
Esterification of methyl glycoside mixtures by lipase catalysis
4
Introduction In terms of biotechnology associated with the pulp and paper industry, hemicelluloses are important wood fibre components. If the starting material for a fractionation process is softwood, the main hemicellulose to be consid- ered is galactoglucomannan. During the fractionation, hemicellulose fragments with a large variation in composition and different chain lengths are produced. In order to modify such fractions hydrophobically, we have searched for enzymatic transformations. Hydrolytic enzymes such as lipases and esterases readily catalyse the acylation of the primary 6-position on carbohydrate substrates with high regioselectivity when organic solvents are used (Therisod and Klibanov, 1986, 1987). We thus decided to study the biocatalysed acylation of methyl pyranosides having the same anomeric configu- ration as that of softwood galactoglucomannan to deter- mine possibility of selectively introducing hydrophobic acyl groups (Ciuffreda et al., 1990; Colombo et al., 1992; de Goede et al., 1994). Materials and methods Materials Lipases from Candida antarctica, Pseudomonas cepacia, Candida cylindracea, Mucor miehei, Humicola lanuginosa, Candida rugosa OF, Aspergillus niger and porcine pancreas were gifts from Novo Nordisk A/S. Candida antarctica lipase B, Novozym 435, was the only immobilised enzyme. Substilisin, porcine liver esterase, methyl a-D- galactopyranoside, methyl b-D-glucopyranoside and methyl b-D-mannopyranoside isopropylate were from Sigma. Esterification of methyl b-D-glucopyranoside. A mixture of methyl b-D-glucopyranoside (43 mM) and octanoic acid (0.5 M) in acetonitrile (1.2 ml) was added to ten vials, each containing 10 mg enzyme, and shaken at 120 rpm for 24 hours at 45°C. Esterification of glycopyranoside mixture. Methyl a-D-galactopyranoside, methyl b-D-glucopyra- noside and methyl b-D-mannopyranoside isopropylate were dissolved in acetonitrile. To remove the isopropanol from the mannoside isopropylate, the acetonitrile solu- tion was evaporated under reduced pressure (repeated three times). Octanoic acid (0.5 M) in acetonitrile was added to the residue to give a solution with a molar ratio of 11:1 between acid and methyl glycosides. Aliquots of the solution (1.2 ml, 15 mM of each sugar) were added to capped vials containing the enzyme (10 mg), and shaken at 120 rpm for different periods of time at 45°C. Octanoyl esters of the methyl glyco- pyranosides were analysed by GC after silylation, using hexadecane as an internal standard. Preparation of 6-O-monoester glycosides. The different methyl 6-O-octanoyl-glycosides were each prepared on a larger scale for NMR and MS analyses. The (C. antarctica lipase B) reaction conditions were similar to those in the enzyme activity test. After silica gel column chromatography fine crystals of methyl 6-O-octanoyl-b-D-glucopyranoside (mp 88°C, Abe et al., 1995 ) and methyl 6-O-octanoyl-a- D-galactopyranoside (mp 137 °C) were obtained from ethyl acetate: hexane. The methyl 6-O-octanoyl-b-D- mannopyranoside had the consistence of syrup. The pure compounds were identified by 1 H and 13 C NMR using a 300 MHz Brücker spectrometer. 1 1 1 1 1 1 Biotechnology Letters, Vol 19, No 1, January 1997, pp. 15–18 © 1997 Chapman & Hall Biotechnology Letters · Vol 19 · No 1 · 1997 15 Esterification of methyl glycoside mixtures by lipase catalysis Armando Córdova 1 , Karl Hult 2 and Tommy Iversen 1 * 1 STFI, Swedish Pulp and Paper Research Institute, PO Box 5604 , S-114 86 Stockholm, Sweden; 2 Department of Biochemistry and Biotechnology, Royal Institute of Technology S-100 44 Stockholm, Sweden The acylation of a mixture of methyl a-D-galacto-, methyl b-D-gluco- and methyl b-D-mannopyranosides by octanoic acid was catalysed by lipases from Candida antarctica, Pseudomonas cepacia, Candida rugosa OF or Mucor miehei in acetonitrile at 45°C. The methyl glycopyranosides had the same anomeric configuration as that of the softwood hemicellulose galactoglucomannan. C. rugosa OF lipase had almost no substrate specificity and P. cepacia lipase had a high substrate specificity for the esterification of methyl a-D-galactopyranoside.
-
Upload
armando-cordova -
Category
Documents
-
view
214 -
download
1
Transcript of Esterification of methyl glycoside mixtures by lipase catalysis

IntroductionIn terms of biotechnology associated with the pulp andpaper industry, hemicelluloses are important wood fibrecomponents. If the starting material for a fractionationprocess is softwood, the main hemicellulose to be consid-ered is galactoglucomannan. During the fractionation,hemicellulose fragments with a large variation incomposition and different chain lengths are produced.In order to modify such fractions hydrophobically, wehave searched for enzymatic transformations. Hydrolyticenzymes such as lipases and esterases readily catalyse theacylation of the primary 6-position on carbohydratesubstrates with high regioselectivity when organicsolvents are used (Therisod and Klibanov, 1986, 1987).We thus decided to study the biocatalysed acylation ofmethyl pyranosides having the same anomeric configu-ration as that of softwood galactoglucomannan to deter-mine possibility of selectively introducing hydrophobicacyl groups (Ciuffreda et al., 1990; Colombo et al., 1992;de Goede et al., 1994).
Materials and methodsMaterialsLipases from Candida antarctica, Pseudomonas cepacia,Candida cylindracea, Mucor miehei, Humicola lanuginosa,Candida rugosa OF, Aspergillus niger and porcine pancreaswere gifts from Novo Nordisk A/S. Candida antarcticalipase B, Novozym 435, was the only immobilisedenzyme. Substilisin, porcine liver esterase, methyl a-D-galactopyranoside, methyl b-D-glucopyranoside andmethyl b-D-mannopyranoside isopropylate were fromSigma.
Esterification of methyl b-D-glucopyranoside.A mixture of methyl b-D-glucopyranoside (43 mM) andoctanoic acid (0.5 M) in acetonitrile (1.2 ml) was addedto ten vials, each containing 10 mg enzyme, and shakenat 120 rpm for 24 hours at 45°C.
Esterification of glycopyranoside mixture.Methyl a-D-galactopyranoside, methyl b-D-glucopyra-noside and methyl b-D-mannopyranoside isopropylatewere dissolved in acetonitrile. To remove the isopropanolfrom the mannoside isopropylate, the acetonitrile solu-tion was evaporated under reduced pressure (repeatedthree times). Octanoic acid (0.5 M) in acetonitrile wasadded to the residue to give a solution with a molarratio of 11:1 between acid and methyl glycosides.Aliquots of the solution (1.2 ml, 15 mM of each sugar)were added to capped vials containing the enzyme (10 mg), and shaken at 120 rpm for different periods oftime at 45°C. Octanoyl esters of the methyl glyco-pyranosides were analysed by GC after silylation, usinghexadecane as an internal standard.
Preparation of 6-O-monoester glycosides.The different methyl 6-O-octanoyl-glycosides were eachprepared on a larger scale for NMR and MS analyses.The (C. antarctica lipase B) reaction conditions weresimilar to those in the enzyme activity test. After silica gel column chromatography fine crystals of methyl 6-O-octanoyl-b-D-glucopyranoside (mp 88°C,Abe et al., 1995 ) and methyl 6-O-octanoyl-a-D-galactopyranoside (mp 137 °C) were obtained fromethyl acetate: hexane. The methyl 6-O-octanoyl-b-D-mannopyranoside had the consistence of syrup. The purecompounds were identified by 1H and 13C NMR usinga 300 MHz Brücker spectrometer.
11
11
11
11
11
11
1
Biotechnology Letters, Vol 19, No 1, January 1997, pp. 15–18
© 1997 Chapman & Hall Biotechnology Letters · Vol 19 · No 1 · 1997 15
Esterification of methyl glycosidemixtures by lipase catalysisArmando Córdova1, Karl Hult2 and Tommy Iversen1*1STFI, Swedish Pulp and Paper Research Institute, PO Box 5604 , S-114 86 Stockholm, Sweden; 2Department ofBiochemistry and Biotechnology, Royal Institute of Technology S-100 44 Stockholm, Sweden
The acylation of a mixture of methyl a-D-galacto-, methyl b-D-gluco- and methyl b-D-mannopyranosides by octanoicacid was catalysed by lipases from Candida antarctica, Pseudomonas cepacia, Candida rugosa OF or Mucor mieheiin acetonitrile at 45°C. The methyl glycopyranosides had the same anomeric configuration as that of the softwoodhemicellulose galactoglucomannan. C. rugosa OF lipase had almost no substrate specificity and P. cepacia lipasehad a high substrate specificity for the esterification of methyl a-D-galactopyranoside.
11
11
11
11
11
11
1
A. Córdova et al.
16 Biotechnology Letters · Vol 19 · No 1 · 1997
Table 1 The activities of ten enzymes with respect to their esterification of methyl b-D-glcp (43 mM) and otanoicacid (0.5 M) in acetonitrile (1.2 ml) at 45˚C
Enzyme I (%) II (%) Enzyme I (%) II (%)
C. antarctica 90 90 H.languiosa 0 0C. rugosa 51 37 C. cylindracea 1 1A. niger 27 11 P. cepacia 81 73M. miehei 77 68 Subtilisin 0 0Porcine pancreas 22 9 Porcine liver esterase 0 0
I = Methyl b-D-glcp (%) reacted and II = methyl 6-O-octanoyl-b-D-glcp (%) produced after 24h. The amount of each enzyme preparationwas 10 mg.
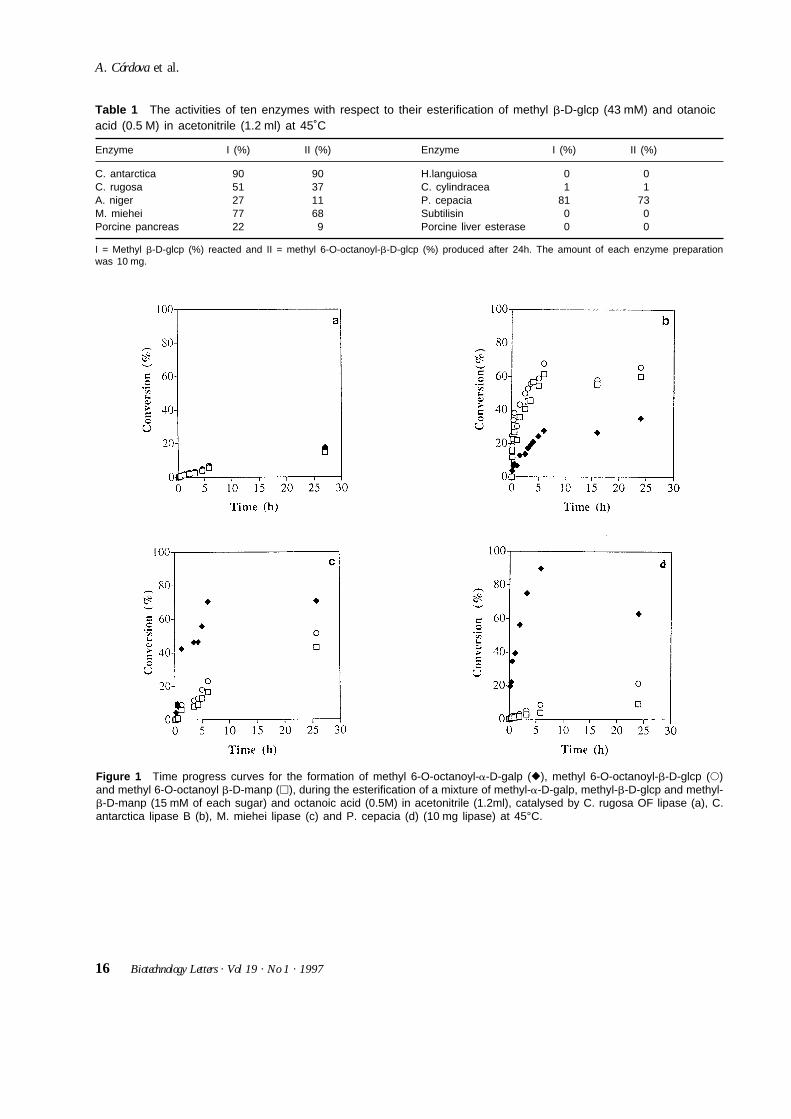
Figure 1 Time progress curves for the formation of methyl 6-O-octanoyl-a-D-galp (r), methyl 6-O-octanoyl-b-D-glcp (s)and methyl 6-O-octanoyl b-D-manp (h), during the esterification of a mixture of methyl-a-D-galp, methyl-b-D-glcp and methyl-b-D-manp (15 mM of each sugar) and octanoic acid (0.5M) in acetonitrile (1.2ml), catalysed by C. rugosa OF lipase (a), C.antarctica lipase B (b), M. miehei lipase (c) and P. cepacia (d) (10 mg lipase) at 45°C.

Results and discussionEnzyme screeningInitially eight lipases, subtilisin and porcine liveresterase were screened for the acylation of methyl b-D-glucopyranoside. The acyl donor was octanoic acid inacetonitrile (Ljunger et al., 1994). The consumption ofthe substrate and the production of 6-O-monoester after24 h. are presented in Table 1. C. antarctica lipase B hadthe highest conversion, the only discernible productbeing the 6-O-monoester. P. cepacia lipase showed arelatively high conversion but also some diester forma-tion. M. miehei and C. rugosa OF lipases both showedactivity with selectivity towards 6-O monoesterformation. The porcine pancreas lipase and the A. nigerlipase had low activities and the other hydrolases noactivity.
Selectivity testAccording to the enzyme screening results, the fourlipases from C. rugosa OF, C. antartica B, M. miehei andP. cepacia, exhibited the highest conversions of methyl b-D-glucopyranoside, and those were chosen for furtherstudies. Since we wished to examine whether theenzymes showed both substrate specificity and regio-selectivity, a mixture of methyl a-D-galactopyranoside,methyl b-D-glucopyranoside and methyl b-D-mannopyranoside was used. The progress of the methyl6-O-octanoyl-b-D-glycoside formation was monitoredby gas chromatography. The results can be seen in Figure1. The C. rugosa OF lipase, Fig. 1a, was unspecific andshowed low initial rates with all the three glycosides.Figure. 1b shows that the C. antarctica lipase B specifically acylated the primary hydroxyl group of methyl b-D-glucopyranoside and the methyl b-D-mannopyranoside substrates. These acylations alsoproceeded with the highest initial rates of all enzyme-substrate combinations. The conversion of methyl b-D-glucopyranoside in mixtures together with the
other glycosides were lower, 70%, than in the experi-ment containing methyl b-D-glucopyranoside as solesubstrate, 90%, (24 hours reaction time). M. mieheilipase, Fig. 1c, was initially selective towards the acyla-tion of the primary hydroxyl group of methyl a-D-galactopyranoside, but as the reaction time proceeded theacylation of the secondary hydroxyl groups of methyl-a-D-galactopyranoside monoester began, and this led todiester formation as indicated by gas chromatography.The other two methyl b-D-glycopyranosides were alsoacylated to a high extent. P. cepacia, Fig. 1d, showed ahigh selectivity for the acylation of methyl a-D-galacto-pyranoside, compared to the acylation of the other twomethyl-b-D-glycosides. The formation of methyl 6-O-octanoyl-a-D-galactopyranoside was high duringthe first six hours of the reaction. At high degrees ofconversion, the methyl 6-O-octanoyl-a-D-galactopyra-noside became a substrate for diester formation.
Selectivity dataUsing the linear part of the formation curves of the three6-O-monoester products the initial rates were calculated.The results are presented in Table 2. The specificity for two competing substrates can be expressed as theratio of their specificity constants (Fersht, 1995). Thespecificity ratio (S) has here been derived as the ratio ofthe initial rates in a combined incubation (S= vi/vj)(Table 2). The equation DDG# = –R*T* ln S relates thespecificity constant to the difference in activation ener-gies of the competing substrates (Fehrst, A., 1995). Itshould be noted that the whole weight of the enzymepreparation was used when the initial rates werecalculated.
ConclusionAcylation catalysed by Pseudomonas cepacia lipase washighly selective for methyla-D-galactopyranoside rela-tive to the acylation of methyl b-D-glucopyranoside and
11
11
11
11
11
11
1
Esterification of methyl glycoside mixtures by lipase catalysis
Biotechnology Letters · Vol 19 · No 1 · 1997 17
Table 2 The initial rates of formation (vi) of the three 6-O-monoester products (A = methyl 6-O-octanoyl-a-D-galactopyranoside, B = methyl 6-O-octanoyl-b-D-glucopyranoside and C = methyl 6-O-octanoyl-b-D-manno-pyranoside) were calculated using the linear parts of the curves (figure 2). The lipase specificity ratio (S) is theratio of the 6-O-monoesters’ initial rates (S = vi/vj). The energy difference in activation energy of the competingsubstrates (DDG#) was calculated using the equation: DDG# = – R*T* ln S (T = 318 K).
Lipase Initial rate Specificity ratio DDG#
vA vB vC A/B A/C B/C A/B A/C B/Cnmol/min,mg kcal/mol
Candida OF 0.6 0.7 0.5 0.9 1.2 1.4 0.1 –0.1 –0.2C. antarctica 7.0 40 32 0.2 0.2 1.3 1.1 1.0 –0.1M. miehei 6.8 1.2 0.7 5.6 9.6 1.7 –1.0 –1.4 –0.3P. cepacia 23 1.0 0.6 21 40 1.9 –1.9 –2.3 –0.4

methyl b-D-mannopyranoside. The initial rate for theformation of methyl 6-O-octanoyl-a-D-galactopyrano-side was also high.
The results of this investigation indicate that there is apotential for using the P. cepacia lipase as a biocatalystfor selective acylation of the galactose units at branchpoints in oligosaccharides derived from softwood galacto-glucomannan.
AcknowledgementThe lipases were generously given by NovoNordisk A/S,Denmark.
ReferencesAbe, Y., Harata, K., Fujiwara, M. and Ohbu, K. (1995) Carbohydr.
Res., 269, 43–51Ciuffreda, P., Ronchetti, F. and Tomma, L. (1990) J. Carbohydr.
Chem., 9, 125–129Colombo, D., Ronchetti, F., Scala, A. and Toma, L. (1992) J.
Carbohydr. Chem., 11, 89–94De Goede, A.T.J.W., Van Oosterom, M., Van Deurzen, M.P.J.,
Sheldon, R.A., Van Bekkum, H. and Van Rantwijk, F. (1994)Biocatalysis, 9, 145–155
Fersht, A. (1985), in: Enzyme Structure and Mechanism, secondedition, 111, New York: Freeman and Company.
Ljunger, G., Adlercreutz, P. and Mattiasson, B. (1994) Biotechnol.Lett., 16, 1167–1172
Therisod, M. and Klibanov A.M. (1986) J. Am. Chem. Soc., 108,5638–5640
Therisod, M. and Klibanov A.M. (1987) J. Am. Chem. Soc., 109,3977–3981
11
11
11
11
11
11
1
A. Córdova et al.
18 Biotechnology Letters · Vol 19 · No 1 · 1997
![BMC Structural Biology BioMed Central€¦ · Candida antarctica lipase B (CALB) is an efficient catalyst for hydrolysis in water and esterification in organic sol-vents [1]. It is](https://static.fdocuments.in/doc/165x107/60e4dfa2ae3684173126f4db/bmc-structural-biology-biomed-central-candida-antarctica-lipase-b-calb-is-an-efficient.jpg)

















