Electron Microscopic Examination of …human disease agents Corynebacterium diph-theriae type gravis...
6
JOURNAL OF BACTERIOLOGY, Mar. 1969, p. 1480-1485 Vol. 97, No. 3 Copyright @ 1969 American Society for Microbiology Printed in U.S.A. Electron Microscopic Examination of Corynebacterium ovis G. C. HARD1 Department of Veterinary Pathology, University of Sydney, New South Wales, Australia Received for publication 26 November 1968 Corynebacterium ovis (C. pseudotuberculosis) was examined by electron micros- copy after being subjected to various methods of fixation. The organism exhibited a fine structure similar to other corynebacterial species in the appearance of its cell wall, plasma membrane, nuclear apparatus, cytoplasmic matrix, wealth and com- plexity of intracytoplasmic membrane systems, and polyphosphate granules. An outstanding structural feature was the existence of an electron-dense, floccular layer external to the cell wall which both ligroin and acetone-methanol extractions demon- strated to be the previously postulated surface lipid of this organism. The only varia- tions in structure evident between virulent and attenuated strains was a quantitative difference in the thickness and appearance of the surface lipid. The observation of this layer provided a basis for explaining the surface properties of C. ovis, with par- ticular respect to its clumping capacity in suspension, the waxiness of its growth on solid media, and its ability to grow as a pellicle on suitable liquid media. The variation in the visible amount of surface lipid between the virulent and avirulent strains ade- quately explained the divergence of these three surface properties between the strains. In the past 8 years, the ultrastructural morphology of a number of bacterial species within the family Corynebacteriaceae has been described, including those of the human and ani- mal pathogen Listeria monocytogenes (5, 15), the human disease agents Corynebacterium diph- theriae type gravis (9) and C. diphtheriae type mitis (17), C. minutissimum, the causal agent of human erythrasma (13, 14), a plant pathogen C. fascians (6), and the nonpathogenic C. xerosis (21). Known pathogens of domestic animals be- longing to the genus Corynebacterium have re- ceived little attention in ultrastructural studies however. Corynebacterium ovis (C. pseudotuberculosis), the causal agent of caseous lymphadenitis in sheep and ulcerative lymphangitis in horses, dis- plays certain unusual characteristics such as the marked propensity for organisms to adhere to- gether closely as firm aggregates in suspension. This property is doubtless associated with the waxy integrity possessed by colonies grown on solid media and with the characteristic ability of C. ovis to grow as a surface pellicle in some liquid media and as submerged clumps or balls in others 1 Present address: Toxicology Research Unit, Medical Re- search Council Laboratories, Woodmansterne Road, Carshal- ton, Surrey, England. (3). It has been shown that the organism possesses a high percentage of easily extractable, and so presumably surface, lipid, which on removal with ligroin does not impair viability (2, 4). Jolly (8) has drawn attention to the possibility that such a large amount of surface lipid might provide the bacterium with a structural barrier against the antibacterial mechanisms of phagocytic host cells. An electron microscopic examination of C. ovis was undertaken to determine the existence of a structural basis for some of the physical and pathogenic properties of this organism. MATERIALS AND METHODS Organisms and culture. Three strains of C. ovis were investigated, one virulent and two attenuated, all initially recovered from sheep in New South Wales. The attenuated strains 137E and 133A were derived from virulent strains by repeated in vitro passage on solid media. They retained some degree of pathogenicity in comparatively high doses, how- ever. The virulent strain 137C/2 was uniformly hemolytic on sheep blood-agar and strongly toxino- genic in suitable media. Strain 133A was also uni- formly hemolytic on sheep blood-agar and tox- inogenic, but less so than 137C/2. Strain 137E was neither hemolytic nor toxinogenic. Colonies were freshly subcultured from lyophilized preparations onto sheep blood-agar with or without a 0.04% content of sodium tellurite, tryptose agar, or viande- 1480 on February 15, 2020 by guest http://jb.asm.org/ Downloaded from
Transcript of Electron Microscopic Examination of …human disease agents Corynebacterium diph-theriae type gravis...

JOURNAL OF BACTERIOLOGY, Mar. 1969, p. 1480-1485 Vol. 97, No. 3Copyright @ 1969 American Society for Microbiology Printed in U.S.A.
Electron Microscopic Examination ofCorynebacterium ovis
G. C. HARD1Department of Veterinary Pathology, University of Sydney, New South Wales, Australia
Received for publication 26 November 1968
Corynebacterium ovis (C. pseudotuberculosis) was examined by electron micros-copy after being subjected to various methods of fixation. The organism exhibiteda fine structure similar to other corynebacterial species in the appearance of its cellwall, plasma membrane, nuclear apparatus, cytoplasmic matrix, wealth and com-plexity of intracytoplasmic membrane systems, and polyphosphate granules. Anoutstanding structural feature was the existence of an electron-dense, floccular layerexternal to the cell wall which both ligroin and acetone-methanol extractions demon-strated to be the previously postulated surface lipid of this organism. The only varia-tions in structure evident between virulent and attenuated strains was a quantitativedifference in the thickness and appearance of the surface lipid. The observation ofthis layer provided a basis for explaining the surface properties of C. ovis, with par-ticular respect to its clumping capacity in suspension, the waxiness of its growth onsolid media, and its ability to grow as a pellicle on suitable liquid media. The variationin the visible amount of surface lipid between the virulent and avirulent strains ade-quately explained the divergence of these three surface properties between thestrains.
In the past 8 years, the ultrastructuralmorphology of a number of bacterial specieswithin the family Corynebacteriaceae has beendescribed, including those of the human and ani-mal pathogen Listeria monocytogenes (5, 15), thehuman disease agents Corynebacterium diph-theriae type gravis (9) and C. diphtheriae typemitis (17), C. minutissimum, the causal agent ofhuman erythrasma (13, 14), a plant pathogen C.fascians (6), and the nonpathogenic C. xerosis(21). Known pathogens of domestic animals be-longing to the genus Corynebacterium have re-ceived little attention in ultrastructural studieshowever.
Corynebacterium ovis (C. pseudotuberculosis),the causal agent of caseous lymphadenitis insheep and ulcerative lymphangitis in horses, dis-plays certain unusual characteristics such as themarked propensity for organisms to adhere to-gether closely as firm aggregates in suspension.This property is doubtless associated with thewaxy integrity possessed by colonies grown onsolid media and with the characteristic ability ofC. ovis to grow as a surface pellicle in some liquidmedia and as submerged clumps or balls in others
1 Present address: Toxicology Research Unit, Medical Re-search Council Laboratories, Woodmansterne Road, Carshal-ton, Surrey, England.
(3). It has been shown that the organism possessesa high percentage of easily extractable, and sopresumably surface, lipid, which on removal withligroin does not impair viability (2, 4). Jolly (8)has drawn attention to the possibility that such alarge amount of surface lipid might provide thebacterium with a structural barrier against theantibacterial mechanisms of phagocytic host cells.An electron microscopic examination of C. oviswas undertaken to determine the existence of astructural basis for some of the physical andpathogenic properties of this organism.
MATERIALS AND METHODSOrganisms and culture. Three strains of C. ovis
were investigated, one virulent and two attenuated,all initially recovered from sheep in New SouthWales. The attenuated strains 137E and 133A werederived from virulent strains by repeated in vitropassage on solid media. They retained some degreeof pathogenicity in comparatively high doses, how-ever. The virulent strain 137C/2 was uniformlyhemolytic on sheep blood-agar and strongly toxino-genic in suitable media. Strain 133A was also uni-formly hemolytic on sheep blood-agar and tox-inogenic, but less so than 137C/2. Strain 137E wasneither hemolytic nor toxinogenic. Colonies werefreshly subcultured from lyophilized preparationsonto sheep blood-agar with or without a 0.04%content of sodium tellurite, tryptose agar, or viande-
1480
on February 15, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

C. OVIS FINE STRUCTURE
foie broth. The period of culture varied from 24 to 96hr.
Preparation of specimens for electron microscopy.Small colonies cultured on solid media were removedin toto and prepared according to the procedure ofKellenberger's standard fixation (10). Samples ofsurface pellicle growth were handled in the sameway, but other broth cultures were centrifuged toproduce small pellets. The organism was also ex-amined after fixation in 0.6% potassium permanga-nate in veronal acetate buffer (12) and after fixationin 6.25% glutaraldehyde in 0.1 M cacodylate buffer(pH 7.4) for 1 hr followed by postosmification for 2hr in 1% OsO in veronal buffer. Some colonies wereimmersed in a sterile solution of ligroin and mechani-cally agitated for 5 hr prior to washing in Kellen-berger's buffer and subsequent fixation. After post-fixation staining in 0.5% uranyl acetate, the specimenswere dehydrated in a graded series of alcohols fol-lowed by propylene oxide, and thence into a 60-40mixture of Araldite and propylene oxide for up to 12hr. Infiltration of the samples was allowed to con-tinue for 16 hr in Araldite before polymerization.Sections were cut on either an LKB ultramicrotomeor Porter-Blum MT-2 microtome with glass knives.All sections were stained on the grid with Reynold'slead citrate (18).
Bacterial cell walls were prepared in a Mickle
disintegrator by the method of Salton and Horne(19) from both a fresh culture of 137C/2 and a freeze-dried, acetone- and methanol-extracted strain ofvirulent 133. Cell walls were spun out at 10,000 X g,and a Gram stain on a smear of the final pellet showed90% of the content to be cell wall ghosts. Samples ofthese preparations were negatively stained in 1%aqueous phosphotungstic acid, according to themethod of Brenner and Horne (1).
Electron microscopy. Specimens were examined ina Philips EM 200 microscope at 80 kv with a 50-Amobjective aperture. Micrographs were made on Kodakprojector slide plates (contrast grade) and enlargedphotographically.
RESULTSThe bacterial shape most commonly observed
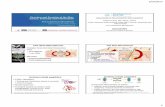
was squatly ovoid (Fig. 1), but elongate formsincluding cigar-shaped and club-shaped individu-als were also frequent (Fig. 2). Viewed in crosssection, bacteria were round or oval. Dimensionsvaried from 0.55 by 0.40 Am in the shorter formsto 1.2 by 0.50 Am in the longest organisms.An outstanding characteristic of the ultra-
structural appearance of C. ovis was the existenceof an electron-dense layer, external to the cellwall, surrounding each organism. The layer con-
IFIG. 1. Cell (in early division) of the virulent C. ovis strain 137C/2,from a 24-hr culture on tellurite-blood-agar.
A thick, electron-dense layer of lipid (arrows) surrounds the cell wall. Denser areas of cytoplasm border the nu-clear apparatus. T indicates crystals of tellurite reduction product. X 74,500.
FIG. 2. Dividing cell of the attenuated strain 137E cultured in viande-foie broth.for 48 hr. An external lipid layeris present (arrows) but is not as prominent as that of the virulent organism. X 74,000.
1481VOL. 97, 1969
.A.-'75 P 4w, -I.:. on February 15, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

J. BACTERIOL.
sisted of floccular material, usuallythe cell wall itself by a narrowabout 4 to 8 nm. There was considein the thickness of this layer witorganisms, but particularly betweezone was widest and most dense istrain 137C/2, ranging from 7 to 3approximate mean of 18 nm (Fiattenuated strains, the range of wi25 nm (approximate mean widtl137E (Fig. 2) and 3 to 18 nm (apprwidth, 9 nm) in 133A. Adjacent oiaggregated together by the envelopof this layer, which was common Iwithin the clump. In colonies ofwashed in ligroin for 5 hr, the exterrin a great majority of organisms wdifficult to see or had disappear(Fig. 3). In bacteria still retaining relayer, its density and thickness wdiminished. Negatively stained prthe pure cell wall samples, froipresumably surface lipid had beenacetone and methanol prior to 4demonstrated the existence of .etched pattern over the entire e
11.1_
FIG. 3. Strain 137C/2 cultured for iagar and subsequently washed for S hr ifaint remnants (arrows) of the surjaceevident. X 37,000.
separated from voluted and intertwining bands of low electronclear space of density (about 5 nm wide) separated by similarrable variation bands of higher density made up the overall:hin individual "crazed" surface pattern (Fig. 4a). Cell walls pre-n strains. The pared from fresh, unextracted organisms werein the virulent clumped together by an enveloping material37 nm with an which had the appearance of lipid and whichig. 1). In the obliterated the surface characteristics of the walldth was 2.5 to itself (Fig. 4b).h, 10 nm) in The detailed structure of cell wall, plasmaroximate mean membrane, polyphosphate granules, intracyto-rganisms were plasmic membrane systems, and nuclear appara-)ing contiguity tus were identical with those described previouslyto all bacteria for C. minutissimum (13, 14) and C. diphtheriaestrain 137C/2 (9), including the appearance of an irregular,nal dense layer dense, granular perinucleoid band of cytoplasm,vas either very as noted by Kawata (9) in the latter species. Thered altogether basic pattern of the mesosome consisted of twoImnants of the closely apposed unit membranes representing anvere markedly invagination of the complete tripartite structurereparations of of the plasmalemma; thus, it conformed in detailm which the to the diagrammatic representation of these or-extracted with ganelles provided by Pate and Ordal (16) in theirdisintegration, ultrastructural study of Chondrococcus columnarisan intricately (Fig. 5). The elaborate structure of membranexterior. Con- systems and their common association with in-
vaginating septa of cells in division were consist-ent with previous descriptions of a number ofgram-positive bacteria (7).
Cell division normaly occurred across thecenter of the bacterial cell, but in a small propor-tion of dividing bacteria a second or third in-vaginating septum was observed. The line ofdivision was not always in one plane but oftenassumed a haphazard pattern. Figure 6 shows amore extreme example with multiple septa. It
.........appeared that as division occurred, daughter celsadhered together by virtue of the cohesive proper-ties of the surface material. Thus, large numbersof organisms of the strain 137C/2 were often in-
'4 volved in the formation of individual clumps,whereas aggregations of 137E contained fewercels, and only 2 to 4 neighboring bacteria ofstrain 133A were usualy clumped together with-in a common surface envelope.
In bacteria cultured on blood agar containingsodium tellurite, smal black crystals appeared inthe cytoplasm subjacent to the plasma mem-brane (Fig. 1). The demonstration of large crystalsoccupying a major proportion of the cytoplasmof apparently unaffected organisms after culturefor 3 days substantiated Winkler's comment on
mW the ability of Corynebacterium to store and toler-ate a considerable quantity of tellurite (24).
24 hr on blood- Apart from the details pertaining to the surfacein ligroin. Only lipid layer, no other structural differences were ob-lipid layer are served between the virulent and attenuated strains
of C. ovis. The only morphological changes
e
1482 HARD
.. !f!
on February 15, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

C. OVIS FINE STRUCTURE
4a - r E4a .i
01
'I ..................I....11FIG. 4. Negatively stained cell wall preparations. (a) Cell wall of virulent C. ovis prepared from organisms
which had been subjected to extraction with methaanol chloroform. The spheroplast presents an intricate suirfacepattern from which the lipid coating is absent. Only clumps of cytoplasmic debris are adherenit (arrow). X 58,000.(b) In contrast, cell walls preparedfrom unextracted virulent organisms are covered with a lipidlike material whichobscures the surface patteriz and is responsible for the clumping together of the spheroplasts. X 58,000.
*: ::r.16. t .. ,s, oSwilES .. ::4b
=&.a. ..wrsa _
X: :S w:
I;
FIG. 5. Bacterium of strain 137C/2 containing a mesosome wlhich arises as an invagination of the tripartiteplasma membrane. X 156,000.
VOL. 97, 1969
.5 [l I
1483
4ba :.
on February 15, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

HARD
.51S
FIG. 6. Bacterium of strain 137C/2 cultured for 24hr on tellutrite-blood-agar showing multiple planes ofdivision. X 65,500.
noted with an increasing period of culturewere an increase in polyphosphate granule con-tent and a decrease in mesosomes.
DISCUSSIONThe outstanding feature of this study was the
observation of an electron-dense layer aroundeach organism, which was responsible for theadherence of many bacteria into clumps. Thatthis layer represented the surface lipid, the exist-ence of which had been postulated on the basisof extraction studies (2, 4), was verified by itspartial or total removal by ligroin or acetone andmethanol treatment. Lacave et al. (11) recentlycharacterized the ligroin extract of C. ovis andfound that the bulk consisted mainly of sodiumcorynomycolate together with sodium salts ofC14, C16, and C18 fatty acids. Some triglyceridesand free fatty acids were also detected. Jolly (8)has shown that quantitative differences exist be-tween the surface lipids of virulent and attenu-ated strains, which supports the hypothesis thatthey are associated with pathogenicity and viru-lence. He found strain 137E to contain 28% lessextractable lipid than 137C/2, although there wasno strain difference in the toxic quality of thematerial. Morphological substantiation of Jolly's
work was demonstrated by the quantitative differ-ence in thickness and appearance of the surfacelayer in the three strains, with a decreasing se-quence of 137C/2 through 137E to 133A.A further manifestation of the variation in sur-
face lipid was observed as a greater clumpingeffect in 137C/2, involving more organisms ineach aggregate than were present in the smallerclumps of the attenuated strains. Such an ob-servation explains some of the differences insurface properties of the strains. For example, themore hydrophilic growth of the attenuated varie-ties on solid media, in comparison with the waxyintegrity of virulent colonies, is doubtless a reflec-tion of the quantitative difference in surface lipidand consequent clump-size. The same observa-tions have application to the variation betweenstrains in ability to grow as a pellicle in someliquid media. The fact that light extraction doesnot impair the viability of the organisms (4)further suggests that only the surface layer is re-moved. The observation of this material also pro-vides a basis for the hypothesis that the bacteriummay possess a barrier which protects it from post-phagocytic degradation by the host cell's lysoso-mal enzyme complement.The existence of material in some form or
other, external to the cell wall, has been recordedin most other taxonomically related bacteria sofar studied. Material of unknown composition atthe periphery of the cell wall of L. monocytogeneshas been reported (5, 6). Glauert (6) described a"thick capsular layer" composed of an extensivenetwork of fibrous material with little organizedstructure around the cell wall of C. fascians. Amoderate amount of floccular material was foundadherent to the external surface of C. diphtheriae(17), whereas extremely electron-dense materialsurrounded C. minutissimum and was usuallyclosely adherent to the cell wall (13, 14). Stanleyand Rose (21) observed that C. xerosis exhibiteda clumping capacity when it was grown at 30 Cand promptly cooled to 15 C with rapid stirring.Electron microscopy showed that electron-densematerial distributed over the bacterial surfacewas responsible for adherence of these organismsinto clumps. Further study indicated that thematerial responsible for the clumping of C. xerosiswas a cell-surface protein. Different types of cell-surface component have been associated with themanifestation of clumping in other microorgan-isms. Such surface material includes carbohydratein Escherichia coli (20), protein in the yeastHansenula wingei (22), and a hyaluronidase-sensitive polysaccharide in Pseudomonas aerugi-nosa (23). The demonstration of a lipid compon-ent responsible for the surface properties of C. ovis
1484 J. BAcrERIoL.
on February 15, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

C. OVIS FINE STRUCIURE
adds a further example to the list and contraststhis organism with C. xerosis. In contrast to mostrelated bacteria which possess a morphologicallydemonstrable surface coat, the external lipid layerof C. ovis was not usually adherent to the cellwall; in fact, it appeared to be separated fromthat structure by a clear space. That this zone ofseparation might have constituted an artifactualchange cannot be discounted, but it is note-worthy that not only was this characteristic aconsistent finding in most bacteria examined butalso its presence was a feature of viable intra-cellular organisms some hours after ingestion bymouse phagocytes (unpublished data).The frequently irregular disposition of the
dividing septa of C. ovis, together with the surfaceproperties of the external lipid layer, are doubtlessresponsible for the palisade and Chinese-lettergroupings which characterize these organisms insmears and histological sections. Similar obliqueand wavy septa have been described in C. diph-theriae (17), and they most probably constitute acorynebacterial characteristic.
ACKNOWLEDGMENTS
This investigation was supported by a grant from the Aus-tralian Wool Board.
I thank D. G. Drummond for the use of an electron micro-scope and, in particular, R. G. Wright for his excellent technicalassistance. I also acknowledge the encouragement of H. R. Carneand C. H. Gallagher throughout this study.
LITERATURE CITED
1. Brenner, S., and R. W. Home. 1959. A negative stainingmethod for high resolution electron microscopy ofviruses. Biochim. Biophys. Acta 34:103-110.
2. Bull, L. B., and C. G. Dickinson. 1935. Studies on infectionby and resistance to the Preisz-Nocard bacillus. IV. Noteson the toxin, the pyogenic action, and the lipase contentof the bacillus. Australian Vet. J. 11:126-138.
3. Carne, H. R. 1939. A bacteriological study of 134 strains ofCorynebacterium ovis. J. Pathol. Bacteriol. 49:313-328.
4. Carne, H. R., N. Wickham, and J. C. Kater. 1956. A toxiclipid from the surface of Corynebacterium ovis. Nature178:701-702.
5. Edwards, M. R., and R. W. Stevens. 1963. Fine structureof Listeria monocytogenes. J. Bacteriol. 86:414-428.
6. Glauert, A. M. 1962. The fine structure of bacteria. Brit.Med. Bull. 18:245-250.
7. Iterson, W. van. 1965. Symposium on the fine structure andreplication of bacteria and their parts. II. Bacterial cyto-plasm. Bacteriol. Rev. 29:299-325.
8. Jolly, R. D. 1966. Some observations on surface lipids ofvirulent and attenuated strains of Corynebacterium ovis.J. Appl. Bacteriol. 29:189-196.
9. Kawata, T. 1961. Electron microscopy of the fine structureof Corynebacterium diphtheriae with special reference tothe intracytoplasmnic membrane system. Japan. J. Micro-biol. 5:441-451.
10. Kellenberger, E., A. Ryter, and J. Sechaud. 1958. Electronmicroscope study of DNA-containing plasms. II. Vege-tative and mature phage DNA as compared with normalbacterial nucleoids in different physiological states. J.Biophys. Biochem. Cytol. 4:671-678.
11. Lacave, C., J. Asselineau, and R. Toubiana. 1967. Sur quelqueconstituants lipidiques de Corynebacterium ovis. EuropeanJ. Biochem. 2:37-43.
12. Luft, J. H. 1956. Permanganate-a new fixative for electronmicroscopy. J. Biophys. Biochem. Cytol. 2:799-802.
13. Montes, L. F., and S. H. Black. 1967. The fine structure ofdiphtheroids of erythrasma. J. Invest. Dermatol. 48:342-351.
14. Montes, L. F., and S. H. Black. 1967. The fine structure ofCorynebacterium minutissimum. Experientia 23:345-346.
15. North, R. J. 1963. Some structural aspects of Listeria mono-cytogenes. J. Ultrastruct. Res. 9:187-197.
16. Pate, J. L., and E. J. Ordal. 1967. The fine structure of Chon-drococcus columnaris. I. Structure and formation of meso-somes. J. Cell Biol. 35:1-13.
17. Reyn, A., A. Birch-Andersen, and S. P. Lapage. 1966. Anelectron microscope study of thin sections of Haemophilusvaginalis (Gardner and Dukes) and some possibly relatedspecies. Can. J. Microbiol. 12:1125-1136.
18. Reynolds, E. S. 1963. The use of lead citrate at high pH asan electron-opaque stain in electron microscopy. J. CellBiol. 17:208-212.
19. Salton, M. R. J., and R. W. Horne. 1951. Studies of thebacterial cell wall. II. Methods of preparation and someproperties of cell walls. Biochim. Biophys. Acta 7:177-197.
20. Sneath, P. H. A., and J. Lederberg. 1961. Inhibition by perio-date of mating in Escherichia coli K-12. Proc. Natl. Acad.Sci. U.S. 47:86-90.
21. Stanley, S. O., and A. H. Rose. 1967. On the clumping ofCorynebacterium xerosis as affected by temperature. J.Gen. Microbiol. 48:9-23.
22. Taylor, N. W. 1964. Specific, soluble factor involved insexual agglutination of the yeast Hansenula winigei. J.Bacteriol. 87:863-866.
23. Warren, G. H., and J. Gray. 1955. Studies on the propertiesof a polysaccharide constituent produced by Pseudomonasaeruginosa. J. Bacteriol. 70:152-157.
24. Winkler, A. 1958. Feinstruktur des Bakterienzellinhaltes.Zentr. Bakteriol. Parasitenk. Abt. I Orig. 173:369-375.
VOL. 97, 1969 1485
on February 15, 2020 by guest
http://jb.asm.org/
Dow
nloaded from