
Enzymatic Adenylylation of and Spectinomycin by R-Factor ...iai.asm.org/content/1/1/109.full.pdf ·...
11
INFECrlON AND IMMUNITY, Jan. 1970, p. 109-119 Vol. 1, No. 1 Copyright © 1970 American Society for Microbiology Printed in U.S.A. Enzymatic Adenylylation of Streptomycin and Spectinomycin by R-Factor-Resistant Escherichia coli R. BENVENISTE, T. YAMADA, AND J. DAVIES Department of Biochemistry, University of Wisconsin, Madison, Wisconsin 53706 Received for publication 22 September 1969 A resistance (R) factor- containing strCain of Escherichia coli which is known to inactivate streptomycin by adenylylation has been shown to be spectinomycin resistant. An osmotic shockate of this strain catalyzes the formation of the bio- logically inactive spectinomycin adenylate, in which the adenylyl residue is probably attached to a D-threo methylamino alcohol moiety of spectinomycin. Both the streptomycin and spectinomycin adenylylating activities show the same temperature inactivation profile, and both are present in a protein fraction purified for the streptomycin inactivating enzyme. Mutants obtained from this strain which were sensitive to either spectinomycin or streptomycin were shown to lack both enzymatic activities when tested in vitro. Revertants of these mutants selected for recovery of either streptomycin resistance or spectinomycin resistance regain both activities. Therefore, we conclude that the inactivation of the two drugs is catalyzed by the same enzyme. Examination of a number of R factor-carrying strains has shown that those strains which are resistant to streptomycin and spectinomycin contain the adenylylating enzyme, whereas strains resistant to streptomycin but sensitive to spectinomycin inactivate streptomycin by phosphorylation. Resistance (R) factors confer antibiotic re- sistance to their host bacteria by a variety of mechanisms. The biochemical bases of R-factor mediated resistances to penicillin, chlorampheni- col, streptomycin, kanamycin, neomycin, and paromomycin have been established (19). With the exception of penicillin, which is hydrolyzed by ,B-lactamases (1), these drugs are inactivated by enzymatic modification of the antibiotic, such as acetylation, phosphorylation, or adenylylation in the R factor-carrying strain. Resistance to sulphonamides and tetracyclines is also deter- mined by R factors, but little is known of the resistance mechanism in these cases. During the course of our recent studies on the inactivation of streptomycin by adenylylation (20), it was found that the bacterial strain used was also resistant to spectinomycin, a recently discovered aminoglycoside antibiotic. R factor- mediated spectinomycin resistance has previously been reported by Smith (15). We wish to report that, like streptomycin, spectinomycin is in- activated by adenylylation (2; R. Benveniste et al., Bacteriol. Proc., p. 48, 1969). MATERIALS AND METHODS Strains. Table 1 contains a list of the Escherichia coli R factor-carrying strains used. Preparation of cell-free extracts. The cells were grown at 37 C in a medium containing the following (per liter): dextrose, 6.7 g; peptone, 8.0 g, MgSO4, 0.3 g; NH4Cl, 1 g; KH2PO4, 3 g; Na2HPO4, 5.8 g; thiamine, 3 mg (pH 7.0). They were harvested in late logarithmic phase of growth and osmotically shocked using a modification of the procedure described by Nossal and Heppel (9). The cells were washed twice at room temperature in 12 volumes of 0.01 M tris- (hydroxymethyl)aminomethane (Tris) and 0.03 M NaCl, pH 7.8. The pellet was suspended in 36 volumes of 20% sucrose, 3 X 10-3 M ethylenediaminetetra- acetic acid (EDTA), and 0.033 M Tris-chloride (pH 7.8), stirred for 15 to 30 min at room temperature, and centrifuged at 16,000 X g for 15 min. The pellet was then suspended in 24 volumes of cold 5 X 10-4 M MgCI2 and stirred vigorously for 15-30 min at 2 to 5 C. The supernatant fluid of a 30-min, 26,000 X g cen- trifugation is referred to as the osmotic shockate. Enzymatic assay. The assay is modified slightly from that previously described (20). The reaction mixture contained osmotic shockate or partially purified enzyme [streptomycin (and spectinomycin) adenylate synthetase], 4.25 nmoles of '4C-adenosine 5'-triphos- phate [(ATP) specific activity, 4.3 ,uc/,umole], 4 nmoles of streptomycin or spectinomycin sulfate, 2.5 ,Amoles of Tris-chloride (pH 8.0 at 30 C), 0.1 jAmole of MgC12, and 0.2 ,umoles of 0-mercaptoethanol in a total volume of 20 ,Aliters. Incubation was at 30 C for various periods of time (1 to 120 min), after which 10 ,lAiters was pipetted onto a 0.75-cm square of phosphocellu- 109 on November 12, 2018 by guest http://iai.asm.org/ Downloaded from
Transcript of Enzymatic Adenylylation of and Spectinomycin by R-Factor ...iai.asm.org/content/1/1/109.full.pdf ·...

INFECrlON AND IMMUNITY, Jan. 1970, p. 109-119 Vol. 1, No. 1Copyright © 1970 American Society for Microbiology Printed in U.S.A.
Enzymatic Adenylylation of Streptomycin andSpectinomycin by R-Factor-Resistant
Escherichia coliR. BENVENISTE, T. YAMADA, AND J. DAVIES
Department of Biochemistry, University of Wisconsin, Madison, Wisconsin 53706
Received for publication 22 September 1969
A resistance (R) factor- containing strCain of Escherichia coli which is known toinactivate streptomycin by adenylylation has been shown to be spectinomycinresistant. An osmotic shockate of this strain catalyzes the formation of the bio-logically inactive spectinomycin adenylate, in which the adenylyl residue is probablyattached to a D-threo methylamino alcohol moiety of spectinomycin. Both thestreptomycin and spectinomycin adenylylating activities show the same temperatureinactivation profile, and both are present in a protein fraction purified for thestreptomycin inactivating enzyme. Mutants obtained from this strain which weresensitive to either spectinomycin or streptomycin were shown to lack both enzymaticactivities when tested in vitro. Revertants of these mutants selected for recovery ofeither streptomycin resistance or spectinomycin resistance regain both activities.Therefore, we conclude that the inactivation of the two drugs is catalyzed by thesame enzyme. Examination of a number of R factor-carrying strains has shown thatthose strains which are resistant to streptomycin and spectinomycin contain theadenylylating enzyme, whereas strains resistant to streptomycin but sensitive tospectinomycin inactivate streptomycin by phosphorylation.
Resistance (R) factors confer antibiotic re-sistance to their host bacteria by a variety ofmechanisms. The biochemical bases of R-factormediated resistances to penicillin, chlorampheni-col, streptomycin, kanamycin, neomycin, andparomomycin have been established (19). Withthe exception of penicillin, which is hydrolyzedby ,B-lactamases (1), these drugs are inactivatedby enzymatic modification of the antibiotic, suchas acetylation, phosphorylation, or adenylylationin the R factor-carrying strain. Resistance tosulphonamides and tetracyclines is also deter-mined by R factors, but little is known of theresistance mechanism in these cases.During the course of our recent studies on the
inactivation of streptomycin by adenylylation(20), it was found that the bacterial strain usedwas also resistant to spectinomycin, a recentlydiscovered aminoglycoside antibiotic. R factor-mediated spectinomycin resistance has previouslybeen reported by Smith (15). We wish to reportthat, like streptomycin, spectinomycin is in-activated by adenylylation (2; R. Benveniste et al.,Bacteriol. Proc., p. 48, 1969).
MATERIALS AND METHODSStrains. Table 1 contains a list of the Escherichia
coli R factor-carrying strains used.
Preparation of cell-free extracts. The cells weregrown at 37 C in a medium containing the following(per liter): dextrose, 6.7 g; peptone, 8.0 g, MgSO4,0.3 g; NH4Cl, 1 g; KH2PO4, 3 g; Na2HPO4, 5.8 g;thiamine, 3 mg (pH 7.0). They were harvested in latelogarithmic phase of growth and osmotically shockedusing a modification of the procedure described byNossal and Heppel (9). The cells were washed twiceat room temperature in 12 volumes of 0.01 M tris-(hydroxymethyl)aminomethane (Tris) and 0.03 MNaCl, pH 7.8. The pellet was suspended in 36 volumesof 20% sucrose, 3 X 10-3 M ethylenediaminetetra-acetic acid (EDTA), and 0.033 M Tris-chloride (pH7.8), stirred for 15 to 30 min at room temperature,and centrifuged at 16,000 X g for 15 min. The pelletwas then suspended in 24 volumes of cold 5 X 10-4 MMgCI2 and stirred vigorously for 15-30 min at 2 to 5 C.The supernatant fluid of a 30-min, 26,000 X g cen-trifugation is referred to as the osmotic shockate.
Enzymatic assay. The assay is modified slightly fromthat previously described (20). The reaction mixturecontained osmotic shockate or partially purifiedenzyme [streptomycin (and spectinomycin) adenylatesynthetase], 4.25 nmoles of '4C-adenosine 5'-triphos-phate [(ATP) specific activity, 4.3 ,uc/,umole], 4 nmolesof streptomycin or spectinomycin sulfate, 2.5 ,Amolesof Tris-chloride (pH 8.0 at 30 C), 0.1 jAmole of MgC12,and 0.2 ,umoles of 0-mercaptoethanol in a total volumeof 20 ,Aliters. Incubation was at 30 C for variousperiods of time (1 to 120 min), after which 10 ,lAiterswas pipetted onto a 0.75-cm square of phosphocellu-
109
on Novem
ber 12, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

BENVENISTE, YAMADA, AND DAVIES
TABLE 1. R-factor strains examined
Host
w4354 (met- B1- F-)K12-w677bK12-w677K12-w677K12-w677K12-w677K12-w677K12-w677K12-w677K12-w677K12-CSH-2 (met- Bg- F-)K12-CSH-2K12-CSH-2K12-CSH-2K12-CSH-2
Antibiotic resistancesa
KanA Str Spc Tet
++
Cam
+
I+
Amp
_-
a KanA, kanamycin A (10 ,ug-disc); Str, streptomycin (20 ,g-disc); Spc, spectinomycin (20 jig-disc);Tet, tetracycline (5 jig-disc); Cam, chloramphenicol (30,ug-disc); Amp, ampicillin (10 ,jg-disc).
b (thr- leui Br- lacr- mall- su- gal7 gal.- xylr- aral- muli Tlr P-).r Inactivation of kanamycin in this strain is believed to be by acetylation, since the strain is resistant
to KanA but sensitive to KanB (10-,ug disc) (10, 11).d Inactivation of kanamycin in this strain is by phosphorylation (5, 12, 13a, 17).
lose paper (Whatman P-81). The squares were allowedto dry for 15 sec and were then immersed in hot,distilled water (70 to 80 C) for 2 min to stop the reac-tion. They were washed several times with largevolumes of distilled water, dried, and counted in aTri-Carb scintillation spectrometer (Packard Instru-ment Co., Inc., Downers Grove, Ill.) using a toluene-based fluid. For initial velocity measurements, thereaction was terminated rapidly by heating at 90 Cfor 1 min, pipetting the mixture onto phosphocellulosepaper squares, and proceeding as described above.Control reactions for nonspecific binding of '4C-ATPto the paper were run in the absence of either enzymeor antibiotic.One unit of enzymatic activity is defined as that
amount of enzyme producing 1 nmole of streptomycinor spectinomycin adenylate per min at 30 C under theabove assay conditions. The radioactivity as 14C-adenosine monophosphate (AMP) bound to thedrug, which is adsorbed on the phosphocellulosepaper, is proportional to the amount of enzyme addedto the assay mixture within the range of 0.10 to 11units/mi. Protein was determined by the method ofLowry et al. (7).
Purification of the adenylylating enzyme. Approxi-mately 1 liter of osmotic shockate was obtained froma 15-liter culture (45 g) of NR73/w677. All steps werecarried out at 0 to 5 C. The osmotic shockate wasmade 0.03 M in Tris-chloride (pH 8.1 at 5 C), con-centrated by (NH4)2SO4 precipitation (700 g/liter),and dialyzed versus 0.03 M Tris-chloride (pH 8.1),10-2 M MgCl2, and 6 X 10-4 M ,-mercaptoethanol.
Nucleic acids were precipitated by the addition ofstreptomycin to a final concentration of 1.5%. The
suspension was centrifuged at 25,000 X g for 20 minand the pellet was discarded.The resulting supernatant fluid was fractionated
with solid (NH4)2SO4, and the precipitate obtainedfrom a 176 to 313 g/liter fraction was resuspended in0.005 M Tris-chloride (pH 8.1), 10-2 M MgC12, 6 X104 M 3-mercaptoethyl alcohol (buffer A) and dialyzedagainst the same buffer.
This dialyzed ammonium sulfate fraction wasapplied to a column of Whatman diethylaminoethylcellulose [(DEAE 32, 1 meq/g) 3 by 23 cm, previouslyequilibrated with buffer A] and eluted with a lineargradient of NaCl from 0.05 M (600 ml) to 0.4M(600 ml) in buffer A. Fractions from the column were
assayed for adenylylating activity as previously de-scribed. The enzyme eluted at 0.25 M NaCl.The peak fractions were collected, concentrated
against polyethylene glycol, dialyzed against buffer A,and placed on a column (2 by 50 cm) of P-200 (Bio-Rad, 50 to 100 mesh) which had been previouslyequilibrated with buffer A. The column was elutedwith the same buffer; the peak of enzymatic activitywas eluted in four void volumes.
Isolation of mutants lacking both enzymatic activi-ties. E. coli JE 254 was treated with ethyl methanesulfonate according to the procedure described byOsborn et al. (13), with the exception that phosphatebuffer (pH 7.0) was used in place of Tris buffer. Thecells were then grown to early logarithmic stage inPenassay broth containing 50 ,Ag of spectinomycinper ml, at which time 250 JAg of ampicillin per ml(Bristol-polycillin-N) was added, and growth was con-
tinued overnight. After plating on antibiotic-freemedium, individual colonies were picked and replica
R factor
R100 (JE254)R100NR84NR85NR86NR73NR79NR1-22NR72NR71NR1NR2-2NR4NR3NR45JR35
110 INFEC. IMMUN.
on Novem
ber 12, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

VOL. 1, 1970 ADENYLYLATION OF STREPTOMYCIN AND SPECTINOMYCIN
plated onto spectinomycin (50 ,g/ml), tetracycline(20 og/ml), streptomycin (20 JAg/ml), and chloram-phenicol (20 ,g/mi) agar plates to score for mutantswhich had lost streptomycin and spectinomycinresistance while retaining tetracycline and chloram-phenicol resistance.
In vitro polypeptide synthesis. Tests of the effect ofdrugs on polynucleotide-directed polypeptide syn-thesis were carried out essentially as described byDavies et al. (3), using an incorporation system withribosomes obtained from a spectinomycin-sensitivestrain.
Electrophoresis. Four Aliters of a 20-,uliter reactionmixture containing 14C-ATP (10 ,Ac/lAmole) werespotted on Sepraphore III cellulose polyacetateelectrophoresis strips (2.54 by 30.48 cm); (GelmanInstrument Co., Ann Arbor, Mich.). The strips weredeveloped in a water-pyridine-glacial acetic acid buffer(400:3:10; pH 3.9) for 2 to 4 hr at 1 ma per strip, andthe radioactivity was monitored with a 7201 Radio-chromatogram scanner (Packard Instrument Co.,Inc., Downers Grove, Ill.).
Electrophoresis of enzymatic reaction products.3H-dihydrospectinomycin was purified by elec-trophoresis on Sepraphore cellulose polyacetate stripsas described above and was reacted with 14C-ATP(413 juc/;imole), purified adenylylating enzyme, andother reaction components. Portions of this reactionmixture were spotted onto Sepraphore strips and runfor 3 hr at 1 ma per strip. Known amounts of 14C-ATPand 3H-dihydrospectinomycin were run separately ascontrols. The strips were cut in 0.5-cm lengths andcounted in a scintillation counter.
Chemicals. 14C-labeled ATP was obtained fromNew England Nuclear Corp., Boston, Mass., a_-32P-ATP from Schwarz BioResearch, Inc., Orangeburg,N.Y., and ?-y-P-ATP from the laboratory of H. G.Khorana. Streptomycin was purchased from Calbio-chem, Los Angeles, Calif. 3H-dihydrospectinomycinwas prepared by one of us (J. D.) by reduction ofspectinomycin with 3H-sodium borohydride. Di-desamidino-dihydrostreptomycin was the gift of L.Anderson, methyl-2-deoxy-2-methylamino-a/13-L-glu-copyranosyl moiety of streptomycin that of C. B.Barlow, N-methyl dihydrostreptomycin that of H.Vanderhaeghe, and 1, 3-diamino-1, 3-dideoxy-myo-inositol that of T. Shier. Spectinomycin, actinamine,methyl streptobiosaminide, streptamine, and deoxy-streptamine were provided by G. B. Whitfield of theUpjohn Co.
RESULTS
Localization of inactivating enzyme in theperiplasmic region of the cell. We have shown, inthe case of streptomycin, that the adenylylatingactivity is released by cold-water shocking ofsucrose-Tris-EDTA-treated cells according tothe methods of Nossal and Heppel (9), and there-fore can be classified as a periplasmic enzyme.This osmotic shock treatment released both thestreptomycin and spectinomycin adenylylatingactivity from the cells (Table 2). In addition, we
TABLE 2. Quantitative release ofenzymatic activityby osmotic shocking of JE 254a
Total enzyme unitsCell treatment
Streptomycin Spectinomycin
Osmotic shock........... 60 33.5Sonically treated, shocked
cellsb.. .......... 1.2 1.1Sonically treated cellsc.. 58.4 36
a Assay was performed as described in Materialsand Methods.
6 Cells (3.2 g) which had been recovered follow-ing osmotic shocking in 5 X 10-4 M MgCl2 (70 ml)were sonically treated in 20 ml of buffer A andcentrifuged to remove cell debris; the supernatantfluid was assayed for enzymatic activity.
c Cells (3.2g) were sonically treated and cen-
trifuged; the supernatant fluid was assayed foradenylylating activity.
11
10
9
8
7
6
W 4
E 3
2-
1
20 40 60 80 100 120 140 160
Minutes of Incubation
FIG. 1. Adenylylation of spectinomycin by a 95-foldpurified enzyme preparation of NR73/w677. At thetimes indicated, 10 ,uliters of a reaction mixture (con-taining 11 nmoles of spectinomycin and 15 nmoles ofATP) was pipetted onto squares of phosphocellulosepaper, which were heated, washed, dried, and countedfor radioactivity as described in Materials and MethodsSymbols: 0, nmoles of Spc-AMP; 0, a reaction run
in the absence of either spectinomycin or enzyme andassayed at the times indicated.
111
on Novem
ber 12, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

BENVENISTE, YAMADA, AND DAVIES
have found that these activities are synthesizedconstitutively, since identical levels of activitywere found in R factor-carrying cells grown inthe presence or in the absence of the drug (20).
Reaction between spectinomycin and ATP. Theformation of streptomycin adenylate can befollowed conveniently by measuring the transferof radioactivity from '4C-ATP or a-'2P-ATP tocation exchange paper (carboxymethyl or phos-phocellulose paper, preferably the latter) in thepresence of streptomycin and the adenylylatingenzyme (20). When streptomycin was replacedby spectinomycin, the same reaction occurred(Fig. 1). No radioactive compound capable ofbinding to phosphocellulose paper was obtainedwhen y-2P-ATP was used. By analogy with theinactivation of streptomycin, this strongly sug-gested that spectinomycin adenylate was beingformed.
Cellulose acetate strip electrophoresis of en-zymatic reaction products. The inactivation of3H-dihydrospectinomycin was carried out in thepresence of '4C-ATP and adenylylating enzyme,and the reaction mixture was electrophoresedas previously described. Under the conditionsused, ATP, adenosine diphosphate, and AMPdo not migrate very far from the origin, whereasdihydrospectinomycin runs toward the cathode.Figure 2 shows that, in addition to the slow run-ning peak of ATP and the fast running peak of
0 1,60
()II
E -112[d
o~~~~~~~~~~~~~~~I I
r~ ~ i
1013
FIG. 2. Electzymatic inactiv,14C-ATP. PurifiATP were reactoenzyme preparastrip, and electand Methods. T)The strip was acounter; the nu
mycin and 14C.moles of 3H-diATP.
STREPTOBIOSAMINE
ISTREPTOSE
N-METHYL-L-GLCOSAMINE
STREPTOMYCIN: R=R1=-NHC R2=-CHONH2
1;NHDIHYDROSTREPTOMYCIN: R= RI=-NHC " , R2=-CH2OHBLUENSOMYIN: R2= -CH2OH
'NH ORR =-NHC , Rg=-OCONH2 VICE
NH2 VERSA
FIG. 3. Arrow points to the OH group on theN-methyl-L-glucosamine residue of streptomycin thathas been shown to be enzymatically adenylylated (18,20). In N-benzyl streptomycylamine, the aldehydegroup ofstreptomycin is reductively coupled to benzyla-mine.
FIG. 4. Structure of spectinomycin. Arrow indicatesthe probable site of adenylylation on the actinaminemoiety.
\/11 \ . ' unchanged dihydrospectinomycin, there is a
; X, L peak running just behind dihydrospectinomycin3 6 t 9 12 is is which contains both 14C and 3H in an equimolar
origin cm ratio, and which is produced only in the presenceIrophoresis of the products of the en- of enzyme. The 14C and 3H counts in the peakation of 3H-dihydrospectinomycin with increased with increasing time of incubation,led 'H-dihydrospectinomycin and 14C- suggesting that a compound of dihydrospectino-led in the presence of a 90-fold purified mycin and adenosine is produced in the enzymaticition, spotted on a cellulose polyacetate reaction. When the peak containing 14C-AMPfrophoresed as described in Materials and 8H-dihydrospectinomycin was eluted and
he cathode lies to the right in the figure.fried, cut, and counted in a scintillation Incubated with snake venom phosphodiesterase
mber of moles of 8H-dihydrospectino- and re-electrophoresed, the "C-AMP and 'H-di-
-ATP were determined. Symbols: * hydrospectinomycin counts were separated.hydrospectinomycin; 0, moles of 14C' Treatment with alkaline phosphatase had no
effect.
112 INFEC. IMMUN.
STREPTOMYCIN ANTIBIOTICS
on Novem
ber 12, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

VOL. 1, 1970 ADENYLYLATION OF STREPTOMYCIN AND SPECTINOMYCIN
3-30
a.
//PECTINOMYCIN
0 20 40 60MINUTES
FIG. 5. Adenylylation of equimolar amounts ofspectinomycin and actinamine. A 40-fold purifiedenzyme was used. See legend of Fig. 1.
Nature of the attachment of the adenylyl residueof streptomycin and spectinomycin. We havepreviously shown that streptomycin is adenyl-ylated on the 3'-OH of the 2-deoxy-2-methyl-amino - L - glucopyranosyl(N - methyl - L - glucosa-mine) residue (20; Fig. 3). If one examines thestructure of spectinomycin as shown in Fig. 4(14), it is clear that there is a D-threo methylaminoalcohol moiety in actinamine with the samestereochemistry as in the N-methyl-L-glucosamineresidue of streptomycin. In the likelihood thatthis particular moiety in spectinomycin was thesubstrate for the adenylylating enzyme, we testedactinamine in the phosphocellulose-paper bind-ing assay and found it to be as effective a sub-strate as spectinomycin (Fig. 5). Thus, one of thehydroxyl groups of actinamine is the site ofattachment of the AMP residue, and we suspectit to be the hydroxyl group indicated by thearrow in Fig. 4 (see Discussion).
Biological activity of inactivated spectinomycin.Samples of spectinomycin adenylate sufficientfor most biological assays have proved difficultto obtain, since spectinomycin, being a somewhatlabile compound, is degraded by prolonged incu-bation at 30 C and pH 8.0 (the conditions forenzymatic inactivation). However, as polypeptidesynthesis in vitro is very sensitive to low con-centrations of the drug, this system was used as atest for the biological activity of spectinomycinadenylate. Spectinomycin-AMP is not effectiveas an inhibitor of polypeptide synthesis (Table 3).
Thermosensitivity of the enzymatic activity. Ifstreptomycin and spectinomycin adenylate werethe products of two enzymes which had not beenseparated by the chromatographic purificationslisted in Table 5, then it might be expected thatthe two reactions would show different heat
TABLE 3. Effect of inactivated spectinomycin onpolypeptide synthesis in vitro
Per centIncubation mixture incorpor-
ation
Complete system.Plus 0.15 ,ug of spectinomycin per 50 ,Aliters of
incubation mixture; preincubation withoutinactivating enzyme..
Plus 0.15 ug of spectinomycin per 50 ,uliters ofincubation mixture; preincubation with in-activatingenzyme.
100
32
99
a Preincubation with inactivating enzyme wasfor 40 min at 30 C. The preincubation mixture con-tained the complete system minus ribosomes andR17 RNA, but with added inactivating enzyme.After 40 min, ribosomes and R17 RNA were addedand the complete system was incubated at 37 C for60 min, precipitated with trichloroacetic acid,filtered, and counted.
inactivation kinetics. An enzyme purified 40-foldfrom an osmotic shockate of strain JE 254 wasdivided into small samples, heated for 5 min attemperatures between 30 and 50 C, and used inthe standard assay system with either strepto-mycin or spectinomycin as substrate. The heatinactivation of both activities followed the samekinetics, as would be expected if the same enzymeperformed both adenylylation reactions (Fig. 6).The heat sensitivity of the adenylylating enzymefrom strain JE 254 in vitro is possibly reflected inthe resistance properties of the strain, since itcan tolerate higher concentrations of streptomycinat 30 C than at 42 C in liquid media.
Mutants with coincident loss of streptomycinand spectinomycin adenylylating activities, andrevertants to both activities. We have obtained 15independently isolated spectinomycin-sensitivemutants of a spectinomycin-resistant strain(JE 254) following mutagenesis and penicillinenrichment, and find that they become con-comitantly streptomycin sensitive. In addition,several temperature-sensitive mutants (includingone obtained from R. Rownd) have been ob-tained which are streptomycin- and spectinomy-cin-resistant at 30 C but streptomycin- andspectinomycin-sensitive at 42 C. In all mutants sofar examined, no adenylylating activity for strep-tomycin and spectinomycin could be detected inosmotic shockates or extracts of the sensitivemutants.
Streptomycin- and spectinomycin-resistant re-vertants were selected from these sensitive mu-tants by plating on media containing streptomycinor spectinomycin. When the resulting resistantclones were tested for resistance to both drugs,it was found that the majority had reverted to the
113
on Novem
ber 12, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

BENVENISTE, YAMADA, AND DAVIES
z
I ~Io I> 40-
~20-
30 35 40 45 54T (OC)
FIG. 6. Heat inactivation of streptomycin andspectinomycin adenylylating activities. Fifty pliters of40-fold purified enzyme (1.0 mg/mI) in 5 mM Tris-chloride, (pH 8.1 at 30 C), I mM MgCI2, and 0.6 mMi-mercaptoethanol were heated for 5 min at varioustemperatures and then cooled in an ice-water mixture.Five ,uditers of heated enzyme were incubated with14C-ATP and drug for 30 min at 30 C as indicated inMaterials and Methods. The results are expressed as
per cent activity remaining compared to the activityofan enzyme sample which had been kept at 5 C.
full wild-type streptomycin and spectinomycinresistance, with full recovery of adenylylatingactivity. However, a very small number were
found to be resistant (at a low level) to eitherstreptomycin or spectinomycin, but not to both.Analysis of the osmotic shockates from theselatter strains showed only partial restoration ofone of the activities. These "single-resistance"revertants were extremely unstable and rapidlydeveloped resistance to both drugs. No attemptshave been made to purify the labile enzymes ofthese strains.
Resistance levels of R factor strains. Seven Rfactors were transferred to strain w677 with theintent of comparing the levels of the enzymaticadenylylation of streptomycin and spectinomycinto the in vivo resistance levels to these drugs(Table 4). In each case, there is a direct correla-tion between the resistance level to each of thedrugs in vivo and the extent of enzymatic inactiva-tion of the drugs in vitro.Although strain w677 carrying R factor R-100
is resistant to 10 to 12 ,ig of streptomycin per ml
only, spontaneous mutants resistant to as high as2 mg of streptomycin per ml can readily be iso-lated. Osmotic shockates of these high resistancemutants show adenylylating activities that alsoincrease in direct proportion to the resistancelevel.The R factors listed in Table 4 are all in the
same host (w677), because transfer of an R factorfrom one host to another can sometimes lead tochanges in the levels of antibiotic resistance andadenylylating activity (R. Benveniste and J.Davies, unpublished data). Changes in resistancelevels have been noted when transferring Rfactors among gram-negative bacteria (16).
Purification of the adenylylating enzyme. Theenzyme has been partially purified approximately100-fold from osmotically shocked cells ofNR73/w677. Table 5 shows that the spectinomycin andstreptomycin adenylylating activities are purifiedto the same extent at each step. The P-200 enzymeyields 4 to 6 bands on polyacrylamide gel elec-trophoresis and, therefore, is not pure by thiscriterion.
Properties of the partially purified enzyme. The100-fold purified enzyme was found to have amolecular weight of approximately 33,000(+ 10%), as judged by the position of the peak
TABLE 4. Relationship between in vivo streptomycin(Str) and spectinomvcin (Spc) resistance of
R-factor strains and in vitroadenylylating activity
MIC Amt of MIC AmtR factora of Strb Str- Ratiod Spc of Spc- Ratiod
(sg/ml) AMPC (ug/mi) AMPc
NR1-22 4 0.5 0.13 40 0.75 0.019NR79 8 1.5 0.19 65 1.1 0.017R100 10 1.7 0.17 85 1.2 0.014NR84 12 2.2 0.18 80 1.4 0.018NR85 13 2.4 0.19 100 1.7 0.017NR86 22 3.7 0.17 135 2.2 0.016NR73 40 6.6 0.17 195 3.3 0.017
All R factors are present in host w677.b Minimum inhibitory concentration was de-
termined by reading the optical density at 550 nmof 5-ml cultures and shaking in Penassay broth at37 C, which contained increasing amounts of theantibiotic weighed as the free base. Values arecorrected for the resistance level of w677 to Str(2 ,g/ml) and Spc (6 Ag/ml).
Expressed as nanomoles per minute per milli-liter of enzyme. Enzymatic activity as determined onosmotic shockates prepared from 40-ml cultures (anaverage of two values) as described in Materials andMethods.
d Expressed as ratio of nanomoles of Str- or Spc-AMP per minute per milliliter to MIC of Str orSpc, respectively.
114 INFEC. IMMUN.
on Novem
ber 12, 2018 by guesthttp://iai.asm
.org/D
ownloaded from
VOL. 1, 1970 ADENYLYLATION OF STREPTOMYCIN AND SPECTINOMYCIN 115
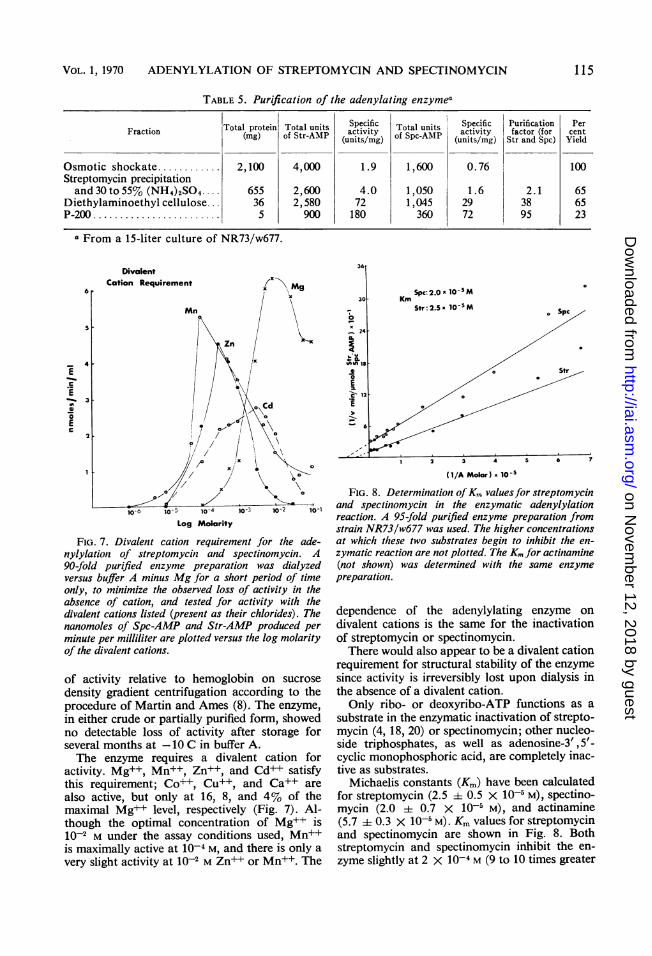
TABLE 5. Purification of the adenylating enzymea
Fraction Total protein Total units Specific Total units Specific Purification PerFraction ~~(mg) of Str-AMP activity of Spc AMP activity factor (for cent(units/mg) - (units/mg) Str and Spc) Yield
Osmotic shockate............ 2,100 4,000 1.9 1,600 0.76 100Streptomycin precipitationand30to55% (NH4)2SO4 655 2,600 4.0 1,050 1.6 2.1 65
Diethylaminoethyl cellulose .. 36 2,580 72 1,045 29 38 65P-200 ...................... 5 900 180 360 72 95 23
From a 15-liter culture of NR73/w677.
DivalentCation Requirement
4
E3
OA Cd
0 0~~~~~
o-6 10-5 10-4 10-3 10-2 10-1
Log Molarity
FIG. 7. Divalent cation requirement for the ade-nylylation of streptomycin and spectinomycin. A90-fold purified enzyme preparation was dialyzedversus buffer A minus Mg for a short period of timeonly, to minimize the observed loss of activity in theabsence of cation, and tested for activity with thedivalent cations listed (present as their chlorides). Thenanomoles of Spc-AMP and Str-AMP produced perminute per milliliter are plotted versus the log molarityof the divalent cations.
of activity relative to hemoglobin on sucrosedensity gradient centrifugation according to theprocedure of Martin and Ames (8). The enzyme,in either crude or partially purified form, showedno detectable loss of activity after storage forseveral months at -10 C in buffer A.The enzyme requires a divalent cation for
activity. Mg++, Mn++, Zn++, and Cd++ satisfythis requirement; Co++, Cu++, and Ca++ arealso active, but only at 16, 8, and 4% of themaximal Mg++ level, respectively (Fig. 7). Al-though the optimal concentration of Mg++ is10-2 M under the assay conditions used, Mn++is maximally active at 10-4 M, and there is only avery slight activity at 10-2 M Zn++ or Mn++. The
(1/A Molar) X 10--5
FIG. 8. Determination ofKm valuesfor streptomycinand spectinomycin in the enzymatic adenylylationreaction. A 95-fold purified enzyme preparation fromstrain NR73/w677 was used. The higher concentrationsat which these two substrates begin to inhibit the en-zymatic reaction are not plotted. The Km for actinamine(not shown) was determined with the same enzymepreparation.
dependence of the adenylylating enzyme ondivalent cations is the same for the inactivationof streptomycin or spectinomycin.There would also appear to be a divalent cation
requirement for structural stability of the enzymesince activity is irreversibly lost upon dialysis inthe absence of a divalent cation.Only ribo- or deoxyribo-ATP functions as a
substrate in the enzymatic inactivation of strepto-mycin (4, 18, 20) or spectinomycin; other nucleo-side triphosphates, as well as adenosine-3', 5'-cyclic monophosphoric acid, are completely inac-tive as substrates.
Michaelis constants (Kin) have been calculatedfor streptomycin (2.5 + 0.5 X 10- M), spectino-mycin (2.0 + 0.7 x 10-5 M), and actinamine(5.7 + 0.3 X 10-5 M). Km values for streptomycinand spectinomycin are shown in Fig. 8. Bothstreptomycin and spectinomycin inhibit the en-zyme slightly at 2 X 10-4 M (9 to 10 times greater
on Novem
ber 12, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

BENVENISTE, YAMADA, AND DAVIES
TABLE 6. Relative efficiency of different antibioticsand related molecules as substrates for the
adenylylating enzyme
Compound'
Streptomycin ...................
Dihydrostreptomycin ...........
Bluensomycin .................
N-benzyl streptomycylamine....Didesamidino dihydrostrepto-mycin ........................
Methyl streptobiosaminide......Methyl-2-deoxy-2-methylamino-
L-glucose .....................Streptamine ....................
Deoxystreptamine .............
Streptidine .....................
N-methyl d4ihydrostreptomycin..Streptomycin-adenylate .........
Streptomycin-phosphate ........
Spectinomycin ................
Actinamine....................1, 3-Diamino-1, 3-dideoxy-myo-
inositol.......................
Antibi-otic
YesYesYesYes
NoNo
NoNoNoNoNoNoNoYesNo
No
a See Fig. 3 and 4 for the structures.bCompared on an equimolar basis with the
concentration of streptomycin giving optimalactivity with a 45-fold purified enzyme preparationfrom strain NR73/w677. The rate of inactivationof those compounds that do not bind to phospho-cellulose paper was determined by electrophoresisof the reaction mixtures as described in Materialsand Methods. The amount of "4C-AMP transferredto the compound was monitored with a radio-chromatogram scanner and compared to theamount of Str-'4C-AMP formed under identicalconditions.
than their Km values) and inhibit more than90% of the reaction at 100 times the Km value.However, at 50 times its Km value, actinaminedoes not inhibit the enzyme.A variety of streptomycin derivatives (see Fig.
3) as well as simpler compounds have beenassayed for enzymatic activity by using the phos-phocellulose paper assay or electrophoresis oncellulose acetate strips (see Materials and Meth-ods) for those compounds that do not bind to thephosphocellulose paper (Table 6).
In addition to the inhibition of adenylylationby high concentrations of the two substratesstreptomycin or spectinomycin, the reaction is50% inhibited by one of the products, inorganicpyrophosphate (PPi), at 10-4 M. Possible inhibi-tion by high concentrations of ATP, and strepto-mycin-AMP or spectinomycin-AMP has notbeen examined.
Tetracycline inhibits the adenylylation reactionby20%at X 10-5Mandby70%at5 X 1O-4M.This inhibition, like that of PPi, is not due tobinding of required divalent cation, since theaddition of excess Mg++ (or streptomycin) tothe enzyme either before or after the additionof tetracycline does not alter the level of inhibi-tion. During an experiment in which the reactioncomponents were added in sequence, it was foundthat preincubation of the enzyme in the presenceof tetracycline, before the addition of ATP, ledto a greater inhibition than did the addition oftetracycline after ATP. At 3.5 X 10-4 M tetra-cycline, with prior addition of ATP, the reactionwas inhibited 62%; adding ATP after tetracyclinegave a 90% inhibition. The presence or absenceof streptomycin did not affect these results.EDTA seems to behave in a similar fashion,
although the extent of inhibition is less (62% at10-2 M EDTA), and in the presence of ATP thisinhibition can be partially reversed by addingmore Mg++.A wide variety of other aminoglycoside anti-
biotics such as gentamicin (and members of itscomplex), nebramycin, kanamycin, neomycin,and paromomycin do not inhibit the enzyme.Some of the more classical enzyme inhibitorssuch as azide, arsenate, iodide, cyanide, orp-chloromercuribenzoate do not inhibit theenzyme at concentrations up to 10-3 M.We have found that NH4CI is an activator of
the enzyme at high concentrations, but its effectis slight at lower concentrations (10% at 10-2 M),and it is omitted from the reaction mixture toavoid complicating the kinetic and inhibitiondata.
Antibiotic resistance pattern of R-factor strains.Fifteen strains carrying independently isolatedR factors have been examined for resistance tostreptomycin and spectinomycin (see Table 1).Eleven of them were resistant to both strepto-mycin and spectinomycin, and assays of theirosmotic shockates revealed high levels of adenyl-ylating activity for both these drugs. Thesestrains presumably contain the same inactivatingenzyme that is present in JE 254 or NR73/w677.The remaining four R-factor strains (JR35, NR71,NR45, and NR3) were resistant only to strepto-mycin, and none had any adenylylating activity.Instead, streptomycin was found to be inactivatedby phosphorylation in the osmotic shockates ofthese strains (2, 13a; R. Benveniste et al., Bac-teriol. Proc., p. 48, 1969).
DISCUSSIONEvidence has been obtained that a single R-
factor enzyme catalyzes the inactivation of strep-tomycin and spectinomycin by adenylylation.
116 INFEC. IMMUN.
on Novem
ber 12, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

VOL. 1, 1970 ADENYLYLATION OF STREPTOMYCIN AND SPECTINOMYCIN
The heat inactivation of a partially purifiedenzyme shows that both enzymatic activities aredestroyed at the same rate, confirming the com-mon nature of the two reactions.
During the purification of the enzyme re-sponsible for the streptomycin inactivation, bothenzymatic activities increase to the same extentat each step. The purest enzyme fraction obtainedto date catalyzes the adenylylation of strepto-mycin and spectinomycin with a divalent cationand ATP as the only other requirements.
Mutants selected for sensitivity to one of theantibiotics always show a complete loss of en-zymatic activity for both streptomycin andspectinomycin when tested in vitro. Revertantsselected for resistance to either drug show fullrestoration of both adenylylating activities, withthe exception of a small number of mutantswhich are extremely unstable both in vivo andin vitro and which have not been studied further.We suspect that these unstable mutants areaffected at the active site of the enzyme, so thatonly one of the antibiotics is recognized.
Further support for a common inactivatingenzyme for spectinomycin and streptomycincomes from the finding that R-factor strainsselected for increased resistance levels to eitherof the antibiotics show an increase of adenylylat-ing activity towards both antibiotics (R.Benveniste and J. Davies, unpublished data).This is also apparent from the data in Table 4,in which the strain most resistant to streptomycin(NR73/w677) is also the one most resistant tospectinomycin and the one that shows the highestadenylylation levels in vitro towards both anti-biotics. This same pattern applies to the secondmost resistant strain (NR86/w677), the third(NR85/w677), and so on.
R-factor expression can presumably be in-fluenced by some property of the host bacteriumor by regulatory functions on the R-factor. Inthose cases where levels of the same R-factorenzymes change when the R factor is transferredamong several hosts, the host bacterium is af-fecting the level of R-factor expression. In addi-tion, we have obtained spontaneous mutants ofR factor-carrying E. coli in which the enzymelevels have increased some 10-fold. We do notknow whether this increase in enzyme activity(and drug resistance) is due to a mutation in thehost bacterium or in the R factor itself. In eithercase, increased enzyme levels could result fromalterations in regulation or structure of R-factorenzymes or from an increase in the number ofcopies of R-factor DNA. Such mutants arecurrently under investigation in our laboratory.The enzymatic assay based on binding of the
substrates to phosphocellulose paper can be
used routinely to detect very small amounts ofstreptomycin or spectinomycin. For example,using 14C-ATP (10 uc/,umole), we have detectedas little as 0.01 ,g of unlabeled streptomycinbound to E. coli ribosomes (J. Davies, unpublisheddata); the specificity and sensitivity of the assaymay also lend itself to clinical assays for thesedrugs.A few tentative conclusions concerning the
structural requirements for substrate activitycan be drawn from the list of substrates whichcan form adenylyl derivatives when incubatedwith ATP and the enzyme (Table 6). As far asthe streptomycin molecule is concerned, altera-tions of the streptidine moiety of the molecule or,indeed, removal of the entire group by methanoly-sis (leaving methyl streptobiosaminide) do notaffect the extent of enzymatic inactivation tothe slightest degree. Removal of the streptoseresidue by methanolysis (leaving the N-methyl-L-glucosamine moiety of streptomycin), reduces thelevel of activity to 5c%O that of streptomycin. Thestreptose ring may be needed for steric reasons,although this is not obvious considering theextent of the enzymatic reaction with actinamine(a degradation product of spectinomycin).The substitution of another methyl group
on the methylamino group of streptomycin(N-methyldihydrostreptomycin) deprives themolecule of any substrate activity. This com-pound is also a weak inhibitor of the enzyme; themechanism of this inhibition is being examined.We have found that N-methyldihydrostrepto-mycin will not inhibit the formation of poly-phenylalanine from polyuridylic acid in an invitro incorporation system using E. coli ribo-somes. This compound, however, can be phos-phorylated by the streptomycin-phosphorylatingenzyme (13a). It should be noted (Table 6) thata compound does not have to be effective as anantibiotic to be inactivated by the adenylylatingenzyme.The importance of a D-threo methylamino-
alcohol moiety in the substrate is shown bycomparing the rate of enzymatic adenylylationof actinamine (50%) to that of 1 ,3-diamino-1 ,3-dideoxy-myo-inositol (7%), a compound con-taining a D-threo amino alcohol moiety.
Streptomycin phosphate, another enzymaticinactivation product of streptomycin, in whichthe 3'OH moiety of the N-methyl-L-glucosamineresidue is phosphorylated (2, 13a; R. Benvenisteet al., Bacteriol. Proc., p. 48, 1969), is, as ex-pected, completely inactive as a substrate for theadenylylating enzyme.We conclude that the adenylylating enzyme
transfers AMP to the D-threo methylaminoalcohol moiety that is present in both strepto-
117
on Novem
ber 12, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

BENVENISTE, YAMADA, AND DAVIES
mycin and spectinomycin. We cannot, at thistime, eliminate the possibility that the hydroxylgroup between the N-methyl groups of the ac-tinamine moiety of spectinomycin is the site ofattachment of the AMP residue, but consider itunlikely sterically, since this hydroxyl group isnot easily susceptible to chemical acetylation.If one accepts that enzymatic adenylylation by Rfactors is specific for a D-threo methylaminoalcohol grouping, the fact that spectinomycin is asubstrate for the enzyme supports the structurein Fig. 4 as the correct stereochemical arrange-ment for the linkages joining actinamine to therest of the molecule in this antibiotic.Although the Km values for streptomycin and
spectinomycin are similar (2 X 10- M)), therate of inactivation of the latter compound(Vmax) is roughly half that of streptomycin.The Km value for actinamine is 5.7 X 10- Mwhich would make it a less effective substratethan spectinomycin, but in fact the rate of itsinactivation is roughly equal to that of strepto-mycin. Although streptomycin and spectino-mycin inhibit the enzyme at 1.8 X 10-4 M (ninetimes their Km values), actinamine does not.Thus, in Fig. 5, the faster rate (apparent) ofinactivation of actinamine can be explained bythe inhibition of the spectinomycin adenylylationreaction by spectinomycin.The requirement of the adenylylating enzyme
for a divalent cation could simply be a require-ment of the enzyme for the divalent cationcomplex of ATP as the substrate; the divalentcation could also be required to activate theenzyme, to form a complex with inorganic pyro-phosphate, or possibly to serve in more thanone of these roles. In an experiment in whichATP and Mg++ were added in limiting equimolarquantities, the addition of 2 mm excess freeATP did not significantly increase the activityseen with any given concentration of Mg-ATP.The conclusion would be that free ATP does notserve as a substrate, and that the enzyme re-quires a divalent cation complex of ATP foractivity. The enzyme seems to have a muchhigher affinity for the Mn++- or Zn++-ATPcomplex than for the Mg++-ATP complex(Fig. 7).
Inhibition of enzymatic adenylylation of thedrugs by tetracycline might be explained by theability of the latter to remove the Mg++ requiredfor the structural stability of the enzyme. Theinhibition of divalent cation-requiring enzymesby tetracycline is well documented, although theexact mechanism is not known (6). The decreasedinhibition that is observed when ATP is addedbefore tetracycline may be due to the formationof an enzyme-Mg-AMP complex that protects this
Mg++ from binding to tetracycline. Experimentsare currently under way to determine whether theenzymatic reaction is an ordered process, withthe binding of ATP to form an enzyme-AMPcomplex with release of PPi occurring as a firststep, followed by the binding of streptomycinand release of streptomycin-AMP.
In the limited number of R factor-carryingstrains of E. coli that we have examined, theonly mechanism of spectinomycin resistance isinactivation by adenylylation. Streptomycin re-sistance should thus be expected to occur simul-taneously with spectinomycin resistance on Rfactors, since the same enzyme catalyzes bothreactions (R. Benveniste et al., Bacteriol. Proc.,p. 48, 1969). In 11 R-factor strains with con-comitant in vivo streptomycin and spectinomycinresistance examined by us so far, the in vitroassay has always revealed the adenylylatingenzyme present in the osmotic shockate. (Thereis no reason why the phosphorylating enzymeshould not also be present in certain R factors.)However, strains that are streptomycin-resistantand spectinomycin-sensitive inactivate the drugby a different mechanism, probably phosphoryla-tion, which has been shown to occur in severalR-factor strains of this nature examined so far(13a). Although the phosphorylating enzymereacts with the same hydroxyl group on the N-methyl-L-glucosamine moiety of streptomycinas the adenylylating enzyme, it does not recog-nize this same site in spectinomycin.
ACKNOWLEDGMENTS
We thank Y. Hirota for strain JE 254, R. Nakaya for other Rfactor-containing strains of E. coli, and L. Anderson and R. D.Wells for their helpful critiques of the manuscript during prepara-tion. Donald Tipper kindly carried out several electrophoresisdeterminations, and many people generously donated liberalquantities of antibiotics and their degradation products.
This investigation was supported by Public Health Servicegrant HD-02362-03 from the National Institute of Child Healthand Human Development.
LITERATURE CITED
1. Datta, N., and M. H. Richmond. 1966. The purification andproperties of a penicillinase whose synthesis is mediated byan R-factor in Escherichia coli. Biochem. J. 98:204-209.
2. Davies, J., R. Benveniste, K. Kvitek, B. Ozanne, and T.Yamada. 1969. Aminoglycosides: biologic effects of molec-ular manipulation. J. Infec. Dis. 119:351-354.
3. Davies, J., L. Gorini, and B. D. Davis. 1965. Misreading ofRNA codewords induced by aminoglycoside antibiotics.Mol. Pharmacol. 1:93-106.
4. Harwood, J. H., and D. H. Smith. 1969. Resistance factor-mediated streptomycin resistance. J. Bacteriol. 97:1262-1271.
5. Kondo, S., M. Okanishi, R. Utahara, K. Maeda, and H.Umezawa. 1968. Isolation of kanamycin and paromamineinactivated by E. coli carrying R-factors. J. Antibiot.(Tokyo) Ser. A 21:22-29.
6. Laskin, A. I. 1967. Tetracycline, p. 331-359. In D. Gottlieb
118 INFEC. IMMUN.
on Novem
ber 12, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

VOL. 1,1970 ADENYLYLATION OF STREPTOMYCIN AND SPECTINOMYCIN
and P. D. Shaw (ed.), Antibiotics: mechanism of action,Vol. 1. Springer-Verlag, Inc., New York, N.Y.
7. Lowry, 0. H., N. J. Rosebrough, A. L. Farr, and R. J.Randall. 1951. Protein measurement with the Folin phenolreagent. J. Biol. Chem. 193:265-275.
8. Martin, R. G., and B. N. Ames. 1960. A method for determin-ing the sedimentation behavior of enzymes: application toprotein mixtures. J. Biol. Chem. 236:1372-1379.
9. Nossal, N. G., and L. A. Heppel. 1966. The release of enzymesby osmotic shock from Escherichia coli in exponentialphase. J. Biol. Chem. 241:3055-3062.
10. Okamoto, S., and Y. Suzuki. 1965. Chloramphenicol, di-hydrostreptomycin, and kanamycin-inactivating enzymesfrom multiple drug resistant Escherichia coli carryingepisome R. Nature 208:1301-1303.
11. Okanishi, M., S. Kondo, Y. Suzuki, S. Okamoto, and H.Umezawa. 1967. Studies in inactivation of kanamycin andresistances of E. coli. J. Antibiot. (Tokyo) Ser. A 20:132-135.
12. Okanishi, M., S. Kondo, R. Utahara, and H. Umezawa. 1967.Phosphorylation and inactivation of aminoglycosidicantibiotics by E. coil carrying R-factors. J. Antibiot.(Tokyo) Ser. A 21:13-21.
13. Osborn, M., S. Person, S. Phillips, and F. Funk. 1967. Adetermination of mutagen specificity in bacteria using
nonsense mutants of bacteriophage T4. J. Mol. Biol. 26:437-447.
13a. Ozanne, B., R. Benveniste, D. Tipper, and J. Davies. 1969.Aminoglycoside antibiotics: inactivation by phosphoryla-tion in Escherichia coli carrying R factors. J. Bacteriol. 100:1144-1146.
14. Rinehart, K. L. 1964. The neomycins and related antibiotics,p. 90. John Wiley & Sons, Inc., New York.
15. Smith, D. H. 1967. R factor infection of Escherichia colilyophilized in 1946. J. Bacteriol. 94:2071-2072.
16. Smith, J. T. 1969. R-factor gene expression in gram-negativebacteria. J. Gen. Microbiol. 55:109-120.
17. Umezawa, H., M. Okanishi, S. Kondo, K. Hamana, R.Utahara, K. Maeda, and S. Mitsuhashi. 1967. Phosphoryla-tive inactivation of aminoglycosidic antibiotics by Esch-erichia coli carrying R-factor. Science 157:1559-1561.
18. Umezawa, H., S. Takasawa, M. Okanishi, and R. Utahara.1968. Adenylylstreptomycin, a product of streptomycininactivation by E. coli carrying R-factor. J. Antibiot.(Tokyo) Ser. A 21:81-92.
19. Weisblum, B., and J. Davies. 1968. Antibiotic inhibitors ofthe bacterial ribosome. Bacteriol. Rev. 32:493-528.
20. Yamada, T., D. Tipper, and J. Davies. 1968. Enzymaticinactivation of streptomycin by R-factor-resistant Esch-erichia coli. Nature 219:288-291.
119
on Novem
ber 12, 2018 by guesthttp://iai.asm
.org/D
ownloaded from










![BMC Biotechnology BioMed Central...1-kb increase in transposon size [23]. To keep the transpo-son under 2 kb the GFPuv reporter and spectinomycin resistance genes were driven by a](https://static.fdocuments.in/doc/165x107/60cd8e2d907b537a414322a8/bmc-biotechnology-biomed-central-1-kb-increase-in-transposon-size-23-to-keep.jpg)







