
Enhancing Sucrose Synthase Activity in Transgenic Potato
12
5 These authors contributed equally to this work. ∗ Corresponding author: E-mail, [email protected]; Fax, +34-948232191. Plant Cell Physiol. 50(9): 1651–1662 (2009) doi:10.1093/pcp/pcp108, available online at www.pcp.oxfordjournals.org © The Author 2009. Published by Oxford University Press on behalf of Japanese Society of Plant Physiologists. All rights reserved. For permissions, please email: [email protected] Sucrose synthase (SuSy) is a highly regulated cytosolic enzyme that catalyzes the conversion of sucrose and a nucleoside diphosphate into the corresponding nucleoside diphosphate glucose and fructose. To determine the impact of SuSy activity in starch metabolism and yield in potato ( Solanum tuberosum L.) tubers we measured sugar levels and enzyme activities in tubers of SuSy- overexpressing potato plants grown in greenhouse and open field conditions. We also transcriptionally characterized tubers of SuSy-overexpressing and -antisensed potato plants. SuSy-overexpressing tubers exhibited a substantial increase in starch, UDPglucose and ADPglucose content when compared with controls. Tuber dry weight, starch content per plant and total yield of SuSy-overexpressing tubers increased significantly over those of control plants. In contrast, activities of enzymes directly involved in starch metabolism in SuSy- overexpressing tubers were normal when compared with controls. Transcriptomic analyses using POCI arrays and the MapMan software revealed that changes in SuSy activity affect the expression of genes involved in multiple biological processes, but not that of genes directly involved in starch metabolism. These analyses also revealed a reverse correlation between the expressions of acid invertase and SuSy-encoding genes, indicating that the balance between SuSy- and acid invertase-mediated sucrolytic pathways is a major determinant of starch accumulation in potato tubers. Results presented in this work show that SuSy strongly determines the intracellular levels of UDPglucose, ADPglucose and starch, and total yield in potato tubers. We also show that enhancement of SuSy activity represents a useful strategy for increasing starch accumulation and yield in potato tubers. Keywords: Carbohydrate metabolism. Abbreviations: ADPG, ADPglucose; SuSy, sucrose synthase; U, unit of enzyme activity; UDPG, UDPglucose. Introduction In many heterotrophic organs sucrose synthase (SuSy) (E.C. 2.4.1.13) is a major determinant of sink strength that highly controls the channeling of incoming sucrose into starch and cell wall polysaccharides (Amor et al. 1995, Herbers and Son- newald 1998, Konishi et al. 2004). SuSy is a highly regulated enzyme that catalyzes the reversible conversion of sucrose and a nucleoside diphosphate into the corresponding nucle- oside diphosphate glucose and fructose (Pontis et al. 1981, Nakai et al. 1998, Purcell et al. 1998, Déjardin et al. 1999, Asano et al. 2002, Ciereszko and Kleczkowksi 2002, Hardin et al. 2004). Although UDP is the preferred nucleoside diphosphate substrate of SuSy to produce UDPglucose (UDPG), ADP is also an effective acceptor molecule of this sucrolytic enzyme to produce ADPglucose (ADPG) (Slabnik et al. 1968, Delmer and Albersheim 1970, Zervosen et al. 1998, Porchia et al. 1999, Baroja-Fernández et al. 2003). Genetic evidence demonstrating the importance of SuSy in starch production and sink strength determination in Enhancing Sucrose Synthase Activity in Transgenic Potato ( Solanum tuberosum L.) Tubers Results in Increased Levels of Starch, ADPglucose and UDPglucose and Total Yield Edurne Baroja-Fernández 1,5 , Francisco José Muñoz 1,5 , Manuel Montero 1 , Ed Etxeberria 1,2 , María Teresa Sesma 1 , Miroslav Ovecka 1,3 , Abdellatif Bahaji 1 , Ignacio Ezquer 1 , Jun Li 1 , Salomé Prat 4 and Javier Pozueta-Romero 1, ∗ 1 Instituto de Agrobiotecnología (CSIC/UPNA/Gobierno de Navarra), Mutiloako etorbidea z/g, 31192 Mutiloabeti, Nafarroa, Spain 2 IFAS Citrus Research and Education Center, University of Florida, 700 Experiment Station Road, Lake Alfred, FL 33850, USA 3 Institute of Botany, Slovak Academy of Sciences, Dubravska cesta 14, SK-84523 Bratislava, Slovakia 4 Centro Nacional de Biotecnología, CSIC, Campus Universidad Autónoma de Madrid, c/Darwin 3, 28049 Madrid, Spain 1651 Plant Cell Physiol. 50(9): 1651–1662 (2009) doi:10.1093/pcp/pcp108 © The Author 2009. Regular Paper Downloaded from https://academic.oup.com/pcp/article/50/9/1651/1854144 by guest on 05 February 2022
Transcript of Enhancing Sucrose Synthase Activity in Transgenic Potato

5 These authors contributed equally to this work. ∗ Corresponding author: E-mail, [email protected] ; Fax, + 34-948232191 .
Plant Cell Physiol. 50(9): 1651–1662 (2009) doi:10.1093/pcp/pcp108, available online at www.pcp.oxfordjournals.org© The Author 2009. Published by Oxford University Press on behalf of Japanese Society of Plant Physiologists.All rights reserved. For permissions, please email: [email protected]
Sucrose synthase (SuSy) is a highly regulated cytosolic enzyme that catalyzes the conversion of sucrose and a nucleoside diphosphate into the corresponding nucleoside diphosphate glucose and fructose. To determine the impact of SuSy activity in starch metabolism and yield in potato ( Solanum tuberosum L.) tubers we measured sugar levels and enzyme activities in tubers of SuSy-overexpressing potato plants grown in greenhouse and open fi eld conditions. We also transcriptionally characterized tubers of SuSy-overexpressing and -antisensed potato plants. SuSy-overexpressing tubers exhibited a substantial increase in starch, UDPglucose and ADPglucose content when compared with controls. Tuber dry weight, starch content per plant and total yield of SuSy-overexpressing tubers increased signifi cantly over those of control plants. In contrast, activities of enzymes directly involved in starch metabolism in SuSy-overexpressing tubers were normal when compared with controls. Transcriptomic analyses using POCI arrays and the MapMan software revealed that changes in SuSy activity affect the expression of genes involved in multiple biological processes, but not that of genes directly involved in starch metabolism. These analyses also revealed a reverse correlation between the expressions of acid invertase and SuSy-encoding genes, indicating that the balance between SuSy- and acid invertase-mediated sucrolytic pathways is a major determinant of starch accumulation in potato tubers. Results presented in this work show that SuSy strongly determines the intracellular
levels of UDPglucose, ADPglucose and starch, and total yield in potato tubers. We also show that enhancement of SuSy activity represents a useful strategy for increasing starch accumulation and yield in potato tubers.
Keywords: Carbohydrate metabolism .
Abbreviations : ADPG , ADPglucose ; SuSy , sucrose synthase ; U , unit of enzyme activity ; UDPG , UDPglucose.
Introduction
In many heterotrophic organs sucrose synthase (SuSy) (E.C. 2.4.1.13) is a major determinant of sink strength that highly controls the channeling of incoming sucrose into starch and cell wall polysaccharides ( Amor et al. 1995 , Herbers and Son-newald 1998 , Konishi et al. 2004 ). SuSy is a highly regulated enzyme that catalyzes the reversible conversion of sucrose and a nucleoside diphosphate into the corresponding nucle-oside diphosphate glucose and fructose ( Pontis et al. 1981 , Nakai et al. 1998 , Purcell et al. 1998 , Déjardin et al. 1999 , Asano et al. 2002 , Ciereszko and Kleczkowksi 2002 , Hardin et al. 2004 ). Although UDP is the preferred nucleoside diphosphate substrate of SuSy to produce UDPglucose (UDPG), ADP is also an effective acceptor molecule of this sucrolytic enzyme to produce ADPglucose (ADPG) ( Slabnik et al. 1968 , Delmer and Albersheim 1970 , Zervosen et al. 1998 , Porchia et al. 1999 , Baroja-Fernández et al. 2003 ). Genetic evidence demonstrating the importance of SuSy in starch production and sink strength determination in
Enhancing Sucrose Synthase Activity in Transgenic Potato ( Solanum tuberosum L.) Tubers Results in Increased Levels of Starch, ADPglucose and UDPglucose and Total Yield Edurne Baroja-Fernández 1 , 5 , Francisco José Muñoz 1 , 5 , Manuel Montero 1 , Ed Etxeberria 1 , 2 , María Teresa Sesma 1 , Miroslav Ovecka 1 , 3 , Abdellatif Bahaji 1 , Ignacio Ezquer 1 , Jun Li 1 , Salomé Prat 4 and Javier Pozueta-Romero 1 , ∗ 1 Instituto de Agrobiotecnología (CSIC/UPNA/Gobierno de Navarra), Mutiloako etorbidea z/g, 31192 Mutiloabeti, Nafarroa, Spain 2 IFAS Citrus Research and Education Center, University of Florida, 700 Experiment Station Road, Lake Alfred, FL 33850, USA 3 Institute of Botany, Slovak Academy of Sciences, Dubravska cesta 14, SK-84523 Bratislava, Slovakia 4 Centro Nacional de Biotecnología, CSIC, Campus Universidad Autónoma de Madrid, c/Darwin 3, 28049 Madrid, Spain
1651Plant Cell Physiol. 50(9): 1651–1662 (2009) doi:10.1093/pcp/pcp108 © The Author 2009.
Regular PaperD
ownloaded from
https://academic.oup.com
/pcp/article/50/9/1651/1854144 by guest on 05 February 2022
heterotrophic organs comes from QTL analyses in maize endosperms ( Thévenot et al. 2005 ), from the substantial reduction of starch levels in the sh1 maize mutants ( Chourey and Nelson 1976 ) and from genetically engineered potato tubers exhibiting a decrease in SuSy activity ( Zrenner et al. 1995 ).
The classical view on sucrose–starch conversion in sink organs of dicotyledonous plants assumes that ADPG pyro-phosphorylase is the sole source of ADPG linked to starch biosynthesis ( Preiss 1988 , Stark et al. 1992 , Müller-Röber et al. 1992 ). According to this view, SuSy catalyzes the con-version of sucrose into fructose and UDPG, which is then converted to glucose-1-phosphate and glucose-6-phosphate by UDPG pyrophosphorylase and cytosolic phosphogluco-mutase. Cytosolic glucose-6-phosphate enters the amylo-plast where it is converted to starch by the coupled reactions of plastid phosphoglucomutase, ADPG pyrophosphorylase and starch synthase ( Tauberger et al. 2000 ). However, in recent years an increasing body of evidence has appeared that points out inconsistencies with such a mechanism ( Muñoz et al. 2006 ), and previews the likelihood of the oper-ation of alternative and/or additional pathway(s) wherein SuSy catalyzes directly the de novo production of ADPG in the cytosol, which is subsequently imported into the amylo-plast by the action of a still unidentifi ed ADPG translocator ( Pozueta-Romero et al. 1991 , Naeem et al. 1997 ) (cf. Fig. 1C , Baroja-Fernández et al. 2003 ). Consistent with this view, using transgenic potato plants expressing an ADPG cleaving enzyme in the cytosol, Muñoz et al. (2008) have recently provided evidence showing that an important pool of ADPG linked to potato tuber starch biosynthesis occurs in the cytosol. Essentially in line with investigations describing the occurrence of processes in bacteria and animals in which gly-cogen synthesis and degradation take place simultaneously ( Bollen et al. 1998 , Guedon et al. 2000 ), active turnover of starch has been shown to occur in non-photosynthetic tis-sues of plants ( Pozueta-Romero and Akazawa 1993 , Neuhaus et al. 1995 , Sweetlove et al. 1996 ). It thus appears that both plastid phosphoglucomutase and ADPG pyrophosphorylase play crucial roles in the scavenging of the glucose units derived from the starch breakdown. The occurrence of an alternative/additional pathway of sucrose–starch conver-sion implies that neither UDPG produced by SuSy, nor cyto-solic hexose-phosphates derived from the action of UDPG pyrophosphorylase on this nucleotide-sugar, are involved in starch biosynthesis (cf. Fig. 1C , Baroja-Fernández et al. 2003 ), which is consistent with the normal starch phenotype of potato tubers silenced for UDPG pyrophosphorylase ( Zrenner et al. 1993 ). This view also agrees with the starch-defi cient phenotype of transgenic potato tubers expressing either bacterial sucrose phosphorylase ( Trethewey et al. 2001 ) or invertase ( Trethewey et al. 1998 ), since the high sucrolytic activity occurring in these tubers leads to a
drastic reduction of sucrose levels, thus limiting the SuSy-dependent production of ADPG linked to starch biosynthesis.
Potato ( Solanum tuberosum L.) is currently the fourth most important crop worldwide after maize, wheat, and rice (FAOSTAT data, http://www.faostat.fao.org ). This crop is notably recognized as a source of proteins, vitamins, minerals, polyphenols and starch ( Al-Saikhan et al. 1995 , Appeldoorn et al. 1997 , Morris et al. 2004 , Andre et al. 2007 ). As part of our current investigation on the impact of SuSy activity on starch metabolism in heterotrophic organs, in this work we biochemically and transcriptionally characterized tubers of potato plants that constitutively overexpress Sus4, a SuSy isoform normally occurring in tubers ( Fu and Park 1995 ). We also transcriptionally characterized starch-defi cient potato tubers silenced for sus4 expression ( Zrenner et al. 1995 ). Our results show that SuSy is a major determinant of the intrac-ellular levels of UDPG and ADPG in potato tubers, and show that changes in SuSy activity exert a strong impact on expres-sion of genes involved in multiple biological processes other than starch synthesis and degradation. Finally, we also show
A
0
50
100
150
200
250
300
Control 4 3 12 6 5 10 7
35S-SuSy-NOS
AD
Pgl
ucos
e pr
oduc
ing
SuS
y ac
tivity
(mU
/ g f
resh
wei
ght)
B
0
200
400
600
800
1000
1200
Control 4 3 12 6 5 10 7
35S-SuSy-NOS
Sta
rch
(µm
ol g
luco
se/ g
fre
sh w
eig
ht)
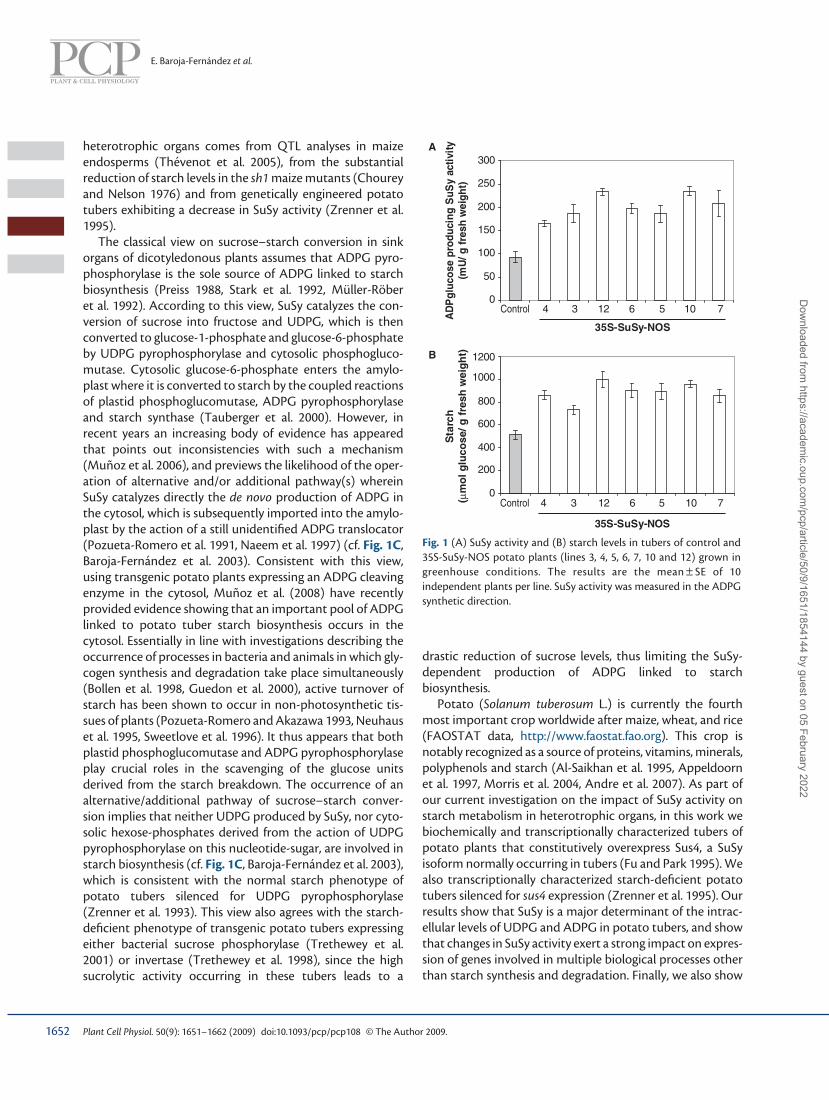
Fig. 1 (A) SuSy activity and (B) starch levels in tubers of control and 35S-SuSy-NOS potato plants (lines 3, 4, 5, 6, 7, 10 and 12) grown in greenhouse conditions. The results are the mean ± SE of 10 independent plants per line. SuSy activity was measured in the ADPG synthetic direction.
1652
E. Baroja-Fernández et al.
Plant Cell Physiol. 50(9): 1651–1662 (2009) doi:10.1093/pcp/pcp108 © The Author 2009.
Dow
nloaded from https://academ
ic.oup.com/pcp/article/50/9/1651/1854144 by guest on 05 February 2022

that enhancement of SuSy activity represents a potentially very useful strategy for increasing starch content and total yield in heterotrophic plant organs.
Results and Discussion
Enhancing SuSy activity results in increased starch, UDPG and ADPG contents in tubers of transgenic potato plants when grown in greenhouse conditions Tubers of sus4 -overexpessing plants ( Muñoz et al. 2005 ) grown in greenhouse conditions were characterized for starch and soluble sugar content. As it is our experience that biochemical analyses are subject to considerable variation, we analyzed 10 plants per line to ensure the attainment of reliable data. As illustrated in Fig. 1A , SuSy activities in con-trol tubers were ca. 90 mU/g FW, whereas activities in SuSy-overexpressing tubers ranged between 170 mU/g FW and 250 mU/g FW. Importantly, analyses of starch content revealed that SuSy-overexpressing tubers contained 55–85 % more starch than control tubers ( Fig. 1B ).
Consistent with the capacity of SuSy to produce UDPG and ADPG from sucrose in vitro ( Delmer and Albersheim 1970 , Baroja-Fernández et al. 2003 , Muñoz et al. 2005 ), SuSy-overexpressing tubers accumulated high levels of UDPG and ADPG when compared with controls ( Table 1 ). These results are also consistent with a previous report showing that potato tubers silenced for SuSy expression accumulate only 25–30 % of the normal UDPG and ADPG ( Baroja-Fernández et al. 2003 ). We must emphasize that transgenic potato tubers with altered activities of ADPG pyrophosphorylase and UDPG pyrophosphorylase have been shown to accumu-late normal levels of ADPG and UDPG ( Sweetlove et al. 1996 ,
Zrenner et al. 1993 ), the overall data indicating that SuSy is the major determinant of intracellular levels of UDPG and ADPG in potato tubers.
SuSy-overexpressing tubers accumulated normal levels of hexose-phosphates ( Table 1 ), which is in agreement with a previous report showing that SuSy-antisensed potato tubers accumulate normal levels of hexose-phosphates ( Baroja-Fernández et al. 2003 ). Noteworthy, levels of 3-phosphoglyceric acid in tubers from all SuSy lines were signifi cantly higher than in control tubers ( Table 1 ), which may refl ect massive channeling of fructose produced by SuSy into the glycolytic pathway ( Geigenberger et al. 1997 ). 3-phosphoglyceric acid is an allosteric activator of ADPG pyrophosphorylase ( Preiss 1988 ). Therefore, although previous reports have shown that potato tubers with elevated levels of 3-phosphoglyceric acid accumulate normal or even reduced starch content ( Geiger et al. 1998 , Sweetlove et al. 2001 ), it is likely that under the present conditions, high 3-phosphoglyceric acid levels favor starch accumulation in the SuSy-overexpressing tubers.
Analyses of enzymes closely associated with starch and sucrose metabolism revealed that SuSy - overexpressing tubers display normal alkaline pyrophosphatase, alkaline invertase, sucrose-phosphate synthase, ADP-sugar pyro-phosphatase, total starch synthase, starch phosphorylase, and total amylolytic activities when compared with control tubers ( Supplementary Table S1 ). ADPG pyrophosphory-lase and UDPG pyrophosphorylase activities in tubers of three of the seven lines were signifi cantly higher than in control tubers, whereas ADPG pyrophosphorylase activities in tubers of one SuSy line were signifi cantly lower than in control tubers ( Supplementary Table S1 ). The overall results thus indicate that the increase in ADPG, UDPG and
Table 1 Metabolite levels in the tubers of control and 35S-SuSy-NOS plants grown in greenhouse conditions
Control 35S-SuSy-NOS
4 3 12 6 5 10 7
UDPG (nmol/g FW) 98.0 ± 4.0 135.1 ± 3.5 144.3 ± 10.3 145.2 ± 13.9 135.5 ± 12.0 146.9 ± 11.0 156.8 ± 5.0 131.0 ± 6.2
ADPG (nmol/g FW) 4.6 ± 0.36 6.0 ± 0.64 7.2 ± 0.67 7.4 ± 0.57 5.7 ± 0.62 7.8 ± 0.99 7.8 ± 1.16 7.3 ± 0.88
3PGA (nmol/g FW) 42.7 ± 2.46 70.3 ± 3.89 57.2 ± 3.16 5 2.9 ± 2.92 65.7 ± 3.63 55.7 ± 3.08 6 1.7 ± 3.41 65.0 ± 3.19
Glucose-1-P (nmol/ g FW) 11.5 ± 1.0 9.6 ± 1.7 9.5 ± 1.2 8.0 ± 0.9 9.7 ± 1.2 9.5 ± 2.1 10.5 ± 1.2 8.9 ± 1.3
Glucose-6-P (nmol/ g FW) 112.7 ± 7.1 98.6 ± 2.1 117.8 ± 1.1 115.9 ± 9.7 101.6 ± 7.9 94.8 ± 12.9 112.6 ± 14.7 97.1 ± 13.0
Fructose-6-P (nmol/ g FW) 44.9 ± 2.4 41.2 ± 2.2 49.9 ± 2.7 48.1 ± 2.6 40.79 ± 2.2 30.5 ± 2.2 40.3 ± 2.2 40.6 ± 2.2
Glucose (µmol/ g FW) 38.5 ± 9.3 25.4 ± 0.9 36.7 ± 7.0 31.3 ± 9.5 23.9 ± 7.1 19.5 ± 4.4 16.4 ± 4.3 20.4 ± 6.7
Fructose (µmol/ g FW) 5.6 ± 1.3 6.3 ± 1.8 5.1 ± 0.7 8.0 ± 2.3 5.9 ± 2.3 8.6 ± 2.7 8.3 ± 3.0 6.0 ± 2.3
Sucrose (µmol/ g FW) 48.8 ± 10.1 37.1 ± 1.1 36.2 ± 3.7 47.4 ± 3.5 42.7 ± 6.5 43.1 ± 2.5 43.3 ± 4,6 35.4 ± 2.3
The results are the mean ± SE of extracts from 10 independent plants per line. Values that are signifi cantly higher than those of control plants are marked in bold. Values that are signifi cantly lower than those of control plants are underlined.
FW: fresh weight; 3PGA, 3-phosphoglyceric acid.
1653
Susy impact in potato tuber starch metabolism
Plant Cell Physiol. 50(9): 1651–1662 (2009) doi:10.1093/pcp/pcp108 © The Author 2009.
Dow
nloaded from https://academ
ic.oup.com/pcp/article/50/9/1651/1854144 by guest on 05 February 2022

starch content in SuSy-overexpressing tubers cannot be ascribed to changes in the activities of enzymes closely con-nected to starch and sucrose metabolisms other than SuSy.
SuSy overexpression leads to increase of tuber starch, ADPG and UDPG content, yield and dry weight per plant when grown in fi eld conditions Because starch metabolism under greenhouse conditions may be directly infl uenced by factors such as pot size and by temperature, light and humidity regimes, we also character-ized SuSy-overexpressing potato tubers from plants grown in the fi eld. We must emphasize that, except in a few instances ( Regierer et al. 2002 ), genetic engineering of potato plants has not been shown to be successful in increasing potato tuber starch and yield in fi eld conditions. As illus-trated in Fig. 2A , SuSy activities in control tubers were ca. 170 mU/g FW, whereas activities in tubers of the SuSy-over-expressing transgenic lines ranged between 220 and 360 mU/g FW. None of the selected transgenic plants showed phenotypic alterations in the aerial parts compared with
control plants (not shown). Average tuber number per plant in SuSy-overexpressing plants also remained unaltered (not shown). Similar to tubers of SuSy-overexpressing plants grown in greenhouse conditions, tubers of SuSy-overex-pressing plants grown in the open fi eld accumulated signifi -cantly higher levels of ADPG and UDPG than control tubers ( Supplementary Fig. S1 ). Moreover, analyses of the starch content revealed that SuSy-overexpressing tubers contained 35–55 % more starch than control tubers ( Fig. 2B ). Tuber dry weight and tuber starch content (both referred to as a percentage of tuber fresh weight) were signifi cantly higher in the SuSy lines than in control plants ( Table 2 ), indicating that the increase in starch resulted at the expense of water. Signifi cantly, the tuber starch content to dry weight ratio was higher in the SuSy lines than in control plants. In addi-tion, although tuber fresh weight per plant was normal in SuSy-overexpressing plants, both tuber dry weight and starch yield per plant were signifi cantly higher in SuSy-over-expressing plants than in control plants ( Table 2 ). In terms of productivity per land surface unit, control plants pro-duced an average of 4.4 tonnes of starch/hectare, whereas SuSy-overexpressing plants produced between 6.1 and 8.2 tonnes of starch/hectare ( Table 2 ).
Taking into account all the limitations inherent in basing conclusions on genetically engineered plants, the overall data show that enhancement of SuSy expression represents a very useful and effective strategy for increasing starch accumulation and total yield in potato tubers.
Changes in SuSy activity do not affect the expression of genes directly involved in starch metabolism Data obtained using SuSy-overexpressing potato tubers (this work), together with those obtained using SuSy-antisensed potato tubers ( Zrenner et al. 1995 , Baroja-Fernández et al. 2003 ) strongly indicate that SuSy is a major determinant of the intracellular levels of ADPG, UDPG and starch in potato tubers. Because cytosolic UDPG serves as a glucosyl donor for the synthesis of glycolypids, glycoproteins, trehalose and cell wall polysaccharides ( Kleczkowski 1994 , Amor et al. 1995 , Neckelmann and Orellana 1998 ), it is highly conceiv-able that changes in SuSy activity will lead to important changes in the tuber metabolome and N -glycome. Further studies will be necessary to confi rm this conjecture.
It is noteworthy that a single enzymatic step is capable of affecting to such a dramatic extent the synthesis and accu-mulation of starch and nucleotide-sugars without affecting enzymes directly involved in starch metabolism (cf. Supple-mental Table S1 ). To better understand this phenomenon, high-throughput transcriptomic analysis of tubers from SuSy-overexpressing plants, and from S-112 and S-129 plants silenced for SuSy expression ( Zrenner et al. 1995 ) was per-formed using the POCI 44K 60-mer oligo array ( http://pgrc.ipk-gatersleben.de/poci ) ( Kloosterman et al. 2008 ). Full
A
Control 4 3 12 6 5 10 7
35S-SuSy-NOS
AD
Pgl
ucos
e pr
oduc
ing
SuS
y ac
tivity
(mU
/g f
resh
wei
ght)
0
50
100
150
200
250
300
350
400
B
Control 4 3 12 6 5 10 7
35S-SuSy-NOS
Sta
rch
(µm
ol g
luco
se/g
fre
sh w
eig
ht)
0
100
200
300
400
500
600
700
800
Fig. 2 (A) SuSy activity, and (B) starch levels in tubers of control and 35S-SuSy-NOS potato plants (lines 3, 4, 5, 6, 7, 10 and 12) grown in the fi eld (May–September 2006). The results are the mean ± SE of 90 independent plants per line. SuSy activity was measured in the ADPG synthetic direction.
1654
E. Baroja-Fernández et al.
Plant Cell Physiol. 50(9): 1651–1662 (2009) doi:10.1093/pcp/pcp108 © The Author 2009.
Dow
nloaded from https://academ
ic.oup.com/pcp/article/50/9/1651/1854144 by guest on 05 February 2022

details of the array hybridization analysis are given in Supplementary Table S2 and Supplementary Table S3 . In SuSy-antisensed tubers 357 genes were found to be deregu-lated ( > 3-fold difference relative to control; P < 0.05), 50 % of them being annotated as ‘no hits found’. Among this popu-lation, 186 genes were down-regulated and 171 genes were up-regulated. In SuSy-overexpressing tubers, 60 genes were down-regulated and 58 genes were up-regulated. Quantita-tive RT–PCR of some of the identifi ed genes ( Supplemen-tary Fig. S2 ) validated the results of the array analyses. To determine the biological processes affected by changes in SuSy activity, an analysis of genes using the MapMan tool ( Thimm et al. 2004 ) ( http://gabi.rzpd.de/projects/MapMan/ ) was carried out. This study revealed that changes in SuSy activity are accompanied by drastic changes in expression of genes involved in multiple processes such as metabolism of carbohydrates, amino acids and lipids, redox status of the cell, development, cell wall biosynthesis, secondary metabo-lism, protein translation and stability, energy production and stress responses ( Fig. 3 , Supplementary Fig. S3 and Supplementary Fig. S4 ). Noteworthy, in line with the results presented in Supplementary Table S1 , no changes were observed in the expression of many genes that code for enzymes directly involved in starch and sucrose metabolism such as starch synthases, plastidial starch phosphorylase, ADPG pyrophosphorylases, α -glucan/water dikinases, hexose-P translocator, plastidial adenylate kinase, UDPG pyrophosphorylases, sucrose-phosphate synthases, alkaline
invertase, β -amylases and α -amylases. In addition, no changes were observed in the expression levels of genes that code for glycolytic enzymes and of important components of carbon sensing/signaling pathways such as the sucrose non-fermenting-1-related protein kinase-1 (SnRK1), hexoki-nase and enzymes involved in trehalose biosynthesis ( Purcell et al. 1998 , Tiessen et al. 2003 , Halford and Paul 2003 ).
The onset of starch accumulation occurring during potato tuber development is accompanied by an increase in SuSy activity and a concomitant decline in acid invertase activity ( Prat et al. 1990 , Appeldoorn et al. 1997 ), the latter being post-translationally regulated by a proteinaceous inhibitor ( Bracho and Whitaker 1990 ). Noteworthy, our RNA profi ling analyses revealed that enhancement of SuSy activity leads to down-regulation of acid invertase expression (2.85-fold decrease) ( Supplementary Table S3 ). By contrast, reduc-tion of SuSy activity resulted in enhancement of acid invertase-encoding transcripts (4.76-fold increase), and in a drastic reduction of the levels of transcripts that code for the inhibitor of this sucrolytic enzyme (16.2-fold decrease) ( Sup-plementary Table S2 ). The occurrence of high levels of expression of acid invertase in SuSy-antisensed tubers is con-sistent with the report of Zrenner et al. (1995) . The reverse correlation between SuSy and acid invertase expression was further supported by our analyses of acid invertase activity in both SuSy-overexpressing and SuSy-antisensed tubers ( Fig. 4 ). It thus appears that (i) SuSy- and acid invertase-mediated sucrolytic pathways may be co-ordinately regulated
Table 2 Fresh weight, dry weight and starch content and yield of control and 35S-SuSy-NOS plants grown under field conditions (May–September 2006)
Control 35S-SuSy-NOS
4 3 12 6 5 10 7
Tuber DW ( % FW) 17.7 ± 0.2 18.9 ± 0.3 19.6 ± 0.3 18.8 ± 0.3 19.0 ± 0.2 19.5 ± 0.3 19.2 ± 0.3 18.9 ± 0.2
Tuber starch ( % FW)
8.2 ± 0.5 11.0 ± 1.2 13.2 ± 1.2 12.2 ± 1.5 11.2 ± 0.1 11.7 ± 0.2 11.1 ± 1.1 11.3 ± 0.5
Tuber starch ( % DW)
46.5 ± 3.7 61.3 ± 3.5 54.5 ± 2.1 64.9 ± 16.0 63.1 ± 4.3 60.1 ± 2.1 57.6 ± 7.9 59.9 ± 4.6
Tuber FW (g/plant) 1,628 ± 270 1,550 ± 290 1,654 ± 210 1,629 ± 330 1,869 ± 250 1,811 ± 260 1,760 ± 360 1,597 ± 300
Tuber DW (g/plant)
282.2 ± 15.9 292.6 ± 26.5 323.2 ± 29.3 306.9 ± 7.6 354.6 ± 31.9 353.1 ± 31.8 337.9 ± 30.4 301.8 ± 27.2
Tuber starch (g/plant)
134.0 ± 7.8 179.5 ± 5.7 176.6 ± 3.8 198.9 ± 24.6 224.0 ± 7.7 212.2 ± 3.4 194.6 ± 19.0 180.7 ± 7.7
Tuber FW (kg/hectare)
54,271 ± 8,929 70,880 ± 6,910 60,683 ± 3,634 54,292 ± 10,464 62,295 ± 8,324 60,368 ± 9,284 58,662 ± 6,055 54,222 ± 5,031
Tuber DW (kg/hectare)
9,606 ± 543 13,382 ± 1,292 11,876 ± 1,146 12,078 ± 1,046 11,824 ± 1,141 11,772 ± 1,136 11,263 ± 1,087 10,248 ± 579
Tuber starch (kg/hectare)
4,468 ± 181 8,209 ± 134 6,478 ± 89 7,851 ± 575 7,466 ± 180 7,072 ± 81 6,485 ± 444 6,135 ± 179
The results are the mean ± SE of extracts from 90 independent plants per line. Values that are signifi cantly different from the control plants are marked in bold.
1655
Susy impact in potato tuber starch metabolism
Plant Cell Physiol. 50(9): 1651–1662 (2009) doi:10.1093/pcp/pcp108 © The Author 2009.
Dow
nloaded from https://academ
ic.oup.com/pcp/article/50/9/1651/1854144 by guest on 05 February 2022

Maj
or C
HO
met
abol
ism
Min
or C
HO
met
abol
ism
TC
A /
org.
tran
sfor
mat
ion
Mito
chon
dria
l ele
ctro
n tr
ansp
ort
Cel
l wal
l
Met
al h
andl
ing
Tet
rapy
rrol
e sy
nthe
sis
Mis
cella
neou
s
Pro
tein
Sig
nalli
ngD
evel
opm
ent
Tra
nspo
rt
RN
A
Str
ess
Hor
mon
e m
etab
olis
m
Sec
onda
ry m
etab
olis
m
Lipi
d m
etab
olis
m
0
2
3
1
4
5
6
7
8
9
10
Num
ber
of d
ereg
ulat
ed g
enes
Aupregulateddownregulated
Pho
tosy
nthe
sis
Bupregulateddownregulated
0
Pho
tosy
nthe
sis
Maj
or C
HO
met
abol
ism
Min
or C
HO
met
abol
ism
TC
A /
org.
tran
sfor
mat
ion
Cel
l wal
l
Met
al h
andl
ing
Tet
rapy
rrol
e sy
nthe
sis
Red
ox r
egul
atio
nP
olya
min
e m
etab
olis
mM
isce
llane
ous
Pro
tein
Sig
nalli
ng
Dev
elop
men
tT
rans
port
Cel
l
DN
A
RN
A
Str
ess
Hor
mon
e m
etab
olis
m
Sec
onda
ry m
etab
olis
m
Lipi
d m
etab
olis
m
Gly
coly
sis
Am
ino
acid
met
abol
ism
2
4
6
8
10
Num
ber
of d
ereg
ulat
ed g
enes
12
14
16
Fig. 3 Functional categorization of the transcripts differentially expressed in (A) SuSy-overexpressing tubers (lines 5, 6 and 12) and (B) SuSy-antisensed tubers (lines S-112 and S-129) that were identifi ed in the transcriptomic analyses using the POCI 44K 60-mer oligo array ( http://pgrc.ipk-gatersleben.de/poci ). Full details of the microarray analysis are given in Supplementary Table S2 and Supplementary Table S3 . Signifi cantly down- and up-regulated transcripts (2-fold difference in SuSy-overexpressing tubers and 3-fold difference in SuSy-antisensed potato tubers) as compared with control were sorted by their putative functional category and compared with the whole genome according to the MapMan software. The number of deregulated genes in each categorical group is presented here. Up-regulated genes are in grey bars and down-regulated genes are in black bars. Genes from the ‘no hits found’ category were not included in the graphic. Plants were grown in greenhouse conditions.
1656
E. Baroja-Fernández et al.
Plant Cell Physiol. 50(9): 1651–1662 (2009) doi:10.1093/pcp/pcp108 © The Author 2009.
Dow
nloaded from https://academ
ic.oup.com/pcp/article/50/9/1651/1854144 by guest on 05 February 2022

in response to identical cues and (ii) the balance between these pathways is a major determinant of starch accumula-tion that explains the high-starch and low-starch pheno-types of the SuSy-overexpressing and SuSy-antisensed potato tubers, respectively.
Changes in SuSy activity do not affect the redox status of ADPG pyrophosphorylase Trehalose-6-phosphate is a sugar signal of emerging signifi -cance that regulates starch synthesis via thioredoxin-mediated post-translational redox activation of ADPG pyrophosphorylase ( Schluepmann et al. 2003 , Kolbe et al. 2005 , Paul 2007 ). When potato tuber extracts are separated on non-reducing SDS gels, ADPG pyrophosphorylase is pres-ent as a mixture of 50 kDa monomers (active form) and 100 kDa dimers formed by intermolecular links involving cysteine bridges ( Tiessen et al. 2002 ). Dimerization leads to inactiva-tion of the enzyme, which can be reactivated in vitro by incubating extracts with dithiothreitol ( Tiessen et al. 2002 ). In leaves, reactivation of ADPG pyrophosphorylase can take place by short-term exposure to light and by feeding trehalose-6-phosphate to intact chloroplasts ( Hendriks et al. 2003 , Kolbe et al. 2005 ). Because UDPG is the direct precursor for trehalose-6-phosphate, we hypothesized that changes in UDPG-producing SuSy activity would lead to concomitant changes in trehalose-6-phosphate levels that, in turn, will affect the redox status of ADPG pyrophosphorylase. To test this assumption, extracts from control and SuSy-antisensed potato tubers were separated on reductive and non-reductive SDS–PAGE and subsequently subjected to western blot analyses of ADPG pyrophosphorylase. As shown in Fig. 5 , these analyses revealed that both control and SuSy-anti-sensed potato tubers accumulate similar amounts of 50 kDa monomer and 100 kDa dimer. When extracts were obtained and separated under reducing conditions (including dithio-threitol), the immunoreactive signal was largely found at 50 kDa. These results indicate that redox status of ADPG pyro-phosphorylase in potato tubers does not depend on intrac-ellular levels of UDPG produced by SuSy. In addition, these results indicate that changes in starch content in tubers with altered SuSy activity ( Figs. 1, 2 ) cannot be ascribed to changes in the redox status of ADPG pyrophosphorylase.
Sus4 is a cytosolic enzyme SuSy is considered to be a cytosolic protein. However, recent studies have shown that some SuSy forms may occur in the mitochondrial compartment ( Subbaiah et al. 2006 ). The fact that some SuSy isoforms may be distributed in organelle(s) not involved in sucrose metabolism suggests that SuSy has novel role(s) beyond sucrose degradation ( Subbaiah et al. 2006 ). Whether results presented in this work are ascribed to SuSy involvement in sucrose metabolism requires subcel-lular localization studies to confi rm (or not) that this enzyme
is located in the cytosol. Therefore, we carried out confocal fl uorescence microscopy analyses of potato plants constitu-tively expressing a translationally fused Sus4–green fl uroes-cent protein (GFP)-encoding gene under the control of the caulifl ower mosaic virus 35S promoter. As shown in Fig. 6 , fl uorescence in Sus4–GFP-expressing cells was distributed uniformly throughout the cytosol. In addition, GFP-positive strands randomly traversed the vacuolar space in Sus4–GFP-expressing cells ( Supplementary Fig. S5 ), the overall data strongly indicating that Sus4–GFP has a cytosolic localization.
Materials and Methods
Plants, growth conditions and sampling The work was carried out using potato ( S. tuberosum L. cv Désirée) plants transformed with 35S-SuSy-NOS (lines 3, 4, 5, 6, 7, 10 and 12) ( Muñoz et al. 2005 ) as well as S-112 and S-129 plants silenced for sus4 expression ( Zrenner et al. 1995 ). 35S-SuSy-NOS plants constitutively express the sus4 gene, the product of which localizes mainly in storage tissues of potato tubers ( Fu and Park 1995 ). We also used potato plants trans-formed with pSus4-GFP, which was produced as indicated in Supplementary Fig. S6 . Transformations of potato plants were conducted as described by Rocha-Sosa et al. (1989) . Transgenic plants were selected on kanamycin-containing medium.
Plants were grown both in greenhouse and fi eld condi-tions. Large-scale greenhouse trials were performed with plants individually grown in 5 liter pots (10 plants/line) under a light regime with a minimum of 250 µmol
Control 1265
35S-SuSy-NOS
0
25
50
75
100
125
150
175
200
225
Aci
d In
vert
ase
(mU
/g f
resh
wei
gh
t)
112
anti-SuSy
Fig. 4 Acid invertase activity in SuSy-overexpressing (lines 5, 6 and 12) and SuSy-antisensed (line S-112) potato tubers. The results are the mean ± SE of three independent experiments.
1657
Susy impact in potato tuber starch metabolism
Plant Cell Physiol. 50(9): 1651–1662 (2009) doi:10.1093/pcp/pcp108 © The Author 2009.
Dow
nloaded from https://academ
ic.oup.com/pcp/article/50/9/1651/1854144 by guest on 05 February 2022

photons m –2 at 22 ° C. Field trials were conducted in the vicinity of Sartaguda (Navarre, Spain) between May and Sep-tember of 2006 and 2007, using 90 plants per line. Plants from each line were randomly distributed in three 50 m 2 par-cels, each containing 30 plants per line. The distance between plants of the same row was 35 cm and the distance between rows was 90 cm. Analyses of enzyme activities and measure-ment of sugar content were carried out using mature tubers of 30–50 g FW immediately after harvest. The term ‘mature tubers’ refers to tubers harvested from senescent plants. Tissue cylinders were obtained from harvested mature tubers using a stainless-steel cork borer, nominally 19.1 mm in diameter. Tissue samples were taken from the center part of cylinders cut perpendicular to the long axis of the tuber, to avoid the large textural and composition differences occurring between the cortex and pith tissues ( Anzaldúa-Morales et al. 1992 ). Three samples were taken from each tuber, immediately freeze-clamped and fi nally ground to a fi ne powder in liquid nitrogen with a pestle and mortar.
Enzyme assays One gram of the frozen powder was resuspended at 4 ° C in 5 ml of 100 mM HEPES (pH 7.5), 2 mM EDTA and 5 mM dithiothreitol. The suspension was desalted and assayed for enzymatic activity. We verifi ed that this procedure did not result in loss of enzymatic activity by comparing activity in extracts prepared from the frozen powder with extracts pre-pared by homogenizing fresh tissue in extraction medium. ADPG pyrophosphorylase, SuSy, sucrose-phosphate syn-thase, ADP-sugar pyrophosphatase, UDPG pyrophosphory-lase, alkaline pyrophosphatase, alkaline invertase, starch synthase, starch phosphorylase and total amylolytic activi-ties were assayed as described by Baroja-Fernández et al. (2004) and Muñoz et al. (2005) . Acid invertase was assayed as described by Zrenner et al. (1995) . All enzymatic reactions were performed at 37 ° C. One unit (U) is defi ned as the amount of enzyme that catalyzes the production of 1 µmol of product per min.
Determination of soluble sugars A 0.5 g aliquot of the frozen powdered tissue was resus-pended in 0.4 ml of 1.4 M HClO 4 , left at 4 ° C for 2 h and cen-trifuged at 10,000 × g for 5 min. The supernatant was neutralized with K 2 CO 3 , centrifuged at 10,000 × g and sub-jected to measurement analyses of nucleotide-sugars, fruc-tose-6-phosphate, glucose-1-phosphate and glucose-6-phosphate. For measurement of sucrose, glucose and fructose, a 0.1 g aliquot of the frozen powder was resuspended in 1 ml of 70 % ethanol, left at 70 ° C for 90 min and centrifuged at 13,000 × g for 10 min. Sucrose, glucose, fructose and hexose-phosphates from supernatants were determined by HPLC with pulsed amperometric detection on a DX-500 Dionex system as described by Baroja-Fernández et al. (2004) . ADPG and UDPG were measured as described by Muñoz et al. (2005) by using either one of the following methods: Assay A: by HPLC on a system obtained from P.E. Waters and Associ-ates fi tted with a Partisil-10-SAX column; Assay B: by HPLC with pulsed amperometric detection on a DX-500 system (Dionex) fi tted to a CarboPac PA 10 column. 3-phosphoglyc-eric acid was measured as described by Muñoz et al. (2005) .
Analytical procedures Protein content was determined by the Bradford method using a Bio-Rad XL-100 prepared reagent (Bio-Rad, San Diego, CA, USA). Starch was measured by using an amylog-lucosydase–based test kit (Sigma-Aldrich Chemical Co., St. Louis, MO, USA).
Western blot analyses For immunoblot analyses, protein samples were separated on 12 % SDS–PAGE, transferred to nitrocellulose fi lters, and immunodecorated by using antisera raised against maize ADPG pyrophosphorylase as primary antibody, and a goat anti-rabbit IgG alkaline phosphatase conjugate (Sigma) as secondary antibody. In the case of western blots of ADPG pyrophosphorylase, samples were extracted and separated on SDS–PAGE under reducing/non-reducing conditions essentially as described by Tiessen et al. (2002) .
Microarrays RNA amplifi cation, labeling and statistical data analysis were performed basically as described by Adie et al. (2007) . Microarray slides Agilent POCI 4 × 44 (015425) that contain 246,000 expressed sequence tags corresponding to 46,345 unigenes ( http://pgrc.ipk-gatersleben.de/poci ) ( Klooster-man et al. 2008 ) were used for hybridization. Labeling and hybridization conditions were those described in ‘The manual two color microarray based gene expression Analy-sis’ of Agilent Technologies. Three independent biological replicates were hybridized for sus4 -overexpressing and -silenced lines as well as for the controls. Images from Cy3
100
Non-reducing Reducing
50
anti-
Sus4
WT
anti-
Sus4
WT
KDa
Fig. 5 Western blot analysis of ADPG pyrophosphorylase in control and SuSy-antisensed tubers in non-reducing and reducing conditions (after addition of 10 mM dithiothreitol).
1658
E. Baroja-Fernández et al.
Plant Cell Physiol. 50(9): 1651–1662 (2009) doi:10.1093/pcp/pcp108 © The Author 2009.
Dow
nloaded from https://academ
ic.oup.com/pcp/article/50/9/1651/1854144 by guest on 05 February 2022

and Hyper5 channels were equilibrated for intensity differ-ences and captured with a GenePix 4000B scanner (Axon). Spots were quantifi ed using GenPix software (Axon) and normalized using the Lowess method. The mean of the three replicate log-ratio intensities and their standard deviations were calculated and the expression data was statistically analyzed using the LIMMA package ( Smyth and Speed 2003 ). Functional characterization of the differentially expressed genes was carried out using the Mapman tool ( http://gabi.rzpd.de/projects/MapMan/ ) .
Real-time quantitative PCR Total RNA was extracted from potato tubers as for microar-ray experiments. RNA was treated with RNase free DNase (Takara). RNA (1.5 µg) was reverse transcribed using poly(T) primers and the Expand Reverse Transcriptase kit (Roche) according to the manufacturer’s instructions. Real time quantitative PCR was performed using a 7900HT sequence
detector system (Applied Biosystems) with the SYBR Green PCR Master Mix (Applied Biosystems) according to the manufacturer’s protocol. Each reaction was performed in triplicate with 0.4 µl of fi rst-strand cDNA in a total volume of 20 µl. The specifi city of the PCR amplifi cation was checked with a heat dissociation curve (from 60 ° C to 95 ° C). Com-parative threshold values were normalized to EF1- α internal control and compared to obtain relative expression levels. The specifi city of the obtained RT–PCR products was con-trolled on 1.8 % agarose gels. Primers used for PCRs are listed in Supplementary Table S4 .
Confocal microscopy Subcellular localization of Sus4–GFP was performed using a D-Eclipse C1 confocal microscope (NIKON, Japan) equipped with standard Ar 488 laser excitation, BA515/30 fi lter for green emission, a BA650LP fi lter for red emission and trans-mitted light detector for bright fi eld images.
Fig. 6 Cytosolic localization of Sus4–GFP. In (A, A1 and A2), general overview of Sus4–GFP localization. In (B), (C, C1) and (D, D1), detailed views of Sus4–GFP localization in leaf epidermal cells, protoplasts and mesophyll cells, respectively. (A, B, C1) Green fl uorescence of GFP. (A1,D) Red chlorophyll autofl uorescence. (A2, D1) Merged images of GFP fl uorescence with red chlorophyll autofl uorescence. (C) Bright-fi eld image. Bar = 10 µm. Results presented in Supplementary Fig. S5 further confi rmed that Sus4–GFP has a cytosolic localization.
1659
Susy impact in potato tuber starch metabolism
Plant Cell Physiol. 50(9): 1651–1662 (2009) doi:10.1093/pcp/pcp108 © The Author 2009.
Dow
nloaded from https://academ
ic.oup.com/pcp/article/50/9/1651/1854144 by guest on 05 February 2022

Funding
This research was supported by grants BIO2004-01922 and BIO2007-63915 from the Comisión Interministerial de Cien-cia y Tecnología and Fondo Europeo de Desarrollo Regional (Spain) and the grant IIM010491.RI1from the Government of Navarra. M.O. was partly supported by grant No. APVV-0432-06 from the Grant Agency APVV.
Acknowledgments
We are very thankful to Dr. R.N. Trethewey who kindly sup-plied the S-112 and S-129 transgenic potato plants defi cient in SuSy. We thank Dr. C. Hannah (University of Florida) who kindly provided us the antibody raised against maize ADPG pyrophosphorylase. E.E. expresses his gratitude to the Span-ish Ministry of Culture and Education, to the Consejo Supe-rior de Investigaciones Científi cas and to the Public University of Nafarroa for their support. We express our gratitude to María Angeles Barado, Jéssica Díaz de Cerio and Cristina Cirauqui for expert technical support. Finally, we all thank Professor T. Akazawa for his enthusiastic and fruitful discus-sions during the initial course of this study.
References
Adie , B.A.T. , Pérez-Pérez , J. , Pérez-Pérez , M. , Godoy , M. , Sánchez-Serrano , J.J. , Schmelz , E.A. , et al . ( 2007 ) ABA is an essential signal for plant resistance to pathogens affecting JA biosynthesis and the activation of defenses in Arabidopsis . Plant Cell 19 : 1665 – 1681 .
Al-Saikhan , M.S. , Howard , L.R. and Miller , J.C. ( 1995 ) Antioxidant activity and total phenolics in different genotypes of potato ( Solanum tuberosum L.) . J . Food Sci. 60 : 341 – 343 .
Amor , Y. , Haigler , C.H. , Johnson , S. , Wainscott , M. and Delmer , D.P. ( 1995 ) A membrane-associated form of sucrose synthase and its potential role in synthesis of cellulose and callose in plants. Proc . Natl Acad . Sci . USA 92 : 9353 – 9357 .
Andre , C.M. , Ghislain , M. , Bertin , P. , Oufi r , M. , Herrera , M.R. , Hoffmann , L. , et al . ( 2007 ) Andean potato cultivars ( Solanum tuberosum L.) as a source of antioxidant and mineral micronutrients. J . Agric . Food Chem. 55 : 366 – 378 .
Anzaldúa-Morales , A. , Bourne , M.C. and Shomer , I. ( 1992 ) Cultivar specifi c gravity and location in tuber affect puncture force of raw potatoes. J . Food Sci. 57 : 1353 – 1356 .
Appeldoorn , N.J.G. , de Bruijn , S.M. , Koot-Gronsveld , E.A.M. , Visser , R.G.F , Vreugdenhil , D. and van der Plas , L.H.W. ( 1997 ) Developmental changes of enzymes involved in conversion of sucrose to hexose phosphate during early tuberisation of potato. Planta 202 : 220 – 226 .
Asano , T. , Kunieda , N. , Omura , Y. , Ibe , H. , Kawasaki , T. , Takano , M. , et al . ( 2002 ) Rice SPK, a calmodulin-like domain protein kinase, is required for storage product accumulation during seed development: phosphorylation of sucrose synthase is a possible factor . Plant Cell 14 : 619 – 628 .
Baroja-Fernández , E. , Muñoz , F.J. , Saikusa , T. , Rodríguez-López , M. , Akazawa , T. and Pozueta-Romero , J. ( 2003 ) Sucrose synthase
catalyzes the de novo production of ADPglucose linked to starch biosynthesis in heterotrophic tissues of plants. Plant Cell Physiol. 44 : 500 – 509.
Baroja-Fernández , E. , Muñoz , F.J. , Zandueta-Criado , A. , Morán-Zorzano , M.T. , Viale , A.M. , Alonso-Casajús , N. , et al . ( 2004 ) Most of ADP-glucose linked to starch biosynthesis occurs outside the chloroplast in source leaves. Proc . Natl Acad . Sci . USA 101 : 13080 – 13085 .
Bollen , M. , Keppens , S. and Stalmans , W. ( 1998 ) Specifi c features of glycogen metabolism in the liver. Biochem . J. 336 : 19 – 31 .
Bracho , G.E. and Whitaker , J.R. ( 1990 ) Purifi cation and partial characterization of potato ( Solanum tuberosum ) invertase and its endogenous proteinaceous inhibitor. Plant Physiol. 92 : 386 – 394 .
Chourey , P.S. and Nelson , O.E. ( 1976 ) The enzymatic defi ciency conditioned by the shrunken -1 mutations in maize. Biochem . Genet. 14 : 1041 – 1055 .
Ciereszko , I. and Kleczkowski , L.A. ( 2002 ) Glucose and mannose regulate the expression of a major sucrose synthase gene in Arabidopsis via hexokinase-dependent mechanisms. Plant Physiol . Biochem. 40 : 907 – 911 .
Déjardin , A. , Sokolov , L.N. and Kleczkowski , L.A. ( 1999 ) Sugar/osmoticum levels modulate differential abscisic acid-independent expression of two stress-responsive sucrose synthase genes in Arabidopsis . Biochem . J. 344 : 503 – 509 .
Delmer , D.P. and Albersheim , P. ( 1970 ) The biosynthesis of sucrose and nucleoside diphosphate glucoses in Phaseolus vulgaris . Plant Physiol. 45 : 782 – 786 .
Fu , H. and Park , W.D. ( 1995 ) Sink- and vascular-associated sucrose synthase functions are encoded by different gene classes in potato. Plant Cell 7 : 1369 – 1385 .
Geigenberger , P. , Reimholz , R. , Geiger , M. , Merlo , L. , Canale , V. and Stitt , M. ( 1997 ) Regulation of sucrose and starch metabolism in potato tubers in response to short-term water defi cit. Planta 201 : 502 – 518 .
Geiger , M. , Stitt , M. and Geigenberger , P. ( 1998 ) Metabolism in slices from growing potato tubers responds differently to addition of sucrose and glucose. Planta 206 : 234 – 244 .
Guedon , E. , Desvaux , M. and Petitdemange , H. ( 2000 ) Kinetic analysis of Clostridium cellulolyticum carbohydrate metabolism: importance of glucose-1-phosphate and glucose-6-phosphate branch points for distribution of carbon fl uxes inside and outside cells revealed by steady-state continuous culture. J . Bacteriol. 182 : 2010 – 2017 .
Halford , N.G. and Paul , M.J. ( 2003 ) Carbon metabolite sensing and signalling. Plant Biotechnol . J. 1 : 381 – 398 .
Hardin , S.C. , Winter , H. and Huber , S.C. ( 2004 ) Phosphorylation of the amino terminus of maize sucrose synthase in relation to membrane association and enzyme activity. Plant Physiol. 134 : 1427 – 1438 .
Hendriks , J.H.M. , Kolbe , A. , Gibon , Y. , Stitt , M. and Geigenberger , P. ( 2003 ) ADP-glucose pyrophosphorylase is activated by posttranslational redox-modifi cation in response to light and to sugars in leaves of Arabidopsis and other plant species. Plant Physiol. 133 : 838 – 849 .
Herbers , K. and Sonnewald , U. ( 1998 ) Molecular determinants of sink strength. Curr . Opin . Plant Biol. 1 : 207 – 216 .
Kleczkowski , L.A. ( 1994 ) Glucose activation and metabolism through UDP-glucose pyrophosphorylase in plants. Phytochemistry 37 : 1507 – 1515 .
Kloosterman , B. , De Koeyer , D. , Griffi ths , R. , Flinn , B. , Steuernagel , B. , Scholz , U. , et al . ( 2008 ) Genes driving potato tuber initiation and
1660
E. Baroja-Fernández et al.
Plant Cell Physiol. 50(9): 1651–1662 (2009) doi:10.1093/pcp/pcp108 © The Author 2009.
Dow
nloaded from https://academ
ic.oup.com/pcp/article/50/9/1651/1854144 by guest on 05 February 2022

growth: identifi cation based on transcriptomal changes using the POCI array. Funct . Integr . Genomcs 8 : 329 – 340 .
Kolbe , A. , Tiessen , A. , Schluepmann , H. , Paul , M. , Ulrich , S. and Geigenberger , P. ( 2005 ) Trehalose 6-phosphate regulates starch synthesis via posttranslational redox activation of ADP-glucose pyrophosphorylase. Proc . Natl Acad . Sci . USA 102 : 11118 – 11123 .
Konishi , T. , Ohmiya , Y. and Hayashi , T. ( 2004 ) Evidence that sucrose loaded into the phloem of a poplar leaf is used directly by sucrose synthase associated with various β -glucan synthases in the stem. Plant Physiol. 134 : 1146 – 1152 .
Morris , W.L. , Ducreux , L. , Griffi ths , D. , Stewart , D. , Davies , H.V. and Taylor , M. ( 2004 ) Carotenogenesis during tuber development and storage in potato. J . Exp . Bot. 399 : 975 – 982 .
Müller-Röber , B. , Sonnewald , U. and Willmitzer , L. ( 1992 ) Inhibition of the ADP-glucose pyrophosphorylase in transgenic potato leads to sugar-storing tubers and infl uences tuber formation and expression of tuber storage protein genes. EMBO J. 11 : 1229 – 1238 .
Muñoz , F.J. , Baroja-Fernández , E. , Ovecka , M. , Li , J. , Mitsui , T. , Sesma , M.T. , et al . ( 2008 ) Plastidial localization of a potato ‘Nudix’ hydrolase of ADPglucose linked to starch biosynthesis. Plant Cell Physiol. 49 : 1734 – 1746 .
Muñoz , F.J. , Baroja-Fernández , E. , Morán-Zorzano , M.T. , Viale , A.M. , Etxeberria , E. , Alonso-Casajús , N. , et al . ( 2005 ) Sucrose synthase controls the intracellular levels of ADPglucose linked to transitory starch biosynthesis in source leaves. Plant Cell Physiol. 46 : 1366 – 1376 .
Muñoz , F.J. , Morán-Zorzano , M.T. , Alonso-Casajús , N. , Baroja-Fernández , E. , Etxeberria , E. and Pozueta-Romero , J. ( 2006 ) New enzymes, new pathways and an alternative view on starch biosynthesis in both photosynthetic and heterotrophic tissues of plants . Biocatal. Biotransform. 24 : 63 – 76 .
Naeem , M. , Tetlow , I.J. and Emes , M.J. ( 1997 ) Starch synthesis in amyloplasts from developing potato tubers. Plant J. 11 : 1095 – 1103.
Nakai , T. , Konishi , T. , Zhang , X.-Q. , Chollet , R. , Tonouchi , N. , Tsuchida , T. , et al . ( 1998 ) An increase in apparent affi nity for sucrose of mung bean sucrose synthase is caused by in vitro phosphorylation or directed mtagenesis of Ser 11 . Plant Cell Physiol. 39 : 1337 – 1341 .
Neckelmann , G. and Orellana , A. ( 1998 ) Metabolism of uridine 5 ′ -diphosphate-glucose in Golgi vesicles from pea stems. Plant Physiol. 117 : 1007 – 1014 .
Neuhaus , H.E. , Henrichs , G. and Scheibe , R. ( 1995 ) Starch degradation in intact amyloplasts isolated from caulifl ower fl oral buds ( Brassica oleracea L.) . Planta 195 : 496 – 504 .
Paul , M. ( 2007 ) Trehalose 6-phosphate. Curr . Opin . Plant Biol. 10 : 303 – 309 .
Pontis , H.G. , Babio , J.R. and Salerno , G. ( 1981 ) Reversible unidirectional inhibition of sucrose synthase activity by disulfi des. Proc . Natl Acad . Sci . USA 78 : 6667 – 6669 .
Porchia , A.C. , Curatti , L. and Salerno , G.L. ( 1999 ) Sucrose biosynthesis in a prokaryotic organism: presence of two sucrose-phosphate synthases in Anabaena with remarkable differences compared with the plant enzymes. Planta 210 : 34 – 40 .
Pozueta-Romero , J. and Akazawa , T. ( 1993 ) Biochemical mechanism of starch biosynthesis in amyloplasts from cultured cells of sycamore ( Acer pseudoplatanus ) . J . Exp . Bot. 44S : 297 – 306 .
Pozueta-Romero , J. , Frehner , M. , Viale , A. and Akazawa , T. ( 1991 ) Direct transport of ADPglucose by an adenylate translocator is linked to
starch biosynthesis in amyloplasts. Proc . Natl Acad . Sci . USA 88 : 5769 – 5773 .
Prat , S. , Frommer , W.B. , Höfgen , R. , Keil , M. , Kossmann , J. , Köster-Töpfer , M. , et al . ( 1990 ) Gene expression during tuber development in potato plants. FEBS Lett. 268 : 334 – 338 .
Preiss , J. ( 1988 ) Biosynthesis of starch and its regulation. In The Biochemistry of Plants . Edited by Preiss , J. pp. 181 – 254. Academic Press , New York .
Purcell , P.C. , Smith , A.M. and Halford , N.G. ( 1998 ) Antisense expression of a sucrose non-fermenting-1-related protein kinase sequence in potato results in decreased expression of sucrose synthase in tubers and loss of sucrose-inducibility of sucrose synthase transcripts in leaves. Plant J. 14 : 195 – 202 .
Regierer , B. , Fernie , A.R. , Springer , F. , Perez-Melis , A. , Leisse , A. , Koehl , K. , Willmitzer , L. , Geigenberger , P. and Kossmann , J. ( 2002 ) Starch content and yield increase as a result of altering adenylate pools in transgenic plants. Nat . Biotechnol. 20 : 1256 – 1260 .
Rocha-Sosa , M. , Sonnewald , U. , Frommer , W.B. , Stratmann , M. , Schell , J. , et al . ( 1989 ) Both developmental and metabolic signals activate the promoter of a class I patatine gene. EMBO J. 8 : 23 – 29 .
Schluepmann , H. , Pellny , T. , van Dijken , A. , Smeekens , S. and Paul , M. ( 2003 ) Trehalose 6-phosphate is indispensable for carbohydrate utilization and growth in Arabidopsis thaliana . Proc . Natl Acad . Sci . USA 100 : 6849 – 6854 .
Slabnik , E. , Frydman , R.B. and Cardini , C.E. ( 1968 ) Some properties of potato tuber UDPG:D-fructose-2-glucosyltransferase (E.C. 2.4.1.14) and UDPG:D-fructose-6-phosphate-2-glucosyltransferase (E.C. 2.4.1.13). Plant Physiol. 43 : 1063 – 1068 .
Smyth , G.K. and Speed , T. ( 2003 ) Normalization of cDNA microarray data. Methods 31 : 265 – 273 .
Stark , D.M. , Timmerman , K.P. , Barry , G.F. , Preiss , J. and Kishore , G.M. ( 1992 ) Regulation of the amount of starch in plant tissues by ADPglucose pyrophosphorylase. Science 258 : 287 – 292 .
Subbaiah , C.C. , Palaniappan , A. , Duncan , K. , Rhoads , D.M. , Huber , S.C. and Sachs , M. M . ( 2006 ) Mitochondrial localization and putative signaling function of sucrose synthase in maize. J . Biol . Chem. 281 : 15625 – 15635 .
Sweetlove , L.J. , Burrell , M.M. and ap Rees , T. ( 1996 ) Starch metabolism in tubers of transgenic potato ( Solanum tuberosum ) with increased ADPglucose pyrophosphorylase. Biochem . J. 320 : 493 – 498 .
Sweetlove , L.J. , Kruger , N.J. and Hill , S.A. ( 2001 ) Starch synthesis in transgenic potato tubers with increased 3-phosphoglyceric acid content as a consequence of increased 6-phosphofructokinase activity. Planta 213 : 478 – 482 .
Tauberger , E. , Fernie , A.R. , Emmermann , M. , Renz , A. , Kossmann , J. , Willmitzer , L. , et al . ( 2000 ) Antisense inhibition of plastidial phosphoglucomutase provides compelling evidence that potato tuber amyloplasts import carbon from the cytosol in the form of glucose-6-phosphate. Plant J. 23 : 45 – 53 .
Thévenot , C. , Simond-Côte , E. , Reyss , A. , Manicacci , D. , Trouverie , J. , Le Guilloux , M. , et al . ( 2005 ) QTLs for enzyme activities and soluble carbohydrates involved in starch accumulation during grain fi lling in maize. J . Exp . Bot. 56 : 945 – 958 .
Thimm , O. , Bläsing , O. , Gibon , Y. , Nagel , A. , Meyer , S. , Krüger , P. , et al . ( 2004 ) MAPMAN: a user-driven tool to display genomics data sets onto diagrams of metabolic pathway and other biological processes. Plant J. 37 : 914 – 939 .
1661
Susy impact in potato tuber starch metabolism
Plant Cell Physiol. 50(9): 1651–1662 (2009) doi:10.1093/pcp/pcp108 © The Author 2009.
Dow
nloaded from https://academ
ic.oup.com/pcp/article/50/9/1651/1854144 by guest on 05 February 2022

Tiessen , A. , Hendriks , J.H.M. , Stitt , M. , Branscheid , A. , Gibon , Y. , Farré , E.M. , et al . ( 2002 ) Starch synthesis in potato tubers is regulated by post-translational redox modifi cation of ADP-glucose pyrophosphorylase: a novel regulatory mechanism linking starch synthesis to the sucrose supply. Plant Cell 14 : 2191 – 2213 .
Tiessen , A. , Prescha , K. , Branscheid , A. , Palacios , N. , McKibbin , R. , Halford , N.G. , et al . ( 2003 ) Evidence that SNF1-related kinase and hexokinase are involved in separate sugar-signalling pathways modulating post-translational redox activation of ADP-glucose pyrophosphorylase in potato tubers. Plant J. 35 : 490 – 500.
Trethewey , R.N. , Fernie , A.R. , Bachmann , A. , Fleischer-Notter , H. , Geigenberger , P. and Willmitzer , L. ( 2001 ) Expression of a bacterial sucrose phosphorylase in potato tubers results in a glucose-independent induction of glycolysis. Plant Cell Environ. 24 : 357 – 365 .
Trethewey , R.N. , Geigenberger , P. Hajirezaei , M. , Sonnewald , U. , Stitt , M. , Riesmeier , J. , et al . ( 1998 ) Combined expression of glucokinase and invertase in potato tubers leads to a dramatic reduction in starch accumulation and a stimulation of glycolysis. Plant J. 15 : 109 – 118 .
Zervosen , A. , Römer , U. and Elling , L. ( 1998 ) Application of recombinant sucrose synthase-large scale synthesis of ADP-glucose. J . Mol . Catalysis B: Enzymat ic 5 : 25 – 28 .
Zrenner , R. , Salanoubat , M. , Willmitzer , L. and Sonnewald , U. ( 1995 ) Evidence for the crucial role of sucrose synthase for sink strength using transgenic potato plants ( Solanum tuberosum L.) . Plant J. 7 : 97 – 107 .
Zrenner , R. , Willmitzer , L. and Sonnewald , U. ( 1993 ) Analysis of the expression of potato uridinediphosphate-glucose pyrophosphorylase and its inhibition by antisense RNA. Planta 190 : 247 – 252 .
(Received June 17, 2009; Accepted July 14, 2009)
1662
E. Baroja-Fernández et al.
Plant Cell Physiol. 50(9): 1651–1662 (2009) doi:10.1093/pcp/pcp108 © The Author 2009.
Dow
nloaded from https://academ
ic.oup.com/pcp/article/50/9/1651/1854144 by guest on 05 February 2022


















