
Ethylene: Role in Fruit Abscission and Dehiscence Processes' 2 · Ethylene at 10 microliters per...
6
Plant Physiol. (1972) 50, 759-764 Ethylene: Role in Fruit Abscission and Dehiscence Processes' 2 Received for publication April 28, 1972 JOHN A. LIPE3 AND PAGE W. MORGAN Department of Plant Sciences, Texas A&M University, College Station, Texas 77843 ABSTRACT Two peaks of ethylene production occur during the develop- ment of cotton fruits (Gossypium hirsutum L.). These periods precede the occurrence of young fruit shedding and mature fruit dehiscence, both of which are abscission phenomena and the latter is generally assumed to be part of the total ripening process. Detailed study of the dehiscence process revealed that ethylene production of individual, attached cotton fruits goes through a rising, cyclic pattern which reaches a maximum prior to dehiscence. With detached pecan fruits (Carya illinoensis [Wang.] K. Koch), ethylene production measured on alternate days rose above 1 microliter per kilogram fresh weight per hour before dehiscence began and reached a peak several days prior to complete dehiscence. Ethylene production by cotton and pecan fruits was measured just prior to dehiscence and then the internal concentration of the gas near the center of the fruit was determined. From these data a ratio of production rate to internal concentration was determined which allowed calculation of the approximate ethylene concentration in the intact fruit prior to dehiscence and selection of appropriate levels to apply to fruits. Ethylene at 10 microliters per liter of air appears to saturate dehiscence of cotton, pecan, and okra (Hibiscus esculentus L.) fruits and the process is com- pleted in 3 to 4 days. In all cases some hasteuing of dehiscence was observed with as little as 0.1 microliter of exogenous ethyl. ene per liter of air. The time required for response to different levels of ethylene was determined and compared to the time course of ethylene production and dehiscence. We concluded that internal levels of ethylene rose to dehiscence-stimulating levels a sufficience time before dehiscence for the gas to have initiated the process. Since our data and calculations indicate that enough ethylene is made a sufficient time before dehis- cence, to account for the process, we propose that ethylene is one of the regulators of natural fruit dehiscence, an im- portant component of ripening in some fruits. Our data also suggest a possible involvement of ethylene in young fruit abscission. Ethylene is now implicated as a natural regulator of leaf abscission (2, 11). Young fruit abscission is a common phe- 1This research was supported in part by Cotton Incorporated. A contribution of the Texas Agricultural Experiment Station. 2Preliminary progress reports of this study have appeared in: 1970 Proceedings of the Beltwide Cotton Production Research Conference. p. 47; 1970 Bulletin Southeastern Biologists. 17: 52-53; 1971 Proceedings Association Southern Agricultural Workers 68: 197-198; 1971 Plant Physiol. 47: S14. 3Present address: Texas A&M University Research and Exten- sion Center at Overton, Drawer E, Overton, Tex. 75684. nomena in many plants. Fruit dehiscence, as a separation of tissues, is an abscission-like process. Thus, we concluded that ethylene may be a natural regulator of young fruit abscission and fruit dehiscence. Horticulturists have long known that ethylene hastens fruit wall (shuck, hull, pericarp, and accessory parts) dehiscence of walnut (6, 19) and pecan (9), but until recently there has been only indirect evidence that ethylene is produced by fruits that dehisce. Lipe and Morgan (13) found that ethylene production by pecan fruits increased to a relatively high level shortly be- fore and during fruit wall dehiscence. Similar results have been observed with detached, dehiscing cotton fruits as well (15, 16). In these studies (15, 16), a smaller peak of ethylene production was also noted near the time of maximum young fruit abscission. While fruit dehiscence is generally considered a component of the ripening process, where the regulatory role of ethylene is well established (3, 17), other considerations indicated the need for the present study. Fruit ripening has been studied most extensively in fleshy fruits which do not dehisce, and some fruits dehisce after they are dry while others open before ap- preciable weight loss occurs (13, 16). In some cases, for ex- ample pecans, dehiscence also involves detachment (abscis- sion) of the seed from its vascular connections. Dehiscence as a physiological process has not received detailed attention, and yet it is a critical phase in the production of some crops where it is the dominant visible feature of the ripening process. Natu- ral abscission of whole, ripe fruits of the species studied here is rare; seed dispersal can promptly follow dehiscence. Production of ethylene by flowers has been rather exten- sively studied, primarily in association with petal fading and abscission (3, 17), and there is a presumptive association of ethylene with the natural separation of fruits from plants fol- lowing ripening. All of this information was useful in our ap- proach to the specific process of young fruit abscission. In the present investigation, a detailed monitoring of the seasonal pattern of ethylene production by intact cotton fruits was conducted, ethylene production by dehiscing cotton and pecan fruits was carefully detailed, and the capacity of physio- logical levels of exogenous ethylene to regulate dehiscence of detached fruits was determined. MATERIALS AND METHODS Pattern of Ethylene Production by Developing Cotton Fruits. In preliminary studies (15, 16) we observed ethylene production by detached cotton (Gossypium hirsutum L., cv. SP23) fruits from anthesis until the completion of fruit de- hiscence. Triplicate samples of cotton fruits from a field on the Texas A&M University Farm, tagged on the same day at anthesis. were collected weekly, and enclosed in groups of three in a 500-ml Erlenmeyer flask on water-saturated filter paper. After 3 to 4 hr, the ethylene content of an air sample from each flask was measured gas chromatographically. Ethylene was measured with a hydrogen flame ionization detector (2). Five-milliliter air samples were injected with a 759 www.plantphysiol.org on August 14, 2019 - Published by Downloaded from Copyright © 1972 American Society of Plant Biologists. All rights reserved.
Transcript of Ethylene: Role in Fruit Abscission and Dehiscence Processes' 2 · Ethylene at 10 microliters per...

Plant Physiol. (1972) 50, 759-764
Ethylene: Role in Fruit Abscission and Dehiscence Processes' 2Received for publication April 28, 1972
JOHN A. LIPE3 AND PAGE W. MORGANDepartment of Plant Sciences, Texas A&M University, College Station, Texas 77843
ABSTRACT
Two peaks of ethylene production occur during the develop-ment of cotton fruits (Gossypium hirsutum L.). These periodsprecede the occurrence of young fruit shedding and maturefruit dehiscence, both of which are abscission phenomena andthe latter is generally assumed to be part of the total ripeningprocess. Detailed study of the dehiscence process revealed thatethylene production of individual, attached cotton fruits goesthrough a rising, cyclic pattern which reaches a maximum priorto dehiscence. With detached pecan fruits (Carya illinoensis[Wang.] K. Koch), ethylene production measured on alternatedays rose above 1 microliter per kilogram fresh weight perhour before dehiscence began and reached a peak several daysprior to complete dehiscence. Ethylene production by cottonand pecan fruits was measured just prior to dehiscence andthen the internal concentration of the gas near the center ofthe fruit was determined. From these data a ratio of productionrate to internal concentration was determined which allowedcalculation of the approximate ethylene concentration in theintact fruit prior to dehiscence and selection of appropriatelevels to apply to fruits. Ethylene at 10 microliters per literof air appears to saturate dehiscence of cotton, pecan, andokra (Hibiscus esculentus L.) fruits and the process is com-pleted in 3 to 4 days. In all cases some hasteuing of dehiscencewas observed with as little as 0.1 microliter of exogenous ethyl.ene per liter of air. The time required for response to differentlevels of ethylene was determined and compared to the timecourse of ethylene production and dehiscence. We concludedthat internal levels of ethylene rose to dehiscence-stimulatinglevels a sufficience time before dehiscence for the gas to haveinitiated the process. Since our data and calculations indicatethat enough ethylene is made a sufficient time before dehis-cence, to account for the process, we propose that ethyleneis one of the regulators of natural fruit dehiscence, an im-portant component of ripening in some fruits. Our data alsosuggest a possible involvement of ethylene in young fruitabscission.
Ethylene is now implicated as a natural regulator of leafabscission (2, 11). Young fruit abscission is a common phe-
1This research was supported in part by Cotton Incorporated. Acontribution of the Texas Agricultural Experiment Station.
2Preliminary progress reports of this study have appeared in:1970 Proceedings of the Beltwide Cotton Production ResearchConference. p. 47; 1970 Bulletin Southeastern Biologists. 17: 52-53;1971 Proceedings Association Southern Agricultural Workers 68:197-198; 1971 Plant Physiol. 47: S14.
3Present address: Texas A&M University Research and Exten-sion Center at Overton, Drawer E, Overton, Tex. 75684.
nomena in many plants. Fruit dehiscence, as a separation oftissues, is an abscission-like process. Thus, we concluded thatethylene may be a natural regulator of young fruit abscissionand fruit dehiscence.
Horticulturists have long known that ethylene hastens fruitwall (shuck, hull, pericarp, and accessory parts) dehiscence ofwalnut (6, 19) and pecan (9), but until recently there has beenonly indirect evidence that ethylene is produced by fruits thatdehisce. Lipe and Morgan (13) found that ethylene productionby pecan fruits increased to a relatively high level shortly be-fore and during fruit wall dehiscence. Similar results havebeen observed with detached, dehiscing cotton fruits as well(15, 16). In these studies (15, 16), a smaller peak of ethyleneproduction was also noted near the time of maximum youngfruit abscission.
While fruit dehiscence is generally considered a componentof the ripening process, where the regulatory role of ethyleneis well established (3, 17), other considerations indicated theneed for the present study. Fruit ripening has been studiedmost extensively in fleshy fruits which do not dehisce, and somefruits dehisce after they are dry while others open before ap-preciable weight loss occurs (13, 16). In some cases, for ex-ample pecans, dehiscence also involves detachment (abscis-sion) of the seed from its vascular connections. Dehiscence asa physiological process has not received detailed attention, andyet it is a critical phase in the production of some crops whereit is the dominant visible feature of the ripening process. Natu-ral abscission of whole, ripe fruits of the species studied hereis rare; seed dispersal can promptly follow dehiscence.
Production of ethylene by flowers has been rather exten-sively studied, primarily in association with petal fading andabscission (3, 17), and there is a presumptive association ofethylene with the natural separation of fruits from plants fol-lowing ripening. All of this information was useful in our ap-proach to the specific process of young fruit abscission.
In the present investigation, a detailed monitoring of theseasonal pattern of ethylene production by intact cotton fruitswas conducted, ethylene production by dehiscing cotton andpecan fruits was carefully detailed, and the capacity of physio-logical levels of exogenous ethylene to regulate dehiscence ofdetached fruits was determined.
MATERIALS AND METHODSPattern of Ethylene Production by Developing Cotton
Fruits. In preliminary studies (15, 16) we observed ethyleneproduction by detached cotton (Gossypium hirsutum L., cv.SP23) fruits from anthesis until the completion of fruit de-hiscence. Triplicate samples of cotton fruits from a field onthe Texas A&M University Farm, tagged on the same day atanthesis. were collected weekly, and enclosed in groups ofthree in a 500-ml Erlenmeyer flask on water-saturated filterpaper. After 3 to 4 hr, the ethylene content of an air samplefrom each flask was measured gas chromatographically.
Ethylene was measured with a hydrogen flame ionizationdetector (2). Five-milliliter air samples were injected with a
759 www.plantphysiol.orgon August 14, 2019 - Published by Downloaded from
Copyright © 1972 American Society of Plant Biologists. All rights reserved.

Plant Physiol. Vol. 50, 1972
gas-sampling valve at an oven temperature of 120 C on anactivated alumina column with helium at 75 ml/min as thecarrier gas. The lower limit of detection with this system wasapproximately 0.003lAof ethylene per liter of air.
Ethylene production by attached (intact organs on the plant),floral buds, flowers, and fruits of cotton (cv. Stoneville 213)was measured daily from 2 days before anthesis until the com-pletion of dehiscence. Greenhouse-grown plants were placed ina controlled environment room during the period when ethyl-ene was measured. Growth room conditions were: 15-hr photo-period of approximately 2000 ft-c beginning at 6 AM, 32 C,and a 9-hr dark period at 24 C with constant 48 ± 5% relativehumidity.
Floral buds were enclosed in 400-ml glass chambers thatsealed around the peduncle. The chambers were constructedfrom the tops of 500-ml Virtis Quickseal freeze-drying flasks.The fruit stem was inserted in a slit in the rubber gasket, andtwo tops were clamped onto the gasket. Each glass chamberwas covered by a loosely draped double layer of cheeseclothto minimize heat buildup inside the chamber during the lightperiod. A thermocouple was used to monitor temperature dif-ferences between the surface of an enclosed fruit and a fruitnot enclosed. The difference was found to be less than 1 C atall times during the light-dark cycle. Leakage of ethylene fromthe chambers was experimentally established as approximately5% of the total content per hour; however, since enchamberedfruits produced ethylene continuously, the actual loss of ethyl-ene during, for example, a 3-hr collection period was probablyconsiderably less than 15% of the final concentration.
Ethylene was collected daily during a single 3-hr period, 9AM to noon. Chambers were open to room air at all other times.Ethylene production was monitored until 2 days after the com-pletion of fruit dehiscence.
Calculations of ethylene production are expressed as micro-liters of ethylene produced per kg fresh weight-hr. For con-venience the production rate term was shortened in the text tomicroliters/kg hr. Weights of attached fruits were estimatedby comparison with sampled fruits of similar size and stage ofdevelopment.Time Sequence of Ethylene Production and Fruit Dehis-
cence. Ethylene production by pecan (Carya illinoensis [Wang.]K. Koch) and cotton fruits wererueasured in detail during thelatter portion of the fruiting season. The time sequence ofnatural fruit dehiscence relative to ethylene production wasobserved.
Pecans for this portion of the study were from 19-year-oldMoore and Stuart variety trees on the Texas A&M UniversityFarm. Sampling for measurement of ethylene production wasbegun a few days before natural dehiscence of the fruit wall.The procedure for measurement of ethylene production bypecans was identical with that described for detached cottonfruit in the previous section. However, as the relative ages ofpecan fruits are quite uniform, randomly selected, untaggedfruits were used.
In addition to the experiments described in the previousparagraphs, a more detailed study of the time sequence ofethylene production and natural dehiscence of attached cottonfruits was conducted. Greenhouse-grown Stoneville 213 plantswere placed in a controlled environment chamber and indi-vidual fruits that were approximately 1 week from dehiscencewere enclosed in gas collection chambers. The growth roomconditions, glass chambers, and technique for measuring ethyl-ene were described in the previous section. Ethylene measure-ments were made at 5-hr intervals during the 15-hr light periodand over a single 9-hr interval during the dark period. Trialsestablished that ethylene production during the night was notsignificantly different whether campling was done twice (two
4.5-hr periods) or once (one 9-hr period). Ethylene produc-tion was monitored until approximately 2 days after the com-
pletion of fruit dehiscence. After each measurement, the glasschambers were opened and flushed with air and then resealedfor the next collection period.A daily pattern of ethylene production was observed. There-
fore, ethylene production was monitored from attached fruitson cotton plants in a controlled environment chamber with:(a) 15-hr photoperiod and constant temperature of 27 C, (b)constant dark at 24 C, and (c) constant light at 24 C. Relativehumidity was constant at 48+ 5% in all tests.
Ratio of Internal Ethylene Concentration to Ethylene Pro-duction Rate. The ratio of the internal concentration of ethyl-ene to production of ethylene by cotton and pecan fruits wasdetermined so that: (a) ethylene production rate data couldbe converted to internal ethylene levels, (b) levels of ethyleneused in fumigation studies could be selected that would ap-proximate those which occur naturally at a later time. Fruitsof Moore pecan and TamcotSP37 cotton were sampled shortlybefore dehiscence. Individual fruits were enclosed in stoppered500-ml Erlenmeyer flasks and ethylene production was meas-ured after 1 to 2 hr. The concentration of ethylene in the flaskair was insignificant relative to the level of ethylene within theenclosed fruits. Internal ethylene was then measured from a0.5-ml air sample withdrawn from each fruit while it was heldunder water. Air samples were taken from cotton fruits by in-serting the needle of a gas-tight syringe into a locule. Airsamples were taken from the vicinity of seeds of pecan fruitsby forcing the needle through the shell. From the data, ratiosof internal ethylene (,ul/l) to ethylene production (,ul/kg hr)were calculated for cotton and pecan fruits.
Fumigation with Ethylene. Detached cotton, pecan, andokra (Hibiscus esculentus L.) fruits were fumigated with physi-ological levels of ethylene and the effect on dehiscence ob-served. Cotton fruits for this portion of the study were fromgreenhouse-grown Stoneville 213 plants. Moore and Stuartpecans from the Texas A&M University Plantation were used.Okra fruits were from field-grown United States Departmentof Agriculture Plant Introduction No. 65.
Fruits were surface-sterilized with commercial sodium hy-pochlorite (diluted 1:4) and placed upright in Petri dishes con-taining a 2% glucose solution. Groups of 10 to 15 fruits eachwere fumigated with ethylene in 54.5-liter Plexiglas chambers.Dishes of freshly dried Drierite were kept in the chambers toreduce humidity. Ethylene was applied at rates of 0, 0.1, 1.0,and 10.0,l/l. Pecans sampled 8 weeks before natural dehis-cence did not respond to any of these levels of ethylene; there-fore, groups of five pecans were also fumigated with 10001ul/l of ethylene. The experiments were conducted in a growthroom with a 15-hr photoperiod and a constant temperature of27 C. Dehiscence was recorded and each chamber was airedand refumigated daily until the completion of dehiscence. Toob,erve dehiscence, slight pressure was placed on the separa-tion zones of each fruit. When any zone of a fruit was partiallyor completely separated, that fruit was recorded as dehisced.Thus, our data represent the beginning of dehiscence whichwas usually completed within 24 hr after the first separationcould be seen.
RESULTS
Pattern of Ethylene Production by Developing CottonFruits. In preliminary work with weekly observations (15, 16),significant amounts of ethylene were produced by developingcotton fruits during two periods, the intervals just precedingyoung fruit abscission and fruit dehiscence. Detached cottonfruits produced approximately 3 --of ethylene per kg1-hr on
760 LIPE AND MORGAN
www.plantphysiol.orgon August 14, 2019 - Published by Downloaded from Copyright © 1972 American Society of Plant Biologists. All rights reserved.

ETHYLENE IN ABSCISSION AND DEHISCENCE
the day of anthesis. Not reported earlier (15, 16) was the ob-servation that 35% of the young fruits present abscised duringthe week following anthesis. This represented almost 90% ofthe total fruit abscission that occurred from these plants. Sub-sequently, ethylene production by cotton fruits was well below1 ,ul/kg -hr until approximately 1 week before the initiation offruit dehiscence when a maximum of almost 7 Al/kg hr wasreached.Our preliminary observations were extended by a daily ethyl-
ene assay of four attached cotton fruits, and the typical patternof ethylene production by two of the fruits is presented in Fig-ure 1. Attached fruits produced ethylene in a pattern basicallysimilar to detached fruits. Ethylene production by young fruitswas highest on the day of anthesis, ranging from 2.1 to 9.2,ul/kg hr for the four fruits. By 4 days after anthesis, ethyleneproduction by each fruit had dropped to generally less than1 pl/kg hr. Young fruit abscission from these plants generallyoccurred from 3 to 8 days after anthesis; however, no fruitswere abscising at the time the data from young fruits in Figure1 were taken. Between 4 and 33 days after anthesis, essentiallythe entire period of fruit enlargement, ethylene production bythe fruits was usually below 1 ,ul/kg hr. At about 5 to 7 daysbefore dehiscence, all of the fruits began a sharp increase inethylene production. Maximum ethylene production occurred37 to 38 days after anthesis, or 1 day before fruit dehiscence.Production peaks for the four fruits ranged from 8.1 to 11.5pi of ethylene per kg *hr.Time Sequence of Ethylene Production and Fruit Dehis-
cence. Our observations on seasonal ethylene production bycotton fruits included the period of fruit dehiscence. We sub-sequently conducted more detailed studies of the time sequenceof ethylene production during dehiscence of cotton and pecanfruits.An increase in ethylene production by Moore (Fig. 2A) and
Stuart (Fig. 2B) pecans preceded fruit wall dehiscence by 2 to6 days and 4 to 10 days, respectively. Moore pecans produceda relatively sharp ethylene peak with a maximum of 33 ,lI/kg-hr during an 8-day dehiscence period. Stuart pecans pro-duced a wide ethylene peak with a maximum of 24 el/kg hrduring a 17-day period of dehiscence.
Ethylene production by three attached cotton fruits of sixthat were monitored during the dehiscence period is presentedin Figure 3 to illustrate the daily pattern and range of ratesobserved. Ethylene production by the fruits began to increasealmost a week before the initiation of dehiscence. Maximumethylene production of 3.0 to 5.7 1ld/kg-hr was attained 1 to 2days before dehiscence began.A diurnal rhythm in ethylene production by cotton fruits
was quite apparent (Fig. 3). Fruits produced considerably lessethylene during the night than during the day. We verifiedthat this reduction was not due to the longer collection periodat night. Night production was well below 1 j1l of ethyleneper kg-hr except during the 2 days before dehiscence whensome fruits produced as much as 1.5 ,lJ/kg hr at night.The contribution of the growth room temperature changes
(32 C day, 24 C night) to the diurnal ethylene production pat-tern was investigated by monitoring ethylene production at aconstant temperature of 27 C with other conditions unchanged.The day-night photoperiod and temperature differences con-tribute to the diurnal pattern (Fig. 4). No diurnal variation ofethylene production was observed under either continual lightor continual darkness at a constant temperature.
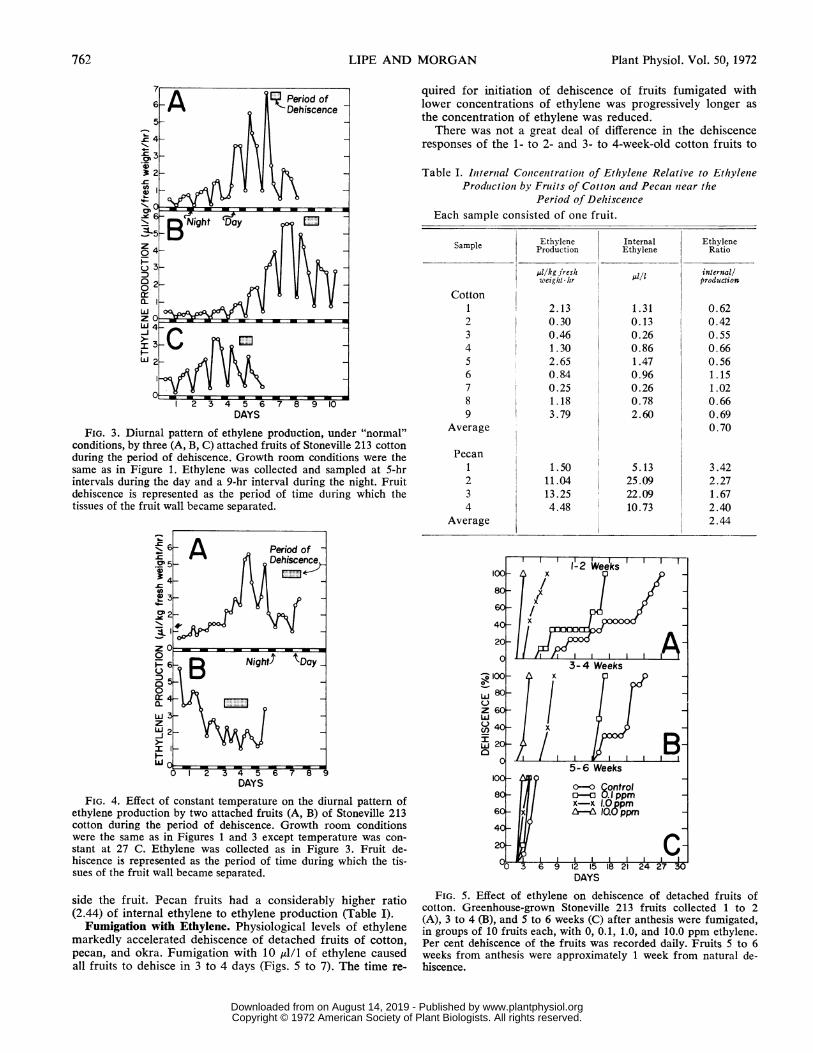
Ratio of Internal Ethylene Concentration to Ethylene Pro-duction. Results showing the ratio of internal ethylene con-centration (,u/l) to ethylene production (pl/kg-hr) are pre-sented in Table I. The ratio was reasonably uniform over awide range, 0.25 to 3.79 1.d/kg hr, of ethylene production by
DAYS FROM BLOOM
FIG. 1. Daily production of ethylene by two attached Stoneville213 cotton fruits (A, B) from 2 days before anthesis (bloom) to thecompletion of dehiscence. Ethylene production was measured atnoon each day after a 3-hr collection period. Fruit dehiscence isrepresented as the period of time during which the tissues of thefruit wall became separated.
DATE
FIG. 2. Pattern of ethylene production and fruit wall dehiscenceof detached Moore (A) and Stuart (B) pecan fruits. Measurementswere made at 2-day intervals beginning shortly before the periodof dehiscence and concluding after all of the fruit walls on trees inthe study had dehisced. Each ethylene production point is the aver-age of duplicate samples of three fruits each collected from eachof four trees. Dehiscence was recorded for walls of fruits in thesamples.
nine cotton fruits. The average ratio of internal ethylene toethylene production by cotton fruits was 0.70. This means thatfor each microliter/kg-hr of ethylene produced by a maturecotton fruit, there are approximately 0.70 ul/l of ethylene in-
Plant Physiol. Vol. 50, 1972761
www.plantphysiol.orgon August 14, 2019 - Published by Downloaded from Copyright © 1972 American Society of Plant Biologists. All rights reserved.
Plant Physiol. Vol. 50, 1972
quired for initiation of dehiscence of fruits fumigated withlower concentrations of ethylene was progressively longer asthe concentration of ethylene was reduced.
There was not a great deal of difference in the dehiscenceresponses of the 1- to 2- and 3- to 4-week-old cotton fruits to
Table I. Internial Conicenztrationt of Ethylenie Relative to EthylenieProduiction by Frutits of Cotton and Pecan nzear the
Period of DehisceniceEach sample consisted of one fruit.
Sample
DAYS
FIG. 3. Diurnal pattern of ethylene production, under "normal"conditions, by three (A, B, C) attached fruits of Stoneville 213 cottonduring the period of dehiscence. Growth room conditions were thesame as in Figure 1. Ethylene was collected and sampled at 5-hrintervals during the day and a 9-hr interval during the night. Fruitdehiscence is represented as the period of time during which thetissues of the fruit wall became separated.
Cotton123456789
Average
Pecan1234
Average
_
0N
LI
zLUI
(I)
LI
0
DAYS
FIG. 4. Effect of constant temperature on the diurnal pattern ofethylene production by two attached fruits (A, B) of Stoneville 213cotton during the period of dehiscence. Growth room conditionswere the same as in Figures 1 and 3 except temperature was con-stant at 27 C. Ethylene was collected as in Figure 3. Fruit de-hiscence is represented as the period of time during which the tis-sues of the fruit wall became separated.
side the fruit. Pecan fruits had a considerably higher ratio(2.44) of internal ethylene to ethylene production (Table I).
Fumigation with Ethylene. Physiological levels of ethylenemarkedly accelerated dehiscence of detached fruits of cotton,pecan, and okra. Fumigation with 10 ul/l of ethylene causedall fruits to dehisce in 3 to 4 days (Figs. 5 to 7). The time re-
EthyleneProduction
2.130.300.461.302.650.840.251.183.79
1.5011.0413.254.48
DAYS
FIG. 5. Effect of ethylene on dehiscence of detached fruits ofcotton. Greenhouse-grown Stoneville 213 fruits collected 1 to 2(A), 3 to 4 (B), and 5 to 6 weeks (C) after anthesis were fumigated,in groups of 10 fruits each, with 0, 0.1, 1.0, and 10.0 ppm ethylene.Per cent dehiscence of the fruits was recorded daily. Fruits 5 to 6weeks from anthesis were approximately 1 week from natural de-hiscence.
InternalEthylene
EthyleneRatio
,gI/kg fresh AIl Iinternallweight* / production
1.310.130.260.861.470.960.260.782.60
5.1325.0922.0910.73
0.620.420.550.660.561.151.020.660.690.70
3.422.271.672.402.44
4)
0'.
CP0
(-)
I-WI,
762 LIPE AND MORGAN
www.plantphysiol.orgon August 14, 2019 - Published by Downloaded from Copyright © 1972 American Society of Plant Biologists. All rights reserved.

ETHYLENE IN ABSCISSION AND DEHISCENCE
each level of ethylene although most fruits in the 1- to 2-week-old group began to dehisce sooner in 0.1 All of ethylene andno added ethylene (control) (Fig. 5, A and B). Although the5- to 6-week-old fruits (Fig. 5C) dehisced quickly, a distinctseparation of the dehiscence rate with each level of ethylenewas still quite evident.
With lower levels of ethylene the difference in dehiscencerates of okra fruits of different ages was much more markedthan occurred with cotton fruits. One-week-old okra controlfruits dehisced completely in only 5 days (Fig. 6A), whereas3- to 4-week-old control fruits required 21 days to completedehiscence (Fig. 6B). The time required for initiation of de-hiscence of fruits was progressively longer as the concentra-tion of ethylene was reduced.
Fruit walls of detached pecan fruits would not dehisce withor without ethylene until approximately 1 month before natu-ral dehiscence. This was determined by periodic fumigationwith up to 1000 yd/I of ethylene for 5 days. Fruit walls onpecan fruits fumigated with ethylene within 2 to 3 weeks ofnatural dehiscence were readily stimulated to dehisce by eachlevel of ethylene used (Fig. 7). Moore pecans sampled 2 to 3weeks and 1 week before natural dehiscence needed only 3to 4 days to dehisce in 10 tul/l of ethylene (Fig. 7, A and B).Stuart pecans sampled 1 week before natural dehiscence took6 days in 10 ,ul/l of ethylene to complete dehiscence (Fig. 7C).Some of the fruits sampled 1 week before natural dehis-
cence (Fig. 7, B and C) dehisced quickly under all levels ofadded ethylene, indicating that these fruits were producingsufficient endogenous ethylene to negate the effect of appliedethylene. Most Stuart fruits (Fig. 7C) dehisced more slowlythan Moore fruits under each level of ethylene applied.
DISCUSSION
The results of this study suggest that ethylene is involvedin both fruit abscission and fruit dehiscence. Data from threedifferent species support the hypothesis that ethylene is amajor regulator of fruit dehiscence, the dominant visible fea-ture of ripening in two of the species. Ethylene was producedin adequate amounts, long enough before fruit dehiscence tocause the response.
Ethylene production by cotton and pecan fruits commonlyreached peak rates of at least 3 to 30 and 20 to 34 1u/kg-hr,respectively, shortly before they dehisced (Figs. 1 to 4) (13).Calculations from the data of Table I indicate that productionof 3 I-l of ethylene per kg *hr by mature cotton fruits is equiva-lent to an internal concentration of 2.1 ,dl/l of ethylene. Pro-duction of 20 pi of ethylene per kg hr by pecan fruits isequivalent to an internal ethylene concentration of almost 50,ul/l. Based on results of ethylene fumigation studies (Figs. 5-7), these internal concentrations of ethylene in cotton andpecan fruits should easily stimulate, if not completely saturatethe dehiscence regulation systems of cotton, pecan, and okrafruits where as little as 0.1 pl/l of ethylene caused a measura-ble stimulation of dehiscence (Figs. 5 to 7). Although detachedfruit may be somewhat more sensitive to ethylene, as indicatedby dehiscence, than attached fruit, natural production of ethyl-ene exceeds the minimum level necessary to promote dehis-cence by a margin that allows confidence in the conclusion ofa causal relationship. Other studies have shown that ethyleneproduction of 3 to 5 ,l/kg hr essentially saturates ethylene-responsive systems such as swelling of stem and root sections,prevention of hook opening, and abscission of debladedpetioles (3). This evidence supports the contention that ethyl-ene produced by dehiscing cotton and pecan fruits was suffi-cient to initiate or regulate dehiscence. It should be pointedout that the ratio of ethylene production to internal concen-
020
a-) _zw00
W 8C_
60
40_
20
( 3 b 9 121IDAYS
18 21 24
FIG. 6. Effect of ethylene on dehiscence of detached fruits ofokra. Field-grown fruits of United States Department of Agricul-ture Plant Introduction 65 were collected approximately 1 week (A)and 3 to 4 weeks (B) after anthesis and were fumigated in groupsof 10 fruits each.
MooAe >J3 IA?4cs '
Moore I Week Conro
80xz60 - g x/ 0.1 ppm
I-40- t '°
KO K-K6 1.0 ppm 1
0
pecan fruits MoorefrItcolceW o3wek A neek
0
(B)befre atual ehsteceart StarWeetkolete we
8 K-
2n40 4 0 1Y~ ~~DY
(B)efor nauraldehScence an Statfriscolced1we
(C) before natural dehiscence were fumigated in groups of 15 fruitseach.
tration varies with the stage of fruit development (Lipe andMorgan, unpublished data); thus, the values here do not ap-ply, for example, to the period of fruit expansion when ethyl-ene production is low (Fig. 1), and sensitivity to ethylene isalso apparently low because abscission is rare.The timing of ethylene production and natural fruit dehis-
cence also supports the proposal that ethylene is a regulator offruit dehiscence. Data in Figures 1 to 4 show accelerated ethyl-ene production by fruits during the week prior to dehiscence.The timing of ethylene production and dehiscence was observed
o-o Control0-c-O 0.1 ppmxKx 1.0 ppm
10.0 ppm
A-
xK/
/
763Plant Physiol. Vol. 50, 1972
www.plantphysiol.orgon August 14, 2019 - Published by Downloaded from Copyright © 1972 American Society of Plant Biologists. All rights reserved.

Plant Physiol. Vol. 50, 1972
in detail in lengthy studies of ethylene production by individualcotton fruits (Fig. 3). Ethylene production from cotton fruitsincreased to at least 2 to 3 Aul/kg hr, 3 days before dehiscence.A maximum of between S to 10 ,ul of ethylene per kg hr wasgenerally produced 1 day before dehiscence. Pecan fruits pro-duced considerably more ethylene than cotton fruits duringthe days immediately preceding dehiscence (Fig. 2). In com-parison fumigation of detached fruits with 10 ul/l of ethyl-ene produced complete dehiscence in only 3 to 4 days and1.0 tl/l1 of ethylene stimulated most fruits to dehisce in 4 to8 days (Figs. 5 to 7). Ethylene production of 1.4 yl/kg hr bycotton fruits and only 0.4 41./kg hr by pecans is roughlyequivalent to fumigation with 1.0 ,Al/l of ethylene accordingto the average ratios from Table I. Most cotton fruits wereproducing near 1.5 ,u of ethylene per kg hr at least 4 daysbefore they dehisced (Figs. 1 and 3). Pecan fruits appear tohave surpassed production of 0.4 /.l of ethylene per kg hr wellover a week before shucks dehisced (Fig. 2).An additional correlation of ethylene production during
natural dehiscence with the effect of ethylene fumigation onfruits was noted. Moore pecans produced a sharp ethylenepeak (Fig. 2A) that spanned an 8- to 10-day period and pre-ceded, by 3 to 5 days, a 10-day period of fruit wall dehiscence.Stuart pecans produced a broad ethylene peak (Fig. 2B) span-ning a 16- to 18-day period and preceding, by 5 to 10 days,an 18-day period of shuck dehiscence. Dehiscence of fruitwalls of detached Moore and Stuart pecan fruits fumigatedwith 10 ,kl/l of ethylene required 1 to 3 days (Figs. 7, A and7B) and 2 to 6 days (Fig. 7C), respectively. This shows thatthe detailed pattern of dehiscence for each variety, althoughquantitatively different for the two varieties, was basically thesame whether in the presence of internally produced ethyleneor applied ethylene.The dehiscence response of several ages of fruits to fumiga-
tion with ethylene (Figs. S to 7) appears to have been governedby the endogenous balance of dehiscence promoters and in-hibitors. Young cotton (Fig. SA) and especially young okrafruits (Fig. 6A) dehisced more quickly than fruits sampled atmid-development (Figs. 5B and 6B). Mature cotton fruits(Fig. SC), as well as young okra fruits, dehisced most rapidly.The young fruits were sampled soon after the period of ethyl-ene production associated with young fruit abscission. It ap-pears that the endogenous ethylene as well as the probabledepletion of juvenile hormones (auxins, gibberellins, andcytokinins) (20), after the fruits were detached, caused a rela-tively quick shift of the hormonal balance in favor of de-hiscence-promoting, senescence factors, i.e., ethylene andpossibly ABA (8, 18). The young okra fruits (Fig. 6A), whichdehisced quickly, were sampled under conditions of minimumfruit set, and young cotton fruits (Fig. SA), which dehiscedmore slowly, were sampled during a period of maximum fruitset. Apparently, detached cotton and okra fruits samplednear mid-development (Figs. SB and 6B) dehisced relativelyslowly because this is the period of rapid growth when juvenilehormones are probably plentiful (7, 10, 20) and ABA (8, 18)and ethylene (Figs. 1 and 3) are at minimum levels. The rapiddehiscence of mature cotton fruits (Fig. SC) probably reflectsa natural decline in juvenile hormones as well as a significantincrease in ABA (8, 18) and ethylene (Fig. 1) within the fruits.
Pecan fruit walls, incapable of dehiscence until approxi-
mately 1 month before natural fruit wall dehiscence, ap-pear to be regulated by both sensitivity to and the level ofethylene. Sensitivity to ethylene probably includes morethan juvenile materials since massive levels of ethylene couldnot cause dehiscence. Thus, an accumulation of an additionalsenescence-promoting material, possibly ABA, may be neces-sary to allow dehiscence. ABA levels in pecans (14) and otherfruits (1) do show massive increases in activity during thetime that dehiscence tissues become sensitive to ethylene.
Although dehiscence is the most obvious aspect of ripen-ing in pecans and cotton and a process of economic importancefor each species, the rapid dehiscence of very young okrafruits with even 0.1 ,ul/l of ethylene is interesting because itsuggests that the dehiscence process may be artificially sep-arated from ripening in some cases.
LITERATURE CITED
1. ADDICOTT, F. T. 1969. Physiology of abscisic acid and related substances. Annoi.Rev. Plant Physiol. 20: 139-164.
2. BEYER, E. M. AND P. W. MIORGAN. 1971. Abscission: The i-ole of etliv enemiiodlificationi of auxin transport. Plant Physiol. 48: 208-212.
3. BURG, S. P. 1962. The physiology of ethylene formation. Annuii. Rev. PlantPhysiol. 13: 265-302.
4. BURG, S. P. 1968. Ethylene, plant senescence and abscission. Plant Physiol. 43:1503-1511.
5. BURG, S. P. AN-D E. A. BURG. 1965. Gas exchange in fruits. Physiol. Plant. 18:870-884.
6. CHACE, E. M. AND D. G. SORBER. 1936. Treating frtuits and ntuts in atmosplherescontaining ethylene. Food Industries 8: 292.
7. CRANE, J. C. 1964. Growth substances in fruit setting and development. Annu.Rev. Plant Physiol. 15: 303-326.
8. DAVIS, L. A. 1968. Gas chromatographic identification and measuremeint ofabscisic acid and othter plant hormones in the developing cotton fulit. Ph.D.Dissertation. University of California at Davis. Also, Plant Physiol. 49:644-648.
9. FINcCH, A. H. 1937. The use of ethylene to improxe pecan harvesting. Proc.Amer. Soc. 1lort. Sci. 34: 74-77.
10. JACKSON, D. I. A-ND B. G. COOMBE. 1966. Gibberellin-like substances in the de-veloping apricot fruit. Science 154: 277-278.
11. JACKSON, M. B. AND D. J. OSBORNE. 1970. Ethylene, tlle natuiral iegulator ofleaf abscission. Nature 225: 1019-1022.
12. LEOIPOLD, A. C. 1964. Plant Growth and Development. McGraw-Hill Book Co.,New York.
13. LIPE, J. A. AND P. W. MORGAN. 1970. Ethylene: Involvement in shuck de-hiscence in pecan fruits (Carya illinocnisis [Wang.] K. Koch). Hortscience5: 266-267.
14. LIPE, J. A., P. W. MORGANN, AND J. B. STOREY. 1969. Growtlh substances andfruit shedding iu the pecan, Carya illinoensis. Proc. Amer. Soc. Hort. Sci.94: 668-671.
15. MORGAN, P. W., D. L. KETRING, E. MI. BEYER, JR., AN-D J. A. LIPE. 1972.Functions of naturally pr-ocluced ethylene in abscission, dleliisceince andseedI germiiination. In: D. J. Carr, ed., Plant Growtth Substances 1970.Springer-Verlag, New York. pp. 502-509.
16. MIORGAN, P. W., E. M. BEYER, J. A. LiPE, AND J. A. AMCAFEE. 1971. Etlhylene,a regulator of natural cotton leaf shed and boll opening. Proceedings of theBeltwide Cotton Production and Research Conference, Cotton Defoliation-Physiology Conference 25: 42-44.
17. PRATT, H. K. AND J. D. GOESCHL. 1969. Physiological roles of ethylene inplants. Annu. Rev. Plant Physiol. 20: 541-584.
18. SMITH, 0. E. 1969. Changes in abscission-accelerating substances witli devel-opnient of cotton fruit. New Phytol. 68: 313-322.
19. SORBER, D. G. AND MI. H. KIMBALL. 1950. Use of ethy-lene in harvesting thePersian walnut (Juglans regia) in California. U. S. Dep. Agr. Tech. Btull.996: 1-80.
20. VAN OVERBEEK, J. 1962. Endogenous regulators of fruit grvowth. Proceedingsof Plant Science Symposium, Campbell Soup Co., Camden, New Jersey. pp.37-58.
21. YANG, S. F. 1968. Biosynthesis of ethylene. In: F. Wightman and G. Setter-field eds., Biochemistry and Physiology of Plant Growth Substanices. RungePress, Ottawa. pp. 1217-1228.
764 LIPE AND MORGAN
www.plantphysiol.orgon August 14, 2019 - Published by Downloaded from Copyright © 1972 American Society of Plant Biologists. All rights reserved.


















