
Ethanol-Induced Leakagein Saccharomyces cerevisiae ...aem.asm.org/content/54/4/903.full.pdf ·...
7
Vol. 54, No. 4 APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Apr. 1988, p. 903-909 0099-2240/88/040903-07$02.00/0 Ethanol-Induced Leakage in Saccharomyces cerevisiae: Kinetics and Relationship to Yeast Ethanol Tolerance and Alcohol Fermentation Productivity SANCHA P. SALGUEIRO, ISABEL SA CORREIA,* AND JULIO M. NOVAIS Laboratorio de Engenharia Bioquimica, Instituto Superior Tecnico, 1000 Lisbon, Portugal Received 2 November 1987/Accepted 5 January 1988 Ethanol stimulated the leakage of amino acids and 260-nm-light-absorbing compounds from cells of Saccharomyces cerevisiae. The efflux followed first-order kinetics over an initial period. In the presence of lethal concentrations of ethanol, the efflux rates at 30 and 36°C were an exponential function of ethanol concentration: kx = k= meE(X-XAJ, where k,j and kxm are the efflux rate constants, respectively, in the presence of a concentration X of ethanol or the minimal concentration of ethanol, Xm, above which the equation was applicable, coincident with the minimal lethal concentration of ethanol. E is the enhancement constant. At 36°C, as compared with the corresponding values at 30°C, the efflux rates were higher and the minimal concentration of ethanol (Xm) was lower. The exponential constants for the enhancement of the rate of leakage (E) had similar values at 30 or 36 °C and were of the same order of magnitude as the corresponding exponential constants for ethanol-induced death. Under isothermic conditions (30°C) and up to 22% (vol/vol) ethanol, the resistance to ethanol-induced leakage of 260-nm-light-absorbing compounds was found to be closely related with the ethanol tolerance of three strains of yeasts, Kluyveromyces marxianus, Saccharomyces cerevisiae, and Saccharomyces bayanus. The resistance to ethanol-induced leakage indicates the possible adoption of the present method for the rapid screening of ethanol-tolerant strains. The addition to a fermentation medium of the intracellular material obtained by ethanol permeabilization of yeast cells led to improvements in alcohol fermentation by S. cerevisiae and S. bayanus. The action of the intracellular material, by improving yeast ethanol tolerance, and the advantages of partially recycling the fermented medium after distillation were discussed. Several ethanol-sensitive rate processes have been identi- fied in yeasts (34). These processes include the enhancement of death at low, intermediate, and high temperatures (30, 34), inhibition of growth (4, 8, 10, 34), and inhibition of several systems for nutrient transport (glucose [12], fructose [28], maltose [19], ammonium [13], and amino acids [14]). The toxicity of ethanol and other drugs in all these rate-sensitive processes was correlated with their lipid-buffer partition coefficients, suggesting their interference with the hydropho- bic regions of membranes (8, 34). The plasma membrane is the possible target for ethanol inhibition of nutrient trans- port, but the inner mitochondrial membrane was determined to be the target region of ethanol-enhanced thermal death (3, 31). Ethanol interacts with membranes possibly by insertion into the hydrophobic interior, increasing the polarity of this region, weakening the hydrophobic barrier to the free ex- change of polar molecules, and weakening the hydrophobic interactions and affecting the positioning of proteins within the membranes (8). The loss of membrane integrity de- creases the ability of the cell to maintain a concentration gradient across the plasma membrane, and several systems involved in solute transport should be affected. Ethanol and other chaotropic factors such as temperature interfere with membrane organization by increasing its fluidity and perme- ability to ions and small metabolites (6, 8). During alcohol fermentation, Saccharomyces carlsber- gensis was found to release nitrogen-containing compounds into the medium (18). These consist mainly of amino acids and related substances and UV-absorbing compounds (sev- * Corresponding author. eral nucleotides, free bases, and nucleosides [16, 17]) and are released by membrane permeabilization during fermenta- tion. They appear to originate in a free intracellular pool (16-18). As, by the end of the fermentation, the leaked material was not reabsorbed by the yeast cells and persisted in the finished product, it was suggested that leakage affects the quality of beer and wine. Ethanol was reported to promote the efflux of Mg2" and 260-nm-light-absorbing com- pounds and, for high ethanol concentrations, of proteins in Zymomonas mobilis (24). Another example of ethanol-in- duced permeabilization of the plasma membrane is the increase of the passive influx of protons in Saccharomyces cerevisiae (9, 15). Ethanol-induced leakage of ions would be expected to interfere with nutrient accumulation because their active transport into the cell is coupled to ion fluxes and based on transmembrane gradients (5). When cellular suspensions of Staphylococcus aureus or Aerobacter aerogenes were heated, leakage of Mg2+, amino acids, 260-nm-light-absorbing compounds, and proteins also occurred (1, 2, 32). The rate and the nature of the material leaked varied with temperature, in part owing to different amounts of protein coagulation and RNA degradation which increased the content of RNA-like material in the metabolic pool and outside the cell (1, 2, 32). No significant degrada- tion of DNA appeared to occur in the temperature-stressed cells (1, 23, 32). In the present work, we studied the kinetics of ethanol- induced leakage of amino acids, 260-nm-light-absorbing compounds, and ribose-containing compounds in S. cerevi- siae at 30 and 36°C. We also correlated the resistance of the plasma membrane to ethanol permeabilization (given by the rate of the efflux of 260-nm-light-absorbing compounds) in S. 903 on August 29, 2018 by guest http://aem.asm.org/ Downloaded from
Transcript of Ethanol-Induced Leakagein Saccharomyces cerevisiae ...aem.asm.org/content/54/4/903.full.pdf ·...

Vol. 54, No. 4APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Apr. 1988, p. 903-9090099-2240/88/040903-07$02.00/0
Ethanol-Induced Leakage in Saccharomyces cerevisiae:Kinetics and Relationship to Yeast Ethanol Tolerance
and Alcohol Fermentation ProductivitySANCHA P. SALGUEIRO, ISABEL SA CORREIA,* AND JULIO M. NOVAIS
Laboratorio de Engenharia Bioquimica, Instituto Superior Tecnico, 1000 Lisbon, Portugal
Received 2 November 1987/Accepted 5 January 1988
Ethanol stimulated the leakage of amino acids and 260-nm-light-absorbing compounds from cells ofSaccharomyces cerevisiae. The efflux followed first-order kinetics over an initial period. In the presence of lethalconcentrations of ethanol, the efflux rates at 30 and 36°C were an exponential function of ethanol concentration:kx =k= meE(X-XAJ, where k,j and kxm are the efflux rate constants, respectively, in the presence of a
concentration X of ethanol or the minimal concentration of ethanol, Xm, above which the equation was
applicable, coincident with the minimal lethal concentration of ethanol. E is the enhancement constant. At36°C, as compared with the corresponding values at 30°C, the efflux rates were higher and the minimalconcentration of ethanol (Xm) was lower. The exponential constants for the enhancement of the rate of leakage(E) had similar values at 30 or 36 °C and were of the same order of magnitude as the corresponding exponentialconstants for ethanol-induced death. Under isothermic conditions (30°C) and up to 22% (vol/vol) ethanol, theresistance to ethanol-induced leakage of 260-nm-light-absorbing compounds was found to be closely relatedwith the ethanol tolerance of three strains of yeasts, Kluyveromyces marxianus, Saccharomyces cerevisiae, andSaccharomyces bayanus. The resistance to ethanol-induced leakage indicates the possible adoption of thepresent method for the rapid screening of ethanol-tolerant strains. The addition to a fermentation medium ofthe intracellular material obtained by ethanol permeabilization of yeast cells led to improvements in alcoholfermentation by S. cerevisiae and S. bayanus. The action of the intracellular material, by improving yeastethanol tolerance, and the advantages of partially recycling the fermented medium after distillation werediscussed.
Several ethanol-sensitive rate processes have been identi-fied in yeasts (34). These processes include the enhancementof death at low, intermediate, and high temperatures (30, 34),inhibition of growth (4, 8, 10, 34), and inhibition of severalsystems for nutrient transport (glucose [12], fructose [28],maltose [19], ammonium [13], and amino acids [14]). Thetoxicity of ethanol and other drugs in all these rate-sensitiveprocesses was correlated with their lipid-buffer partitioncoefficients, suggesting their interference with the hydropho-bic regions of membranes (8, 34). The plasma membrane isthe possible target for ethanol inhibition of nutrient trans-port, but the inner mitochondrial membrane was determinedto be the target region of ethanol-enhanced thermal death (3,31). Ethanol interacts with membranes possibly by insertioninto the hydrophobic interior, increasing the polarity of thisregion, weakening the hydrophobic barrier to the free ex-change of polar molecules, and weakening the hydrophobicinteractions and affecting the positioning of proteins withinthe membranes (8). The loss of membrane integrity de-creases the ability of the cell to maintain a concentrationgradient across the plasma membrane, and several systemsinvolved in solute transport should be affected. Ethanol andother chaotropic factors such as temperature interfere withmembrane organization by increasing its fluidity and perme-ability to ions and small metabolites (6, 8).
During alcohol fermentation, Saccharomyces carlsber-gensis was found to release nitrogen-containing compoundsinto the medium (18). These consist mainly of amino acidsand related substances and UV-absorbing compounds (sev-
* Corresponding author.
eral nucleotides, free bases, and nucleosides [16, 17]) and arereleased by membrane permeabilization during fermenta-tion. They appear to originate in a free intracellular pool(16-18). As, by the end of the fermentation, the leakedmaterial was not reabsorbed by the yeast cells and persistedin the finished product, it was suggested that leakage affectsthe quality of beer and wine. Ethanol was reported topromote the efflux of Mg2" and 260-nm-light-absorbing com-pounds and, for high ethanol concentrations, of proteins inZymomonas mobilis (24). Another example of ethanol-in-duced permeabilization of the plasma membrane is theincrease of the passive influx of protons in Saccharomycescerevisiae (9, 15). Ethanol-induced leakage of ions would beexpected to interfere with nutrient accumulation becausetheir active transport into the cell is coupled to ion fluxes andbased on transmembrane gradients (5).When cellular suspensions of Staphylococcus aureus or
Aerobacter aerogenes were heated, leakage of Mg2+, aminoacids, 260-nm-light-absorbing compounds, and proteins alsooccurred (1, 2, 32). The rate and the nature of the materialleaked varied with temperature, in part owing to differentamounts of protein coagulation and RNA degradation whichincreased the content of RNA-like material in the metabolicpool and outside the cell (1, 2, 32). No significant degrada-tion of DNA appeared to occur in the temperature-stressedcells (1, 23, 32).
In the present work, we studied the kinetics of ethanol-induced leakage of amino acids, 260-nm-light-absorbingcompounds, and ribose-containing compounds in S. cerevi-siae at 30 and 36°C. We also correlated the resistance of theplasma membrane to ethanol permeabilization (given by therate of the efflux of 260-nm-light-absorbing compounds) in S.
903
on August 29, 2018 by guest
http://aem.asm
.org/D
ownloaded from

904 SALGUEIRO ET AL.
cerevisiae and Kluyveromyces marxianus (with differentethanol tolerances as previously characterized by conven-tional methods [25, 27-29]) and Saccharomyces bayanus.Improvements in alcoholic fermentation by supplementationof media with amino acids, nucleotides, proteins, vitamins,and metal ions (4, 10) and the protection given to S. bayanusand K. marxianus toward lethal concentrations of ethanol bythe compounds leaked from yeast cells subjected to ethanolpermeabilization (22) also justified our interest in the presentstudy.
MATERIALS AND METHODS
Microorganisms. The strains used in this study were S.cerevisiae IGC 3507 III, the main strain used by van Udenand co-workers in studies on ethanol toxicity (31); K.marxianus IGC 2671, previously Kluyveromyces fragilis,isolated by van Uden et al. (33), subjected to studies onethanol toxicity and tolerance and used in Jerusalem arti-choke juice fermentation (25, 27-29); and S. bayanus, anindustrial strain used in the production of sparkling wines(35, 36).Growth conditions. Yeast cells used in ethanol permeabi-
lization experiments were pregrown for 16 h in an orbitalshaker (200 rpm) at 30°C in 500-ml conical flasks closed withcotton plugs and containing 200 ml of a medium with (perliter) 5 g of yeast extract (Difco Laboratories, Detroit,Mich.), 5 g of KH2PO4, 5 g of (NH4)2S04, 1 g of MgSO4, and100 g of glucose (initial optical density at 640 nm, 0.025, in aPYE UNICAM SP6-550 spectrophotometer). These cellswere used to inoculate a similar liquid medium with 20 g ofglucose per liter (the initial optical density at 640 nm was 0.7)and grown under the same conditions up to an opticaldensity of 3 (exponential phase).
Ethanol permeabilization and sampling. Cells of S. cerevi-siae IGC 3507 III grown as described above were collectedby centrifugation, washed twice with sterile phosphatebuffer (50 mM, pH 5.0), and suspended in sterile buffer plusdifferent ethanol concentrations contained in conical flasksclosed with a rubber bung. The optical density at 640 nm ofthese cellular suspensions was 3 (approximately 5 x 107 cellsper ml, corresponding to 0.94 mg [dry weight]/ml). Thecultures were incubated in a thermostated shaking waterbath at 30 and 36°C. Samples (5 ml) were centrifuged for 3min, and the supernatants (A) were removed and immedi-ately frozen for further analysis of UV-absorbing com-pounds, amino acids, and ribose-containing compounds. Thepelleted cells in the centrifuge tubes were suspended in 5 mlof 0.25 M perchloric acid at 4°C and maintained at thistemperature for 30 min. They were later centrifuged for 3min, and the supernatants (B) (containing the cold-acid-soluble compounds) were removed and immediately frozenfor further analysis of UV-absorbing compounds, aminoacids, and ribose-containing compounds as described below.After cold-acid extraction, the pelleted cells were suspendedin 5 ml of 0.3 M potassium hydroxide, placed in a water bathat 37°C, and maintained at this temperature for 1 h for RNAhydrolysis (7). After this treatment, 5 ml of 1 M perchloricacid at 4°C was added to precipitate the DNA and proteins.The supernatants (C) containing the result of cellular RNAhydrolysis were removed after centrifugation and immedi-ately frozen for further RNA analysis by measuring theirribose content (7).
Analysis. Supernatants A and B were examined for 260-nm-light-absorbing compounds in a PYE UNICAM SP6-550spectrophotometer. Amino acids in supernatants A and B
were determined by the ninhydrin method (21), using glycineas the standard. The supernatant (B) containing the cold-acid-soluble compounds was neutralized with sodium hy-droxide before amino acid analysis. The ribose content ofthe supernatants (A, B, and C) was determined by theorcinol method (7), using ribose as the standard.Comparison of yeast ethanol tolerance by leakage of 260-
nm-light absorbing compounds. For comparison of yeastethanol tolerance, cells of S. cerevisiae IGC 3507 III, K.marxianus IGC 2671, and S. bayanus were harvested at theexponential phase. The cells were pregrown under standardconditions as described above for permeabilization experi-ments and subjected to ethanol-induced permeabilization at30°C, and the supernatants (A) from the centrifuged sampleswere immediately examined for 260-nm-light-absorbing com-pounds.
Alcohol fermentations. Batch fermentations were done inan orbital shaker at 30°C by S. cerevisiae and S. bayanus inbase medium containing yeast extract (Difco) (1 g/liter),MgSO4 (1 g/liter), (NH4)2SO4 (5 g/liter), KH2PO4 (5 g/liter),and glucose (350 g/liter), and the fermentation profiles werecompared with those of fermentations carried out in (i)similar medium including the material obtained after ethanol-induced permeabilization of yeast cells incubated with 18%(vol/vol) ethanol for 4 and 18 h, or (ii) similar mediumincluding the cell debris obtained in the same permeabiliza-tion experiments.These fermentation supplements were obtained by etha-
nol-induced permeabilization at 30°C of yeast suspensionscontaining 5 x 107 cells per ml, corresponding to 0.94 g (dryweight)/liter. After 4 or 18 h of incubation with 18% (vol/vol)ethanol, the cells were centrifuged and the debris waswashed with sterile water before addition to the fermentationmedia. The ethanol in the supernatants was removed byevaporation under vacuum at 40°C, until a final concentra-tion below 0.5% (vol/vol) ethanol was obtained. Duringevaporation, the volume of the supernatants was reduced to50% of the initial volume. The fermentation media weremade up with all the material leaked or with the cell debriscorresponding to the permeabilized yeasts in such a way thatthe concentration of base medium nutrients was maintained.Glucose was analyzed with the dinitrosalicylic acid re-
agent (20), and ethanol was analyzed by gas chromatographyin an HP 5710 A gas chromatograph (equipped with a0.125-in. (0.32-cm) by 2-m column packed with Chromosorb101 and a flame ionization detector). The temperature of theinjector and of the detector was 250°C, and the oven wasoperated isothermically at 160°C. The carrier gas was N2with a flow rate of 60 ml/min. Butanol was used as theinternal standard.
RESULTS AND DISCUSSION
Kinetics of ethanol-induced leakage. When S. cerevisiaecells were suspended in buffer (pH 5.0) at 30 or 36°C, leakageof amino acids and 260-nm-light-absorbing compounds oc-curred, as described for S. carlsbergensis (18). The leakagefollowed first-order kinetics over an initial period (Fig. laand b). When ethanol was added to the buffer, the effective-ness of the plasma membrane in retaining these intracellularcompounds against a concentration gradient decreased. Un-der isothermic conditions, the rate of the efflux of aminoacids and 260-nm-light-absorbing compounds increased asan exponential function of ethanol concentration:
k= IXmeE(X-Xm) (1)
APPL. ENVIRON. MICROBIOL.
on August 29, 2018 by guest
http://aem.asm
.org/D
ownloaded from
ETHANOL-INDUCED LEAKAGE IN SACCHAROMYCES CEREVISIAE
EC
C>%O
c
.0 0.10
.0
E
0
.eKU
0 2 4 6 0 2 4 6Ti me (hou rs)
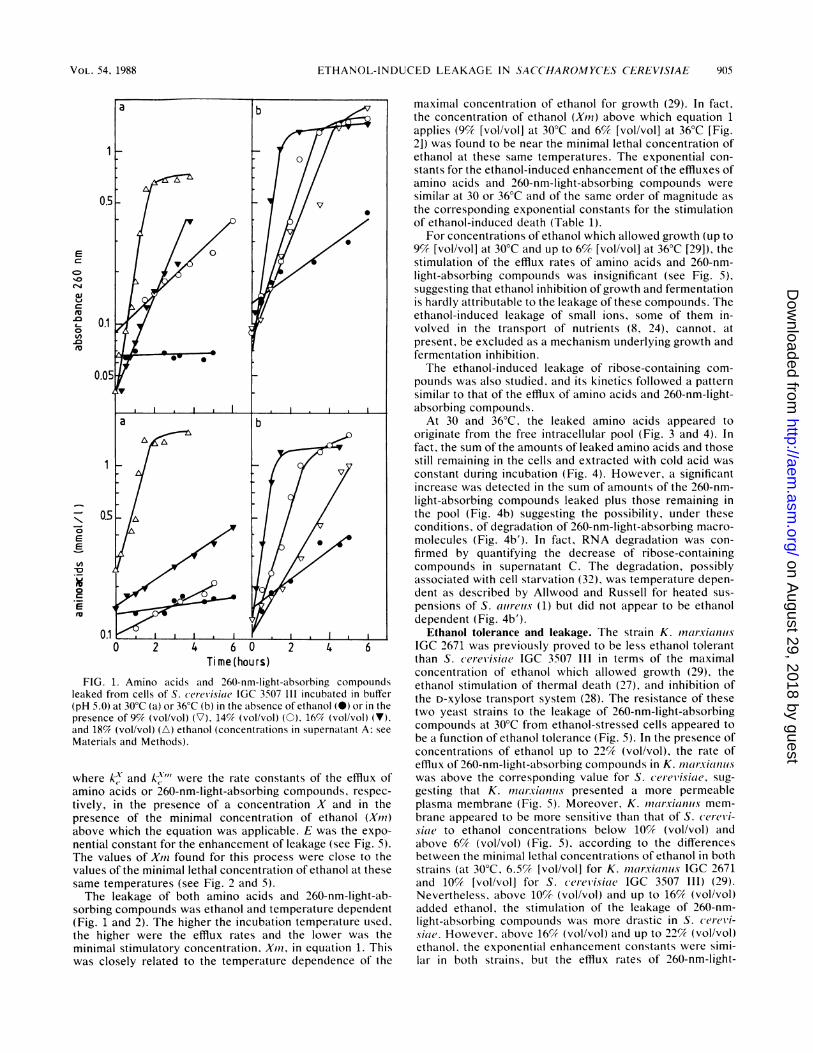
FIG. 1. Amino acids and 260-nm-light-absorbing compoundsleaked from cells of S. (erevisiae IGC 3507 III incubated in buffer(pH 5.0) at 30°C (a) or 36°C (b) in the absence of ethanol (S) or in thepresence of 9% (vol/vol) (V), 14% (vol/vol) (0), 16% (vol/vol) (V),and 18% (vol/vol) (A) ethanol (concentrations in supernatant A: see
Materials and Methods).
where k-x and k-,Y"' were the rate constants of the efflux ofamino acids or 260-nm-light-absorbing compounds, respec-tively, in the presence of a concentration X and in thepresence of the minimal concentration of ethanol (Xiii)above which the equation was applicable. E was the expo-nential constant for the enhancement of leakage (see Fig. 5).The values of Xiii found for this process were close to thevalues of the minimal lethal concentration of ethanol at thesesame temperatures (see Fig. 2 and 5).The leakage of both amino acids and 260-nm-light-ab-
sorbing compounds was ethanol and temperature dependent(Fig. 1 and 2). The higher the incubation temperature used,the higher were the efflux rates and the lower was theminimal stimulatory concentration. Xiii, in equation 1. Thiswas closely related to the temperature dependence of the
maximal concentration of ethanol for growth (29). In fact,the concentration of ethanol (Xmn) above which equation 1applies (9% [vol/vol] at 30°C and 6% [vol/vol] at 36°C [Fig.21) was found to be near the minimal lethal concentration ofethanol at these same temperatures. The exponential con-stants for the ethanol-induced enhancement of the effluxes ofamino acids and 260-nm-light-absorbing compounds weresimilar at 30 or 36°C and of the same order of magnitude asthe corresponding exponential constants for the stimulationof ethanol-induced death (Table 1).
For concentrations of ethanol which allowed growth (up to9% [vol/vol] at 30°C and up to 6% [vol/vol] at 36°C [29]), thestimulation of the efflux rates of amino acids and 260-nm-light-absorbing compounds was insignificant (see Fig. 5),suggesting that ethanol inhibition of growth and fermentationis hardly attributable to the leakage of these compounds. Theethanol-induced leakage of small ions, some of them in-volved in the transport of nutrients (8, 24), cannot, atpresent, be excluded as a mechanism underlying growth andfermentation inhibition.The ethanol-induced leakage of ribose-containing com-
pounds was also studied, and its kinetics followed a patternsimilar to that of the efflux of amino acids and 260-nm-light-absorbing compounds.At 30 and 36°C, the leaked amino acids appeared to
originate from the free intracellular pool (Fig. 3 and 4). Infact, the sum of the amounts of leaked amino acids and thosestill remaining in the cells and extracted with cold acid wasconstant during incubation (Fig. 4). However, a significantincrease was detected in the sum of amounts of the 260-nm-light-absorbing compounds leaked plus those remaining inthe pool (Fig. 4b) suggesting the possibility, under theseconditions, of degradation of 260-nm-light-absorbing macro-molecules (Fig. 4b'). In fact, RNA degradation was con-firmed by quantifying the decrease of ribose-containingcompounds in supernatant C. The degradation, possiblyassociated with cell starvation (32), was temperature depen-dent as described by Allwood and Russell for heated sus-pensions of S. aureus (1) but did not appear to be ethanoldependent (Fig. 4b').
Ethanol tolerance and leakage. The strain K. rnarxiamiisIGC 2671 was previously proved to be less ethanol tolerantthan S. (ceievisiae IGC 3507 III in terms of the maximalconcentration of ethanol which allowed growth (29), theethanol stimulation of thermal death (27), and inhibition ofthe D-xylose transport system (28). The resistance of thesetwo yeast strains to the leakage of 260-nm-light-absorbingcompounds at 30°C from ethanol-stressed cells appeared tobe a function of ethanol tolerance (Fig. 5). In the presence ofconcentrations of ethanol up to 22%c (vol/vol), the rate ofefflux of 260-nm-light-absorbing compounds in K. mzarxlU1ia1nSwas above the corresponding value for S. cerevisiae, sug-gesting that K. nlZarxianl1lIs presented a more permeableplasma membrane (Fig. 5). Moreover, K. inar1vianus mem-brane appeared to be more sensitive than that of S. cereivi-siaie to ethanol concentrations below 10% (vol/vol) andabove 6% (vol/vol) (Fig. 5), according to the differencesbetween the minimal lethal concentrations of ethanol in bothstrains (at 30°C, 6.5% [vol/voll for K. inaia.vius IGC 2671and 10% [vol/voll for S. (erev'isiae IGC 3507 III) (29).Nevertheless, above 10% (vol/vol) and up to 16% (vol/vol)added ethanol, the stimulation of the leakage of 260-nm-light-absorbing compounds was more drastic in S. cerevi-sie. However, above 16% (vol/vol) and up to 22'% (vol/vol)ethanol, the exponential enhancement constants were simi-lar in both strains, but the efflux rates of 260-nm-light-
VOL. 54, 1988 905
on August 29, 2018 by guest
http://aem.asm
.org/D
ownloaded from

906 SALGUEIRO ET AL.
10
5
3
0
0.5~ ~
- .11 .
8 12 16 20 8 12 1 20Ethanol % (v /v )
FIG. 2. Efflux rates of 260-nm-light-absorbing compounds at 30°C(O) or 36°C (O)or ofamino acids at 30°C(A) or 36°C(A) in S.cerevisiaeIGC 3507 III in the presence of increasing concentrations of added ethanol.
absorbing compounds of K. marxianus were always abovethe corresponding values for S. cerevisiae. a)b)
Trying to generalize the correspondence betweenyeasttresistance to the leakage of 260-nm-light-absorbingcom-epounds induced by ethanol and ethanol tolerance, we also 0.6ox.quantified the efflux rates of these compounds from cells of \a strain of S. bayanus used for secondary fermentation in the \ 4sparkling wine industry and known to have a high ethanol °0_\
0.3~~~~~~~~~C
tolerance. The rates of efflux of 260-nm-light-absorbing w 5 \[compounds in S. bayanus were above the equivalents in S. ,cerevisiae for concentrations of added ethanol below 169o D \\(vol/vol) (Fig. 5), indicating a more permeable plasmamem- °O 0.2 _ \ \brane. However, 16%o(vol/vol) ethanol seemed to be, for S. X \ 3bayanus, the minimal stimulatory concentration of ethanol Rin equation 1. -Above 16%o(vol/vol) added ethanol, the exponential en- 0a)E.|f b)
hancement constants of ethanol-induced leakage for thethree -fermenting yeast strains were identical, but the rates ofleakage were higher in K. marxianus and lower in S. 1,5h)Sbayanus, closely related with their tolerance to ethanol- _ Xiinduced death. X \
0~~~~~~~~~
Although it is an important property in alcohol fermenta- E 11tion, at present there is no method generally used or ac- E 11 0 t
TABLE 1. Constants for the exponential enhancement by ethanol m \\at 30 and 36°C of the specific death rates (values calculated from .' 0 0\\\
reference 26) and the rates of the efflux of amino acids and E \\260-nm-light-absorbing compounds in S. cerevisiae IGC 3507 III ')3
Efflux of
Temp Death Amino acid 260-nm-light-(0C) (1/mol) efflux (1/mol) absorbing
compounds(1/mol)
30 2.03 1.74 1.7436 2.48 1.06 1.52
Time (hours)FIG. 3. Amino acids and 260-nm-light-absorbing compounds ex-
tracted with cold acid from cells of S. cerevisiae IGC 3507 IIIincubated in buffer (pH 5.0) at 30°C (a) and 36°C (b) with 16%(vol/vol) (E), 18% (vol/vol) (0), and 20% (vol/vol) (A) ethanol(concentrations in supernatant B; see Materials and Methods).
APPL. ENVIRON. MICROBIOL.
on August 29, 2018 by guest
http://aem.asm
.org/D
ownloaded from

ETHANOL-INDUCED LEAKAGE IN SACCHAROMYCES CEREVISIAE
2
In
10
a)
02o AtA 8o0
I .. . ..
I I * ~~~~~b)
0 0-00 0
5I -I I I I
1.b I . I . I I
0 2 4 6Timo (hotrs)
FIG. 4. Sum of the concentrations of amino acids and 260-nm-
light-absorbing compounds in supernatants A and B (see Materials
and Methods) from cellular suspensions of S. cerevisiae IGC 3507III at 30°C (a and a') and 36°C (b and b') in buffer (pH 5.0) with 16%
(vol/vol) (I), 18% (vol/vol) (0), and 20% (vol/vol) (A) ethanol.
cepted to characterize ethanol tolerance. This is understand-
able because ethanol tolerance deeply depends on mediumcomposition (4; M. F. Rosa,I. Si-Correia, and J. M. Novais,
Biotechnol. Bioeng., in press), temperature (30), and other
environmental conditions. Recently, a method was proposed
5
3
C
0..3/
0 4 8 12 16 20
Ethanol% (v/v)
FIG. 5. Rates of efflux of 260-nm-light-absorbing compoundsfrom cells of S. cerevisiae IGC 3507 III (0), K. marxianus IGC 2671(0), and S. bayanus (A) suspended in buffer (pH 5.0) at 30°C withincreasing concentrations of ethanol.
(9), based on the enhancement by ethanol of passive influx ofprotons through membranes, in which differences in ethanoltolerance were associated with the effect of ethanol on theacidification curves (9). The method proposed here is alsobased on the resistance of the plasma membrane to permea-bilization by ethanol. As compared with the stimulation ofproton influx (9), this method is not sensitive to the stimu-lation induced by concentrations of ethanol below the max-imal for growth, probably because nucleotides, owing totheir electric charge and higher volume, are less susceptibleto leakage than protons.The multiple differences observed among these three yeast
species indicate that further work needs to be done in testinga significant number of fermenting strains to address in moredetail the relationship between the resistance to the ethanol-induced leakage of 260-nm-light-absorbing compounds andethanol tolerance.Improvements in alcohol fermentation by the addition of the
material leaked from ethanol-permeabilized cells. S. cerevi-siae IGC 3507 III and S. bayanus were incubated in a basefermentation medium supplemented or not supplementedwith the material leaked from cells incubated at 30°C inbuffer (pH 5.0) with 18% (vol/vol) ethanol for 4 or 18 h. andtheir alcohol fermentation profiles were compared. Thesupplementation led to an increase of the fermentation ratesof the two yeast strains (Fig. 6), the improvements beingmore significant with the material released after 18 h whichcontained a higher percentage of intracellular compounds(Table 2). S. cerevisiae IGC 3507 III led to higher yields inethanol possibly because it is a respiratory mutant, but S.bayanus produced ethanol with lower yields (Fig. 6). There-fore, the higher sugar consumption rates observed with S.bayanus were understandable and so were the insignificantdifferences observed in the medium supplemented with thematerial released after 4 or 18 h (Fig. 6).Owing to the utilization of a large inoculum, with cells in
the exponential phase, it was possible to attain 15 and 13%(vol/vol) ethanol, respectively, after 100 h of fermentation inthe base medium carried out by S. cerevisiae IGC 3507 III orS. bayanus. Some glucose (29 and 17%, respectively) re-mained unfermented during the process carried out by S.cerevisiae and S. bayanus. Supplementation with the mate-rial released by cells permeabilized with ethanol led to theexhaustion of 350 g of glucose per liter in 96 h and to theproduction of 17% (vol/vol) ethanol by S. cerevisiae IGC3507 III and 14% (vol/vol) ethanol by S. bayanus. Theincorporation of the cell debris which remained after ethanolpermeabilization for 4 or 18 h led to the acceleration ofglucose consumption by both strains but with no counter-balance in terms of the ethanol produced (Fig. 6). Thedecrease in the yield of ethanol production during the first 48h of fermentation can probably be attributed to the high
TABLE 2. Amino acids and 260-nm-light-absorbing compoundsleaked from cellular suspensions of S. cerevisiae IGC 3507 III
and S. bayanus incubated at 30°C in buffer (pH 5.0)with 18% (vol/vol) ethanol for 4 or 18 h"
Time of Amino acids(%) A260 ()incubation
(h) S. cerevisiae S. bayanus S.cerevisiae S. bayanus
4 59 b 72 7518 83 88 83
"Amino acids or 260-nm-light-absorbing compounds released were consid-ered as the percentage of the material released from the same yeast suspen-sions incubated at 100°C for 20 min (16).
b_, No results available.
VOL. 54, 1988 907
ecc:,V%OCI*4
0
29- 10W.0m 0.1
on August 29, 2018 by guest
http://aem.asm
.org/D
ownloaded from

908 SALGUEIRO ET AL.
3ba b0 lo-C
c a) b)
3000
0~~~~0
LV~~~
100~~~~~~~~~~~~~
V 0
0I0 24 48 72 96 120 0 24 48 72 96 120
Time (hours)FIG. 6. Fermentations carried out by S. cerevisiae IGC 3507 III (a) or S. bayanus (b) of 350 g of glucose per liter in a base medium (0),
in medium supplemented with the compounds released from yeast cells incubated with 18% (vol/vol) ethanol for 4 h (A) or 18 h (V) orsupplemented with the cell debris obtained after 4 h (A) or 18 h (V) of ethanol permeabilization.
concentration of the cell debris added, but it is also possiblethat we are dealing with a physical phenomenon of glucoseadsorption by the debris since it was possible to attain anidentical maximal concentration of ethanol (Fig. 6) after 100h of fermentation.The cell debris used here probably corresponded to the
"yeast ghosts" referred to by Lafon-Lafourcade et al. (11)as being involved in the restart of stopped fermentations. Ifthis is so, the role of the yeast ghosts in fermentationimprovement cannot be attributed only to the adsorption oftoxic subproducts of alcohol fermentation as was suggested(11) but mainly to the presence of intracellular compoundsstill present inside the ethanol-permeabilized cells.The improvements in alcohol fermentation discussed so
far might *be attributed to the increase of yeast ethanoltolerance owing to the presence of several intracellularcompounds. This was recently proved (22) for the strain ofS. bayanus used here. The ethanol-induced specific deathrate was higher for sparse yeast suspensions but decreasedto the value calculated for dense suspensions when deathexperiments were undertaken in a medium containing theintracellular compounds released from dense cell suspen-sions during incubation with ethanol (22). Defined additivesreported to be involved in alcohol fermentation improve-ments such as amino acids, nucleotides, vitamins, and metalions (4) are present intracellularly. Therefore, it is possiblethat the material released from yeast cells by ethanol per-
meabilization can protect the remaining living cells fromethanol toxicity, according to several reports on fermenta-tion stimulation by the supplementation of media with com-mercial yeast extract (4). Besides their role in decreasingethanol-induced death, the presence of those protectors ledto the decrease of growth and fermentation inhibition byethanol concentrations near the maximal for growth, butthey probably have no effect in the kinetics of growth andfermentation for ethanol concentrations below the minimallethal concentration as was proved for peptone (C. A. Viegasand I. Sa-Correia, unpublished data) and Ca2" supplemen-tation (R. C. Nabais, I. Sa-Correia, and J. M. Novais,submitted for publication).The results reported here partially explain two current
practices in industrial alcohol fermentation: the possibility ofincreasing the maximal concentration of ethanol by increas-ing the amount of the inoculum and by the partial recycling(of about 30% [vol/vol]) of the fermented medium after yeastremoval and ethanol distillation. The higher initial cellconcentration, besides allowing a faster production of etha-nol during the initial stages of fermentation, might, throughthe compounds released during fermentation by ethanol-induced cell permeabilization, increase the tolerance of thecells still metabolically active to higher concentrations ofethanol. These yeasts, less ethanol inhibited, could continuea more rapid fermentation up to a higher concentration ofethanol.
APPL. ENVIRON. MICROBIOL.
on August 29, 2018 by guest
http://aem.asm
.org/D
ownloaded from

ETHANOL-INDUCED LEAKAGE IN SACCHAROMYCES CEREVISIAE
The partial recycling of the fermented medium after cellremoval and ethanol distillation, a medium containing thecompounds released from the cells during the fermentation,may have a positive effect that surpasses the toxicity of lessvolatile subproducts of alcohol fermentation such as octa-noic and decanoic acids (26, 36).
ACKNOWLEDGMENTS
We acknowledge the Instituto Nacional de Investigacao Cientificafor financial support. The work was supported, in part, by theCommission of the European Communities (contract EN 3B-0081-P).
LITERATURE CITED
1. Allwood, M. C., and A. D. Russell. 1968. Thermally inducedribonucleic acid degradation and leakage of substances from themetabolic pool in Staphylococcus aureus. J. Bacteriol. 95:345-349.
2. Allwood, M. C., and A. D. Russell. 1970. Influence of ionic andnonionic materials on thermally-induced ribonucleic acid degra-dation and leakage in Staphylococcus aureus. J. Pharm. Sci.59:180-183.
3. Cabeca Silva, C., A. Madeira Lopes, and N. van Uden. 1982.Temperature relations of ethanol-enhanced petite mutation inSaccharomyces cerevisiae: mitochondria as targets of thermaldeath. FEMS Microbiol. Lett. 15:149-151.
4. Casey, G. P., and W. M. Ingledew. 1986. Ethanol tolerance inyeasts. Crit. Rev. Microbiol. 13:219-290.
5. Eddy, A. A. 1982. Mechanism of solute transport in selectedeukaryotic microorganisms. Adv. Microb. Physiol. 23:1-78.
6. Gutkneat, J., and D. C. Toteson. 1970. Ionic permeability of thinlipid membranes. J. Gen. Physiol. 55:359-374.
7. Herbert, D., P. J. Phipps, and R. E. Strange. 1971. Chemicalanalysis of microbial cells. Methods Microbiol 5B:209-344.
8. Ingram, L. O., and T. M. Buttke. 1984. Effects of alcohols onmicroorganisms. Adv. Microb. Physiol. 25:253-300.
9. Jimknez, J., and N. van Uden. 1985. Use of extracellularacidification for the rapid testing of ethanol tolerance in yeasts.Biotechnol. Bioeng. 27:1596-1598.
10. Jones, R. P., N. Pamment, and P. F. Greenfield. 1981. Alcoholfermentation by yeasts-the effect of environmental and othervariables. Proc. Biochem. 16:42-49.
11. Lafon-Lafourcade, S., C. Geneix, and P. Ribereau-Gayon. 1984.Inhibition of alcoholic fermentation of grape must by fatty acidsproduced by yeasts and their elimination by yeast ghosts. Appl.Environ. Microbiol. 47:1246-1249.
12. Leio, C., and N. van Uden. 1982. Effects of ethanol and otheralkanols on the glucose transport system of Saccharomycescerevisiae. Biotechnol. Bioeng. 24:2601-2604.
13. Ledo, C., and N. van Uden. 1983. Effects of ethanol and otheralkanols on the ammonium transport system of Saccharomycescerevisiae. Biotechnol. Bioeng. 25:2085-2090.
14. Ledo, C., and N. van Uden. 1984. Effects of ethanol and otheralkanols on the general amino acid permease of Saccharomycescerevisiae. Biotechnol. Bioeng. 26:403-405.
15. Leao, C., and N. van Uden. 1984. Effects of ethanol and otheralkanols on passive proton influx in the yeast Saccharomycescerevisiae. Biochim. Biophys. Acta 774:43-48.
16. Lee, T. C., and M. J. Lewis. 1968. Mechanism of release ofnucleotidic material by fermentating brewer's yeast. J. FoodSci. 33:124-128.
17. Lee, T. C., and M. J. Lewis. 1968. Identifying nucleotidicmaterials released by fermenting brewer's yeast. J. Food Sci.
33:119-123.18. Lewis, M. J. 1964. Release of substances absorbing ultraviolet
light by brewer's yeast suspended in diluted wort. Am. Soc.Brew. Chem. Proc. 1964:207-210.
19. Loureiro-Dias, M. C., and J. M. Peinado. 1982. Effect of ethanoland other alkanols on the maltose transport system of Saccha-romyces cerevisiae. Biotechnol. Lett. 4:721-724.
20. Miller, G. L. 1959. Use of dinitrosalicylic acid reagent fordetermination of reducing sugar. Anal. Chem. 31:426-428.
21. Moore, S., and W. H. Stein. 1954. A modified ninhydrin reagentfor the photometric determination of amino acids and relatedcompounds. J. Biol. Chem. 211:907-913.
22. Nabais, R. C., I. Sa-Correia, and J. M. Novais. 1987. Improvedyeast tolerance to ethanol-induced death by centrifugates ofdense cell suspensions incubated with ethanol. Biotechnol.Lett. 9:6-9.
23. Nash, C. H., and N. A. Sinclair. 1968. Thermal injury and deathin an obligately psychrophilic yeast, Candida nivalis. Can. J.Microbiol. 14:691-697.
24. Osman, Y. A., and L. 0. Ingram. 1985. Mechanism of ethanolinhibition of fermentation in Zymomonas mobilis CP4. J. Bac-teriol. 164:173-180.
25. Rosa, M. F., I. Sa-Correia, and J. M. Novais. 1987. Productionof ethanol at high temperatures in the fermentation of Jerusalemartichoke juice and a simple medium by Kluyveromyces mar-xianus. Biotechnol. Lett. 9:441-444.
26. Sa-Correia, I. 1986. Synergistic effects of ethanol, octanoic anddecanoic acids on the kinetics and the activation parameters ofthermal death in Saccharomyces bayanus. Biotechnol. Bioeng.28:761-763.
27. Sa.-Correia, I., and N. van Uden. 1982. Effects of ethanol on thethermal death and the maximum and the minimum temperaturefor growth of the yeast Kluyveromyces fragilis. Biotechnol.Lett. 4:805-808.
28. Sa-Correia, I., and N. van Uden. 1983. Effect of ethanol on thefructose transport system of Kluyveromyces fragilis. Biotech-nol. Lett. 5:413-418.
29. Sa-Correia, I., and N. van Uden. 1983. Temperature profiles ofethanol tolerance: effects of ethanol on the minimum and themaximum temperatures for growth of the yeasts Saccha-romyces cerevisiae and Kluyveromyces fragilis. Biotechnol.Bioeng. 25:1665-1667.
30. Sa-Correia, I., and N. van Uden. 1986. Ethanol-induced death ofSaccharomyces cerevisiae at low and intermediate growth tem-peratures. Biotechnol. Bioeng. 28:301-303.
31. Simoes-Mendes, B., A. Madeira Lopes, and N. van Uden. 1978.Kinetics of petite mutation and thermal death in Saccharomycesgrowing at supraoptimal temperatures. Z. Allg. Mikrobiol.18:275-279.
32. Strange, R. E., and M. Shon. 1964. Effects of thermal stress onviability and ribonucleic acid of Aerobacter aerogenes in aque-ous suspension. J. Gen. Microbiol. 34:99-114.
33. van Uden, N., L. Carmo Sousa, and M. Farinha. 1958. On theintestinal yeast flora of horses, sheep, goats and swine. J. Gen.Microbiol. 19:435-445.
34. van Uden, N. 1985. Ethanol toxicity and ethanol tolerance inyeasts. Annu. Rep. Ferm. Proc. 8:11-58.
35. Viegas, C. A., I. Sa-Correia, and J. M. Novais. 1985. Nutrient-enhanced production of remarkably high concentrations ofethanol by Saccharomyces bayanus through soy flour supple-mentation. Appl. Environ. Microbiol. 50:1333-1335.
36. Viegas, C. A., I. Sa.-Correia, and J. M. Novais. 1985. Synergisticinhibition of the growth of Saccharomyces bayanus by octanoicand decanoic acids. Biotechnol. Lett. 7:611-614.
VOL. 54, 1988 909
on August 29, 2018 by guest
http://aem.asm
.org/D
ownloaded from


















