Electrophysiology of Neuromuscular Transmission
35
Electrophysiology of Neuromuscular transmission Dr. Fiaz Hamsath Moderator: Dr. Maya.
-
Upload
fiaz-hamsath -
Category
Education
-
view
78 -
download
0
Transcript of Electrophysiology of Neuromuscular Transmission

Electrophysiology of Neuromuscular transmissionDr. Fiaz HamsathModerator: Dr. Maya.

Impulse generation in a Neuron Resting State:
Membrane of axon is more permeable to K+ ions and impermeable to Na+ ions and the negatively charged proteins present in axoplasm.
Plasma in the axon: Rich in K+ ions and proteins and low concentration of Na+.
Fluid outside the axon: Rich in Na+ and low concentration of K+ ions. Concentration gradient formed. Active transport of ions occurs across the membrane by the sodium-
potassium pump where 3 Na+ are transported outwards and 2 K+ move into the cell.
This makes the outer surface of the membrane positively charged while the inner surface is negatively charged.
The electrical potential difference across the resting plasma membrane is known as resting potential.

Depolarised state: When we apply a stimulus at a site on the polarised membrane, the
membrane at the site becomes freely permeable to Na+ ions. Due to this, Na+ move inside the cell and the outer side of the
membrane becomes negatively charged and the inner side becomes positively charged.
The electrical potential difference generated across the plasma membrane at this site is known as action potential or nerve impulse.
This area becomes a stimulus for the neighbouring area of the membrane which becomes depolarised.
The former membrane becomes repolarised due to the movement of sodium ions outside the cell.

After the stimulus goes above the threshold level, more gated ion channels open and allow more Na+ inside the cell. This causes complete depolarization of the neuron and an action potential is created.
All-or-none phenomenon: If a stimulus doesn’t exceed the threshold level and cause all the gates to open, no action potential results. However, after the threshold is crossed, complete depolarization occurs and the stimulus will be transmitted.

Repolarization: After the inside of the cell becomes flooded with Na+, the gated ion
channels on the inside of the membrane open to allow the K+ to move to the outside of the membrane.
Just after the K+ gates open, the Na+ gates close, otherwise, the membrane couldn’t repolarize.


Hyperpolarization: When the K+ gates finally close, the neuron has slightly more K+ on
the outside than it has Na+ on the inside. This causes the membrane potential to drop slightly lower than the
resting potential, and the membrane is said to be hyperpolarized because it has a greater potential.
After the impulse has travelled through the neuron, the action potential is over, and the cell membrane returns to normal (that is, the resting potential).

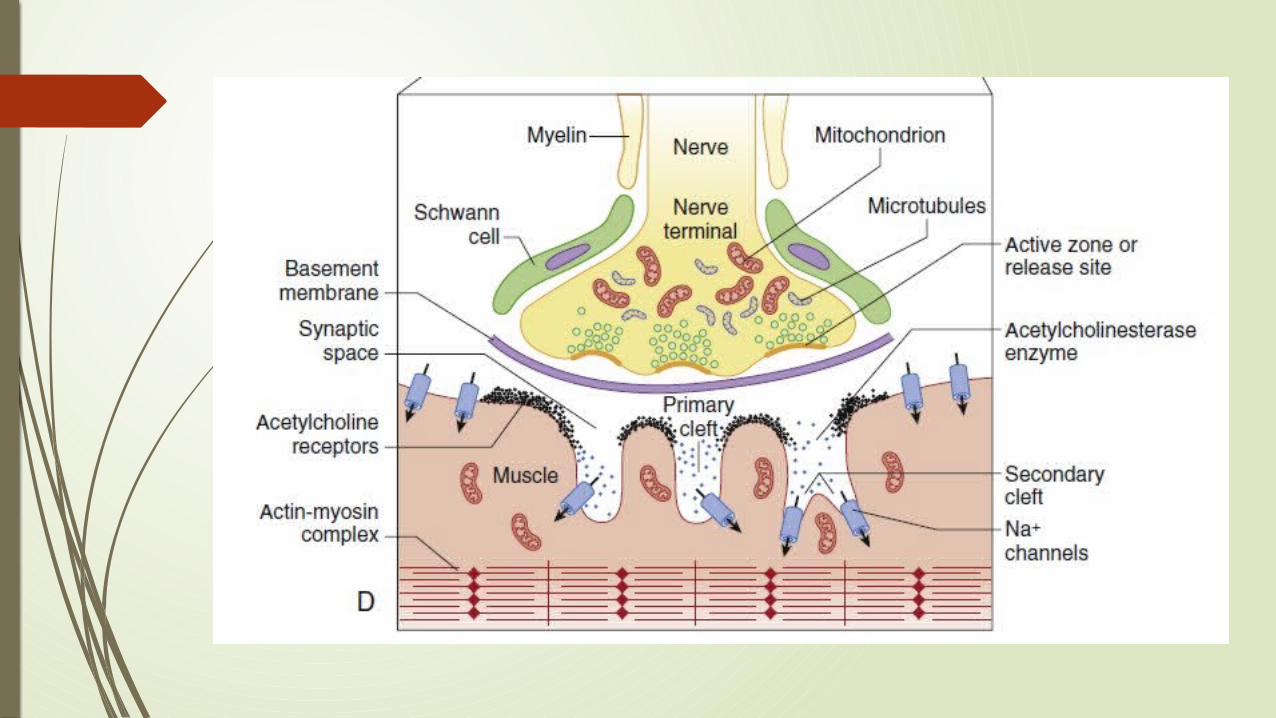

Formation of Acetyl Choline
Simple products Choline and Acetate are obtained from the environment of nerve ending.
Choline is transported by a special system from ECF to cytoplasm. Acetate is transported in the form of ACoA from mitochondria. The enzyme Choline acetyl transferase brings about the reaction of
Choline and acetate to form acetylcholine. After synthesis, ACh is stored in cytoplasm, until it is transported and
incorporated into vesicles.


Role of Calcium in nerve impulse transmission During a nerve action potential, Na+ from outside flows across the
membrane. Resulting depolarizing voltage opens Ca2+ channels, which allows entry
of Ca2+ ions > Causes Acetyl Choline to be released. If calcium is not present, then depolarization of the nerve, even by
electrical stimulation, will not produce the release of transmitter. Doubling the ECF Ca2+ results in a 16-fold increase in the quantal
content of an end-plate potential. Calcium current persists until the membrane potential is returned to
normal by outward fluxes of K+ from inside the nerve cell.

Function of K+ channel is to limit entry of calcium into the nerve and therefore depolarization [The Calcium current can be prolonged by potassium channel blockers]
Post-tetanic potentiation: An effect of increasing calcium in the nerve ending. Occurs after a nerve of a patient paralyzed with an NDMR is
stimulated at high, tetanic frequencies. Ca2+ enters the nerve with every stimulus, but it accumulates
during the tetanic period because it cannot be excreted as quickly as the nerve is stimulated.
More than the usual amount of Ca2+ causes the release of more than the normal amount of ACh.
Abnrmally large amount of acetylcholine antagonizes the relaxant and causes the characteristic increase in the size of the twitch.

Calcium channels: Important for the release of transmitter: P channels and the slower L
channels. Voltage dependent and are opened and closed by changes in
membrane voltage caused by the nerve action potential. Alterations in entry of calcium into the nerve ending can also alter
the release of transmitter. Eaton-Lambert myasthenic syndrome: Autoimmune disease in
which antibodies are directed against voltage-gated calcium channels at nerve endings > Decreased release of transmitter > Inadequate depolarization > Muscle weakness.

Higher-than-normal concentrations of bivalent inorganic cations (e.g., magnesium, cadmium, manganese) can also block the entry of calcium through P channels > Profoundly impair NMT.
This mechanism is behind the typical muscle weakness and potentiation of the effect of muscle relaxants in a mother and fetus when MgSO4 is administered to treat preeclampsia.
CCBs have no effect on P type Ca2+ channels, they have profound effect on slower L-type channels instead.
So CCBs have no significant effect at therapeutic doses on the normal release of acetylcholine or on the strength of normal neuromuscular transmission.

SNARE proteins: Release occurs when calcium ions enter the nerve through the P channels
lined up on the sides of the active zones by soluble N-ethylmaleimide–sensitive attachment protein receptor (SNARE) proteins.
Involved in fusion, docking, and release of acetylcholine at the active zone. Calcium activate these proteins on the vesicle wall - Docking. The Calcium sensor in the synaptic vesicle is Synaptotagmin, a SNARE
protein.


Model of protein mediated Exocytosis Action potential > calcium ions enter > synapsin becomes
phosphorylated > Frees the vesicle from its attachment to the cytoskeleton.
Synaptobrevin on the vesicle forms ternary complex with syntaxin and SNAP-25 (complexes attached to the plasma membrane).
Synaptotagmin, the protein on the vesicular membrane that acts as a calcium sensor, localizes the synaptic vesicles to synaptic zones rich in calcium channels, and stabilizes the vesicles in the docked state.
Assembly of the ternary complex forces the vesicle to move close to the underlying nerve terminal membrane.
Close proximity of release sites, calcium channels, and synaptic vesicles lead to a burst of release of new transmitter synchronous with the stimulus.

Three types of vesicles based on release of NT:1. “Kiss and run”: Stay open briefly before retrieval and do not
completely collapse.2. Compensatory: Stay open longer and probably do not completely
collapse.3. Stranded: Completely collapse and are not retrieved until another
stimulus is delivered.


Action of Botulinum toxin on Exocytosis Botulinum neurotoxin selectively digests one or all of these SNARE
proteins and blocks exocytosis of the vesicles. Ultimately results in muscle weakness or more profound muscle
paralysis. Botulinum toxin consists of two protein segments known as heavy and
light chains. Heavy chain interacts with polysialogangliosides in the cell membrane
and synaptotagmin on the vesicle to enter the vesicle. In the vesicle, the light chain inactivates neuromuscular transmission by
breakdown and thereby inhibits the function of SNARE proteins.

Clinical uses of Botulinum toxin: Treat spasticity or spasm in several neurologic and surgical diseases. To prevent hyperhidrosis in patients with excessive sweating. Cosmetically to correct wrinkles.

Binding of Acetyl Choline to Post junctional AChR: AChRs are synthesized in muscle cells. Anchored to the end-plate membrane by a special protein rapsyn. The binding site for acetylcholine is on each of the α1-subunits, which is
located on the extracellular component. This is the site of competition between receptor agonists and
antagonists.

Depolarization of Endplate by Acetyl Choline: Normally, pore of the channel surrounded by 5 subunits of nAChR is
closed by approximation of the cylinders (i.e., subunits). When an agonist occupies both α-subunit sites, the protein molecule
undergoes a conformational change with a twisting movement along the central axis of the receptor.
Results in the opening of the central channel through which ions can flow along a concentration gradient.
When the central channel is open, Na+ and Ca2+ flow from the outside of the cell to the inside and K+ flows from the inside to the outside.
The net current is depolarizing and creates the end-plate potential that stimulates the muscle to contract.
The pulse stops when the channel closes by a reversed mechanical conformation.


Conformational changes in Sodium Channel during depolarization Sodium channel is a cylindrical transmembrane protein through which
sodium ions can flow. Two parts of its structure act as gates that allow or stop the flow of
sodium ions. Both gates must be open if sodium is to flow through the channel;
closing of either cuts off the flow. Because these two gates act sequentially, a sodium channel has three
functional conformational states and can progressively move from one state to another
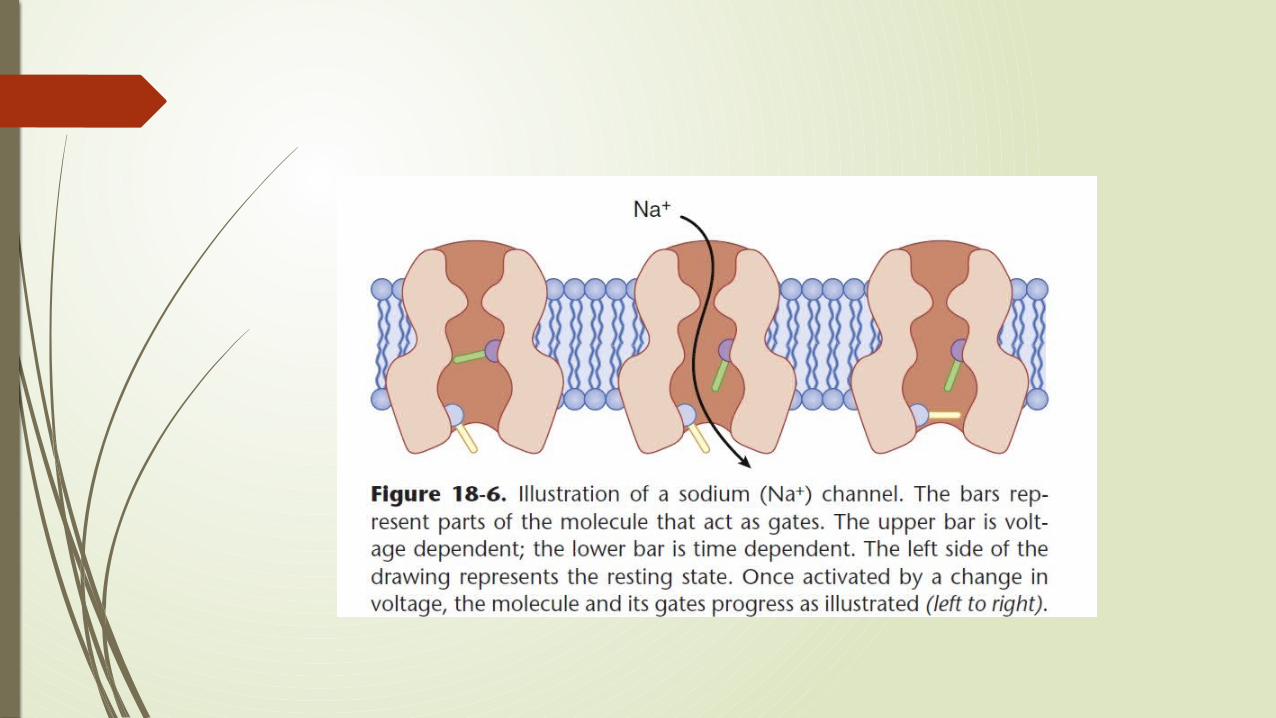

Resting state: Lower gate, the time-dependent or inactivation gate is open, but the upper gate, the voltage-dependent gate is closed, and sodium ions cannot pass.
When adjacent membrane is depolarized, the top gate opens. Because the bottom (time-dependent) gate is still open, sodium flows through the channel.
Voltage-dependent gate stays open as long as the molecule is experiencing a depolarizing influence from the membrane around it,.
Shortly after the voltage dependent gate opens, the bottom gate closes and again cuts off the flow of ions.

The bottom gate cannot open until the voltage-dependent gate closes. When depolarization of the end plate stops, the voltage-dependent gate
closes, the time-dependent gate opens, and the sodium channel returns to its resting state.

End Plate Potential
Depolarizations of skeletal muscle fibers caused by neurotransmitters binding to the postsynaptic membrane.
Called as "end plates" because the postsynaptic terminals of muscle fibers have a large, saucer-like appearance.
Two types of EPPs:1. Miniature EPP2. Stimulus Evoked EP

Miniature End Plate Potentials (MEPPs) The small (~0.4mV) depolarizations of the postsynaptic terminal caused
by the release of a single vesicle into the synaptic cleft. Neurotransmitter vesicles containing acetylcholine collide spontaneously
with the nerve terminal and release acetylcholine into the neuromuscular junction even without a signal from the axon.
These small depolarizations are not enough to reach threshold and so an action potential in the postsynaptic membrane does not occur.
They are unitary responses; that is, there is a minimum size for the MEPP, and the sizes of all MEPPs are equal to or multiples of this minimum size.
They are produced by uniformly sized packages, or quanta, of transmitter released from the nerve (in the absence of stimulation).
So, MEPP is the smallest possible depolarization which can be induced in a muscle.

Stimulus Evoked End Plate Potential
Additive depolarization produced by the synchronous discharge of quanta from several hundred vesicles.
When SEPPs cause the membrane to reach threshold (-mV), the voltage gated ion channels in the postsynaptic membrane open causes in influx of sodium ions and a sharp spike in depolarization.
This spike causes an action potential to occur and propagate down the postsynaptic membrane leading to muscle contraction.
In a normal muscular contraction, approximately 100-200 acetylcholine vesicles are released causing a depolarization that is 100 times greater in magnitude than a MEPP.
This causes the membrane potential to depolarize.


Physiology of Muscle Contraction
Voltage gated Na+ channels in the peri-junctional membrane opens when enough voltage is developed across them.
Resulting AP propagates along the muscle membrane and T tubule system, which are the invaginations of the muscle membrane.
This AP results in opening of Na+ channels and release Calcium from sarcoplasmic reticulm.
Intracellullar Ca2+ allows contractile protein actin and myosin to interact, which results in muscle contraction.

THANK YOU







![Physiology and Pathophysiology of Neuromuscular Transmission€¦ · 1 Physiology and Pathophysiology of Neuromuscular Transmission “The neuromuscular junction... [is] an experimentally](https://static.fdocuments.in/doc/165x107/5b1acdb37f8b9a41258e0a00/physiology-and-pathophysiology-of-neuromuscular-1-physiology-and-pathophysiology.jpg)