
Epigenetics In the 1940’s (Waddington et al.) The sum of the genes and their products and how they...
58
Epigenetics In the 1940’s (Waddington et al.) • The sum of the genes and their products and how they define a phenotype (dividing kidney and skin cells are identical in their DNA but give rise to cells of different phenotype). Today • Changes in gene function that are mitotically and/or meiotically heritable and that do not entail a change in DNA sequence. What does this mean?
-
date post
21-Dec-2015 -
Category
Documents
-
view
214 -
download
0
Transcript of Epigenetics In the 1940’s (Waddington et al.) The sum of the genes and their products and how they...

Epigenetics
In the 1940’s (Waddington et al.)• The sum of the genes and their products and how they define a phenotype (dividing kidney and skin cells are identical in their DNA but give rise to cells of different phenotype).
Today• Changes in gene function that are mitotically and/or meiotically heritable and that do not entail a change in DNA sequence.
What does this mean?

•• Production of genetically normal mosaic micefrom malignant teratocarcinoma cells (Mintzand Illmense 1975).•• Some carcinogens (tumor promoters, i.e. TPA) are not genotoxic.
Therefore, modification of gene expression, function due to developmental changes orexogenous factors could lead to cancer.
Evidence for Epigenetic Mechanisms of Carcinogenesis

Importance of Epigenetics
1. DNA methylation2. Imprinting3. X-chromosome inactivation4 Development/Reprogramming of somatic nucleus5. Cancer6. Rett syndrome1. DNA methylation7. Non-coding RNAs & heterochromatin

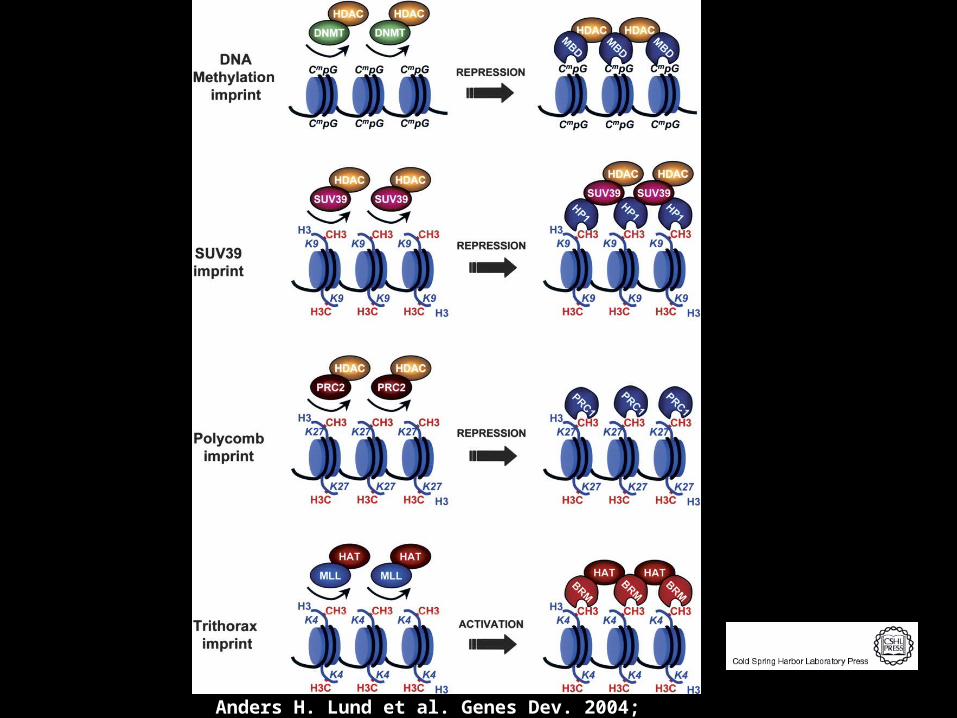
Anders H. Lund et al. Genes Dev. 2004; 18: 2315-2335

trx-GPc-G
Homeotic gene expression
Cloned Members of the Pc-G and trx-G
(PRE/TRE)
Pc M33/MPc1, MPc2, MPc3ph Rae28/Mph1Psc Mel18, Bmi-1Scm Scmh1, Scmh2Pcl M96Pho YY1Su(z)2 Mel18, Bmi-1dRING Ring1a, Ring1bE(z) Enx-1, -2/Ezh-1, -2esc eedAsx Asxl1, Asxl2crm
trx Mll, Mll2trl brm Brm, Brg1mo Baf155, Baf170osa Baf250snr1 Baf47/Snf5zash-1 Ash1lash-2 Ash2llidkislawc
PRC1
PRC2
SWI/SNF

Ying-Yang Theory
A natural homoestasis occurs between protein
isoforms with and without SET domains. This
balance is perturbed in human cancer development
by either the loss of the SET domain isoform or a busion gene lacking the SET domain.
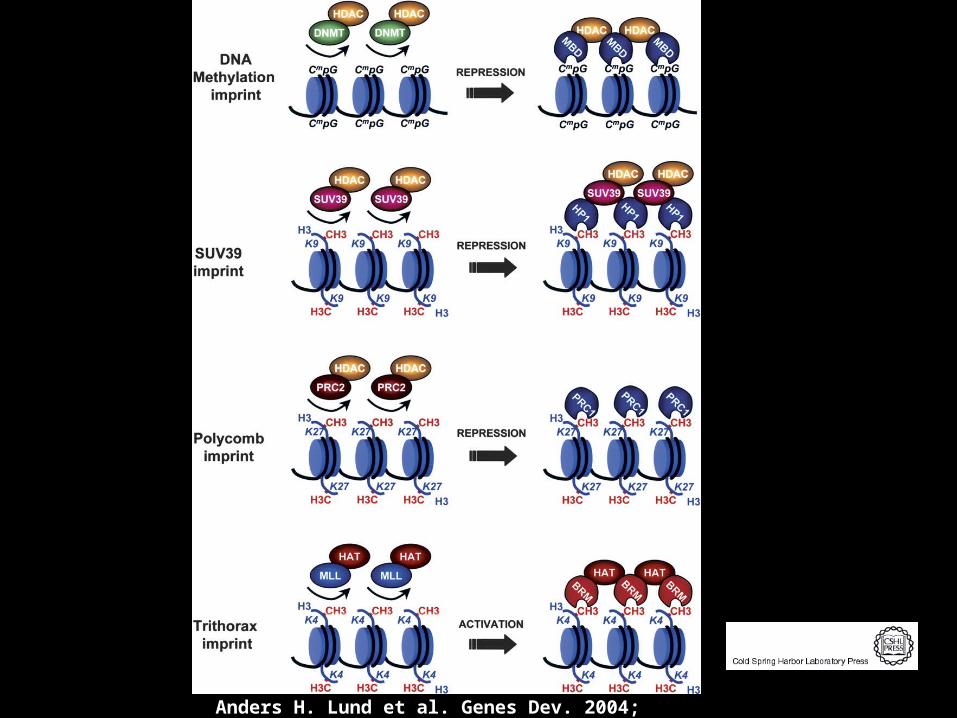
Anders H. Lund et al. Genes Dev. 2004; 18: 2315-2335

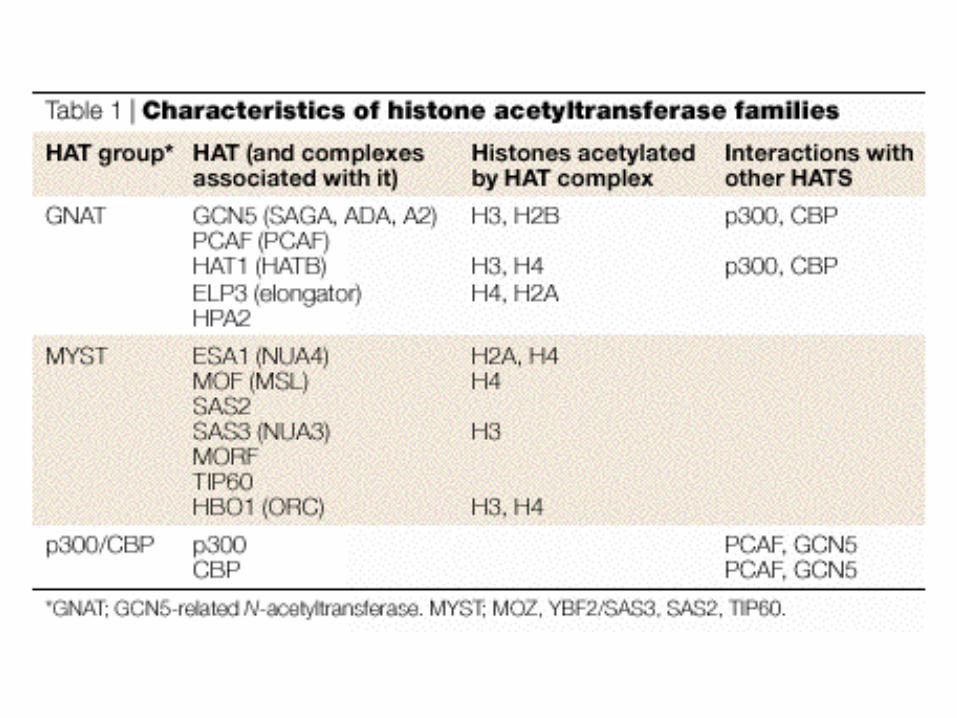
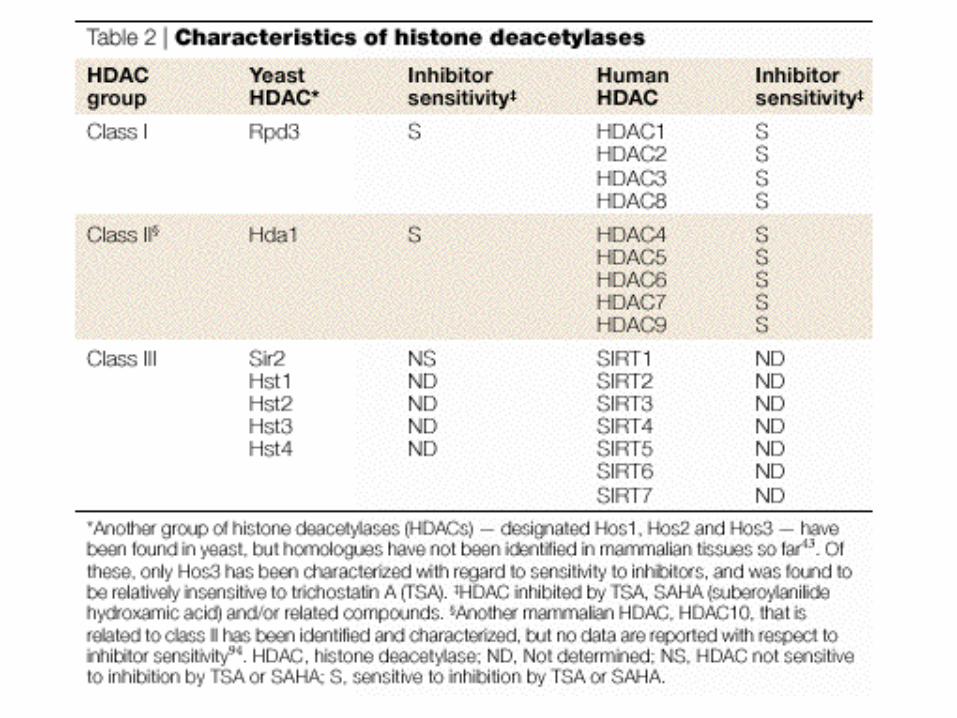
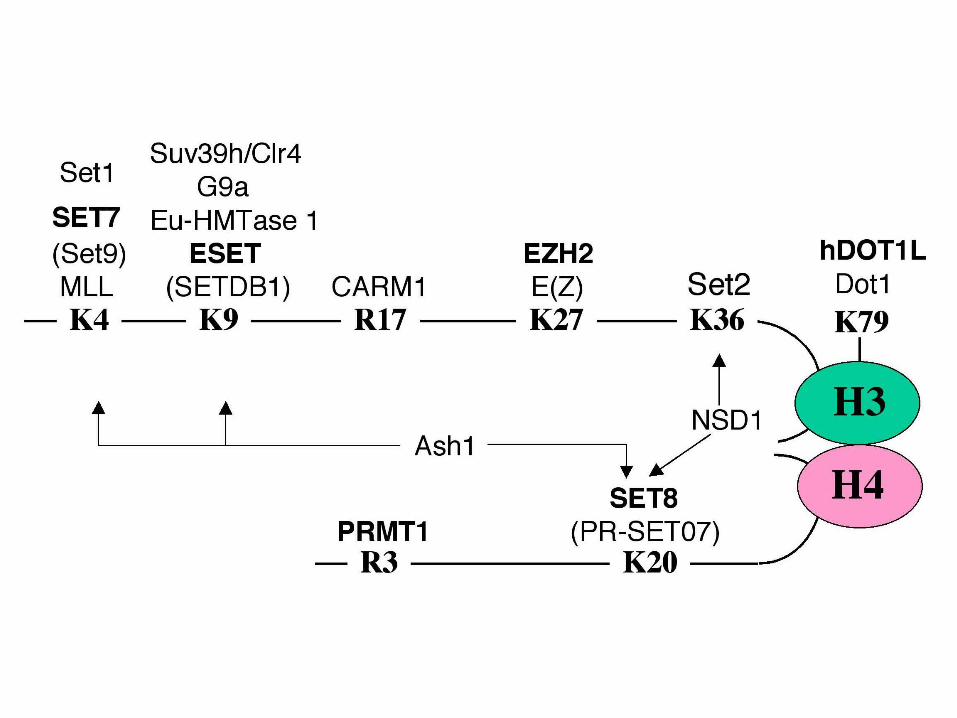

Disease Link to Chromatin (ref.)
Myeloid leukemias Translocation of the MOZ HAT (Smith, 1998)
Promyelocytic leukemias Fusion of a HDAC to transcription factors
(Grignani, 1998; Lin, 1998)
Acute Myeloid leukemia CBP-HRX fusion (Cairns, 1996)
Rubinstein-Taybi Syndrome CBP point mutations (Petrij, 1995)
Colorectal/gastric p300 missense mutations (Muraoka, 1996)
carcinomas
Breast cancer Amplification/Overexpression of
AIB1/HAT (Sobulo, 1997; Anzick, 1997)
Prostate cancer Overexpression of EZH2 (Varambally, 2002)

Anders H. Lund et al. Genes Dev. 2004; 18: 2315-2335

Characteristics of Hetero- and Eu-chromatin
Euchromatin Heterochromatin(constitutive)
Staining Dispersed Condensed (speckles)
DNA sequence Predominantly unique Repetitive & gene poor & gene rich
Replication Early to Late S-Phase Late S-Phase
Recombination Abundant Limited
Chromatin Irregular nucleosomes Regular nucleosomesAccessible to nucleases Less accessible to nucleasesHypersensitive Sites Few Hypersensitive Sites
Modifications H3-K4 methylation H3-K9 methylation Low CpG methylation More CpG methylation

DNA Methylation• In mammals, ~1% of DNA bases methylated on carbon-5 of cytosine pyrimidine ring (5-methylcytosine).• Most frequent at 5’-CpG-3’ dinucleotides (~70% of all CpGs).• In general, CpGs are under-represented (suppressed)
CpG islands• Regions (500-1 kb) of higher G+C than genome average.• Relatively devoid of methylation.• ~ 60% mammalian RNA Pol II promoters found in CpG islands.

DNA Methylation and Gene Expression
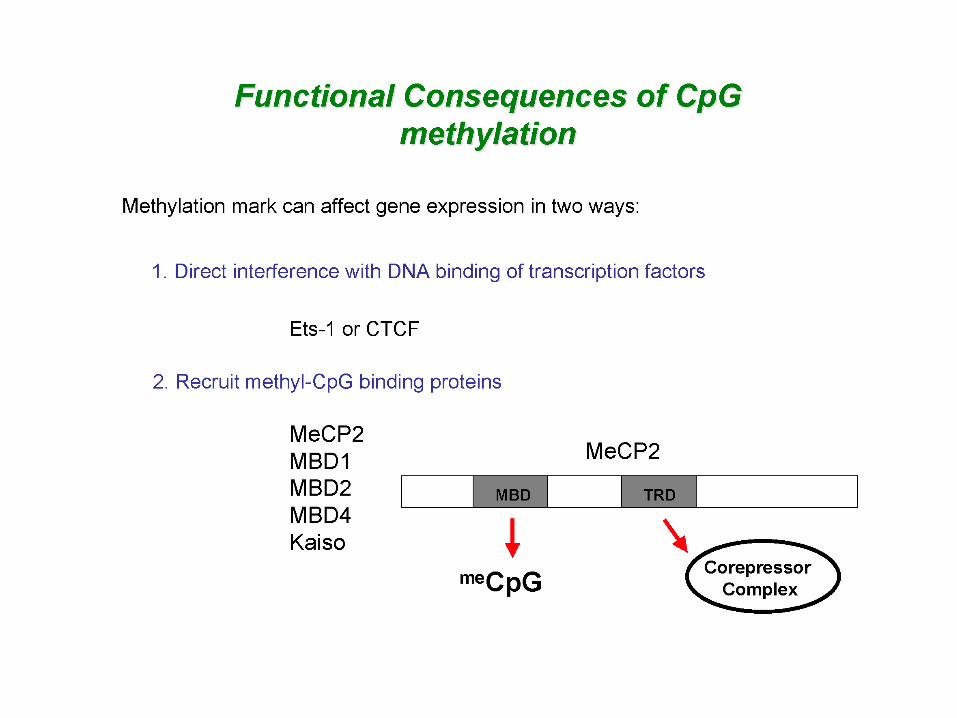
•• Cytosine residues in 5’CpG are often postsynthetically methylated•• CpG methylation is involved in long-term silencing of certain gene during development•• The methyl-CpG-binding proteins MeCP1 and MeCP2 interact specifically with methylated DNA and mediate transcriptional repression.




DNA Methylation and Cancer•• Hypomethylation
•• Human tumors normally exhibit decreased overall genome methylation
•• Cancer Res. 48:1159-1161, 1988; 64:4238-4243,2001•• Adv. Cancer Res. 54:1023,1990•• Gene. Cytogenet. 97:83-89,1987
•• Hypomethylation occurs relatively early in carcinogenesis
•• New Engl J./med. 319:525-532,1988.•• Cancer Res. 48:1159-1161, 1988•• Gut 39:434-438, 1996.

A significant reduction in 5- methylcytosinecontent in adenoma and adenocarcinomacompared with normal, paired colonicmucosa, whereas no difference existedbetween benign adenoma and malignantadenocarcinoma.
Feinberg A.P. et al. Cancer Res.48:1159-1161, 1988.
•• Hypermethylation•• Tumor Suppressor genes hypermethylated in human cancerCancer Res. 61:3225-3229,2001.

A Gene Hypermethylation Profile of Human CancerM. Esteller et al. Cancer Res.61:3225-3229, 2001.
Analyzed promoter hypermethylation changes in 12genes from 600 primary tumor samplesrepresenting 15 major tumors.
P16INK4a, P15INK4b - Cell cycle regulationBRCA1, hMLH1, MGMT - DNA repairTIMP3, CDH1, DAPK - Cell adherence, metastasisGSTP1 - Metabolic enzymeP14ARF, P73 - P53 networkAPC - β - catenin function

Principle of Bisulfite Modification
Ggg gcg gac cgc
Ggg gug gau ugu
Ggg gcmg gacm cmgcm
Ggg gcmg gacm cmgcm
Bisulfite modification
Bisulfite modification
Unmethylated DNA
Methylated DNA
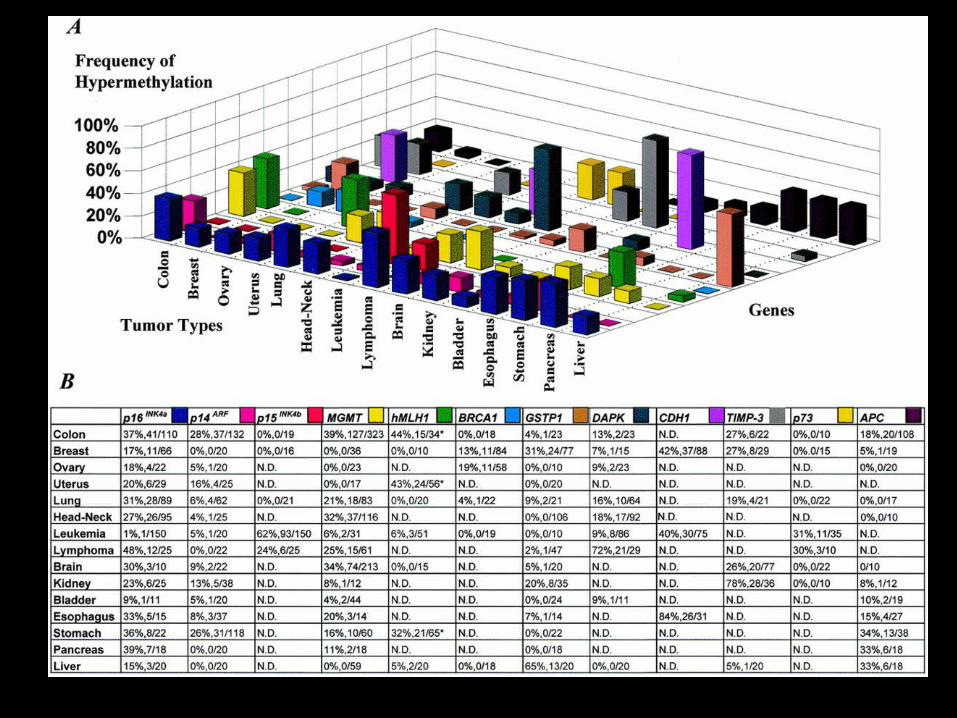

A unique profile of promoterhypermethylation may exist for humancancers where some gene changes are
shared and others are cancer-type specific:
•• P16 INK4a - Many tumor types•• APC - Liver, pancreas, stomach
•• P14 ARF - Colon, stomach•• GSTP - Liver, breast, prostate
•• MGMT - Colon, brain•• P15 INK4b - Leukemia

Simultaneous inactivation ofmultiple tumor suppressor/promoter
pathways may also occur:
Colon : P16 INK4a, hMLH1, TIMP3Breast : P16 INK4a, BRCA1, CDH1Lung : P16 INK4a, MGMT, DAPK

Sir2 (S.c.) =Clr4/Suv39 (S.p.,human)Sir3/4 (S.c.) =Swi6/HP1 (S.p.,human)

trx-GPc-G
Homeotic gene expression
Cloned Members of the Pc-G and trx-G
(PRE/TRE)
Pc M33/MPc1, MPc2, MPc3ph Rae28/Mph1Psc Mel18, Bmi-1Scm Scmh1, Scmh2Pcl M96Pho YY1Su(z)2 Mel18, Bmi-1dRING Ring1a, Ring1bE(z) Enx-1, -2/Ezh-1, -2esc eedAsx Asxl1, Asxl2crm
trx Mll, Mll2trl brm Brm, Brg1mo Baf155, Baf170osa Baf250snr1 Baf47/Snf5zash-1 Ash1lash-2 Ash2llidkislawc
PRC1
PRC2
SWI/SNF






Copyright ©1999 by the National Academy of Sciences
Toyota, Minoru et al. (1999) Proc. Natl. Acad. Sci. USA 96, 8681-8686


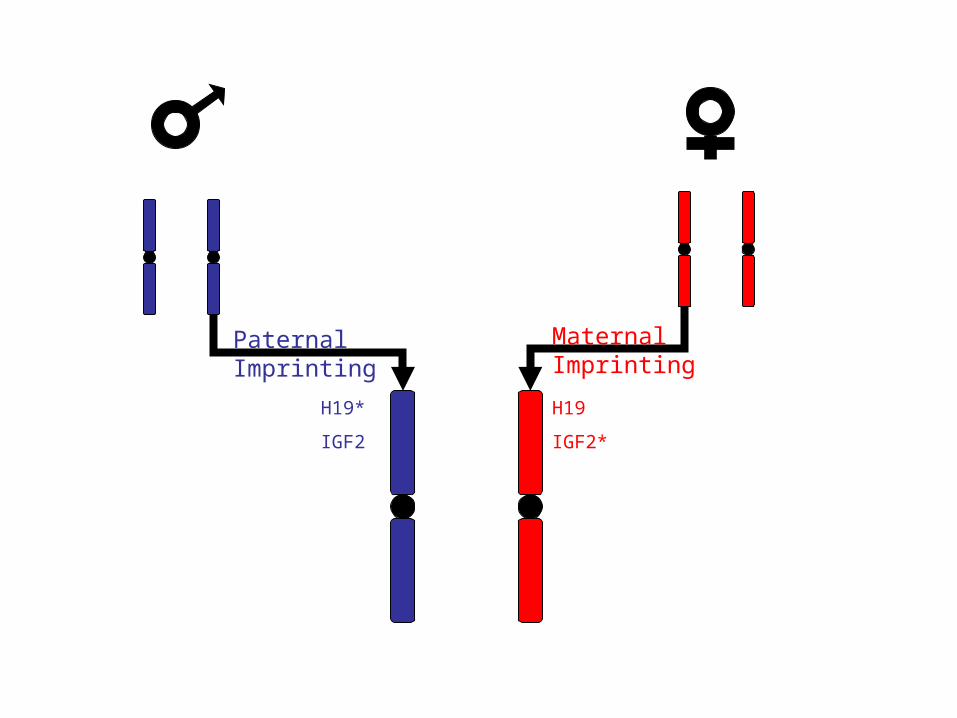
H19
IGF2*
H19*
IGF2
Paternal Imprinting
Maternal Imprinting

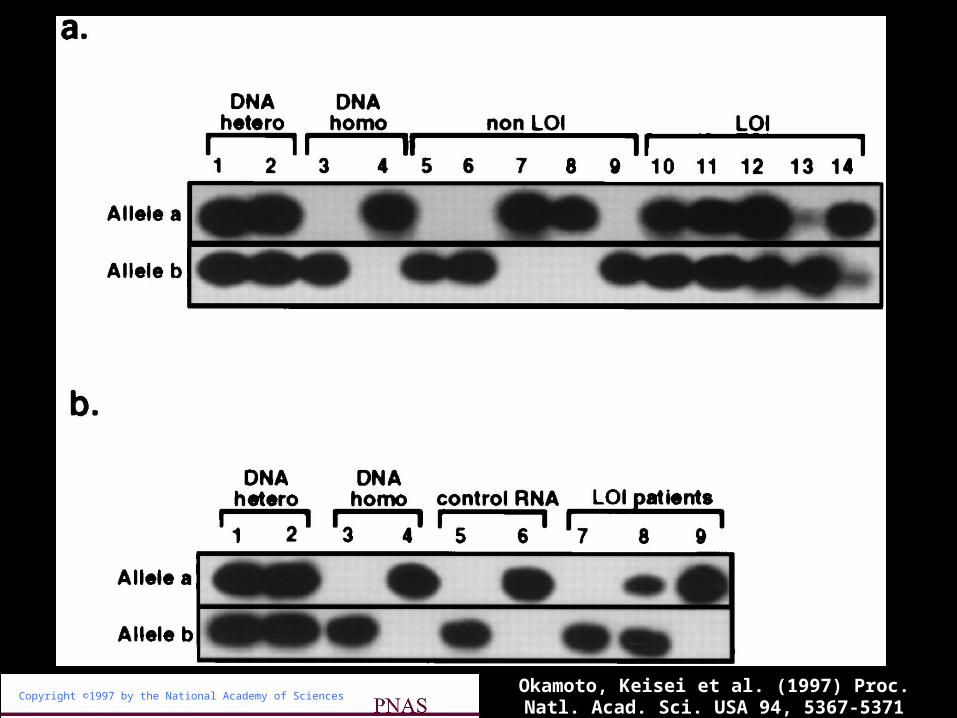
Copyright ©1997 by the National Academy of Sciences
Okamoto, Keisei et al. (1997) Proc. Natl. Acad. Sci. USA 94, 5367-5371

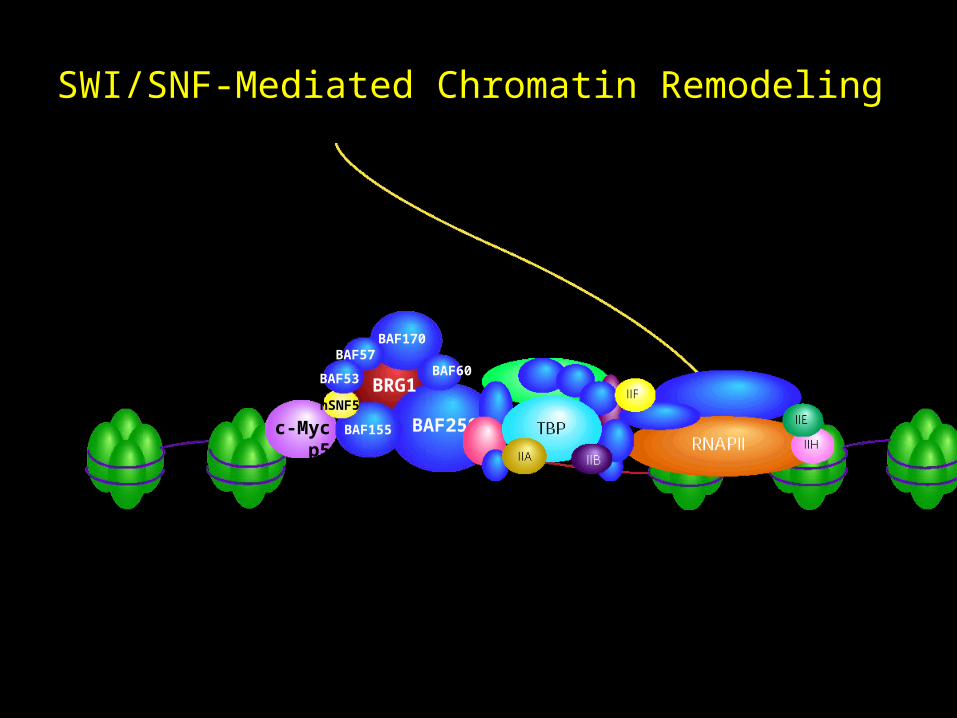
The SWI/SNF Complex
• ATP-dependent chromatin remodeling complex
• Yeast mating type switching (SWI) and sucrose non-
fermenting (SNF)
• Expression of 6% of yeast genes require SWI/SNF function
• 9-12 subunits - 2MDa
• Each subunit is required for function of the entire complex
• Evolutionarily conserved: C. elegans, yeast, drosophila, rat,
mouse, human
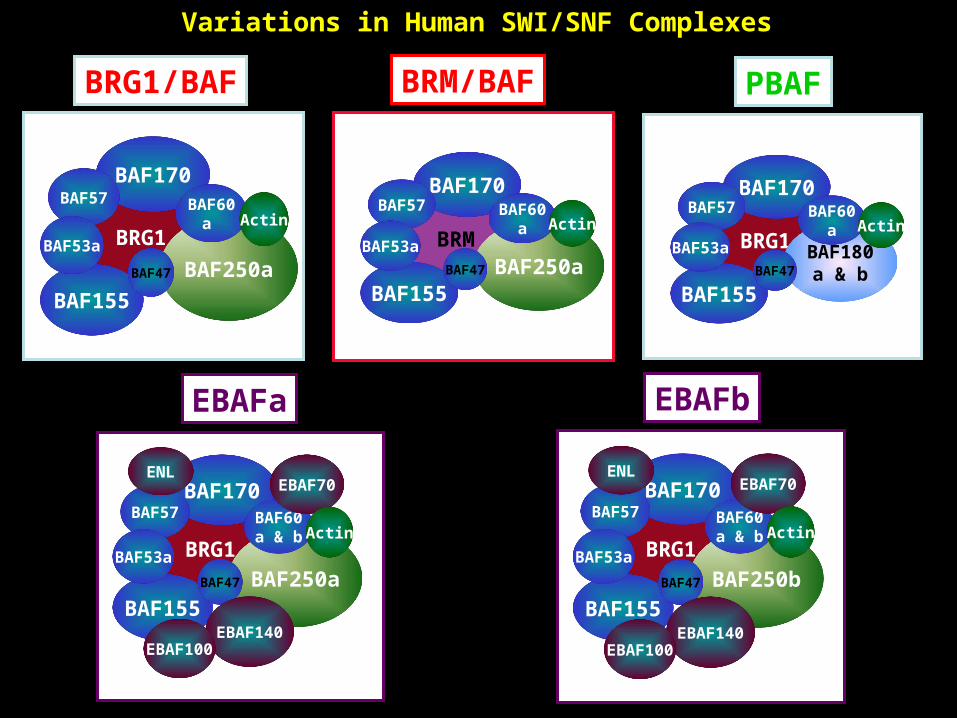
Variations in Human SWI/SNF Complexes
Actina
BRG1/BAF
BRG1
BAF155
BAF170BAF57
BAF53a
BAF250a
BAF60a
BAF47
Actin
PBAF
BRG1
BAF155
BAF170BAF57
BAF53a BAF180a & b
BAF60a
BAF47
Actin
BRM/BAF
BRM
BAF155
BAF170BAF57
BAF53aBAF250a
BAF60a
BAF47
Actin
BRG1
BAF155
BAF170BAF57
BAF53a
BAF250b
BAF60a & b
BAF47
EBAF70
EBAF140EBAF100
Actin
ENL
EBAFb
BRG1
BAF155
BAF170BAF57
BAF53a
BAF250a
BAF60a & b
BAF47
EBAF70
EBAF140EBAF100
Actin
ENL
EBAFa

BRG1
BAF250
BAF170
BAF60BAF57
BAF53
BAF155
hSNF5
c-Myc p53
SWI/SNF-Mediated Chromatin Remodeling

c-Myc p53
BRG1
BAF250
BAF170
BAF60BAF57
BAF53
BAF155
hSNF5
SWI/SNF-Mediated Chromatin Remodeling
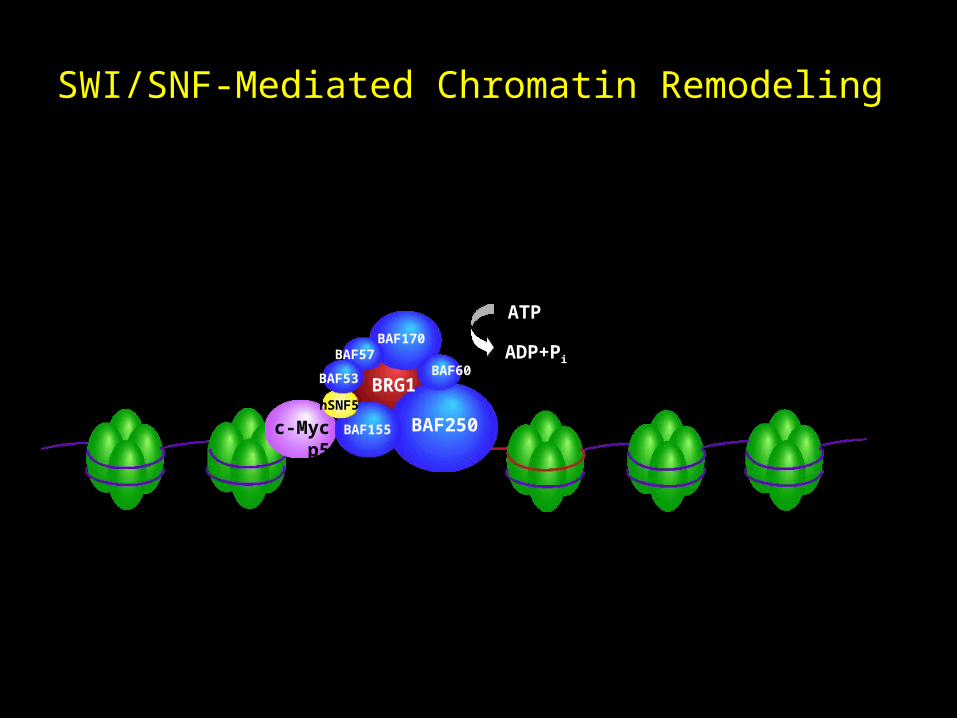
c-Myc p53
BRG1
BAF250
BAF170
BAF60BAF57
BAF53
BAF155
hSNF5
SWI/SNF-Mediated Chromatin Remodeling
ATP
ADP+Pi
c-Myc p53
BRG1
BAF250
BAF170
BAF60BAF57
BAF53
BAF155
hSNF5
SWI/SNF-Mediated Chromatin Remodeling


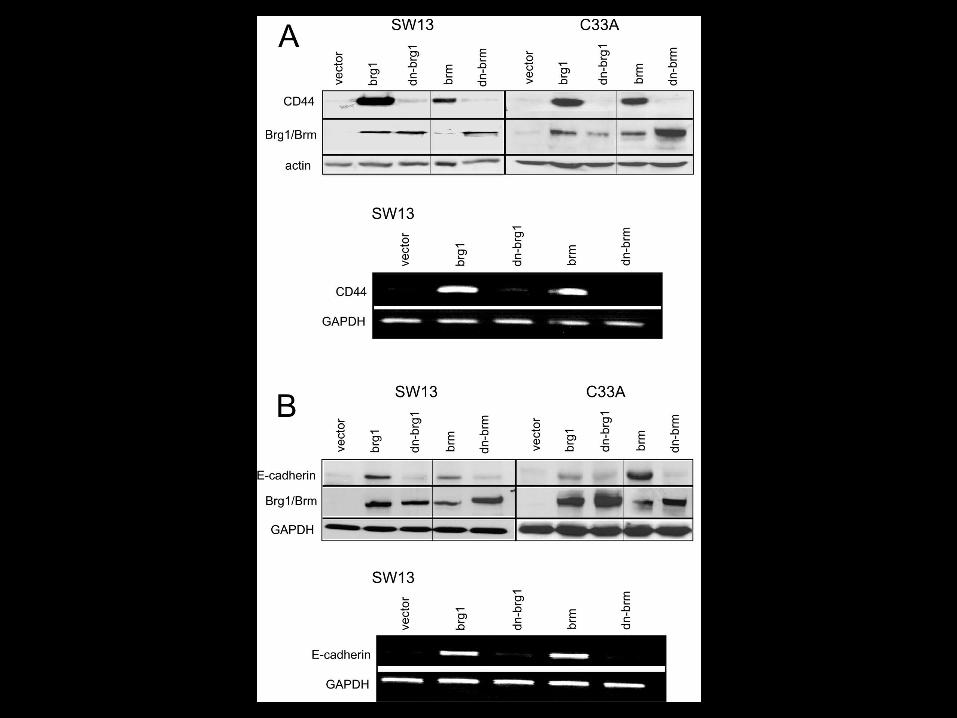
Evidence for BRG1’s Role in Human Tumor Development
- Required for RB-mediated growth arrest
- Mutations found in 10% of human tumor cell lines
- Expression lost in ~50% of NSCLC tumor cell lines, ~20% of adenocarcinoma cell lines & 10% of primary tumors.
- BRG1+/- mice develop adenocarcinomas
- LOH in region surrounding BRG1 locus (19p13.2)

How does loss of BRG1/BRM promote tumor development?
1. Abrogation of Rb mediated growth arrest?
2. Alterated DNA methylation patterns?
3. Aberrant cytoskeletal organization/signaling?


How does loss of BRG1/BRM promote tumor development?
1. Abrogation of Rb mediated growth arrest?
2. Alterated DNA methylation patterns?
3. Aberrant cytoskeletal organization/signaling?




Summary
• Chromatin structure plays a central role many cellularprocesses.• Relevant to most aspects of gene expression as well aschromosome stability, DNA replication, recombination and repair.• Chromatin regulated at local and global levels.• Misregulation of chromatin or DNA methylation states leads to cancer.• Chromatin is highly dynamic both in terms of structure &chemical composition.• CpG methylation and heterochromatin formation partition the genome.


















