
European Law and Italy’s Tax Shield - European Policy Forum
14
858 J. N. Am. Benthol. Soc., 2005, 24(4):858–871 q 2005 by The North American Benthological Society Chimney construction by Chironomus riparius larvae in response to hypoxia: microbial implications for freshwater sediments PETER STIEF 1 Max Planck Institute for Marine Microbiology, Celsiusstrasse 1, 28359 Bremen, Germany LARISA NAZAROVA 2 Ecological Faculty, Kazan State University, Kremlyevskaya Str. 18, 420008 Kazan, Russia DIRK DE BEER 3 Max Planck Institute for Marine Microbiology, Celsiusstrasse 1, 28359 Bremen, Germany Abstract. Many shallow aquatic ecosystems with high nutrient loads experience periods of O 2 depletion that evoke behavioral responses by macrobenthic organisms. The sediment-dwelling midge larva Chironomus riparius reduces its deposit-feeding activity and allocates more time to burrow ven- tilation during periods of hypoxia. We investigated another striking behavioral adaptation of this species, i.e., the elongation of U-shaped sediment burrows to chimneys that tower above the sediment surface. Chironomus riparius larvae gradually abandoned burrow construction and took up chimney construction when exposed to hypoxic conditions in laboratory microcosms. Microsensors were used to show that the chimneys were oxic sediment compartments that were periodically irrigated by the larvae with oxygenated surface water. O 2 uptake rates per unit interface area were significantly higher for chimneys than for the flat sediment surface. This observation was consistent with the dense colonization of the chimneys by bacteria. Chimneys may facilitate the larval acquisition of both O 2 for respiration and microbial biomass for food. Given the mass abundance of C. riparius in many polluted and O 2 -deficient habitats, the chimneys also may contribute significantly to the patchiness of the benthic microbial community in terms of structure and function. In particular, the presence of chimneys might favor aerobic bacterial populations and their metabolism. Key words: Chironomidae, macrofauna, freshwater sediment, oxygen depletion, biogenic structure, sediment bacteria, diffusive oxygen uptake, animal–microbe interaction, sediment microcosm, micro- sensor. Periods of O 2 depletion are common in many aquatic ecosystems, including coastal upwelling regions (Levin 2003), fjords (Rosenberg et al. 2001), streams receiving sewage water (Hell- mann 1994), and lake hypolimnia during tem- perature stratification (Hamburger et al. 2000). Common features of ecosystems susceptible to hypoxia or anoxia are high nutrient concentra- tions, extensive organic sedimentation, and poor exchange of bottom water (Gray et al. 2002, Lev- in 2003). High nutrient concentrations can be of natural origin (e.g., coastal upwelling regions, Bru ¨chert et al. 2003), or they can result from anthropogenic eutrophication and pollution 1 Present address: Department of Microbiology, In- stitute of Biological Sciences, Aarhus University, Ny Munkegade, Building 540, 8000 Aarhus C, Denmark. E-mail: [email protected] 2 E-mail addresses: [email protected] 3 [email protected] (e.g., in streams, ponds, and lakes, Hellmann 1994, Hamburger et al. 2000). Benthic macro- and microorganisms must cope with the consequences of hypoxic events, including shortage of O 2 and accumulation of toxic substances (e.g., sulfide). Macrofauna may have physiological adaptations to these condi- tions (e.g., hemoglobin, Choi et al. 2000; anaero- biosis, Hamburger et al. 2000), or they may re- spond to hypoxia with changes in behavior. Ventilation of sediment burrows is intensified in shallow waters (Leuchs 1986, Heinis and Crom- mentuijn 1992), bioturbation activity is reduced (Levin 2003), and sediment burrows are elon- gated above the sediment surface (Kon and Hi- daka 1983, Jørgensen and Revsbech 1985). How- ever, stress avoidance and behavioral compen- sation are of little importance to sediment bac- teria (except for mobile taxa). Instead, external conditions regulate the metabolism of bacteria
Transcript of European Law and Italy’s Tax Shield - European Policy Forum

858
J. N. Am. Benthol. Soc., 2005, 24(4):858–871q 2005 by The North American Benthological Society
Chimney construction by Chironomus riparius larvae in response tohypoxia: microbial implications for freshwater sediments
PETER STIEF1
Max Planck Institute for Marine Microbiology, Celsiusstrasse 1, 28359 Bremen, Germany
LARISA NAZAROVA2
Ecological Faculty, Kazan State University, Kremlyevskaya Str. 18, 420008 Kazan, Russia
DIRK DE BEER3
Max Planck Institute for Marine Microbiology, Celsiusstrasse 1, 28359 Bremen, Germany
Abstract. Many shallow aquatic ecosystems with high nutrient loads experience periods of O2
depletion that evoke behavioral responses by macrobenthic organisms. The sediment-dwelling midgelarva Chironomus riparius reduces its deposit-feeding activity and allocates more time to burrow ven-tilation during periods of hypoxia. We investigated another striking behavioral adaptation of thisspecies, i.e., the elongation of U-shaped sediment burrows to chimneys that tower above the sedimentsurface. Chironomus riparius larvae gradually abandoned burrow construction and took up chimneyconstruction when exposed to hypoxic conditions in laboratory microcosms. Microsensors were usedto show that the chimneys were oxic sediment compartments that were periodically irrigated by thelarvae with oxygenated surface water. O2 uptake rates per unit interface area were significantly higherfor chimneys than for the flat sediment surface. This observation was consistent with the densecolonization of the chimneys by bacteria. Chimneys may facilitate the larval acquisition of both O2
for respiration and microbial biomass for food. Given the mass abundance of C. riparius in manypolluted and O2-deficient habitats, the chimneys also may contribute significantly to the patchinessof the benthic microbial community in terms of structure and function. In particular, the presence ofchimneys might favor aerobic bacterial populations and their metabolism.
Key words: Chironomidae, macrofauna, freshwater sediment, oxygen depletion, biogenic structure,sediment bacteria, diffusive oxygen uptake, animal–microbe interaction, sediment microcosm, micro-sensor.
Periods of O2 depletion are common in manyaquatic ecosystems, including coastal upwellingregions (Levin 2003), fjords (Rosenberg et al.2001), streams receiving sewage water (Hell-mann 1994), and lake hypolimnia during tem-perature stratification (Hamburger et al. 2000).Common features of ecosystems susceptible tohypoxia or anoxia are high nutrient concentra-tions, extensive organic sedimentation, and poorexchange of bottom water (Gray et al. 2002, Lev-in 2003). High nutrient concentrations can be ofnatural origin (e.g., coastal upwelling regions,Bruchert et al. 2003), or they can result fromanthropogenic eutrophication and pollution
1 Present address: Department of Microbiology, In-stitute of Biological Sciences, Aarhus University, NyMunkegade, Building 540, 8000 Aarhus C, Denmark.E-mail: [email protected]
2 E-mail addresses: [email protected] [email protected]
(e.g., in streams, ponds, and lakes, Hellmann1994, Hamburger et al. 2000).
Benthic macro- and microorganisms mustcope with the consequences of hypoxic events,including shortage of O2 and accumulation oftoxic substances (e.g., sulfide). Macrofauna mayhave physiological adaptations to these condi-tions (e.g., hemoglobin, Choi et al. 2000; anaero-biosis, Hamburger et al. 2000), or they may re-spond to hypoxia with changes in behavior.Ventilation of sediment burrows is intensified inshallow waters (Leuchs 1986, Heinis and Crom-mentuijn 1992), bioturbation activity is reduced(Levin 2003), and sediment burrows are elon-gated above the sediment surface (Kon and Hi-daka 1983, Jørgensen and Revsbech 1985). How-ever, stress avoidance and behavioral compen-sation are of little importance to sediment bac-teria (except for mobile taxa). Instead, externalconditions regulate the metabolism of bacteria

2005] 859CHIRONOMID RESPONSE TO HYPOXIA
from full activity to dormancy. On the micro-scale, sediments can be considered model habi-tats for studying the bacterial response to hyp-oxia and anoxia because sharp O2 gradients de-termine the metabolic activity of aerobic and an-aerobic bacteria within the sediment (Wieringaet al. 2000, Altmann et al. 2003).
Here we present an example of the behavioralresponse of an abundant macrofaunal organismto hypoxic conditions and the implications ofthis behavior for bacterial colonization patternsin freshwater sediments. Chironomus riparius, anonbiting midge with aquatic larval stages, is atypical inhabitant of polluted and temporarilyO2-deficient shallow waters (Pinder 1986, Bair-lein 1989, Penttinen and Holopainen 1995). Thelarvae regularly occur in masses, construct U-shaped burrows within soft sediments, and feedon the detritus particles and the attached bac-teria that are deposited onto the sediment sur-face (Pinder 1986). Chironomus riparius larvae tol-erate O2 deficiency because their hemolymphcontains hemoglobin with a high affinity for O2
(Choi et al. 2000). The larvae also can switch toanaerobiosis, but they are less efficient than lar-vae of other species (e.g., Chironomus anthracinus,Penttinen and Holopainen 1995, Hamburger etal. 2000). Chironomus riparius larvae increasetheir undulation activity during periods of lowO2 concentrations in the water column (Leuchs1986). Occasionally, the larvae elongate theirburrows to reach above the sediment surface, abehavior described as chimney-projecting behavior(Kon and Hidaka 1983, von Danwitz 1983). Bio-genic structures that pierce through the diffu-sive boundary layer are thought to improve O2
availability for the inhabiting animal (Jørgensenand Revsbech 1983). Projecting biogenic struc-tures also may generate small-scale habitat het-erogeneity for the benthic microbial communityby promoting advective nutrient transport andtrapping organic particles (Eckman and Nowell1984, Eckman 1985, Eckman and Thistle 1991,Soltwedel and Vopel 2001).
Our study addressed the following questions:1) Does a threshold O2 concentration induce thechimney-projecting behavior at the expense ofsediment burrow construction or is the responseto increasing hypoxia graded? 2) Does the erectshape of chimneys make them a residual oxicsediment compartment under hypoxic condi-tions? 3) Are chimneys preferential sites for bac-terial colonization? 4) How does sedimentary O2
uptake vary between inhabited and uninhabitedsediments and the chimneys? Laboratory micro-cosm experiments were conducted using ho-mogenized freshwater sediments that were ex-posed to different O2 concentrations (factor 1)and to which C. riparius larvae (factor 2) wereadded or not. The number and size of burrowsand chimneys were quantified, total bacterialabundance was determined, and O2 concentra-tions were measured on a microscale in all treat-ment combinations.
Methods
Origin of sediment and animals
Sediment was sampled in a freshwater ditch;20 km west of Koln (North-Rhine Westphalia,Germany). The ditch runs through an intenselyfarmed area. The water column is characterizedby high nutrient concentrations and transient O2
depletions. Large populations of various Chiron-omus species inhabit the silty and organic-richsediment of the ditch. A flat shovel (10 3 15 cm)was pushed 10 cm into the sediment and theload of sediment was lifted carefully out of itsplace. Five loads were placed in a 10-L plasticbucket and covered with 2 L of ditch water. Thesediment was sieved through a 1-mm-meshscreen, homogenized, and stored at 48C in thedark until use. Egg masses of C. riparius wereprovided by the Institute for Inland Water Man-agement and Waste Water Treatment (RIZA) inLelystad, The Netherlands. Larvae were bred inlaboratory tanks (40 3 50 3 15 cm) filled withaerated, aged tap water and a 5-mm thick layerof sediment. A food suspension of groundleaves of stinging nettle (Urtica sp.) and fishfood (Trouvit, Tetramint) was added 3 times aweek. The culture was kept at 208C under alight:dark cycle of 16:8 h. Chironomus riparius lar-vae grew to a body length of 5 to 7 mm within10 to 15 d (3rd-instar larvae) and were thenready for use in the hypoxia experiments. Agroup of larvae was tested for its sensitivity toa standard toxicant (K2Cr2O7, Grootelaar et al.1996) before the actual experiment. The sensitiv-ity of larvae taken from our culture lay withinthe frame of acceptability for use in laboratoryexposure experiments.
Laboratory microcosms
Sediment was added to the center compart-ment (10 3 5 3 5 cm) of transparent aquaria

860 [Volume 24P. STIEF ET AL.
FIG. 1. Microcosms used for hypoxia experiments (side view). Numbered parts are acrylic aquarium (1), O2
macroelectrode (2), oxymeter (3), O2 control unit (4), magnetic valve (5), gas dispenser (6), stir bar (7), andmagnetic stirrer (8).
(20 3 5 3 10 cm; Fig. 1). The remaining volumeof the aquarium was filled with aged tap water.Two stir bars, rotating on either side of the cen-tral sediment compartment, homogenized thewater. The O2 content of the water body wasregulated using an O2 control unit (Aquastar V2.0, iks ComputerSysteme GmbH, Karlsbad,Germany). The O2 concentration of the waterwas measured next to the sediment compart-ment with a calibrated macrosensor. A magneticvalve opened automatically and N2 gas wasblown into the water via a gas dispenser whenO2 concentrations were above the selected value.The valve closed as soon as the selected valuewas reached. The O2 concentration (expressed as% air saturation) of the water could be held con-stant with an accuracy of 5%.
Experimental schedule
Experiments were carried out in 6 consecutiveruns. Three runs with chironomid larvae (ani-mal treatments) and 3 runs without larvae (con-
trol treatments) were done in random order.Each run began with a 2-wk preincubation pe-riod during which 4 aquaria were filled withsediment and allowed to equilibrate at 100% airsaturation and 158C in the dark. After this pre-incubation period, fifty 3rd-instar larvae (1 lar-va/cm2) were added to each aquarium in ani-mal treatment runs. No larvae were added toaquaria in control treatment runs. Chironomus ri-parius typically dug into the sediments in ,30min. The O2 content of the aquaria was adjustedafter 2 d of acclimation. One aquarium was keptat 100% air saturation, and one each was kept at45, 30, or 15% air saturation. Larvae were fedevery 3rd day with 5 mL of food suspension.Aquarium water was replaced the day afterfood addition to prevent accumulation of detri-mental NO2
2 concentrations. Daily checks of theoverlying water showed that NO2
2 concentra-tions remained well below the tolerance level forthe larvae (20 mmol/L; Neumann et al. 2001) atall times. All manipulations carried out in ani-mal treatments also were carried out in control

2005] 861CHIRONOMID RESPONSE TO HYPOXIA
FIG. 2. Representative positions of O2 microsensor during measurements for O2 concentration profiles nearburrows and chimneys constructed by Chironomus riparius larvae in hypoxia experiments. Letters refer to probepositions when measuring vertical profiles across the sediment–water interface (A), profiles within the diffusiveboundary layer surrounding a chimney (B), and time-series profiles at the opening of a chimney (C). Repre-sentation of the chimney (C) is a simplification: chimney is actually an extension of the U-shaped burrow.Microsensor drawings are not to scale.
treatments. Larval performance, O2 uptake ratesof sediments and chimneys, and total bacterialabundance in sediments and chimneys werestudied during the 3-wk experimental incuba-tion period after the addition of larvae.
Larval performance
The 2 types of larval tubes (burrows andchimneys) were quantified in terms of totalnumber and average size (length, diameter, andthickness of walls) on day 14 of the experimen-tal incubations. A cylindrical shape was as-sumed for the chimneys, and the dimensionswere used to calculate the relative gain in sed-iment–water interface area and oxic sedimentvolume. A few C. riparius larvae left the sedi-
ments and were found in the water on eitherside of the sediment compartment. These larvaewere regarded as having escaped from adverseconditions in the sediment, and their numberwas recorded during daily inspections. Deadlarvae and pupae and the exuviae left behindfrom emerged animals were collected andcounted. The cumulative % of escaped, dead,and emerged animals was calculated over theduration of the experiments. The periodic un-dulation behavior of the larvae inside the bur-rows was verified qualitatively with observa-tions through the transparent aquarium wall.Larval undulation inside chimneys was docu-mented in the form of fluctuating O2 concentra-tions near the chimney opening (Fig. 2C). Mi-crosensors (see below) were used to make sev-

862 [Volume 24P. STIEF ET AL.
eral 240-min time-series recordings (temporalresolution 5 3 s) of O2 concentrations near chim-ney openings.
O2 uptake rates
Microsensors were used to quantify the dif-fusive O2 uptake by sediments and chimneysduring weeks 2 and 3 of the experimental in-cubations. O2 microsensors were constructed,calibrated, and used for concentration profilingas described previously (Revsbech 1989, Stiefand de Beer 2002). The sensors had tip diame-ters of 10 to 20 mm and allowed measurementswith a spatial resolution of 50 to 200 mm. Sensortips were positioned relative to the sediment orchimney surface (Fig. 2) with a dissection mi-croscope and a motorized micromanipulatorthat drove the sensors stepwise into the sedi-ments or towards the chimney surface. Sensorsignals were logged on a computer using theprogram InSight (R. Thar, University of Copen-hagen, Denmark). Diffusive O2 uptake by thesediments and chimneys was calculated fromlinear concentration gradients within theboundary layers covering their surfaces usingFick’s 1st Law of Diffusion. The diffusion coef-ficient of O2 at 158C was taken as 1.83 3 1025
cm2/s (Broecker and Peng 1974). O2 micropro-files were repeated at as many different posi-tions as possible to average out lateral hetero-geneities of the samples. H2S and total S22 con-centrations and pH were measured using mi-crosensors (Revsbech et al. 1983, Jeroschewski etal. 1996) in 100% and 15% air saturation treat-ments.
Bacterial abundance
Total bacterial abundance on sediments andchimneys was measured at the end of each ex-periment. Up to three 2.5-cm-diameter sedimentcores were taken from each aquarium. Sedimentcores were positioned to avoid inclusion ofchimneys in the sample. A 2-mm-thick sedimentslice (i.e., the feeding layer of C. riparius larvae,Stief and de Beer 2002) was removed from thetop of each core and fixed in 4% paraformal-dehyde for 1 h. Chimneys .3 mm in lengthwere removed as whole units using tweezersand fixed. Fixed samples were washed 33 inphosphate-buffered saline (3 mmol/L NaH2PO4,7 mmol/L Na2HPO4 [pH 7.2], 130 mmol/L
NaCl) and stored in 3 volumes of a 1:1 mixtureof phosphate-buffered saline and 96% ethanol at2208C. Samples were diluted 1003 with phos-phate-buffered saline and sonicated with anUW70 probe (Bandelin Electronic GmbH, Ber-lin, Germany) at a setting of 3 3 60 s, 20% pulse,and 109 mm amplitude. A volume of 75 mL ofthe sonicated dilutions was immobilized onblack polycarbonate membrane filters (pore size5 0.2 mm; Osmonics, Minnetonka, Minnesota).Cells were stained with 49,6-diamino-2-pheny-lindole (DAPI) at a concentration of 0.5 mg/mLfor 5 to 10 min. Bacterial cells were counted in20 to 30 randomly chosen microscopic fields oneach filter, giving a total of 700 to 1500 cellscounted/filter. Cell counts were related to thewet volumes of the sediment slices and thechimneys. In treatments with larvae, the chim-ney dimensions were used to calculate the rel-ative increase in oxic sediment volume (seeabove) and the relative increase in the numberof bacteria in the oxic sediment volume.
Statistical analysis
One-way analysis of variance (ANOVA) wasused to identify significant effects of % air sat-uration on the number of burrows and chim-neys built by the larvae. Each experimental rundelivered a single number of burrows and chim-neys in each % air saturation treatment and wastreated as a replicate. Homogeneity of varianceswas checked with Levene’s test. Scheffe andGames–Howell post hoc tests were run for pair-wise comparisons when variances were homog-enous or nonhomogenous, respectively. Two-way ANOVA was used to account for the blockdesign of the experiment whenever multiple val-ues were obtained in a single treatment (i.e.,depth of burrows, height of chimneys, O2 up-take rates, and bacterial numbers). In these cas-es, the experimental run (block) was used as a2nd fixed factor for the analysis. Significant in-teractions between block and % air saturationwere not detected. The dependence of cumula-tive percentages of escaping, emerging, and dy-ing larvae on % air saturation was analyzed us-ing a x2 test. The dependence of the % gain inbacterial cell abundance in the oxic sedimentvolume of animal treatments on % air saturationalso was analyzed using a x2 test. Multiple pair-wise comparisons were made, so a (5 0.05) wasBonferroni-corrected (a/6 5 0.0083).

2005] 863CHIRONOMID RESPONSE TO HYPOXIA
Results
Larval performance
Burrows and chimneys were observed insideand on top of the sediments, respectively, with-in 24 h after introducing the larvae into theaquaria. Surface sculpturing of the sediment bythe larvae was fully developed after 2 wk. Someof the burrows were filled with sediment andsome of the chimneys had disappeared by the3rd week of the experiments. Therefore, the 2types of tubes were routinely quantified on day14 of all experimental runs (Fig. 3). The numberof U-shaped burrows was not affected by % airsaturation (ANOVA, F3,8 5 0.710, p 5 0.573),whereas the number of chimneys increased sig-nificantly as % air saturation decreased (ANO-VA, F3,8 5 6.306, p 5 0.017; Fig. 3A). The numberof chimneys was significantly greater at 15%than at 100% air saturation (Scheffe, p 5 0.020).
The depth of burrows and height of chimneysvaried significantly with % air saturation (AN-OVA, burrows: F3,140 5 6.542, p , 0.0001, chim-neys: F3,192 5 7.540, p , 0.0001; Fig. 3B). Burrowswere significantly less deep under all hypoxicconditions than at 100% air saturation (Games–Howell, 45% air saturation: p 5 0.050, 30% airsaturation: p 5 0.003, and 15% air saturation: p5 0.002). Chimneys built at 15% air saturationwere significantly taller than those built at allother air saturations (Games–Howell, 30% airsaturation: p 5 0.013, 45% air saturation: p 50.001, and 100% air saturation: p 5 0.035). At100% air saturation, burrows reached throughthe black, sulfidic sediment layer (located at adepth of 4–7 mm) and extended down to a max-imum depth of 25 mm. Under all hypoxic con-ditions, the deepest point of the burrows gen-erally was in the sulfidic sediment layer, i.e., theburrows never reached .7 mm into the sedi-ment. Chimneys did not persist in their size andshape over the 3 wk of the experiment. Instead,some of them decreased in height or disap-peared completely only a few days after theirconstruction. However, new chimneys were con-structed until the end of the experimental in-cubation. Whether the larvae destroyed or atethe chimneys and what the larvae did after theremoval of the chimneys could not be deter-mined.
The cumulative proportion of escaped larvaewas independent of % air saturation (x2 5 4.555,
df 5 3, p 5 0.207). At $30% air saturation, onlya small number of larvae (,10%) escaped fromthe sediment compartment (Table 1). At 15% airsaturation, the cumulative proportion of es-caped larvae was 23.6%. Emergence of adultmidges was independent of % air saturation (x2
5 0.317, df 5 3, p 5 0.957) and was ,13.1% inall % air saturation treatments. Mortality wasindependent of % air saturation (x2 5 5.827, df5 3, p 5 0.120). Mortality was ;25% at 45% and100% air saturation, ;38% at 30% air saturation,and ;18% at 15% air saturation.
Spurts of larval undulation in burrows oc-curred every 3 to 4 min and lasted for ;20 seach (PS, unpublished data). Spurts of undula-tion in chimneys, inferred from O2 concentrationmeasurements made at the opening of the chim-neys, were less frequent (every 7–8 min) butlasted longer (3–6 min). A current of oxygenat-ed water was drawn into the chimney duringeach ventilation spurt, whereas anoxic condi-tions prevailed at the chimney opening duringresting periods (Fig. 4).
O2 uptake rates
Vertical O2 concentration profiles were mea-sured across the sediment–water interface (Fig.5A, B), and O2 microdistribution profiles weremeasured around chimneys (Fig. 5C). Theseprofiles were used to calculate the oxic sedimentvolume (Table 2) and the diffusive O2 uptakerates by sediments and chimneys (Fig. 6). O2
penetration into the sediments decreased withdecreasing % air saturation in the water column,but O2 penetration was not affected by the pres-ence or absence of larvae (Fig. 5A, B). The oxicsediment volumes were 2200, 800, 800, and 300cm3/m2 for the 100%, 45%, 30%, and 15% airsaturation treatments, respectively. The pres-ence of chimneys increased the oxic sedimentvolume substantially (Table 2). The diffusive O2
uptake rates by the sediments decreased signif-icantly with decreasing O2 concentration in thewater column (ANOVA, control treatment: F3,17
5 19.887, p , 0.0001, animal treatment: F3,75 5108.213, p , 0.0001; Fig. 6). Diffusive O2 uptakerates by the chimneys also were significantly in-fluenced by % air saturation (ANOVA, F2,20 516.829, p , 0.0001; Fig. 6). Rates were the sameat 45% and 30% air saturation, but rates weresignificantly lower at 15% than at 30% and 45%air saturation (Scheffe, 30% air saturation: p ,

864 [Volume 24P. STIEF ET AL.
FIG. 3. Mean (11 SE) number (A) and depths and heights (B) of burrows and chimneys of Chironomusriparius larvae. Within each tube type, columns that share the same letter are not significantly different.
0.0001, 45% air saturation: p 5 0.007). Moreover,within each air saturation treatment, the diffu-sive O2 uptake per unit interface area was al-ways significantly higher for chimneys than forthe sediment surface (control and animal treat-ments) (ANOVA, 45% air saturation: F2,22 5
4.412, p 5 0.024, 30% air saturation: F2,25 532.112, p , 0.0001, and 15% air saturation: F2,38
5 11.112, p , 0.0001).The representative H2S measurements in the
100% and 15% air saturation treatments showedthat free H2S was not present in the sediments

2005] 865CHIRONOMID RESPONSE TO HYPOXIA
TABLE 1. Mean (61 SE) % of larvae added to microcosms that escaped, emerged, or died in laboratorymicrocosms under different O2 (% air saturation) conditions. Treatments sharing the same letter within a columnwere not significantly different.
% air saturation
Cumulative %
Escaped larvae Emerged larvae Dead larvae and pupae
100 7.3 6 2.0a 10.7 6 5.9a 27.3 6 8.9a
45 7.5 6 3.5a 11.8 6 7.9a 25.3 6 8.3a
30 9.4 6 5.0a 13.1 6 7.1a 37.7 6 12.7a
15 23.6 6 12.3a 10.5 6 6.0a 18.2 6 9.0a
FIG. 4. A.—Time series of O2 concentrations at the opening of a chimney in a 15% air saturation (;40 mmolO2/L) treatment. The O2 microsensor was positioned level with the rim of the 5-mm-high chimney (Fig. 2C).At the start of each larval ventilation spurt, oxygenated water was drawn into the chimney and passed themicrosensor tip; between spurts, anoxic conditions prevailed at the opening of the chimney. B.—Expansion ofthe temporal scale showing the first 30 min from (A).
or in the water column in animal or controltreatments (data not shown). Vertical profiles ofpH typically showed a decrease from pH 8.3 inthe water column to pH 7.5 in the sediments(data not shown).
Bacterial abundance
Bacterial abundance in the sediment de-creased significantly with decreasing % air sat-uration in control treatments (ANOVA, F3,8 5
8.874, p 5 0.006; Fig. 7), whereas bacterial abun-dance in the sediment increased significantlywith decreasing % air saturations in the animaltreatments (ANOVA, F3,35 5 7.668, p , 0.0001).Percent air saturation did not affect total bacte-rial counts in chimneys (ANOVA, F2,22 5 2.569,p 5 0.099). However, total bacterial counts weresignificantly higher in chimneys (animal treat-ments) than in sediments (control treatments) at30% and 15% air saturation (ANOVA, 30% airsaturation: F2,16 5 10.640, p 5 0.001, 15%: F2,18 5

866 [Volume 24P. STIEF ET AL.
FIG. 5. Mean (61 SE) O2 concentrations across the sediment–water interface in control (A) and animaltreatments (B) and toward the surfaces of chimneys in hypoxic animal treatments (C). Dotted lines in A andB indicate the sediment–water interface. Dotted line in C indicates the surface of a chimney.
TABLE 2. Mean (61 SE) % changes in sediment O2 dynamics and bacterial distribution associated with thepresence of chironomid chimneys in laboratory microcosms. Percent air saturation treatments sharing the sameletter were not significantly different.
% air saturation
Chimney-related gain (%)
Sediment–waterinterface area
Oxic sedimentvolume
Bacterial abundance in oxic sediment(relative to sediment bacteria
in animal treatment)
100 2.3 6 0.3 0.5 6 0.1 0.0 6 0.0a
45 11.8 6 2.2 7.4 6 1.4 7.4 6 1.2b
30 8.3 6 3.0 11.4 6 1.9 14.6 6 1.8b
15 31.5 6 7.7 52.4 6 12.8 57.5 6 5.0c
9.120, p 5 0.002). Moreover, chimneys increasedthe abundance of bacteria in oxic sediment ofthe animal treatments (Table 2). The relativegain was dependent on air saturation (x2 test, x2
5 28.39, df 5 3, p , 0.0001; Table 2).
Discussion
Importance of chimneys for larvae
Chironomus riparius larvae switched graduallyfrom burrow to chimney construction when ex-posed to a range of decreasing % air satura-tions. The burrows inside the sediment becameless deep, whereas the chimneys on top of thesediments became more abundant and taller.However, small chimneys were observed even at100% air saturation. Therefore, we conclude thatC. riparius chimney-projecting behavior has no
threshold % air saturation below which the be-havior is induced (Kon and Hidaka 1983).
The microsensor measurements showed thatthe O2 concentration at the sediment–water in-terface (i.e., at the level of burrow openings) was;50% that of the mixed water column, regard-less of the % air saturation in the water column(Fig. 5A, B). Conversely, the O2 concentration atthe rim of actively irrigated chimneys was aboutthe same as that of the mixed water column (Fig.4). Thus, chimneys pierced through the 400- to600-mm-thick diffusive boundary layer (DBL)and made higher concentrations of O2 availablefor the larvae (Kon and Hidaka 1983, Jørgensenand Revsbech 1985). The apparent O2 concen-tration at the sediment–water interface seemedto determine the size of the chimney to be con-structed. However, whether the larvae respond

2005] 867CHIRONOMID RESPONSE TO HYPOXIA
FIG. 6. Mean (61 SE) O2 uptake rates of sediments and chimneys in laboratory hypoxia experiments. Rateswere calculated from the concentration profiles presented in Fig. 5. Within each sample type, columns thatshare the same letter are not significantly different.
to the actual O2 concentration or to a proxy ofhypoxic conditions (e.g., CO2 or H2S) is unclear.O2 depletion may favor the production of H2Sclose to the sediment surface (Holmer andStorkholm 2001), and H2S could make the larvaeshift their burrows from within the sediment toabove the sediment surface, but the microsensormeasurements showed that H2S was not presentin our sediments (data not shown). CO2 also canbe ruled out as a proximate factor influencingchimney construction in our study because hyp-oxic conditions were achieved artificially byblowing O2 out with N2 gas. Thus, the low O2
concentrations should not have been associatedwith elevated CO2 concentrations.
One might ask why the chimneys were muchhigher (up to 8 mm) than the thickness of theDBL (0.4–0.6 mm), and why their height in-creased with decreasing % air saturation. Thesmallest chimneys found were tall enough topierce through the DBL. One reason could bethat the thickness of the DBL is inversely cor-related with the % air saturation of the watercolumn under natural conditions. Hypoxia nearthe sediment surface typically develops under
stagnant conditions (Gray et al. 2002), and lowflow velocities lead to the expansion of the DBLfrom a few hundred mm to .1 mm (Jørgensenand Revsbech 1985). Thus, chironomid larvaemay somehow associate the perceived % air sat-uration with a certain thickness of DBL to bepenetrated. If so, the inherent response by thelarvae to natural DBL dimensions was meaning-less in our experiments because the DBL alwayshad the same thickness.
Chimneys also may provide larvae with food.Larvae might elongate their burrows above thesediment surface if they were in need of bacte-rial biomass that could be scraped from the bur-row walls (Olafsson and Paterson 2004). Threelines of evidence support this hypothesis. 1) Ex-trapolation of the O2 microprofiles indicated thatthe 500-mm-thick chimney walls were not com-pletely anoxic (Fig. 5C), and the lumens of thechimneys were flushed periodically with oxy-genated water because of larval undulation (Fig.4). Chimneys might allow the attachment andgrowth of aerobic bacteria rather than the an-aerobic bacteria that would grow in less oxy-genated burrows. Aerobic bacteria have shorter
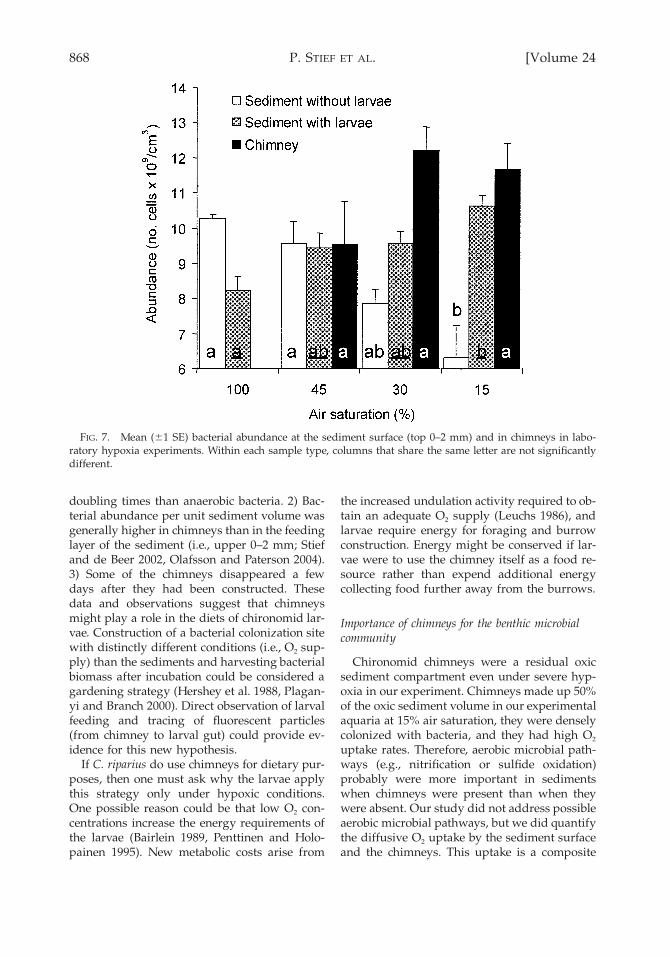
868 [Volume 24P. STIEF ET AL.
FIG. 7. Mean (61 SE) bacterial abundance at the sediment surface (top 0–2 mm) and in chimneys in labo-ratory hypoxia experiments. Within each sample type, columns that share the same letter are not significantlydifferent.
doubling times than anaerobic bacteria. 2) Bac-terial abundance per unit sediment volume wasgenerally higher in chimneys than in the feedinglayer of the sediment (i.e., upper 0–2 mm; Stiefand de Beer 2002, Olafsson and Paterson 2004).3) Some of the chimneys disappeared a fewdays after they had been constructed. Thesedata and observations suggest that chimneysmight play a role in the diets of chironomid lar-vae. Construction of a bacterial colonization sitewith distinctly different conditions (i.e., O2 sup-ply) than the sediments and harvesting bacterialbiomass after incubation could be considered agardening strategy (Hershey et al. 1988, Plagan-yi and Branch 2000). Direct observation of larvalfeeding and tracing of fluorescent particles(from chimney to larval gut) could provide ev-idence for this new hypothesis.
If C. riparius do use chimneys for dietary pur-poses, then one must ask why the larvae applythis strategy only under hypoxic conditions.One possible reason could be that low O2 con-centrations increase the energy requirements ofthe larvae (Bairlein 1989, Penttinen and Holo-painen 1995). New metabolic costs arise from
the increased undulation activity required to ob-tain an adequate O2 supply (Leuchs 1986), andlarvae require energy for foraging and burrowconstruction. Energy might be conserved if lar-vae were to use the chimney itself as a food re-source rather than expend additional energycollecting food further away from the burrows.
Importance of chimneys for the benthic microbialcommunity
Chironomid chimneys were a residual oxicsediment compartment even under severe hyp-oxia in our experiment. Chimneys made up 50%of the oxic sediment volume in our experimentalaquaria at 15% air saturation, they were denselycolonized with bacteria, and they had high O2
uptake rates. Therefore, aerobic microbial path-ways (e.g., nitrification or sulfide oxidation)probably were more important in sedimentswhen chimneys were present than when theywere absent. Our study did not address possibleaerobic microbial pathways, but we did quantifythe diffusive O2 uptake by the sediment surfaceand the chimneys. This uptake is a composite

2005] 869CHIRONOMID RESPONSE TO HYPOXIA
of overall microbial O2 consumption and thechemical re-oxidation of reduced compounds(Canfield et al. 1993). Diffusive O2 uptake by theflat sediment surface decreased with decreasing% air saturation in the water column regardlessof the presence of larvae. This positive correla-tion probably was caused by the decreasing vol-ume of sediment in which oxidative respirationand chemical oxidation could take place (Fig.5A, B). However, chimneys had significantlyhigher diffusive O2 uptake rates than the sedi-ment surface at all % air saturations. The erectshape, hollow structure, and periodic irrigationof the chimneys made these biogenic structuresaccessible to dissolved O2 in the water column.Moreover, the chimneys are likely to be enrichedin easily degradable organics from larval salivaexcretions used for stabilization of the chimney(Leuchs and Neumann 1990) and from trappedsuspended organic particles (Soltwedel and Vo-pel 2001). The dense bacterial colonization onchimneys might have been both a consequenceof O2 and organic substrate availability and acause of the significantly higher diffusive O2 up-take rates of the chimneys than sediment (Eck-man 1985). In any case, chimneys contribute tothe patchy distribution of bacterial biomass andsmall-scale O2 dynamics in the sediment (Eck-man 1985, Soltwedel and Vopel 2001).
Larvae changed their feeding behavior whenexposed to hypoxic conditions. Deposit-feedingat the sediment surface was visibly reduced atlower % air saturations. In normoxic conditions,C. riparius larvae collect and ingest organic par-ticles deposited on the sediment surface (Ras-mussen 1984). Larvae reduce the abundance andmetabolic activity of particle-attached bacteriawithin the feeding layer, and they reduce thediffusive O2 uptake by the sediment (Stief andde Beer 2002, Altmann et al. 2004). At 100% airsaturation in our study, bacterial abundance inthe sediment was lower when larvae were pre-sent than when they were absent (Fig. 7), butthe diffusive O2 uptake rate did not differ be-tween inhabited and uninhabited sediments.The addition of organic food particles to thesediments in both inhabited and uninhabitedsediments may have obscured larval effects onthe metabolic activity of sediment bacteria.
Bacterial abundances in inhabited sedimentsin hypoxic conditions were similar to bacterialabundances in uninhabited sediments in nor-moxic conditions. Larvae did not reduce bacte-
rial abundance in the sediment in hypoxic con-ditions, probably because they showed reducedgrazing at the sediment surface. If reducedgrazing in hypoxic conditions were the only rea-son for similar bacterial abundances in inhabit-ed hypoxic sediment and uninhabited normoxicsediment, then bacterial abundances shouldhave been similar in inhabited and uninhabitedhypoxic sediments. However, under hypoxicconditions, bacterial abundance was lower inuninhabited sediment than in inhabited sedi-ment. Therefore, we assume that physicalchanges in the sediment were induced by thelarvae and contributed to the observed patternof bacterial abundance. Hypoxia causes othersediment-dwelling macrofauna to favor advec-tive over diffusive water exchange with the wa-ter column because of compensatory bioirriga-tion (Forster et al. 1995). This shift also shouldoccur for chironomids that increase their venti-lation activity when exposed to low O2 concen-trations (Leuchs 1986). Advective transport ofO2 and nutrients stimulates the metabolic activ-ity of bacteria that are otherwise diffusion lim-ited (Ziebis et al. 1996). Thus, inhabited sedi-ment may have been a favorable site for bacte-rial colonization because of intense irrigation inthe absence of significant grazing effects.
Altered behavior of C. riparius larvae underhypoxic conditions causes spatial rearrange-ment of sediment bacteria and diffusive O2 up-take by the sediment. Chimney-projecting be-havior creates distinct biogenic structures withhigh bacterial abundance and high diffusive O2
uptake. In contrast, reduced feeding activity un-der hypoxic conditions leads to high bacterialabundance, but low diffusive O2 uptake in thesurficial feeding layer. We hypothesize that theparticular geometry of chironomid chimneys(i.e., an erect and hollow structure that is peri-odically irrigated with oxygenated water) favorsthe growth of metabolically active aerobic bac-teria. Thus, chimneys may not only facilitate thelarval acquisition of both O2 for respiration, butalso of microbial biomass for food.
Acknowledgements
Gaby Eickert and Ines Schroder (Max PlanckInstitute, Bremen, Germany) kindly provided uswith microsensors. We thank Armin Kureck(University of Cologne, Germany) who lent ustechnical equipment and Raphaela Schoon and

870 [Volume 24P. STIEF ET AL.
Sven Schulz who counted bacteria. This studywas supported financially by Deutscher Aka-demischer Austauschdienst, Bonn, Germany,and the Max Planck Society.
Literature Cited
ALTMANN, D., P. STIEF, R. AMANN, AND D. DE BEER.2004. Nitrification in freshwater sediments as in-fluenced by insect larvae: quantification by micro-sensors and fluorescence in situ hybridization.Microbial Ecology 48:145–153.
ALTMANN, D., P. STIEF, R. AMANN, D. DE BEER, AND
A. SCHRAMM. 2003. In situ distribution and activ-ity of nitrifying bacteria in freshwater sediment.Environmental Microbiology 5:798–803.
BAIRLEIN, F. 1989. The respiration of Chironomus larvae(Diptera) from deep and shallow waters underenvironmental hypoxia and at different tempera-tures. Archiv fur Hydrobiologie 115:523–536.
BROECKER, W. S., AND T.-H. PENG. 1974. Gas exchangerates between air and sea. Tellus 26:21–35.
BRUCHERT, V., B. B. JøRGENSEN, K. NEUMANN, D.RIECHMANN, M. SCHLOSSER, AND H. N. SCHULZ.2003. Regulation of bacterial sulfate reductionand hydrogen sulfide fluxes in the central Na-mibian coastal upwelling zone. Geochimica etCosmochimica Acta 67:4505–4518.
CANFIELD, D. E., B. B. JøRGENSEN, H. FOSSING, R.GLUD, J. GUNDERSEN, N. B. RAMSING, B. THAM-DRUP, J. W. HANSEN, L. P. NIELSEN, AND P. O. J.HALL. 1993. Pathways of organic-carbon oxida-tion in 3 continental-margin sediments. MarineGeology 113:27–40.
CHOI, J. H., H. ROCHE, AND T. CAQUET. 2000. Effectsof physical (hypoxia, hyperoxia) and chemical(potassium dichromate, fenitrothion) stress on an-tioxidant enzyme activities in Chironomus ripariusMg. (Diptera, Chironomidae) larvae: potentialbiomarkers. Environmental Toxicology andChemistry 19:495–500.
ECKMAN, J. E. 1985. Flow disruption by an animaltube mimic affects sediment bacterial coloniza-tion. Journal of Marine Research 43:419–435.
ECKMAN, J. E., AND A. R. M. NOWELL. 1984. Boundaryskin friction and sediment transport about an an-imal-tube mimic. Sedimentology 31:851–862.
ECKMAN, J. E., AND D. THISTLE. 1991. Effects of flowabout a biologically produced structure on har-pactoid copepods in San Diego trough. Deep SeaResearch Part A 38:1397–1416.
FORSTER, S., G. GRAF, J. KITLAR, AND M. POWILLEIT.1995. Effects of bioturbation in oxic and hypoxicconditions: a microcosm experiment with a NorthSea sediment community. Marine Ecology Pro-gress Series 116:153–161.
GRAY, J. S., R. S. S. WU, AND Y. Y. OR. 2002. Effects of
hypoxia and organic enrichment on the coastalmarine environment. Marine Ecology ProgressSeries 238:249–279.
GROOTELAAR, E. M. M., J. L. MAASAND, AND C. VAN
DE GUCHTE. 1996. Protocol for testing of substanc-es in chronic sediment bioassays with the fresh-water dipteran Chironomus riparius. Working doc-ument 94.086X, Report No. WSCE 94–05. Divi-sion for Ecotoxicology, Institute for Inland WaterManagement and Waste Water Treatment, Lelys-tad, The Netherlands.
HAMBURGER, K., P. C. DALL, C. LINDEGAARD, AND I.B. NILSON. 2000. Survival and energy metabolismin an oxygen deficient environment. Field andlaboratory studies on the bottom fauna from theprofundal zone of Lake Esrom, Denmark. Hydro-biologia 432:173–188.
HEINIS, F., AND T. CROMMENTUIJN. 1992. Behaviouralresponses to changing oxygen concentrations ofdeposit feeding chironomid larvae (Diptera) of lit-toral and profundal habitats. Archiv fur Hydro-biologie 124:173–185.
HELLMANN, H. 1994. Load trends of selected chemicalparameters of water quality and of trace sub-stances in the River Rhine between 1955 and1988. Water Science and Technology 29:69–75.
HERSHEY, A. E., A. L. HILTNER, M. A. J. HULLAR, M.C. MILLER, J. R. VESTAL, M. A. LOCK, S. RUNDLE,AND B. J. PETERSON. 1988. Nutrient influence on astream grazer: Orthocladius microcommunities re-spond to nutrient input. Ecology 69:1383–1392.
HOLMER, M., AND P. STORKHOLM. 2001. Sulphate re-duction and sulphur cycling in lake sediments: areview. Freshwater Biology 46:431–451.
JEROSCHEWSKI, P., C. STEUCKART, AND M. KUHL. 1996.An amperometric microsensor for the determi-nation of H2S in aquatic environments. AnalyticalChemistry 68:4351–4357.
JøRGENSEN, B. B., AND N. P. REVSBECH. 1983. Colorlesssulfur bacteria, Beggiatoa spp. and Thiovulum spp.,in O2 and H2S microgradients. Applied and En-vironmental Microbiology 45:1261–1270.
JøRGENSEN, B. B., AND N. P. REVSBECH. 1985. Diffusiveboundary layers and the oxygen uptake of sedi-ments and detritus. Limnology and Oceanogra-phy 30:111–122.
KON, M., AND T. HIDAKA. 1983 Chimney projectingbehaviour of chironomid larvae (Chironomus yosh-imatsui; Diptera, Chironomidae). Journal of Ethol-ogy 1:111–113.
LEUCHS, H. 1986. The ventilation activity of Chirono-mus larvae (Diptera) from shallow and deep lakesand the resulting water circulation in correlationto temperature and oxygen conditions. Archiv furHydrobiologie 108:281–299.
LEUCHS, H., AND D. NEUMANN. 1990. Tube texture,spinning and feeding behaviour of Chironomus

2005] 871CHIRONOMID RESPONSE TO HYPOXIA
larvae. Zoologisches Jahrbuch fur Systematik 117:31–40.
LEVIN, L. A. 2003. Oxygen minimum zone benthos:adaptation and community response to hypoxia.Oceanography and Marine Biology 41:1–45.
NEUMANN, D., M. KRAMER, I. RASCHKE, AND B. GRAFE.2001. Detrimental effects of nitrite on the devel-opment of benthic Chironomus larvae, in relationto their settlement in muddy sediments. Archivfur Hydrobiologie 153:103–128.
OLAFSSON, J. S., AND D. M. PATERSON. 2004. Alterationof biogenic structure and physical properties bytube-building chironomid larvae in cohesive sed-iments. Aquatic Ecology 38:219–229.
PENTTINEN, O. P., AND I. J. HOLOPAINEN. 1995. Physi-ological energetics of a midge, Chironomus ripariusMeigen (Insecta, Diptera): normoxic heat outputover the whole life cycle and response of larva tohypoxia and anoxia. Oecologia (Berlin) 103:419–424.
PINDER, L. C. V. 1986. Biology of freshwater Chiron-omidae. Annual Review of Entomology 31:1–23.
PLAGANYI, E. E., AND G. M. BRANCH. 2000. Does thelimpet Patella cochlear fertilize its own algal gar-den? Marine Ecology Progress Series 194:113–122.
RASMUSSEN, J. B. 1984. Comparison of gut contentsand assimilation efficiency of fourth instar larvaeof two coexisting chironomids, Chironomus ripar-ius (Meigen) and Glyptotendipes paripes (Edwards).Canadian Journal of Zoology 62:1022–1026.
REVSBECH, N. P. 1989. An oxygen microsensor with aguard cathode. Limnology and Oceanography 34:474–478.
REVSBECH, N. P., B. B. JøRGENSEN, T. H. BLACKBURN,AND Y. COHEN. 1983. Microelectrode studies ofthe photosynthesis and O2, H2S, and pH profilesof a microbial mat. Limnology and Oceanogra-phy 28:1062–1074.
ROSENBERG, R., H. C. NILSSON, AND R. J. DIAZ. 2001.Response of benthic fauna and changing sedi-ment redox profiles over a hypoxic gradient. Es-tuarine, Coastal and Shelf Science 53:343–350.
SOLTWEDEL, T., AND K. VOPEL. 2001. Bacterial abun-dance and biomass in response to organism-gen-erated habitat heterogeneity in deep-sea sedi-ments. Marine Ecology Progress Series 219:291–298.
STIEF, P., AND D. DE BEER. 2002. Bioturbation effects ofChironomus riparius on the benthic N-cycle asmeasured using microsensors and microbiologi-cal assays. Aquatic Microbial Ecology 27:175–185.
VON DANWITZ, B. 1983. Chironomiden und andereBodentiere eines hochbelasteten Flachsees (Poel-venn, Nettetal). Diploma Thesis, University ofKoln, Koln, Germany.
WIERINGA, E. B. A., J. OVERMANN, AND H. CYPIONKA.2000. Detection of abundant sulphate-reducingbacteria in marine oxic sediment layers by a com-bined cultivation and molecular approach. Envi-ronmental Microbiology 2:417–427.
ZIEBIS, W., M. HUETTEL, AND S. FORSTER. 1996. Impactof biogenic sediment topography on oxygen flux-es in permeable seabeds. Marine Ecology Pro-gress Series 140:227–237.
Received: 11 November 2004Accepted: 2 June 2005


















