure ofspermatozoa of different species of Neogoveidae, Sironidae,and Stylocellidae (Cyphophthalmi: Opiliones). Contributionsto Natural History – Scientific papers from the NaturalHistory Museum Bern 12: 53–69.
Alberti G, Lipke E, Giribet G. 2008. On the ultrastructureand identity of the eyes of Cyphophthalmi based on a studyof Stylocellus sp. (Opiliones, Stylocellidae). Journal ofArachnology 36: 379–387.
Ali JR, Aitchison JC. 2008. Gondwana to Asia: plate tec-tonics, paleogeography and the biological connectivity of theIndian sub-continent from the Middle Jurassic throughlatest Eocene (166–35 Ma). Earth-Science Reviews 88: 145–166.
Allwood J, Gleeson D, Mayer G, Daniels S, Beggs JR,Buckley TR. 2010. Support for vicariant origins of the NewZealand Onychophora. Journal of Biogeography 37: 669–681.
Araujo MB, Rahbek C. 2006. How does climate changeaffect biodiversity? Science 313: 1396–1397.
Benavides LR, Giribet G. 2007. An illustrated catalogue ofthe South American species of the cyphophthalmid familyNeogoveidae (Arthropoda, Opiliones, Cyphophthalmi) witha report on 37 undescribed species. Zootaxa 1509: 1–15.
de Bivort B, Clouse RM, Giribet G. 2010. Amorphometrics-based phylogeny of the temperate Gond-wanan mite harvestmen (Opiliones, Cyphophthalmi, Pettal-idae). Journal of Zoological Systematics and EvolutionaryResearch 48: 294–309.
de Bivort B, Giribet G. 2010. A systematic revision of theSouth African Pettalidae (Arachnida : Opiliones : Cyphoph-thalmi) based on a combined analysis of discrete and con-tinuous morphological characters with the description ofseven new species. Invertebrate Systematics 24: 371–406.
de Bivort BL, Giribet G. 2004. A new genus of cyphoph-thalmid from the Iberian Peninsula with a phylogeneticanalysis of the Sironidae (Arachnida : Opiliones : Cyphoph-thalmi) and a SEM database of external morphology. Inver-tebrate Systematics 18: 7–52.
Boyer SL, Baker JM, Giribet G. 2007a. Deep geneticdivergences in Aoraki denticulata (Arachnida, Opiliones,Cyphophthalmi): a widespread ‘mite harvestman’ defiesDNA taxonomy. Molecular Ecology 16: 4999–5016.
Boyer SL, Clouse RM, Benavides LR, Sharma P, Sch-wendinger PJ, Karunarathna I, Giribet G. 2007b. Bio-geography of the world: a case study from cyphophthalmidOpiliones, a globally distributed group of arachnids. Journalof Biogeography 34: 2070–2085.
Boyer SL, Giribet G. 2003. A new Rakaia species (Opiliones,Cyphophthalmi, Pettalidae) from Otago, New Zealand.Zootaxa 133: 1–14.
Boyer SL, Giribet G. 2007. A new model Gondwanan taxon:systematics and biogeography of the harvestman familyPettalidae (Arachnida, Opiliones, Cyphophthalmi), with ataxonomic revision of genera from Australia and NewZealand. Cladistics: The International Journal of the WilliHennig Society 23: 337–361.
Boyer SL, Giribet G. 2009. Welcome back New Zealand:regional biogeography and Gondwanan origin of threeendemic genera of mite harvestmen (Arachnida, Opiliones,Cyphophthalmi). Journal of Biogeography 36: 1084–1099.
Boyer SL, Karaman I, Giribet G. 2005. The genusCyphophthalmus (Arachnida, Opiliones, Cyphophthalmi) inEurope: a phylogenetic approach to Balkan Peninsulabiogeography. Molecular Phylogenetics and Evolution 36:554–567.
Buckley TR, Marske K, Attanayake D. 2010. Phylogeog-raphy and ecological niche modelling of the New Zealandstick insect Clitarchus hookeri (White) support survival inmultiple coastal refugia. Journal of Biogeography 37: 682–695.
Castresana J. 2000. Selection of conserved blocks from mul-tiple alignments for their use in phylogenetic analysis.Molecular Biology and Evolution 17: 540–552.
32 G. GIRIBET ET AL.
© 2011 The Linnean Society of London, Biological Journal of the Linnean Society, 2011, ••, ••–••

Channell JET, D’Argenio B, Horváth F. 1979. Adria, theAfrican Promontory, in Mesozoic Mediterranean Paleogeog-raphy. Earth Science Reviews 15: 213–292.
Clouse RM. 2010. Molecular and morphometric phylogenet-ics and biogeography of a Southeast Asian arachnid family(Opiliones, Cyphophthalmi, Stylocellidae). PhD Thesis,Department of Organismic and Evolutionary Biology,Harvard University, Cambridge.
Clouse RM, de Bivort BL, Giribet G. 2009. A phylogeneticanalysis for the South-east Asian mite harvestman familyStylocellidae (Opiliones : Cyphophthalmi) – a combinedanalysis using morphometric and molecular data. Inverte-brate Systematics 23: 515–529.
Clouse RM, General DM, Diesmos AC, Giribet G.2011. An old lineage of Cyphophthalmi (Opiliones) discov-ered on Mindanao highlights the need for biogeographicalresearch in the Philippines. Journal of Arachnology 39:147–153.
Clouse RM, Giribet G. 2007. Across Lydekker’s Line – firstreport of mite harvestmen (Opiliones : Cyphophthalmi :Stylocellidae) from New Guinea. Invertebrate Systematics21: 207–227.
Clouse RM, Giribet G. 2010. When Thailand was an island– the phylogeny and biogeography of mite harvestmen(Opiliones, Cyphophthalmi, Stylocellidae) in Southeast Asia.Journal of Biogeography 37: 1114–1130.
Cooper A, Cooper RA. 1995. The Oligocene bottleneck andNew Zealand biota: genetic record of a past environmentalcrisis. Proceedings of the Royal Society of London Series B,Biological Sciences 261: 293–302.
Crisp MD, Trewick SA, Cook LG. 2011. Hypothesistesting in biogeography. Trends in Ecology & Evolution 26:66–72.
DaSilva MB, Pinto-da-Rocha R, Giribet G. 2010. Cangarenatae, a new genus and species of Cyphophthalmi fromBrazilian Amazon caves (Opiliones: Neogoveidae). Zootaxa2508: 45–55.
De Laet J. 2010. A problem in POY tree searches (and itswork-around) when some sequences are observed to beabsent in some terminals. Cladistics: The InternationalJournal of the Willi Hennig Society 26: 453–455.
Diniz JAF, De Marco P, Hawkins BA. 2010. Defying thecurse of ignorance: perspectives in insect macroecology andconservation biogeography. Insect Conservation and Diver-sity 3: 172–179.
Drummond AJ, Ho SY, Phillips MJ, Rambaut A. 2006.Relaxed phylogenetics and dating with confidence. PLoSBiology 4: e88.
Drummond AJ, Rambaut A. 2007. BEAST: Bayesian evo-lutionary analysis by sampling trees. BMC EvolutionaryBiology 7: 214.
Dunlop JA. 2007. Paleontology. In: Pinto-da-Rocha R,Machado G, Giribet G, eds. Harvestmen: the biology ofOpiliones. Cambridge, MA: Harvard University Press, 247–265.
Dunlop JA, Anderson LI, Kerp H, Hass H. 2003.Preserved organs of Devonian harvestmen. Nature 425:916.
Dunlop JA, Anderson LI, Kerp H, Hass H. 2004. for 2003).A harvestman (Arachnida: Opiliones) from the Early Devo-nian Rhynie cherts, Aberdeenshire, Scotland. Transactionsof the Royal Society of Edinburgh: Earth Sciences 94: 341–354.
Dunlop JA, Giribet G. 2003. The first fossil cyphophthalmid(Arachnida: Opiliones), from Bitterfeld amber, Germany.Journal of Arachnology 31: 371–378.
Dunlop JA, Mitov PG. 2011. The first fossil cyphophthalmidharvestman from Baltic amber. Arachnologische Mitteilun-gen 40: 47–54.
Edgar RC. 2004. MUSCLE: multiple sequence alignmentwith high accuracy and high throughput. Nucleic AcidsResearch 32: 1792–1797.
Edgecombe GD, Giribet G. 2008. A New Zealand species ofthe trans-Tasman centipede order Craterostigmomorpha(Arthropoda : Chilopoda) corroborated by molecular evi-dence. Invertebrate Systematics 22: 1–15.
Eernisse DJ, Kluge AG. 1993. Taxonomic congruence versustotal evidence, and amniote phylogeny inferred from fossils,molecules, and morphology. Molecular Biology and Evolu-tion 10: 1170–1195.
Elith J, Graham CH, Anderson RP, Dudík M, Ferrier S,Guisan A, Hijmans RJ, Huettmann F, Leathwick JR,Lehmann A, Li J, Lohmann LG, Loiselle BA, ManionG, Moritz C, Nakamura M, Nakazawa Y, Overton JM,Peterson AT, Phillips SJ, Richardson K, Scachetti-Pereira R, Schapire RE, Soberón J, Williams S, WiszMS, Zimmermannm NE. 2006. Novel methods improveprediction of species’ distributions from occurrence data.Ecography 29: 129–151.
Espeland M, Johanson KA. 2010. The effect of environmen-tal diversification on species diversification in New Cale-donian caddisflies (Insecta: Trichoptera: Hydropsychidae).Journal of Biogeography 37: 879–890.
Evans ME, Smith SA, Flynn RS, Donoghue MJ. 2009.Climate, niche evolution, and diversification of the ‘bird-cage’ evening primroses (Oenothera, sections Anogra andKleinia). American Naturalist 173: 225–240.
Farris JS, Albert VA, Källersjö M, Lipscomb D, KlugeAG. 1996. Parsimony jackknifing outperforms neighbor-joining. Cladistics: The International Journal of the WilliHennig Society 12: 99–124.
Forster RR. 1948. The sub-order Cyphophthalmi Simon inNew Zealand. Dominion Museum Records in Entomology 1:79–119.
Forster RR. 1952. Supplement to the sub-order Cyphoph-thalmi. Dominion Museum Records in Entomology 1: 179–211.
Gibbs G. 2006. Ghosts of Gondwana. The history of life inNew Zealand. Nelson: Craig Potton Publishing.
Gillespie RG, Roderick GK. 2002. Arthropods on islands:colonization, speciation, and conservation. Annual Review ofEntomology 47: 595–632.
Giribet G. 2000. Catalogue of the Cyphophthalmi of theWorld (Arachnida, Opiliones). Revista Ibérica de Arac-nología 2: 49–76.
Giribet G. 2002. Stylocellus ramblae, a new stylocellid
EVOLUTIONARY AND BIOGEOGRAPHICAL HISTORY OF ARACHNIDS 33
© 2011 The Linnean Society of London, Biological Journal of the Linnean Society, 2011, ••, ••–••

(Opiliones, Cyphophthalmi) from Singapore, with a discus-sion of the family Stylocellidae. Journal of Arachnology 30:1–9.
Giribet G. 2003a. Karripurcellia, a new pettalid genus(Arachnida : Opiliones : Cyphophthalmi) from Western Aus-tralia, with a cladistic analysis of the family Pettalidae.Invertebrate Systematics 17: 387–406.
Giribet G. 2003b. Stability in phylogenetic formulations andits relationship to nodal support. Systematic Biology 52:554–564.
Giribet G. 2007a. Efficient tree searches with available algo-rithms. Evolutionary Bioinformatics 3: 1–16.
Giribet G. 2007b. Neogoveidae Shear, 1980. In: Pinto-da-Rocha R, Machado G, Giribet G, eds. Harvestmen: thebiology of Opiliones. Cambridge, MA: Harvard UniversityPress, 95–97.
Giribet G. 2008. On the identity of Pettalus cimiciformis andP. brevicauda (Opiliones, Pettalidae) from Sri Lanka.Journal of Arachnology 36: 199–201.
Giribet G. 2010. A new dimension in combining data? Theuse of morphology and phylogenomic data in metazoansystematics. Acta Zoologica (Stockholm) 91: 11–19.
Giribet G. 2011. Shearogovea, a new genus of Cyphophthalmi(Arachnida, Opiliones) of uncertain position from Oaxacancaves, Mexico. Breviora 528: 1–7.
Giribet G, Boyer SL. 2002. A cladistic analysis of thecyphophthalmid genera (Opiliones, Cyphophthalmi).Journal of Arachnology 30: 110–128.
Giribet G, Boyer SL. 2010. ‘Moa’s Ark’ or ‘GoodbyeGondwana’: is the origin of New Zealand’s terrestrial inver-tebrate fauna ancient, recent, or both? Invertebrate System-atics 24: 1–8.
Giribet G, Edgecombe GD. 2006. The importance of lookingat small-scale patterns when inferring Gondwanan bio-geography: a case study of the centipede Paralamyctes(Chilopoda, Lithobiomorpha, Henicopidae). BiologicalJournal of the Linnean Society 89: 65–78.
Giribet G, Edgecombe GD, Wheeler WC, Babbitt C. 2002.Phylogeny and systematic position of Opiliones: a combinedanalysis of chelicerate relationships using morphologicaland molecular data. Cladistics: The International Journal ofthe Willi Hennig Society 18: 5–70.
Giribet G, Kury AB. 2007. Phylogeny and biogeography. In:Pinto-da-Rocha R, Machado G, Giribet G, eds. Harvestmen:the biology of Opiliones. Cambridge, MA: Harvard Univer-sity Press, 62–87.
Giribet G, Prieto CE. 2003. A new Afrotropical Ogovea(Opiliones, Cyphophthalmi) from Cameroon, with a discus-sion on the taxonomic characters in the family Ogoveidae.Zootaxa 329: 1–18.
Giribet G, Rambla M, Carranza S, Baguñà J, Riutort M,Ribera C. 1999. Phylogeny of the arachnid order Opiliones(Arthropoda) inferred from a combined approach of complete18S and partial 28S ribosomal DNA sequences and mor-phology. Molecular Phylogenetics and Evolution 11: 296–307.
Giribet G, Shear WA. 2010. The genus Siro Latreille, 1796(Opiliones, Cyphophthalmi, Sironidae), in North America
with a phylogenetic analysis based on molecular data andthe description of four new species. Bulletin of the Museumof Comparative Zoology 160: 1–33.
Giribet G, Tsurusaki N, Boyer SL. 2006. Confirmation ofthe type locality and the distributional range of Suzukielussauteri (Opiliones, Cyphophthalmi) in Japan. Acta Arach-nologica 55: 87–90.
Giribet G, Vogt L, Pérez González A, Sharma P,Kury AB. 2010. A multilocus approach to harvestman(Arachnida: Opiliones) phylogeny with emphasis on bioge-ography and the systematics of Laniatores. Cladistics: TheInternational Journal of the Willi Hennig Society 26: 408–437.
Goldberg J, Trewick SA, Paterson AM. 2008. Evolutionof New Zealand’s terrestrial fauna: a review of mole-cular evidence. Philosophical Transactions of the RoyalSociety of London Series B, Biological Sciences 363: 3319–3334.
Goloboff PA, Catalano SA, Mirande JM, Szumik CA,Arias JS, Källersjö M, Farris JS. 2009. Phylogeneticanalysis of 73 060 taxa corroborates major eukaryoticgroups. Cladistics: The International Journal of the WilliHennig Society 25: 211–230.
Goloboff PA, Farris JS, Nixon KC. 2008. TNT, a freeprogram for phylogenetic analysis. Cladistics: The Interna-tional Journal of the Willi Hennig Society 24: 774–786.
Grandcolas P, Murienne J, Robillard T, DeSutter-Grandcolas L, Jourdan H, Guilbert E. 2008. NewCaledonia: a very old Darwinian island? PhilosophicalTransactions of the Royal Society of London Series B, Bio-logical Sciences 363: 3309–3317.
Hadly EA, Spaeth PA, Li C. 2009. Niche conservatism abovethe species level. Proceedings of the National Academy ofSciences of the United States of America 106: 19707–19714.
Hall R. 2002. Cenozoic geological and plate tectonic evolutionof SE Asia and the SW Pacific: computer-based reconstruc-tions, model and animations. Journal of Asian EarthSciences 20: 353–431.
Hansen HJ, Sørensen W. 1904. On two orders of Arachnida:Opiliones, especially the suborder Cyphophthalmi, andRicinulei, namely the family Cryptostemmatoidae. Cam-bridge: Cambridge University Press.
Heads M, Craw R. 2004. The Alpine Fault biogeographichypothesis revisited. Cladistics: The International Journalof the Willi Hennig Society 20: 184–190.
Heino J, Soininen J. 2007. Are higher taxa adequate sur-rogates for species-level assemblage patterns and speciesrichness in stream organisms? Biological Conservation 137:78–89.
Hejnol A, Obst M, Stamatakis A, Ott M, Rouse GW,Edgecombe GD, Martinez P, Baguñà J, Bailly X, Jon-delius U, Wiens M, Müller WEG, Seaver E, WheelerWC, Martindale MQ, Giribet G, Dunn CW. 2009. Assess-ing the root of bilaterian animals with scalable phyloge-nomic methods. Proceedings of the Royal Society of LondonSeries B, Biological Sciences 276: 4261–4270.
Hijmans RJ, Cameron SE, Parra JL, Jones PG, Jarvis A.2005. Very high resolution interpolated climate surfaces for
34 G. GIRIBET ET AL.
© 2011 The Linnean Society of London, Biological Journal of the Linnean Society, 2011, ••, ••–••

global land areas. International Journal of Climatology 25:1965–1978.
Juberthie C. 1956. Une nouvelle espèce d’Opilions Sironidaede France et d’Espagne: Parasiro coiffaiti n. sp. Bulletin duMuséum National d’Histoire Naturelle, 2e série 28: 394–400.
Juberthie C. 1958. Révision du genre Parasiro (Opilions,Sironidae) et descriptions de Parasiro minor n. sp. Bulletindu Muséum National d’Histoire Naturelle, 2e série 30: 159–166.
Juberthie C. 1960a. Contribution a l’étude des opilionscyphophthalmes: description de Metasiro gen. n. Bulletin duMuséum National d’Histoire Naturelle, 2e série 32: 235–241.
Juberthie C. 1960b. Sur la biologie d’un opilion endogé, Sirorubens Latr. (Cyphophthalmes). Comptes rendus des séancesde L’Académie des Sciences 251: 1674–1676.
Juberthie C. 1961. Étude des opilions cyphophthalmes(arachnides) du Portugal: description d’Odontosiro lusitani-cus g. n., sp. n. Bulletin du Muséum National d’HistoireNaturelle, 2e série 33: 512–519.
Juberthie C. 1962. Étude des opilions cyphophthalmes Sty-locellinae du Portugal. Description de Paramiopsalis ramu-losus gen. n., sp. n. Bulletin du Muséum National d’HistoireNaturelle, 2e série 34: 267–275.
Juberthie C. 1969. Sur les opilions cyphophthalmes Stylo-cellinae du Gabon. Biologia Gabonica 5: 79–92.
Juberthie C. 1970a. Les genres d’opilions Sironinae(Cyphophthalmes). Bulletin du Muséum National d’HistoireNaturelle, 2e série 41: 1371–1390.
Juberthie C. 1970b. Sur Suzukielus sauteri (Roewer, 1916)opilion cyphophthalme du Japon. Revue d’Écologie et deBiologie du Sol 7: 563–569.
Juberthie C. 1979. Un cyphophthalme nouveau d’une grottede Nouvelle-Caledonie: Troglosiro aelleni n. gen., n. sp.(Opilion, Sironinae). Revue suisse de Zoologie 86: 221–231.
Juberthie C, Manier JF. 1976. Les grands traits de laspermiogenese chez les opilions Comptes Rendus de laTroisième Réunion des Arachnologistes d’ExpressionFrançaise. Station biologique des Eyzies. Les Eyzies:Académie de Paris, 74–82.
Juberthie C, Manier JF. 1978. Étude ultratructurale com-parée de la spermiogenèse des opilions et son intérêt phylé-tique. Symposium of the Zoological Society of London 42:407–416.
Juberthie C, Manier JF, Boissin L. 1976. Étude ultra-structurale de la double espermiogenèse chez l’opilioncyphophthalme Siro rubens Latreille. Journal deMicroscopie et de Biologie Cellulaire 25: 137–148.
Juberthie C, Massoud Z. 1976. Biogéographie, taxonomie etmorphologie ultrastructurale des opilions cyphophthalmes.Revue d’Écologie et de Biologie du Sol 13: 219–231.
Juberthie C, Muñoz-Cuevas A. 1970. Revision [sic] deChileogovea oedipus Roewer (Opiliones: Cyphophthalmi:Sironinae). Senckenbergiana biologica 51: 109–118.
Karaman IM. 2009. The taxonomical status and diversity ofBalkan sironids (Opiliones, Cyphophthalmi) with descrip-tions of twelve new species. Zoological Journal of theLinnean Society 156: 260–318.
Karamata S. 2006. The geological development of the BalkanPeninsula related to the approach, collision and compres-sion of Gondwanan and Eurasian units. In: Robertson AHF,Mountrakis D, eds. Tectonic development of the EasternMediterranean region. London: Geological Society, 155–178.
Knapp M, Mudaliar R, Havell D, Wagstaff SJ, LockhartPJ. 2007. The drowning of New Zealand and the problem ofAgathis. Systematic Biology 56: 862–870.
Kodandaramaiah U. 2011. Tectonic calibrations in molecu-lar dating. Current Zoology 57: 116–124.
Landis CA, Campbell HJ, Begg JG, Mildenhall DC,Paterson AM, Trewick SA. 2008. The WaipounamuErosion Surface: questioning the antiquity of the NewZealand land surface and terrestrial fauna and flora. Geo-logical Magazine 145: 173–197.
Lawrence RF. 1931. The harvest-spiders (Opiliones) ofSouth Africa. Annals of the South African Museum 29:341–508.
Lawrence RF. 1933. The harvest-spiders (Opiliones) ofNatal. Annals of the Natal Museum 7: 211–241.
Lawrence RF. 1939. A contribution to the opilionid fauna ofNatal and Zululand. Annals of the Natal Museum 9: 225–243.
Lawrence RF. 1963. The Opiliones of the Transvaal. Annalsof the Transvaal Museum 24: 275–304.
Legg G. 1990. Parogovia pabsgarnoni, sp. n. (Arachnida,Opiliones, Cyphophthalmi) from Sierra Leone, with notes onother African species of Parogovia. Bulletin of the BritishArachnological Society 8: 113–121.
Legg G, Pabs-Garnon EB. 1989. The life history of a tropi-cal forest cyphophthalmid from Sierra Leone (Arachnida,Opiliones). In: Haupt J, ed. Xi europäisches arachnologis-ches colloquium. Berlin: Technische Universität Berlin,222–230.
Linton EW. 2005. MacGDE: Genetic Data Environment forMacOS X, Version 2.0. Available at: http://www.msu.edu/~lintone/macgde/
Marshall BA. 2011. A new species of Latia Gray, 1850 (Gas-tropoda: Pulmonata: Hygrophila: Chilinoidea: Latiidae)from Miocene Palaeo-lake Manuherikia, southern NewZealand, and biogeographic implications. MolluscanResearch 31: 47–52.
Marske KA, Leschen RAB, Barker GM, Buckley TR.2009. Phylogeography and ecological niche modelling impli-cate coastal refugia and trans-alpine dispersal of a NewZealand fungus beetle. Molecular Ecology 18: 5126–5142.
Márton E, Cosovic V, Moro A, Zvocak S. 2008. The motionof Adria during the Late Jurassic and Cretaceous: newpaleomagnetic results from stable Istria. Tectonophysics454: 44–53.
Murienne J. 2009. Testing biodiversity hypotheses in NewCaledonia using phylogenetics. Journal of Biogeography 36:1433–1434.
Murienne J, Edgecombe GD, Giribet G. 2010a. Includingsecondary structure, fossils and molecular dating in thecentipede tree of life. Molecular Phylogenetics and Evolution57: 301–313.
EVOLUTIONARY AND BIOGEOGRAPHICAL HISTORY OF ARACHNIDS 35
© 2011 The Linnean Society of London, Biological Journal of the Linnean Society, 2011, ••, ••–••

Murienne J, Edgecombe GD, Giribet G. 2011. Compara-tive phylogeography of the centipedes Cryptops pictus andC. niuensis in New Caledonia, Fiji and Vanuatu. OrganismsDiversity & Evolution 11: 61–74.
Murienne J, Giribet G. 2009. The Iberian Peninsula:ancient history of a hot spot of mite harvestmen (Arachnida:Opiliones: Cyphophthalmi: Sironidae) diversity. ZoologicalJournal of the Linnean Society 156: 785–800.
Murienne J, Grandcolas P, Piulachs MD, Bellés X,D’Haese C, Legendre F, Pellens R, Guilbert E. 2005.Evolution of a shaky piece of Gondwana: is local endemismrecent in New Caledonia? Cladistics: The InternationalJournal of the Willi Hennig Society 21: 2–7.
Murienne J, Karaman I, Giribet G. 2010b. Explosive evo-lution of an ancient group of Cyphophthalmi (Arachnida:Opiliones) in the Balkan Peninsula. Journal of Biogeogra-phy 37: 90–102.
Murienne J, Pellens R, Budinoff R, Wheeler WC, Grand-colas P. 2008. Phylogenetic analysis of the endemic NewCaledonian cockroach Lauraesilpha. Testing competinghypotheses of diversification. Cladistics: The InternationalJournal of the Willi Hennig Society 24: 802–812.
Nix HA. 1986. A biogeographic analysis of Australian elapidsnakes. In: Longmore R, ed. Australian flora and faunaseries number 7. Atlas of elapid snakes of Australia. Can-berra: Australian Government Publishing Service, 4–15.
Nixon KC, Carpenter JM. 1996. On simultaneous analysis.Cladistics: The International Journal of the Willi HennigSociety 12: 221–241.
Page TJ, Baker AM, Cook BD, Hughes JM. 2005. Histori-cal transoceanic dispersal of a freshwater shrimp: the colo-nization of the South Pacific by the genus Paratya (Atyidae).Journal of Biogeography 32: 581–593.
Pamic J, Gusic I, Jelaska V. 1998. Geodynamic evolution ofthe Central Dinarides. Tectonophysics 297: 251–268.
Paris JP, Andreieff P, Coudray J. 1979. Sur l’âge Eocènesupérieur de la mise en place de la nappe ophiolitique deNouvelle-Calédonie. Comptes rendus de l’Académie des sci-ences, série B 288: 1659–1661.
Pearson RG, Raxworthy CJ, Nakamura M, Peterson AT.2007. Predicting species distributions from small numbersof occurrence records: a test case using cryptic geckos inMadagascar. Journal of Biogeography 34: 102–117.
Pelletier B. 2006. Geology of the New Caledonia region andits implications for the study of the New Caledonian biodi-versity. In: Payri C, Richer de Forges B, eds. Compendiumof marine species from New Caledonia. Doc. Sci. Tech. IRD.IRD, II 7, 17–30.
Phillips MJ, Gibb GC, Crimp EA, Penny D. 2010.Tinamous and moa flock together: mitochondrial genomesequence analysis reveals independent losses of flightamong ratites. Systematic Biology 59: 90–107.
Phillips SJ, Anderson RP, Schapire RE. 2006. Maximumentropy modeling of species geographic distributions. Eco-logical Modelling 190: 231–259.
Phillips SJ, Dudik M. 2008. Modeling of species distribu-tions with Maxent: new extensions and a comprehensiveevaluation. Ecography 31: 161–175.
Pocock RI. 1897. Descriptions of some new Oriental Opil-iones recently received by the British Museum. Annals andMagazine of Natural History, series 6 19: 283–292.
Poinar G. 2008. Palaeosiro burmanicum n. gen., n. sp., afossil Cyphophthalmi (Arachnida: Opiliones: Sironidae) inEarly Cretaceous Burmese amber. In: Makarov SE, Dimi-trijevic RN, eds. Advances in arachnology and developmen-tal biology. Papers dedicated to Prof. Dr. Bozidar Curcic.Vienna, Belgrade, Sofia: Faculty of Life Sciences, Universityof Vienna, and Serbian Academy of Sciences and Arts,267–274.
Posada D. 2005. Modeltest, Version 3.7. Available at: http://darwin.uvigo.es
Posada D, Crandall KA. 1998. MODELTEST: testing themodel of DNA substitution. Bioinformatics (Oxford,England) 14: 817–818.
Pyron RA. 2011. Divergence time estimation using fossils asterminal taxa and the origins of Lissamphibia. SystematicBiology 60: 466–481.
Rambaut A, Drummond AJ. 2007. Tracer, Version 1.5.Available at: http://beast.bio.ed.ac.uk/Tracer
Rambla M. 1974. Consideraciones sobre la biogeografía de losOpiliones de la Península Ibérica. Miscellanea AlcobéSpecial volume: 45–56.
Rambla M. 1991. A new Stylocellus from some caves ofBorneo, Malaysia (Opiliones, Cyphophthalmi, Stylocellidae).Mémoires de Biospéologie 18: 227–232.
Rambla M. 1994. Un nouveau Cyphophthalme du sud-estasiatique, Fangensis leclerci n. gen. n. sp. (Opiliones,Sironidae). Mémoires de Biospéologie 21: 109–114.
Rambla M, Fontarnau R. 1984. Les Opilions Cyphoph-thalmes (Arachnida) de la faune ibérique: I. Sur Paramiop-salis ramulosus Juberthie, 1962. Revue Arachnologique 5:145–152.
Rambla M, Fontarnau R. 1986. Les Opilions Cyphoph-thalmes (Arachnida) de la faune ibérique: III. Sur Odonto-siro lusitanicus Juberthie, 1961. Mémoirs de la Sociétéroyale belge d’Entomologie 33: 171–178.
Raven PH, Axelrod DI. 1972. Plate tectonics and Australa-sian paleobiogeography. Science 176: 1379–1386.
Ree RH, Moore BR, Webb CO, Donoghue MJ. 2005. Alikelihood framework for inferring the evolution of geo-graphic range on phylogenetic trees. Evolution 59: 2299–2311.
Ree RH, Smith SA. 2008. Maximum likelihood inference ofgeographic range evolution by dispersal, local extinction,and cladogenesis. Systematic Biology 57: 4–14.
Roewer CF. 1916. 7 neue Opilioniden des Zoolog. Museumsin Berlin. Archiv für Naturgeschichte 81: 6–13.
Roewer CF. 1961. Opiliones aus Süd-Chile. Senckenbergianabiologica 42: 99–105.
Sanmartín I. 2002. A paleogeographic history of the SouthernHemisphere. Uppsala: Uppsala University.
Sanmartín I, Ronquist F. 2004. Southern hemisphere bio-geography inferred by event-based models: plant versusanimal patterns. Systematic Biology 53: 216–243.
Scheibner C, Speijer RP. 2008. Late Paleocene-earlyEocene Tethyan carbonate platform evolution – a response
36 G. GIRIBET ET AL.
© 2011 The Linnean Society of London, Biological Journal of the Linnean Society, 2011, ••, ••–••

to long- and short-term paleoclimatic change. Earth ScienceReviews 90: 71–102.
Schoener TW. 1968. The Anolis lizards of Bimini: resourcepartitioning in a complex fauna. Ecology 49: 704–726.
Schwendinger PJ, Giribet G. 2005. The systematics of thesouth-east Asian genus Fangensis Rambla (Opiliones :Cyphophthalmi : Stylocellidae). Invertebrate Systematics 19:297–323.
Sharma P, Giribet G. 2005. A new Troglosiro species(Opiliones, Cyphophthalmi, Troglosironidae) from NewCaledonia. Zootaxa 1053: 47–60.
Sharma P, Giribet G. 2006. A new Pettalus species (Opil-iones, Cyphophthalmi, Pettalidae) from Sri Lanka with adiscussion on the evolution of eyes in Cyphophthalmi.Journal of Arachnology 34: 331–341.
Sharma P, Giribet G. 2009a. A relict in New Caledonia:phylogenetic relationships of the family Troglosironidae(Opiliones: Cyphophthalmi). Cladistics: The InternationalJournal of the Willi Hennig Society 25: 279–294.
Sharma P, Karunarathna I, Giribet G. 2009. On theendemic Sri Lankan genus Pettalus (Opiliones, Cyphoph-thalmi, Pettalidae) with the description of a new species anda discussion on the magnitude of its diversity. Journal ofArachnology 37: 60–67.
Sharma PP, Giribet G. 2009b. The family Troglosironidae(Opiliones: Cyphophthalmi) of New Caledonia Zoologia Neo-caledonica 7. Biodiversity Studies in New Caledonia. Paris:196, 83–123.
Sharma PP, Giribet G. 2011. The evolutionary and biogeo-graphic history of the armoured harvestmen – Laniatoresphylogeny based on ten molecular markers, with thedescription of two new families of Opiliones (Arachnida).Invertebrate Systematics 25: 106–142.
Sharma PP, Vahtera V, Kawauchi GY, Giribet G. 2011.Running WILD: the case for exploring mixed parameter setsin sensitivity analysis. Cladistics: The InternationalJournal of the Willi Hennig Society 27: 538–549.
Shear WA. 1977. The opilionid genus Neogovea Hinton, witha description of the first troglobitic cyphophthalmid fromthe western hemisphere (Opiliones, Cyphophthalmi).Journal of Arachnology 3: 165–175.
Shear WA. 1979a. Huitaca ventralis, n. gen., n. sp., with adescription of a gland complex new to cyphophthalmids(Opiliones, Cyphophthalmi). Journal of Arachnology 7: 237–243.
Shear WA. 1979b. Stylocellus sedgwicki n.sp., from PenangIsland, Malaysia (Opiliones, Cyphophthalmida, Stylocel-lidae). Bulletin of the British Arachnological Society 4: 356–360.
Shear WA. 1980. A review of the Cyphophthalmi of theUnited States and Mexico, with a proposed reclassificationof the suborder (Arachnida, Opiliones). American MuseumNovitates 2705: 1–34.
Shear WA. 1985. Marwe coarctata, a remarkable newcyphophthalmid from a limestone cave in Kenya (Arach-nida, Opiliones). American Museum Novitates 2830: 1–6.
Shear WA. 1993a. The genus Chileogovea (Opiliones,
Cyphophthalmi, Petallidae [sic]). Journal of Arachnology21: 73–78.
Shear WA. 1993b. The genus Troglosiro and the new familyTroglosironidae (Opiliones, Cyphophthalmi). Journal ofArachnology 21: 81–90.
Shear WA. 1993c. New species in the opilionid genus Stylo-cellus from Malaysia, Indonesia and the Philippines (Opil-iones, Cyphophthalmi, Stylocellidae). Bulletin of the BritishArachnological Society 9: 174–188.
Shear WA, Gruber J. 1996. Cyphophthalmid opilionids newto Madagascar: two new genera (Opiliones, Cyphophthalmi,?Pettalidae). Bulletin of the British Arachnological Society10: 181–186.
Shultz JW, Regier JC. 2001. Phylogenetic analysis of Pha-langida (Arachnida, Opiliones) using two nuclear protein-encoding genes supports monophyly of Palpatores. Journalof Arachnology 29: 189–200.
Simon E. 1872. Notice sur les Arachnides cavernicoles ethypogés. Annales de la Société entomologique de France, 5esérie 2: 215–244.
Smith SA, Donoghue MJ. 2010. Combining historical bio-geography with niche modeling in the Caprifolium clade ofLonicera (Caprifoliaceae, Dipsacales). Systematic Biology59: 322–341.
de Souza Muñoz ME, De Giovanni R, Ferreira deSiqueira M, Sutton T, Brewer P, Scachetti Pereira R,Llange Canhos DA, Perez Canhos V. 2011. openMod-eller: a generic approach to species’ potential distributionmodelling. Geoinformatica 15: 111–135.
Spagna JC, Álvarez-Padilla F. 2008. Finding an upperlimit for gap costs in direct optimization parsimony. Cladis-tics: The International Journal of the Willi Hennig Society24: 787–801.
Stamatakis A. 2006. RAxML-VI-HPC: maximum likelihood-based phylogenetic analyses with thousands of taxa andmixed models. Bioinformatics (Oxford, England) 22: 2688–2690.
Stamatakis A, Hoover P, Rougemont J. 2008. A rapidbootstrap algorithm for the RAxML Web servers. SystematicBiology 57: 758–771.
Suzuki S, Ohrui M. 1972. Opiliones of the Izu Peninsula,Central Japan. Acta Arachnologica 24: 41–50. [In Japanese.
Thorell T. 1882–1883. Descrizione di alcuni Aracnidi inferioridell’ Arcipelago Malese. Annali del Museo civico di Storianaturale di Genova 18: 21–69.
Thorell T. 1890–1891. Aracnidi di Pinang raccolti nel 1889dai Sig.ri L. Loria e L. Fea. Annali del Museo civico diStoria naturale di Genova (Ser. 2a) 10: 269–383.
Trewick SA, Paterson AM, Campbell HJ. 2007. Hello NewZealand. Journal of Biogeography 34: 1–6.
Varón A, Sy Vinh L, Wheeler WC. 2010. POY version 4:phylogenetic analysis using dynamic homologies. Cladistics:The International Journal of the Willi Hennig Society 26:72–85.
Vlahovic I, Tisljar J, Velic I, Maticec D. 2005. Evolution ofthe Adriatic carbonate platform: palaeogeography, mainevents and depositional dynamics. Palaeogeography Palaeo-climatology Palaeoecology 220: 333–360.
EVOLUTIONARY AND BIOGEOGRAPHICAL HISTORY OF ARACHNIDS 37
© 2011 The Linnean Society of London, Biological Journal of the Linnean Society, 2011, ••, ••–••

Wallis GP, Trewick SA. 2009. New Zealand phylogeography:evolution on a small continent. Molecular Ecology 18: 3548–3580.
Warren DL, Glor RE, Turelli M. 2008. Environmental nicheequivalency versus conservatism: quantitative approachesto niche evolution. Evolution 62: 2868–2883.
Warren DL, Glor RE, Turelli M. 2010. ENMTools: a toolboxfor comparative studies of environmental niche models.Ecography 33: 607–611.
Waters JM, Craw D. 2006. Goodbye Gondwana? NewZealand biogeography, geology, and the problem of circular-ity. Systematic Biology 55: 351–356.
Westwood JO. 1874. Thesaurus entomologicus oxoniensis; or,illustrations of new, rare, and interesting insects, for themost part contained in the collections presented to the Uni-versity of Oxford by the Rev. F.W. Hope . . . with forty platesfrom drawings by the author. Oxford: Clarendon Press.
Wheeler WC. 1995. Sequence alignment, parameter sen-sitivity, and the phylogenetic analysis of molecular data.Systematic Biology 44: 321–331.
Wheeler WC. 1996. Optimization alignment: the end of mul-tiple sequence alignment in phylogenetics? Cladistics: TheInternational Journal of the Willi Hennig Society 12: 1–9.
Wheeler WC. 2005. Alignment, dynamic homology, and opti-mization. In: Albert VA, ed. Parsimony, phylogeny, andgenomics. Oxford: Oxford University Press, 71–80.
Wiens JJ. 2003. Missing data, incomplete taxa, and phylo-genetic accuracy. Systematic Biology 52: 528–538.
Wiens JJ. 2009. Paleontology, genomics, and combined-dataphylogenetics: can molecular data improve phylogeny esti-mation for fossil taxa? Systematic Biology 58: 87–99.
Wortmann UG, Weissert H, Funk H, Hauck J. 2001.Alpine plate kinematics revisited: the Adria problem.Tectonics 20: 134–147.
SUPPORTING INFORMATION
Additional Supporting information may be found in the online version of this article:
Appendix S1. Specimen sampling. Detailed accounts of specimens and collecting data.
Please note: Wiley-Blackwell are not responsible for the content or functionality of any supporting materialssupplied by the authors. Any queries (other than missing material) should be directed to the correspondingauthor for the article.
APPENDIXTable A1. Size of data matrices for each gene before and subsequent to treatment with GBLOCKS
Data partition
Number ofpositions aftertreatment withMUSCLE
Number ofpositions aftertreatment withGBLOCKS
16S rRNA 598 40318S rRNA 1769 176928S rRNA 2267 2016COI 820 657Histone H3 327 327
38 G. GIRIBET ET AL.
© 2011 The Linnean Society of London, Biological Journal of the Linnean Society, 2011, ••, ••–••

Table A2. Lagrange analyses.
Subtree: PettalidaeAreas:(a) South Africa(b) Chile(c) Eastern Australia(d) Western Australia(e) New Zealand, Australian plate(f) New Zealand, Pacific plate(g) Sri Lanka
Geological intervals:(1) 0–35 Ma (disconnection of all landmasses)(2) 35–60 Ma (fragmentation of transantarctic connections between Australian plate and temperate South America)(3) 60–75 Ma (disconnection of Australia and Zealandia)(4) 75–110 Ma (disconnection of South America and West Africa)(5) 110–120 Ma (Sri Lanka + Madagascar + India separated from Africa)(6) 120–167 Ma (East Gondwana separated from West Gondwana)(7) 167–184 Ma (connection of all landmasses)
Subtree: SternophthalmiAreas:(a) Southeast USA(b) Amazonia(c) Tropical West Africa(d) New Caledonia
Geological intervals:(1) 0–35 Ma (disconnection of all three landmasses)(2) 35–45 Ma (submersion of New Caledonia)(3) 45–60 Ma (New Caledonia emergent and disconnected)(4) 60–75 Ma (submersion of New Caledonia)(5) 75–110 Ma (transantarctic connections between the Australian plate and temperate South America; disconnection of South
America and West Africa)(6) 110–206 Ma (connection of all landmasses)
Subtree: BoreophthalmiAreas:(a) Thai-Malay Peninsula(b) Eastern Himalayas(c) Borneo(d) Indo-Malay Archipelago(e) North America(f) Western Europe(g) Mediterranean(h) Balkans(i) Iberia(j) Japan
Geological intervals0–35 Ma (separation of Mediterranean plate from Western Europe; separation of Japan from Eurasia; connection of Iberia to Eurasia)35–45 Ma (separation of Borneo and Indo-Malay Archipelago from Eurasia)45–60 Ma (Balkans connected to Western Europe; Iberia connected to Mediterranean plate, Balkans and Japan)60–75 Ma (Iberia separated from Mediterranean plate, Balkans and Japan; North America separated from Western Europe;
emergence of Indo-Malay Archipelago)75–110 Ma (Mediterranean plate separated from North America; Iberia connected to western Laurasia; Balkans separated from North
America and Western Europe)110–120 Ma (Iberia disconnected from other landmasses; Western Europe, Mediterranean plate and North America separated from
Eastern Laurasia; emergence of Borneo; Indo-Malay Archipelago nonexistent)120–180 Ma (Iberia disconnected from other landmasses; Western Europe, Mediterranean plate and North America separated from
Eastern Laurasia; Borneo and Indo-Malay Archipelago nonexistent)180–250 Ma (Thai-Malay Peninsula disconnected from other landmasses; Eastern Himalayas disconnected from North America,
Western Europe and Iberia; Borneo and Indo-Malay Archipelago nonexistent)250–296 Ma (Borneo and Indo-Malay Archipelago nonexistent; other landmasses connected)
EVOLUTIONARY AND BIOGEOGRAPHICAL HISTORY OF ARACHNIDS 39
© 2011 The Linnean Society of London, Biological Journal of the Linnean Society, 2011, ••, ••–••


















