
ESTRUCTURA DE LA COMUNIDAD DE MACROALGAS DE LA … · Distribución vertical estacional deSpyridia...
109
INSTITUTO POLITECNICO NACIONAL CENTRO INTERDISCIPLINARIO DE CIENCIAS MARINAS DEPARTAMENTO DE PESQUERIAS Y BIOLOGIA MARINA CWMAR W;N’ I’JLO INTERDISCIPLiNARIO DE CIENCIAS MARINAS ‘BIBLIOTECA 1. P. N. ‘ DONATIVO ESTRUCTURA DE LA COMUNIDAD DE MACROALGAS DE LA LAGUNA SAN IGNACIO, B.C.S., MÉXICO (1992-93). TESIS QUE COMO REQUISITO PARA OBTENER EL GRADO DE MAESTRO EN CIENCIAS PRESENTA: ROBERTO AURELIO NÚÑEZ LÓ PEZ LA PAZ, B.C.S. MAYO 1996
-
Upload
phunghuong -
Category
Documents
-
view
223 -
download
0
Transcript of ESTRUCTURA DE LA COMUNIDAD DE MACROALGAS DE LA … · Distribución vertical estacional deSpyridia...

INSTITUTO POLITECNICO NACIONALCENTRO INTERDISCIPLINARIO DE CIENCIAS MARINAS
DEPARTAMENTO DE PESQUERIAS Y BIOLOGIA MARINACWMAR
W;N’I’JLO INTERDISCIPLiNARIO DECIENCIAS MARINAS
‘BIBLIOTECA1. P. N.‘ DONATIVO
ESTRUCTURA DE LA COMUNIDAD DE MACROALGASDE LA LAGUNA SAN IGNACIO, B.C.S., MÉXICO (1992-93).
TESIS
QUE COMO REQUISITO PARA OBTENER EL GRADODE MAESTRO EN CIENCIAS
PRESENTA:
ROBERTO AURELIO NÚÑEZ LÓ PEZ
LA PAZ, B.C.S. MAYO 1996

t
1.
2.
3.
4.
5.
GLOSARIO . . . . . .
LISTA DE FIGURAS
LISTA DE TABLAS .
RESUMEN . . . . . .
ABSTRACT . . . . . .
INTRODUCCION . .
ANTECEDENTES . .
JUSTIF ICACION . .
OBJETIVOS . . . . .
AREA DE ESTUDIO
5. 1. Ubicación Y5. 2. Ubicación Y
.......................................... III
.......................................v
...................................... Vii
.... . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . VIII
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . iX
. . . . . . . . . . . . . . . . . . . . . . ..; . . . . . . . . . . . . . . 1
....................................... 3
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 7
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 8
caracterización fisiográfica .................... 8descripción de las localidades estudiadas . . . . . . . . . 1 1
6. METODOLOGIA . . . . . . . . . . . . . . . . . . .
6. 1.6. 2.
6. 3.6. 4.6. 5.
Flora ficológica .............Distribución ................6. 2. 1. Similitud entre localidades
Estacionalidad ..............Abundancia ................
6. 6.6. 7.
Principales componentes ficológicos:vertical . . . . . . . . . . . . . . . . . . . .Especies epi l í t icas y epífitas . . . . . .Factores ambientales: temperatura y
7. RESULTADOS . . . . . . . . . . . . . . . . . . . .
CONTENIDO
CIENCIAS MARINAS -BIBL IOTECA
LRN.DONATIVO
7. 1.7. 2.
7. 3.
Flora ficológica . . . . , . . . . . . . .Distribución . . . . . . . . . . . . . . . .7. 2. 1. Similitud entre localidades
Estacionalidad . . . . . . . . . . . . . .
ii
. . . . . . . . . .
..........
..........
..........
..........
..........estacionalidad....................s a l i n i d a d . . .
. . . . . . . . . .
..........
..........
..........
..........
........... 13
. . . . . . . . . . . 14
. . . . . . . . . . . 15
. . . . . . . . . . . 15
. . . . . . . . . . . 16
........... 17y distribución. . . . . . . . . . . 18. . . . . . . . . . . 19. . . . . . . . . . . 19
. . . . . . . . . . . 2 0
. . . . . . . . . . . 2 0
. . . . . . . . . . . 21
. . . . . . . . . . . 31
........... 3 4

7. 4. Abundancia ...................................... 377. 5. Principales componentes ficológicos: estacionalidad y distribución
vertical ......................................... 437. 5. 1. Gracilaria pacifica ........................... 437. 5. 2. Sp yridia filamentosa .......................... 4 47. 5. 3. Hypnea valentiae ............................ 467. 5. 4. Sargassum sinicola .......................... 477. 5. 5. Codium cuneatum ........................... 4 9
-7. 6. Especies epilíticas y epífitas ........................... 507. 7. Factores ambientales ............................... 52
7. 7. 1. Temperatura ............................... 527. 7. 2. Salinidad ................................. 53
8. DISCUSION . . . . . . . . . . . . . . . . . . . . . . . . . . . . ._ . . . . . . . . . . . . . . 55
8. 1. Flora ficológica ................................... 558. 2. Distribución ...................................... 59
8. 2. 1. Similitud entre localidades ...................... 6 48. 3. Estacionalidad .................................... 668. 4. Abundancia ...................................... 708. 5. Principales componentes ficológicos ..................... 73
8. 5. 1. Gracilaria pacifica ........................... 748. 5. 2. Sp yridia filamen tosa .......................... 758. 5. 3. Hypnea valentiae ............................ 768. 5. 4. Sargassum sinicola .......................... 778. 5. 5. Codium cuneatum ........................... 78
8. 6. Especies epilíticas y epífitas ........................... 798. 7. Factores ambientales ............................... 81
8. 7. 1. Temperatura ............................... 818. 7. 2. Salinidad ................................. 82
9. CONCLUSIONES . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 83
10. RECOMENDACIONES . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 86
ll. LITERATURA CITADA . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 88
iii

_GLOSARIO
Anual. Organismo que vive menos de un año. En el caso de las plantas son aquellasque nacen, se desarrollan, florecen y fructifican durante un sólo período
i vegetativo, su duración no pasa de un año y mueren después de fructificar.
Ambiente. Conjunto de elementos naturales, bióticos y abióticos, o inducidos por elhombre que interactúan en un espacio y tiempo determinados.
Biomasa. Peso total de materia que constituye a un ser vivo y que se encuentra enforma de proteínas, carbohidratos, lípidos y otros compuestos orgánicos. Esexpresada por unidad de área o volumen.
Comunidad. Conjunto de poblaciones de organismos vivos en un sitio o hábitat dado;unidad ecológica empleada en sentido amplio para inclÚir grupos de organismosde diversos tamaños y grados de integración.
Clasificación. Agrupación de objetos en clases sobre la base de atributos que poseenen común y/o sus relaciones.
Determinación. Ubicación de un objeto no identificado en la clase o grupo al quecorresponde conforme a una clasificación previa.
Dinámica. En ecología de poblaciones, se refiere al estudio de las razones de cambiosen el tamaño de la población.
Epífita. Planta que se desarrolla sobre la superficie de otra planta, generalmente noparasítica.
Epilítica. Planta que se desarrolla directamente sobre la superficie de las rocas. En estetrabajo se cons.idera epilíticas a las algas que se desarrollan tanto en sustratosduros como en sustratos blandos.
Fenología. Estudio de los fenómenos biológicos de los organismos, sincronizados acierto ritmo periódico y relacionados con el clima.
Florística. Relativo o perteneciente a la flora: estudios florísticos. // f. Parte de lafitogeografía consagrada a inventariar las entidades sistemáticas de un país,dando el área de cada una de ellas e indicaciones relativas a su hábitat,abundancia o escasez, época de floración, etc.
Indice de similitud. Razón de semejanza entre dos conjuntos de elementos. Mide lasimilitud en cada par de comunidades con base a las especies que compartenen común.
iv

Macroalga. Organismo pluricelular constituído por células indiferenciadas, deestructura generalmente tallosa, fijo al litoral o a fondos rocosos, que presentauna gran variedad morfológica y cuyos tamaños van desde unos cuantos
: centímetros hasta más de 60 m de longitud. Incluye especies de las divisionesRhodophyta, Phaeophyta y Chlorophyta.
Perenne. Planta que vive más de un año.
Perturbación. Es un evento que origina cambios en las condiciones normales de unaComunidad natural alterando las condiciones del sistema.
Reserva de la biosfera. Area biogeográfica relevante mayor a 10 000 hectáreas,sujeta a un régimen de protección. Es un área representativa de uno o másecosistemas no alterados significativamente por la actìuldad del hombre en quehabitan especies consideradas endémicas, amenazadas o en peligro deextinción.
Senescente. Adj. Que envejece.
Sistemática. Se emplea para definir el estudio científico de las clases y diversidad deorganismos y sus interrelaciones; comprende clasif icación, taxonomía ydeterminación.
V

LISTA DE FIGURAS
Fig. 1.\‘F ig . 2 .
Fig. 3.
Fig. 4.
Fig. 5.
Fig. 6.
Fig. 7.
Fig. 8.
Fig. 9.
Fig. 10.
F ig. ll.
Fig. 12.
Fig. 13.
Fig. 14.
Fig. 15.
Fig. 16.
Laguna San Ignacio, B. C. S. y localidades de muestreo.
Caracterización fisiográfica de la laguna San Ignacio, B. C. S. segúnSwartz y Cummings (1978).
Riqueza específica de macroalgas por división en la laguna San Ignacio,B. C.S (1992-93).
Riqueza específica de macroalgas por división en las diferenteslocalidades (1992-93).
Dendrograma de similitud ficoflorística entre las 19calidades de la lagunaSan Ignacio, B. C. S. a partir del Indice de Jaccard durante 1992-93.
Dendrograma de similitud ficoflorística estacional entre las localidadesde la laguna San Ignacio, B. C. S. a partir del Indice de Jaccard durante1992-93.
Riqueza específica estacional de las macroalgas por división (1992-93).
Riqueza específica espacio-temporal de las macroalgas (1992-93).
Contribución de las macroalgas por división a la abundancia total( 1992-93).
Abundancia estacional de las macroalgas por división (1992-93).
Abundancia espacio-temporal de las macroalgas (1992-93).
Abundancia espacial de las macroalgas por división (1992-93).
Abundancia relativa de los principales componentes ficológicos(1992-l 993).
Contribución de los principales componentes ficológicos (PCF) a laabundancia total de cada localidad (1992-l 993)
Abundancia de los principales componentes ficológicos en cada localidad( 1992-93).
Patrón de abundancia estacional de Gradaría pacifica (1992-93).
vi

_
. . Fig. 17.
Fig. 18.
‘Fig. 1 9 .
Fig. 20.
Fig. 21.‘
Fig. 22.
Fig. 23.
Fig. 24.
Fig. 25.
Fig. 26.
Fig. 27.
Fig. 28.
Fig. 29.
Fig. 30.
Fig. 31.
Distribución vertical estacional de Gracilaria pacifica (1992-93).
Patrón de abundancia estacional de Spyridia filamentosa (1992-93).
Distribución vertical estacional de Spyridia flamentosa (1992-93).
Patrón de abundancia estacional de Hypnea valentiae (1992-93).
Distribución vertical estacional de Hypnea valentiae (1992-93).
Patrón de abundancia estacional de Sargassum sinicola (1992-93).
Distribución vertical estacional de Sargassum sinicola (1992-93).
Patrón de abundancia estacional de Codium cuniatum (1992-93).
Distribución vertical estacional de Codium cuneatum (1992-93).
Estacionalidad de las comunidadades de algas epilíticas y epífitas( 1992-93).
Abundancia estacional de macroalgas epilíticas y epífitas (1992-I 993).
Variación estacional de la temperatura del agua en la laguna SanIgnacio, B. C. S. (1992-93).
Temperatura del agua en diferentes localidades de la laguna San Ignacio,B. C. S. (1992-93).
Variación estacional de la salinidad en la laguna San Ignacio, B. C. S.( 1992-93).
Salinidad en diferentes localidades de la laguna San Ignacio, B. C. S.( 1992-93).
vii

* LISTA DE TABLASr
Tabla 1. Trabajos ficológicos en donde se citan especies de algas colectadas en
: la laguna San Ignacio, B. C. S.
Tabla 2. Lista sistemática de la flora ficológica de la laguna San Ignacio, B.C.S.,durante 1992-93 (Arreglo de acuerdo a Wynne 1986) y afinidad de lasespecies.
Tabla 3. Especies exclusivas o de distribución restringida a una localidad en lalaguna San Ignacio, B. C. S. (1992-93).
Tabla 4. Distribución espacial y temporal de la flora ficológica de la laguna SanIgnacio, B. C. S. (1992-93).
Tabla 5. Especies anuales estacionales que ocurren en una época (1992-93).
Tabla 6. Abundancia estacional de las especies de macroalgas en la laguna de SanIgnacio, B. C. S. (1992-l 993).
viii

,.RESUMEN
: Este estudio representa la primera investigación sistemática en la laguna San
Ignacio, B. C. S. para determinar la composición específica de algas marinas
bentónicas, su distribución, estacionalidad y abundancia. Los muest reos se
realiza.ron en cada época del año durante 1992-l 993 (mayo, agosto y noviembre de
1992, y febrero de 19931, en ocho localidades ubicadas en la boca, parte media y
cabecera de la laguna. Las algas fueron colectadas por buceo libre o empleando
equipo SCUBA, a lo largo de un transecto de 200 m que abarcó tanto la zona litoral
como la sublitoral. La unidad de muestreo fue un cuadro de-un metro por lado. Se
identificaron 97 especies; 59 fueron de la división Rhodophyta, 15 de la Phaeophyta
y 23 de la Chlorophyta, de las cuales 31 % fueron algas epífitas. La ficoflora de la
laguna fue representativa de una zona de transición templada-cálida, sin embargo, la
mayoria de las especies fueron de afinidad templada. Las localidades con sustrato duro
presentaron mayor riqueza que las de sustrato blando, siendo Punta Choya la de
mayor número de especies (71) mientras que Los Médanos la de menor (10). De las
especies, 28 ocurrieron sólo en una localidad y generalmente en una época del año.
Estacionalmente, los
registrando el menor
verano. En cuanto a
cambios en la riqudza específica fueron poco notables,
número de especies (57) en primavera y el mayor (65) en
abundancia, las algas rojas fueron las de mayor biomasa,
contribuyendo con el 78.5% de la biomasa total, seguidas por las pardas con 13 %
y las verdes con el 8.5 % restante. La menor biomasa se registró en primavera con
67 g peso húmedo/m* ,mientras que en verano la mayor, con 290 g peso húmedo/m*.
Las especies más abundantes fueron; Gracilaria pacifica, Sp yridia filamen tosa e
Hypnea valentiae, las cuales aportaron el 65 % de la biomasa total. Otras especies
características de la comunidad de macroalgas de la laguna fueron; GraciIariopsW
lemaneiformis, Sargassum sinicola, Gracilaria subsecunda ta y Codium cunea tum.
ix

‘_ ABSTRACT
: This study is the first systematic survey in Laguna San Ignacio, B. C. S. to
determinate the composition of benthic marine algae, their distribution, seasonality and
abundance. Sample collection was carried out once each season during 1992-93
(May, %August and November 1992 and February 1993) at eight locations, placed at
the moúth, middle and head of the lagoon. The algae were collected by free or SCUBA
diving, along a 200 m transect in both eulittoral and sublittoral zones. The sample
unit was a one-meter square. Ninety seven species were identified; 59 were
Rhodophyta, 15 Phaeophyta and 23 Chlorophyta, 31 % of them were epiphytic algae.
The phycologic flora in the lagoon was representative of a temperate-warm transition
zone, however, most species were of temperate affinity. Localities with hard
substrate showed greater species richness than those with soft substrate; Punta
Choya being the site with the greatest number of species (71) while Los Medanos
showed the smallest number (10). Twenty eight species ocurred only in one place and
often in just one season. The species richness varied little from season to season,
however, the minimum (57) was in the spring and the maximum (65) was in summer.
With respect to abundance; the greatest biomass corresponded to the red algae,
which was 78.5% of the biomass, followed by brown algae (13%) and green algae
(8.5%). The smallest biomass was measured in spring with 679 wet weightl m2
while the greatest biomass ocurred in summer with 290 g wet weigthl m2. The most
abundant species were; Gracilaria pacifica, Sp yridia filamentosa and H y p n e a
valentiae, which was 65 % of the biomass. Other characteristic species of the
seaweed community in the lagoon were; Gracilariopsis lemaneiformis, Sargassum
sinicola, and Codium cuneatum .
X

_v 1. INTRODUCCION
L La República Mexicana dispone aproximadamente de 10 000 Km de litoral con
rasgos fisiográficos variables y
ficológica muy diversa a lo largo
atributos comunitarios diferentes1
diferentes tipos de clima, que favorecen una flora
de sus costas con características morfológicas y
(Guzmán del Proo et al. 1986).
Particularmente, en la Península de Baja California, existe una importante
reserva de algas de interés comercial, por lo que se ha establecido en este lugar la
pesquería del recurso algal mexicano (Robledo 1990). kas componentes más
importantes son: Macrocystis pyrifera (Linnaeus) C. Agardh, Gelidium robustum
(Gardner) Hollenberg y Abbott, y Chondrachantus canaliculatus (Harvey) Guiry, antes
Gigartina canaliculata Harvey (Hommersand et al. 19931, mismos que han sido
explotados de manera ininterrumpida desde 1956. A partir de los ochenta y en forma
esporádica se han explotado Egregia menziesii (Turner) Areschoug, Porphyra
perforata J. Agardh, y Euchema uncinatum Setchell y Gardner. Recientemente
Gracilaria pacifica Abbott se integra a las algas comercialmente explotadas (Zertuche
1993a).
La extracción de algas, básicamente sustentada por mantos naturales, genera
empleos y divisas que contribuyen al mejoramiento de la economía nacional y al
desarrollo regional. Por su volúmen de explotación, el recurso algal es uno de los
principales recursos pesqueros del país, ocupando, para el periódo de noviembre de
1992 a octubre de 1993, el quinto lugar de producción después de la sardina,
mojarra, camarón y atún (Garcia 1994). En los últimos años, México contribuyó con
3.2 % a la producción mundial de algas, siendo los ingresos por exportación de 1.6,
1.5 y 2.6 millones de dólares para 1990, 1991 y 1992 respectivamente (Zertuche
1993a, 1993b).
1

Actualmente, de las algas explotadas, Macrocystis es la especie más estudiada
y la única que cuenta con medidas de regulación (Diario oficial, 26 de junio de 1987).
: Para el resto, existe gran desconocimiento de aspectos biológicos y ecológicos.
Además de las algas económicamente importantes de la península, tanto en la
costa pacífica como en el Golfo de California, existen otras que por su abundancia yL
usos qbe se les atribuyen, representan recursos potenciales, como: Sargassum C.
Agardh spp, Spyridia filamentosa (Wulfen) Harvey, Hypnea valentiae (Turner)
Montagne, Laurencia Lamoroux spp, Enteromorpha Link spp, Uva Linnaeus spp y
Caulerpa sertularioides (Gmelin) Howe, entre otras. Sin embargo, falta mucho por
conocer sobre su biología y ecología y en general en las diferentes disciplinas de la
investigación ficológica.
En nuestro país, las investigaciones se han orientado básicamente a la
obtención de listados florísticos (Serviere 1993), siendo escasas las de tipo biológico,
ecológico y cuantitativo. Los estudios florísticos, en su mayoría, se han realizado en
la zona supralitoral y eulitoral (intermareal), sobre plataformas rocosas y pozas de
mareas, existiendo pocos realizados en la zona sublitoral (submareal). Además, gran
parte de ellos no presentan un método de muestreo sistemático y las colectas se
hacen en una o dos épocas del año.
Los estudios florísticos son de gran importancia ya que representan el punto de
partida para conocer la riqueza de determinado lugar y para detectar posibles recursos
potenciales, mientras que los biológicos, ecológicos (encaminados a caracterizar a las
comunidades de algas) y cuantitativos (para conocer la disponibilidad del recurso)
constituyen una necesidad cuando se trata de establecer las medidas de regulación
requeridas para su explotación.
Aún cuando las investigaciones ficológicas en México han sido básicamente
florísticas, todavía existen lugares no explorados, como es el caso de la laguna San
2

Ignacio en Baja California Sur. Dicha laguna, representa uno de los principales cuerpos
costeros del Estado por la actividad pesquera y turística que en ella se desarrolla.
: Además, reviste singular importancia ecológica debido a que forma parte de la Reserva
de la Biosfera El Vizcaíno, el área protegida más grande del país (Breceda et al.
1991).
.Por
el manejo
naturales,
lo anterior y considerando que el enfoque ecológico es indispensable para
de áreas protegidas, y en general para el aprovechamiento de recursos
este trabajo describe algunos aspectos relacionados con la estructura de la
comunidad de macroalgas; composición espacial y temporal de la flora fìcológica,
abundancia, y distr ibución horizontal y vert ical de las principales especies
componentes.
2. ANTECEDENTES
El conocimiento que se tiene de la ficoflora de la península de Baja California
se debe en gran medida a estudios y contribuciones de investigadores extranjeros,
principalmente a E. Y. Dawson (1944, 1948, 1953, 1960a, 1960b, 1961 a, 1961 b,
1962, 1963) quien dedicó gran parte de su obra científica a la vegetación marina de
la costa del Pacífico. Aguilar-Rosas et a/. (1984, 1990), Aguilar-Rosas y Aguilar-
Rosas 1986, Agu i l a r -Rosas y Pacheco-Ruíz 1985, Huer ta -Muzqu iz y
Mendoza-González 1985, Mendoza-González y Mateo-Cid 1985, Sánchez-Rodríguez
et al. 1989, Riosmena et al. 1991, y Mateo-Cid et al. 1993, entre otros, son
investigadores nacionales que también han enriquecido el conocimiento ficológico de
la región.
Particularmente, en la laguna San Ignacio, B. C. S., las investigaciones
biológicas realizadas en ella son escasas. A la fecha sólo se ha trabajado con
mamíferos marinos (Swartz y Cummings 1978, Jones y Swartz 1984), aves
(Reitherman y Storrer 1981, Danemann 1994) y peces (Danemann y De La Cruz
3

‘. 1993). En el campo ficológico, únicamente se tiene conocimiento de 13 registros de
algas: 12 especies y una variedad (Tabla l), colectadas de manera esporádica,
!desconociéndose el sitio y temporada de colecta.
TABLA 1. TRABAJOS FICOLOGICOS EN DONDE SE CITAN ESPECIES DE ALGASCOLECTADAS EN LA LAGUNA SAN IGNACIO, B. C. S.
leferencia Especie
)awson 1948 Colpomenia sinuosa (Roth) Derbes y Solier 1851Hydroclathrus clathratus (C. Agardh) Howe 1920Scinaia johnstoniae Setchell 19 13Amphiroa valonioides Yendo 1902
(Amphiroa annulata Lemoine 1929)Lomentaria hakodatensis Yendo 1920Griffitsia furcellata J. Agardh 1842
(Griffitsia multiramosa Taylor 1939)Hypoglossum attenuatum Gardner 1927Dasya baillouviana (Gmelin) Montagne 1824
(Dasya pedycellata Dawson 1944)Polysiphonia johnstonii Setchell y Gardner 1924
var concinna (Hollenberg) Hollenberg 1961(Polysiphonia concinna Hollenber 1944)
Polysiphonia mollis Hooker y Harvey 1847(Polysiphonia eastwoodae Setchell y Gardner 1930)(Polysiphonia snyderae Kylin 1941)
Iawson 1961 a Lomentaria hakodatensis Yendo 1920Griffitsia furcellata J. Agardh 1842
(Griffithsia multiramosa Taylor 1939)
>awson 1962 Callithamnion marshallense Dawson 1957Hypoglossum attenuatum Gardner 1927
var abyssicolum (Taylor) Dawson 1962
rguilar-Rosas yiguilar-Rosas993a Sargassum muticum (Yendo) Fensholt 1955lota: Dentro del paréntesis se indican los nombres con que se reportaron algunas especies y que ahora
representan sinonimias.
4

_, 3. JUSTIFICACION
: El presente estudio se originó del proyecto “Evaluación Estacional de la
Ictiofauna y Macroalgas de la laguna San Ignacio, B. C. S.“, el cual se enmarco dentro
del programa: Inventario y Sistemática de Recursos Marinos de Baja California llevado
a cabo por el Centro Interdisciplinario de Ciencias Marinas (CICIMAR-IPN).
La laguna San Ignacio junto con la laguna Ojo de Liebre, bahía Magdalena,
bahía Concepción y bahía de la Paz son los principales cuerpos costeros, desde el
punto de vista pesquero, de Baja California Sur. Además, Se encuentra sujeta a un
régimen de protección debido a que se incluye en la Reserva de la Biosfera El Vizcaíno
(Diario oficial, 30 de noviembre de 1988). Con anterioridad, la laguna, f u econsiderada como “Zona de reserva y refugio de aves migratorias y fauna silvestre”
(Diario oficial, 1 1 de septiembre de 1972) y como “Refugio para ballenas grávidas
y ballenatos y Zona de atracción turístico marítima” (Diario oficial, 16 de julio de
1976).
En general, con la creación de las Reservas de la Biosfera se busca la
conservación de especies y ecosistemas representativos del orbe, y la integración del
hombre a estos espacios. Para esto, se tienen que implementar medidas de manejo
adecuadas a las condiciones prevalecientes del lugar. Nuestra legislación estipula en
el artículo 68 de la Ley General de Equilibrio Ecológico y Protección al Ambiente que
cada Reserva de la Biosfera debe contar con un plan de manejo, en donde se
establezcan los lineamentos generales específicos que permitan la conservación,
preservación y utilización de los elementos naturales del área.
Para elaborar un plan de manejo, se requiere identificar necesidades de manejo,
establecer prioridades, organizar las acciones a diferentes t iempos según la
problemática particular de cada área y determinar las necesidades de coordinación
entre diferentes instituciones, autoridades y organizaciones no gubernamentales que
5

_
:- intervengan en la reserva. Además, se deben contemplar también las actividades
económicas que puedan desarrollarse en el área protegida, con particular atención a
,necesidades y sugerencias de la población local (Breceda 1992).
En este contexto, resalta la importancia que pueden tener los estudios
ecológicos en la laguna puesto que este lugar prácticamente ha sido “inexplorado
científikamente”. Conociendo aspectos de su estructura comunitaria se podrá inferir
respecto a su funcionamiento. Cabe mencionar por ejemplo, que las algas forman
parte de la base de la cadena trófica de pastoreo en los sistemas acuáticos, por lo que
su presencia o ausencia condiciona a su vez la presencs o ausencia de otros
organismos incluso de mayor importancia económica. Así, el enfoque ecológico para
la preservación y conservación de áreas naturales protegidas es indispensable para su
administración, y en general para el aprovechamiento de sus recursos naturales.
Por tanto, la información sobre composición florística, distribución, abundancia
y estacionalidad, generada al realizar esta investigación aportará en primera instancia
elementos necesarios a considerar en el Plan de Manejo de la Reserva de la Biosfera
El Vizcaíno, particularmente en lo que se refiere a el uso y aprovechamiento de uno
de sus recursos marinos, las macroalgas.
6

_4. OBJETIVOS
GENERAL:
Determinar la estructura de la comunidad de macroalgas de la laguna San
Ignacio, B. C. S. y analizar su variación estacional en un ciclo anual.
PARTICULARES:
al
b)
cl4
el
d)
Conformar la lista sistemática de la flora ficológica~de la laguna.
Determinar la distribución espacial de los componentes ficológicos.
Determinar la estacionalidad de los componentes ficológicos.
Estimar la abundancia de los componentes ficológicos.
Conocer la variación estacional de los componentes epilíticos y epífitos de la
flora ficológica.
Determinar el patrón de abundancia estacional y distribución vertical de
los principales componentes ficológicos.

. .5. AREA DE ESTUDID
,5.1 UBICAClON Y CARACTERIZACION FISIOGRAFICA
La laguna San Ignacio, se localiza en la costa occidental de la peninsula de Baja
California, entre los 26” 43’ y 26” 88’ N y los ll 3” 08’ y ll 3” 16’ 0 (Fig. l), Se
encuerltra rodeada por el desierto El Vizcaíno y pertenece al municipio de Mulegé, al
norte del estado de Baja California Sur.
Fig. 1. Laguna San Ignacio, B. C. S. y localidades de muestreo.
8

Tiene un área aproximada de 17 500 ha (Contreras 1988), con extensión de
casi 35 km de largo por 6 de ancho. Es somera, presenta una profundidad de 2 a 4
m en su mayor parte, llegando hasta los 20 m en los canales que la comunican con
el océano (Swartz y Cummings 1978). El interior de la laguna tiene un sistema de
canales separados por grandes y extensos bajos de arena (Fig. 2), muchos de los
cuales quedan expuestos durante las mareas bajas (Swartz y Cummings 1978,
Reitherman y Storrer 1981). Su costa esta representada por playas arenosas, áreas
de conglomerados roca-concha (coquina) y zonas de manglares compuestos por
Rhizophora mangle (Swartz y Cummings 1978).
El clima es semicálido, muy seco, con temperatura media anual entre 18 y 22°C;
presenta una oscilación térmica diaria de 7 a 14 “C (Contreras 1988). Las lluvias caen
predominantemente en invierno con una precipitación anual de 56 mm en promedio;
no hay agua de desagüe, ni arroyos o ríos de agua dulce fluyendo hacia la laguna
(Jones y Swartz 1984). Presenta mareas semidiurnas que varían entre 0.9 y 2.4 m.
El máximo rango mareal ocurre en los meses de febrero y marzo; las corrientes de
marea en la entrada y los canales son muy turbulentas (Jones y Swartz 1984).
Con base a las características fisiográficas que presenta, Swartz y Cummings
(1978) dividen a la laguna en tres áreas (Fig. 2):
a) laguna inferior. Comprende la entrada que la comunica con el mar. Presenta
una línea de rompiente, desde Punta Bronaugh hasta Punta Holcombe, y se extiende
al noroeste, entre Punta Piedra y la costa opuesta. En esta área, se encuentran
canales de aproximadamente 10 m de profundidad y hasta de 20 m cerca de la
entrada.
b) laguna media. Se extiende desde los límites de la laguna inferior, apartir de
Punta Piedra, hasta Campo Pachico y su orilla opuesta, a la altura del Cerro Doble.
En esta, se encuentran tres canales separados por dos grandes bajos. Del noroeste al
9

DONA$TVO , ;;,,$_
sureste, los canales se designan como exterior, medio, e interior, y tienen una
profundidad de casi 9 m, mientras que los bajos menos de 1.5 m cuando las mareas
,son altas.
c) laguna superior. Comprende la cabecera de la laguna, se extiende al norte a
partir del Cerro Doble y su orilla opuesta. Presenta dos zonas expuestas llamadas Islas
Ballena. Casi la mitad de esta área es muy somera, alcanzan 2 m o menos de
profundidad durante la máxima pleamar. La porción central de la laguna superior tiene
una profundidad máxima de 4.5 m.
Fig. 2. Caracterización fisiográfica de la laguna San Ignacio,B. C. S., según Swartz y Cummingas (1978).

_5. 2 UBICACION Y
: El presente
DESCRIPCION DE LAS LOCALIDADES ESTUDIADAS
trabajo se llevó a cabo en ocho localidades de la laguna,
seleccionadas con base: a su ubicación a lo largo y ancho de la misma, para tener
representados ambientes de la boca, parte media y cabecera; a los diferentes tipos de
sustrato que presentaron, y, a la accesibilidad para llegar a ellas.
Considerando la división propuesta por Swartz y Cummings (1978), las
observaciones de campo y los resultados obtenidos al analizar los sedimentos por el
método de Diagrama Ternario de Clasificación de Folk (19547, se caracterizaron las
localidades de estudio:
La Boca (Localidad 1). Se ubica al suroeste, en la parte inferior de la laguna.
Presenta una pendiente que gradualmente se hace pronunciada hasta alcanzar 16 m
de profundidad. El sustrato es fundamentalmente arenoso (arena fina). Se alternan
lugares protegidos por barreras de arena con zonas de rompientes (estas últimas en
el exterior de la laguna), sujetas a un fuerte oleaje y gran turbulencia provocada por
las corrientes de mareas. En áreas protegidas, el nivel intermareal superior queda
expuesto durante las mareas bajas. En el sublitoral, el movimiento del agua es
moderado en los primeros metros pero conforme se acerca al canal de entrada, el
movimiento es considerable.
Punta Piedra (Localidad 2). Se localiza al sureste, y constituye los límites de la
porción inferior de la laguna. Presenta una pendiente que se hace pronunciada (6 m
aprox.) conforme se acerca al canal de entrada. Constituye una punta de rompiente,
que alterna con lugares semiprotegidos; en los primeros 10 metros el sustrato es
rocoso, en adelante, es arenoso (arena media) con conchilla (conchas fragmentadas)
y algunos cantos rodados. En la orilla rocosa la acción del oleaje es intenso. En
general, es un área influenciada por las corrientes de marea con una franja intermareal
estrecha.
11

Campo Manuela (Localidad 3). Se localiza al oeste, en la parte media de la
laguna. Es un área protegida con una planicie uniforme de poca profundidad (hasta
,0.70 m). El sustrato es predominantemente arenoso (arena fina) y una gran extensión
queda expuesto durante las mareas bajas. Es una zona en donde el movimiento de
agua no es considerable (no esta sujeta a corrientes de mareas).
La Base (Localidad 4). Se encuentra ubicada al este, en la parte media de la
laguna. Es un área somera, protegida, con poca influencia de corrientes de mareas. El
sustrato en los primeros 30 m es fango-arenoso (limo con arena fina) y a partir de allí
es arenoso (arena fina) con conchilla, con algunos guijarrós y cantos rodados.
Presenta una franja intermareal ancha y una parte extensa de la superficie de la orilla
de la playa queda expuesta durante las mareas bajas.
Cantil Cristal (Localidad 5). Se localiza al noroeste, en la parte superior de la
laguna. Es una zona protegida, sin mucho movimiento de agua, la franja intermareal
es extensa y queda expuesta durante las bajamares. En los primeros dos metros
sobresalen de manera irregular proyecciones (arenisca), algunas veces planas, del
peñasco Cantil Cristal, que se suaviza hasta formar una planicie uniforme de poca
profundidad. El sustrato es predominantemente fango-arenoso (limo con arena fina).
Punta Choya (Localidad 6). Se encuentra en el extremo sur de Isla Ballena.
Comprende áreas de rompientes moderadas y protegidas, las primeras son
representadas por salientes o puntas de la isla, mientras las segundas son pequeñas
ensenadas. En las primeras, el sustrato es predominantemente arenoso (arena media)
con bastante presencia de conchas y conchilla, en las segundas, es fango-arenoso
(limo con arena fina) con conchilla.
Isla Garza (Localidad 7). Se ubica en el extremo norte de Isla Ballena.
Aproximadamente hasta los 5 m presenta sustrato pedregoso, apartir de ahi, es
arenoso con conchilla y algunos cantos rodados. La franja intermareal es angosta y
12

queda expuesta durante las bajamares. El movimiento del agua por las corrientes es
moderado en los primeros metros y mayor cerca de los canales de circulación.
Isla Ballena es conocida también como Isla Zopilotes, Pelícanos, Garza, Whale,
0 simplemente “Islotes”. Se ubica en la porción superior de la laguna y cubre una
superficie de 1 .3 km2. Está dividida en dos secciones separadas por un canal de
maread de aproximadamente 500 m de ancho, que se seca durante las bajamares. La
sección sur es prácticamente plana, mientras que la norte presenta acantilados de
hasta 10 m de altura. La mayor parte de la isla está constituida por playas arenosas
y planicies lodosas, encontrándose algunas porciones rocosãs (Danemann 1994).
Los Médanos (Localidad 8). Se localiza al noreste de la laguna, en la zona
superior o cabecera. Es una planicie uniforme con sustrato fango-arenoso (limo con
arena fina). Representa una zona protegida y somera, con poca circulación; la franja
intermareal es ancha.
6. METODOLOGIA
La colecta de algas y registro de parámetros ambientales se efectuó
trimestralmente durante mayo, agosto y noviembre de 1992 y febrero de 1993,
correspondientes a primavera, verano, otoño e invierno de un ciclo anual.
Los muestreos se llevaron a cabo durante las mareas muertas y la colecta se
realizó manualmente en las zonas eulitoral (intermareal) y sublitoral (submareal),
mediante buceo libre o autónomo, con equipo SCUBA cuando las algas se encontraron
a profundidades mayores de tres metros.
La metodología aplicada fue por transectos, empleados frecuentemente en
estudios de tipo ecológico para conocer la distribución y abundancia de macroalgas
(Mann 1972, Santelices 1980, Kendrick eta/. 1990, Vásquez y González 1994). La
13

unidad de muestreo fue un cuadro de un metro por lado y se eligió porque se ha
demostrado que este tamaño es más eficiente, en estudios cuanti tat ivos de
,macroalgas, para hacer mediciones de biomasa debido a las dimensiones que alcanzan
estos organismos (Santelices 1980, Vásquez y González 1994). En cada localidad se
colocaron dos transectos de 200 m de longitud, perpendiculares a la línea de costa
y separados a una distancia aproximada de 30 m. A lo largo de estos, cada 10 m de
distanc!ia se colocó la unidad de muestreo y se colectaron todas las algas contenidas
en su área, las cuales se fijaron en solución de formaldehído al 4 % con agua de mar.
En las diferentes épocas, los transectos se situaron en la misma área pero no en el
mismo lugar.
6. 1 FLORA FICOLOGICA
Las algas se separaron y se determinaron considerando los caracteres
morfológicos, citológicos y reproductivos de interés sistemático para cada especie. Las
características morfológicas externas incluyen; forma de crecimiento, color, talla, y
tipo de ramificación entre otras, mientras que las internas; tipo y arreglo de células y
plastos, presencia o ausencia de estructuras especiales como f i lamentos,
engrosamientos, etc., y tipo y organización de estructuras reproductivas. Para
conocer la disposición y arreglo estructural interno de los talos y su morfología
reproductiva, se realizaron cortes transversales y longitudinales, los cuales se
observaron en el microscopio óptico marca Will Wetzlar, a 10 y 40x.
Las claves y listas empleadas en la determinación fueron las de Abbott y
Hollenberg (1976), Dawson (1944, 1953, 1960b, 1961 b, 1962, 19631, y Taylor
(1960). Ejemplares representativos de las especies encontradas se incorporaron a la
colección ficológica del Centro Interdisciplinario de Ciencias Marinas y otros se
depositaron en la sección ficológica del Herbario de la Escuela Nacional de Ciencias
Biológicas del 1. P.N. [ENCBI y en el Herbario de la Universidad Autónoma de Baja
California Sur [HFBCS]. Algunas especies que presentaron dificultad para su
14

_determinación se enviaron a
L El arreglo sistemático
‘( 1986). Además del trabajo
especialistas para identificar su posición taxonómica.
de las especies sigue el orden propuesto por Wynne
anterior, se consultaron los de Scagel (1989), Stewart
(1991) y Hommersand et al. (1993) para actualizar la nomenclatura de algunas
especies.
Asimismo, se consideró pertinente determinar la afinidad biogeográfica de las
especies, para caracterizar la ficoflora de la laguna, consultando la literatura disponible
sobre los límites de distribución latitudinal de las especies. L-as fuentes consultadas
fueron los trabajos de Taylor (1960), Abbott y Hollenberg (19761, Schneider y Searles
(1991) y la serie de Dawson sobre las algas marinas del Pacífico Mexicano. Para fines
prácticos, se agrupó a las especies en tres categorias:l) de afinidad templada,
aquellas que se desarrollan mejor en aguas frías y son características de la región
biogeográfica templada fria, aunque también pueden distribuirse en la región templada
cálida (Lüning 1990); 2) de afinidad tropical, aquellas que tienen una preferencia
márcada por aguas cálidas y que son típicas de la región tropical; y 3) de amplia
distribución, las que se desarrollan tanto en ambientes frios como en cálidos y se
distribuyen desde la región templada fría hasta la tropical.
6. 2 DISTRIBUCION.
Se elaboraron gráficos y tablas que presentan la distribución de las especies en
las distintas localidades y épocas.
6. 2. 3 SIMILITUD ENTRE LOCALIDADES
El análisis de la estructura espacial de la comunidad de macroalgas puede
hacerse a través de la estratificación vertical de las formas de crecimiento o de la
diferenciación horizontal, relacionada con los modelos de distribución de los grupos
15

de especies (Chapman 1979). En este trabajo, La estructura espacial de la comunidad
se analizó desde el segundo enfoque debido a que es de mayor interés en estudios
:de comunidades litorales. Para ello, se empleó la técnica de clasificación por similitud
relativa o clasificación numérica. El grado de semejanza ficofloristica entre las
localidades, de manera general y para cada época del año, se determinó por medio del
índice de similitud de Jaccard. Este, se eligió porque es ampliamente recomendado en
estudids ecológicos basados en la presencia o ausencia de organismos (Ludwing y
Reynolds 1988). Con la matriz de similitud resultante se realizó un análisis de grupo
(cluster analysis) empleando como estrategia de agrupación la técnica de ligamentos
promedios no ponderados (UPGMA: Unweighted Pair Group Method Using Arithmetic
Averages), también de uso generalizado en este tipo de estudios (Crisci y López
1983). Los resultados se presentan en dendrogramas realizados por medio de l
paquete estadístico NTSYS-PC, versión 1.6.
6. 3 ESTACIONALIDAD.
Se determinó comparando los cambios en la composición florística de cada
localidad en las diferentes épocas del año. Además, con base a los criterios
establecidos por Sears y Wilce (19751, a los cambios de abundancia estacional,
como al apoyo bibliográfico para algunos casos, se caracterizó a las especies
acuerdo a su periodicidad estacional.
así
de
La categorización de las especies de algas de acuerdo a su estacionalidad o
longevidad resulta complicada debido a las diferentes manifestaciones de las especies
a lo largo de su ciclo de vida (p. e. generaciones heteromórficas con diferente período
de longevidad) y al desconocimiento biológico de la mayoría de estos organismos.
Sears y Wilce (1975), considerando la longevidad del talo, su forma durante la época
adversa y el período en que se manifiestan las especies, clasifican a las algas en:
16

Anuales estacionales: aquellas que persisten por menos de un año, pasan su
estación adversa en latencia, en estado juvenil o estan completamente ausentes de la
kcomunidad en una parte del año. Estas especies pueden ocurrir en una o dos épocas
‘y se les conoce como especies anuales estacionales de una y dos épocas
repectivamente.
Anuales no estacionales: persisten menos de un año, aunque estan presentes
en todas las épocas debido a el traslape de sus generaciones.
Pseudoperenes: persisten por más de un año, sin embargo, la mayoría de las
plantas de la población pasan su estación (es) adversa (s) en forma deteriorada.
Perennes: plantas que persisten más de un año y presentan pocas variaciones
morfológicas. La porción principal del talo se mantiene durante todo el año.
6. 4 ABUNDANCIA
La valoración de abundancia para las comunidades de algas, resulta complicada
puesto que se comparan individuos y poblaciones con diferentes niveles de
organización: costrosas, f i lamentosas y talosas entre otras, las cuales crecen
entremezcladas en asociaciones microambientales (Serviere 1993). En muchos casos
no es posible reconocer organismos individuales debido a que se fusionan para formar
organismos modulares. Por lo anterior, la abundancia se determinó en términos de
biomasa, considerando sólo a las especies que pesaron mas de 0.1 g. El peso de algas
calcáreas, articuladas y costrosas, no se consideró debido a la gran cantidad de calcio
fijada por ellas que hace impreciso los valores de biomasa. Tampoco se consideró a
las costrosas no calcáreas por la dificultad que implica su medición. La biomasa (peso
húmedo) se registró con una balanza eléctrica portátil con precisión de 0.1 gramo.
17

., Para determinar la biomasa promedio (gramos en peso húmedo/m2), se sumo la
biomasa de la especie x colectada en cada cuadrante y se dividió entre el número total
.de puntos muestreados por transecto (Brower y Zar 1984).
B = tPx/A
en donde B es igual a la biomasa expresada en g peso húmedo/m2, P es el peso de
la especie x en cada punto de colecta y A es el área total áe muestreo.
Con lo anterior fue posible describir el patrón de abundancia estacional de
macroalgas por división para cada localidad. Asimismo, se Ealculó la biomasa relativa
(contribución de la abundancia de cada especie a la abundancia total) para detectar
a las especies mas abundantes:
Br = Px/Bt * 700
en donde Br es igual a la biomasa relativa (%) de la especie x, Px es la biomasa de
la especie x y Bt es la biomasa total de las especies.
6. 5 PRINCIPALES COMPONENTES FICOLOGICOS: ESTACIONALIDAD Y
DISTRIBUCION VERTICAL.
Se definió como principales componentes ficológicos de la comunidad, a las
especies más abundantes que en suma contribuyeron con el 85 % de la biomasa total.
El patrón estacional de cada uno de los principales componentes se determinó
estimando su biomasa promedio en las diferentes épocas del año, considerando sólo
a las localidades que concentraron su mayor abundancia.
En cuanto a su distribución vertical, se calculó la biomasa promedio en cada
punto del transecto y a partir de ahi la abundancia relativa, para determinar la zona
18

de mayor concentración a lo largo del gradiente vertical.
6. 6. ESPECIES EPILITICAS Y EPIFITAS.
Para facilitar la descripción de la estructura comunitaria se consideró pertinente
hacer una diferenciación entre la comunidad de especies epilíticas y la de epífitas. Las
algas que no crecieron sobre otras plantas se consideraron epilíticas, las cuales se
desarrollaron tanto en sustratos duros (formaciones rocosas, guijarros, cantos
rodados, conchas y fragmentos de concha o conchilla), como en sustratos blandos
(arena y fango). Tomando en cuenta que algunas especies pueden ocurrir tanto como
epilíticas como epífitas, se le asignó la categoria con base a su modo más frecuente
de aparición.
La estacionalidad de las comunidades epilítica y epífita se determinó a través
de sus variaciones, tanto en riqueza específica como en abundancia, entre épocas
del año.
6. 7 FACTORES AMBIENTALES: TEMPERATURA Y SALINIDAD.
En cada localidad, en el último punto del transecto se registró la temperatura
del agua superficial y se tomaron muestras de agua para determinar salinidad. Esto
se efectuó alrededor del medio día para fines comparativos. La temperatura se midió
con un termómetro de cubeta con presión de 0.1 OC y la salinidad se determinó por
el método de Culkin (Grasshoff 1983). Para establecer si la diferencia entre épocas
del año y localidades fue significativa, se realizó un análisis de variancia (ANDEVA) de
una vía, a través del paquete STATISTICA versión 4. 3. El límite de significancia se
estableció con un error alfa de 0.05.
1 9

7. RESULTADOS
:7.1. FLORA FICOLOGICA.
Se determinaron 97 especies de algas: 59 fueron rojas, 23 verdes y 15 pardas
(Fig. 3), que representan 61, 24 y 15 % respectivamente . El arreglo sistemático y
afinidad de las especies se presenta en la Tabla 2.
Fig. 3. Riqueza específica de macroalgas por división en lalaguna San Ignacio, B. C. S. (1992-931.
Las especies pertenecieron a 25 familias: 14 fueron de la división Rhodophyta,
seis de la Phaeophyta y cinco de la Chlorophyta. De estas 13 presentaron un sólo
registro, mientras que las mejor representadas para la primera división fueron,
Rhodomelaceae y Corall inaceae, con 18 y 12 especies; para la segunda,
Scytosiphonaceae, Dictyotaceae y Sargassaceae, todas con cuatro especies; y para
la tercera, Ulvaceae y Cladophoraceae, con ll y siete especies repectivamente.
20

‘” En general, la ficoflora de la laguna estuvo constituída por una mezcla de
componentes de afinidad templada y tropical. Los primeros representaron 43 %,
,mientras que los segundos, 16 %. El 41% restante correspondió a especies de amplia
distribución latitudinal, es decir, especies que se desarrollan tanto en ambientes
templados como tropicales.
7. 2. ISETIWK~~N
En términos generales, la localidad con mayor riqueza específica fue Punta
Choya (711, seguida por Punta Piedra (63) e Isla Garza (581, mientras que Los
Medanos (10) y Cantil Cristal (12) fueron las localidades con menor riqueza. Respecto
a las diferentes divisiones, el mayor número de Rhodophyta (44) y Chlorophyta (18)
se encontró en Punta Choya, y el de Phaeophyta (í2) en Punta Piedra (Fig. 4).
1 2 3 4 5 6 7 6
LOCALIDAD
a RHODOPHYTA 0 P H A E O P H Y T A m C H L O R O P H Y T A -e- T O T A L
Fig. 4. Riqueza específica de macroalgas por división en lasdiferentes localidades (1992-93).
21

TABLA 2. LISTA SISTEMATICA DE LA FLORA FICOLOGICA DE LA LAGUNA SANIGNACIO, B. C. S. DURANTE 1992-93 (Arreglo de acuerdo a Wynne1986) Y AFINIDAD DE LAS ESPECIES.
RHODOPHYTA
Bangiophycidae AFINIDAD
PORPHYRIDIALES
Goniotrichaceae
“Chroodactylon ornatum (C. Agardh) Basson(Asterocytis ramosa (Thwaites) Schmitz)
Florideophycidae
GELIDIALES
Gelidiaceae
Selidium pusillum (Stackhouse) Le JolisQterocladia caloglossoides (Howe) Dawson
CORALLINALES
Corallinaceae
4mphiroa beauvoisi Lamouroux A(Amphiroa zonata Yendo)
4mphiroa Lamouroux spJorallina frondescens Postels et RuprechtZorallina vancouveriensis YendoJorallina Linnaeus spYydrolíthon decipiens (Foslie) AdeyJania adhaerens Lamouroux
ATTTAC
(Jania capillacea Harvey)“Jania Lamouroux splithoph yllum imitans Foslielithoph yllum Philippi spVeogoniolithon setchelli (Foslie) Adey
(Hydrolithon setchelli (Foslie) Setchell et Mason)Neogoniolithon Setchell et Mason sp
CAAT
A
T
AA
22

GIGARTINALES
Hypneaceae
Hypnea valentiae (Turner) Montagne C
Plocamiaceae
Plocan$m Lamouroux sp T
Gracilariaceae
Gracilaria marcialana DawsonGracilaria pacifica Ab bott
(Graclaria verrucosa Hudson et Papenfus)Gracilaria subsecundata Setchell et GardnerGracilaria textorii (Suringar) J. Agardh
var. cunninghamii (Farlow) DawsonGracJaria Greville sp*Gracilariophi/a gardnerii SetchellGraciiariopsis lemaneiformis (Dawson) Acleto et Foldvik
(Gracilaria lemaneiformis (Bory) Weber van Bosse)
CA
C
TAAA
Gigartinaceae
Chondrachantus canaliculatus (Harvey) Guiry(Gigartina canaliculata Harvey)
CRYPTONEMIALES
Peyssonnelliaceae
Peyssonnellia rubra (Greville) J. Agardhvar. orientalis Weber-van Bosse
Halymeniaceae
Grateloupia versicolor (J. Agardh) J. Agardh
T
A
C
RHODYMENIALES
Rhodymeniaceae
Leotofauchea pacifica Dawson T
23

Ceramiaceae
*Anotrichium tenue (C. Agardh) Nägeli(Griffthsia tenuis C. Agardh)
“Ceramium caudatum Setchell et Gardner“Ceramium flaccidum (Kützing) Ardissone“Ceramium serpens Setchel et Gardner“Ceramium Roth sp“Centroceras clavulatum (C. Agardh) Montagne“Griffithsia furcellata J. AgardhSp yridia filamentosa (Wulfen) Harvey“Tiffaniella saccorhiza (Setchel et Gardner) Doty et Meñez _
Delesseriaceae
4crosorium venulosum (Zanardini) Kylin(Acrosorium uncinatum (Turner) Kylin)
“Cryptopleura Kützing sp“Hypoglossum attenuatum Gardner
var. abyssicolum (Taylor) Dawson
Dasyaceae
Dasya baillouviana (S. G. Gmelin) Montagne(Dasya pedicellata Dawson)
Rhodomelaceae
Chondria dasyphylla (Woodward) C. Agardh“Herposiphonia secunda ( C. Agardh) Falkenberg
f. tenella (C. Agardh) Wynne(Herposiphonia tenella f. secunda (C. Agardh) Hollenberg)
“Herposiphonia Nägeli spLaurencia hancockii DawsonLaurencia gardnerii HollenbergLaurencia lajolla DawsonLaurencia masonii Sechell et GardnerLaurencia pacifica KylinLaurencia sinicola Setchell et GardnerLaurencia sn yderiae DawsonLaurencia sp Lamouroux“Polysiphonia flaccidissima Hollenberg
24
A
AAAAATCT
A
T
C
A
A
A
ATTTTTCTTT

“Polysiphonia johnstonii Setchell et Gardnervar. johnstonii Setchell et Gardner A
“Polysiphonia mollis Hooker et Harvey T“Polysiphonia pacifica Hollenberg T*Polysiphonia simplex Hollenberg C“Polysiphonia Greville sp A“Pterosiphonia dendroidea (Montagne) Falkenberg A
PHAEOPHYTA1
Phaeophyceae
ECTOCARPALES
Ectocarpaceae
“Ectocarpus parvus (Saunders) Hollenberg
Ralfsiaceae
Salfsia confusa Hollenberg
SCYTOSIPHONALES
Scytosiphonaceae
Zolpomenia sinuosa (Roth) Derbes et SolierZolpomenia tuberculata SaundersHydroclathrus clathratus (C. Agardh) HoweRosenvingea intrincata (J. Agardh) Boergesen
SPHACELARIARIALES
Sphacelariaceae
Sphacelaria californica
Dictyotaceae
(Sauvageau) Setchell et Gardner
T
T
TACC
T
Oictyota flabellata (Collins) Setchell et GardnerPadina crispa ta ThivyPadina durvilaei BoryPach ydictyon coriaceum (Holmes) Okamura
ACAT
25

FUCALES
Sargassaceae
Sargassum agardhianum J. Agardh;argassum muticum (Yendo) Fensholtgargassum sinicola Setchell et Gardner;argassum C. Agardh sp
‘CHLOROPHYTA
Chlorophyceae
ULVALES
Monostromaceae
‘Kornmannia leptoderma (Kjellman) Bliding(Monostroma zoster-kola Tilden)
Ulvaceae
Fnteromorpha clathrata (Roth) Greville(Enteromorpha clathrata var. clathratra (Roth) Greville)
kteromorpha compressa (Linnaeus) GrevilleFnteromorpha flexuosa (Roth) J. AgardhFnteromorpha intestinalis (Linnaeus) LinkFnteromorpha prolifera (0. F. Müller) J. Agardh:nteromorpha ramulosa (J. E. Smith) Carmichael
(En teromorpha cla thra ta var. crinita (Roth) Hau k)Jlva californica WilleJlva dactylifera Setchell et GardnerJlva lactuca LinnaeusJlva lobata (Kützing) Setchell et Gardner‘Jlva rigida C. Agardh
CLADOPHORALES
Cladophoraceae
+Chaetomorpha californica CollinsThaetomorpha linum (0. F. Müller) Kützing“Cladophora albida (Hudson) Kützing“Cladophora graminea Collins
TTCT
T
A
AATTA
TTAAA
TAAT
26

“Cladophora microcladiodes Collins“Cladophora sericea (Hudson) Kützing“Rhizoclonium riparium (Roth) Kützing
(Rhizoclonium implexum (Dillwyn) Kützing)
TTT
CAULERPALES
Codiaceae
1Zodium cuneatum Setchell et Gardner AZodium decorticatum (Woodward) Howe TTodium Stackouse sp T
DASYCLADALES
Polyphysaceae
ketabularia calyculus Quoy et Gaimard C__. . . . . . - - ..- - ..- _ _ . . .Afmldad Qe especles: I = Templada; C = Tropical; A = especies de amplia distribución* Especies epífitas.( 1 Sinonimias.
Del total de especies, 28 ocurrieron sólo en una localidad (12 en Punta Choya,
10 en Punta Piedra, 5 en Isla Garza y una en Los Medanos) (Tabla 3); 24 fueron
comunes en dos localidades; 18 en 3; 10 en 4; 6 en 5; 2 en 6; 6 en 7; y 3 en 8. Las
especies de amplia distribución (aquellas que se distribuyeron en 7 y 8 localidades)
fueron: Gracilaria pacifica, Sp yridia filamentosa, Cladophora microcladiodes,
Chroodactylum ornatum, Hypnea valentiae, Das ya baillouviana, Ectocarpus parvus,
Enteromorpha clathrata y Cladophora albida. Cabe señalar que su distribución cambió
en las diferentes épocas del año; por ejemplo, Chroodactylum ornatum, Dasya
baillouviana y Cladophora spp ampliaron su distribución en otoño y Ectocarpusparvus
en invierno. La distribución de los componentes ficológicos en la laguna, por localidad
y época del año, asi como su periodicidad estacional se presenta en la Tabla 4.
2 7

TABLA 3. ESPECIES EXCLUSIVAS 0 DE DISTRIBUCION RESTRINGIDA A UNALOCALIDAD EN LA LAGUNA SAN IGNACIO, B. C. S. (1992-93).
ocalidad
unta Choya
‘
Especie Estación
Corallina frondescens otoñoCoralfina sp PrimaveraHydrolithon decipiens PrimaveraPlocamium sp Verano
* Gracilarioph ylla gardneri VeranoGra teloupia versicolor PrimaveraLep to fauchea pacifica otoñoLaurencia gardnerii InviernoH ydrocla thrus cla thra tus Ver-lnvDic t yo ta flabella ta _ InviernoEn teromorpha prolifera Prim-Oto-lnvUlva loba ta otoño
unta Piedra Gracilaria textorii* Ceramium cauda tum* Ceramium serpens* Cryp topleura sp* Herphosiphonia secunda
Colpomenia tubercula taSphacelaria californicaPadina crispa taPach ydic t yum coriaceumSargassum muticum
InviernoPrimaveraPrimaveraVeranoVeranoPrim-Oto-lnvPrim-VerPrimaveraVeranoInvierno
;la Garza Pterocladia caloglossoides+ Cen troceras cla vala tum+ Tiffaniella saccorhiza“Herposiphonia spLaurencia hancockii
InviernootoñoVeranootoñoVerano
os Medanos* Especies epífitas.
Rosenvingea in trinca ta otoño
28

TABLA 4. DISTRIBUCION ESPACIAL Y TEMPORAL DE LA FLORA FICOLOGICA DE LAGUNA SAN IGNACIO S.C.S. (1992-1993).
Acmsodum venulosum1 Amphhoa beauvoisiiAmphiroa spAnobichium tenueCentroceras clavalatumCeramium caudatumCaramium flaccidumCeramrum serpens
~Ceramium sp RcthChondracnantus canaliculattisChondria dasyphyllaChroodacrylon omatumCorallina fmndescensCorallina spCorallina vancouvenerwsCIyptopkwa spDasya baillrwana ‘Gelidium ~‘,~siilumGratilana maroaranaGracilatia paoficaGracttana spGractlaña subsecundataGracilatia texbo: vac cunnmghamirGmctlancphJa gamnedGracfladcpsis remanerfcmisGrateloupfa versrcolorGdfithsia funxilataHerpostphoma s-cunda f. tenellaHerposiphonia spHydmlithon denpiensHypnea valentia aHypogtossum atkwaturn var. abyssicolumJanta adhaeransJania spLaurencia gardneniLaumncia hancockiiLaurencia /ajo//aLaurencta masoniiLaurãncia pacificaLaurancia sinicolaLaumncia snydedaeLaurencia spLeptofauchea paci8caLtthophyllum imdanstithophyllum spNeogoniolithon setchelliNeogoniolithon spPayssonnellia cubra var. otientabsPlocamium spPolystphonia flacodissimaPotysiphonia johnstoné wr. johnstoniiPotysiphonia mollisPolysiphonia pacnìcaPolysiphonia simplexPolysiphonia spPterocladia caloglossoidesPtemstphonia dendmideaSpyddia lilamenfosa7iffaniella saccorhiza
PHAEOPHYTA
Colpomenia sinuosaCoIpomenta tuberculataLMtyuta flabetlataEctocarpus pawsHydmclathrus clatiratusPachydictyon codaceum
IA” 2%=!Ratia confusaRosentingea inbtncataSargassum agardhianumSaqassum mticum6argassum sinicolaSaqassum spSphacelaria califom!ca
0
01
IVOI
0
I
01
I0101
01
:,
01
0
VO
I
JO
V O
1
I
0
VO
I
PVOII
V
! 0iv IPVOI
;P 01‘P 0
PVOI
VOII
VOIVOI101V O
fVOI,*01
VOI0VOIVOI
I
v IV
0VOI,V
fO101
v IVOI01
/
101100
I0
VOI
I
I0 1I I
V O
I
VOI01
01
0
AEP
:PjAF4sIAE
i;;
lACIANsAPic.ANsPS
‘PSP
IAEPS
iAt&‘PS/PS‘PSiPSIAEIAEPS
#AEANsAEAEPSPSAE
IPPAEAEANsAEANsANsAEANsAEPPPPPAEAEANsANsANsAEANsAEANs
IPSAE
AEANsAEANsAEAEAEPSPAEAEAEPSPS
IAE
29

Acetabulada ca/yC~l~sCbaetmo!pha californica
‘VO1Codium decorbcatumCodium spEntemmorpha clatbrata
.
Vv I v I
Entemmofpha compresa JEntemmorpha nexoosaEntemmorpha intesbri3lis IEntemmorpha pr0ltf.wEnteromotpha ramïlosa 01Kommannia leptodemn IRhizoclonium npanxn ’ 01Uva cahfomica 0Uva dacty/ffera VUva tactwa 1 VUva lobakUva ti rda
0
0
0 -
01
001
0
!
,l‘P
L
11010
1/000
01J I
0I
0
010
0
0
VOIVOI
VOIVOIv IVOI
0101
V01
VOIVv I0
l
Localidad: 1) La Boca, 2) Punta Piedra. 3) Campo Manuela, 4) La Base, 5) Cantil Cristal, 6) Punta Choya. 7) Isla Gana, 8) LOS Madanos.Estacion: (P) Primavera, (V) Verano, (0) Otorlo, (1) InviernoPeriodicidad Estacional: (AE) Anual estacional, (ANs) Anual no estacional, (PS) Pseudoperenne, (P) Perenne.
0I
V OVVOIv IVv 1
0
I
0V
V
I- -
0 IAE‘AE‘ANsANsAE
I ‘Al&Ai&ANS
‘ANsAPNSANs,AEANS
;ANsAEANS,AEAE__--- -
30

_
‘_ 7. 2. 1. SIMILITUD
En términos
ENTRE LOCALIDADES
generales, a un nivel de similitud del 29%, se formaron dos
grandes grupos (Fig. 5), que corresponden a localidades con mayor presencia de
sustratos duros (2, 6 y 7) y localidades con sustratos fundamentalmente blandos (1,
3, 4, 5 y 8). A un mayor nivel de similitud (45 %) se evidenciaron cuatro grupos: 1)
lo integran las localidades 6, 7 y 2, que presentan sustratos duros (rocas, piedras o
cantos rodados, guijarros, conchas y conchilla); II) formado por la 3 y 4, con sustrato
arenoso y fango-arenoso respectivamente; III) por la 5 y 8, con sustrato fango-
arenoso; y IV) constituído sólo por la localidad 1, que presentãsustrato arenoso.
7 6 2 8 5 4 3 1
Fig. 5. Dendrograma de similitud ficoflorísticaentre las localidades de la Laguna de San Ignacio,B. C. S., a partir del Indice de Jaccard durante1992-93.
0.00
0.25
0.50
0.75
1 .oo
31

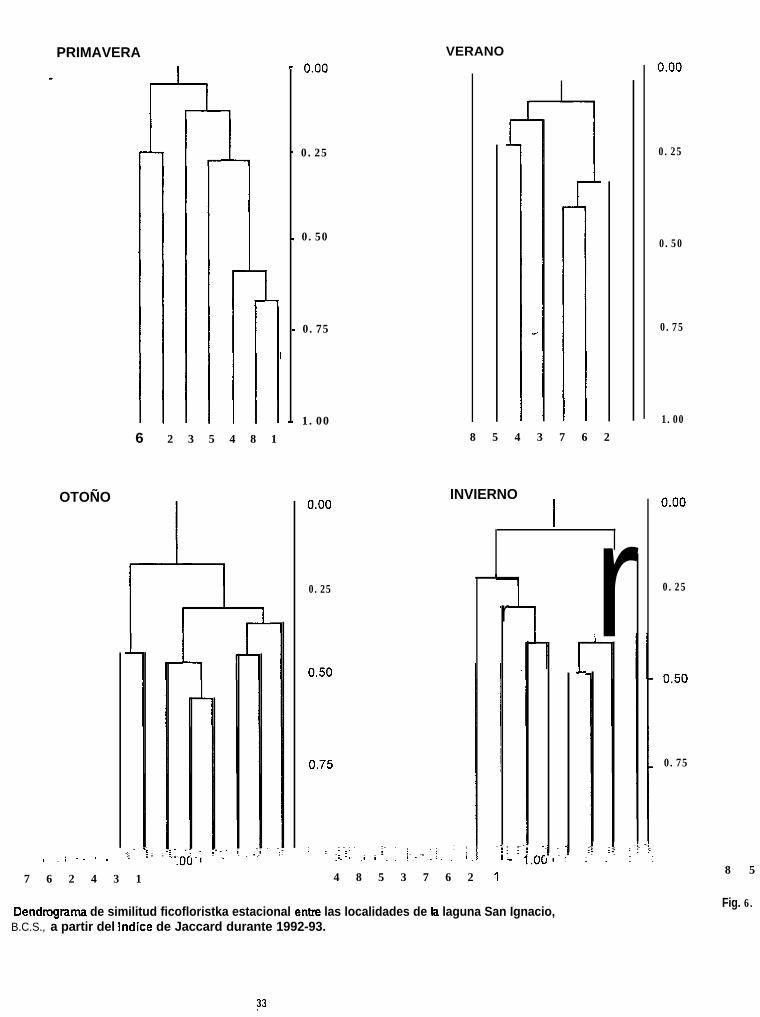
A lo largo del año, con excepción del primer grupo (localidades con sustratos
duros) que se mantuvo constante, la integración de las localidades en los diversos
%grupos que se formaron fue variable (Fig. 6).
En primavera, a un nivel bajo de similitud (10%) se reconocieron dos grupcs
(Fig. 6a). El primero estuvo constituído por las localidades 1, 8, 4, 5 y 3, todas con.
sustratos blandos. La mayor similitud se observó entre las localidades 1 y 8, con un
valor de 70 %. El segundo grupo lo formaron las localidades 2 y 6, las cuales
presentaron sustratos duros. La localidad 7 se excluye del análisis debido a que, por
diversas razones, no fué posible colectar algas en este lugar.
En verano, con una similitud de 15%, se formaron cuatro grupos (Fig. 6b); uno
integrado por las localidades 2, 6 y 7, con sustratos duros, el cual presentó los
valores más altos de similitud (30 - 40 %); otro, integrado por las localidades 3, 4 y
5, con sustratos blandos, y los dos restantes fueron constituídos sólo por una
localidad que corresponden a la 1 y 8.
En otoiio, a un mayor nivel de similitud (33%) que en épocas anteriores se
evidenciaron tres grupos (Fig. 6c), compuestos por las localidades 2, 6 y 7; 1, 3 y 4;
5 y 8. Igual que sucedie en verano, la mayor similitud (46-56 %) la presentó el primer
grupo, que corresponde a localidades con sustratos duros .
Finalmente en invierno, con un nivel de 20%, también se hicieron evidentes
tres grupos (Fig. 6d), cambiando únicamente la agrupación de las localidades con
sustratos blandos. Como sucedió en épocas anteriores, las localidades 2, 6 y 7 fueron
las más similares (40 a 49 %), seguidas por el grupo de las localidades 3, 5, 8 y 4,
y el grupo formado por la localidad 1.
32
PRIMAVERA- 0.00
- 0.25
_ 0.50
- 0.75
l
~ - 1.00
OTOÑO
6 2 3 5 4 8 1 8 5 4 3 7 6 2
,~ mi -. . .
7 6 2 4 3 1
0.00
VERANO
INVIERNO
I
0.25
4 8 5 3 7 6 2
r
Dendrograma de similitud ficofloristka estacional entre las localidades de la laguna San Ignacio,B.C.S., a partir del Indice de Jaccard durante 1992-93.
L
r
0.00
0.25
0.50
0.75
1.00
0.00
0.25
0.50
0.75
8 5
Fig. 6.

_7. 3. ESTACIONALIDAD
A lo largo del año no se presentaron grandes cambios en la riqueza específica;
el número de especies varió entre 57 y 65, correspondiendo el mínimo a primavera y
el máximo a verano. Por división, la riqueza fue similar en las diferentes épocas,
encontrando el mayor número de Rhodophyta (39) y Phaeophyta (10) en verano, y el
de Chlórophyta (18) en invierno (Fig. 7). A nivel localidad, se observó un mayor
número de especies durante otoño e invierno (Fig. 8).
-70 ,
B 40iiw0 30
2 20
10
0PRIM UER OTO INU
c:;:.::i:jl RHOOOPHYTFI 0 PHKOPHYTh mCHLOROPHYTFI -TOTAL
Fig. 7. Riqueza específica estacional de las macroalgas pordivisión (1992-93).
Respecto a la temporalidad de las especies, 32 ocurrieron sólo en una época,
ll en dos, 25 en tres y 29 todo el año. De estas, 72 fueron anuales (estacionales y
no estacionales), 14 pseudoperennes y ll perennes. Las especies anuales
estacionales que ocurrieron en una época (Tabla 5) en su mayoría fueron especies que
se encontraron sólo en una localidad.
34

Entre las especies anuales estacionales que ocurrieron en dos épocas se
encuentran: Laurencia masonii, Polysiphonia simplex, Sphacelaria californica,
Cladophora sericea y Monostroma zostericola; entre las anuales no estacionales (que
ocurrieron en tres y cuatro épocas): Laurencia sinicola, Polysiphonia johnstonii, P.
mollis, Ectocarpus parvus, En teromorpha cla thra ta, E. compresa, Codium cunea tum,
Cladophora graminea, C. albida y Acetabularia calyculus; entre pseudoperennes:
Hypnea valentiae, Gracilariopsis lemaneiformis, Gracilaria pacifica, G. subsecunda ta,
Sp yridia filamentosa, Dasya baillouviana y Sargassum sinicola; y entre perennes:
Jania adhaerens, Corallina vancouveriensis, Neogoniolithon sp y Amphiroa spp. Con
excepción de las algas calcáreas, cuyos cambios esta’Tcionales fueron menos
perceptibles, el resto de las especies presentaron cambios notables en su distribución
y abundancia en las diferentes épocas.
mpRIn m(JERm INU - TOTAL
OTO
Fig. 8. Riqueza específica espacio-temporal de las macroalgas (1992-93).
35

TABLA 5. ESPECIES ANUALES ESTACIONALES QUE OCURREN EN UNA EPOCA(1992-93).
Estación
Primavera
Especie
Corallina sp’Hydrolithon decipiens ’Gra teloupia versicolor ’Acrosorium venulosum
* Ceramium cauda tum ’* Ceramium serpens ’Polysiphonia flaccidisimaPadina crispa ta ’
Verano
otoño
Invierno
Plocamium sp ’ ** Gracilarioph ylla gardnerii ’* Tiffaniella saccorhiza ’* Cryptopleura sp’* Herphosiphonia secunda ’
Laurencia hancockii’Laurencia sn yridaePach ydictyum coriaceum ’Sargassum agardhianumUlva dac t yliferaCodium sp
Corallina frondescens ’Lep tofauchea pacifica ’
+ Ceramium sp+ Cen troceras cla vula tum ’+ Herposiphonia s p ’
Rosenvingea in trinca ta ’Uva loba ta ’
Laurencia gardnerii ’Dictyota flabellata ’Gracilaria tex torii ’Sargassum muticum ’Pterocladia caloglossoides ’Uva rlqida
* Especies epífitas.’ Especies exclusivas a una localidad
36

_7. 4. ABUNDANCIA.
De 97 especies determinadas, sólo 66 presentaron una biomasa mayor a 0.1
g peso húmedo/m’ , por lo que el análisis respectivo corresponde a estas especies
(Tabla6). Las algas rojas fueron las más abundantes, seguidas por las pardas y
ve rdes (Fig. 9).
CHLOZOPHYTA (8.47
PHAEOPHYTA (13.1%)
Fig. 9. Contribución de las macroalgas por división a laabundancia total (1992-93).
Estacionalmente se detectó un patrón general de abundancia estacional: la
biomasa fué menor en primavera, se incrementó hasta alcanzar el máximo en verano
y disminuyó a partir de otoño. Las algas rojas siguieron este patrón, no asi las verdes
y pardas que presentaron sus valores máximos en invierno (Fig. 10).
Entre localidades, los valores mas altos de biomasa anual (247.6 g peso
húmedo/m2) los presentó Punta Choya (loc. 61, seguida por La Boca (1) y Punta Piedra
(21, con 156.3 y 155 g peso húmedo/m2 respectivamente, mientras que Los Médanos
37

(B), Cantil Cristal (5) y Campo Manuela (3), presentaron los mínimos, con biomasa de
0.2, 1.8 y 2.3 g peso húmedo/m2 respectivamente. El patrón de abundancia estacional
:fue diferente en cada localidad; por ejemplo, la mayor biomasa en La Boca ocurrió en
otoño, en Punta Piedra en invierno y en Punta Choya en verano (Fig. 1 1).
PRII1 UEí)
r:i:::.::l] RHOOOPHYTR 0 PHREOPHYTA
OTO INU
CHLOROPHYTR -TOTAL
Fig. 10. Abundancia estacional de las macroalgas por division(1992-93).
260:240: 220;200,; 180= 160* 140d 1200100" 805 60g 40; 20
0
LOCALIDAO
BpRIn BUER OTOm INU - TOTAL
Fig. Il. Abundancia espacio-temporal de las macroalgas(1992-93).
38

TABLA 9. ABUNDANCIA ESTACIONAL (g peso humedolm’) DE LAS ESPECIES DE
MACROALGAS DE LA LAGUNA SAN IGNACIO, 0. C. S. DURANTE UN CICLO ANUAL
(1992-93).
Uva lactuca
TESPECIE ‘R I M A V E R A V E R A N O 1 OTONO 1 I N V I E R N O
RHODOPHYTA
Acrosorium venulosumAnotrichium tenueCeramium spChondria dasyphyllaDasya baillouvianaGracilaria marcialanaGracilaria pacificaGracilaria spGraoilaria subsecundataGracilariopsis lemaneifonnGrifhthsia furcellatatiypnea valentiaeLaurencia lajollaLaurencia masoniiLaurencia pacificaLaurencia sinicolaLaurencia snyderiaeLaurencia spPolysiphonia flaccidissimaPolysiphonia johnstonii va!Polysiphonia mollisPolysiphonia pacificaPolysiphonia simplexPolysiphonia spPterosiphonia dendroideaSpyridia filamentosa
I
0.2400
3.730.100.08
18.760
1.081.80
07.370.033.570.150.10
0t
0.150.100.030.16
*
0.03t
20.33
PHAEOPHYTA
Colpomenia sinuosa 0Colpomenia tuberculata 1.22Ectocarpus parvus 0.08Wydrocla thrus cla thra tus 0Pachydictyon coriaceum 0Padina crispata 0.30Padina durvillaei 0.40Sargassum agardhianum 0Sargassum muticum 0Sargassum sinicola 1.60Sargassum sp 0
CHLOROPHYTA
4cetabulana calyculusChaetomorpha linumCladophora albidaCladophora gramineaCladophora microcladiodeCodium cuneatumCodium decorticatumEnteromorpha clathrataEnteromotpha compressaEnteromotpha flexuosaEnteromorpha intestinalisEnteromomha proliferaEnteromorpha ramulosaRhizoclonium ripariumUva califomicaUlva dactylifera
00
0.130.03
04.78
*
0.03*
0.03*
0.68*?
00?
00.65
05.060.160.52
83.290.6
0.9719.58
t
40.920.029.17
?
0.230.720.07
00.120.57
00
0.01*
00.070.130.120.420.04
77.360.75l l . 65.720.11
20.5800
0.730.16
00.48
00.03
3.55.05
04.31
*93.53 48.55
00.06
00.760.320.0410.40.050.512.091.840.150.25
00
0.040
0.050
0.10.21
0.90.660.071.253.57
0.010t*
0.620
9.130.35
012.33
0.4
00.240.13
000000
4.71.97
0.50.633.630.39
00
18.410
25.53.78
?
0.08*
0.410
0.1835.310.030.011.542.65
?
0.590
6.240.867.310.52
?
000
0.150.16
0.040.02
0.30.01
0.2;0.360.03
0
0.030.29
00.01
03.440.662.441.796.160.534.032.730.010.32
00.081 01 O.OS]
_ . .._??Especie presente pero con biomasa menor a 0.1 g peso numeaolmz.
39

La aportación por división de algas a la abundancia, también fue diferente en
cada localidad; por ejemplo, La Boca (1) y Punta Piedra (2) presentaron valores
,similares de biomasa, sin embargo, la biomasa de la primera fue sustentada
prácticamente sólo por algas rojas, mientras que en Punta Piedra, contribuyeron rojas,
pardas y en menor proporción verdes (Fig. 12).
260240
::g 2202 200$ 180': 160g 140å 120z LO@
u 80:: 60i 40m 20
012 3 4 5 6 7 8
L0CflL10Atl
m RHOOOPHYTF1 0 PMEOPHYTPI =CHLOROPHYTA - TOTRL
Fig. 12. Abundancia espacial de las macroalgas por división(1992-93).
Las especies más abundantes que representaron el 85 % de la abundancia total
fueron: Gracilaria pacifica, Sp yridia filamentosa, Hypnea valentiae, Gracilariopsis
lemaneiformis, Padina durvillae, Sargassum muticum, Sargassum sinícola, Gracílaría
subsecundata y Codíum cuneatum. Las tres primeras contribuyeron con 65 %,
mientras que las otras seis con el 20 % (Fig. 13). La aportación de estas a la
abundancia total de cada localidad se presenta en la Fig. 14.
4 0

Fig. 13. Abundancia relativa de los principales componentesficológicos (1992-93).
260r\ 240=$2204 200: 18013c 160u 140å120$100$ 80c 600 40i 20
0
-BIORASA TOTAL --.s.--. BIOMASA DE PCF
Fig. 14. Contribución de los principales componentesficológicos (PCF) a la abundancia total de cadalocalidad (1992-93).
41

La abundancia de las principales especies de algas fue diferente entre
localidades (Fig. 15); por ejemplo, Gracilaria pacifica se concentró básicamente en La
,Boca (78 %) y Punta Piedra (21 %); Spyridia filamentosa en Punta Choya (64 %) e Isla
Garza (20 %); Hypnea valentiae en Punta Choya (74 %) y La Base (10 %) y
Sargassum muticum sólo en Punta Piedra. De igual manera, la contribución de dichas
especies en cada localidad fue diferente, como el caso de Gracilaria pacifica q u e
representó prácticamente la totalidad de la biomasa encontrada en La Boca, mientras
que para el resto de las localidades, la biomasa fué sustentada por varias de las
especies.
Fig. 15. Abundancia de los principales componentes ficológicos en cadalocalidad (1992-93).
42

_7. 5. PRINCIPALES COMPONENTES FICOLOGICOS: ESTACIONALIDAD Y
DISTRIBUCION VERTICAL.
Para este análisis se consideró a Gracilaria pacifica, Spyridia filamentosa,
Hypnea valentiae, Sargassum sinicola y Codium cuneatum. Las tres primeras se
seleccionaron por ser las especies que contribuyeron con mayor porcentaje a la
abundáncia total (69 %), mientras que las dos últimas, para tener representadas a las
divisiones Phaeophyta y Chlorophyta.
7. 5. 1 Gracilaria pacifica
Su patrón de abundancia estacional coincide con el descrito con anterioridad
para algas rojas, con valores mínimos en invierno (9.2 g peso húmedo/m2) y máximos
en verano (41.5 g peso húmedo/m2) (Fig. 16).
Fig. 16. Patrón de abundancia estacional deGradada pacifica (1992-93).
43

La mayor concentración de Gracilaria pacifica se presentó entre los 50 y 1 OO
m a partir de la línea de costa. Durante primavera, se distribuyó básicamente hasta
,los 70 m; en verano, se extendió hasta los 100 m y su mayor concentración se
presentó entre los 90 y 1 OO m; otoño fué la época de mas amplia distribución,
encontrándose algas hasta los 170 m; en invierno su distribución disminuyó, y sólo
se presentó entre los 50-I OO m (Fig. 17).1
10 20 30 70 80 w loa 110 120 m 140 150 160 170 180 190 zu9
DISTANCIA 13 PFIRTIR OE L A C O S T A (m)
Fig. 17. Distribución vertical estacional deGracilaria pacifica (1992-93).
7. 5. 2 Sp yridia filamentosa
Su estacionalidad también concuerda con el patrón de abundancia de algas
rojas: la biomasa mínima promedio se observó en invierno (1.8 g peso húmedo/m2) y
la máxima en verano (30 g peso húmedo/m2) (Fig. 18).
Spyridia filamentosa prácticamente se distribuyó a todo lo largo del gradiente
vertical; en primavera, la mayor concentración se manifestó entre los 130-I 90 m; en
verano, amplió su distribución y fue la época en donde ocurrió casi a todo lo largo del
4 4

gradiente vertical, la mayor concentración fue entre los 80 y 170 m; en otoño,
presentó una distribución discontínua o irregular, con su concentración más alta en los
,primeros 70 m y en invierno, disminuyó tanto su distribución como su concentración,
restringiéndose sólo hasta los primeros 80 m de la orilla costera (Fig. 19).
0PRIM UER OTO INU
Fig.18. Patrón de abundancia estacional deSpyridia filamentosa (1992-93).
010 m 40 M 60 70 60 m im110uon3womuormi6uis2m
DISTANCIA 6 PFIRTIR DE LA COSTA (n)
Fig. 19.Distribución vertical estacional deSp yridia filamentosa (1992-93).
4 5

7. 5. 3 Hypnea valentiae
Su patrón estacional fue muy evidente: alcanzando su pico de abundancia en
verano (10.2 g peso húmedo/m*) hasta prácticamente desaparecer en invierno
(Fig.20).
12]
0’ IPRiF UER OTO INU
Fig. 20. Patrón de abundancia estacional deHypnea valentiae (1992-93).
Aún con sus cambios estacionales, de manera general, es una especie que
presentó una amplia distribución vertical. En primavera, su distribución fue muy
homogénea y ocurrió a partir de los 40 m de la línea costera hasta los 180 m. En
verano, también presentó una distribución regular pero fue más amplia que en
primavera y comprendió hasta los 200 m, su mayor concentración ocurrió entre los
120 y 190 m. En otoño, se encontró hasta los 200 m, pero su distribución fue más
irregular o discontínua. En invierno, su presencia fue muy escasa (Fig. 21).
46

Fig. 21. Distribución vertical estacional deHypnea valentiae (1992-93).
7. 5. 4 Sargassum sinicola
La abundancia máxima se presentó en verano (6.1 g peso húmedo/m’),
disminuyó en otoño y se mantuvo similar en invierno y primavera (Fig. 22).
En general, la mayor concentración de Sargassum sinkola ocurrió entre los
120-200 m a partir de la línea de costa, presentándo variaciones estacionales en su
distribución. Con excepción de primavera, también se encontró de los 20 hasta los
40 m. Verano fué la época de más amplia distribución, mientras que en otoño,
prácticamente desapareció, ocurriendo sólo a los 20 y 200 m (Fig. 23).
47




_
L epilíticas se reduce hasta sus niveles más bajos en otoño y comienza a incrementarse
a partir de invierno (Fig. 27).
w 50Eiw . .; 40 -
0; 30 -
I,_._._.,._,......,._...*....~.~~-,,~.'~.."~
.....,....*.........~...._.%. '.... “.‘.yl
20 JPRIil UER OTO INU
- EPILITICAS --.w.-- EPIFITAS
Fig. 26. Estacionalidad de las comunidades de algas epiiíticasy epífitas (1992-93).
60-
50-
40-
30-
zo-Y........ .10 - ____._...~J~ . ...' ""--".........._*
_...O- Y......__................_..~J~
PRItl UER OTO INU
- EPILITICFIS --++EPIFITAS
27. Abundancia estacionalepífitas (1992-93).
51
de macroalgas epilíticas y

I
7. 7. FACTORES AMBIENTALES
;7. 7. 1. TEMPERATURA
La temperatura presentó un patrón estacional muy evidente: la temperatura
promedio más baja (18 “CI se registró en invierno, aumentó en primavera y alcanzó
su máximo (28 OC) en verano (Fig. 28). Estadísticamente las diferencias fueron
significativas (ANDEVA; p < 0.05).
.I
28
A 260
is w; 22
ã 243c
aii
5t- 20
18 -
16 ’PRIR UER OTO iNU
Fig. 28. Variación estacional de la temperatura del agua en lalaguna San Ignacio, B. C. S. (1992-93).
Entre localidades, no fue posible reconocer un patrón general a lo largo de la
laguna debido a que las fluctuaciones fueron diferentes en cada época del año.
Estadísticamente las diferencias no fueron significativas (ANDEVA; p > 0.05), sin
embargo, se observaron algunas tendencias para cada época: en primavera y verano,
la temperatura fue más baja cerca de la boca de la laguna y se incrementó hacia el
52

_ DONATIVOinterior de la misma; en otoño, por el contrario, las temperaturas más altas se
registraron cerca de la boca y disminuyó hacia el interior de la misma, mientras que
,en invierno, !a temperatura fue similar en las distintas localidades (Fig. 29).
.1 2 3 4 5 6 7 8
LOCALIDALl
-c- PR11 - UER - O T O- INU - PROllEOIO ANUAL
Fig. 29. Temperatura del agua en diferentes localidades de lalaguna San Ignacio, B. C. S.
7. 7. 2. SALINIDAD
Igual que sucedió con la temperatura, las variaciones estacionales de salinidad
fueron notables; la mínima (32 O/oo) se registró en invierno y la máxima (37 O/oo) en
otoño (Fig. 30). Estadísticamente las diferencias fueron significativas (ANDEVA;
p-CO.05).
Entre localidades no se detectó un patrón evidente a lo largo de la laguna para
todo el período de estudio debido a las diferencias estacionales que se presentaron,
sin embargo, se observó una tendencia a incrementar desde la boca al interior de la
laguna (Fig. 31). Estadísticamente las diferencias no fueron significativas (ANDEVA;
p > 0.5).
53
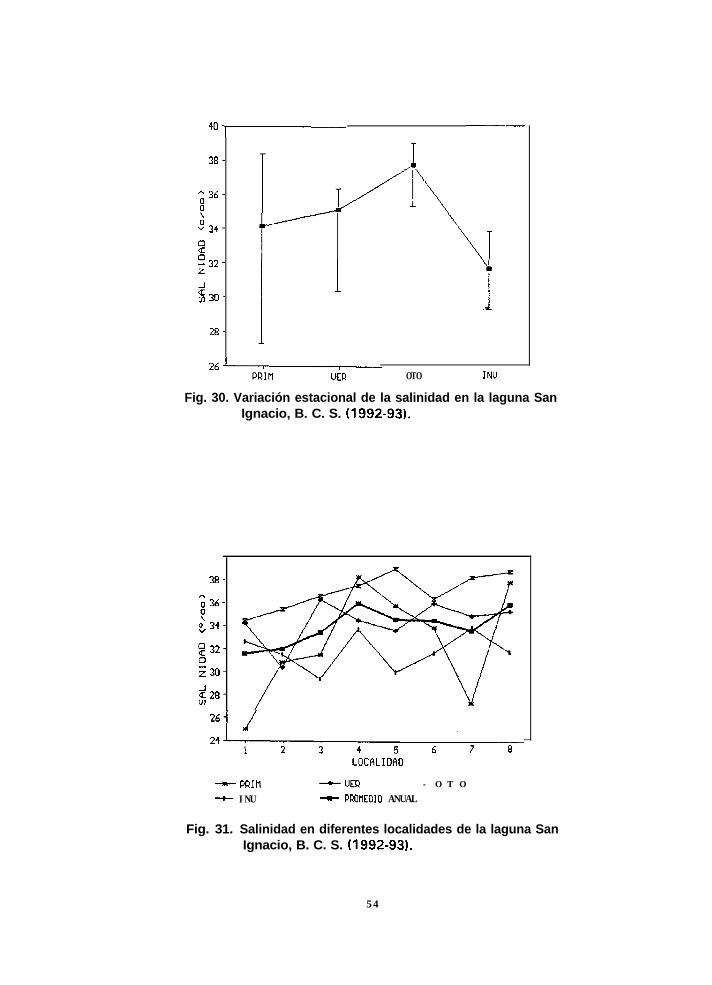
4or------ --
38 -
;; 36 -
Yt 34-
Q; 32 -
2cn 30 -
28 -
OTO INU
Fig. 30. Variación estacional de la salinidad en la laguna SanIgnacio, B. C. S. (1992-93).
38 -
; 36 -?$ 34 -
z 32 -clz 30 _
$ 28-
“1”24
1 2 3 6 7 8
- PRIM - UER - O T O- INU - PROKDIO ANUAL
Fig. 31. Salinidad en diferentes localidades de la laguna SanIgnacio, B. C. S. (1992-93).
54

_8. DISCUSION
L Cuando se habla de la estructura de una comunidad vegetal se puede hacer
referencia a la estructura física o biológica: la primera, es en esencia lo que se observa
en ella, es decir, sus formas de crecimiento y estratificación o patrones de distribución
espaci.al de las mismas, mientras que la biológica, abarca composición y abundancia
de espécies, cambios temporales y relaciones entre sus componentes (Krebs 1985).
8. 1. FLORA FICOLOGICA.
Las comunidades difieren en cuanto al número de especies que incluyen y una
de las áreas de investigación de mayor actividad en ecología, actualmente es el
estudio de la riqueza o diversidad de especies (Krebs 1985). En el presente trabajo,
para algunos casos no fue posible determinar a algunas especies debido a que se
encontraron en diferentes estados de desarrollo (plántulas, juveniles o senescentes)
y los caracteres necesarios para reconocerlas fueron insuficientes. En otros casos, las
algas presentaron caracteres morfológicos que no coincidían con las especies
descritas en las claves disponibles, por lo que sólo se determinó a nivel genérico.
De las 97 especies determinadas, 90 representan primeros registros en la
laguna en virtud de que siete ya habían sido citados con anterioridad (Dawson 1948,
1961 a, 1962, Aguilar-Rosas y Aguilar-Rosas 1993a). Si se toman en cuenta los
registros previos y los actuales, la flora ficológica potencial de la laguna está formada
por 103 especies. Los seis taxa registrados en la literatura que no se encontraron,
probablemente fueron especies escasas o inconspicuas, que quedaron fuera del área
de muestreo, o especies que por su ciclo de vida no se manifestaron en las épocas de
colecta o lo hicieron de una manera muy efímera. También, puede ser que las
condiciones ambientales, prevalecientes durante el periódo de estudio, fueron
desfavorables para su desarrollo.
55

_
‘_ Entre las especies, se encontraron representantes que por su distribución
geográfica han sido caracterizadas como de afinidad temp‘lada (Chondrachantus
: canalicula tus, Lep to fauchea pacifica, Ec tocarpus parvus, Sargassum agardhianum,
Cladophora microcladiodes y Kornmannia leptoderma entre otras) y especies
caracterizadas como de afinidad tropical (Hypnea valentiae, Spyridiafilamentosa, Jania
adhaerens, Gracilaria subsecunda ta, Hydroclathrus clathratus, Acetabularia calyculos,
etc.). Ásimismo, se encontraron especies de amplia distribución latitudinal, es decir,
especies que se desarrollan tanto en ambientes templados como tropicales.
Considerando la afinidad de los taxa, puede decirse que la ficoflora de la laguna
es más característica de ambientes templados que tropicales puesto que se observó
mayor proporción de especies de afinidad templada en comparación con las de
afinidad tropical. Esto puede explicarse porque la laguna se localiza en una región
geográfica con características más templadas que cálidas. Además, se encuentra en
áreas de surgencia (Roden y Groves 1959 citado por Aguilar-Rosas y Aguilar-Rosas
1993b, Espinoza-Avalos 19931, con aguas frías y altas concentraciones de nutrientes,
que favorecen la ocurrencia de algas de afinidad templada.
El gran porcentaje de especies de amplia distribución detectado en la laguna se
atribuye a las condiciones ambientales propias de la región Templado Cálida del
Hemisferio Norte, región biogeográfica (Lüning 1990) que incluye el área de la laguna,
y a las condiciones determinadas por la zona de transición, característica de sus aguas
circundantes. A lo largo de la costa pacífico de la península de Baja California, de
norte a sur fluyen aguas frías y de baja salinidad (< 34.6 ‘/OO) por la corriente de
California y de sur a norte aguas más cálidas y de salinidad intermedia (entre 34.7 -
34.9 “/OO) por la corriente Nor-ecuatorial (Fernández et al. 1993). La convergencia de
estas corrientes originan una zona de transición que ha sido denominada
Templada-Tropical (Dawson 1960a, Fernández et al. 1993, Zertuche 1993a). Dichas
características, aunado a la presencia de bahías protegidas, con aguas cálidas,
provocan la existencia de una mezcla de biota templada y tropical a lo largo del litoral
56

occidental de la península (Dawson 1960a, Bakus 1989 en Aguilar-Rosas y Aguilar-
Rosas 1993b).
Durante 1992/93, periodo en que se realizó este estudio, en costas del Pacifico
se registraron condiciones de El Niño o ENSO (El Niño/Southern Oscillation)
(Anónjmo 1993) provocando, entre otros efectos, un aumento en la temperatura
superficial del agua. Aún cuando es díficil precisar los efectos de dicho fenómeno
sobre comunidades de macroalgas (Tegner y Dayton 1987) debido a la gran
variabilidad temporal intrínseca que éstas experimentan, se conoce que puede afectar,
directa o indirectamente, y de manera diferencial, a las pobla&nes de algas marinas.
Murray y Horn (1989) no encontraron cambios drásticos en la distribución y
abundancia de macrofitas en la costa central de California; sin embargo, observaron
que la cobertura de algas calcáreas costrosas se incrementó más de lo normal durante
el periodo crítico de El Niño de 1982-83 (invierno 19831, al mismo tiempo que
disminuyó la cobertura de algas rojas no calcáreas. Por otra parte, Tegner y Dayton
(1987) encontraron que El Niño en cuestión provocó una reducción considerable en
la cobertura y crecimiento de Macrocystis pyrífera.
Las condiciones de El Niño 92/93 probablemente favorecieron la ocurrencia de
algas tropicales y afectaron a algunas algas de afinidad templada, como pudo ser el
caso de Scínaía johnstoniae Setchell y Polysiphonia jonstonii var conccina
(Hollenberg) Hollenberg, especies registradas con anterioridad (Dawson 1948) que no
se encontraron en este trabajo. Sin embargo, el efecto de este evento no puede ser
evaluado porque no existe información previa sobre la composición ficoflorística en
condiciones de No Niño.
En cuanto a riqueza, igual que en otros lugares de la región de la península de
Baja California (Huerta-Múzquiz y Mendoza-González 1985, Sánchez-Rodríguez et al.
1989, Aguilar-Rosas et al. 1990, Mateo-Cid eta/. 1993) y en otras partes del mundo
(Bolton y Stegenga 1987, Mathieson 1989, Kendrick et al. 1990, y Villal$rd-Bohnsack
57

‘. y Harlin í992, entre otros), el mayor número de especies correspondió a las algas
rojas. Lo anterior puede comprenderse porque éste grupo constituye el más diverso
,de macroalgas (Woelkerling 19901, cuyas formas de vida y estrategias reproductivas
les permiten persistir y manifestarse durante todas las épocas del año (Sears y Wilce
1975, Abbott y Hollenberg 1976, Darley 1987, Mathieson 1989). Las algas rojas en
su mayoría presentan reproducción asexual, que tiene la ventaja de requerir menor
gasto bnergético para la formación de esporas, las cuales son diseminadas en forma
rápida y permiten asi una dispersión más eficiente de las especies (Mateo-Cid et al.
1993). Las algas pardas generalmente presentan reproducción sexual, con mayor
gasto energético, mientras que las verdes, en su mayoría preséntan estado vegetativo,
reproduciéndose a través de propágulos, estolones y rizoides (Fritsch 1935 en Mateo-
Cid et al. 1993).
Respecto a las algas verdes y pardas, las primeras fueron más numerosas que
las segundas, con una diferencia de ocho especies que representa el 21%. En la
península de Baja California, la mayor riqueza de algas pardas se presenta en la costa
occidental y se incrementa de sur a norte debido pricipalmente a factores como el
gradiente de temperatura, corrientes y tipo de sustrato (Aguilar-Rosas y Aguilar-Rosas
1993b). Esto, coincide con el patrón de distribución a escala latitudinal, en donde se
establece de manera general que las algas pardas prefieren aguas más frías (lugares
templados y polares) y son menos conspícuas en áreas tropicales (Darley 1987,
Whittick et al. 1989). Por la localización geográfica de la laguna, pudiera esperarse
mayor riqueza de algas pardas que de algas verdes, como ocurre en islas Todos
Santos (Aguilar-Rosas et al. 1990) y Bahía Tortugas (Mendoza-González y Mateo-Cid
1985), localizadas en la parte norte y centro de la península respectivamente, sin
embargo, ocurre lo contrario. Lo anterior, puede reflejar el efecto del calentamiento
del agua provocada por el Niño 92/93, aunque pudo ser más determinante la
disponibilidad de sustrato. Las algas pardas se desarrollan preferentemente en
sustratos rocosos (Espinoza-Avalos 1993) y en la laguna este fue escaso. Por otra
parte, se conoce que las algas verdes presentan una amplia distribución porque se
58

desarrollan en diferentes tipos de sustrato y se adaptan bien a hábitats extremos
(Darley 1987, Whittick et al. 1989).
8. 2. DISTRIBUCION
Antes de iniciar el análisis de distribución de macroalgas, conviene señalar que
los pastos, particularmente Zostera marina (Linnaeus) se distribuyeron ampliamente
a lo largo de la laguna. En Punta Piedra, Punta Choya, e Isla Garza su distribución
vertical comenzaba aproximadamente a partir de los 1 OO m desde la orilla costera
hasta la zona de canales. En áreas protegidas, con menor circulación de agua, alejadas
de los canales, se encontró Philloxpadix sp, una especie de pasto filamentoso. Este,
se restringió a las localidades con sustrato fango-arenoso ubicadas en la parte central
(La Base y una parte de Punta Choya) y en la cabecera de la laguna (Cantil Cristal y
Los Medanos). De esta manera, y entre otras causas, puede decirse que la distribución
de macroalgas en el interior de la laguna estuvo Iímitada por el efecto de
competencia de los pastos marinos, que enc&kntraron en los bajos y canales (fondo
arenoso) las condiciones necesarias para constituir la vegetación dominante. La
competencia es uno de principales factores biológicos que afectan la distribución de
macroalgas (Round 1984 en Torres 1991, Dawes í986, Carpenter 1990, Stewart
1989). Además, las especies dominantes tienen un elevado indice de éxito ecológico
y determinan en gran parte las condiciones de las especies con las que coexisten
(Krebs 1985). En otros sistemas, con características ambientales semejantes a las de
la laguna, de poca profundidad y con sustrato arenoso y fango-arenoso, se ha
señalado también a los pastos marinos como vegetación dominante (Trono y Saraya
1987, Kendrick et al. 1990).
de la
Las condiciones ambientales en un hábitat determinado, regulan la estructura
comunidad y son resultado de los efectos combinados y sinergéticos de
diferentes factores como temperatura, luz, salinidad, tipo de sustrato, disponibilidad
de nutrientes, y condiciones de disturbio tanto físicas (intensidad de corrientes y
59

oleaje, mareas, movimiento y acumulación de arena o sedimentación, vientos, y
cambios climáticos anómalos) como biológicas (depredación, competencia y efectos
,antropogénicos) (Dawes 1986, Trono y Saraya 1987, Trono 1988, Kendrick et al.
1990). Murray y Littler (1989) indican que la distribución, composición, y abundancia
de macroalgas serán determinadas por las características propias del lugar en que se
encuentran.1
A escala latitudinal o geográfica, los factores más importantes que determinan
la distribución de algas marinas son la la temperatura y la luz (Abbott y North 1971,
Santelices 1977, Bolton y Stegenga 1987, Wolfe y Harlin 1988á, Whittick eta/. 1989,
Lüning 19901, mientras que a un nivel local, son factores como el tipo de sustrato,
salinidad y régimen de mareas (Dawes 1986, Kendrick et al. 1990). Además, se
presupone que los factores físicos (oleaje o desecación entre otros) son más
importantes en las zonas someras, en tanto los biológicos (competencia y
depredación) lo son en zonas más profundas (Torres 1991, Stewart 1989).
En la Laguna San Ignacio, como sucede en otros lugares (Aguilar-Rosas 1982,
Trono y Saraya 1987, Mateo-Cid et al. 19931, el tipo de sustrato fue el factor principal
que determinó la distribución de macroalgas. Las localidades con sustratos duros
(Punta Choya, Isla Garza y Punta Piedra), que incluyen rocas, cantos rodados,
guijarros, conchas y conchilla, tuvieron mayor riqueza específica que las que
presentaron sustratos blandos (La Boca, Campo Manuela, La Base, Cantil Cristal y Los
Médanos), constituídos por arena y mezcla de fango arena (fango-arenoso). Cabe
señalar que la mayor parte de algas bentónicas se adhieren al sustrato por medio de
una célula basal, un disco de fijación, hápteras o con todo el talo, por lo que su
presencia en la mayoría de los casos está restringida a sustratos duros (Santelices
1977, Mateo-Cid et al. 1993).
Aparte del sustrato, el disturbio físico y biológico altera la diversidad algal en
las comunidades bentónicas (Villalard-Bohnsack y Harlin 1992). ‘Entre estas
60











En invierno se presentaron condiciones óptimas para el desarrollo de algas
verdes, lo cual se hizo evidente porque lograron su biomasa máxima. Esto se
:relaciona con las bajas temperaturas, haciéndose manifiesto la preferencia que tienen
‘las especies de algas verdes de la laguna por el agua fria, que se corrobora al
constatar que la mayoria de las especies determinadas han sido caracterizadas como
de afinidad templada. Además, es probable que los nutrientes tengan un efecto
importante en su producción puesto que se ha encontrado que las algas verdes,
particularmente Uva spp y Enteromorpha spp se desarrollan mejor en ambientes con
altas concentraciones, debido a que presentan una alta cinética de absorción y rápido
crecimiento en respuesta al insumo de compuestos nitrógenádos (Harlin 1995). A
pesar de que no se midió la concentración de amonio (NH,), nitratos (NO,), nitritos
(NO,) y ortofosfatos (PO,), nutrientes inorgánicos más importantes en el crecimiento
de las algas marinas (barley 19871, se conoce que en invierno hay un enriquecimiento
debido a la influencia de aguas de la corriente de California (Fernández et al. 1993).
También, puede ser que el deslave de los terrenos circundantes vertidos a la laguna
provocados por las lluvias características de invierno, contribuyeron a incrementar la
concentración de nutrientes. Soto-Balderas y Alvarez-Borrego (1993) discuten sobre
la dinámica general (remineralización, estacionalidad) de nutrientes en algunas lagunas
costeras mexicanas y mencionan, entre otras cosas, que los altos niveles de nitratos
estan asociados al aporte de agua dulce. Si bien en la laguna no hay influencia de
agua dulce por rios o arroyos, el aporte provocado por las lluvias pudo ser
considerable. En un sistema estuarino, Coutinho (1987, en Coutinho y Zingmark 1993)
encontró que las plantas de Uva curvata crecen, se reproducen y alcanzan su máxima
producción durante invierno-primavera, cuando las temperaturas fueron bajas y la
concentración de nutrientes alta, esta última asociada con las lluvias de primavera. De
esta manera, los nutrientes pueden determinar la abundancia de algas, sin embargo,
su efecto dependerá de la cantidad y calidad de los mismos, asi como de los
requerimientos particulares de cada especie (Harlin 1995).
71

Respecto a algas pardas, estas presentan un patrón con mínimos valores de
abundancia en primavera y máximos en invierno, que refleja la afinidad de sus
,especies por las aguas frías, aunque también pudiera ser resultado de sus
‘adaptaciones cuando la luz es escasa.
cuando la temperatura fue menor y los
En la laguna, la mayor abundancia coincidió
días fueron más cortos.
Durante el periódo de estudio, algunas localidades presentaron gran abundancia
en comparación con las demás, que las señala como lugares con condiciones
ambientales más favorables para el desarrollo de las algas, muchas de las cuales ya
se han discutido en el capítulo de distribución. Asimismo, lá abundancia de cada
localidad tiene que ver más con la composición que con la riqueza específica. Lo
anterior se establece porque, si bien se observó mayor abundancia en Punta Choya
y Punta Piedra, localidades con mayor número de especies, también se detectó gran
abundancia en La Boca, una de las localidades con menor riqueza. Como la
comunidad ficológica de cada localidad fue diferente, cada una de ellas presentó un
patrón particular de abundancia estacional, determinado por las características
fenológicas de sus especies, y probablemente por las interacciones entre ellas
(competencia, efectos alelopáticos ) y otros organismos (depredación).
Las estrategias adaptativas de las especies a diversos habitats, permiten
comprender las diferencias que se presentaron en la contribución por grupos de algas
a la abundancia de cada localidad. Las algas rojas fueron las de mayor abundancia en
todas las localidades por su capacidad de desarrollarse tanto en sustratos duros como
en blandos, mientras que la mayor abundancia de algas pardas se encontró en Punta
Piedra, con sustrato rocoso y ambientes más frios debido a la influencia de aguas
oceánicas, condiciones más favorables para su desarrollo.
Los componentes representativos de la comunidad de macroalgas en la laguna
fuero n : Gracilaria pacifica, Spyridia filamentosa e Hypnea valentiae, ya que se
distribuyeron ampliamente ‘y fueron los más abundantes. También, G>acilariopsis
72

‘.. lemaneiformis, Padina durvillae, Sargassum muticum, Sargassum sinicola, Gracilaria
subsecundata, y Codium cuneatum, fueron abundantes, aunque algunas sólo en cierta
:época y otras en cierta localidad.
En forma general, los principales componentes ficológicos representaron un
porcentaje importante de la biomasa total y su contribución fue diferente en cada
localidad. Por ejemplo, Spyridia filamentosa e Hypnea valentiae, sustentaron la mayor
biomasa en Punta Choya; G. pacifica, Padina durvillae, y Sargassum muticum en
Punta Piedra, y G. pacifica sustenta casi la totalidad de la biomasa de La Boca. Estas
diferencias se atribuyen al éxito competitivo de las especies representativas de cada
localidad para adaptarse y explotar las condiciones ambientales favorables y lograr su
dominancia. Sears y Wilce (1975) mencionan que las especies que habitan lugares
donde el movimiento de arena es considerable colonizan estos hábitats, en donde
muchas algas no pueden competir y se adaptan de tal manera que su talo permanece
viable aún cuando la arena lo cubre por periodos frecuentes y prolongados. Dicen
además, que la capacidad para resistir a la abrasión y al cubrimiento por arena son
probablemente los factores más importantes que explican la baja diversidad y la
dominancia de algas adaptadas a estas condiciones.
8. 5. PRINCIPALES COMPONENTES FICOLOGICOS: ESTACIONALIDAD Y
DISTRIBUCION VERTICAL.
Como se ha mencionado, la estacionalidad de las algas es resultado del efecto
combinado de los factores ambientales tanto físicos (luz, temperatura, nutrientes,
sustrato) como biológicos (competencia, depredación), y de la respuesta intrinseca
(ciclos circanuales, periodicidad estacional) de las especies de macroalgas a tales
factores. Así, cada especie exhibirá un patrón de abundancia estacional de acuerdo
a sus características fenológicas y requerimientos ambientales necesarios para su
desarrollo. La abundancia máxima refleja la época óptima de crecimiento para cada una
de ellas y la transición a su época de senescencia.
73

En cuanto a distribución vertical, en la Laguna San Ignacio, B. C. S., no se
observó un patrón de zonación como el descrito para zonas templadas (Chapman
: 1979, Santelices 1980, Lüning 1990).
8. 5. 1 . Gracilaria pacifica.
Se observó una relación directa entre el ciclo de producción de Gracílaria
pacifica con la temperatura: cuando el agua fue más cálida hubo mayor biomasa,
mientras que cuando fue más fría la biomasa fue menor. Otros investigadores
coínciden en señalara la temperatura como uno de los principales factores que regulan
la estacionalidad de Gracilaria spp (Mayer 1981, Molloy y Bolton 1995). Mayer
(19811, en una bahía de Argentina, también detectó una relación directa de la
temperatura con la cobertura y biomasa de Gracilaria sp.
En verano, época de máxima abundancia, G. pacifica se distribuyó en los
primeros 1 OO m a partir de la costa, mientras que en otoño ocurrió de los 90 m hasta
los 140 m. En la primera época, se observaron talos en buen estado, de textura firme
y sin epífitas, mientras que en otoño la mayoría fueron senescentes, ya que eran
quebradizos y estaban epifitados. Con base en lo anterior, puede decirse que G.
pacífica se desarrolló más rápido cerca de la costa, en donde la profundidad fue
menor, y más lento hacia el interior de la laguna en donde hubo mayor profundidad.
Molloy y Bolton (1995) encontraron resultados similares en el sistema lagunar de
Lüdertiz, Namibia (Sudáfrica) y explican que en aguas poco profundas Gracilaria se
desarrolla y alcanza su talla máxima más rápido que las de aguas profundas, porque
tienen una tasa de crecimiento más alta debido a que el agua es más cálida y hay
mayor disponibilidad de luz. Asimismo, mencionan que los talos de la ‘orilla costera,
son más suceptibles al disturbio causado por el movimiento del agua a través de las
mareas y vientos. En términos generales, durante el período de estudio, la mayor
concentración de G. pacifica ocurrió entre los 50 y 100 m, a una profundidad
aproximada de 3 m. En los primeros 50 m la escasa presencia de G;aci/aria, con
74

excepción de verano cuando los talos fueron mas sanos y por lo tanto más
resistentes, puede ser consecuencia del efecto de la desecación, ya que el área
,quedaba expuesta durante las bajamares. Después de los 140 m, se observó que el
movimiento del agua por corrientes era más intenso debido a la cercania de uno de los
canales de entrada, lo que seguramente impidió el establecimiento y desarrollo de esta
especie.
8. 5. 2. Spyridia filamentosa
Presentó un ciclo de producción anual, con su bioma5a máxima en verano y
mínima en invierno. Su patrón estacional, también se relacionó directamente con la
temperatura. Como lo indican los valores de biomasa, en verano, cuando el agua fue
más cálida se presentaron las mejores condiciones para su desarrollo, mientras que
en invierno, cuando el agua fue más fría, las condiciones le fueron desfavorables ya
que prácticamente desapareció. Sears y Wilce (19751, en cabo Cod, U. S. A.,
observarón también un mejor desarrollo de S. fílamentosa en verano, señalando que
es un reflejo de su afinidad tropical.
S. filamentosa es una especie con adaptaciones morfológicas para establecerse
y desarrollarse en una gran variedad de sustratos como rocas, guijarros, conchas,
arena, otras algas, raíz de pasto y raíz de mangle (Sears y Wilce 1975, Mateo-Cid et
al. 1993). Puesto que dichos sustratos, con excepción del manglar, fueron
característicos en las localidades donde se encontró, se explica la amplia distribución
de S. filamentosa a lo largo del gradiente vertical. Espinoza-Avalos (1993) menciona
que esta especie domina en lagunas costeras por su capacidad para adaptarse a vivir
en fondos arenosos y en condiciones de disturbio a través de su’ reproducción
vegetativa. En invierno, sólo se distribuyó en los primeros 90 m a partir de la costa,
en donde la temperatura del agua, probablemente, fue más elevada debido a un
mayor calentamiento del agua por efecto de la poca profundidad. En primavera,
además de incrementar su biomasa, se nota una mayor distribución bertical de S.
75

_filamen tosa, lo que permite suponer que es en esta época cuando inicia su
reclutamiento.
8. 5. 3. Hypnea valentiae
Presentó un ciclo de producción anual, relacionado principalmente con las
fluctua’ciones estacionales de temperatura. H. valentiae alcanzó su biomasa máxima
en verano, cuando la temperatura fue más elevada y su biomasa fue casi nula en
invierno, cuando la temperatura fue más baja, reflejándose así su afinidad tropical
(Dawson 1961 b, Abbott y Hollenberg 1976). Por el contrario;Schenkaman (19891,
observó una relación inversa entre la biomasa de H. muscíformis con la temperatura
(mayor biomasa cuando la temperatura fue más fria y menor cuando fue más cálida)
en costas de Ubatuba, al norte de Sao Pablo, Brasil. Seguramente, estas diferencias
se deben a los requerimientos particulares de cada especie, aunque el efecto de otros
factores pudo también influir; Además de la temperatura del agua, las mareas,
disponibilidad de sustrato, herbivoria (Schenkaman 19891, temperatura ambiental,
húmedad relativa, velocidad del viento (Murthy et al. 19891, salinidad (Tokoya y de
Oliveira 1992, Velkanni y Rengasamy 1992b), intensidad luminosa (Velkanni y
Rengasamy 1992a) y movimiento del agua (Berchez et al. 1993) se han relacionado
con la producción estacional de Hypnea spp.
La amplia distribución vertical de H. valentiae en la laguna se atribuyó a la
disponibilidad de sustrato y a sus adaptaciones morfológicas, que le permitieron
establecerse y desarrollarse sobre conchas, guijarros, arena, y otras algas, sustratos
característicos de las localidades donde se encontró. Asimismo, sus características
reproductivas le confieren un mayor éxito competitivo en la colonizaciórí de espacios
libres: generalmente, las especies de Hypnea se reproducen asexualmente a través de
la producción de esporas y vegetativamente por fragmentos del talo dispersos en la
columna de agua (Berchez et al. 1993).
76

8. 5. 4. Sargassum sinicola
: Presentó un ciclo de producción anual que coíncide con sus cambios
fenológicos, loi cuales a su vez se relacionaron con las fluctuaciones de temperatura.
En localidades del golfo de California también se ha descrito un patrón de biomasa
anual o monomodal para S. sinicola (McCourt 1983, Núñez-López 19931, sin embargo,
presentan una estacionalidad diferente a la que se observó en la laguna San Ignacio,
6. C. S,. mientras que en el golfo de California la abundancia máxima de Sargassum
ocurre a finales de primavera, en la laguna (del lado del pacífico) ocurre en verano.
Tales evidencias, concuerdan con lo establecido por McCourt T”r 983) quien señala que
el pico de abundancia para este género en aguas templadas ocurre en los meses
cálidos, mientras que en zonas tropicales y subtropicales ocurre en los meses frios.
Cabe mencionar, que en la laguna S. sinkola no formó mantos como en bahía
Concepción, B. C. S. (Núñez-López 19931, por lo que su biomasa por metro cuadrado
fue significativamente menor; mientras que en la laguna se registró una biomasa de
6 g peso húmedo/m2, en la bahía fue de 4 kg peso húmedo/m2. Además, a diferencia
de la bahía, cuyos organismos fueron modulares, en la laguna se observaron
organismos individuales, adheridos a conchas o cantos rodados.
S. sinkola siempre se encontró sumergido, a una profundidad aproximada de
3 m. Su mayor concentración se restringió entre los 120 y 200 m a partir de la línea
de costa, aunque también se encontró entre los 20 y 40 m. Debido a que se observó
que la profundidad y transparencia del agua (turbidez) fue muy similar a lo largo del
gradiente vertical, se considera que su distribución estuvo determinada básicamente
por la disponibilidad de sustrato. Como se sabe, Sargassum tiene pieferencia por
sustrato rocoso o sustratos duros, (0 que permítíósu presentía en (os prímeros 40 m,
donde la frecuencia de este tipo de sustrato fue mayor. Es posible también que las
interacciones biológicas, como la competencia interespecífica, tengan un papel
importante en la distribución vertical de Sargassum, lo que podría explicai su ausencia
77

entre los 50 y 120 m, donde se observó una dominancia de Spyridia filamentosa,
Hypnea valentíae y Zostera marina (pasto). En verano, S. sinicola presentó su mayor
,distribución porque encontró las condiciones óptimas para su desarrollo, mientras que
en otoño sólo se encontró en los 30 y 200 m y los talos estuvieron epifitados por
Jania sp y Laurencia sínicola entre otras, señalando su época de decadencia.
8. 5. 5. Codium cuneatum
El patrón de abundancia estacional de Codíum cuneatum, parece estar más
relacionado con la salinidad que con la temperatura. La biofiasa máxima ocurrió en
primavera, cuando la salinidad promedio fue baja (32. 5 ‘/OO) y la mínima, en otoño,
cuando fue más alta (37 ‘/OO), haciéndose manifiesto una relación inversa. Las
especies de Codium presentan una gran área superfificial de pigmentos fotosintéticos
que les permiten captar o absorber gran cantidad de luz (Lobban 19811, por lo que
probablemente este parámetro también influyó en su patrón de producción estacional.
En invierno, cuando los dias fueron más cortos, se registró un incremento en la
biomasa de Codium, lo que hace suponer mejores condiciones de luz (cantidad y
calidad), las cuales se mantienen hasta primavera cuando alcanzó su mayor
producción. Por el contrario, en verano, cuando los dias fueron más largos, las
condiciones fueron desfavorables para su desarrollo, como lo indica la disminución de
su biomasa.
La distribución vertical de C. cuneatum sigue su patrón de abundancia
estacional. En otoño, cuando fue más escaso, la mayor concentración se restringió a
los primeros 20 m, mientras que en invierno se distribuyó casi regularmente.
Tomando en cuenta que en invierno se incrementó la biomasa y’ se amplió la
distribución vertical de Codium, se considera a esta época como de reclutamiento. En
primavera y verano también se distribuyó ampliamente gracias a la disponibilidad de
sustratos duros. Asi, parece ser que el principal factor fisico que regula la distribución
vertical de esta especie es el sustrato, ya que en las áreas donde se enkontró fueron
78

abundantes las conchas, guijarros y tapetes de algas calcáreas (Jania spp y Amphiroa
spp) donde también se estableció. La luz, probablemente también influyó en la
&distribución vertical de Codium. En la laguna, el agua cerca de la costa era más
transparente que la de las porciones más internas en donde, aún cuando habia
visibilidad hasta el fondo, el agua era más turbia. Lo anterior, pudiera explicar la
mayor concentración de esta especie en los primeros 130 m durante el periodo de
estudio.
8. 6. ESPECIES EPILITICAS Y EPIFITAS.
Los diversos grupos que conforman una comunidad pueden separarse para su
estudio ecológico. En este caso, la comunidad de macroalgas se dividió en comunidad
d’e algas epilíticas y comunidad de algas epífitas para comprender mejor su dinámica,
particularmente en lo que se refiere al desarrollo y sucesión de las poblaciones
integrantes.
Entre las especies epífitas, algunas sólo pueden ocurrir con este modo de vida,
es decir, tienen una forma obligadamente epífita y en muchos casos forman
asociaciones permanentes o casi permanentes con otras especies de algas (Santelices
1977). Por otro lado, existen especies, principalmente pardas y verdes, que pueden
ser epilíticas, pero también ocurren como epífitas debido a la escasez de sustrato
(Sears y Wilce 1975, Kendrick et al. 1988). En el caso particular de la laguna, en
Punta Choya e Isla Garza se observaron mantos o tapetes de algas coralinas y sobre
ellas algas como Spyridia filamentosa, Laurencia spp, Hypnea valentíae, Dasya
bai//ouviana (se encontró también sobre pasto), las cuales se consideraron epilíticas
por ser el modo de ocurrencia más frecuente. Kendrick et al. (1988) sugieren que la
disponibilidad de sustrato colonizable parece ser más importante en la estructura de
la comunidad de epífitas que el disturbio y modelos competitivos aplicados a las
comunidades algales bentónicas.
79

Con base en las observaciones sobre fenología de las macroalgas y sus cambios
estacionales en distribución y abundancia, se detectó un patrón estacional diferente
ctanto para la comunidad de algas epilíticas como para la de epífitas, indicando un
desfasamiento en el desarrollo de cada comunidad que puede explicarse por las
características intrínsecas de los organismos, las interacciones entre ellos y su
relación con los factores físicos.
La comunidad de algas epilíticas presentó, en otoño, una disminución
considerable tanto en riqueza como en abundancia, lo cual se atribuyó a la
periodicidad estacional de las especies y a su estado de desarróllo. En la misma epoca,
la comunidad de epífitas alcanzó su máxima riqueza y abundancia, cuando la mayoría
de especies epilíticas estuvieron decadentes o senescentes. Ducker y Knox (1984)
observaron que las epífitas oportunistas o no específicas son sólo capaces de colonizar
algas maduras o las partes más viejas del talo y que en algunos casos sustituyen a las
epífitas específicas cuando las hospederas estan viejas o moribundas. Señalan
además, que las epífitas oportunistas no colonizan algas jovenes o en desarrollo
debido a los mecanismos de defensa activos que presentan, mientras que las epífitas
específicas son capaces de establecerse en plantas jóvenes o en crecimiento porque
tienen mecanismos de reconocimiento celular con las plantas hospederas. Kendrick y
hawkes (1988) encontraron altas densidades de especies epífitas en las porciones
más viejas del talo hospedero y mencionan que no es sorprendente encontrar más
epífitas en talos más grandes, maduros o senescentes. Por su parte, Sears y Wilce
(1975) observaron que las plantas jóvenes, fueron más coloridas y estuvieron libres
de epífitas, a diferencia de las plantas senescentes que fueron descoloridas y llenas
de epífitas. De esta manera puede establecerse que el estado de desarrollo de la
comunidad epilítica determina el establecimiento y desarrollo de la comunidad de
epífitas y que estas a su vez son indicadores del estado de salud de una planta,
población o comunidad de especies epilíticas.
Posiblemente la salinidad también fue un factor determinante en el desarrollo
80

de las epífitas, puesto que su mayor riqueza y abundancia ocurrió en otoño, cuando
la salinidad fue mayor y menor en primavera cuando la salinidad fue baja. Además,
tla reducción de epífitas observada en invierno, cuando se registró la salinidad más
baja, posiblemente estuvo relacionada con la periodicidad estacional de las especies,
debido a que son algas anuales, de ciclo de vida corto. Probablemente la salinidad
tiene un efecto diferencial en las poblaciones de algas, sin embargo, se requiere
considerar a otros factores y mas estudios experimentales o ecofisiológicos que
evaluen esta relación.
8. 7. FACTORES AMBIENTALES
8 .7. 1. TEMPERATURA
Los cambios climáticos estacionales y la dinámica oceanográfica de la región
determinaron las amplias variaciones de temperatura que se observaron en la laguna.
En verano, la mayor temperatura se explica porque, además de que hubo mayor
calentamiento del agua debido a que los dias fueron mas largos, fluyen aguas más
cálidas (tropicales) a través de la contracorriente Ecuatorial ya que en esta época se
intensifica hacia el norte (Fernández et al. 1993). Por el contrario, en invierno la
menor temperatura se atribuye al menor calentamiento del agua puesto que los dias
fueron más cortos y a la influencia de la Corriente de California que, en esta época,
se intensifica y se desplaza más al sur de la península transportando aguas frias.
Entre localidades no fue posible detectar un gradiente de temperatura general
debido a que los registros fueron insuficientes y a que presentó un comportamiento
diferente en cada época. Sin embargo, en primavera y verano, se registraron
temperaturas más bajas cerca de la boca y tendió a incrementarse hacia el interior de
la laguna, lo cual puede explicarse por el efecto de la evaporación, que presenta un
comportamiento particular en cada localidad determinado por las características
fisiográficas de cada una de ellas. En la cabecera y parte media de’la laguna, la

profundidad es menor en comparación con la boca, por lo mismo, el calentamiento del
agua es mayor. En otoño se registró mayor temperatura cerca de la boca que en el
interior de la laguna, señalando *la entrada de aguas más cálidas, que pudieran ser
aporte de la contracorriente Norecuatorial o bien ser efecto de el Niño 92/93. En
invierno la temperatura fue similar en todas las localidades.
8. 7. 2. SALINIDAD
Las fluctuaciones estacionales de este parámetro se relacionan también con las
variaciones climáticas y condiciones oceanográficas. En otoñóse registró la salinidad
más alta, probablemente como consecuencia de la acumulación de sales a partir de
verano, causada por la mayor evaporación cuando la temperatura fue elevada, asi
como por el aporte de la corriente Norecuatorial. En tanto, en invierno la salinidad fue
menor, como consecuencia de la baja evaporación provocada por la disminución de
la temperatura cuando los dias fueron menos largos y por la influencia de las aguas
de la corriente de California que son de baja salinidad, < a 34 o/oo (Fernández et al.
1993). Cabe señalar que en primavera se presenta una gran variación de la salinidad
como resultado de las diferencias fisiográficas que se presentan en cada localidad;
unas son menos profundas que otras y por lo tanto la evaporación es mayor.
Aún cuando las diferencias no fueron significativas, a lo largo de la laguna se
detectó un gradiente de salinidad, con salinidades menores en la boca y mayores en
la parte media y cabecera. La profundidad de las localidades y la intensidad de la
evaporación explican este resultado.
82

9. CONCLUSIONES.
De las especies determinadas 90 representan primeros registros para la laguna San
Ignacio, 6. C. S, que sumados a los previos, integran una lista de 103 especies de
macroalgas.
La comunidad de macroalgas está compuesta por especies de afinidad templada y
tropical, así como componentes de amplia distribución, por lo que es representativa
de una zona de transición templada-cálida, sin embargo, la mayoría de las especies
fueron de afinidad templada.
La mayor riqueza específica correspondió a las algas rojas, seguidas por las verdes
y pardas.
La composición ficoflorística entre localidades fue diferente.
Las localidades con sustratos duros (rocas, guijarros, conchas) presentaron mayor
riqueza específica que las que se caracterizaron por tener sustratos blandos (arenoso
y fango-arenoso).
En la cabecera o parte más interna de la laguna hubo escasa presencia de algas.
El mayor número de algas rojas y verdes ocurrió en la parte media de la laguna (Punta
Choya), mientras que el de pardas ocurrió cerca de la boca (Punta Piedra).
De las especies, 28 restringieron su distribución a sólo una localidad y’generalmente
se desarrollaron en una época el año, mientras que Gracilaria pacífica, Spyrídia
filamen tosa, H. valen tiae, Das ya baíllouviana, En teromorpha cla thra ta y E. ramulosa
presentaron una amplia distribución tanto espacial como temporal.5
83

Las localidades que presentaron mayor semejanza ficoflorística fueron Punta Choya,
Isla Garza y Punta Piedra, encontrando entre sus componentes especies perennes
estructuralmente más complejas.
El número de especies varió poco entre épocas del año (no se detectó un patrón
estacional evidente en riqueza específica) .
La mayoría de las algas fueron especies anuales, encontrando en verano el mayor
número.
Cada localidad presentó una estacionalidad
número de especies en otoño e invierno.
particular, manifestándose el mayor
En cuanto abundancia, las algas rojas fueron las de mayor biomasa, seguidas por las
pardas y verdes. En forma general, el patrón de abundancia estacional fue muy
evidente, con su máximo en verano y mínimo en primavera. Las algas rojas se ajustan
a este patrón no asi las pardas y verdes que logran su pico de abundancia en invierno.
La abundancia estacional de cada localidad fue diferente.
De las localidades; Punta Choya, Punta Piedra y La Boca, concentraron la mayor
abundancia de algas.
Las especies más abundantes fueron Gracilariapacifica, Spyridia filamentosa, Hypnea
valen tiae, Gracilariposis lemaneiformis, Padina durvillaei, Sargassum muticum,
Sargassum sinicola, Gracilaria subsecundata y Codium cuneatum, aunque algunos
en cierta época y otros en cierta localidad.
Se considera que los principales componentes que caracterizan la comunidad de
macroalgas de la laguna fueron Gracilaria pacifica, Spyridia filamentosa’ e Hypnea
84

valentiae, por presentar una amplia distribución y ser los más abundantes.
:Por su abundancia, las especies que constituyen un recurso potencial son Gracilaria
pacifica, Spyridia filamentosa e Hypnea valentiae, mientras que por los usos que se
les atribuyen y su frecuencia, otras especies potenciales son: Gracilariopsis
lemaneiformis, Laurencia spp, Dasya baillouviana, Polysiphonia spp, Sargassum
muticu~, Enteromorpha spp, Uva spp y Cladophora spp.
La estacionalidad de la comunidad de algas epilíticas es diferente a la de algas epífitas,
la primera alcanza su máximo desarrollo en verano, mientras qÚe la segunda lo logra
en otoño.
No se observaron patrones de zonación como los descritos para lugares templados.
85

_10. RECOMENDACIONES
: Considerando la importancia que reviste la laguna como parte de la Reserva de
la Biósfera El Vizcaíno, las investigaciones a realizarse en ella deben tener una
orientación ecológica, encaminada a aportar elementos para hacer un buen manejo y
aprovechamiento de sus recursos. A continuación, con base a las observaciones
resultantes en el presente estudio y a las posibilidades de aprovechamiento del recurso
algal, se mencionan algunos posibles temas a desarrollar en trabajos posteriores:
Realizar colectas mensuales para detectar con mayor pre%ión la composición
específica y sus cambios estacionales, y por lo menos durante dos años, ya que se
ha encontrado en otros lugares que la estacionalidad tanto en riqueza, composición
y abundancia puede variar interanualmente.
Considerando que el periódo de estudio se caracterizó por la presencia de un Niño
(ENSO) se recomienda monitorear la estructura de la comunidad ó llevar a cabo otro
estudio similar en condiciones climáticas de no Niño (“normales”).
Realizar estudios autoecológicos y sinecológicos con las especies algales dominantes
o características y otros organismos. Entre los primeros se recomienda: a) Estudiar el
efecto de la luz, temperatura, nutrientes y salinidad sobre
(abundancia) de los principales componentes ficológicos de la
Gracilaria pacifica, Sp yridia flamen tosa e Hypnea valen tiae.
la estacionalidad
comunidad como
b) Realizar estudios ecofisiológicos con los principales
para conocer su tolerancia y respuesta a las variaciones de
mencionados.
componentes ficológicos
los factores físicos antes
c) Realizar estudios de productividad de las principales componentes ficológicos.
86

d) Efectuar estudios fenológicos y de longevidad de los componentes ficológicos
de la comunidad de macroalgas de la laguna que representan un recurso potencial.
Entre trabajos sinecológicos: a) Estudiar la relación alga-pasto, puesto que se observó
que los pastos presentan una amplia distribución y gran abundancia en la laguna.
b) Realizar estudios de competencia intraespecífica (efectos alelopáticos) en
las localidades con comunidades estructuralmente más complejas como Punta Piedra,
Punta Choya e Isla Garza, entre las algas características y otras especies para conocer
el proceso de dominancia.
c) Estudiar el efecto de los depredadores sobre la comunidad de macroalgas.
d) Evaluar la importancia de la comunidad de macroalgas como procesadora de
energía através de la cadena trófica y como productora de materia orgánica.
En las localidades con baja riqueza especifica, que se caracterizan por presentar
sustrato arenoso, realizar estudios ambientales y de factibilidad para el cultivo de
algas de interés comercial como Gracilaria pacifica y otras agarofitas, que tienen
preferencia por este tipo de sustrato.
Para cuestiones de manejo del recurso alga, se hace necesario evaluar la biomasa
cosechable de las especies potenciales como Gracilaria pacifica, Spyridia filamentosa,
Hypnea valentiae, Sargassum muticum, En teromorpha s pp y Uva s p p.
Finalmente, también pueden realizarse estudios de tipo farmacológico eón Laurencia
spp, Dasya baillouviana, Polysiphonia spp, y Cladophora spp que si bien no son
abundantes, son especies frecuentes y pueden ser empleadas para obtener
compuestos antibióticos.
87

ll. LITERATURA CITADA
)Abbott 1. A. and W. J. North. í971. Temperature influentes on floral composition in
California Coastal Waters. Proceedings of the Seventh International Seaweed
Symposium. University of Tokyo Press. 72-79
Abbott‘ 1. A. and G. J. Hollenberg. 1976. M a r i n e
University Press, Stanford California 827 pp.
algae of California. Stanford
Aguilar-Rosas L. E. 1982. Ocurrencia de algas cafés (Phaeoph’yta) en la Bahía Todos
Santos, Baja California. Ciencias Marinas, 8(2): 25-34.
Aguilar-Rosas L. E. e 1. Pacheco-Ruíz. 1985. Nuevos registros y ampliación del rango
geográfico para algas marinas de la costa dei pacífico de Baja California,
México. ll. Ciencias Marinas, ll (2): 69-76.
Aguilar-Rosas R., 1. Pacheco-Ruíz, y L. E. Aguilar-Rosas. 1984. Nuevos registros y
algunas notas para la flora algal marina de la costa noroccidental de Baja
California, México. Ciencias Marinas, 1 O(2): 149-l 58.
Aguilar-Rosas R. y M. A. Aguilar-Rosas. 1986. Nuevos registros de algas marinas
para la flora de Baja California, México. Ciencias Marinas, 12(2): 17-20
Aguilar-Rosas R., 1. Pacheco-Ruíz y L. E. Aguilar-Rosas. 1990. Algas marinas de las
Islas Todos Santos, Baja California, México. Ciencias Marinas, 16(2): 1 17-l 29.
Aguilar-Rosas R. y L. E. Aguilar-Rosas. 1993a. Cronología de la colonización de
Sargassum muticum (Phaeophyta) en las costas de la Península de Baja
California, México (1971-l 990). Revista de Investigación Cientifica UABCS,
4(1):41-51.t
88

Aguilar-Rosas R. y M. A. Aguilar-Rosas. 1993b. Fitogeografía de las algas pardas
(Phaeophyta) de la Península de Baja California, pp 197-206. En: Biodiversidad
\ marina y costera de México. S. 1. Salazar Vallejo y N. E. Gónzalez (eds.) Com.
Nac. Biodiversidad y CIQRO, México, 865 pp.
Anónimo. 1993. Informe anual. Comisión Interamericana del Atún Tropical. La Jolla,
California. 315 pp.
Berchez F. A., R. T. L. Pereira and N. F. Kamiya. 1993. Culture of H y p n e a
musciformis (Rhodophyta, Gigartinales) on artificial súbstrates attached to
linear ropes. Hydrobiologia, 260/261: 415-420.
Bolton J. J. and H. Stegenga. 1987. The marine algae of Hluleka (Transkei) and the
warm temperate/subtropicaI transition on the east coast of southern Africa.
Helgoländer Meeresunters, 41: 165-l 83.
Breceda A., A. Castellanos, L. Arriaga y A. Ortega. 1991. Conservación y áreas
protegidas en Baja California Sur, 21-32 pp. En: La Reserva de la Biosfera El
Vizcaíno en Baja California. A . Ortega y L. Arriaga (Eds.) . Centro de
Investigaciones Biológicas de Baja California Sur A. C. México. 317 pp.
Breceda A. 1992. Plan de manejo para la reserva de la biósfera El Vizcaíno. Reporte
Preeliminar, Centro de Investicaciones
México.
Biológicas de Baja California Sur A. C.
Brower J. E. and J. H. Zar. 1984. Field and laboratory methods for genkral ecology.
Second Ed. Wm. C. Brown Publishers. Dubuque, Iowa, USA, 226 pp.
Carpenter R. C. 1990. Competition among marine macroalgae: a physiological
perspective. Journal of Phycology, 26: 6-l 2.c
89

_
. . Chapman A. R. 0. 1979. Biology of seaweeds: levels of organization. University Park
Press, Baltimore. 134 pp.
Contreras F. 1988. Las lagunas costeras mexicanas. Centro de Ecodesarrollo,
Secretaría de Pesca. 263 pp.
Coutinho R. and R. Zingmark. 1993. In teract ions of l ight and nitrogen on
photosynthesis and growth of the marine macroalga Vulva curvata (Kützing) De
Toni. Journal of Experimental Marine Biology and Ecology, 167: 1 l-l 9.
Crisci J. V. y M. F. López. 1983. Introducción a la teoría y práctica de la taxonomía
numérica. Secretaría General de la Organización de los Estados Americanos.
Serie de Biología, Monografía No. 26. 132 pp.
Danneman G. D. 1994. Biología reproductiva del aguila pescadora (Pandion haliaetus)
en Isla Ballena, Laguna San Ignacio, Baja California Sur. Te& de Maestría.
Centro Interdisciplinario de Ciencias Marinas, La Paz, B. C. S. México. 77 p.
Danneman G. D. y J. De La Cruz. 1993. Ictiofauna de Laguna San Ignacio, Baja
California Sur, México. Ciencias Marinas, 1 g(3): 333-341.
Darley W. M. 1987. Biología de las algas; enfoque fisiológico. Primera Edición, Ed.
Limusa, México, 236 pp.
Dawes C. J. 1986. Botánica Marina. Primera Edición. Ed. Limusa. México. 673 pp.
Dawson E. Y. 1944. The marine algae of the Gulf of California. Allan Hancock Pacific
Expeditions, 3: 189-454.
90

Dawson E. Y. 1948. Resultados preliminares de un reconocimiento de las algas
marinas de la costa Pacífica de México. Revista de la Sociedad Mexicana de
Historia Natural, 7: 167-215.
Dawson E. Y. 1953. Marine red algae of Pacific Mexico. 1. Bangiales to Corallinaceae
subfamily Corallinoideae. Allan Hancock Pacific Expeditions, 17( 1): l-239..
Dawson E. Y. 1960a.
benthic flora. In:
ll, Marine Biotas
Dawson E. Y. 1960b.
A review of the ecology, distribution and affinities of the
The biogeography of Baja California and adjacent seas. Part
Systematic Zoology. 9:93-l OO. -
Marine red algae of Pacific Mexico. III. Cryptonemiales,
Corallinaceae subfamily Melobesioideae. Pacific Naturalist, 2(l): l-l 25.
Dawson E. Y. 1961a. A guide to the literature and distributions of Pacific benthic
algae from Alaska to the Galapagos Islands. Pacific Science, 15: 371-461.
Dawson E. Y. 1961 b. Marine red algae of Pacific Mexico. IV. Gigartinales. Pacific
Naturalist, 2(5): 191-343.
Dawson E. Y. 1962. Mar ine red algae of Paci f ic Mexico. VI I . Ceramia les:
Ceramiaceae, Delesseriaceae. Allan Hancock Pacífic Expeditions, 26(2): l-207.
Dawson E. Y. 1963. Marine red algae of Pacific Mexico. VIII. Ceramiales: Dasyaceae,
Rhodomelaceae. Nova Edwigia, 6: 400-481.
Ducker S. C. and R. B. Knox. 1984. Epiphytism at the cellular level with special
referente to algal epiphytes, pp 1 13-I 33. En: Encyclopedia of plant physiology
New Series, Vol. 17. H. F. Linskens y J. Heslop-Harrinson (eds). Springer-
Verlag Berlin Heidelberg.L
91

Espinoza-Avalos J. 1993. Macroalgas marinas del Golfo de California, pp 328-357.
En: Biodiversidad marina y costera de México. S. 1. Salazar Vallejo y N. E.
. Gónzalez (eds.) Com. Nac. Biodiversidad y CIQRO, México, 865 pp.
Fernández E. A., A. Gallegos G. y J. Zavala H. 1993. Oceanografía física de México.
Ciencia y Desarrollo, CONACyT 18(108): 25-35.
Folk R. L. 1954. The distinction between grain size and mineral composition in
sedimentary rock nomenclatura. Journal of Geology, 62: 344-359.
Garcia M. S. 1994. Productos pesqueros, una industria nutritiva. Revista del
Consumidor, 207: 9-l 6.
Grassoff K. 1983. Determination of salinity, pp 31-59.in: Methods of seawater
analysis. K. Grassoff, M. Ehrhardt, K. Kremling (eds.). Verlag Chemie,
Alemania, 419 pp.
Guzmán del Proó S. A., M. Casas, C. A. Díaz, L. Ma. Díaz, B. J. Pineda y Ma. L.
Sánchez R. 1986. Diagnóstico sobre las investigaciones y explotación de las
algas marinas en México. Investigaciones Marinas CICIMAR, 3 (2): l-63.
Harlin M. M. 1995. Changes in major plants groups following nutrient enrichment, pp
173-l 87. En: Eutrophic shallow estuaries and lagoons. A. J. McComb (ed.).
CRC Press.
Huerta-Muzquiz L. y C. Mendoza-González. í985. Algas marinas de la’parte sur de la
Bahía de la Paz, Baja California Sur. Phytologia, 59(l): 35-57.
Hommersand M. H., M. D. Guiry, S. Fredericq and G. L. Leister. 1993. New
perspective in the taxonomy of the Gigartinaceae (Gigartinales, Rhodophyta).
92

Hydrobiologia, 260/261: 105-l 20.
,Jones M. L. and S. L. Swartz. 1984. Demography and phenology of gray whales and
evaluation of whale-watching activities in Laguna San Ignacio, Baja California
Sur, México. In: The gray whale Eschrictiusrobustus. M. L. Jones, S. L. Swartz
.y S. Leatherwood Eds. Academic Press, 600 pp.
Kendrick G. A. and M. W. Hawkes. 1988. The epiphyte Macrocladia coulteri
(Rhodophyta): changes in population structure with spatial and temporal
variation in availability of host species. Marine Ecology Pfogress Series, 43: 79-
86.
Kendrick G. A., D. 1. Waiker and A. J. McComb. 1988. Changes in distribution of
macro-algal epiphytes on stems of seagrass Amphibolís antarctica along a
salinity gradient in Shark Bay, Western Australia. Phycologia, 27(2): 201-208.
Kendrick G. A., J. M. Huisman and D. 1. Walker. 1990. Benthic macroalgae of Shark
Bay, western Australia. Botanica Marina, 33: 47-54.
Krebs C. J. 1985. Ecología: Estudio de la distribución y la abundancia. Segunda
Edición. Harla, Harper 81 Row Latinoamericana, México, 753 pp.
León H. D. Fragoso, D. León, C. Candelaria, E. Serviere and J. González-González.
1993. Characterizacion of tidal pool algae in the Mexican Tropical Pacific coast.
Hydrobiologia, 260/261: 197-205.
Littler M. M. and D. S. Littler. 1981. Intertidal macrophyte communities from Pacific
Baja California and the upper Gulf of California: relatively constan V S
environmentally fluctuating systems.Marine Ecology Progress Series,4: 145-I 58.
93

Littler M. M. and D. S. Littler. 1984. Relationships between macroalgal funtional forms
groups and sustrata stability in a subtropycal rocky-intertidal system. Journal
of Experimental Marine Biology and Ecology, 74: 13-34.
Ludwing J. A. and J. F. Reynolds. 1988. Statistical Ecology. John Wiley & Sons,
.New York, N. Y. 337 pp.
Lüning K. 1990. Seaweeds: their environment, biogeography, and ecophysiology.
Wiley Interscience, U. S. A., 527 pp.
Lüning K. 1993. Environmental and interna1 control of seasonal growth in seaweeds.
Hydrobiologia, 260/26 1: l-l 4.
McCourt R. M. 1983. Zonation and phenology of three species of Sargassum in the
intertidal zone of northern Gulf of California. Ph. D. Thesis University of
Arizona. 144 pp.
Mann K. H. 1972. Ecological energetics of the seaweed zone in a marine bay on the
Atlantic Coast of Canada. 1. Zonation and biomass of seaweeds. Marine
Biology, 12: l-l 0.
Mateo-Cid L. E., 1. Sánchez R., Y. E. Rodríguez M. y M. M. Casas V. 1993. Estudio
florístico de las algas marinas bentónicas de Bahía Concepción, B. C. S. México.
Ciencias Marinas, 19( 1): 41-60.
Mathieson A. C. 1989. Phenological patterns of Northern New England seaweeds.
Botanica Marina, 32: 419-438.
Mayer A. M. S. 1981. Studies on Gracilaria sp in bahia Arredondo, Chubut province,
Argentina. Proceedings of the Xth International Seaweed Symposiu’m, 704-710.
94

Mendoza-González C. y L. E. Mateo-Cid. 1985. Contribución al estudio florístico
ficológico de la costa occidental de Baja California, México. Phytologia, 59(l):
17-33.:
Molloy F. J. and J. J. Bolton. 1995. Distribution, biomass and production oí Gracilaria
in Lüderitz, Namibia. Journal of Applied Phycology, 381-392.
Murray S. N. and M. H. Horn. 1989. Variations in standing stocks of central California
macrophytes from a rocky intertidal habitat before and during the 1982-I 983
El Niño. Marine Ecology Progress Series, 58: 1 13-l 22:
Murray S. N. and M. M. Littler. 1989. Seaweeds and seagrasses of Southern
California: Distributional lists for twenty-one rocky intertidal sites. Bulletin of
Southern California Academy of Sciences, 88 (2): 61-79.
Murthy M. S., T. Ramakrishna, Y. N. Rao, and D. K. Ghose. 1989. Ecological studies
on some agarophytes from Veraval coast (India). 1. Effects of aerial conditions
on the biomass dynamics. Botanica Marina, 32 (6): 515-520.
Núñez-López R. A. 1993. Biomasa estacional específica de Sargassum spp
(Sargassaceae, Phaeophyta) en tres zonas de Bahía Concepción, 8. C. S. Tesis
de Licenciatura. Universidad Autónoma Metropolitana Unidad Xochimilco. 55~.
Reitherman B. and J. Storrer. 1981. A preliminary report on the reproductive biology
and ecology of the Whale Island osprey (Pandion halietus) population, San
Ignacio Lagoon, B. C. S. Research Associates of the Western Foundation of
Vertebrate Zoology. Los Angeles, California. 28 p.
Riosmena R. R., D. A. Siqueiros-Beltrones, 0. Garcia de la Rosa, y V. Rocha-Ramirez.
1991. The extension geographic range of selected seaweeds ‘on the Baja
95

‘. California Per-insula. Revista de Investigación Científica, UABCS, 2(2): 13-20.
,Robledo R. D. 1990. Las macroalgas marinas, un recurso desconocido. Instituto de
Ciencia y Tecnología (ICyT), 12(169): 3-8.
Sanchéz-Rodríguez l., C. Fajardo L. y C. Oliveiro P. 1989. Estudio florístico estacional
de las algas en Bahía Magdalena, B. C. S. México. Investigaciones Marinas
CICIMAR, 4( 1): 16-35.
Santelices B. 1977. Ecología de algas marinas bentóntias: efecto de factores
ambientales. Documento de la dirección general de investigaciones, Pontificia
Universidad Católica de Chile, Santiago de Chile. 488 p.
Santelices B. 1980. Muestreo cuantitativo de comunidades intermareales de Chile
central. Archivos de Biologia y Medicina Experimentales, 13: 413-424
Scagel, R. F., P. W. Gabrielson, D. G. Garbary, L. Golden, M. W. Hawkes, S. C.
Lindstrom, J. C. Oliera and T. B. Widdoson. 1989. A synopsis of the benthic
marine algae of the British Columbia, Southest Alaska, Washington and Oregon.
Phycological Contrib. No.3. University of British Columbia, Vancouver. 532 p.
Sears J. R. and R. T. Wilce. 1975. Sublittoral, benthic marine algae of southern Cape
Cod and adjacent islands: seasonal periodicity, associations, diversity, and
floristic composition. Ecological Monographs, 45: 377-365.
Serviere Z. E. 1993. Descripción y análisis del litoral rocoso de Bahía de Banderas,
Jalisco-Nayarit. Tesis Doctoral. Facultad de Ciencias, Universidad Nacional
Autónoma de México. 71 p.
96

Schenkaman R. F. 1989. Hypnea musciformis (Rhodophyta): ecological influente on
growth. Journal of Phycology, 25: 192-I 96.
Schneider C. W. and R. B. Searles. 1991, Seaweeds of the Southern United States:
Cape Hatteras to Cape Canaveral. Duke University Press. 563 pp.
Soto-Bálderas M. G. y S. Alvarez-Borrego. 1991. Nutrientes inorgánicos en los canales
de inundación de las marismas de una laguna costera del noreste de Baja
California. Ciencias Marinas, 17(3): I-20.
Stewart, J. G. 1989. Maintenance of balanced, shifting boundary between the
seagrass Phyllospadix and algal turf. Aquatic Botany, 33: 223-241.
Stewart, J. G. 1991. Marine algae and seagrasses of San Diego County. California
Sea Grant College, University of California. 197 pp.
Swartz S. L. and W. C. Cummings. 1978. Gray whales, Eschrichtius robustus, in
Laguna San Ignacio, Baja California Sur, México. Final Report. Marine Mammal
Commission, Whashinton, D. C. 38 p.
Taylor W. R. 1960. Marine Algae of the eastern tropical and subtropical coasts of the
Americas. The Universidad of Michigan Press. Scientific Series,Vol. Xx1.780 pp.
Tegner M. J. and P. K. Dayton. 1987. El Niño effects on Southern California kelp
forest communities. Advances in Ecological Research, 17: 243-279.
Torres M. E. 1991. Zonación de macrolgas bentónicas en el arrecife de Puerto
Morelos, Quintana Roo. Tesis Profesional. Universidad Nacional Autónoma de
México. 64 pp.
97

Tokoya N. S. and E. C. de Oliveira. 1992. Efectos de la salinidad en la tasa de
crecimiento, morfología y contenido de agua de algunas algas rojas brasileñas
: de importancia económica. Ciencias Marinas, 18 (2): 49-64.
Trono G. C. 1988. Seaweed ecology: influente of environmental factors on the
structure, phenology and distribution oí seaweeds communities. Report on the
training course on seaweed farming. ASEAN/UNDP/FAO.Manila Philippines. 14-
16 p.
Trono G. C. Jr. and A. Saraya. 1987. The structure and distri6ution of macrobentic
algal comunities on the reef of Santiago Island, Bolinao, Pangansinan. The
Philippine Journal of Science, 17: 63-81.
Vásquez J. y J. González. 1994. Métodos de evaluación de macroalgas submareales.
En: Manual de métodos ficológicos. En prensa. Universidad de Concepción,
Chile.
Velkanni K. and R. Rengasamy. 1992a. lnfluence of light on growth and pigment
composition of Hypnea valentiae (Turner) Montagne.Mahasagar,25(1):45-49.
Velkanni K. and R. Rengasamy. 1992b. Influente of salinity on growth, pigments and
k-carragenan contents of Hypnea valentiae (Turner) Montagne. Mahasagar,
25(2): 123-I 28.
Villalard-Bohnsack M. and M. M. Harlin. 1992. Seasonal distribution and reproductive
status of macroalgae in Narragansett Bay and associated waters, Rhode Island,
USA. Botanica Marina, 35: 205-214.
Whittick A., R. G. Hooper and G. R. South. 1989. Latitude, distribution and
phenology: reproductiv’e strategies in some Newfoundland Seawee¿is. Botanica
98

Marina, 32: 407-417.
,Woelkerling W. J. 1990. An introduction. pp. 1-6. En: Biology of red algae. K, M. Cole
y R. G. Sheath (eds.). Cambridge University Press. 517 pp.
Wolfe J. M. and M. M. Harlin. 1988a. Tidepools in Southern Isiand, U. S. A. 1.
Distribution and seasonality of macroalgae. Botanica Marina, 31: 525-536.
Wolfe J. M. and M. M. Harlin. 1988b. Tidepools in Southern Island, U. S. A. ll.
Species diversity and similarity analisis of macroalgal communities. Botanica
Marina, 31: 537-546 .
Wynne M. J. 1986. A checklist of benthic marine algae of the tropical and subtropical
western Atlantic. Canadian Journal of Botany, 64: 2239-2281.
Zertuche G. J. 1993a. Situación actual de la industria de las algas marinas
productoras de ficocoloides en México, pp 33-38. En: Situación actual de la
industria de macroalgas productoras de ficocoloides en America Latina y El
Caribe. J. Zertuche G. (Ed.). Organización de las Naciones Unidas para la
Agricultura y la Alimentación. FAO. Documento de campo No. 13, 56 pp.
Zertuche G. J. 1993b. Situación actual de la industria de las algas agarofitas en
America Latina y El Caribe, pp 5-l 5. En: Situación actual de la industria de
macroalgas productoras de ficocoloides en America Latina y El Caribe. J.
Zertuche G. (Ed.). Organización de las Naciones Unidas para la Agricultura y la
Alimentación. FAO. Documento de campo No. 13, 56 pp.
99


















