
Energy metabolism and thermoregulation in Chinchilla brevicaudata
7
Journal of Thermal Biology 28 (2003) 489–495 Energy metabolism and thermoregulation in Chinchilla brevicaudata Arturo Cort ! es a, *, Carlos Tirado a , Mario Rosenmann b a Departamento de Biolog ! ıa, Facultad de Ciencias, Universidad de La Serena, Casilla 599, La Serena, Chile b Departamento de Ciencias Ecol ! ogicas, Facultad de Ciencias, Universidad de Chile, Casilla 653, Santiago, Chile Received 6 November 2002; accepted 6 June 2003 Abstract Chinchilla brevicaudata lives at 3500–5000 m, with high ambient temperatures during the day but cold at night. In this Andean habitat there is also low availability of food and water resources. Physiological attributes that may minimize their energetic cost as well as the water requirements are: (1) Low values of basal metabolic rate (67.2%) and thermal conductance (51.0%) compared to predicted values. (2) The aerobic metabolic expansivity was 5.1, while the calculated theoretical critical lethal temperature was extremely low (67.8 C). (3) The energetic cost for maintenance of water balance was 85.3% of the predicted value for xeric rodents of similar size. r 2003 Elsevier Ltd. All rights reserved. Keywords: Chinchilla brevicaudata; Basal metabolism; Maximum metabolism; Thermal conductance; Evaporative water loss 1. Introduction In xeric environments, the ambient temperature (T a ), the photoperiod and the availability of food and water are the main variables that present the largest daily and seasonal variations (Degen, 1997; Kronfeld-Schor et al., 2000). Consequently, organisms that live in these environments should be able to maintain homeostatic conditions, particularly in their water and energy equilibrium. In mammals, one of the most utilized energetic parameters is the basal metabolic rate (BMR), which is the main component of the energy spent under laboratory as well as in natural conditions (Cruz-Neto et al., 2001). For some wild mammals BMR may represent 50% of the daily energy expenditure (Nagy et al., 1999; Speakman, 2000). Variations of BMR among homeotherms have been basically explained by allometric relations of body mass (Kleiber, 1961). More recently the residual variance of this correlation has been applied at taxonomic level (Hayssen and Lacy, 1985) or in relation to food habits (McNab, 1986), ambient temperature (MacMillen and Garland, 1989), and habitat (McNab and Morrison, 1963; Hulbert and Dawson, 1974; Shkolnik and Schmidt-Nielsen, 1976; Lovegrove, 1986; Lovegrove et al., 1991). In nature most homeotherms keep their body tem- perature (T b ) within certain limits. This condition depends on some physiological characteristics, such as the metabolic rate (MR) and thermal conductance (C). McNab (1979) found that granivorous rodents from xeric habitats have low BMR, avoiding risks of hyperthermia and maintaining their water economy by minimizing evaporative water loss (EWL). This last feature has been greatly neglected in studies in South American mesic and xeric rodents, in spite of the fact that EWL plays an important role in thermoregulation and water balance (Degen, 1997). A few exceptions are the observations in some Chilean rodents (Rosenmann, 1977; Cort! es, 1985; Cort! es et al., 1988, 1990, 2000a, b; Bozinovic et al., 1995). In rodents, the metabolic water production (MWP) and the EWL are utilized to evaluate the efficiency ARTICLE IN PRESS *Corresponding author. E-mail addresses: [email protected] (A. Cort ! es), mrosen- [email protected] (M. Rosenmann). 0306-4565/03/$ - see front matter r 2003 Elsevier Ltd. All rights reserved. doi:10.1016/S0306-4565(03)00049-4
-
Upload
arturo-cortes -
Category
Documents
-
view
212 -
download
0
Transcript of Energy metabolism and thermoregulation in Chinchilla brevicaudata

Journal of Thermal Biology 28 (2003) 489–495
Energy metabolism and thermoregulation inChinchilla brevicaudata
Arturo Cort!esa,*, Carlos Tiradoa, Mario Rosenmannb
aDepartamento de Biolog!ıa, Facultad de Ciencias, Universidad de La Serena, Casilla 599, La Serena, ChilebDepartamento de Ciencias Ecol !ogicas, Facultad de Ciencias, Universidad de Chile, Casilla 653, Santiago, Chile
Received 6 November 2002; accepted 6 June 2003
Abstract
Chinchilla brevicaudata lives at 3500–5000m, with high ambient temperatures during the day but cold at night. In this
Andean habitat there is also low availability of food and water resources. Physiological attributes that may minimize
their energetic cost as well as the water requirements are: (1) Low values of basal metabolic rate (67.2%) and thermal
conductance (51.0%) compared to predicted values. (2) The aerobic metabolic expansivity was 5.1, while the calculated
theoretical critical lethal temperature was extremely low (�67.8�C). (3) The energetic cost for maintenance of waterbalance was 85.3% of the predicted value for xeric rodents of similar size.
r 2003 Elsevier Ltd. All rights reserved.
Keywords: Chinchilla brevicaudata; Basal metabolism; Maximum metabolism; Thermal conductance; Evaporative water loss
1. Introduction
In xeric environments, the ambient temperature (Ta),
the photoperiod and the availability of food and water
are the main variables that present the largest daily and
seasonal variations (Degen, 1997; Kronfeld-Schor et al.,
2000). Consequently, organisms that live in these
environments should be able to maintain homeostatic
conditions, particularly in their water and energy
equilibrium. In mammals, one of the most utilized
energetic parameters is the basal metabolic rate (BMR),
which is the main component of the energy spent under
laboratory as well as in natural conditions (Cruz-Neto
et al., 2001). For some wild mammals BMR may
represent 50% of the daily energy expenditure (Nagy
et al., 1999; Speakman, 2000).
Variations of BMR among homeotherms have been
basically explained by allometric relations of body mass
(Kleiber, 1961). More recently the residual variance of
this correlation has been applied at taxonomic level
(Hayssen and Lacy, 1985) or in relation to food habits
(McNab, 1986), ambient temperature (MacMillen and
Garland, 1989), and habitat (McNab and Morrison,
1963; Hulbert and Dawson, 1974; Shkolnik and
Schmidt-Nielsen, 1976; Lovegrove, 1986; Lovegrove
et al., 1991).
In nature most homeotherms keep their body tem-
perature (Tb) within certain limits. This condition
depends on some physiological characteristics, such as
the metabolic rate (MR) and thermal conductance (C).
McNab (1979) found that granivorous rodents from xeric
habitats have low BMR, avoiding risks of hyperthermia
and maintaining their water economy by minimizing
evaporative water loss (EWL). This last feature has been
greatly neglected in studies in South American mesic and
xeric rodents, in spite of the fact that EWL plays an
important role in thermoregulation and water balance
(Degen, 1997). A few exceptions are the observations in
some Chilean rodents (Rosenmann, 1977; Cort!es, 1985;
Cort!es et al., 1988, 1990, 2000a, b; Bozinovic et al., 1995).
In rodents, the metabolic water production (MWP)
and the EWL are utilized to evaluate the efficiency
ARTICLE IN PRESS
*Corresponding author.
E-mail addresses: [email protected] (A. Cort!es), mrosen-
[email protected] (M. Rosenmann).
0306-4565/03/$ - see front matter r 2003 Elsevier Ltd. All rights reserved.
doi:10.1016/S0306-4565(03)00049-4

of water regulation by means of the relation
Ta@MWP=EWL (MacMillen and Hinds, 1983), where
Ta@ is the ambient temperature when MWP/EWL=1.
If temperature and humidity are kept constant this index
is not affected by the animal’s activity (Raab and
Schmidt-Nielsen, 1972). The efficiency of water regula-
tion has also been expressed considering the energetic
cost of maintenance of water balance (MR-WB), taking
the value of Ta@ that permits the calculation MR-WB
from the relation between MR and the species-specific
ambient temperature (Cort!es et al., 2000b). Further-
more, the value of MR-WB permits the regulatory
efficiency between mesic and xeric rodents to be
determined (Cort!es et al., 2000b).
Chinchilla brevicaudata (Waterhouse, 1848) is an hystri-
comorphic nocturnal rodent dwelling between 3500 to
5000m above sea level in the Andes Range (Munoz and
Y!anez, 2000). Due to the special quality of the fur, the
species was intensively hunted and a great part of the
Chilean populations was extinguished or significantly
diminished (Jim!enez, 1996). Currently its distribution is
restricted to Southern Per!u, Northeast Argentina and to
the (I–III Regi!on) of Northern Chile (Redford and
Einsenberg, 1989). Conservation problems have been
noted and it has been considered in critical danger in
Argentina (Garc!ıa et al., 1997) and in risk of extinction in
Chile (Glade, 1993; SAG, 2000). Ecophysiological studies
in C. brevicaudata are few, but some ecological and
conservation aspects have been reported (Jim!enez, 1996).
The combined effects of physiological, morphological,
behavioral and ecological attributes allow desert
rodents to minimize the energetic costs and assure their
survival in these harsh environments (Bozinovic and
Contreras, 1990; Prakash, 2001). Because of the extreme
climatic conditions of the Andean range, we hypothe-
sized that C. brevicaudata would present most, if not all
of these features. To test this idea we measured oxygen
consumption in normal air and under He–O2 (80–20%)
atmosphere. EWL and body temperature (Tb) were also
measured at different ambient temperatures (Ta). In
addition, two indices Ta@ (MWP=EWL), that repre-
sents the efficiency of water regulation, and MR-WB,
representing the energetic cost of maintaining the water
balance, were estimated.
2. Materials and methods
2.1. Experimental animals
Five individuals of C. brevicaudata (2##, 3~~) were
captured with National traps in the locality of El Morro
Negro (25�000S; 69�450W) in the National Park Llul-
laillaco (II Regi !on, South East of Antofagasta, Chile),
between 3000 and 5000m of altitude. This area is
characterized by a perarid climate (di Castri and Hajek,
1976), with an annual precipitation of 20–50mm
(Messerli et al., 1993) and a mean annual temperature
of 2�C (Luebert, 1998). At the site of capture we found
scant vegetation, covering less than 9% of the ground
surface. One of us (AC) found the following proportions
of shrubs and herbs: Baccharis tola (0.05%), Adesmia
eranicea (0.08%), Adesmia caespitosa (0.05%), Cristaria
andicola (0.06%), Fabiana bryoides (5.10%) and Stipa
chrysophylla (3.55%). The captured animals were
transported to the laboratory and maintained under
natural photoperiod in individual cages, with water and
food (barley and alfalfa) ad libitum. At the laboratory,
ambient temperature was 21.073�C, while relative
humidity averaged 60%.
2.2. Energy metabolism
All oxygen consumption measurements were conducted
individually with a modified automatic closed-system
respirometer, based on the manometric design of Morrison
(1951). Animals were in postabsortive state (2–3h after
feeding). Different ambient temperatures (Ta) were main-
tained within70.1�C in a water–glycol bath in which themetabolic chambers were submerged. Average body mass
of our five experimental animals was 454.4762.5 g. Bodytemperature (Tb) was recorded before and after each
ARTICLE IN PRESS
Nomenclature
BMR basal metabolic rate (ml O2/g h)
C thermal conductance (ml O2/g h�C)
CHe thermal conductance in He–O2 atmosphere
(ml O2/g h�C)
EWL evaporative water loss (mg H2O/g h)
mb body mass (g)
MR metabolic rate (ml O2 /g h)
MMR maximum metabolic rate (ml O2/g h)
MWP metabolic water production (mg H2O/g h)
Ta ambient temperature (�C)
Ta@ ambient temperature (�C), when MWP/
EWL=1
Tb body temperature (�C)
Tic calculated theoretical lower critical tempera-
ture (�C)
TLL calculated theoretical lower lethal temperature
(�C)
MR-WB energetic cost of maintaining the water balance
(cal/g h)
DTm minimum thermal differential between Tb and
Tic (�C)
A. Cort!es et al. / Journal of Thermal Biology 28 (2003) 489–495490
metabolic run with a Cole Parmer, Model 8500-40 copper-
constant thermocouple. O2 consumption was also mea-
sured in 80% He–20% O2 atmosphere, which has been
used to obtain the maximum metabolic rate of thermo-
regulation (MMR) (Rosenmann and Morrison, 1974;
Holloway and Geiser, 2001). For this purpose we used a
Ta range of 5�C to �7.5�C (Bozinovic and Rosenmann,
1989). Metabolic values were determined from the average
of the three minimum periods of 3–5min of each
experimental trial which lasted 1–3h (Rosenmann and
Morrison, 1974). BMR was similarly estimated from the
lowest three MR values when independence of Ta was
determined. Thermal conductance (C) was calculated from
the slope of the regression (MR vs. Ta), below thermo-
neutrality. The lower critical temperature (Tic) was inferred
by the intersection of C with BMR. Values of BMR, C;and the endothermic limit were compared with the
expected values for mammals of similar size using
the relations: BMR=3.42m�0:25b (Kleiber, 1961) and C ¼
1:0m�0:50b (McNab and Morrison, 1963). The ratio BMR/
C that indicates the minimum thermal differential (DTm),
between Tb and Tic; was also calculated: DTm¼ BMR=C
(�C)=3.42m0:25b ¼ Tb2Tic (McNab, 1979).
2.3. Evaporative water loss
Values of EWL were obtained gravimetrically
(70.1mg) from the average of three minimum 5min
periods during 2–3 h of measurements, following the
method of Hainsworth (1968). EWL trials were con-
ducted at different Ta (5�C, 15�C, 20�C, 25�C, 30�C
and 32.5�C), with an air flow of 5 l/min. All measure-
ments started after 1 h of thermal equilibrium (Cort!es
et al., 1990). Results of minimum EWL were compared
with the expected values for mesic (EWL ¼ 7:272m�0:532b )
and desert rodents (EWL ¼ 5:968m�0:416b ) of similar size
(Cort!es et al., 2000b).
2.4. Efficiency of water regulation
To evaluate the efficiency of water regulation we
utilized two indices: Ta@MWP ¼ EWL (MacMillen
and Hinds, 1983) and MR-WB (Cort!es, 2000b). MWP
was assessed from MR data vs. Ta; assuming that 1mlO2 consumed produce 0.62mg of water (Schmidt-
Nielsen, 1979) and 4.8 cal (Schmidt-Nielsen, 1990). The
magnitude of MR-WB was compared with the expected
values for mesic and xeric rodents, using the relations
MR-WB=34.627m�0:339b and MR-WB=68.132m�0:381
b ;respectively (Cort!es et al., 2000b).
2.5. Statistical analyses
Regression equations were calculated using least-
squares analyses (Steel and Torrie, 1985). Values are
given as means7SD.
3. Results
3.1. Energy metabolism
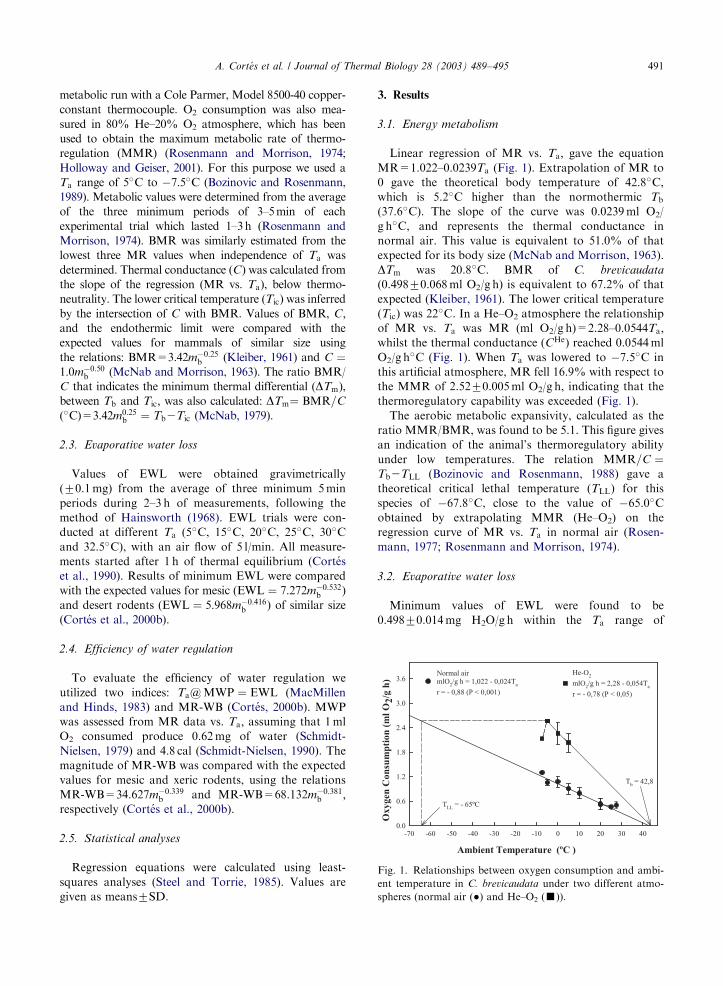
Linear regression of MR vs. Ta; gave the equationMR=1.022–0.0239Ta (Fig. 1). Extrapolation of MR to
0 gave the theoretical body temperature of 42.8�C,
which is 5.2�C higher than the normothermic Tb(37.6�C). The slope of the curve was 0.0239ml O2/
g h�C, and represents the thermal conductance in
normal air. This value is equivalent to 51.0% of that
expected for its body size (McNab and Morrison, 1963).
DTm was 20.8�C. BMR of C. brevicaudata
(0.49870.068ml O2/g h) is equivalent to 67.2% of that
expected (Kleiber, 1961). The lower critical temperature
(Tic) was 22�C. In a He–O2 atmosphere the relationship
of MR vs. Ta was MR (ml O2/g h)=2.28–0.0544Ta;whilst the thermal conductance (CHe) reached 0.0544ml
O2/g h�C (Fig. 1). When Ta was lowered to �7.5�C in
this artificial atmosphere, MR fell 16.9% with respect to
the MMR of 2.5270.005ml O2/g h, indicating that thethermoregulatory capability was exceeded (Fig. 1).
The aerobic metabolic expansivity, calculated as the
ratio MMR/BMR, was found to be 5.1. This figure gives
an indication of the animal’s thermoregulatory ability
under low temperatures. The relation MMR=C ¼Tb2TLL (Bozinovic and Rosenmann, 1988) gave a
theoretical critical lethal temperature (TLL) for this
species of �67.8�C, close to the value of �65.0�Cobtained by extrapolating MMR (He–O2) on the
regression curve of MR vs. Ta in normal air (Rosen-
mann, 1977; Rosenmann and Morrison, 1974).
3.2. Evaporative water loss
Minimum values of EWL were found to be
0.49870.014mg H2O/g h within the Ta range of
ARTICLE IN PRESS
Fig. 1. Relationships between oxygen consumption and ambi-
ent temperature in C. brevicaudata under two different atmo-
spheres (normal air (�) and He–O2 (’)).
A. Cort!es et al. / Journal of Thermal Biology 28 (2003) 489–495 491

p20�C (Fig. 2), while the averages of EWL at 25�C,
30�C and 32.5�C were 0.604, 0.820 and 1.105mg H2O/
g h, representing increases of 20%, 65% and 120%,
respectively, over the minimum. At the highest experi-
mental temperature EWL was equivalent to only 24.6%
of the basal rate of heat production (2.39 cal/g h). The
low cooling capability was reflected in a Tb increase of
1.1�C above the normothermic condition (38.7�C vs.
37.6�C) (Fig. 2).
3.3. Efficiency of water regulation
Fig. 3 shows the relation of LogMWP/EWL vs.
Ta; which gave the equation MWP=EWL ¼ 1:705�ð0:951ÞTa : Taking MWP/EWL=1, we obtained a valueof 10.6�C, that represents the efficiency index of
water regulation for C. brevicaudata (see Hinds and
MacMillen, 1985). Replacing Ta ¼ 10:6�C in the regres-sion equation (MR=1.022–0.0239Ta), we obtained a value
of 3.71 cal/gh, that is the energetic cost of maintaining the
water balance (MR-WB) (Cort!es et al., 2000b).
4. Discussion
4.1. Energy metabolism
C. brevicaudata showed an average BMR of
0.49870.068ml O2/g h, which is equivalent to 67.3%of the predicted value for a mammal of similar size
(Kleiber, 1961) and to 34% of the average given for
South American octodontid and murid rodents (Rosen-
mann, 1977; Bozinovic and Rosenmann, 1988; Bozino-
vic, 1992; Bozinovic et al., 1995); but in these
comparisons we should consider that our species is
twice the body size of the largest octodontid measured.
A comparison with a closer species indicated that BMR
in C. brevicaudata reached up to 75% of that reported
for Chinchilla lanigera (Cort!es et al., 2000a). Both
species are herbivorous (Cort!es et al., 2002), and
following the proposition of McNab (1986), their
BMR should be somewhat higher than that expected
for granivorous heteromyids and murids from North
American and Australian deserts (McNab, 1979; Daw-
son, 1955; Carpenter, 1966; MacMillen and Lee, 1970),
and also higher than some murids from Asian deserts
(Shkolink and Borut, 1969). Our data did not confirm
McNab’s proposition. Here BMR values were similar to
those reported for the other groups of desert rodents.
The fact that some species may possess a lower than
expected BMR is favorable for the maintenance of
energy and water balance, especially for those species
inhabiting arid environments with extreme climatic
conditions as found in the highlands of Northern Chile
ARTICLE IN PRESS
Ambient Temperature (ºC )
0 5 10 15 20 25 30 35
Eva
pora
tive
Wat
er L
oss
(mg/
g h )
0.0
0.5
1.0
1.5
2.0
Bod
y T
empe
ratu
re (
ºC)
37
38
39
40
Eva
pora
tive
Hea
t L
oss
(% o
f he
at p
rodu
ctio
n)
0
10
20
30
40
Normothermic37.6 + 0.28ºC.
Fig. 2. Relationships between evaporative water loss, evapora-
tive heat loss, body temperature and ambient temperature in
C. brevicaudata.
Ambient Temperature (ºC)
0 5 10 15 20 25
MW
P/E
WL
0.4
0.6
0.8
1
1.2
1.4
MWP/EWL = 1.705 (0.951)Ta
r = - 0.88 (P < 0.01)
Ta @ = 10.6ºC
Fig. 3. Relation between MWP/EWL and ambient temperature
in C. brevicaudata.
A. Cort!es et al. / Journal of Thermal Biology 28 (2003) 489–495492

where C. brevicaudata dwells. A low thermal conduc-
tance was also found in this species (C ¼ 0:0239ml O2/g h�C), equivalent to half of that predicted for body size
(McNab and Morrison, 1963). In fact, this is the lowest
conductance reported for South American rodents,
including murids, octodontids and chinchillids (Rosen-
mann, 1977; Bozinovic and Rosenmann, 1988; Bozino-
vic, 1992; Bozinovic et al., 1995; Cort!es et al., 2000a).
Both the high thermal insulation as well as the low Ticshould allow this nocturnal species to maintain a normal
Tb at low ambient temperatures. We should note that
the mean annual Ta at the capture site is about 2�C
(Luebert, 1998). The high thermal insulation is also
reflected by a DTm ¼ 22�C. This figure is 16�C and
6.4�C higher than the minimum and maximum DTmvalues reported for cricetids from South America
(Bozinovic and Rosenmann, 1988), and also higher than
the reported for some North American heteromyids
(Hinds and MacMillen, 1985). The aerobic metabolic
expansivity, (MMR/BMR) in C. breviaudata, was 5.1,
somewhat lower than the value of 5.8 reported for
C. lanigera (Cort!es et al., 2000a), and is clearly lower
than 8.2, found in the Andean murid Calomys ducilla
(Rosenmann and Morrison, 1974), but is of similar
magnitude to the values reported for Microtus oecono-
mus (5.1), Uromys caudimaculatus (4.7) (Hinds et al.,
1993) and the octodontid Octodon degus (4.9) (Rosen-
mann, 1977).
The critical lethal temperature that was calculated
for C. brevicaudata (TLL ¼ �67:8�C), was similar to theone for C. lanigera (�65.5�C) (Cort!es et al., 2000a), butwas 51.8�C lower than that (TLL¼ �16�C) given forC. ducilla (Rosenmann and Morrison, 1974).
4.2. Evaporative water loss
Minimum EWL in our chinchilla was 0.498mg H2O/
g h (Fig. 2). This value is within that expected for xeric
rodents (Cort!es et al., 2000b). It is probable that similar
morphological respiratory nasal adaptations occur as
reported for other xeric and desert rodents (Cort!es et al.,
1990). In any case, the low EWL may have unfavorable
consequences at high temperatures where evaporative
heat loss is important; for example, at Ta ¼ 32:5�C,C. brevicaudata was able to loose by EWL only 14 of the
metabolic heat production, which was reflected by a Tbincrease of 1.1�C. This moderate increase was seen to
affect its normal conditions. Similar responses to
moderate high temperatures have been reported in
O. degus (Rosenmann, 1977) and in C. lanigera
(Cort!es et al., 2000a). The low EWL of the chinchilla
may appear unfavorable at high temperatures, but this
physiological response is valuable for the maintenance
of body water, considering that this species inhabits
highly xeric environments where the only water source
may be found in the few plants that are normally
consumed (e.g., S. chrysophylla, A. eranicea, F. bryoides,
C. andicola and B. tola). In the same context, it is quite
likely that the high ambient temperatures occurring
around noon may not have significant consequences on
the animal’s thermoregulation, because of the nocturnal
habits described in this chinchilla (Munoz and Y!anez,
2000). Moreover, one of us (AC) found during several
summer days a relatively stable Ta range of 20–25�C
inside the dens or shelters, which are build under or
between large (2–3m) rocks.
4.3. Efficiency of water regulation
A Ta@ of 10.6�C was calculated for C. brevicaudata
(Fig. 3). This value may indicate a slightly lower
efficiency compared with other Chilean rodents from
mesic and xeric habits: Abrothrixs olivaceus (18.6�C), A.
andinus (12.5�C), Phyllotis darwini (14�C), P. magister
(10.5�C), P. rupestris (12.1�C), Oligoryzomys longicau-
datus (12.1�C), C. lanigera (12.7�C) and O. degus
(16.6�C) (Cort!es et al., 2000b). Nevertheless, the
energetic cost of maintaining the water balance, MR-
WB=3.71 cal/g h, was similar or lower than the values
reported for the other rodent species. In fact, the
energetic cost in our studied chinchilla is the lowest so
far described, being only 82% of the predicted value
(Cort!es et al., 2000b). Because of the extreme environ-
mental conditions endured by C. brevicaudata (low
availability of food and water), we found that this
physiological feature was not surprising.
Acknowledgements
We thank the Corporaci !on Nacional Forestal (CON-
AF, II Regi !on, Chile) for logistic assistance, mainly to
the wildlife keepers Rodrigo Araya and Alfonso Tapia
(Parque Nacional Llullaillaco). We also thank Dr. Jaime
Jim!enez (Universidad de Los Lagos) for his valuable
collaboration in the field. This work was financed by the
projects FONDECYT 5960017, Programa Sectorial
Biomas y Climas del Norte de Chile, Comisi !on Nacional
de Investigaci !on Cient!ıfica y Tecnol !ogica de Chile
(CONICYT), and Project FONDECYT 1981122.
References
Bozinovic, F., 1992. Rates of basal metabolism of grazing
rodents from different habitats. J. Mammal. 73, 379–384.
Bozinovic, F., Rosenmann, M., 1988. Comparative energetics
of South American cricetid rodents. Comp. Biochem.
Physiol. 91A, 195–202.
Bozinovic, F., Rosenmann, M., 1989. Maximum metabolic rate
of rodents: physiological and ecological consequences on
distributional limits. Funct. Ecol. 3, 173–181.
ARTICLE IN PRESSA. Cort!es et al. / Journal of Thermal Biology 28 (2003) 489–495 493

Bozinovic, F., Contreras, L.C., 1990. Basal rate of metabolism
and temperature regulation of two desert hervivorous
octodontid: Octomys mimax and Tympanoctomys barrerae.
Oecologia 84, 567–570.
Bozinovic, F., Rosenmann, M., Novoa, F.F., Medel, R.G.,
1995. Mediterranean type of climatic adaptation in physio-
logical ecology of rodent species. In: Arroyo, M.T.K.,
Zedler, H., Fox, M.D. (Eds.), Ecology and Biogeography of
Mediterranean Ecosystems in Chile. California and Aus-
tralia. Springer, New York, pp. 347–361.
Carpenter, R.E., 1966. A comparison of thermoregulation and
water metabolism in the Kangoroo rats Dipodomys agilis
and Dipodomys merriami. Univ. Calif. Publ. Zool. 78, 1–36.
Cort!es, A., 1985. Adaptaciones fisiol !ogicas y morfol !ogicas de
pequenos mam!ıferos de ambientes semi!aridos. Tesis de
Mag!ıster, Universidad de Chile, 121pp.
Cort!es, A., Zuleta, C., Rosenmann, M., 1988. Comparative
water economy of sympatric rodents in a Chilean semi-arid
habitat. Comp. Biochem. Physiol. 91A, 711–714.
Cort!es, A., Rosenmann, M., B!aez, C., 1990. Funci!on del rin !on
y del pasaje nasal en la conservaci !on del agua corporal en
roedores simp!atridos de Chile central. Rev. Chil. Hist. Nat.
63, 279–291.
Cort!es, A., Rosenmann, M., Bozinovic, F., 2000a. Relaci !on
costo-beneficio en la termorregulaci !on de Chinchilla lani-
gera. Rev. Chil. Hist. Nat. 73, 351–357.
Cort!es, A., Rosenman, M., Bozinovic, F., 2000b. Water
economy in rodent: evaporative water loss and metabolic
water production. Rev. Chil. Hist. Nat. 73, 311–321.
Cort!es, A., Miranda, E., Jim!enez, J.E., 2002. Seasonal food
habits of the endangered long-tailed chinchilla (Chinchilla
lanigera): the effect of precipitation. Mamm. Biol. 67,
1–9.
Cruz-Neto, A., Garland, T., Shynya, A., 2001. Diet, phylogeny,
and basal metabolic rate in phyllostomid bats. Zoology 104,
49–58.
Dawson, W.R., 1955. The relation of oxygen consumption to
temperature in desert rodents. J. Mammal. 36, 543–553.
Degen, A., 1997. Ecophysiology of small desert mammals, 1st
Edition. Springer, Berlin, 296pp.
Di Castri, F., Hajek, E.R., 1976. Bioclimatolog!ıa de Chile.
Editorial de la Universidad Cat !olica de Chile, Santiago,
Chile, 128pp.
Garc!ıa, J., Ojeda, R., Fraga, R., D!ıaz, G., Baig !un, R., 1997.
Libro rojo de mam!ıferos y aves amenazados de la
Argentina. Fundaci!on para la Conservaci !on de las Especies
y el Medio Ambiente. Buenos Aires, Argentina, 221pp.
Glade, A., 1993 Libro rojo de los vertebrados terrestres de
Chile. Corporaci !on Nacinal Forestal, Ministerio de Agri-
cultura, Santiago, 65pp.
Hainsworth, F.R., 1968. Evaporative water loss from rats in the
heat. Am. J. Physiol. 214, 979–982.
Hayssen, V., Lacy, R.C., 1985. Basal metabolic rates in
mammals: taxonomic differences in the allometry of BMR
and body mass. Comp. Biochem. Physiol. 81A, 741–754.
Hinds, D.S., MacMillen, R.E., 1985. Scaling of energy
metabolism and evaporative water loss in heteromyid
rodents. Physiol. Zool. 58, 282–298.
Hinds, D.S., Baudinette, R.V., MacMillen, R.E., Halpern,
E.A., 1993. Maximum metabolism and the aerobical
factorial scope of endotherms. J. Exp. Biol. 182, 41–56.
Holloway, J.C., Geiser, F., 2001. Effects of helium/oxygen and
temperature on aerobic metabolism in the marsupial sugar
glider, Petaurus breviceps. Physiol. Biochem. Zool. 74 (2),
219–225.
Hulbert, A.J., Dawson, T.J., 1974. Standard metabolism and
body temperature of perameloid marsupials from different
environments. Comp. Biochem. Physiol. 47A, 583–590.
Jim!enez, J., 1996. The extirpation and current status of wild
chinchillas Chinchilla lanigera and C. brevicaudata. Biol.
Conserv. 77, 1–6.
Kleiber, M., 1961. The Fire of Life, an Introduction to Animal
Energetics. Wiley, New York, USA, 454pp.
Kronfeld-Schor, N., Haim, A., Dayan, T., Zisapel, N.,
Klingespor, M., Heldmaier, G., 2000. Seasonal thermogenic
acclimation of diurnally and nocturnally active desert spiny
mice. Physiol. Biochem. Zool. 73, 37–44.
Lovegrove, B.G., 1986. The metabolism of social subterranean
rodent: adaptation to aridity. Oecologia (Berlin) 69,
551–555.
Lovegrove, B.G., Heldmaier, G., Knigth, M., 1991. Seasonal
and circadian energetic patterns in an arboreal rodent,
Thallomys paedulcus, and a burrow-dwelling rodent, Aeth-
omys namaquensis, from the Kalahari desert. J. Therm. Biol.
16, 199–209.
Luebert, F., 1998. Relaciones biogeogr!aficas de la flora del
Parque Nacional Llullaillaco. XI. Reuni !on Anual de la
Sociedad de Bot!anica de Chile. Abstr. Noticiero Biol.
6 (5), 197.
MacMillen, R.E., Lee, A.K., 1970. Energy metabolism and
pulmocutaneus water loss of Australian hopping mice.
Comp. Biochem. Physiol. 35, 355–369.
MacMillen, R.E., Hinds, D.S., 1983. Water regulatory effi-
ciency in heteromyid rodents: a model and its application.
Ecology 64, 152–164.
MacMillen, R.E., Garland, T., 1989. Adaptative physiology. In:
Lane, J.N., Kirkland, G.L. (Eds.), Advances in the Study of
Peromyscus (Rodentia). Texas Technical University Press,
Lubbock, pp. 143–168.
McNab, B.K., 1979. Climatic adaptation in the energetics of
heteromyid rodents. Comp. Biochem. Physiol. 62, 813–820.
McNab, B.K., 1986. The influence of food habits on the
energetics of eutherian mammals. Ecol. Monogr. 56, 1–19.
McNab, B.K., Morrison, P.R., 1963. Body temperature and
metabolism in subspecies of Peromyscus from arid and
mesic environment. Ecol. Monogr. 33, 63–82.
Messerli, B.M., Grosjean, G., Bonani, A., Burgi, M., Geyh, K.,
Graf, K., Rameyer, K., Romero, H., Schotterer, U.,
Schreier, H., Vuille, M., 1993. Climate change and natural
recourse dynamics of the Atacama Altiplano during the
18,000 last years: a preliminary synthesis. Mt. Res. Dev. 13,
117–127.
Morrison, P.R., 1951. An automatic manometric respirometer.
Rev. Sci. Instrum. 2, 264–267.
Munoz, A., Y!anez, J., 2000. Mam!ıferos de Chile. Ediciones
CEA, Valdivia, Chile, 455pp.
Nagy, K.A., Girard, I.A., Brown, T.K., 1999. Energetics of
free-ranging mammals, reptiles, and birds. Annu. Rev.
Nutr. 19, 247–277.
Prakash, I., 2001. Survival strategies of desert vertebrates. In:
Prakash, I. (Ed.), Ecology of Desert Environments.
Scientific Publishers, India, pp. 459–471.
ARTICLE IN PRESSA. Cort!es et al. / Journal of Thermal Biology 28 (2003) 489–495494

Raab, J., Schmidt-Nielsen, K., 1972. Effect of running on water
balance of kangaroo rat. Am. J. Physiol. 222, 1230–1235.
Redford, K.H., Einsenberg, J.F., 1989. Mammals of the
Neotropics: The Southern Cone, Vol. 2: Chile, Argentina,
Uruguay and Paraguay. University of Chicago Press,
Chicago and London. 430pp.
Rosenmann, M., 1977. Regulaci!on t!ermica en Octodon degus.
Medio Ambiente. (Chile) 3, 127–131.
Rosenmann, M., Morrison, P.R., 1974. Maximum oxygen
consumption and heat loss facilitation in small home-
otherms by He–O2. Am. J. Physiol. 226, 490–495.
SAG, 2000. Cartilla de Caza. Ministerio de Agricultura,
Servicio Agr!ıcola Ganadero, Departamento de Protecci !on
de los Recursos Naturales Renovables, Chile, 84pp.
Shkolink, A., Borut, A., 1969. Temperature and water relations in
two species of spiny mice (Acomys). J. Mammal. 50, 245–255.
Shkolnik, A., Schmidt-Nielsen, K., 1976. Temperature regula-
tion in hedgehogs from temperate and desert environment.
Physiol. Zool. 49, 54–56.
Schmidt-Nielsen, K., 1979. Desert Animals: Physiological
Problems of Heat and Water. Dover Publications, Inc.,
New York, 277pp.
Schmidt-Nielsen, K., 1990. Animal Physiology: Adaptation and
Environment. Cambridge University Press, Cambridge, 583pp.
Speakman, J.R., 2000. The cost of living: field metabolic rates
of small mammals. Adv. Ecol. Res. 30, 178–297.
Steel, R.G.D., Torrie, J.H., 1985. Bioestad!ıstica: Principios y
Aplicaciones. McGraw-Hill, Bogot!a, Colombia, 340pp.
ARTICLE IN PRESSA. Cort!es et al. / Journal of Thermal Biology 28 (2003) 489–495 495


















