
Ethylene- and shade-induced hypocotyl elongation share transcriptome patterns and functional...
48
1 Short title: Ethylene- and shade-induced stress escape 1 Corresponding authors: Rashmi Sasidharan, Ronald Pierik 2 Address: Padualaan 8, 3584 CH, Utrecht, The Netherlands 3 Telephone number: +31 30 2536838 4 Email: [email protected], [email protected] 5 6 Ethylene- and shade-induced hypocotyl elongation share transcriptome 7 patterns and functional regulators 8 Das D. 1 , St. Onge K.R. 1,2 , Voesenek L.A.C.J. 1 , Pierik R. 1 * and Sasidharan R. 1 * 9 *shared senior and corresponding authors 10 1 Plant Ecophysiology, Institute of Environmental Biology, Utrecht University, 3584CH Utrecht, The 11 Netherlands 12 2 Department of Biological Sciences, CW405 BioSci Building, University of Alberta, T6J2E9, Canada 13 14 Summary: Ethylene and shade share a conserved set of molecular regulators to 15 control plasticity of hypocotyl elongation in Arabidopsis. 16 17 AUTHOR CONTRIBUTIONS 18 R.P., R.S and L.A.C.J.V. conceived the original research plans and project. R.S., 19 R.P, L.A.C.J.V., K.R.S. supervised the experiments. D.D. performed most of the 20 experiments. D.D., K.R.S., R.P. and R.S. designed the experiments. D.D. and 21 K.R.S. analyzed the data. D.D. wrote the article with contributions of all the authors. 22 FUNDING INFORMATION 23 This research was supported by the Netherlands Organisation for Scientific 24 Research (grant nos. ALW Ecogenomics 84410004 to L.A.C.J.V., ALW VENI 25 86312013 and ALW 82201007 to R.S., ALW VIDI 86412003 to R.P.) and a Utrecht 26 University Scholarship to D.D. 27 28 CORRESPONDING AUTHOR EMAIL: [email protected], [email protected] 29 30 31 32 Plant Physiology Preview. Published on June 21, 2016, as DOI:10.1104/pp.16.00725 Copyright 2016 by the American Society of Plant Biologists www.plantphysiol.org on July 4, 2016 - Published by www.plantphysiol.org Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.
-
Upload
debatosh-das -
Category
Science
-
view
44 -
download
2
Transcript of Ethylene- and shade-induced hypocotyl elongation share transcriptome patterns and functional...

1
Short title: Ethylene- and shade-induced stress escape 1
Corresponding authors: Rashmi Sasidharan, Ronald Pierik 2
Address: Padualaan 8, 3584 CH, Utrecht, The Netherlands 3
Telephone number: +31 30 2536838 4
Email: [email protected], [email protected] 5
6
Ethylene- and shade-induced hypocotyl elongation share transcriptome 7
patterns and functional regulators 8
Das D.1, St. Onge K.R.1,2, Voesenek L.A.C.J.1, Pierik R.1* and Sasidharan R.1* 9
*shared senior and corresponding authors 10 1Plant Ecophysiology, Institute of Environmental Biology, Utrecht University, 3584CH Utrecht, The 11 Netherlands 12 2 Department of Biological Sciences, CW405 BioSci Building, University of Alberta, T6J2E9, Canada 13
14
Summary: Ethylene and shade share a conserved set of molecular regulators to 15
control plasticity of hypocotyl elongation in Arabidopsis. 16
17
AUTHOR CONTRIBUTIONS 18
R.P., R.S and L.A.C.J.V. conceived the original research plans and project. R.S., 19
R.P, L.A.C.J.V., K.R.S. supervised the experiments. D.D. performed most of the 20
experiments. D.D., K.R.S., R.P. and R.S. designed the experiments. D.D. and 21
K.R.S. analyzed the data. D.D. wrote the article with contributions of all the authors. 22
FUNDING INFORMATION 23
This research was supported by the Netherlands Organisation for Scientific 24
Research (grant nos. ALW Ecogenomics 84410004 to L.A.C.J.V., ALW VENI 25
86312013 and ALW 82201007 to R.S., ALW VIDI 86412003 to R.P.) and a Utrecht 26
University Scholarship to D.D. 27
28
CORRESPONDING AUTHOR EMAIL: [email protected], [email protected] 29
30
31
32
Plant Physiology Preview. Published on June 21, 2016, as DOI:10.1104/pp.16.00725
Copyright 2016 by the American Society of Plant Biologists
www.plantphysiol.org on July 4, 2016 - Published by www.plantphysiol.orgDownloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

2
ABSTRACT 33
Plants have evolved shoot elongation mechanisms to escape from diverse 34
environmental stresses such as flooding and vegetative shade. The apparent 35
similarity in growth responses suggests possible convergence of the signalling 36
pathways. Shoot elongation is mediated by passive ethylene accumulating to high 37
concentrations in flooded plant organs and by changes in light quality and quantity 38
under vegetation shade. Here we study hypocotyl elongation as a proxy for shoot 39
elongation and delineated Arabidopsis hypocotyl length kinetics in response to 40
ethylene and shade. Based on these kinetics, we further investigated ethylene and 41
shade-induced genome-wide gene expression changes in hypocotyls and cotyledons 42
separately. Both treatments induced a more extensive transcriptome reconfiguration 43
in the hypocotyls compared to the cotyledons. Bioinformatics analyses suggested 44
contrasting regulation of growth promotion- and photosynthesis-related genes. 45
These analyses also suggested an induction of auxin, brassinosteroid and gibberellin 46
signatures and the involvement of several candidate regulators in the elongating 47
hypocotyls. Pharmacological and mutant analyses confirmed the functional 48
involvement of several of these candidate genes and physiological control points in 49
regulating stress-escape responses to different environmental stimuli. We discuss 50
how these signaling networks might be integrated and conclude that plants, when 51
facing different stresses, utilise a conserved set of transcriptionally regulated genes 52
to modulate and fine tune growth. 53
54
INTRODUCTION 55
All organisms, including plants, assess and respond to both biotic and abiotic factors 56
in their environments (Pierik and De Wit, 2014; Pierik and Testerink, 2014; Franklin 57
et al., 2011; Quint et al., 2016; Osakabe et al., 2014; Voesenek and Bailey-Serres, 58
2015). However, unlike animals, plants, cannot move away from extremes in their 59
surrounding environment but rather rely on various plastic morphological and 60
metabolic responses. Such response traits include changes in plant architecture to 61
escape the stress and optimize resource capture (Pierik and Testerink, 2014; 62
Mickelbart et al., 2015). With energy reserves being invested in escape traits, plants 63
often have lower plant biomass and crop yield (Casal, 2013). Molecular investigation 64
www.plantphysiol.org on July 4, 2016 - Published by www.plantphysiol.orgDownloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

3
of the different signalling pathways controlling these traits along with the 65
characterization of underlying molecular components would not only enhance 66
fundamental knowledge of stress-induced plasticity but also benefit crop 67
improvement. 68
Plants are highly sensitive to changes in their light environment. Young plants 69
growing in a canopy experience changes in light quality and quantity due to 70
neighbouring plants and compete to harvest optimum light (Pierik and De Wit, 2014; 71
Casal et al., 2013). When a plant cannot outgrow its neighbours, it experiences 72
complete vegetation shade (hereafter termed as “shade”) which, in addition to low 73
red: far-red (R:FR), is marked by a significant decline in blue light and overall light 74
quantity. These changes initiate so-called Shade Avoidance Syndrome (SAS) 75
responses consisting of petiole, hypocotyl and stem elongation; reduction of 76
cotyledon and leaf expansion; upward movement of leaves (hyponasty), decreased 77
branching and increased apical dominance (Vandenbussche et al., 2005; Franklin, 78
2008; Casal, 2012; Pierik and De Wit, 2014). Shade-induced elongation comprises a 79
complex network of photoreceptor-regulated transcriptional and protein-level 80
regulation involving BASIC HELIX LOOP HELIX (BHLH) and HOMEODOMAIN-81
LEUCINE ZIPPER, (HD-ZIP) transcription factors and auxin, gibberellin and 82
brassinosteroid hormone genes (Casal, 2012; 2013). Flooding often leads to partial 83
or complete submergence of plants. Water severely restricts gas diffusion and the 84
consequent limited exchange of O2 and CO2 restricts respiration and photosynthesis. 85
Another consequence is the rapid accumulation of the volatile hormone ethylene. 86
Ethylene is considered an important regulator of adaptive responses to flooding, 87
including accelerated shoot elongation responses that bring leaf tips from the water 88
layer into the air (Sasidharan and Voesenek, 2015; Voesenek and Bailey-Serres, 89
2015). In deepwater rice, this flooding-induced elongation response involves 90
ethylene-mediated induction of members of the group VII Ethylene Response 91
Factors (ERF-VII) family, a decline in active abscisic acid (ABA) and consequent 92
increase in gibberellic acid (GA) responsiveness and promotion of GA biosynthesis 93
(Hattori et al., 2009). In submerged Rumex palustris petioles, ethylene also rapidly 94
stimulates cell wall acidification and transcriptional induction of cell wall modification 95
proteins to facilitate rapid elongation (Voesenek and Bailey-Serres, 2015). Shade 96
cues are reported to enhance ethylene production resulting in shade avoidance 97
www.plantphysiol.org on July 4, 2016 - Published by www.plantphysiol.orgDownloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

4
phenotypes (Pierik et al., 2004). However, these responses are mediated by 98
ethylene concentrations of a much lower magnitude than that occuring in flooded 99
plant organs (1µl L-1) (Sasidharan and Voesenek, 2015). 100
So far, it is largely unknown to what extent these growth responses to such highly 101
diverse environmental stimuli, share physiological and molecular components 102
through time. A preliminary study in R. palustris showed that GA is a common 103
regulator of responses to both submergence and shade (Pierik et al. 2005). Although 104
submergence is a compound stress, rapid ethylene accumulation is considered an 105
early and reliable flooding signal triggering plant adaptive responses. High ethylene 106
concentrations as occur within submerged plant organs, promote rapid shoot 107
elongation (Voesenek and Bailey-Serres, 2015). This submergence response, which 108
has been extensively characterised in rice and Rumex (Hattori et al. 2009; van Veen 109
et al., 2013), can be almost completely mimicked by the application of saturating (1µl 110
L-1) ethylene concentrations (Sasidharan and Voesenek, 2015). Saturating ethylene 111
concentrations were therefore used here as a submergence mimic. Shade was given 112
as true shade which combines the three known key signals that trigger elongation 113
(red and blue light depletion with relative far-red enrichment). 114
A hypocotyl elongation assay in Arabidopsis thaliana (Col-0) was used as a proxy for 115
shoot elongation under ethylene and shade in order to study to what extent ethylene 116
and shade responses share molecular signalling components. Although ethylene 117
suppresses Arabidopsis hypocotyl elongation in dark, high ethylene concentrations 118
in light (as occurs during submergence) stimulate hypocotyl elongation in 119
Arabidopsis (Smalle et al., 1997; Zhong et al., 2012). Also upon simulated shade, 120
Arabidopsis demonstrates pronounced hypocotyl elongation (Morelli and Ruberti, 121
2000). To capture early physiological responses and gene expression changes in 122
response to ethylene and shade, Arabidopsis seedling hypocotyl elongation and 123
cotyledon expansion were examined over time. The two treatments elicited 124
characteristic hypocotyl growth kinetics. To uncover the transcriptomic changes 125
regulating the elongation response to these signals, an organ-specific genome-wide 126
investigation was carried out on hypocotyls and cotyledons separately at three time-127
points corresponding to distinct hypocotyl elongation phases. Clustering analyses in 128
combination with biological-enrichment tests allowed identification of gene clusters 129
with expression patterns matching the hypocotyl growth trends across the three time-130
www.plantphysiol.org on July 4, 2016 - Published by www.plantphysiol.orgDownloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

5
points in both the treatments. Correlation of genome-wide hypocotyl and cotyledon-131
specific transcriptomic changes to publicly available microarray data on hormone 132
treatments identified enriched hormonal signatures of auxin, brassinosteroid and 133
gibberellin in hypocotyl tissues and several potential growth regulatory candidate 134
genes. Using hormone mutants and chemical inhibitors, we confirmed the combined 135
involvement of these hormones and candidate regulators in the hypocotyl elongation 136
response to ethylene and shade. We suggest that growth responses to diverse 137
environmental stimuli like ethylene and shade converge on a common regulatory 138
module consisting of both positive and negative regulatory proteins that interact with 139
a hormonal triad to achieve a controlled fine-tuned growth response. 140
141
142
RESULTS 143
Delineation of hypocotyl elongation kinetics under ethylene and shade in 144
Arabidopsis seedlings 145
Exogenous application of ethylene (1 µl L-1) in light-grown seedlings resulted in thick, 146
yet elongated hypocotyls and smaller cotyledons as compared to untreated controls. 147
Shade, achieved by the use of a green filter, stimulated strong hypocotyl elongation 148
in seedlings but resulted in mildly smaller cotyledons compared to controls (Fig. 1A). 149
Hypocotyl length increments in the two treatments relative to control were around 2-150
fold under ethylene and greater than 3-fold under shade (Figs. 1, A and B). For both 151
treatments growth stimulation was strongest in the first two days of treatment (Fig. 152
1C) and faded out in the subsequent days. When combined, shade and ethylene 153
exposure resulted in hypocotyl lengths that were intermediate to the individual 154
treatments (Supplemental Fig. S1). 155
To get a more detailed time-line of the early elongation kinetics, we performed a 156
follow-up experiment with 3 h measurement intervals for the first 33 h (Fig. 1D). 157
Ethylene-mediated stimulation of hypocotyl elongation started only in the middle of 158
the dark period after 15 h. However, under shade, longer hypocotyls were recorded 159
already after 3 h and this rapid stimulation continued until the start of the dark period. 160
Interestingly, accelerated elongation was again observed at around 24 h after the 161
start of the treatments (when the lights were switched on). Based on this time-line, 162
we determined epidermal cell lengths at time-points 0, 3, 7.5, 15 and 27 h (Gendreau 163
www.plantphysiol.org on July 4, 2016 - Published by www.plantphysiol.orgDownloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

6
et al., 1997; Fig. 1E), the latter four of which are either prior to start of accelerated 164
growth or during it in response to ethylene and shade. The rapid stimulation of 165
elongation under shade starts at the base of the hypocotyl (3 h) and then progresses 166
all along the hypocotyl with maximum elongation occurring in the middle segment 167
while under ethylene accelerated elongation is observed at the middle-bottom of 168
hypocotyl (27 h). 169
170 Organ-specific transcriptomics in hypocotyl and cotyledon under ethylene and 171
shade 172
The transcriptome response to ethylene and shade in hypocotyl and cotyledon 173
tissues was characterised using Affymetrix Arabidopsis Gene 1.1 ST arrays at three 174
time points of hypocotyl length kinetics (1.5 h, 13.5 h and 25.5 h) (Fig. 1D). Principal 175
component analysis (PCA) (Abdi and Williams, 2010) of all replicate samples for 176
hypocotyl and cotyledon exposed to control, ethylene or shade conditions showed 177
that replicate samples generally clustered together (Fig. 2A). The first principal 178
component (34.2%) separates tissue-specific samples, whereas the second principal 179
component (13.0%) showed separate clustering of the 13.5h samples which falls 180
during the dark period. 181
Hierarchical clustering (HC) (Eisen et al., 1998) of mean absolute expression 182
intensities for the different main samples (combination of 3 replicates) revealed 183
similar trends (Fig. 2B). Fig. 2C shows the distribution of up- and down-regulated 184
differentially expressed genes (DEGs) (genes with adjusted p-value ≤ 0.01) in 185
hypocotyl and cotyledon for ethylene and shade at the three harvest time points 1.5 186
h, 13.5 h, 25.5 h respectively. In both the conditions and tissues, the number of both 187
up- and down-regulated DEGs increased with time. Ethylene regulated substantially 188
more DEGs in the hypocotyl as compared to shade at all harvest time points (Fig. 189
2C). Data analysis identified 6668 and 4741 genes (hereafter termed as “total 190
DEGs”) that were differentially expressed in hypocotyl at one or more of the three 191
tissue harvest time points by ethylene and shade respectively. Interestingly, in the 192
cotyledon, at 1.5 h, the number of significant DEGs under ethylene was higher than 193
under shade but at the subsequent two time points, shade regulated more genes. In 194
the cotyledon, 1197 and 2173 DEGs were identified that were differentially 195
expressed at one or more of the three tissue harvest time points by ethylene and 196
shade respectively. 197
www.plantphysiol.org on July 4, 2016 - Published by www.plantphysiol.orgDownloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

7
Interestingly, at 1.5 h, there was more transcriptional regulation in ethylene- than 198
shade-exposed hypocotyls even though subsequent hypocotyl elongation was much 199
more rapid in shade (Figs. 2C and 1D). For ethylene-specific downregulated DEGs 200
at 1.5 h, the topmost enriched GO term was cell wall organization (containing 30 201
genes), which suggested a repression of growth-promoting genes and possible lack 202
of ethylene-mediated elongation at 1.5 h (Supplemental Fig. S2). We also found 6 203
genes (AT1G65310, XYLOGLUCAN ENDOTRANSGLUCOSYLASE / HYDROLASE 204
17 (XTH17); AT5G23870, pectin acetyl esterase; AT3G06770, glycoside hydrolase; 205
AT5G46240, POTASSIUM CHANNEL IN ARABIDOPSIS THALIANA 1 (KAT1); 206
AT1G29460, SMALL AUXIN UPREGULATED RNA 65 (SAUR65) and AT3G02170, 207
LONGIFOLIA 2 (LNG2)) in the shade-specific upregulated 32 genes at 1.5 h, with 208
implied cell expansion roles and could possibly be associated with the rapid 209
elongation response (Sasidharan et al., 2010; Philippar et al., 2004; Nozue et al., 210
2015; Chae et al., 2012; Lee et al., 2006). 211
212
Different gene expression clusters contributing to hypocotyl growth in 213
ethylene and shade 214
In order to find specific genes regulating the elongation phenotype under both 215
treatments, we used temporal clustering of DEGs based on expression values. Due 216
to distinct hypocotyl length kinetics in response to ethylene and shade (Fig. 1D), we 217
searched for a set of temporally co-expressed genes that could potentially contribute 218
to this treatment-specific kinetics. Time-point based clustering was performed for the 219
6668 ethylene and 4741 shade total DEGs based on the positive or negative 220
magnitude of log2FC for DEGs at the three time points (Fig. 3, A and D). The gene 221
expression patterns in clusters-1 and 5 across the three time points matched the 222
ethylene hypocotyl growth kinetics closely (Fig. 3, B and C). Similarly, gene 223
expression kinetics in clusters-1 and 3 matched the hypocotyl length kinetics in 224
shade (Fig. 3, E and F). These growth pattern matching clusters were termed 225
“positive”. All the clusters with mirror image of gene expression profiles to that of the 226
positive clusters (clusters-8 and 4 in ethylene and clusters-8 and 6 in shade) were 227
termed as “negative” clusters. 228
www.plantphysiol.org on July 4, 2016 - Published by www.plantphysiol.orgDownloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

8
Next, a hypergeometric over-representation test for selected MapMan bins (“stress”, 229
“hormone”, “signalling”, “RNA.Regulation of transcription” and “cell wall”) was carried 230
out for the temporal gene clusters (Fig. 3G). Interestingly, “cell wall”, “hormone” and 231
“signalling” were highly co-enriched in positive clusters (cluster-1 and 5 for ethylene 232
and cluster-1 for shade) which hints at co-regulation of genes mapped to these terms 233
during transcriptomic response to ethylene and shade in the hypocotyl. 234
To identify growth promotion related DEGs, we identified DEGs common to both the 235
treatments in the clusters previously designated as ‘positive’ and ‘negative’ clusters 236
(Fig. 3, H and I). 997 DEGs were obtained from a venn diagram between the 237
treatment-specific positive clusters, upregulated at atleast two time-points, and 238
hereafter called “Common Up”. Similarly, 824 DEGs were shared between ethylene 239
and shade negative clusters, were downregulated at atleast two time points and 240
hereafter called “Common Down”. 241
Enriched functional categories in the different gene sets from Venn diagrams of 242
positive and negative clusters, were identified using the GeneCodis tool (Tabas-243
Madrid et al., 2012) (Fig. 4). In the Common Up set, we found a variety of growth 244
associated GO categories including cell wall modification, hormone (auxin and 245
brassinosteroid) signaling and metabolism, transport processes, tropisms, response 246
to abiotic stimuli and signal transduction. The ethylene-specific set for positive 247
clusters was enriched for ethylene-associated terms as expected, but also for 248
various sugar metabolic, endoplasmic reticulum (ER)-related and protein post-249
translational modification-related processes. Some of the enriched GO terms in the 250
ethylene-specific set for positive clusters were also found in the Common Up set, but 251
caused by different genes in the same GO category, including those associated with 252
growth, hormones and transport processes. The shade-specific set for positive 253
clusters showed only few clear GO enrichments such as trehalose metabolism, 254
secondary cell wall biogenesis and amino acid metabolism, but also shared some 255
with the Common Up set like shade avoidance and protein phosphorylation and with 256
both Common Up and ethylene-specific set for positive clusters, such as response to 257
auxin and unidimensional growth. 258
In the Common Down set, GO terms associated with photosynthesis, primary and 259
secondary metabolism, response to biotic and abiotic stress, as well as 260
www.plantphysiol.org on July 4, 2016 - Published by www.plantphysiol.orgDownloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

9
photomorphogenesis were enriched. The latter is striking given that ethylene does 261
not alter the light environment. The ethylene-specific set for negative clusters 262
included strong enrichment of circadian rhythm and a variety of photosynthesis-and 263
chloroplast-associated GO terms, partially shared with the Common Down set. In the 264
shade-specific set for negative clusters, flavonoid/anthocyanin biosynthesis and 265
response to UV-B and heat were enriched. The shade-specific set for negative 266
clusters shared terms from defence-associated GO categories and cadmium, karrikin 267
response with Common Down set. Terms common to all three sets of negative 268
clusters were related to photomorphogenesis and metabolic process. 269
270
Functional characterization: shared components in ethylene -and shade-271
mediated regulation of hypocotyl length 272
We classified Common Up and Down set genes into transcriptional regulators, 273
hormone metabolism genes, signalling genes and cell wall genes and also applied a 274
logFC filter (see “Materials and Methods” sub-section “LogFC Filter and Gene 275
Classification”) to obtain a final list of 53 and 8 genes in the two sets respectively 276
(Fig. 5, A and B). We selected a subset of candidate regulators for functional testing 277
and made sure to include two transcription factors since these may be relatively 278
upstream in the convergence of signaling pathways. Obvious targets for these 279
regulators would be different plant hormones, and before zooming in on these, we 280
ran a hormonometer analysis with our transcriptome data to further subset our 281
candidate regulators. 282
283
Transcription factor candidates: 284
ARABIDOPSIS THALIANA HOMEOBOX PROTEIN 28 (ATHB28) is a zinc-finger 285
homeodomain (ZF-HD) transcription factor that showed upto 2-fold induction in the 286
hypocotyls in ethylene and shade respectively. A homozygous null mutant 287
(Supplemental Fig. S3, A-D), “athb28”, showed a significant reduction in hypocotyl 288
lengths under both ethylene and shade (Fig. 6A), consistent with its induction upon 289
both the treatments. 290
www.plantphysiol.org on July 4, 2016 - Published by www.plantphysiol.orgDownloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

10
Both shade and ethylene also significantly induced transcript levels of the bHLH 291
transcription factor INCREASED LEAF INCLINATION1 BINDING bHLH1 like-1 292
(IBL1) Arabidopsis hypocotyls. However, the ibl1 (SALK T-DNA insertion line) 293
showed wild-type responses to the treatments (Fig. 6B). The bHLH transcription 294
factors IBL1 and its homolog IBH1 have been implicated in repressing BR-mediated 295
cellular elongation. In an ibl1 mutant, IBH1 is still present and may negatively 296
regulate cell elongation independently of IBL1. The 35S overexpression line, IBL1OE 297
(Zhiponova et al., 2014), had shorter hypocotyls than the wild-type under control 298
conditions. IBL1OE also lacked ethylene and shade-induced hypocotyl elongation 299
implying an inhibitory role for IBL1 (Fig. 6C). 300
301
Hormone candidates: auxin, brassinosteroid and gibberellin 302
To further investigate the significant hormone related changes amongst the growth 303
related DEGs we analysed our data using Hormonometer (Volodarsky et al., 2009). 304
For both treatments, the hormonal signatures across the three time-points for BR 305
and GA most closely matched the hypocotyl elongation kinetics (Figs. 1D and 7A). 306
The analysis also showed significant correlations with auxin responses for all data 307
sets. 308
In the Common Up set 49 genes were present that were all also auxin-regulated, 309
whereas there were 14 that were also BR-regulated. In the Common Down set 16 310
genes were present that can be regulated by auxin, 16 that can also be ABA-311
regulated and 13 genes that are also JA-regulated. Interestingly, there were no 312
genes for BR in the Common Down set. In addition, genes involved in auxin-313
conjugation genes (GRETCHEN HAGEN 3 FAMILY PROTEIN 3.17 (GH3.17) and 314
AT5G13370), GA catabolising genes (GIBBERELLIN 2 OXIDASE (GA2OX) 2, 315
GA2OX4 and GA2OX7) and JA augmenting genes (LIPOXYGENASE (LOX) 1, 316
LOX2 and LOX3) were down-regulated. 317
In order to test the possible role of auxin, GA and BR, in mediating shade and 318
ethylene-induced hypocotyl elongation, we first tested the effects of pharmacological 319
inhibitors of these hormones on shade and ethylene-induced hypocotyl elongation. 320
To visualize the auxin effect we treated the pIAA19:GUS auxin response marker line 321
with the auxin transport inhibitor, 1-N-Naphthylphthalamic acid (NPA). As shown in 322
www.plantphysiol.org on July 4, 2016 - Published by www.plantphysiol.orgDownloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

11
Fig. 7B, application of NPA (25 µM) inhibited hypocotyl elongation and also strongly 323
reduced staining in the hypocotyl region. This inhibition was rescued by IAA (10 µM). 324
The auxin perception inhibitor, α-(phenylethyl-2-one)-indole-3-acetic acid, PEO-IAA 325
(100 µM), strongly reduced staining in the whole pIAA19:GUS seedling and inhibited 326
elongation in response to both treatments. The significant inhibition of ethylene and 327
shade-induced hypocotyl elongation by the different auxin inhibitors is quantitated in 328
Fig. 7,C-E. The addition of NPA, yucasin (auxin biosynthesis inhibitor) and α-329
(phenylethyl-2-one)-indole-3-acetic acid (PEO-IAA; auxin antagonist) inhibited 330
hypocotyl elongation under both the treatments confirming that all three aspects of 331
auxin are required for ethylene- and shade-induced hypocotyl elongation. BR 332
biosynthesis inhibitor brassinazole (BRZ) and GA biosynthesis inhibitor paclobutrazol 333
(PBZ) fully inhibited these elongation responses as well (Fig. 7, F and G). 334
To further validate the involvement of auxin, BR and GA in ethylene and shade-335
induced hypocotyl elongation we tested hypocotyl elongation responses in a variety 336
of hormone mutants, including mutants for candidate genes from Figure 5. 337
Both the auxin receptor (tir1-1) and biosynthesis (wei8-1) mutants showed 338
significantly impaired hypocotyl elongation responses compared to the wild type 339
ethylene and shade response (Fig. 8, A and B). A similar effect was seen in the 340
auxin transport mutant pin3pin4pin7 which had severely reduced hypocotyl 341
elongation in both treatments (Fig. 8C). 342
The GA biosynthesis (ga1-3) and insensitive (gai) mutant both showed a complete 343
lack of hypocotyl elongation in both treatments (Fig. 8, D and E). We also tested the 344
GA biosynthesis mutant ga20ox1-3, since it was identified as a ‘common Up’ gene, 345
induced in response to both treatments (Fig. 5A). The ga20ox1-3 mutant showed a 346
significantly reduced elongation phenotype in both treatments compared to the wild 347
type response (Fig. 8F). 348
The BR receptor (bri1-116) and biosynthesis (dwf4-1) mutants both showed severe 349
hypocotyl elongation phenotypes, and did not respond to either treatment (Fig. 8, G 350
and I). In another biosynthesis mutant, rot3-1, while the ethylene elongation 351
response was absent there was a severely reduced shade response (Fig. 8H). Two 352
BR-metabolism related genes were identified in the common Up set (Fig. 5A): 353
BR6OX1 and BAS1. The bas1-2 mutant showed constitutive elongation in all 354
treatments ((Fig. 8J) confirming a negative role of BR catabolism through BAS1 in 355
www.plantphysiol.org on July 4, 2016 - Published by www.plantphysiol.orgDownloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

12
hypocotyl elongation control. Although the BR biosynthetic mutant br6ox1 (cyp85a1-356
2) mutant did not show any phenotypic alteration (Fig. 8K), a double mutant of 357
BR6OX1 and BR6OX2 (cyp85a1cyp85a2) showed a complete lack of elongation in 358
response to both ethylene and shade (Fig. 8L). 359
360 361
Discussion 362
Accelerated shoot elongation is a common mode of stress escape that allows plants 363
to grow away from stressful conditions (Pierik and Testerink, 2014). Stress escape 364
does, however, come at an energetic cost and is only beneficial if improved 365
conditions are achieved. Here our goal was to establish to what extent shade and 366
ethylene elicit similar responses through shared or distinct molecular pathways. In 367
our study we found distinct elongation kinetics in ethylene and shade for Arabidopsis 368
hypocotyls, differing in both temporal regulation and the degree of response. Shade 369
treatment evoked a rapid, strong and persisting hypocotyl elongation, whereas 370
ethylene initially inhibited elongation and only in the first night period started to 371
promote hypocotyl length (Fig. 1). In both treatments, hypocotyl growth involved 372
enhanced epidermal cell elongation. Previous studies have shown that low R:FR 373
induces ethylene biosynthesis in Arabidopsis (Pierik et al., 2009) and it could be 374
argued that the shade response might act through ethylene. However, ethylene 375
marker genes were not induced in shade, and the ethylene-insensitive ein3eil1 376
mutant retained a full response to shade (Supplemental Fig. S4), ruling out a role for 377
ethylene in the shade response. Interestingly, combining shade and ethylene 378
treatments did not lead to an additive response and instead dampened shade-379
induced hypocotyl elongation (Supplemental Fig. S1). The growth inhibitory effect of 380
ethylene under shaded conditions could function similar to its effects in limiting 381
hypocotyl elongation in the dark i.e. via induction of negative growth regulators such 382
as ERF1 (Zhong et al., 2012). Transcriptome characterisation of the elongating 383
hypocotyl upon exposure to single shade and ethylene stresses indicated 384
considerable overlap between the two treatments. Thus, a large portion of DEGs 385
under both treatments may contribute to similar processes implying that they target 386
shared genetic components but have treatment-specific upstream regulatory factors. 387
388
www.plantphysiol.org on July 4, 2016 - Published by www.plantphysiol.orgDownloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

13
Hypocotyl growth promotion and photosynthesis repression occur 389
concurrently under ethylene and shade 390
By identifying gene clusters with expression patterns closely matching the distinct 391
ethylene and shade growth kinetics, we identified positive and negative clusters for 392
the respective treatments. These clusters were also among the bigger clusters that 393
contributed to most of the transcriptomic changes, suggesting that a large part of 394
transcriptomic response are associated with the hypocotyl growth and concurrent 395
biological processes. Functional enrichment analysis for the Common Up set (shared 396
between positive clusters of ethylene and shade) (Fig. 4), suggested an involvement 397
of growth promoting genes. Cell wall genes are all involved in mediating cellular 398
expansion in growing hypocotyls. However, they need to be controlled by either the 399
environmental signal directly or by upstream factors in the signal transduction 400
pathway. The Common Down set (shared between negative clusters of ethylene and 401
shade) (Fig. 4), was highly enriched in photosynthesis-related terms and proteins. 402
The effects of ethylene on photosynthesis can be positive or negative depending on 403
the context (Iqbal et al., 2012; Tholen et al., 2007). Low R:FR treated stems of 404
tomato showed reduced expression of photosynthetic genes (Cagnola et al., 2012). 405
This reduction was mainly due to a decrease in expression of Calvin cycle genes, 406
which we also observed for our Common Down set (Supplemental Table S1). In 407
addition, under ethylene specifically, Photosystem II and I genes were mostly 408
repressed. Thus acceleration of hypocotyl elongation is accompanied by repression 409
of genes associated with non-elongation processes like metabolism and 410
photosynthesis. This was also shown to be true for low R:FR treated elongating 411
stems of tomato (Cagnola et al., 2012). Light capture and carbon fixation are 412
minimized and energy is apparently invested in stimulating growth (Sulpice et al., 413
2014; Henriques et al., 2014; Lilley et al., 2012). It would be interesting to investigate 414
how photomorphogenic responses are associated with and influence photosynthesis 415
and growth promotion. 416
417
Convergence of signaling pathways in response to ethylene and shade in 418
control of hypocotyl elongation 419
In shade avoidance responses, photoreceptors like phyB and cry1/2 would regulate 420
the elongation phenotype via control of Phytochrome Interacting Factor (PIF) levels. 421
However, since these are photoreceptors, it seems unlikely that these proteins 422
www.plantphysiol.org on July 4, 2016 - Published by www.plantphysiol.orgDownloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

14
themselves would integrate information from the ethylene pathway as well (Park et 423
al., 2012; Li et al., 2012). It was shown by van Veen et al. (2013), that in the 424
submerged petioles of R. palustris (which displays petiole elongation under complete 425
submergence), early molecular components of light signalling (KIDARI, COP1, PIFs, 426
HD-ZIP IIs) are induced by ethylene independently of any change in light quality. 427
Over expression of PIF5 on the other hand leads to increased ethylene production in 428
etiolated Arabidopsis seedlings, causing inhibition of hypocotyl length (Khanna et al., 429
2007). Interestingly, the downstream ethylene signal transduction protein EIN3 was 430
shown to physically interact with PIF3 (Zhong et al., 2012). We, therefore, suggest 431
that ethylene and shade might both induce this shared gene pool by, for example, 432
targeting (different) members of the PIF family of transcription factors. Since different 433
PIFs likely regulate the expression of at least partly shared target genes (Leivar and 434
Monte, 2014), this would explain our observed partial overlap in the transcriptional 435
response to shade and ethylene. PIFs are also known to directly bind and regulate 436
expression of other transcription factors like homeodomain (HD) TFs (Capella et al., 437
2015; Kunihiro et al. 2011) in the control of shade avoidance responses. Indeed, our 438
TAIR motif analysis hinted towards the presence of significantly enriched binding 439
signatures of PIF/MYC proteins (CACATG) as well as HD proteins (TAATTA) in the 440
upstream promoter sequences of the Common Up set genes (Pfeiffer et al., 2014; 441
Kazan and Manners, 2013; Supplemental Fig. S5). 442
Several potential transcriptional regulators were identified in the narrowed down 443
Common Up gene set. A growth-promoting role of KIDARI in regulating elongation in 444
response to shade and ethylene was suggested previously (Hyun and Lee 2006; van 445
Veen et al., 2013). Upregulation of another bHLH encoding gene and a negative 446
regulator of elongation, IBL1 was observed for both treatments (Fig. 6C)). While 447
PIF4 induces IBH1 and IBL1, IBH1 represses PIF4 targets (Zhiponova et al., 2014). 448
IBH1 and its homolog IBL1 collectively regulate expression of a large number of BR-, 449
GA- and PIF4-regulated genes and this might their mode of action in shade- and 450
ethylene-induced hypocotyl elongation. In addition to these bHLH proteins, we show 451
that the ZF-HD transcription factor ATHB28 is also involved in regulating hypocotyl 452
elongation under ethylene and shade (Fig. 6A). Hong et al. (2011) showed that 453
another ZF-HD protein, MIF1 interacts strongly with four other ZF-HD proteins 454
including ATHB33 and ATHB28. This leads to non-functional MIF1-ATHB 455
heterodimers and inhibition of e.g. ATHB33-regulated expression and growth 456
www.plantphysiol.org on July 4, 2016 - Published by www.plantphysiol.orgDownloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

15
promotion. Transcriptomics data for 35S:MIF1 (displaying short hypocotyl 457
phenotype), shows downregulation of auxin, BR and GA responsive genes and 458
upregulation of ABA genes (Hu and Ma, 2006). We can speculate that MIF1 on one 459
hand and ATHB33 and ATHB28 on the other hand, might target the same set of 460
hormone genes, but in an opposite manner, to control growth. What remains to be 461
studied is how ethylene and shade regulate ZF-HD transcription factors and this will 462
be an important topic for future studies. 463
Well-established targets for above-mentioned PIFs and HD TFs are various aspects 464
of auxin signalling and homeostasis, such as YUCCA biosynthetic enzymes and 465
AUX/IAA proteins for signalling (Kunihiro et al. 2011; Li et al., 2012; Sun et al., 2012; 466
De Smet et al., 2013). Our list of candidate genes (Fig. 5) contained auxin-467
responsive transcriptional regulator IAA3 and many of the auxin-responsive SAUR 468
genes which have been shown to positively modulate hypocotyl elongation (Kim et 469
al., 1998; Sun et al., 2012; Chae et al., 2012; Spartz et al., 2012) and may act 470
individually or in concert to regulate the phenotype. With reference to elongation 471
responses under shade in Arabidopsis seedlings, auxin seems to play a major role. 472
An increase in free auxin levels and its transport towards epidermal cells in hypocotyl 473
is necessary for low R:FR-mediated hypocotyl elongation (Tao et al., 2008; 474
Keuskamp et al., 2010; Zheng et al., 2016). The importance of YUCCAs and TAA1 in 475
low R:FR responses has been previously demonstrated (Li et al., 2012). It is 476
generally assumed that auxin synthesized in the cotyledons is required to regulate 477
hypocotyl elongation in response to e.g. low R:FR light conditions (Procko et al. 478
2014). Indeed, cotyledons are key regulators of hypocotyl elongation in a 479
phytochrome-dependent way (Endo et al., 2005; Warnasooriya and Montgomery, 480
2009; Estelle, 1998; Tanaka et al., 2002). In our data, hormonometer analysis 481
identified strong induction of auxin-associated genes in the cotyledons in both 482
treatments (Fig. 7). We speculate that the physiological regulation of hypocotyl 483
elongation in our study depends on cotyledons via auxin dynamics. However, we 484
also show that auxin is certainly not the only shared physiological regulator between 485
the ethylene and shade response. 486
GA20OX1 and BR6OX1 expression were up-regulated in patterns that closely 487
matched the hypocotyl elongation profiles (Fig. 5A), and hormonometer analysis also 488
revealed enrichment of GA and BR hormonal signatures in ethylene and shade 489
www.plantphysiol.org on July 4, 2016 - Published by www.plantphysiol.orgDownloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

16
exposed hypocotyls. (Fig. 7). The positive role of GA in flooding-associated shoot 490
elongation (Voesenek and Bailey-Serres, 2015) and shade avoidance (Djakovic-491
Petrovic et al., 2007) is well established. It is well known that GA20OX1 specifically 492
affects plant height without having any other major phenotypic effects (Barboza et 493
al., 2013; Rieu et al., 2008), and it has been shown to be involved in shade 494
avoidance (Nozue et al. 2015; Hisamatsu et al. 2005). In our data, ga20ox1 knockout 495
showed reduced elongation to shade as well as ethylene, extending its function from 496
controlling shade avoidance to ethylene mediated elongation responses (Fig. 8F). 497
GA biosynthesis via GA20OX1 can be induced by brassinosteroids, suggesting a 498
possible cross-talk between the two growth-promoting hormones (Unterholzner et 499
al., 2015). Although future studies are needed to establish if this crosstalk occurs 500
under the conditions tested here, we do confirm that BR is an important hormone 501
involved in both the responses since several BR mutants showed disturbed 502
elongation responses to ethylene and shade (Fig. 8G-8L). Interestingly, also auxin 503
and brassinosteroids have partially overlapping roles in hypocotyl elongation control 504
(Chapman et al., 2012; Nemhauser et al., 2004), further extending the crosstalk 505
towards a tripartite network. Among the BR mutants tested is the cyp85a1cyp85a2 506
mutant, encoding a double mutant for BR6OX2 and BR6OX1 which showed a 507
complete lack of elongation to both treatments (Fig. 8L) similar to a BRZ treatment 508
(Figs. 7F). BR6OX1 was one of the direct candidate genes identified from the 509
transcriptomics analysis (Figure 5A). A tripartite bHLH transcription factor module 510
consisting of IBH1, PRE and HBI1, has been previously implicated in regulating cell 511
elongation in response to hormonal and environmental signals (Bai et al., 2012). 512
Several BR biosynthesis and signaling genes are direct targets of HBI1, including 513
BR6OX1 (Fan et al. 2014), indicating the possible involvement of this bHLH 514
regulatory module in promoting BR responses during shade and ethylene exposure. 515
Why is there be such an elaborate network of regulators and even hormones 516
involved in controlling unidrectional cell expansion in hypocotyl growth responses? 517
To achieve a controlled growth, feedback loops are likely required, and crosstalk 518
between different routes are probably a necessity to deal with multiple environmental 519
inputs simultaneously. We found BAS1 transcriptional upregulation in hypocotyls in 520
response to ethylene and shade. BAS1 may act to balance the hypocotyl growth 521
promotion mediated by brassinosteroids (castasterone (CS) and brassinolide (BL)) 522
www.plantphysiol.org on July 4, 2016 - Published by www.plantphysiol.orgDownloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

17
as it inactivates both CS and BL (Neff et al., 1999; Turk et al., 2005). In shade 523
avoidance research, likewise, HFR1 is induced by PIFs to suppress the growth 524
promotion induced the same PIF proteins, and putative control of DELLAs would 525
modulate GA responses. 526
Conclusion 527
Hypocotyl elongation in response to ethylene and shade treatments is likely 528
regulated at the upstream level by (a) a bHLH module consisting of positive growth 529
regulators, PIFs (PIF3 in ethylene and PIF4 and PIF5 in shade) and inhibitory factors 530
like IBL1 and (b) a homeodomain module, where ZF-HD TFs like ATHB28 may 531
either act in parallel to the bHLH TFs or are regulated by PIFs (similar to induction of 532
HD-ZIP TFs) to transcriptionally target genes related to the growth promoting 533
hormone module (auxin, BR and GA), as hinted by promoter motif analysis. We 534
hypothesize that in Arabidopsis seedlings, shade and ethylene stimulate auxin 535
synthesis in the cotyledons, which is then transported to the hypocotyl to epidermal 536
cell layers where it interacts with both GA and BR to co-ordinately induce hypocotyl 537
elongation. This increased auxin response, indicated by elevated SAUR levels, and 538
likely increased levels of GA and BR as indicated by increased GA20OX1 and 539
BR6OX expression in the hypocotyl, likely act to induce unidirectional epidermal cell 540
wall elongation via upregulation of genes encoding cell-wall modifying proteins, 541
which promote cellular expansion leading to hypocotyl elongation. 542
543
MATERIALS AND METHODS 544
Plant Material and Growth Conditions 545
Around 30 Arabidopsis thaliana (Col-0) and mutant seeds were sown per agar plate 546
containing 1.1 g L-1 Murashige-Skoog (1/4 MS) and 8 g L-1 Plant-agar (0.8%w/v) 547
(both Duchefa Biochemie, The Netherlands). Mutants or overexpression lines used 548
in this work were: pin3pin4pin7 (Blilou et al., 2005); wei8-1 (Stepanova et al., 2008), 549
tir1-1 (Ruegger et al., 1998), dwf4-1 (Azpiroz et al., 1998), rot3-1 (Kim et al., 1998), 550
ga1-3 (Wilson et al., 1992), gai (Talon et al., 1990), ein3eil1 (Binder et al., 2004); 551
bri1-116 (van Esse et al., 2012); ibl1 and IBL1OE (Zhiponova et al., 2014); 552
cyp85a1/cyp85a2 and cyp85a1-2 (Nomura et al., 2005); bas1-2 (Turk et al., 2005); 553
ga20ox1-3 (Hisamatsu et al., 2005). 554
www.plantphysiol.org on July 4, 2016 - Published by www.plantphysiol.orgDownloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

18
ibl1 (N657437), cyp85a1 (N681535), wei8-1 (N16407) and ga20ox1-3 (N669422) 555
were obtained from NASC seed-stock centre; bri1-116 was kindly provided by Sacco 556
de Vries Lab, Wageningen University while cyp85a1/cyp85a2 and dwf4-1 were 557
kindly provided by Sunghwa Choe Lab, Seoul National University. IBL1OE 558
overexpression lines were kindly provided by the Jenny Russinova lab (VIB Ghent, 559
Belgium). bas1-2 mutant was kindly provided by Michael Neff lab (Washington State 560
University, USA). Some GA mutants used were in the Ler background. All other 561
mutants were in the Col-0 background. After stratification at 4°C for 3 d in the dark, 562
seeds were transferred for 2 h to control light conditions (see below) and then kept in 563
dark (at 20°C) again for another 15 h. Subsequently seedlings were allowed a period 564
of 24 h growth under control light conditions in Short-Day photoperiod conditions (15 565
h dark/9h light) before being transferred to 22.4 L glass desiccators with air-tight lids 566
for specific treatments. Col-0 genotypes were grown at 21 ± 1 °C. Ler genotypes 567
were grown at 19 ± 1 °C (Supplemental Figure S6). 568
For athb28 (GK-326G12), lines were obtained from NASC (UK). Genotyping was 569
performed using the following primers: for athb28, athb28_fwd 570
(CTAAGTACCGGGAATGTCAGAAG); athb28_rev (TAACCAACTGAGCTATTCC 571
AGCTA) and LB primer o8474: (ATAATAACGCTG CGGA CATCTACATTTT). For 572
verifying transcript levels, ATHB28_fwd (GGAGAAGATGAAGGAATTTGCA) and 573
ATHB28_rev (TGTTTCTCTTCA TTGCTTGCT) were used. 574
575
Treatments 576
Control and ethylene desiccators were kept in control light conditions 577
(Photosynthetically Active Radiation, PAR = 140 μmol m-2 s-1, blue light (400 nm-500 578
nm) = 29 μmol m-2 s-1 and R:FR = 2.1). Ethylene treatments were started by injecting 579
ethylene into the desiccators (with 1 µl L-1 final concentration in the dessicator) and 580
levels were verified with a Gas Chromatograph (GC955; Synspec, The Netherlands). 581
Shade treatment was started by putting desiccators under a single layer of Lee Fern 582
Green Filter (Lee Hampshire, UK) (PAR = 40 μmol m-2s-1, blue light (400 nm-500 583
nm) = 3 μmol m-2 s-1 and R:FR = 0.45). For growth curve experiments, two plates 584
with 15 seedlings per treatment per genotype distributed over two desiccators were 585
used. For mutant analyses, one plate with 15 seedlings per treatment per genotype 586
was used. 587
588
www.plantphysiol.org on July 4, 2016 - Published by www.plantphysiol.orgDownloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

19
Imaging and Hypocotyl Length Measurements 589
For hypocotyl elongation assays, experiments were replicated twice. Seedling plates 590
were collected from the dessicators. Seedlings were flattened on the agar plates to 591
reveal the full extent of their hypocotyl, and images of the seedlings were obtained 592
by scanning the plates using EPSON Perfection v370 Photo scanner (Epson Europe 593
BV, The Netherlands). Hypocotyl lengths were measured from these images using 594
ImageJ (http://rsbweb.nih.gov/ij/) and values (per data point) were obtained for n ≥ 595
30-60 seedlings. Final values for the data was obtained by taking mean ± S.E. for 596
values from the two independent experiments. 597
598
Epidermal Cell Length Measurements 599
Seedlings were mounted on microscopic slides and covered with a cover slip. 600
Hypocotyl epidermal cells were imaged using Olympus AX70 (20x objective, Nikon 601
DXM1200 camera), after which cell lengths were measured using ImageJ software 602
tool (http://rsbweb.nih.gov/ij/). 603
604
Microarray Tissue Harvest, RNA Isolation and Array Hybridization 605
Seedlings were dissected using BD PrecisionGlide Hypodermic 27 Gauge 1 1/4" 606
Grey Needle with outer diameter = 0.41 mm (Becton Dickinson B.V., The 607
Netherlands) to separately harvest the hypocotyl and cotyledon + shoot apical 608
meristem (hereafter termed “cotyledon”). The roots were discarded. Samples were 609
harvested at 1.5 h, 13.5 h and 25.5 h after the start of the treatments. For 13.5 h time 610
point, which occurs during the dark period, dissection was carried out under low 611
intensity green safelight (≈ 5 μmol m-2 s-1). For minimizing the effects of green light 612
on gene expression, seedlings were kept in the dark until dissected and dissection 613
was carried out in several rounds with each round involving maximum 2-3 seedlings. 614
In total, 3 replicate experiments were carried out. In each replicate experiment, 615
tissues were harvested from two independent technical replicates (each with 25 616
seedlings from the two technical replicate plates as mentioned in “Treatments” 617
section above). Harvested material was immediately frozen in liquid nitrogen and 618
stored at -80°C until further use. 619
Frozen tissue was ground using tissue lyser and total RNA was isolated using 620
RNeasy Mini Kit (QIAGEN, Germany). QIAGEN RNase-Free DNase set was used to 621
eliminate Genomic DNA contamination by performing on-column DNase digestion. 622
www.plantphysiol.org on July 4, 2016 - Published by www.plantphysiol.orgDownloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

20
Extracted RNA was verified (for quality check) and quantified using NanoDropTM ND-623
1000 Spectrophotometer (Isogen Life Science, De Meern, The Netherlands). 624
RNA samples were sent to AROS (AROS Applied Biotechnology A/S, Aarhus, 625
Denmark). RNA was repurified on low-elution QIAGEN RNeasy columns, re-626
quantified with NanoDropTM 8000 UV-Vis Spectrophotometer and checked for quality 627
with Agilent 2100 Bioanalyzer (Agilent Technologies, Palo Alto, CA). RNA samples 628
with RNA Integrity Number (RIN) value of > 7.5 were considered for further use. 50 629
ng RNA from each of the two independent technical replicates was mixed in the ratio 630
1:1 and the pooled sample was considered as one biological replicate for 631
hybridization experiments. Thus, 3 biological replicates were obtained for the 3 632
replicate experiments. 100 ng of RNA sample was processed for cDNA synthesis, 633
fragmentation and labelling was carried out for the RNA samples. The samples were 634
hybridized to the Affymetrix Gene 1.1 ST Arabidopsis Array Plate and washed on an 635
Affymetrix GeneAtlas system followed by scanning of arrays at AROS Applied 636
Biotechnology (http://arosab.com/services/microarrays/gene-expression/). 637
638
Microarray Data Analysis 639
Scanned arrays in the form of .cel files (provided by AROS) were checked for quality 640
control using Affymetrix Expression Console Software and an in-house script in R 641
and Bioconductor (http://www.r-project.org./; http://www.bioconductor.org/) 642
(Bioconductor “oligo” and “pd.aragene.1.1.st” ). Bioconductor was used for Robust 643
Multi-array Average (RMA) normalization of raw data at Gene level to obtain 644
summarized signal intensity values for all genes present on the array (log2 format). 645
Principal component analysis was carried out using Affymetrix Expression Console 646
Software (http://www.affymetrix.com/) and dendrogram of all microarray samples 647
according to the mean signal intensity values was generated using R (“plot” 648
package). Bioconductor (“Limma” package) was used for carrying out differential 649
expression analysis. 650
651
Temporal Clustering And Bioinformatics Analysis 652
We clustered the list of total DEGs (defined as number of DEGs that were regulated 653
at atleast one of the three time points) under ethylene and shade based on positive 654
or negative regulation at each of the three time-points. With three time-points and 655
two directions of expression (positive or negative), 23 = 8 possible trends can occur 656
www.plantphysiol.org on July 4, 2016 - Published by www.plantphysiol.orgDownloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

21
and accordingly as many clusters were obtained. DEGs were clustered temporally 657
based on log2FC. 658
659
MapMan bin overrepresentation using hypergeometric test was done using R (“stats” 660
package) and adjusted p-values for the statistical significance of enrichment were 661
converted into negative logarithm score and plotted as a heatmap. A score above 662
1.3 for this score was considered significant. 663
GeneCodis (http://genecodis.cnb.csic.es/compareanalysis) webtool was used for GO 664
analysis of different sets obtained from venn of Positive clusters and of Negative 665
Clusters. A score above 1.3 for negative log of adjusted p-values was considered 666
significant. 667
668
LogFC Filter and Gene Classification 669
In order to narrow down the genes for functional characterization (from the list of 670
classified genes), we utilised a logFC filter. To narrow down the Common Up set, a 671
filter of log2FC < 0.5 at 1.5 h and log2FC ≥ 0.5 at both 13.5 h and 25.5 h for ethylene 672
and log2FC ≥ 0.5 at both 1.5 h and 25.5 h for shade was applied. To narrow down 673
the Common Down set, we applied a filter of log2FC > - 0.5 at 1.5 h and log2FC ≤ - 674
0.5 at both 13.5 h and 25.5 h under ethylene and of log2FC ≤ - 0.5 at both 1.5 h and 675
25.5 h under shade. These resulting group of genes were then classified based on 676
MapMan based classification for the terms: “RNA.regulation of transcription”, “Cell 677
wall”, “Signalling” and “Hormone metabolism”. Genes from Plant TFDB 678
(http://planttfdb.cbi.pku.edu.cn/index.php?sp=Ath) and Potsdam TFDB 679
(http://plntfdb.bio.uni-potsdam.de/v3.0/index.php?sp_id=ATH) were also included as 680
a source for gene-classification to select for additional transcriptional regulators. 681
682
Hormone Correlational Analysis 683
Hormonometer software (http://genome.weizmann.ac.il/hormonometer/) was used to 684
evaluate transcriptional similarities between the transcriptome data obtained here 685
and the published, indexed list of those elicited by exogenous application of plant 686
hormones. Arabidopsis gene locus IDs were converted to Affymetrix GeneChip 687
identifiers using the “at to AGI converter” tool (The Bio-Analytic Resource for Plant 688
Biology, http://bar.utoronto.ca/). We used the new Affymetrix aragen1.1st arrays (28k 689
genes) for transcriptomics but the hormonometer data is based on 3’ ATH1 arrays 690
www.plantphysiol.org on July 4, 2016 - Published by www.plantphysiol.orgDownloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

22
(22k genes). Accordingly, many locus IDs could not be included (those which were 691
newly incorporated in the aragene1.1st arrays) in this analysis. In few other cases 692
where multiple ATH1 GeneChip IDs for one locus ID was obtained, all IDs were 693
retained. 694
695
Pharmacological Treatments 696
Auxin transport was inhibited by use of 25 µM NPA (Duchefa Biochemie, The 697
Netherlands) (Petrásek et al., 2003). Auxin perception was blocked by use of 100 µM 698
PEO-IAA (Hayashi et al., 2008). Auxin biosynthesis was blocked by use of 50 µM 699
yucasin (Nishimura et al., 2014). 2 µM brassinazole (TCI Europe, Japan) was used 700
to inhibit BR biosynthesis (Asami et al., 2000). 2 µM paclobutrazol (Duchefa 701
Biochemie, The Netherlands) was used to inhibit GA biosynthesis (Rademacher, 702
2000). All chemicals were dissolved in respective solvents (DMSO or ETOH) with 703
final solvent concentration in media < 0.1% to prevent toxicity due to solvents. All 704
chemicals were applied by pipetting 150 µl of chemical solutions or mock solvents as 705
a thin film over the MS-agar media in the petri plates and then allowing the solution 706
to diffuse through the medium before starting the treatments. 707
708
GUS Staining and Imaging 709
For GUS assays, seedlings were transferred immediately from treatments to a GUS 710
staining solution (1 mM X-Gluc (5-bromo-4-chloro-3-indolyl-D-glucoronide) (Duchefa 711
Biochem, The Netherlands) in 100 mM Sodium phosphate buffer (pH=7.0) along with 712
0.1 % Triton X-100, 0.5 mM each of Potassium ferrocyanide (K4Fe(CN)6 and 713
Potassium ferricyanide (K3Fe(CN)6 and 10 mM of EDTA (Merck Darmstadt, 714
Germany)) and kept at 37°C overnight. Seedlings were bleached in 70% ethanol for 715
1 day before capturing images. 716
717
Statistical Analysis and Graphing 718
1-way ANOVAs followed by Tukey's HSD Post-Hoc tests were performed on the 719
measurements obtained in hypocotyl length/cotyledon area kinetics to assess 720
statistically significant differences between mean hypocotyl length/cotyledon area 721
under ethylene or shade relative to control at the same time point. 722
A two-way ANOVA followed by a Tukey's HSD Post-Hoc test was used for pairwise 723
multiple comparison. For hypocotyl elongation assays, statistical significance was 724
www.plantphysiol.org on July 4, 2016 - Published by www.plantphysiol.orgDownloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

23
indicated by the use of different letters. All statistical analyses were done in the R 725
software environment. Graphs were plotted using Prism 6 software (GraphPad 726
Software, USA). 727
728
Accession numbers 729
.CEL files utilised in the organ-specific transcriptomics for hypocotyl and cotyledon 730
tissues are posted with the GEO accession series GSE83212. 731
732
Supplemental data 733
The following materials are available in the online version of this article. 734
Supplemental Figure S1. Hypocotyl elongation response in wild-type Col-0 under 735
control, ethylene, shade and combination (ethylene + shade) treatments. 736
Supplemental Figure S2. Venn diagram of gene intersection between up- and 737
down- regulated DEGs of ethylene and shade separately at 1.5 h. 738
Supplemental Figure S3. Genotyping and transcript level verification for (A)-(D) 739
athb28. 740
Supplemental Figure S4. Hypocotyl elongation response in ethylene signalling 741
mutant, ein3eil1 under ethylene and shade. 742
Supplemental Figure S5. TAIR motif analysis for Common Up and Down genesets. 743
Supplemental Figure S6. Hypocotyl length of Ler under control and ethylene 744
conditions when grown at 220 C day / 200 C night and 200 C day / 180 C night 745
temperature regime. 746
Supplemental Table S1. Photosynthesis gene proportion in genesets from Venn of 747
negative clusters. 748
ACKNOWLEDGMENTS 749
We would like to thank all the group members of Plant Ecophysiology, Utrecht 750
University, The Netherlands for their help in the harvest for the transcriptomics 751
experiment. 752
www.plantphysiol.org on July 4, 2016 - Published by www.plantphysiol.orgDownloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

24
753
FIGURE LEGENDS 754
Figure 1. Physiological responses, hypocotyl length and epidermal cell length 755
kinetics under ethylene and shade in Arabidopsis (Col-0) seedlings. 1-day old 756
seedlings were exposed to control conditions (PAR = 140 μmol m-2 s-1, blue light = 757
29 μmol m-2 s-1and R:FR = 2.1), elevated ethylene (1 µl L-1, PAR = 140 μmol m-2 s-1, 758
blue light = 29 μmol m-2 s-1 and R:FR = 2.1) or shade (PAR = 40 μmol m-2 s-1, blue 759
light = 3 μmol m-2 s-1 and R:FR = 0.45). A, Figure shows representative seedlings 760
displaying typical phenotypes after 96 h of exposure to control, ethylene or shade 761
conditions. B, Mean hypocotyl lengths at 0 h, 24 h, 48 h, 72 h and 96 h under control 762
(open circle), ethylene (closed circle) and shade (open triangle) are shown. C, Rate 763
of increase in hypocotyl length. Differences between mean hypocotyl lengths of 764
subsequent time points averaged over 1 d time interval for control, ethylene and 765
shade are shown. D, Detailed hypocotyl length kinetics. 1-day old seedlings were 766
exposed to control (open circle), ethylene (closed circle) and shade (open triangle) 767
and measured at 3 h time intervals (s, e: first time point at which shade (s) or 768
ethylene (e) treatment lead to significantly longer hypocotyls). Data are mean ± S.E. 769
(n=60) for (A)-(D). Shaded area denotes the 15 h dark period in the 15 h dark/9 h 770
light photoperiodic growth condition. e and s denote the first point of statistically 771
significant differences in hypocotyl length or cotyledon area relative to control for 772
ethylene and shade respectively. E, Epidermal cell length kinetics. 1-day old 773
seedlings were exposed to control, ethylene or shade. Mean cell length ± S.E. (n ≥ 774
10) for epidermal cells of the Arabidopsis hypocotyl at 0 h control (black line) and at 775
3 h, 7.5 h, 15 h and 27 h control (orange line), ethylene (blue line) and shade (green 776
line) is shown. Apex denotes the hypocotyl-cotyledon junction and base denotes the 777
hypocotyl-root junction. 778
Figure 2. Overall description of microarray data. A, Principal component analysis 779
(PCA) and hierarchical clustering (HC) was used to describe the structure in the 780
microarray data. Expression intensities for all genes on the array for all 54 hypocotyl 781
and cotyledon samples (3 time points, 3 treatments and 3 replicates) were projected 782
onto the first three principal components. B, Hierarchical clustering was used to 783
group 18 main samples (according to mean expression intensity of 3 replicates for 784
www.plantphysiol.org on July 4, 2016 - Published by www.plantphysiol.orgDownloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

25
each main sample) into a dendrogram. C, Distribution of differentially expressed 785
genes (DEGs) in hypocotyl and cotyledon samples at three time-points in response 786
to ethylene and shade. DEGs obtained for each time-point were plotted separately 787
as up-regulated, down-regulated (adjusted p-value ≤ 0.01 and log2FC > or < 0) or 788
non-significant (adjusted p-value > 0.01). Bar length denotes total DEGs obtained 789
after combining DEGs from all 3 time points. 790
Figure 3. Temporal gene expression clusters for ethylene (A) and shade (D); 791
hypocotyl growth curve for ethylene (B) and shade (E); clusters with gene expression 792
matching hypocotyl growth kinetics in ethylene (C) and shade (F). Heatmap for 793
temporal clusters (A, D) based on the log2FC at the three microarray time points 794
(grey box represents dark; arrows indicate heatmap time-points and treatments 795
include control (open circle), ethylene (closed circle), shade (open triangle)). Yellow 796
denotes up-regulation and blue denotes down-regulation. Two gene expression 797
clusters (C, F) with mean log2FC temporal pattern resembling the hypocotyl length 798
kinetics (B, E) were named positive clusters. G, Heatmap for hypergeometric 799
enrichment of selected MapMan bins for temporal clusters under ethylene and 800
shade. Horizontal-axis denotes the cluster number. More intense colours indicate 801
higher statistical significance. Grey color indicates non-significant score or absence 802
of genes in the bin. H, Venn intersection for positive clusters (clusters with gene 803
expression pattern matching the hypocotyl length kinetics) from ethylene and shade 804
to obtain “Common Up” genes. I, Venn intersection for negative clusters (mirror 805
image to Positive clusters) from ethylene and shade to obtain “Common Down” 806
genes. 807
Figure 4. Gene Ontology (GO) enrichment analysis using GeneCodis for positive 808
and negative clusters. Adjusted p-values for statistical significance of GO enrichment 809
were converted into negative logarithm score (base10) (> 1.3 are considered 810
significant). Heatmap colours denote this score and more intense colours indicate 811
higher statistical significance. White color indicates non-significant score or absence 812
of genes in the GO terms. 813
814
Figure 5. Heatmap of (A) Common Up and (B) Common Down set of genes 815
classified into the categories: transcriptional regulator, hormone metabolism, 816
www.plantphysiol.org on July 4, 2016 - Published by www.plantphysiol.orgDownloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

26
signalling and cell wall genes after application of a log2fold-change (log2FC) filter. 817
Log2FC at the three time-points 1.5 h, 13.5 h, 25.5 h for ethylene and shade 818
microarray dataset identified in the color scheme of the heatmap. Genes were 819
categorized according to Mapman bin gene classification (shown on left side). 820
821
Figure 6. Hypocotyl length measurements for (A) athb28 (B) ibl1 (C) IBL1-OE 822
following 96 h of control, ethylene and shade treatments. Data represents mean ± 823
S.E. (n=30 seedlings). Different letters above the bars indicate significant differences 824
(2-way ANOVA followed by Tukey’s HSD Post-Hoc pairwise comparison). 825
826
Figure 7. A, Identification of enriched hormonal signatures in the ethylene- and 827
shade-induced Arabidopsis transcriptome. Ethylene and shade-induced hypocotyl 828
and cotyledon transcriptomes were analyzed for hormonal signatures using the tool 829
Hormonometer (Valdorsky et al., 2009) to establish correlations with expression data 830
in an established hormonal transcriptome database. Positive correlations were 831
colored yellow and negative correlations blue. Significant correlations were identified 832
with absolute correlation values of 0.3 and higher. Numbers in the cells represent the 833
exact correlation values. Rows denote hormone treatments that are indicated by the 834
name of the hormone and the duration of hormone treatment. Columns denote 835
ethylene and shade transcriptome in the hypocotyl and cotyledon at the three time-836
points of tissue harvest. The magnitude of correlation in gene expression is indicated 837
by the color scale at top right side. B, Effect of auxin transport inhibitor NPA (25 µM), 838
IAA (10 µM), NPA (25 µM) + IAA (10 µM) and auxin perception inhibitor PEO-IAA 839
(100 µM) on GUS staining of the pIAA19:GUS lines. For NPA and PEO-IAA effect in 840
GUS assay, seedlings were exposed to 2 days of treatment conditions. Arabidopsis 841
(Col-0) seedlings were treated with chemical inhibitors for (C) auxin transport (NPA), 842
(D) auxin biosynthesis (Yucasin), (E) auxin perception (PEO-IAA), (F) 843
brassinosteroid biosynthesis (BRZ) and (G) gibberellin biosynthesis (PBZ) at the 844
indicated concentrations in the legend and length was measured following 96h of 845
ethylene and shade. Hypocotyl length was measured following 96 h of ethylene and 846
shade. Mean ± S.E. was calculated for 30 seedlings. Different letters above the bars 847
indicate significant differences from a 2-way ANOVA followed by Tukey’s HSD Post-848
Hoc pairwise comparison. Abbreviations: PEO-IAA is α-(phenylethyl-2-one)-indole-3-849
www.plantphysiol.org on July 4, 2016 - Published by www.plantphysiol.orgDownloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

27
acetic acid; NPA is 1-N-Naphthylphthalamic acid; BRZ is Brassinazole; PBZ is 850
Paclobutrazol. 851
852
Figure 8. Hypocotyl elongation response in response to ethylene and shade in (B)–853
(D) auxin, (E)–(G) gibberellin (GA) and (H)–(J) brassinosteroid (BR) mutants. Mean 854
± S.E. was calculated for 30 seedlings. Different letters above the bars indicate 855
significant differences from a 2-way ANOVA followed by Tukey’s HSD Post-Hoc 856
pairwise comparison. 857
858
LITERATURE CITED 859 860 Abdi, H., and Williams, L. J. (2010). Principal component analysis. Wiley 861
Interdisciplinary Reviews: Computational Statistics, 2(4), 433-459. 862 Achard, P., Cheng, H., Grauwe, L. De, Decat, J., Schoutteten, H., Moritz, T., 863
Straeten, D. Van Der, Peng, J., and Harberd, N.P. (2006). Integration of Plant 864 Responses to Environmentally Activated Phytohormonal Signals. Science 311: 865 91–94. 866
Asami, T., Min, Y.K., Nagata, N., Yamagishi, K., Takatsuto, S., Fujioka, S., 867 Murofushi, N., Yamaguchi, I., and Yoshida, S. (2000). Characterization of 868 brassinazole, a triazole-type brassinosteroid biosynthesis inhibitor. Plant 869 Physiol. 123: 93–100. 870
Azpiroz, R., Wu, Y., LoCascio, J.C., and Feldmann, K.A. (1998). An Arabidopsis 871 brassinosteroid-dependent mutant is blocked in cell elongation. The Plant Cell 872 10: 219–230. 873
Bai, M.Y., Fan, M., Oh, E., and Wang, Z.Y. (2012) A triple helix-loop-helix/basic 874 helix-loop-helix cascade controls cell elongation downstream of multiple 875 hormonal and environmental signaling pathways in Arabidopsis. Plant Cell. 24: 876 4917-4929 877
Barboza, L., Effgen, S., Alonso-blanco, C., Kooke, R., Keurentjes, J.J.B., and 878 Koornneef, M. (2013). Arabidopsis semidwarfs evolved from independent 879 mutations in GA20ox1, ortholog to green revolution dwarf alleles in rice and 880 barley. Proc. Natl. Acad. Sci. U. S. A. 110: 15818–15823. 881
Binder, B.M., Mortimore, L.A., Stepanova, A.N., Ecker, J.R., and Bleecker, A.B. 882 (2004). Short-Term Growth Responses to Ethylene in Arabidopsis. Plant 883 Physiol. 136: 2921–2927. 884
Blilou, I., Xu, J., Wildwater, M., Willemsen, V., Paponov, I., Friml J, Heidstra R., 885 Aida M., Palme K. & Scheres, B. (2005). The PIN auxin efflux facilitator 886 network controls growth and patterning in Arabidopsis roots. Nature, 433(7021), 887 39-44. 888
Cagnola, J. I., Ploschuk, E., Benech-Arnold, T., Finlayson, S. A., and Casal, J. 889 J. (2012). Stem transcriptome reveals mechanisms to reduce the energetic cost 890 of shade-avoidance responses in tomato. Plant Physiol., 160(2), 1110-1119. 891
Capella, M., Ribone, P. A., Arce, A. L., and Chan, R. L. (2015). Arabidopsis 892 thaliana HomeoBox 1 (AtHB1), a Homedomain‐Leucine Zipper I (HD‐Zip I) 893
www.plantphysiol.org on July 4, 2016 - Published by www.plantphysiol.orgDownloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

28
transcription factor, is regulated by PHYTOCHROME‐INTERACTING FACTOR 894 1 to promote hypocotyl elongation. New Phytologist, 207, 669-682. 895
Casal, J.J. (2013). Photoreceptor signaling networks in plant responses to shade. 896 Annu. Rev. Plant Biol. 64: 403–27. 897
Casal, J.J. (2012). Shade avoidance. Arabidopsis Book 10: e0157. 898 Chae, K., Isaacs, C.G., Reeves, P.H., Maloney, G.S., Muday, G.K., Nagpal, P., 899
and Reed, J.W. (2012). Arabidopsis SMALL AUXIN UP RNA63 promotes 900 hypocotyl and stamen filament elongation. Plant J. 71: 684–697. 901
Chapman, E.J., Greenham, K., Castillejo, C., Sartor, R., Bialy, A., Sun, T.P., and 902 Estelle, M. (2012). Hypocotyl transcriptome reveals auxin regulation of growth-903 promoting genes through GA-dependent and -independent pathways. PLoS 904 One 7: e36210. 905
De Smet, I., Lau, S., Ehrismann, J.S., Axiotis, I., Kolb, M., Kientz, M., Weijers, 906 D., and Jurgens, G. (2013). Transcriptional repression of BODENLOS by HD-907 ZIP transcription factor HB5 in Arabidopsis thaliana. J. Exp. Bot. 64: 3009–3019. 908
Djakovic-Petrovic, T., Wit, M. De, Voesenek, L.A.C.J., and Pierik, R. (2007). 909 DELLA protein function in growth responses to canopy signals. Plant J. 51: 117–910 126. 911
Eisen, M.B., Spellman, P.T., Brown, P.O., and Botstein, D. (1998). Cluster 912 analysis and display of genome-wide expression patterns. Proc. Natl. Acad. Sci. 913 U. S. A. 95: 14863–14868. 914
Endo, M., Nakamura, S., Araki, T., Mochizuki, N., and Nagatani, A. (2005). 915 Phytochrome B in the Mesophyll Delays Flowering by Suppressing 916 FLOWERING LOCUS T Expression in Arabidopsis Vascular Bundles. The Plant 917 Cell 17: 1941–1952. 918
Estelle, M. (1998). Polar Auxin Transport: New Support for an Old Model. The Plant 919 Cell 10: 1775–1778. 920
Fan, M., Bai, M. Y., Kim, J. G., Wang, T., Oh, E., Chen, L., Park, C.H., Son S., 921 Kim S., Mudgett M.B. and Wang, Z. Y. (2014). The bHLH transcription factor 922 HBI1 mediates the trade-off between growth and pathogen-associated 923 molecular pattern–triggered immunity in Arabidopsis. The Plant Cell Online, 924 26(2), 828-841. 925
Franklin, K. A., Lee, S.H., Patel, D., Kumar, S.V., Spartz, A.K., Gu, C., Ye, S., Yu, 926 P., Breen, G., Cohen, J.D., Wigge, P.A., and Gray, W.M. (2011). 927 Phytochrome-interacting factor 4 (PIF4) regulates auxin biosynthesis at high 928 temperature. Proc. Natl. Acad. Sci. U. S. A. 108: 20231–5. 929
Franklin, K. A. (2008). Shade avoidance. New Phytol. 179: 930–944. 930 Gendreau, E., Traas, J., Desnos, T., Grandjean, O., Caboche, M., and Höfte, H. 931
(1997). Cellular basis of hypocotyl growth in Arabidopsis thaliana. Plant Physiol. 932 114: 295–305. 933
Hattori, Y., Nagai, K., Furukawa, S., Song, X., Kawano, R., Sakakibara, H., Wu, 934 J., Matsumoto, T., Yoshimura, A., Kitano, H., Matsuoka, M., Mori, H., 935 Ashikari, M. (2009). The ethylene response factors SNORKEL1 and 936 SNORKEL2 allow rice to adapt to deep water. Nature 460: 1026–1030. 937
Hayashi, K., Tan, X., Zheng, N., Hatate, T., Kimura, Y., Kepinski, S., and Nozaki, 938 H. (2008). Small-molecule agonists and antagonists of F-box protein-substrate 939 interactions in auxin perception and signaling. Proc. Natl. Acad. Sci. U. S. A. 940 105: 5632–5637. 941
www.plantphysiol.org on July 4, 2016 - Published by www.plantphysiol.orgDownloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

29
Henriques, R., Bögre, L., Horváth, B., and Magyar, Z. (2014). Balancing act: 942 Matching growth with environment by the TOR signalling pathway. J. Exp. Bot. 943 65: 2691–2701. 944
Hisamatsu, T., King, R.W., Helliwell, C. a, and Koshioka, M. (2005). The 945 involvement of Gibberellin 20-Oxidase genes in phytochrome-regulated petiole 946 elongation of Arabidopsis. Plant Physiol. 138: 1106–1116. 947
Hong, S.Y., Kim, O.K., Kim, S.G., Yang, M.S., and Park, C.M. (2011). Nuclear 948 import and DNA binding of the ZHD5 transcription factor is modulated by a 949 competitive peptide inhibitor in Arabidopsis. J. Biol. Chem. 286: 1659–1668. 950
Hu, W. and Ma, H. (2006). Characterization of a novel putative zinc finger gene 951 MIF1: Involvement in multiple hormonal regulation of Arabidopsis development. 952 Plant J. 45: 399–422. 953
Hyun, Y. and Lee, I. (2006). KIDARI, encoding a non-DNA Binding bHLH protein, 954 represses light signal transduction in Arabidopsis thaliana. Plant Mol. Biol. 61: 955 283–96. 956
Iqbal, N., Khan, N. A., Nazar, R., and da Silva, J. A. T. (2012). Ethylene-stimulated 957 photosynthesis results from increased nitrogen and sulfur assimilation in 958 mustard types that differ in photosynthetic capacity. Environ. Exp. Bot. 78: 84–959 90. 960
Kazan, K. and Manners, J.M. (2013). MYC2: The master in action. Mol. Plant 6: 961 686–703. 962
Keuskamp, D.H., Pollmann, S., Voesenek, L.A.C.J., Peeters, A.J.M., and Pierik, 963 R. (2010). Auxin transport through PIN-FORMED 3 (PIN3) controls shade 964 avoidance and fitness during competition. Proc. Natl. Acad. Sci. U. S. A. 107: 965 22740–22744. 966
Keuskamp, D.H., Sasidharan, R., Vos, I., Peeters, A.J.M., Voesenek, L.A.C.J., 967 and Pierik, R. (2011). Blue-light-mediated shade avoidance requires combined 968 auxin and brassinosteroid action in Arabidopsis seedlings. Plant J. 67: 208–217. 969
Khanna, R., Shen, Y., Marion, C.M., Tsuchisaka, A., Theologis, A., Schafer, E., 970 and Quail, P.H. (2007). The basic helix-loop-helix transcription factor PIF5 acts 971 on ethylene biosynthesis and phytochrome signaling by distinct mechanisms. 972 Plant Cell. 19: 3915-3929. 973
Kim, B.C., Soh, M.S., Hong, S.H., Furuya, M., and Nam, H.G. (1998). 974 Photomorphogenic development of the Arabidopsis shy2-1D mutation and its 975 interaction with phytochromes in darkness. Plant J. 15: 61–68. 976
Kunihiro, A., Yamashino, T., Nakamichi, N., Niwa, Y., Nakanishi, H., and 977 Mizuno, T. (2011). PHYTOCHROME-INTERACTING FACTOR 4 and 5 (PIF4 978 and PIF5) activate the Homeobox ATHB2 and auxin-inducible IAA29 genes in 979 the coincidence mechanism underlying photoperiodic control of plant growth of 980 Arabidopsis thaliana. The Plant Cell Physiol. 52: 1315–1329. 981
Lee, Y.K., Kim, G., Kim, I., Park, J., Kwak, S., Choi, G., and Chung, W. (2006). 982 LONGIFOLIA1 and LONGIFOLIA2, two homologous genes, regulate 983 longitudinal cell elongation in Arabidopsis. Development 133: 4305–4314. 984
Leivar, P. and Monte, E. (2014). PIFs: systems integrators in plant development. 985 The Plant Cell 26: 56–78. 986
Li, L. et al. (2012). Linking photoreceptor excitation to changes in plant architecture. 987 Genes Dev. 26: 785–790. 988
Lilley, J.L.S., Gee, C.W., Sairanen, I., Ljung, K., and Nemhauser, J.L. (2012). An 989 Endogenous Carbon-Sensing Pathway Triggers Increased Auxin Flux and 990 Hypocotyl Elongation. Plant Physiol. 160: 2261–2270. 991
www.plantphysiol.org on July 4, 2016 - Published by www.plantphysiol.orgDownloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

30
Mickelbart, M. V., Hasegawa, P.M., and Bailey-Serres, J. (2015). Genetic 992 mechanisms of abiotic stress tolerance that translate to crop yield stability. Nat. 993 Rev. Genet. 16: 237–251. 994
Morelli, G., Ruberti, I., Molecolare, B., Sapienza, L., and Moro, P.A. (2000). 995 Update on Light Signaling Shade Avoidance Responses. Driving Auxin along 996 Lateral Routes. Plant Physiol. 122: 621–626. 997
Neff, M.M., Nguyen, S.M., Malancharuvil, E.J., Fujioka, S., Noguchi, T., Seto, H., 998 Tsubuki, M., Honda, T., Takatsuto, S., Yoshida, S., and Chory, J. (1999). 999 BAS1: A gene regulating brassinosteroid levels and light responsiveness in 1000 Arabidopsis. Proc. Natl. Acad. Sci. 96: 15316–15323. 1001
Nemhauser, J.L., Mockler, T.C., and Chory, J. (2004). Interdependency of 1002 brassinosteroid and auxin signaling in Arabidopsis. PLoS Biol. 2: E258. 1003
Nishimura, T., Hayashi, K.I., Suzuki, H., Gyohda, A., Takaoka, C., Sakaguchi, Y., 1004 Matsumoto, S., Kasahara, H., Sakai, T., Kato, J.I., Kamiya, Y., and Koshiba, 1005 T. (2014). Yucasin is a potent inhibitor of YUCCA, a key enzyme in auxin 1006 biosynthesis. Plant J. 77: 352–366. 1007
Nomura, T., Kushiro, T., Yokota, T., Kamiya, Y., Bishop, G.J., and Yamaguchi, 1008 S. (2005). The last reaction producing brassinolide is catalyzed by cytochrome 1009 P-450s, CYP85A3 in tomato and CYP85A2 in Arabidopsis. J. Biol. Chem. 280: 1010 17873–17879. 1011
Nozue, K., Tat, A. V., Kumar Devisetty, U., Robinson, M., Mumbach, M.R., 1012 Ichihashi, Y., Lekkala, S., and Maloof, J.N. (2015). Shade Avoidance 1013 Components and Pathways in Adult Plants Revealed by Phenotypic Profiling. 1014 PLOS Genet. 11: e1004953. 1015
Osakabe, Y., Osakabe, K., Shinozaki, K., and Tran, L. S. P. (2014). Response of 1016 plants to water stress. Front. Plant Sci. 5(86): 10-3389. 1017
Park, E., Park, J., Kim, J., Nagatani, A., Lagarias, J.C., and Choi, G. (2012). 1018 Phytochrome B inhibits binding of phytochrome-interacting factors to their target 1019 promoters. Plant J. 72: 537–546. 1020
Petrásek, J., Cerná, A., Schwarzerová, K., Elckner, M., Morris, D. A., and 1021 Zazímalová, E. (2003). Do phytotropins inhibit auxin efflux by impairing vesicle 1022 traffic? Plant Physiol. 131: 254–263. 1023
Pfeiffer, A., Shi, H., Tepperman, J.M., Zhang, Y., and Quail, P.H. (2014). 1024 Combinatorial Complexity in a Transcriptionally Centered Signaling Hub in 1025 Arabidopsis. Mol. Plant 7: 1598–1618. 1026
Philippar, K., Ivashikina, N., Ache, P., Christian, M., Lüthen, H., Palme, K., and 1027 Hedrich, R. (2004). Auxin activates KAT1 and KAT2, two K+-channel genes 1028 expressed in seedlings of Arabidopsis thaliana. Plant J. 37: 815–827. 1029
Pierik, R., Cuppens, M.L., Voesenek, L.A.C.J. and Visser, E.J. (2004) Interactions 1030 between ethylene and gibberellins in phytochrome-mediated shade avoidance 1031 responses in tobacco. Plant Physiol. 136: 2928-2936. 1032
Pierik, R., Djakovic-Petrovic, T., Keuskamp, D.H., de Wit, M., and Voesenek, 1033 L.A.C.J. (2009). Auxin and ethylene regulate elongation responses to neighbor 1034 proximity signals independent of gibberellin and DELLA proteins in Arabidopsis. 1035 Plant Physiol. 149: 1701–1712. 1036
Pierik, R., Millenaar, F.F., Peeters, A. J.M., and Voesenek, L.A.C.J. (2005). New 1037 perspectives in flooding research: The use of shade avoidance and Arabidopsis 1038 thaliana. Ann. Bot. 96: 533–540. 1039
Pierik, R. and Testerink, C. (2014). The art of being flexible: how to escape from 1040 shade, salt and drought. Plant Physiol. 166: 5–22. 1041
www.plantphysiol.org on July 4, 2016 - Published by www.plantphysiol.orgDownloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

31
Pierik, R. and De Wit, M. (2014). Shade avoidance: Phytochrome signalling and 1042 other aboveground neighbour detection cues. J. Exp. Bot. 65: 2815–2824. 1043
Procko, C., Crenshaw, C.M., Ljung, K., Noel, J.P., and Chory, J. (2014). 1044 Cotyledon-Generated Auxin Is Required for Shade-Induced Hypocotyl Growth in 1045 Brassica rapa. Plant Physiol. 165: 1285–1301. 1046
Quint, M., Delker, C., Franklin, K. A., Wigge, P. A., Halliday, K. J., and van 1047 Zanten, M. (2016). Molecular and genetic control of plant 1048 thermomorphogenesis. Nature Plants. 2: 15190. 1049
Rademacher, W. (2000). GROWTH RETARDANTS: Effects on Gibberellin 1050 Biosynthesis and Other Metabolic Pathways. Annu. Rev. Plant Physiol. Plant 1051 Mol. Biol. 51: 501–531. 1052
Rieu, I., Ruiz-Rivero, O., Fernandez-Garcia, N., Griffiths, J., Powers, S.J., Gong, 1053 F., Linhartova, T., Eriksson, S., Nilsson, O., Thomas, S.G., Phillips, A.L., 1054 and Hedden, P. (2008). The gibberellin biosynthetic genes AtGA20ox1 and 1055 AtGA20ox2 act, partially redundantly, to promote growth and development 1056 throughout the Arabidopsis life cycle. Plant J. 53: 488–504. 1057
Ruegger, M., Dewey, E., Gray, W.M., Hobbie, L., Turner, J., and Estelle, M. 1058 (1998). The TIR1 protein of Arabidopsis functions in auxin response and is 1059 related to human SKP2 and yeast Grr1p. Genes Dev. 12: 198–207. 1060
Sasidharan, R., Chinnappa, C.C., Staal, M., Elzenga, J.T.M., Yokoyama, R., 1061 Nishitani, K., Voesenek, L.A.C.J., and Pierik, R. (2010). Light quality-1062 mediated petiole elongation in Arabidopsis during shade avoidance involves cell 1063 wall modification by xyloglucan endotransglucosylase/hydrolases. Plant Physiol. 1064 154: 978–990. 1065
Sasidharan, R. and Voesenek, L.A.C.J. (2015). Ethylene-mediated acclimations to 1066 flooding stress. Plant Physiol.: pp.00387. 1067
Smalle, J., Haegman, M., Kurepa, J., Van Montagu M, and Straeten, D. V (1997). 1068 Ethylene can stimulate Arabidopsis hypocotyl elongation in the light. Proc. Natl. 1069 Acad. Sci. U. S. A. 94: 2756–2761. 1070
Spartz, A.K., Lee, S.H., Wenger, J.P., Gonzalez, N., Itoh, H., Inzé, D., Peer, W.A., 1071 Murphy, A.S., Overvoorde, P.J., and Gray, W.M. (2012). The SAUR19 1072 subfamily of SMALL AUXIN UP RNA genes promote cell expansion. Plant J. 70: 1073 978–990. 1074
Stepanova, A.N., Robertson-Hoyt, J., Yun, J., Benavente, L.M., Xie, D.Y., 1075 Doležal, K., Schlereth, A., Jürgens, G., and Alonso, J.M. (2008). TAA1-1076 Mediated Auxin Biosynthesis Is Essential for Hormone Crosstalk and Plant 1077 Development. Cell 133: 177–191. 1078
Sulpice, R., Flis, A., Ivakov, A. A., Apelt, F., Krohn, N., Encke, B., Abel, C., Feil, 1079 R., Lunn, J.E., and Stitt, M. (2014). Arabidopsis coordinates the diurnal 1080 regulation of carbon allocation and growth across a wide range of Photoperiods. 1081 Mol. Plant 7: 137–155. 1082
Sun, J., Qi, L., Li, Y., Chu, J., and Li, C. (2012). PIF4-mediated activation of 1083 YUCCA8 expression integrates temperature into the auxin pathway in regulating 1084 Arabidopsis hypocotyl growth. PLoS Genet. 8: e1002594. 1085
Tabas-Madrid, D., Nogales-Cadenas, R., and Pascual-Montano, A. (2012). 1086 GeneCodis 3: a non-redundant and modular enrichment analysis tool for 1087 functional genomics. Nucleic Acids Res. 40: W478–W483. 1088
Talon, M., Koornneef, M., and Zeevaart, J. A. D. (1990). Accumulation of C19-1089 gibberellins in the gibberellin-insensitive dwarf mutant gai of Arabidopsis 1090 thaliana (L.) Heynh. Planta 182: 501–505. 1091
www.plantphysiol.org on July 4, 2016 - Published by www.plantphysiol.orgDownloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

32
Tanaka, S., Nakamura, S., Mochizuki, N., and Nagatani, A. (2002). Phytochrome 1092 in cotyledons regulates the expression of genes in the hypocotyl through auxin-1093 dependent and -independent pathways. The Plant Cell Physiol. 43: 1171–1181. 1094
Tao, Y., Ferrer, J.L., Ljung, K., Pojer, F., Hong, F., Long, J.A., Li, L., Moreno, 1095 J.E., Bowman, M.E., Ivans, L.J., Cheng, Y., Lim, J., Zhao, Y., Ballaré, C.L., 1096 Sandberg, G., Noel, J.P. and Chory, J. (2008). Rapid Synthesis of Auxin via a 1097 New Tryptophan-Dependent Pathway Is Required for Shade Avoidance in 1098 Plants. Cell 133: 164–176. 1099
Tholen, D., Pons, T.L., Voesenek, L.A.C.J., and Poorter, H. (2007). Ethylene 1100 insensitivity results in down-regulation of rubisco expression and photosynthetic 1101 capacity in tobacco. Plant Physiol. 144: 1305–1315. 1102
Turk, E.M. et al. (2005). BAS1 and SOB7 act redundantly to modulate Arabidopsis 1103 photomorphogenesis via unique brassinosteroid inactivation mechanisms. Plant 1104 J. 42: 23–34. 1105
Unterholzner, S.J., Rozhon, W., Papacek, M., Ciomas, J., Lange, T., Kugler, 1106 K.G., Mayer, K.F., Sieberer, T., and Poppenberger, B. (2015). 1107 Brassinosteroids Are Master Regulators of Gibberellin Biosynthesis in 1108 Arabidopsis. The Plant Cell 27: 2261–2272. 1109
Van Esse, G.W., van Mourik, S., Stigter, H., ten Hove, C. a., Molenaar, J., and de 1110 Vries, S.C. (2012). A Mathematical Model for BRASSINOSTEROID 1111 INSENSITIVE1-Mediated Signaling in Root Growth and Hypocotyl Elongation. 1112 Plant Physiol. 160: 523–532. 1113
Vandenbussche, F., Pierik, R., Millenaar, F.F., Voesenek, L.A.C.J., and Van Der 1114 Straeten, D. (2005). Reaching out of the shade. Curr. Opin. Plant Biol. 8: 462–1115 468. 1116
van Veen, H., Mustroph, A., Barding, G.A., Vergeer-van Eijk, M., Welschen-1117 Evertman, R.A.M., Pedersen, O., Visser, E.J.W., Larive, C.K., Pierik, R., 1118 Bailey-Serres, J., Voesenek, L.A.C.J., and Sasidharan, R. (2013). Two 1119 Rumex species from contrasting hydrological niches regulate flooding tolerance 1120 through distinct mechanisms. The Plant Cell 25: 4691–707. 1121
Voesenek L.A.C.J.; and Bailey-serres, J. (2015). Flood adaptive traits and 1122 processes : an overview. New Phytol. 206: 57–73. 1123
Volodarsky, D., Leviatan, N., Otcheretianski, A., and Fluhr, R. (2009). 1124 HORMONOMETER: a tool for discerning transcript signatures of hormone 1125 action in the Arabidopsis transcriptome. Plant Physiol. 150: 1796–1805. 1126
Warnasooriya, S.N. and Montgomery, B.L. (2009). Detection of spatial-specific 1127 phytochrome responses using targeted expression of biliverdin reductase in 1128 Arabidopsis. Plant Physiol. 149: 424–33. 1129
Wilson, R.N., Heckman, J.W., and Somerville, C.R. (1992). Gibberellin Is Required 1130 for Flowering in Arabidopsis thaliana under Short Days. Plant Physiol. 100: 403–1131 408. 1132
Zhiponova, M.K., Morohashi, K., Vanhoutte, I., Machemer-Noonan, K., 1133 Revalska, M., Van Montagu, M., Grotewold, E., and Russinova, E. (2014). 1134 Helix-loop-helix/basic helix-loop-helix transcription factor network represses cell 1135 elongation in Arabidopsis through an apparent incoherent feed-forward loop. 1136 Proc. Natl. Acad. Sci. U. S. A. 111: 2824–9. 1137
Zheng, Z., Guo, Y., Novak, O., Chen, W., Ljung, K., Noel, J.P., and Chory, J. 1138 (2016). Local auxin metabolism regulates environment-induced hypocotyl 1139 elongation. Nat. Plants. 21: 16025. doi: 10.1038/nplants.2016.25 1140
www.plantphysiol.org on July 4, 2016 - Published by www.plantphysiol.orgDownloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

33
Zhong, S., Shi, H., Xue, C., Wang, L., Xi, Y., Li, J., Quail, P.H., Deng, X.W., and 1141 Guo, H. (2012). A molecular framework of light-controlled phytohormone action 1142 in Arabidopsis. Curr. Biol. 22: 1530–1535. 1143
1144 1145 1146 1147 1148 1149
www.plantphysiol.org on July 4, 2016 - Published by www.plantphysiol.orgDownloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

www.plantphysiol.org on July 4, 2016 - Published by www.plantphysiol.orgDownloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

www.plantphysiol.org on July 4, 2016 - Published by www.plantphysiol.orgDownloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.
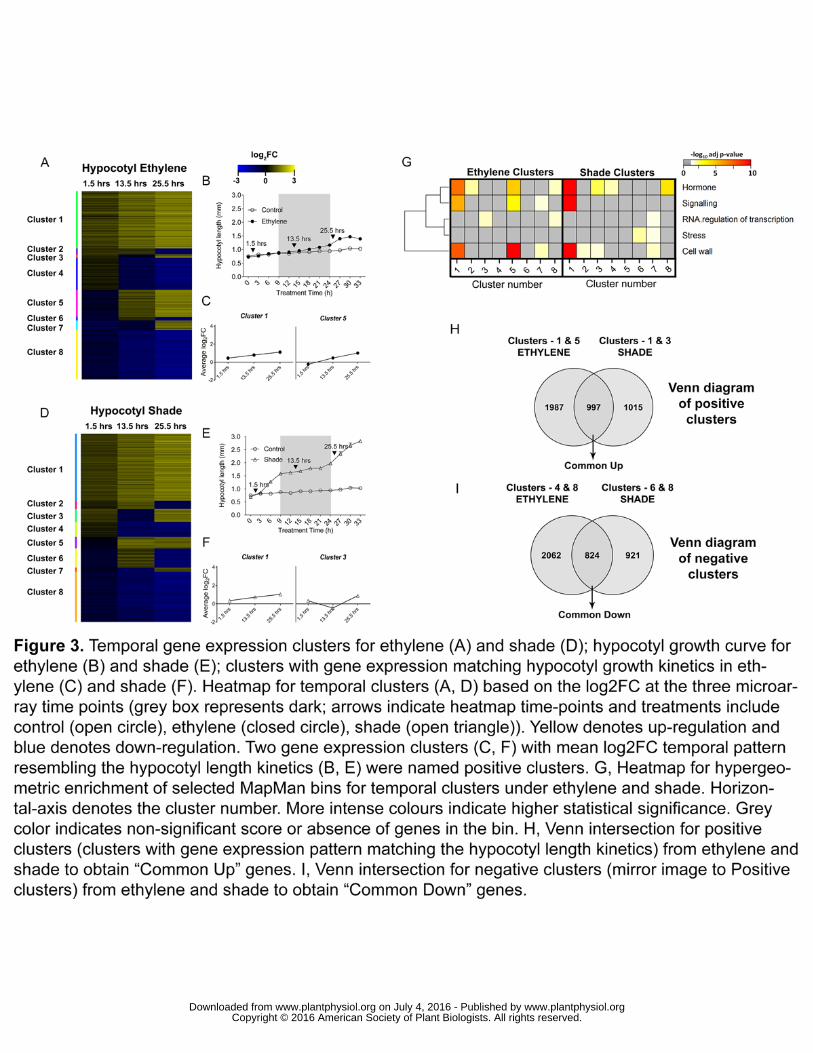
www.plantphysiol.org on July 4, 2016 - Published by www.plantphysiol.orgDownloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

www.plantphysiol.org on July 4, 2016 - Published by www.plantphysiol.orgDownloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

www.plantphysiol.org on July 4, 2016 - Published by www.plantphysiol.orgDownloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

www.plantphysiol.org on July 4, 2016 - Published by www.plantphysiol.orgDownloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.
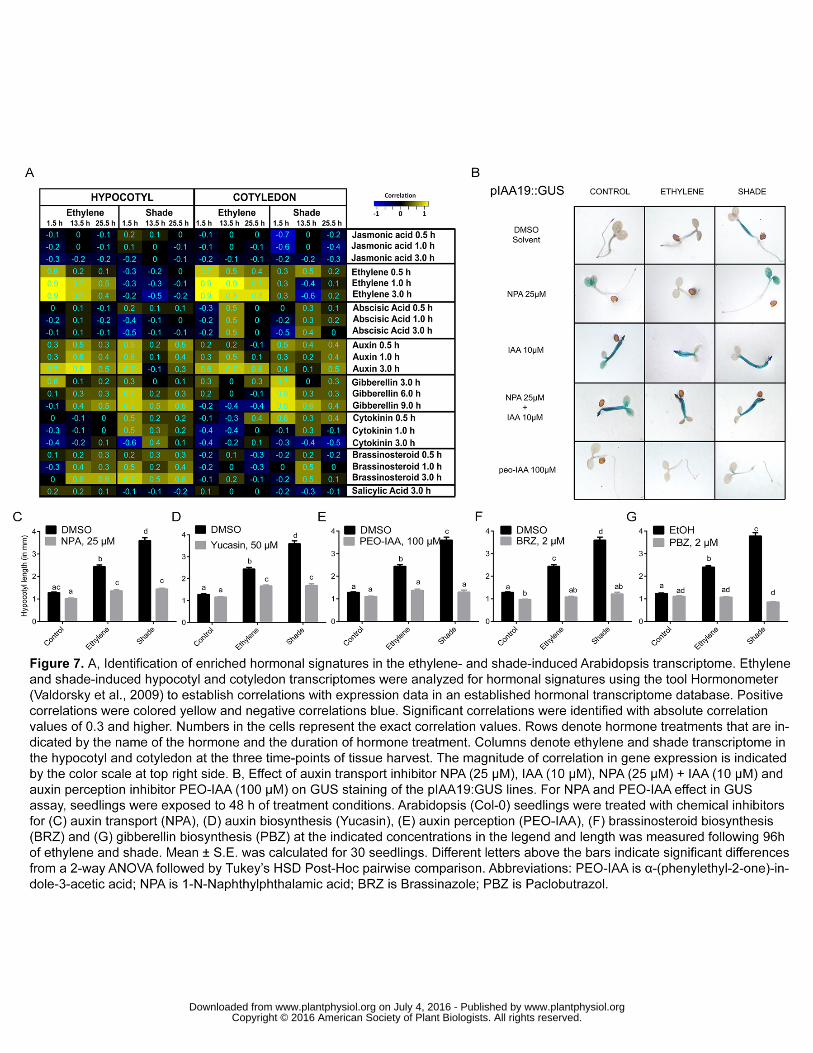
www.plantphysiol.org on July 4, 2016 - Published by www.plantphysiol.orgDownloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.
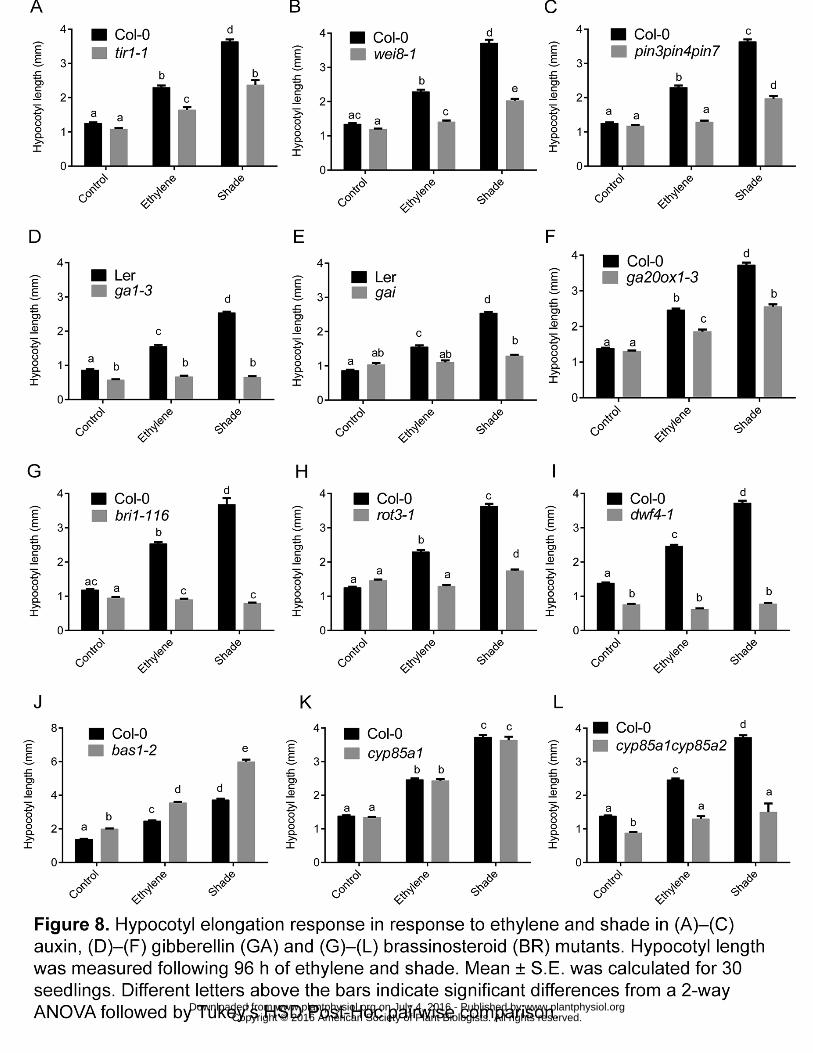
www.plantphysiol.org on July 4, 2016 - Published by www.plantphysiol.orgDownloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

Parsed CitationsAbdi, H., and Williams, L. J. (2010). Principal component analysis. Wiley Interdisciplinary Reviews: Computational Statistics, 2(4),433-459.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Achard, P., Cheng, H., Grauwe, L. De, Decat, J., Schoutteten, H., Moritz, T., Straeten, D. Van Der, Peng, J., and Harberd, N.P.(2006). Integration of Plant Responses to Environmentally Activated Phytohormonal Signals. Science 311: 91-94.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Asami, T., Min, Y.K., Nagata, N., Yamagishi, K., Takatsuto, S., Fujioka, S., Murofushi, N., Yamaguchi, I., and Yoshida, S. (2000).Characterization of brassinazole, a triazole-type brassinosteroid biosynthesis inhibitor. Plant Physiol. 123: 93-100.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Azpiroz, R., Wu, Y., LoCascio, J.C., and Feldmann, K.A. (1998). An Arabidopsis brassinosteroid-dependent mutant is blocked in cellelongation. The Plant Cell 10: 219-230.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Bai, M.Y., Fan, M., Oh, E., and Wang, Z.Y. (2012) A triple helix-loop-helix/basic helix-loop-helix cascade controls cell elongationdownstream of multiple hormonal and environmental signaling pathways in Arabidopsis. Plant Cell. 24: 4917-4929
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Barboza, L., Effgen, S., Alonso-blanco, C., Kooke, R., Keurentjes, J.J.B., and Koornneef, M. (2013). Arabidopsis semidwarfsevolved from independent mutations in GA20ox1, ortholog to green revolution dwarf alleles in rice and barley. Proc. Natl. Acad.Sci. U. S. A. 110: 15818-15823.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Binder, B.M., Mortimore, L.A., Stepanova, A.N., Ecker, J.R., and Bleecker, A.B. (2004). Short-Term Growth Responses to Ethylenein Arabidopsis. Plant Physiol. 136: 2921-2927.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Blilou, I., Xu, J., Wildwater, M., Willemsen, V., Paponov, I., Friml J, Heidstra R., Aida M., Palme K. & Scheres, B. (2005). The PINauxin efflux facilitator network controls growth and patterning in Arabidopsis roots. Nature, 433(7021), 39-44.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Cagnola, J. I., Ploschuk, E., Benech-Arnold, T., Finlayson, S. A., and Casal, J. J. (2012). Stem transcriptome reveals mechanisms toreduce the energetic cost of shade-avoidance responses in tomato. Plant Physiol., 160(2), 1110-1119.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Capella, M., Ribone, P. A., Arce, A. L., and Chan, R. L. (2015). Arabidopsis thaliana HomeoBox 1 (AtHB1), a Homedomain-LeucineZipper I (HD-Zip I) transcription factor, is regulated by PHYTOCHROME-INTERACTING FACTOR 1 to promote hypocotylelongation. New Phytologist, 207, 669-682.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Casal, J.J. (2013). Photoreceptor signaling networks in plant responses to shade. Annu. Rev. Plant Biol. 64: 403-27.Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Casal, J.J. (2012). Shade avoidance. Arabidopsis Book 10: e0157.Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Chae, K., Isaacs, C.G., Reeves, P.H., Maloney, G.S., Muday, G.K., Nagpal, P., and Reed, J.W. (2012). Arabidopsis SMALL AUXIN UPRNA63 promotes hypocotyl and stamen filament elongation. Plant J. 71: 684-697.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
www.plantphysiol.org on July 4, 2016 - Published by www.plantphysiol.orgDownloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

Chapman, E.J., Greenham, K., Castillejo, C., Sartor, R., Bialy, A., Sun, T.P., and Estelle, M. (2012). Hypocotyl transcriptome revealsauxin regulation of growth-promoting genes through GA-dependent and -independent pathways. PLoS One 7: e36210.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
De Smet, I., Lau, S., Ehrismann, J.S., Axiotis, I., Kolb, M., Kientz, M., Weijers, D., and Jurgens, G. (2013). Transcriptional repressionof BODENLOS by HD-ZIP transcription factor HB5 in Arabidopsis thaliana. J. Exp. Bot. 64: 3009-3019.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Djakovic-Petrovic, T., Wit, M. De, Voesenek, L.A.C.J., and Pierik, R. (2007). DELLA protein function in growth responses to canopysignals. Plant J. 51: 117-126.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Eisen, M.B., Spellman, P.T., Brown, P.O., and Botstein, D. (1998). Cluster analysis and display of genome-wide expression patterns.Proc. Natl. Acad. Sci. U. S. A. 95: 14863-14868.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Endo, M., Nakamura, S., Araki, T., Mochizuki, N., and Nagatani, A. (2005). Phytochrome B in the Mesophyll Delays Flowering bySuppressing FLOWERING LOCUS T Expression in Arabidopsis Vascular Bundles. The Plant Cell 17: 1941-1952.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Estelle, M. (1998). Polar Auxin Transport: New Support for an Old Model. The Plant Cell 10: 1775-1778.Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Fan, M., Bai, M. Y., Kim, J. G., Wang, T., Oh, E., Chen, L., Park, C.H., Son S., Kim S., Mudgett M.B. and Wang, Z. Y. (2014). The bHLHtranscription factor HBI1 mediates the trade-off between growth and pathogen-associated molecular pattern-triggered immunity inArabidopsis. The Plant Cell Online, 26(2), 828-841.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Franklin, K. A., Lee, S.H., Patel, D., Kumar, S.V., Spartz, A.K., Gu, C., Ye, S., Yu, P., Breen, G., Cohen, J.D., Wigge, P.A., and Gray,W.M. (2011). Phytochrome-interacting factor 4 (PIF4) regulates auxin biosynthesis at high temperature. Proc. Natl. Acad. Sci. U. S.A. 108: 20231-5.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Franklin, K. A. (2008). Shade avoidance. New Phytol. 179: 930-944.Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Gendreau, E., Traas, J., Desnos, T., Grandjean, O., Caboche, M., and Höfte, H. (1997). Cellular basis of hypocotyl growth inArabidopsis thaliana. Plant Physiol. 114: 295-305.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Hattori, Y., Nagai, K., Furukawa, S., Song, X., Kawano, R., Sakakibara, H., Wu, J., Matsumoto, T., Yoshimura, A., Kitano, H.,Matsuoka, M., Mori, H., Ashikari, M. (2009). The ethylene response factors SNORKEL1 and SNORKEL2 allow rice to adapt to deepwater. Nature 460: 1026-1030.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Hayashi, K., Tan, X., Zheng, N., Hatate, T., Kimura, Y., Kepinski, S., and Nozaki, H. (2008). Small-molecule agonists and antagonistsof F-box protein-substrate interactions in auxin perception and signaling. Proc. Natl. Acad. Sci. U. S. A. 105: 5632-5637.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Henriques, R., Bögre, L., Horváth, B., and Magyar, Z. (2014). Balancing act: Matching growth with environment by the TORsignalling pathway. J. Exp. Bot. 65: 2691-2701.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Hisamatsu, T., King, R.W., Helliwell, C. a, and Koshioka, M. (2005). The involvement of Gibberellin 20-Oxidase genes in www.plantphysiol.org on July 4, 2016 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2016 American Society of Plant Biologists. All rights reserved.

phytochrome-regulated petiole elongation of Arabidopsis. Plant Physiol. 138: 1106-1116.Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Hong, S.Y., Kim, O.K., Kim, S.G., Yang, M.S., and Park, C.M. (2011). Nuclear import and DNA binding of the ZHD5 transcriptionfactor is modulated by a competitive peptide inhibitor in Arabidopsis. J. Biol. Chem. 286: 1659-1668.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Hu, W. and Ma, H. (2006). Characterization of a novel putative zinc finger gene MIF1: Involvement in multiple hormonal regulationof Arabidopsis development. Plant J. 45: 399-422.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Hyun, Y. and Lee, I. (2006). KIDARI, encoding a non-DNA Binding bHLH protein, represses light signal transduction in Arabidopsisthaliana. Plant Mol. Biol. 61: 283-96.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Iqbal, N., Khan, N. A., Nazar, R., and da Silva, J. A. T. (2012). Ethylene-stimulated photosynthesis results from increased nitrogenand sulfur assimilation in mustard types that differ in photosynthetic capacity. Environ. Exp. Bot. 78: 84-90.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Kazan, K. and Manners, J.M. (2013). MYC2: The master in action. Mol. Plant 6: 686-703.Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Keuskamp, D.H., Pollmann, S., Voesenek, L.A.C.J., Peeters, A.J.M., and Pierik, R. (2010). Auxin transport through PIN-FORMED 3(PIN3) controls shade avoidance and fitness during competition. Proc. Natl. Acad. Sci. U. S. A. 107: 22740-22744.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Keuskamp, D.H., Sasidharan, R., Vos, I., Peeters, A.J.M., Voesenek, L.A.C.J., and Pierik, R. (2011). Blue-light-mediated shadeavoidance requires combined auxin and brassinosteroid action in Arabidopsis seedlings. Plant J. 67: 208-217.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Khanna, R., Shen, Y., Marion, C.M., Tsuchisaka, A., Theologis, A., Schafer, E., and Quail, P.H. (2007). The basic helix-loop-helixtranscription factor PIF5 acts on ethylene biosynthesis and phytochrome signaling by distinct mechanisms. Plant Cell. 19: 3915-3929.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Kim, B.C., Soh, M.S., Hong, S.H., Furuya, M., and Nam, H.G. (1998). Photomorphogenic development of the Arabidopsis shy2-1Dmutation and its interaction with phytochromes in darkness. Plant J. 15: 61-68.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Kunihiro, A., Yamashino, T., Nakamichi, N., Niwa, Y., Nakanishi, H., and Mizuno, T. (2011). PHYTOCHROME-INTERACTING FACTOR4 and 5 (PIF4 and PIF5) activate the Homeobox ATHB2 and auxin-inducible IAA29 genes in the coincidence mechanism underlyingphotoperiodic control of plant growth of Arabidopsis thaliana. The Plant Cell Physiol. 52: 1315-1329.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Lee, Y.K., Kim, G., Kim, I., Park, J., Kwak, S., Choi, G., and Chung, W. (2006). LONGIFOLIA1 and LONGIFOLIA2, two homologousgenes, regulate longitudinal cell elongation in Arabidopsis. Development 133: 4305-4314.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Leivar, P. and Monte, E. (2014). PIFs: systems integrators in plant development. The Plant Cell 26: 56-78.Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Li, L. et al. (2012). Linking photoreceptor excitation to changes in plant architecture. Genes Dev. 26: 785-790.Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title www.plantphysiol.org on July 4, 2016 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2016 American Society of Plant Biologists. All rights reserved.

Lilley, J.L.S., Gee, C.W., Sairanen, I., Ljung, K., and Nemhauser, J.L. (2012). An Endogenous Carbon-Sensing Pathway TriggersIncreased Auxin Flux and Hypocotyl Elongation. Plant Physiol. 160: 2261-2270.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Mickelbart, M. V., Hasegawa, P.M., and Bailey-Serres, J. (2015). Genetic mechanisms of abiotic stress tolerance that translate tocrop yield stability. Nat. Rev. Genet. 16: 237-251.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Morelli, G., Ruberti, I., Molecolare, B., Sapienza, L., and Moro, P.A. (2000). Update on Light Signaling Shade Avoidance Responses.Driving Auxin along Lateral Routes. Plant Physiol. 122: 621-626.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Neff, M.M., Nguyen, S.M., Malancharuvil, E.J., Fujioka, S., Noguchi, T., Seto, H., Tsubuki, M., Honda, T., Takatsuto, S., Yoshida, S.,and Chory, J. (1999). BAS1: A gene regulating brassinosteroid levels and light responsiveness in Arabidopsis. Proc. Natl. Acad.Sci. 96: 15316-15323.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Nemhauser, J.L., Mockler, T.C., and Chory, J. (2004). Interdependency of brassinosteroid and auxin signaling in Arabidopsis. PLoSBiol. 2: E258.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Nishimura, T., Hayashi, K.I., Suzuki, H., Gyohda, A., Takaoka, C., Sakaguchi, Y., Matsumoto, S., Kasahara, H., Sakai, T., Kato, J.I.,Kamiya, Y., and Koshiba, T. (2014). Yucasin is a potent inhibitor of YUCCA, a key enzyme in auxin biosynthesis. Plant J. 77: 352-366.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Nomura, T., Kushiro, T., Yokota, T., Kamiya, Y., Bishop, G.J., and Yamaguchi, S. (2005). The last reaction producing brassinolide iscatalyzed by cytochrome P-450s, CYP85A3 in tomato and CYP85A2 in Arabidopsis. J. Biol. Chem. 280: 17873-17879.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Nozue, K., Tat, A. V., Kumar Devisetty, U., Robinson, M., Mumbach, M.R., Ichihashi, Y., Lekkala, S., and Maloof, J.N. (2015). ShadeAvoidance Components and Pathways in Adult Plants Revealed by Phenotypic Profiling. PLOS Genet. 11: e1004953.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Osakabe, Y., Osakabe, K., Shinozaki, K., and Tran, L. S. P. (2014). Response of plants to water stress. Front. Plant Sci. 5(86): 10-3389.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Park, E., Park, J., Kim, J., Nagatani, A., Lagarias, J.C., and Choi, G. (2012). Phytochrome B inhibits binding of phytochrome-interacting factors to their target promoters. Plant J. 72: 537-546.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Petrásek, J., Cerná, A., Schwarzerová, K., Elckner, M., Morris, D. A., and Zazímalová, E. (2003). Do phytotropins inhibit auxin effluxby impairing vesicle traffic? Plant Physiol. 131: 254-263.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Pfeiffer, A., Shi, H., Tepperman, J.M., Zhang, Y., and Quail, P.H. (2014). Combinatorial Complexity in a Transcriptionally CenteredSignaling Hub in Arabidopsis. Mol. Plant 7: 1598-1618.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Philippar, K., Ivashikina, N., Ache, P., Christian, M., Lüthen, H., Palme, K., and Hedrich, R. (2004). Auxin activates KAT1 and KAT2,two K+-channel genes expressed in seedlings of Arabidopsis thaliana. Plant J. 37: 815-827.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
www.plantphysiol.org on July 4, 2016 - Published by www.plantphysiol.orgDownloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

Pierik, R., Cuppens, M.L., Voesenek, L.A.C.J. and Visser, E.J. (2004) Interactions between ethylene and gibberellins inphytochrome-mediated shade avoidance responses in tobacco. Plant Physiol. 136: 2928-2936.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Pierik, R., Djakovic-Petrovic, T., Keuskamp, D.H., de Wit, M., and Voesenek, L.A.C.J. (2009). Auxin and ethylene regulate elongationresponses to neighbor proximity signals independent of gibberellin and DELLA proteins in Arabidopsis. Plant Physiol. 149: 1701-1712.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Pierik, R., Millenaar, F.F., Peeters, A. J.M., and Voesenek, L.A.C.J. (2005). New perspectives in flooding research: The use of shadeavoidance and Arabidopsis thaliana. Ann. Bot. 96: 533-540.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Pierik, R. and Testerink, C. (2014). The art of being flexible: how to escape from shade, salt and drought. Plant Physiol. 166: 5-22.Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Pierik, R. and De Wit, M. (2014). Shade avoidance: Phytochrome signalling and other aboveground neighbour detection cues. J.Exp. Bot. 65: 2815-2824.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Procko, C., Crenshaw, C.M., Ljung, K., Noel, J.P., and Chory, J. (2014). Cotyledon-Generated Auxin Is Required for Shade-InducedHypocotyl Growth in Brassica rapa. Plant Physiol. 165: 1285-1301.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Quint, M., Delker, C., Franklin, K. A., Wigge, P. A., Halliday, K. J., and van Zanten, M. (2016). Molecular and genetic control of plantthermomorphogenesis. Nature Plants. 2: 15190.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Rademacher, W. (2000). GROWTH RETARDANTS: Effects on Gibberellin Biosynthesis and Other Metabolic Pathways. Annu. Rev.Plant Physiol. Plant Mol. Biol. 51: 501-531.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Rieu, I., Ruiz-Rivero, O., Fernandez-Garcia, N., Griffiths, J., Powers, S.J., Gong, F., Linhartova, T., Eriksson, S., Nilsson, O.,Thomas, S.G., Phillips, A.L., and Hedden, P. (2008). The gibberellin biosynthetic genes AtGA20ox1 and AtGA20ox2 act, partiallyredundantly, to promote growth and development throughout the Arabidopsis life cycle. Plant J. 53: 488-504.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Ruegger, M., Dewey, E., Gray, W.M., Hobbie, L., Turner, J., and Estelle, M. (1998). The TIR1 protein of Arabidopsis functions inauxin response and is related to human SKP2 and yeast Grr1p. Genes Dev. 12: 198-207.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Sasidharan, R., Chinnappa, C.C., Staal, M., Elzenga, J.T.M., Yokoyama, R., Nishitani, K., Voesenek, L.A.C.J., and Pierik, R. (2010).Light quality-mediated petiole elongation in Arabidopsis during shade avoidance involves cell wall modification by xyloglucanendotransglucosylase/hydrolases. Plant Physiol. 154: 978-990.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Sasidharan, R. and Voesenek, L.A.C.J. (2015). Ethylene-mediated acclimations to flooding stress. Plant Physiol.: pp.00387.Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Smalle, J., Haegman, M., Kurepa, J., Van Montagu M, and Straeten, D. V (1997). Ethylene can stimulate Arabidopsis hypocotylelongation in the light. Proc. Natl. Acad. Sci. U. S. A. 94: 2756-2761.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Spartz, A.K., Lee, S.H., Wenger, J.P., Gonzalez, N., Itoh, H., Inzé, D., Peer, W.A., Murphy, A.S., Overvoorde, P.J., and Gray, W.M. www.plantphysiol.org on July 4, 2016 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2016 American Society of Plant Biologists. All rights reserved.

(2012). The SAUR19 subfamily of SMALL AUXIN UP RNA genes promote cell expansion. Plant J. 70: 978-990.Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Stepanova, A.N., Robertson-Hoyt, J., Yun, J., Benavente, L.M., Xie, D.Y., Doležal, K., Schlereth, A., Jürgens, G., and Alonso, J.M.(2008). TAA1-Mediated Auxin Biosynthesis Is Essential for Hormone Crosstalk and Plant Development. Cell 133: 177-191.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Sulpice, R., Flis, A., Ivakov, A. A., Apelt, F., Krohn, N., Encke, B., Abel, C., Feil, R., Lunn, J.E., and Stitt, M. (2014). Arabidopsiscoordinates the diurnal regulation of carbon allocation and growth across a wide range of Photoperiods. Mol. Plant 7: 137-155.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Sun, J., Qi, L., Li, Y., Chu, J., and Li, C. (2012). PIF4-mediated activation of YUCCA8 expression integrates temperature into theauxin pathway in regulating Arabidopsis hypocotyl growth. PLoS Genet. 8: e1002594.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Tabas-Madrid, D., Nogales-Cadenas, R., and Pascual-Montano, A. (2012). GeneCodis 3: a non-redundant and modular enrichmentanalysis tool for functional genomics. Nucleic Acids Res. 40: W478-W483.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Talon, M., Koornneef, M., and Zeevaart, J. A. D. (1990). Accumulation of C19-gibberellins in the gibberellin-insensitive dwarf mutantgai of Arabidopsis thaliana (L.) Heynh. Planta 182: 501-505.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Tanaka, S., Nakamura, S., Mochizuki, N., and Nagatani, A. (2002). Phytochrome in cotyledons regulates the expression of genes inthe hypocotyl through auxin-dependent and -independent pathways. The Plant Cell Physiol. 43: 1171-1181.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Tao, Y., Ferrer, J.L., Ljung, K., Pojer, F., Hong, F., Long, J.A., Li, L., Moreno, J.E., Bowman, M.E., Ivans, L.J., Cheng, Y., Lim, J., Zhao,Y., Ballaré, C.L., Sandberg, G., Noel, J.P. and Chory, J. (2008). Rapid Synthesis of Auxin via a New Tryptophan-Dependent PathwayIs Required for Shade Avoidance in Plants. Cell 133: 164-176.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Tholen, D., Pons, T.L., Voesenek, L.A.C.J., and Poorter, H. (2007). Ethylene insensitivity results in down-regulation of rubiscoexpression and photosynthetic capacity in tobacco. Plant Physiol. 144: 1305-1315.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Turk, E.M. et al. (2005). BAS1 and SOB7 act redundantly to modulate Arabidopsis photomorphogenesis via unique brassinosteroidinactivation mechanisms. Plant J. 42: 23-34.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Unterholzner, S.J., Rozhon, W., Papacek, M., Ciomas, J., Lange, T., Kugler, K.G., Mayer, K.F., Sieberer, T., and Poppenberger, B.(2015). Brassinosteroids Are Master Regulators of Gibberellin Biosynthesis in Arabidopsis. The Plant Cell 27: 2261-2272.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Van Esse, G.W., van Mourik, S., Stigter, H., ten Hove, C. a., Molenaar, J., and de Vries, S.C. (2012). A Mathematical Model forBRASSINOSTEROID INSENSITIVE1-Mediated Signaling in Root Growth and Hypocotyl Elongation. Plant Physiol. 160: 523-532.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Vandenbussche, F., Pierik, R., Millenaar, F.F., Voesenek, L.A.C.J., and Van Der Straeten, D. (2005). Reaching out of the shade. Curr.Opin. Plant Biol. 8: 462-468.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
van Veen, H., Mustroph, A., Barding, G.A., Vergeer-van Eijk, M., Welschen-Evertman, R.A.M., Pedersen, O., Visser, E.J.W., Larive,C.K., Pierik, R., Bailey-Serres, J., Voesenek, L.A.C.J., and Sasidharan, R. (2013). Two Rumex species from contrasting hydrological
www.plantphysiol.org on July 4, 2016 - Published by www.plantphysiol.orgDownloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

niches regulate flooding tolerance through distinct mechanisms. The Plant Cell 25: 4691-707.Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Voesenek L.A.C.J.; and Bailey-serres, J. (2015). Flood adaptive traits and processes?: an overview. New Phytol. 206: 57-73.Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Volodarsky, D., Leviatan, N., Otcheretianski, A., and Fluhr, R. (2009). HORMONOMETER: a tool for discerning transcript signaturesof hormone action in the Arabidopsis transcriptome. Plant Physiol. 150: 1796-1805.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Warnasooriya, S.N. and Montgomery, B.L. (2009). Detection of spatial-specific phytochrome responses using targeted expressionof biliverdin reductase in Arabidopsis. Plant Physiol. 149: 424-33.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Wilson, R.N., Heckman, J.W., and Somerville, C.R. (1992). Gibberellin Is Required for Flowering in Arabidopsis thaliana under ShortDays. Plant Physiol. 100: 403-408.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Zhiponova, M.K., Morohashi, K., Vanhoutte, I., Machemer-Noonan, K., Revalska, M., Van Montagu, M., Grotewold, E., andRussinova, E. (2014). Helix-loop-helix/basic helix-loop-helix transcription factor network represses cell elongation in Arabidopsisthrough an apparent incoherent feed-forward loop. Proc. Natl. Acad. Sci. U. S. A. 111: 2824-9.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Zheng, Z., Guo, Y., Novak, O., Chen, W., Ljung, K., Noel, J.P., and Chory, J. (2016). Local auxin metabolism regulates environment-induced hypocotyl elongation. Nat. Plants. 21: 16025. doi: 10.1038/nplants.2016.25
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Zhong, S., Shi, H., Xue, C., Wang, L., Xi, Y., Li, J., Quail, P.H., Deng, X.W., and Guo, H. (2012). A molecular framework of light-controlled phytohormone action in Arabidopsis. Curr. Biol. 22: 1530-1535.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
www.plantphysiol.org on July 4, 2016 - Published by www.plantphysiol.orgDownloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.




![Transcriptome and metabolome profiling provide insights into … · 2021. 3. 1. · elongation by restricting signals that control stem elong-ation [13]. The establishment of vascular](https://static.fdocuments.in/doc/165x107/61206886edb8ca0e3f524b6f/transcriptome-and-metabolome-profiling-provide-insights-into-2021-3-1-elongation.jpg)













