
End-Guadalupian extinction of the Permian gigantic bivalve ...
11
End-Guadalupian extinction of the Permian gigantic bivalve Alatoconchidae: End of gigantism in tropical seas by cooling Yukio Isozaki a, ⁎, Dunja Aljinović b a Department of Earth Science and Astronomy, The University of Tokyo, Meguro, Tokyo 153-8902, Japan b Faculty of Mining Geology and Petroleum Engineering, The University of Zagreb, Zagreb 10000, Croatia abstract article info Article history: Received 14 May 2007 Received in revised form 4 November 2008 Accepted 15 August 2009 Available online 9 September 2009 Keywords: Gigantism Capitanian Permian Mass extinction photosymbiosis Eutrophication The unique Permian bivalve family Alatoconchidae has aberrant shell forms and extraordinary size up to 1 m, representing the largest bivalve group in the Paleozoic. Their occurrence is reported sporadically from Lower–Middle Permian shallow-marine carbonates in 9 areas in the world (Tunisia, Croatia, Oman, Iran, Afghanistan, Thailand, Malaysia, the Philippines, and Japan) that cover low-latitudes of both the Tethyan and Panthalassan domains. Alatoconchids almost always occurred in close association with large-tested fusulines (Verbeekinidae) and/or rugose corals (Waagenophyllidae) of the typical Tethyan assemblage, suggesting their preferential adaptation to shallow warm-water (tropical) environments. This “tropical trio” (Alatoconchidae, Verbeekinidae, and Waagenophyllidae) became extinct either during the Late Guadalupian or around the Guadalupian–Lopingian boundary (G–LB). Their intimate association and occurrence range suggest that these 3 taxonomically distinct clades may have shared not only a common habit but also a common cause of extinction. The shell structure of alatoconchids suggests their symbiosis with photosynthetic organisms (algae + cyanobacteria) in order to maintain their large body size that required high energy-consuming metabolism in contrast to smaller forms. The Alatoconchidae attained their largest size in the Wordian (Middle Guadalupian), probably maximizing the benefits of photosymbiosis. The subsequent extinction of the warm-water-adapted “tropical trio” both in Tethys and Panthalassa positively supports the explanation that a critical cooling took place on a global scale, including low-latitude oceans. The end of the gigantism in fusulines and bivalves in the Capitanian (Late Guadalupian) was likely caused by the collapse of photosymbiotic systems during a temporary temperature drop of seawater (Kamura cooling event) coupled with eutrophication that was detrimental to the tropical fauna adapted particularly to oligotrophic conditions. Gigantism of bivalves occurred several times in the Phanerozoic; e.g., Siluro- Devonian, Permian, Triassic–Early Jurassic, and Jurassic–Creataceous, mostly in warm periods. The sea-level change in the Phanerozoic apparently synchronized with the intermittent rise and decline of bivalve gigantism, suggesting that the photosymbiosis-related gigantism in low-latitudes may serve as a potential monitor of global warming/cooling in the past. © 2009 Elsevier B.V. All rights reserved. 1. Introduction Size variations of fossil animals and their changes over geological time have long attracted intense curiosity of paleontologists ever since the 19th century. For example, an apparent trend recognized in fossil records led to the empirical concept known as Cope's rule, i.e., orthogenesis towards large size. Stanley (1973) re-interpreted this classic perspective in a modern way by emphasizing the common pattern recognized in many higher taxa (order and class) with small- size ancestors and large-size descendants, such as bivalves, ammo- noids, and fusulines. Later, Jablonski (1996) challenged this view by pointing out sampling bias with respect to the skewed population structure. Although the actual mechanisms of driving body size and their meanings in evolution have not been sufficiently explained, biotic interactions among various organisms and nutrient availability likely contributed to the determination of the size of fossil animals (e.g., Vermeij, 1977; Brasier, 1995). As to the balance between body size and population size, MacArthur and Wilson (1967) tried to formulate the ecological structure of modern animal communities in terms of the dichotomy of K-selection and r-selection. This classic approach was further emphasized by Pianka (1978), which, however, was not favored by contemporary ecologists because the suggested trade-off relations between the two selections cannot be recognized in many modern cases. Nonetheless, fossil records suggest that some ancient taxa did deploy the r-strategy under extremely stressful conditions in the geologic past, particularly in the aftermaths of major mass extinction events (e.g., Urbaneck, 1993; Schubert and Bottjer, 1995; Twitchett, Palaeogeography, Palaeoclimatology, Palaeoecology 284 (2009) 11–21 ⁎ Corresponding author. Dept. of Earth Science and Astronomy, Univ. Tokyo, Japan. Tel.: +81 3 5454 6608; fax: +81 3 5465 8244. E-mail address: [email protected] (Y. Isozaki). 0031-0182/$ – see front matter © 2009 Elsevier B.V. All rights reserved. doi:10.1016/j.palaeo.2009.08.022 Contents lists available at ScienceDirect Palaeogeography, Palaeoclimatology, Palaeoecology journal homepage: www.elsevier.com/locate/palaeo
Transcript of End-Guadalupian extinction of the Permian gigantic bivalve ...

Palaeogeography, Palaeoclimatology, Palaeoecology 284 (2009) 11–21
Contents lists available at ScienceDirect
Palaeogeography, Palaeoclimatology, Palaeoecology
j ourna l homepage: www.e lsev ie r.com/ locate /pa laeo
End-Guadalupian extinction of the Permian gigantic bivalve Alatoconchidae:End of gigantism in tropical seas by cooling
Yukio Isozaki a,⁎, Dunja Aljinović b
a Department of Earth Science and Astronomy, The University of Tokyo, Meguro, Tokyo 153-8902, Japanb Faculty of Mining Geology and Petroleum Engineering, The University of Zagreb, Zagreb 10000, Croatia
⁎ Corresponding author. Dept. of Earth Science and ATel.: +81 3 5454 6608; fax: +81 3 5465 8244.
E-mail address: [email protected] (Y. Isozak
0031-0182/$ – see front matter © 2009 Elsevier B.V. Aldoi:10.1016/j.palaeo.2009.08.022
a b s t r a c t
a r t i c l e i n f oArticle history:Received 14 May 2007Received in revised form 4 November 2008Accepted 15 August 2009Available online 9 September 2009
Keywords:GigantismCapitanianPermianMass extinctionphotosymbiosisEutrophication
The unique Permian bivalve family Alatoconchidae has aberrant shell forms and extraordinary size up to 1 m,representing the largest bivalve group in the Paleozoic. Their occurrence is reported sporadically fromLower–Middle Permian shallow-marine carbonates in 9 areas in the world (Tunisia, Croatia, Oman, Iran,Afghanistan, Thailand, Malaysia, the Philippines, and Japan) that cover low-latitudes of both the Tethyan andPanthalassan domains. Alatoconchids almost always occurred in close association with large-tested fusulines(Verbeekinidae) and/or rugose corals (Waagenophyllidae) of the typical Tethyan assemblage, suggestingtheir preferential adaptation to shallow warm-water (tropical) environments. This “tropical trio”(Alatoconchidae, Verbeekinidae, and Waagenophyllidae) became extinct either during the Late Guadalupianor around the Guadalupian–Lopingian boundary (G–LB). Their intimate association and occurrence rangesuggest that these 3 taxonomically distinct clades may have shared not only a common habit but also acommon cause of extinction. The shell structure of alatoconchids suggests their symbiosis withphotosynthetic organisms (algae+cyanobacteria) in order to maintain their large body size that requiredhigh energy-consuming metabolism in contrast to smaller forms. The Alatoconchidae attained their largestsize in the Wordian (Middle Guadalupian), probably maximizing the benefits of photosymbiosis. Thesubsequent extinction of the warm-water-adapted “tropical trio” both in Tethys and Panthalassa positivelysupports the explanation that a critical cooling took place on a global scale, including low-latitude oceans.The end of the gigantism in fusulines and bivalves in the Capitanian (Late Guadalupian) was likely caused bythe collapse of photosymbiotic systems during a temporary temperature drop of seawater (Kamura coolingevent) coupled with eutrophication that was detrimental to the tropical fauna adapted particularly tooligotrophic conditions. Gigantism of bivalves occurred several times in the Phanerozoic; e.g., Siluro-Devonian, Permian, Triassic–Early Jurassic, and Jurassic–Creataceous, mostly in warm periods. The sea-levelchange in the Phanerozoic apparently synchronized with the intermittent rise and decline of bivalvegigantism, suggesting that the photosymbiosis-related gigantism in low-latitudes may serve as a potentialmonitor of global warming/cooling in the past.
stronomy, Univ. Tokyo, Japan.
i).
l rights reserved.
© 2009 Elsevier B.V. All rights reserved.
1. Introduction
Size variations of fossil animals and their changes over geologicaltime have long attracted intense curiosity of paleontologists eversince the 19th century. For example, an apparent trend recognized infossil records led to the empirical concept known as Cope's rule, i.e.,orthogenesis towards large size. Stanley (1973) re-interpreted thisclassic perspective in a modern way by emphasizing the commonpattern recognized in many higher taxa (order and class) with small-size ancestors and large-size descendants, such as bivalves, ammo-noids, and fusulines. Later, Jablonski (1996) challenged this view bypointing out sampling bias with respect to the skewed population
structure. Although the actual mechanisms of driving body size andtheir meanings in evolution have not been sufficiently explained,biotic interactions among various organisms and nutrient availabilitylikely contributed to the determination of the size of fossil animals(e.g., Vermeij, 1977; Brasier, 1995).
As to the balance between body size and population size,MacArthur and Wilson (1967) tried to formulate the ecologicalstructure of modern animal communities in terms of the dichotomy ofK-selection and r-selection. This classic approach was furtheremphasized by Pianka (1978), which, however, was not favored bycontemporary ecologists because the suggested trade-off relationsbetween the two selections cannot be recognized in many moderncases. Nonetheless, fossil records suggest that some ancient taxa diddeploy the r-strategy under extremely stressful conditions in thegeologic past, particularly in the aftermaths of major mass extinctionevents (e.g., Urbaneck, 1993; Schubert and Bottjer, 1995; Twitchett,

12 Y. Isozaki, D. Aljinović / Palaeogeography, Palaeoclimatology, Palaeoecology 284 (2009) 11–21
1999; Fraiser and Bottjer, 2004). The term “Lilliput effect”was coinedby Urbaneck (1993) for describing such remarkable minimalism inthe fossil record.
On the other hand, as large fossils are eye-catchers not only foramateur fossil collectors but also for professional paleontologists,gigantism in fossil records has been recognized and discussed withrespect to modern examples. Among modern bivalves, Tridacna gigasin the Indo-Pacific region is known as the biggest (a single valve mayreach weights of more than 110 kg; Yonge, 1936) that is clearly out ofthe 2-sigma range in the normal distribution of modern and ancientbivalve sizes (e.g., Nicol, 1964). In fossil records, gigantism in bivalvesappeared repeatedly at least 4 times; i.e. Siluro-Devonian mega-lodonts, Permian alatoconchids, Late Triassic–Early Jurassic mega-lodonts/lithiotids, and Late Jurassic–Cretaceous rudists/inoceramids,although most of themwere not necessarily related in direct lineages.
In general, the advantage of attaining a large body size includesefficiency/advantage in 1) collecting food of various sizes, 2)predation, 3) protection from other animal's predation, 4) reproduc-tion, and 5) maintaining internal-body homeostasis (e.g., Pianka,1978). However, these advantages detract from small populationsizes, long maturation time, specialized habit, and/or large amount ofrequired nutrients/energy, which may act as disadvantages whenenvironmental conditions change into different modes. Thus, in orderto achieve unusual gigantism out of the normal size range of aclade, animals need to overcome these disadvantages by trading offsome benefits in being small and by deploying unusual strategies.Yonge (1936) first pointed out the occurrence of symbiotic algaezooxanthellae (= dinoflagellate genus Symbiodium) in Tridacna, andother modern examples (e.g., Corculum, Fragum, and Hippopus) wereadded later in the list (e.g., Kawaguti, 1983). As for ancient bivalves,the gigantism in rudists and other aberrant gigantic fossil clams was
Fig. 1. Morphology of Alatoconchidae. A–C: Reconstructed general shell morphology of Alatocostored in the Croatian Natural History Museum in Zagreb.
likewise explained in terms of photosymbiosis (e.g., Vogel, 1975;Yancey and Boyd, 1983; Kauffman and Johnson, 1988; Seilacher,1990; de Freitas et al., 1993) on the basis of indirect lines of evidence,i.e., analogy with modern examples in large size, aberrant form, andunique shell structures.
Among the above-listed fossil giant clams, the Permian bivalvefamily Alatoconchidae (Fig. 1) appears the most noteworthy notmerely because it represents the largest fossil bivalve ever since theCambrian radiation of metazoans but also because it became extinctabruptly near the end of the Guadalupian (Middle Permian) thatmarks the timing of one of the major mass extinction events in thePhanerozoic (Stanley and Yang, 1994; Jin et al., 1994; Isozaki and Ota,2001; Isozaki, 2007a,b, 2009).
This article reviews the occurrences of the Permian bivalve familyAlatoconchidae to date, and discusses the geological implications oftheir gigantism and extinction pattern. By emphasizing the intimateassociation of tropical-adapted Alatoconchidae, Verbeekinidae (large-tested fusulines) and Waagenophyllidae (rugose corals), possibleclimate-tuning of the intermittent rise/decline of bivalve gigantism inthe Phanerozoic is also discussed in the light of photosymbiosis intropical shallow-marine environments.
2. Permian giant clam Alatoconchidae
Alatoconchidae is a unique Permian bivalve family characterizedby an aberrant shell form (Fig. 1) and an extraordinarily large size upto 1 m in length. The occurrence of this bivalve family was firstdescribed from a Middle Permian limestone in Japan with a newgenus/species name Shikamaia akasakaensis Ozaki; however, theoriginal specimen was treated as a paleontological problematicasimply because its bizarre shape appeared too distinct from those of
nchidae, D–E: the holotype of Shikamaia (=Tanchintongia) ogulineci (Kochansky-Devidé)

13Y. Isozaki, D. Aljinović / Palaeogeography, Palaeoclimatology, Palaeoecology 284 (2009) 11–21
ordinary bivalves (Ozaki, 1968). Despite such taxonomic confusionin the beginning, additional occurrences of Alatoconchidae weresporadically reported later from Lower and Middle Permian rocks inMalaysia, Afghanistan, Iran, Croatia, and Tunisia (Termier et al., 1973;Runnegar and Gobbett, 1975; Kochansky-Devidé, 1978; Boyd andNewell, 1979; Thiele and Ticky, 1980). The family name Alatoconchidaewas given by Termier et al. (1973) who first identified them as bivalvesand demonstrated their overall shell morphology with ligamentstructure, although their reconstruction was oriented up-side-down.Runnegar and Gobbett (1975) correctly illustrated a general morphol-ogy for alatoconchids, and Kochansky-Devidé (1978) discussed theirunique morphological adaptation with respect to ecological habitat. Onthe basis of all these previous data and interpretations plus newmaterial added later, Yancey and Boyd (1983) and Yancey and Ozaki(1986) summarized the paleobiology and taxonomy of the entirefamily, as briefly described below.
The family Alatoconchidae comprises two subfamilies; i.e.,Alatoconchinae and Saikraconchinae. The former includes the genusShikamaia Ozaki and the subgenus Alatoconcha (Yancey & Boyd),whereas the latter contains the genus Saicraconcha Yancey & Boydand the subgenus Dereconcha Yancey & Boyd. Genus TanchintongiaRunnegar & Gobbett was regarded as a synonym of Shikamaia Ozaki.
Due to their three-dimensionally twisted and fragile shell form(Fig. 1), most of the described specimens occur as broken fragments(Fig. 2). In addition, as shells are often embedded tightly withinlimestone matrices, it is generally difficult to isolate free individualvalves. Nonetheless, by virtue of some exceptionally well-preservedspecimens (Fig. 1C and D) and intensive studies by the above-mentioned workers, their general three-dimensional shell form wasreasonably reconstructed (Fig. 1A–C). The length of individualalatoconchid shell mostly ranges from 5 to 60 cm with an averagearound 30 cm (Fig. 1A). Yancey and Ozaki (1986) mentioned that themaximum size of adults likely had reached nearly 1 m in length. Oneof the present authors (Y.I.) observed a nearly 0.8 m-long specimenwith a shell thickness over 5 cm in Japan. There is no known bivalvefamily in the Permian or in the Paleozoic that reached this size except
Fig. 2. Field occurrence of Alatoconchidae. Left: Alatoconchidae gen. et sp. indet. from the Neothe Pseudofusulina ambigua Zone in Neo (Japan), lower middle: juvenile form of Alatoconchidbed of the Yabeina Zone at Akasaka (Japan). Scale for the left photo is 20 cm long. Note thatagitated shallow-marine environment for their habitat. The example in Akasaka (right) occ
Alatoconchidae, the size of which can be compared with that of thelargest modern bivalve Tridacna gigas.
Alatoconchidae is equivalved; two valves are arranged symmet-rically with respect to the orthothetic plane of commissure (Fig. 1).The bivalve is elongated in the anterior–posterior direction, butcompressed in the dorsal–ventral direction. The most striking featureis the lateral flange formed by the horizontal extension of umbonalcarinae. The ventral surface forms a large planer surface, whereas thedorsal side forms a crest extending perpendicular to the flat ventralsurface. There is a remarkable contrast in shape between the anteriorand posterior halves. The height of dorsal crest reaches the apex in theanterior half and decreases sharply in the mid-shell, whereas thewidth of shell decreases gradually towards the posterior. Thus theposterior half of the bivalve has pronounced flat lateral flanges. Anoverall profile of the bivalve across the main axis forms a triangle inthe anterior and an extremely flat oval in the posterior half.
Such a large size and aberrant form of shell suggest thatalatoconchids were epifaunal suspension-feeders, and that theywere not very capable either of moving around or of frequentopening/closing of their shells (Termier et al., 1973; Runnegar andGobbett, 1975; Kochansky-Devide, 1978). It is noteworthy thatalatoconchids preferentially occur in a bituminous black limemudstone facies (Yancey and Boyd, 1983; Sano, 1988; Kiessling andFlügel, 2000; Isozaki, 2006; Aljinović et al., 2008; Fig. 2 left andmiddle). Alatoconchids may have adapted to the soft fine-grainedsediment surface of wackestone- to lime mudstone-dominant faciesby developing a flat-bottom in order to maintain their position on softsubstrates against their heavy shell weight, i.e., like snowshoe oroutrigger (e.g., Seilacher, 1990). In-situ shell concentrations indicate agregarious habitat in low to middle-energy environments of a shallowsea (Yancey and Boyd, 1983; Fig. 2 left and middle). Some cases withrandom alignment of shell fragments suggest a reworked origin instorm beds (Fig. 2 right).
As to the shell structure, Alatoconchidae is characterized by thedevelopment of a unique external layer in a double-layered shell(Yancey and Boyd, 1983; Isozaki, 2006; Fig. 3). The external layer is
schwagerina Zone in Kamura (Japan), upper middle: Shikamaia akasakaensis Ozaki fromae from the Yabeina Zone in Brušane, the Velebit Mountains (Croatia), right: Shikamaiaalatoconchids occur preferentially in wackestone and lime mudstone, suggesting a lessurs in a debris flow bed in which alatoconchid shells are scattered in randomly.

Fig. 3. Shell structure of Alatoconchidae. Left: large specimen of alatoconchid from the Yabeina Zone in Brusane, Croatia (upper) and prismatic calcite layer of the external shell on abroken surface (lower). Right: large specimen of alatoconchid from the Neoschwagerina Zone in Kamura, Japan (upper) and prismatic calcite layer of the external shell in thin section(lower).
14 Y. Isozaki, D. Aljinović / Palaeogeography, Palaeoclimatology, Palaeoecology 284 (2009) 11–21
composed of prismatic calcite aligned perpendicularly to the shellsurface, whereas the internal one consists of mosaic granular calcite.The development of long prismatic calcite, sometimes reaching over1 cm in length (fig. 2e in Isozaki, 2006), is significant with respect tothe bivalve's ecological habit of photosymbiosis, as these crystals canserve as optic fibers that let sunlight penetrate into the interior ofbivalves even when closed. A similar structure is known in modernphotosymbiotic bivalve groups, such as Corculum (e.g., Kawaguti,1983; Seilacher, 1990). The extraordinarily large size, generally flatshell form, and the transparent shell structure suggest that thePermian aberrant alatoconchids performed symbiosis with photosyn-thetic microorganisms (algae or cyanobacteria; just like modernzooxanthellae) in the photic zone in shallow-marine environmentsless than 50 m deep (Yancey and Boyd, 1983; Seilacher, 1990).
3. Paleogeographic distribution
The occurrence of the Permian alatoconchids has been reported todate from 9 areas in the world as shown in Table 1 and Fig. 4; i.e., fromwest to east, Tunisia (Boyd and Newell, 1979), Croatia (Kochansky-Devidé, 1978; Aljinović et al., 2008), Iran (Thiele and Ticky, 1980),
Table 1List of the occurrences of the Permian bivalve family Alatoconchidae. Pa: Pseudofusulinacraticulifera Zone, Nm: N. margaritae Zone), Zone, Y: Yabeina Zone, L: Lepidolina Zone, Art:
Locality Unit(Fm)
Biozone
List of occurrences of Alatoconchidae1. Japan
a. Akasaka Upper Pk–Yb. Neo Funabuseyama Pac. Kamura Iwato Nc–Ld. Shiroiwayama – Ne. Tomochi Hashirimizu N
2. Philippines (Palawan) Minilog Nc3. Malaysia (Kinta Valley) – Pa4. Thailand (Khao Khwang) Saraburi N–L5. Afghanistan (Al-e Say Pass) – Nm6. Iran
a. Kuh e Ahmad – ?b. Mahallt – ?c. Shahpoor – ?
7. Oman (Saih Harat) Saiq N8. Croatia (Velebit Mtn.) Velebit Nc–Y9. Tunisia (Saikra) Saikra N
Afghanistan (Termier et al., 1973), Oman (Pillevat, 1993; Wiedlichand Bernecker, 2007), Thailand (Udchachon et al., 2007), Malaysia(Runnegar and Gobbett, 1975), the Philippines (Kiessling and Flügel,2000), and Japan (Ozaki, 1968; Yancey and Ozaki, 1986; Saito et al.,2005; Isozaki, 2006; Matsuda et al., 2006). The first 7 cases werereported all from shallow-marine carbonates deposited aroundTethys along the eastern coast of Pangea, and they are characterizedby their co-occurrence with typical Tethyan faunal assemblages thatinclude large-tested fusulines of Verbeekinidae and rugose corals ofWaagenophyllidae (Yancey and Boyd, 1983). Most of the bivalvelocalities were apparently in low-latitude domains near the paleo-equator. Oman and Malaysia localities were rather southerlypositioned around 30° S with respect to the rest, nonetheless, stillwithin a narrow Neo-Tethys probably washed by warm waters.
In contrast, the last two cases in the Philippines and Japan werereported from Permian shallow-marine limestones contained inJurassic accretionary complexes (Isozaki et al., 1987; Isozaki, 1997b;Zamoras and Matsuoka, 2004). In Japan, all alatoconchid-bearinglimestones represent Permian mid-oceanic, atoll-type carbonatebuildups developed on seamounts that were primarily located inthe middle of the superocean Panthalassa. Their tectono-sedimentary
ambigua Zone, Pk: Parafusulina kaerimizuensis Zone, N: Neoschwagerina Zone (Nc: N.Artinskian, Word: Wordian, Cap: Capitanian.
Age References
Road–Cap Ozaki (1968), Ota and Isozaki (2006)Art Yancey and Ozaki (1986); Sano (1988)Word–Cap Isozaki (2006)Word Matsuda et al. (2006)Word–Cap? Saito et al. (2005)Word Kiessling and Flügel (2000)Art Runnegar and Gobbett (1975)Word–Cap Udchachon et al. (2007)Word Termier et al. (1973)
? Thiele and Ticky (1980)? Thiele and Ticky (1980)? Thiele and Ticky (1980)Word Pillevat (1993), Wiedlich and Bernecker (2007)Word–Cap Kochansky-Devidé (1978), Aljinović et al. (2008)Word Boyd and Newell (1979)

Fig. 4. Paleogeographic distribution of Alatoconchidae (compiled from the references listed in the text and Table 1; map based on Scotese and Langford (1995) and modified fromIsozaki (2006)). Note that the occurrences are concentrated in the low-latitude domains of both Tethys and Panthalassa.
15Y. Isozaki, D. Aljinović / Palaeogeography, Palaeoclimatology, Palaeoecology 284 (2009) 11–21
history, from the mid-Panthalassa deposition in the Permian to thefinal accretion to the Asian continental margins in the Jurassic, wasclearly documented by detailed field mapping and radiolarian dating(e.g., Wakita, 1988; Sano, 1988). In addition to Akasakawhere the firstalatoconchid specimen was described (Ozaki, 1968), their occurrencehas been reported from 4 different areas in SW Japan; i.e. Neo (Yanceyand Ozaki, 1986; Sano, 1988), Kamura (Isozaki, 2006), Shiroiwayama(Matsuda et al., 2006), and Tomochi (Saito et al., 2005). These mid-Panthalassan carbonate buildups are characterized by the dominantoccurrence of Tethyan fauna (e.g., Ozawa and Nishiwaki, 1992; ZawWin, 1999) as well as the above-mentioned 7 examples of bona fideTethyan origin. This suggests that at least the western domain of low-latitude Panthalassa had a strong link with Paleo-Tethys, and thatalatoconchids were no doubt adapted to such a tropical shallow-marine environment, as well as Verbeekinidae and Waagenophylli-dae. The paleo-latitude of the Permian atoll limestone at Kamura inJapanwas recently revealed to be 12° S (Kirschvink and Isozaki, 2007).
In North Palawan, west Philippines, Kiessling and Flügel (2000)reported the occurrence of alatoconchids from the Guadalupian part oftheMinilog Formation composed of shallow-marine carbonates. Judgingfrom their geological description plus geotectonic setting and continuityto the neighboring Calamian islands to the north (Isozaki et al., 1987;Zamoras and Matsuoka, 2004), the alatoconchid-bearing Permianlimestone of the Minilog Formation probably occurs as exotic blockswithin a Mesozoic (probably Jurassic) accretionary complex. If thisexample is reasonable, the Permian Minilog limestone may have beendepositednot along the easternmarginof SouthChina (Yangtzeblock) asoriginally reported but somewhere in the midst of Panthalassa, as inJapan. As to the Minilog Formation, the complete absence of corallinesponges, a major dissimilarity to coeval Tethyan reef fauna pointed outby Kiessling and Flügel (2000), can be better explained in the above-mentioned paleogeographic framework and context. Although theprecise paleo-latitude of that in the Philippines is unknown, theassociated Tethyan fusulines and corals likewise suggest a warm-waterdomain, as in the same low-latitude tropical sea in western Panthalassa.
Thus the distribution of Alatoconchidae was restricted largely tothe shallow-marine environments in warm, low-latitude domainsboth of Tethys and Panthalassa. There is no report of alatoconchidsfrom South China and North and South America despite their low-latitude positions during the Permian (Fig. 4). The absence of
alatoconchids in these two American continental blocks needs to beexplained somehow, e.g., in terms of paleo-provincialism under aunique setting with one supercontinent and one superocean.
4. Stratigraphic range
The biostratigraphic range of alatoconchids is apparently restrictedto the Cisuraian and Guadalupian, and no large bivalves have beenreported from the Lopingian, although the fossil localities are notmany in total (Table 1). Fig. 5 depicts the biostratigraphic range ofAlatoconchidae reported from the 9 areas mentioned above. Theoldest known alatoconchid is the Artinskian (Pseudofusulina ambiguaZone) Shikamaia (= Tanchintongia) perakensis (Runnegar & Gobbett)described fromMalaysia (Runnegar andGobbett, 1975) and Shikamaiaakasakaensis Ozaki from Neo (Japan). The latter was originallyreported from the Neoschwagerina craticulifera Zone (Yancey andOzaki, 1986) but Sano (1988) later corrected the biostratigraphichorizon of the same section into the P. ambigua Zone. The occurrenceslimited to Malaysia and Japan may indicate that Alatoconchidaeoriginated in western Panthalassa and secondarily migrated towardsthe west through Tethys.
On the other hand, the youngest is Alatoconchidae gen. et sp. indet.from the Capitanian (Yabeina–Lepidolina Zone) Iwato Formation inJapan, Saraburi Group in Thailand, and Velebit Formation in Croatia(Isozaki, 2006; Udchachon et al., 2007; Aljinović et al., 2008).Alatoconchids occur most frequently from theWordian (or Murgabian;Middle Guadalupian) Neoschwagerina Zone (Termier et al., 1973;Kochansky-Devidé, 1978; Boyd and Newell, 1979; Yancey and Ozaki,1986; Kiessling and Flügel, 2000; Isozaki, 2006). It is noteworthy thattheir size and shell thickness also reached a maximum in the Neosch-wagerina Zone. The highest horizon of Alatoconchidae to date is in theCapitanian LepidolinaZone (fusuline) in Japan (Isozaki, 2006), and in theCapitanian Yabeina Zone in Croatia (Aljinović et al., 2008). Alatoconch-idae from the Lepidolina Zone are much smaller than those from theNeoschwagerina Zone.
Because no occurrence has ever been reported from the Lopingian,alatoconchids likely became extinct at the end of the Guadalupianboth in Tethys and western Panthalassa. In other words, the Permiangigantic bivalve family was likely involved in the end-Guadalupianmass extinction.
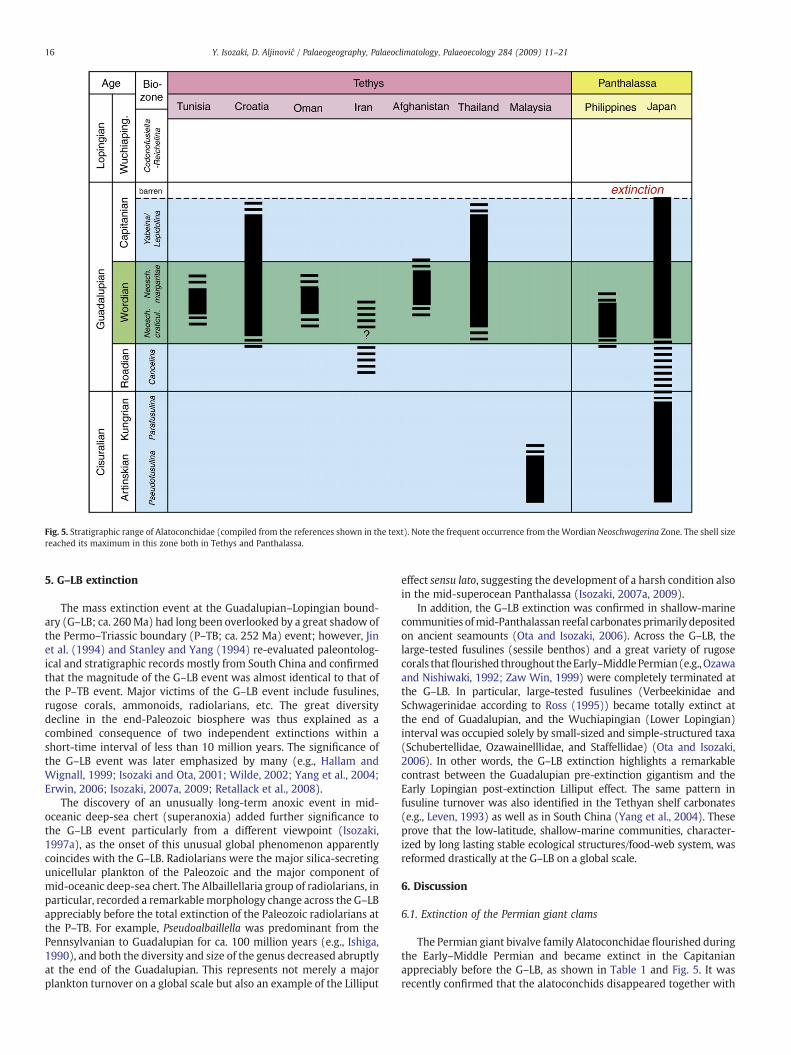
Fig. 5. Stratigraphic range of Alatoconchidae (compiled from the references shown in the text). Note the frequent occurrence from theWordian Neoschwagerina Zone. The shell sizereached its maximum in this zone both in Tethys and Panthalassa.
16 Y. Isozaki, D. Aljinović / Palaeogeography, Palaeoclimatology, Palaeoecology 284 (2009) 11–21
5. G–LB extinction
The mass extinction event at the Guadalupian–Lopingian bound-ary (G–LB; ca. 260Ma) had long been overlooked by a great shadow ofthe Permo–Triassic boundary (P–TB; ca. 252 Ma) event; however, Jinet al. (1994) and Stanley and Yang (1994) re-evaluated paleontolog-ical and stratigraphic records mostly from South China and confirmedthat the magnitude of the G–LB event was almost identical to that ofthe P–TB event. Major victims of the G–LB event include fusulines,rugose corals, ammonoids, radiolarians, etc. The great diversitydecline in the end-Paleozoic biosphere was thus explained as acombined consequence of two independent extinctions within ashort-time interval of less than 10 million years. The significance ofthe G–LB event was later emphasized by many (e.g., Hallam andWignall, 1999; Isozaki and Ota, 2001; Wilde, 2002; Yang et al., 2004;Erwin, 2006; Isozaki, 2007a, 2009; Retallack et al., 2008).
The discovery of an unusually long-term anoxic event in mid-oceanic deep-sea chert (superanoxia) added further significance tothe G–LB event particularly from a different viewpoint (Isozaki,1997a), as the onset of this unusual global phenomenon apparentlycoincides with the G–LB. Radiolarians were the major silica-secretingunicellular plankton of the Paleozoic and the major component ofmid-oceanic deep-sea chert. The Albaillellaria group of radiolarians, inparticular, recorded a remarkablemorphology change across the G–LBappreciably before the total extinction of the Paleozoic radiolarians atthe P–TB. For example, Pseudoalbaillella was predominant from thePennsylvanian to Guadalupian for ca. 100 million years (e.g., Ishiga,1990), and both the diversity and size of the genus decreased abruptlyat the end of the Guadalupian. This represents not merely a majorplankton turnover on a global scale but also an example of the Lilliput
effect sensu lato, suggesting the development of a harsh condition alsoin the mid-superocean Panthalassa (Isozaki, 2007a, 2009).
In addition, the G–LB extinction was confirmed in shallow-marinecommunities ofmid-Panthalassan reefal carbonates primarilydepositedon ancient seamounts (Ota and Isozaki, 2006). Across the G–LB, thelarge-tested fusulines (sessile benthos) and a great variety of rugosecorals thatflourished throughout theEarly–MiddlePermian(e.g.,Ozawaand Nishiwaki, 1992; Zaw Win, 1999) were completely terminated atthe G–LB. In particular, large-tested fusulines (Verbeekinidae andSchwagerinidae according to Ross (1995)) became totally extinct atthe end of Guadalupian, and the Wuchiapingian (Lower Lopingian)interval was occupied solely by small-sized and simple-structured taxa(Schubertellidae, Ozawainelllidae, and Staffellidae) (Ota and Isozaki,2006). In other words, the G–LB extinction highlights a remarkablecontrast between the Guadalupian pre-extinction gigantism and theEarly Lopingian post-extinction Lilliput effect. The same pattern infusuline turnover was also identified in the Tethyan shelf carbonates(e.g., Leven, 1993) as well as in South China (Yang et al., 2004). Theseprove that the low-latitude, shallow-marine communities, character-ized by long lasting stable ecological structures/food-web system, wasreformed drastically at the G–LB on a global scale.
6. Discussion
6.1. Extinction of the Permian giant clams
The Permian giant bivalve family Alatoconchidae flourished duringthe Early–Middle Permian and became extinct in the Capitanianappreciably before the G–LB, as shown in Table 1 and Fig. 5. It wasrecently confirmed that the alatoconchids disappeared together with

17Y. Isozaki, D. Aljinović / Palaeogeography, Palaeoclimatology, Palaeoecology 284 (2009) 11–21
Verbeeknidae fusuline; i.e., at the top of the Lepidolina Zone inKamura, and at the top of the Yabeina Zone in Akasaka, Japan (Isozaki,2006; Ota and Isozaki, 2006). Particularly at Kamura, the extinctionboth of Alatoconchidae and Verbeekinidae occurred at ca. 11 m belowthe G–LB, leaving a barren interval in the uppermost Capitanian (Otaand Isozaki, 2006). Also in the Velebit Mountains in Croatia,alatoconchids occur throughout the N100 m-thick Yabeina Zone anddisappear together with Yabeina below a dolomite that probablybelongs to the Lopingian (Aljinović et al., 2008). The additionaloccurrence from the Lepidolina Zone in Thailand (Udchachon et al.,2007) likewise supports the above-mentioned extinction horizon,though it is not yet well-constrained biostratigraphically. Otheralatoconchid localities (Table 1) have no direct constraint on theupper range limit; however, it is apparent that Alatoconchidaesurvived up to the Capitanian but could not extend their lineageinto the Lopingian both in Panthalassa and Tethys.
It is noteworthy that the extinction of alatoconchids took placesimultaneously with that of Verbeekinidae in mid-Panthalassa(Isozaki, 2006) and also in western Tethys (Aljinović et al., 2008),located almost on the opposite side of the globe (Fig. 4). Theextinction of Verbeekinidae was also accompanied by the sharpdiversity decline in the rugose coral family Waagenophyllidae and oflarge gastropods (e.g., Bellerophon, Pleurotomaria, etc. over 10 cm indiameter) at Akasaka (Hayasaka and Hayasaka, 1953; Minato andKato, 1965). Alatoconchidae, Verbeekinidae, and Waagenophyllidae,i.e. “tropical trio” by Isozaki (2006), occurred in intimate association(Yancey and Boyd, 1983), representing the typical Tethyan (low-latitude and tropical) shallow-marine fauna. The synchronousextinction/decline of the Guadalupian “tropical trio” in Panthalassaand Tethys indicates that they were involved in a major environ-mental change of global context; i.e. the G–LB event. It is noteworthythat the Capitanian global change appreciably before the G–LB mayhave preferentially affected low-latitude shallow-marine communi-ties and consequently led them to extinction.
6.2. Collapse of photosymbiosis
The common pattern of expansion/extinction indicates that theGuadalupian “tropical trio” essentially shared not only the samehabitat but also the same extinction scenario regardless of theirmutual distances in taxonomy. In this respect, living habit such asphotosymbiosis was the most likely significant common featureamong the three distinct clades. As the tropical shallow-marineenvironment inmid-ocean is generally poor in nutrients, animals withrelatively weak, diffusion-dependent metabolism generally cannotafford large body size. In order to pursue gigantism in such anoligotrophic condition, the bivalves and foraminifers need to deployunusual but appropriate strategies. Just like the modern giant clamTridacna and large benthic foraminifera hosting photosymbiontalgae (e.g., Cowen, 1983; Hallock, 1999), Alatoconchidae andVerbeekinidae probably performed photosymbiosis, as their shell/test structures are analogous to those of modern counterpartsfunctioning for hosting symbiont algae/cyanobacteria (Ross, 1972;Yancey and Boyd, 1983; Seilacher, 1990; Wilde, 2002; Vachard et al.,2004). The history of photosymbiosis in metazoans likely tracks backto the Cambrian archeothyatids (Cowen, 1988) that are known as thefirst reef-building metazoans, and the Paleozoic shallow-marineenvironments may have fostered various photosymbiotic couplesmany times. In addition to the large bivalves and fusulines, somePaleozoic rugose corals may likewise have performed photosymbiosisin the shallow-marine photic zone like tabulate corals, although thishas yet to be confirmed.
The “tropical trio” probably performed photosymbiosis during theEarly-Middle Permian both in Panthalassa and Tethys. Alatoconchidaelikely optimized the photosymbiotic system particularly in theWordian(Middle Guadalupian), as their size reached their maximum (up to 1 m
in length) simultaneously both in Tethys and Panthalassa. In the mid-Capitanian (the Lepidolina Zone), however, their shell size decreasedremarkably down to less than 30 cm in length, and they became extinctin the late Capitanian together with the other two members of the“tropical trio”. This suggests that the stable conditions in shallow-marineenvironments were perturbed in the Capitanian; in particular theoptimized photosymbiotic systems for the “tropical trio”were halted bycertain changes in conditions, such as temperature and nutrient supply.
For shallow-marine biota in low-latitudes in general, there is athreshold temperature for survival, particularly on the lower side.When global climate turns into cooling, there is no escape for tropical-adapted organisms, and they may suffer from severe diversity loss oreven extinction, owing to inefficient metabolism under low tempera-tures and possibly to a malfunction in photosymbiotic systems. Ingeneral, a drop in sea surface temperature may have loweredmetabolic rates in various marine invertebrates (Hochachka andSomero, 1984), and probably halted photosymbiotic communitiesefficiently by killing symbiont algae. Modern reef-forming corals withsymbiotic algae have a strict temperature limit, i.e., winter-minimumof 18–19 °C (Johnston, 1980). Wilde (2002), Yang et al. (2004), andOta and Isozaki (2006) suggested that the end-Guadalupian decline oflarge-tested fusulines (Verbeekinidae and Schwagerinidae) wasprobably related to certain problems in maintaining the photosym-biotic system. Wang and Sugiyama (2002) demonstrated the sharpdiversity loss of Permian corals at the G–LB, and these possiblephotosymbiotic organisms may have suffered in the same manner.
The Permian, in particular, the mid-Permian has been generallyregarded as an interval of global warming after the Gondwanaglaciation that lasted for nearly 50 million years from the LateCarboniferous to the Early Permian (e.g., Crowell, 1999; Jones andFielding, 2004). Throughout the late Early and Middle Permian, thebiodiversity stayed almost constant at the highest level of thePaleozoic (Sepkoski, 1984) until the end of the Guadalupian when amajor mass extinction occurred for the first time since the LateDevonian. The ultimate cause of the G–LB extinction has not beendeciphered yet; however, the recent documentation of a N3 millionyear-long interval of high positive δ13Ccarb values in the Capitaniancarbonates suggests that a temporary cooling (Capitanian Kamuraevent) appeared and worked as a potential kill mechanism for theshallow-marine biota in low latitudes at the end-Guadalupian (Isozakiet al., 2007a,b, 2009). For the Guadalupian “tropical trio” too adaptedto warm-water conditions, the Capitanian Kamura cooling event wasparticularly crucial.
The appearance of a temporary cooling in the Capitanian issupported by the eustatic sea-level change that records the Phaner-ozoic lowest stand straddling the G–LB (Hallam and Wignall, 1999;Tong et al., 1999; Haq and Schutter, 2008), i.e., the termination of themajor reef complex in Texas and extensive unconformity in SouthChina, by the mid-Permian chert event in high latitudes (Beauchampand Baud, 2002), and also by the short-time glaciation in Ver-khoyansk–Omolon (northeastern Russia) and in eastern Australia(Ustritsky, 1973; Fielding et al., 2008). In addition, some Early–MiddlePermian brachiopods derived from middle-high latitude domains(e.g., Attenuatella, Waagenites, Strophalosiina, and Comuqia) migratedtowards low latitudes and first appeared in the tropical zone in theCapitanian (Shen and Shi, 2002), supporting the development ofcooling. In general, a low-latitude sea is insensitive to temperaturechange compared with a high latitude sea due to its high heatcapacity; therefore, a temperature drop in low latitudes directlyindicates the appearance of global cooling. These relations suggestthat the late Middle Permian cooling was not restricted to the low-latitude domains but was global in extent.
Accordingly, all these observations suggest that the extinction ofthe “tropical trio” was related to the Capitanian global cooling(Kamura event; Fig. 6). Temporary cooling in the middle of thepost-Gondwanan long-term warming trend may have been

18 Y. Isozaki, D. Aljinović / Palaeogeography, Palaeoclimatology, Palaeoecology 284 (2009) 11–21
influential particularly for tropical-adapted, photosymbiosis-depen-dant biota. Stanley (1988) once discussed that a cooling event causedthe P–TB extinction but his idea is likely applicable to the G–LB eventrather than P–TB per se. The collapse of photosymbiotic systems mayhave played a significant role not only in the mass killing of the“tropical trio” but also in disturbing the Late Guadalupian ecologicalstructures including food web composed of other shallow-marine,sessile benthos.
6.3. Eutrophication shock
Hallock and Schlager (1986) pointed out that eutrophication isfatal to the survival of coral reefs and carbonate platforms. When theworld climate changes into cooling, global oceanic circulationbecomes accelerated and enhances upwelling of abundant nutrientsfrom the deep-sea to the ocean surface. Additional input of nutrientsto the surface will elevate the level of primary productivity throughalgal blooming that will make surface waters less transparent. Bydimming the photic zone in shallow-water carbonate facies, sucheutrophication can damage photosymbiosis-dependent sessile ben-thos fully adapted to oligotrophic conditions with a clear water-column. In the Holocene mid-oceanic reefs and/or carbonate plat-forms, various carbonate-secreting organisms were terminated byeutrophication associated with cooling (Hallock and Schlager, 1986).By demonstrating another example in the Eocene, Brasier (1995)pointed out that a similar change from an oligotrophic to a eutrophiccondition in shallow-marine environments triggered the extinction oflarge-tested foraminifera.
The extinction pattern of the Guadalupian “tropical trio” appearsanalogous to these cases in several aspects (Fig. 6). The extinction ofAlatoconchidae and Verbeekinidae in the middle of the Kamuracooling event was not likely a matter of contingency but a new line ofevidence for intimate cause–effect linkage between the cooling andextinction (Isozaki et al., 2007a,b). The topmost part of the Capitanianin mid-oceanic paleo-atoll carbonates at Kamura, even above theextinction horizon of Alatoconchidae and Verbeekinidae, is composed
Fig. 6. Schematic diagram showing the extinction pattern of the “tropical trio” (Alatoconcchange in the Capitanian (Late Guadalupian). The extinction of the “tropical trio” occurred inwas responsible for collapse of photosymbiotic systems in shallow-marine environments in
of black bituminous limestone with TOC over 0.1 wt.% (remarkablyhigher than the gray limestone below and above with TOC less than0.01 wt.%). This proves that the demise of the “tropical trio” occurredin the midst of high primary productivity associated with thedominant burial of organic material, i.e., eutrophication of a shallowsea. The extinction of Alatoconchidae under eutrophic conditions mayrepresent an actual case in the fossil record of the “paradox ofenrichment” predicted from numerical modeling (Rosenzweig, 1971).The environmental stress(es) may have appeared accumulativelyrather than abruptly. The “tropical trio”were terminated by and largeat the same time when the stress(es) exceeded the threshold forsurvival.
In the Capitanian Kamura cooling event, the tropical-adaptedgigantic clams plus large-tested fusulines became extinct, and thecontemporary rugose corals also faced a sharp diversity loss. Althoughit is not yet clear whether the die-off of symbiont algae by lowtemperature, eutrophication-driven dimming in the water-column, ora combination of the two was critically responsible for the killing, theGuadalupian photosymbiosis-dependent communities characterizedby the unusual gigantism disappeared dramatically in low-latitudedomains both in Tethys and Panthalassa. This might have consider-ably changed the ecological structure, including the food web, in theaftermath of the extinction. For a possible cause of the Capitaniancooling, and of the G–LB event per se, please refer to Isozaki (2007b,2009) who discussed a contemporary mantle plume activity andrelevant effects on the surface environments.
6.4. Bivalve gigantism tuned by climate
Gigantism in bivalves has had a long history since the middlePaleozoic. Giant bivalves appeared intermittently at least 5 times inthe Phanerozoic, i.e. 1) the Silurian–Devonian megalodonts, 2)Permian Alatoconchidae, 3) Late Triassic to Early Jurassic mega-lodonts/lithiotids, 4) Late Jurassic to Cretaceous rudists/inoceramids,and 5) Miocene–modern tridacnids (Fig. 7). Most of these aberrantbivalves lived in shallow-marine environments of low-latitude
hidae, Verbeekinidae, and Waagenophyllidae) and relevant environmental/ecologicalthe midst of the Kamura cooling event, suggesting that the low sea surface temperaturelow-latitude Tethys and Panthalassa.

Fig. 7. History of bivalve gigantism. Ranges of giant bivalves are after Kauffman and Johnson (1988), de Freitas et al. (1993), and this study. Sea-level curve is modified from Hallam(1992) based on Hallam andWignall (1999), Tong et al. (1999), and Haq and Schutter (2008). Note the 5 intervals of bivalve gigantism punctuated by cool/cold periods that includethewell-known Pennsylvanian–Early Permian Gondwana glaciation and recently documented Capitanian (Late Guadalupian) Kamura cooling event. The Toarcian event at the end ofthe Early Jurassic and the K–TB event at the end of the Cretaceous may have accompanied short-term cool spikes.
19Y. Isozaki, D. Aljinović / Palaeogeography, Palaeoclimatology, Palaeoecology 284 (2009) 11–21
domains or under extremely warm climates (e.g., Yancey and Boyd,1983; Kauffman and Johnson, 1988; Seilacher, 1990; de Freitas et al.,1993). This suggests that the gigantism represents a common styleof adaptation to warm-water (probably tropical) shallow-marineenvironments for bivalves regardless of age, and that bivalvesprobably attained their extraordinary size through symbiosis. Mostof them, including the oldest, most likely performed photosymbiosis,whereas others (e.g., lithiotids and inoceramids) may have deployeddifferent (non-photosymbiotic) strategies, such as chemosymbiosis(Seilacher, 1990) like modern Calyptogena. As demonstrated inFig. 7, the rise of photosymbiotic giant bivalves preferentiallyoccurred during warming periods. Ohno et al. (1995) postulatedthat algal–bivalve symbiosis was established several times in his-tory, and that the bivalves changed their habit from an infaunal toan epifaunal style via sciaphilous (shade-loving) stages once theyacquired photosymbiosis.
The gigantism of bivalves, specialized for the tropical domain,appears to have been fragile, because their survival was highlydependent on the stability of warm environments. Although thesymbiosis between bivalves and photosynthetic bacteria/algae likelystarted in the Early Silurian (de Freitas et al., 1993), it is noteworthythat none of the gigantic bivalves lasted a long time, not longer thantwo geologic periods (Fig. 7). The historical pattern in occurrence ofthe photosymbiotic giant bivalves in the Phanerozoic suggests thattheir intermittent rise and decline may have been controlled by along-term change in global climate.
Judging from the Phanerozoic sea-level curve, large photosymbio-tic bivalves likely flourished during warm periods, whereas theydeclined in diversity or died off during cool intervals (Fig. 7).Megalodonts appear to be the champion of a long lineage; however,
their total range consists of 2 distinct peaks, one in the Siluro-Devonian and the other in the Late Triassic–Early Jurassic, which areseparated by a remarkably long interval of decline/disappearance inthe Late Paleozoic that included the well-known Gondwana glacialperiod. Their disappearance in the Early Devonian (de Freitas et al.,1993) may also have been related to a short-term sea-level drop thatwas probably driven by cooling. As to the extinction of the PermianAlatoconchidae during the Kamura event, cooling even with a shortduration, appears to have been a crucial cause for terminatinggigantism. The extinction of megalodonts at the end of the EarlyJurassic and that of rudists at the end of the Cretaceous likewise mayhave been related to short-term cooling/eutrophication possiblyassociated with the Toarcian event and K–T boundary event,respectively. Likewise, the gradual increase in size of rudist apparentlyoccurred during a period of warming, whereas their sharp declinetook place during times of short-term cooling, as across the Jurassic/Cretaceous boundary, in the Middle–Late Aptian, in the Early–LateCenomanian, and in the Middle–Late Maastrichtian (Kauffman andJohnson, 1988; personal communication from Bogdan Jurkovsek).
Further study of the fundamental processes of bivalve gigantism inthe light of climate-tuned radiation/extinction patterns is definitelyneeded; however, the gigantism in tropical shallow-marine environ-ments may potentially serve as a sensitive monitor for seawatertemperature in low-latitude areas in the past.
7. Conclusions
This study summarizes all hitherto known occurrences of thePermian giant bivalve family Alatoconchidae, confirming its strati-graphic range, paleogeographic distribution, and association with

20 Y. Isozaki, D. Aljinović / Palaeogeography, Palaeoclimatology, Palaeoecology 284 (2009) 11–21
other fossils. By analyzing these datasets, the following conclusionsare made.
1) Alatoconchidae started and evolved in the Early–Middle Permian,and became extinct in the late Capitanian (Late Guadalupian).
2) The distribution of Alatoconchidae was nearly global but restrictedto low-latitude (tropical) domains both in Panthalassa and Paleo-Tethys.
3) The intimate association of Alatoconchidae with Verbeekinidae(large-tested fusulines) andWaagenophyllidae (rugose corals), i.e. a“tropical trio”, suggests a common ecological position in the tropics.
4) Alatoconchidae and Verbeekinidae performed photosymbiosisunder oligotrophic environments in tropical shallow seas, andprobably so did Waagenophyllidae.
5) The simultaneous extinction of the “tropical trio” was probablycaused by the Kamura cooling event and associated eutrophicationin the late Capitanian.
6) The recurrent rise and decline of bivalve gigantism in thePhanerozoic was fine-tuned by long-term global warming/cooling.
Acknowledgements
Michael Hautmann and Steven M. Stanley provided constructivereview comments on the originalmanuscript. Katarina Krizmanic, SanjaJapundzic, Jakov Radovcic, and Lucija Hrvat helped us in takingphotographs of the holotype specimen of Shikamaia (= Tanchintongia)ogulineci (Kochansky-Devide) stored in the Croatian Natural HistoryMuseum in Zagreb. Mongkol Udchachon, Wolfgang Kiessling, MichaelaBernecker, Bogdan Jurkovsek, Hiroshi Nishi, Hisayoshi Igo, TaketoYoshida, Masakazu Shimada, and Motomi Itoh, provided usefulreferences on the occurrence of Alatoconchidae, rudists, photosymbio-sis, and modern ecological structures. Brian F. Windley corrected theEnglish language. This study was supported by the Grant-in-Aid fromthe Japan Society for Promoting Sciences (no. 16204040, 20224012).
References
Aljinović, D., Isozaki, Y., Sremac, J., 2008. The occurrence of giant bivalve Alatoconchidaefrom the Yabeina Zone (Upper Guadalupian, Permian) in European Tethys.Gondwana Research 13, 275–287.
Beauchamp, B., Baud, A., 2002. Growth and demise of Permian biogenic chert alongnorthwest Pangea: evidence for end-Permian collapse of thermohaline circulation.Palaeogeography, Palaeoclimatology, Palaeoecology 187, 37–63.
Brasier, M.D., 1995. Fossil indicators of nutrient levels. 2. Evolution and extinction inrelation to oligotrophy. In: Bosence, D.W.J., Allison, P.A. (Eds.), Marine Palaeoenvi-ronmental Analysis from Fossils: Geological Society (London) Special Publication,vol. 83, pp. 133–150.
Boyd, D.W., Newell, N.D., 1979. Permian pelecypods from Tunisia. American MuseumNovitates 2686 22pp.
Cowen, R., 1983. Algal symbiosis and its recognition in the fossil record. In: Tevesz,M.J.S.,McCall, P.L. (Eds.), Biotic Interactions in Recent and Fossil Benthic Communities.Plenum, New York, pp. 431–478.
Cowen, R., 1988. The role of algal symbiosis in reefs through time. Palaios 3, 221–227.Crowell, J.C., 1999. Pre-Mesozoic ice ages: their bearing on understanding the climate
system. Geological Society of America Memoir 192 106 pp.de Freitas, T.A., Brunton, F., Bernecker, T., 1993. Silurian megalodont bivalves of the
Canadian Arctic and Australia: paleoecology and evolutionary significance. Palaios8, 450–464.
Erwin, D.H., 2006. Extinction. Princeton University Press, Princeton. 296 pp.Fielding, C.R., Frank, T.D., Birgenheier, L.P., Rygel, M.C., Jones, A.T., Roberts, J., 2008.
Stratigraphic imprint of the Late Palaeozoic Ice Age in eastern Australia: a record ofalternating glacial and nonglacial climate regime. Journal of the Geological Society(London) 165, 129–140.
Fraiser, M.L., Bottjer, D.J., 2004. The non-actualistic Early Triassic gastropod fauna: acase study of the Lower Triassic Sinbad limestone member. Palaios 19, 259–275.
Hallam, A., 1992. Phanerozoic Sea-Level Changes. Columbia University Press, New York.266 pp.
Hallam, A., Wignall, P.B., 1999. Mass extinctions and sea-level changes. Earth-ScienceReview 48, 217–250.
Hallock, P., 1999. Symbiont-bearing foraminifera. In: Gupta, B.K.S. (Ed.), ModernForaminifera. Kluwer, Dordrecht, pp. 123–139.
Hallock, P., Schlager, W., 1986. Nutrient excess and the demise of coral reefs andcarbonate platforms. Palaios 1, 389–398.
Haq, B.U., Schutter, S.R., 2008. A chronology of Paleozoic sea-level changes. Science 322,64–68.
Hayasaka, I., Hayasaka, S., 1953. Fossil assemblages of mollusks and brachiopods ofunusually large sizes from the Permian of Japan. Transaction Proceedings ofPalaeontological Society of Japan, new series, vol. 30, pp. 37–44.
Hochachka, P.W., Somero, G.N., 1984. Biochemical Adaptation. Princeton UniversityPress, Princeton. 250 pp.
Ishiga, H., 1990. Paleozoic radiolarians. In: Ichikawa, K., Mizutani, S., Hara, I., Hada, S.,Yao, A. (Eds.), Pre-Cretaceous Terranes of Japan. Nihon-Insatsu, Osaka, pp. 285–295.
Isozaki, Y., 1997a. Permo–Triassic boundary superanoxia and stratified superocean:records from lost deep-sea. Science 276, 235–238.
Isozaki, Y., 1997b. Jurassic accretion tectonics in Japan. The Island Arc 6, 25–52.Isozaki, Y., 2006. Guadalupian (Middle Permian) giant bivalve Alatoconchidae from a
mid-Panthalassa paleo-atoll complex in Kyushu, Japan: a unique communityassociated with Tethyan fusulines and corals. Proceedings of Japan Academy 82B,25–32.
Isozaki, Y., 2007a. Guadalupian–Lopingian boundary event in mid-Panthalassa:correlation of accreted deep-sea chert and mid-oceanic atoll carbonate. In:Wong, T.E. (Ed.), Proceedings of the XVth International Congress on Carboniferousand Permian Stratigraphy, Royal Netherlands Academy of Arts and Sciences, SpecialPublication, pp. 111–124.
Isozaki, Y., 2007b. Plume Winter scenario for biosphere catastrophe: the Permo–Triassic boundary case. In: Yuen, D., Maruyama, S., Karato, S., Windley, B.F. (Eds.),Superplumes: Beyond Plate Tectonics. Springer, Dordrecht, pp. 409–440.
Isozaki, Y., 2009. Integrated “plume winter” scenario for the double-phased extinctionduring the Paleozoic-Mesozoic transition: G-LB and P-TB events from a Pantha-lassan perspective. Journal of Asian Earth Sciences 36, 459–480.
Isozaki, Y., Amiscaray, E.A., Rillon, A., 1987. Permian, Triassic and Jurassic beddedradiolarian cherts in North Palawan Block, Philippines: evidence of Late Mesozoicsubduction–accretion. Journal of Geological Society of Philippines 41, 79–93.
Isozaki, Y., Ota, A., 2001. Middle/Upper Permian (Maokouan/Wuchapingian) boundaryin mid-oceanic paleo-atoll limestone in Kamura and Akasaka, Japan. Proceedings ofJapan Academy 77B, 104–109.
Isozaki, Y., Kawahata, H., Ota, A., 2007a. A unique carbon isotope record across theGuadalupian–Lopingian (Middle-Upper Permian) boundary in mid-oceanicpaleoatoll carbonates: the high-productivity “Kamura event” and its collapse inPanthalassa. Global and Planetary Change 55, 21–38.
Isozaki, Y., Kawahata, H., Minoshima, K., 2007b. The Capitanian (Permian) Kamuracooling event: the beginning of the Paleozoic–Mesozoic transition. Palaeoworld 16,16–30.
Jablonski,D., 1996. Body size andmacroevolution. In: Jablonski,D., Erwin,D.H., Lipps, J. (Eds.),Evolutionary Paleobiology. Univ. Chicago Press, Chicago, pp. 256–289.
Jin, Y.G., Zhang, J., Shang, Q.H., 1994. Two phases of the end-Permian massextinction. In: Embry, A.F., Beauchamp, B., Glass, D.J. (Eds.), Pangea: GlobalEnvironments and Resources: Memoir Canadian Society of Petroleum Geolo-gists, vol. 17, pp. 813–822.
Johnston, I.S., 1980. Theultrastructureof skeletogenesis inhermatypic corals. InternationalReview of Cytology 67, 171–188.
Jones, A.T., Fielding, C.R., 2004. Sedimentological record of the late Paleozoic glaciationin Queensland, Australia. Geology 32, 153–156.
Kauffman, E.G., Johnson, C.C., 1988. The morphological and ecological evolution ofMiddle and Upper Cretaceous reef-building rudists. Palaios 3, 194–216.
Kawaguti, S., 1983. The third record of association between bivalve mollusks andzooxanthellae. Proceedings of Japan Academy 59B, 17–20.
Kiessling, W., Flügel, E., 2000. Late Paleozoic and Late Triassic limestones from NorthPalawan Block (Philippines): microfacies and paleogeographical implications. Facies43, 6–16.
Kirschvink, J.L., Isozaki, Y., 2007. Extending the sensitivity of paleomagnetictechniques: Magnetostratigraphy of weakly-magnetized, organic-rich blacklimestone from the Permian of Japan. Abst. American Geophisical Union Fallmeeting, pp. GP43B–1223.
Kochansky-Devidé, V., 1978. Tanchintongia — eine aberrante permische Bivalve inEuropa. Paläontologische Zeitschrift 52, 213–218.
Leven, E.Y., 1993. Main events in Permian history of the Tethys and fusulinids.Startigraphy and Geological Correlation 1, 59–75.
Matsuda, K., Yamamoto, T., Akimoto, H., 2006. Molluscan fossils from the Shiroiwayamalimestone, Miyazaki prefecture. Bulletin of Miyazaki Prefectural Museum of Natureand History 27, 41–44 (in Japanese).
MacArthur, R.H., Wilson, E.O., 1967. The Theory of Island Biogeography. PrincetonUniversity Press, Princeton. 203 pp.
Minato, M., Kato, M., 1965. Waagenophyllidae. Journal of Faculty of Science, HokkaidoUniversity, ser. 4 12, 1–241.
Nicol, D., 1964. An essay on size of marine pelecypods. Journal of Paleontology 38,968–974.
Ohno, T., Katoh, T., Yamasu, T., 1995. The origin of algal-bivalve photo-symbiosis.Palaeontology 38, 1–21.
Ota, A., Isozaki, Y., 2006. Fusuline biotic turnover across the Guadalupian–Lopingian(Middle-Upper Permian) boundary inmid-oceanic carbonate buildups: biostratigraphyof accreted limestone in Japan. Journal of Asian Earth Sciences 26, 353–368.
Ozaki, K., 1968. Problematical fossils from the Permian limestone of Akasaka, Gifuprefecture. Yokohama National University Science Report (sect. 2). Biological andGeological Sciences 14, 27–33.
Ozawa, T., Nishiwaki, N., 1992. Permian Tethyan biota and sedimentary facies of theAkasaka Limestone Group. Field Trip Guidebook B13 of International GeologicalCongress, Kyoto, pp. 189–195.
Pianka, E.R., 1978. Evolutionary Ecology, 2nd ed. Harper & Row, New York. 397 pp.Pillevat, A., 1993. Les bloc exotics du Sultanat d'Oman. Evolution paleogeographique
d'une marge massive flexurale. Memoir Geologique Lausanne 17 249 pp.

21Y. Isozaki, D. Aljinović / Palaeogeography, Palaeoclimatology, Palaeoecology 284 (2009) 11–21
Retallack, G.J., Metzger, C.A., Greaver, T., Jahren, A.H., Smith, R.M.H., Sheldon, N.D., 2008.Middle–Late Permian extinction on land. Geological Society of America Bulletin118, 1398–1411.
Rosenzweig, M.L., 1971. Paradox of enrichment: destabilization of exploitationecosystems in ecological time. Science 171, 385–387.
Ross, C.A., 1972. Paleobiological analysis of fusulinacean (Foraminiferida) shellmorphology.Journal of Paleontology 46, 719–728.
Ross, C.A., 1995. Permian fusulinaceans. In: Scholle, P.A., Peryt, T.M., Ulmer-Scholle,D.S. (Eds.), The Permian of Northern Pangea, 1: Paleogeography, Paleoclimates,Stratigraphy. Springer, New York, pp. 167–185.
Runnegar, B., Gobbett, D., 1975. Tanchintongia gen. nov., a bizarre Permian myalinidbivalve from West Malaysia and Japan. Palaeontology 18, 315–322.
Saito, M., Miyazaki, K., Toshimitsu, S., Hoshizumi, H., 2005. Geology of the TomochiDistrict. Quadrangle Series 1:50,000, Geological Survey of Japan, AIST. 218 pp. (inJapanese with English abstract).
Sano, H., 1988. Permian oceanic rocks of Mino terrane, central Japan, part II. Limestonefacies. Journal of Geological Society of Japan 94, 963–976.
Schubert, J.K., Bottjer, D.J., 1995. Aftermath of the Permian–Triassic mass extinctionevent: paleoecology of Lower Triassic carbonates in the western USA. Palaeogeo-graphy, Palaeoclimatology, Palaeoecology 116, 1–39.
Scotese, C.R., Langford, R.P., 1995. Pangea and the paleogeography of the Permian. In:Scholle, P.A., Peryt, T.M., Ulmer-Scholle, D.S. (Eds.), The Permian of NorthernPangea, vol. 1. Springer, New York, pp. 3–19.
Seilacher, A., 1990. Aberrations in bivalve evolution related to photo- and chemo-symbiosis. Historical Biology 3, 289–311.
Sepkoski Jr., J.J., 1984. A kinetic model of Phanerozoic taxonomic diversity. III. Post-Paleozoic families and mass extinctions. Paleobiology 10, 246–267.
Shen, S.Z., Shi, G.R., 2002. Paleobiogeographical extinction patterns of Permianbrachiopods in the Asian–Western Pacific region. Paleobiology 28, 449–463.
Stanley, S.M., 1973. An explanation for Cope's rule. Evolution 27, 1–26.Stanley, S.M., 1988. Climatic cooling andmass extinction of Paleozoic reef communities.
Palaios 3, 228–232.Stanley, S.M., Yang, X.N., 1994. A double mass extinction at the end of the Paleozoic era.
Science 266, 1340–1344.Termier, H., Termier, G., De Lapparent, A.F., 1973. Grands bivalves récifaux du Permien
supérier de l'Afghanistan central. Annales de la Societe Géologique de Nord 93,75–80.
Thiele, O., Ticky, G., 1980. Über das Aufreten von Tanchintongia (Bivalvia) im Perm desIran. Der Karinthin 82, 166–173.
Tong, J.N., Yin, H.F., Zhang, K.X., 1999. Permian and Triassic sequence stratigraphy andsea level changes of eastern Yangtze platform. Journal of China University ofGeosciences 10, 161–169.
Twitchett, R.J., 1999. Palaeoenvironments and faunal recovery after the end-Permianmass extinction. Palaeogeography, Palaeoclimatology, Palaeoecology 154, 27–37.
Udchachon, M., Chonglakmani, C., Campbell, H., Thanee, N., 2007. Late Middle Permianalatoconchid-bearing limestones from the south of the Khao Khwang platform,central Thailand. In: Tantiwanit, W. (Ed.), Proceedings of the InternationalConference on Geology of Thailand: Towards Sustainable Development andSufficiency Economy. Bangkok, pp. 169–176.
Urbaneck, A., 1993. Biotic crises in the history of Upper Silurian graptoloids: apalaeobiological model. Historical Biology 7, 29–50.
Ustritsky, V.I., 1973. Permian climate. In: Logan, A., Hills, L.V. (Eds.), The Permian andTriassic Systems and their mutual boundary, Canadian Society of PetroleumGeologists Memoire, vol. 2, pp. 733–744.
Vachard, D., Munnecke, A., Servais, T., 2004. New SEM observations of keriothecal walls:implications for the evolution of fusulinida. Journal of Foraminiferal Research 34,232–242.
Vermeij, G.J., 1977. The Mesozoic marine revolution: evidence from snails, predatorsand grazers. Paleobiology 3, 245–258.
Vogel, K., 1975. Endosymbiotic algae in rudists? Palaeogeography, Palaeoclimatology,Palaeoecology 17, 327–332.
Wakita, K., 1988. Origin of chaotically mixed rock bodies in the Early Jurassic to EarlyCretaceous sedimentary complex of the Mino terrane, central Japan. Bulletin ofGeological Survey of Japan 39, 675–757.
Wang, X.D., Sugiyama, T., 2002. Diversity and extinction patterns of Permian coralfaunas of China. Lethaia 33, 285–294.
Wiedlich, O., Bernecker, M., 2007. Differential severity of Permian–Triassic environ-mental changes on Tethyan shallow-water carbonate platforms. Global andPlanetary Change 55, 209–235.
Wilde, G.L., 2002. End Permian: end fusulinaceans. In: Hills, L.V., Henderson, C.M.,Bamber, E.W. (Eds.), Carboniferous and Permian of the World. Memoir CanadianSociety of Petroleum Geologists, vol. 19, pp. 616–629.
Yancey, T.E., Boyd, D.W., 1983. Revision of the Alatoconchidae: a remarkable family ofPermian bivalve. Palaeontology 26, 497–520.
Yancey, T.E., Ozaki, K., 1986. Redescription of the genus Shikamaia and clarification of thehinge characters of the family Alatoconchidae (bivalvia). Journal of Paleontology 60,116–125.
Yang, X.N., Liu, J.R., Shi, G.J., 2004. Extinction process and patterns of Middle Permianfusulimaceans in southwest China. Lethaia 37, 139–147.
Yonge, C.M., 1936. Mode of life, feeding, digestion and symbiosis with zooxanthellae inthe Tridacnidae. Great Barrier Reef Expedition, Scientific Reports, 283–321. BritishMuseum, London.
Zamoras, L.R., Matsuoka, A., 2004. Accretion and postaccretion tectonics of the CalamianIslands, North Palawan Block, Philippines. The Island Arc 12, 506–519.
Zaw Win, 1999. Fusuline biostratigraphy and palaeontology of the Akasaka Limestone,Gifu Prefecture, Japan. Bulletin of Kitakyushu Museum of Natural History 18, 1–76.


















