
El ideotipo del girasol (Helianthus annuus L.)€¦ · El ideotipo del girasol (Heilanthus annuus L...
12
AGRISCIENTIA , 1994, VOL. XI : 87-98 REVISIÓN El ideotipo del girasol (Helianthus annuus L.) Hernández , L.F. y G.A. Orioli RESUMEN Se analizan las características morfológicas y fisiológicas que son importantes para definir el potencial del rendimiento de la planta de girasol En base a ello se propone un tipo de planta ideal (ideotipo), la cual sea capaz de aprovechar con máxima eficiencia la disponibilidad de recursos provistos con las actuales prác- ticas agronómicas. Se realiza una descripción del desarrollo del cultivo, ponien- do énfasis en los diferentes factores que pueden afectar cada uno de los esta- dos del desarrollo Dentro de las características morfológicas, se propone que las plantas posean un sistema radical extenso y profundo, tallos de porte mediano con alta capaci- dad de reserva de fotoasimilados, los cuales puedan ser redistribuidos al grano durante el período de su llenado Hojas con un ritmo de senescencia lento y ca- paces de alcanzar un IAF no menor de 4 en corto tiempo Dentro de las caracte- rísticas asociadas con la formación del rendimiento, se propone que la superf i- cie meristemática del receptáculo sea máxima previo al inicio de la diferenciación de los primordios florales, y que el capítulo maduro posea un bajo ritmo respira- torio. Se deben lograr capítulos con frutos más grandes y más pesados y con ma- yor contenido porcentual de aceite. Dentro de la respuesta a practicas agronómicas, el cultivo deberá ser capaz de producir mejor bajo nuevas técnicas de manejo tales como "intercropping" o la- branza "cero", tener una mayor resistencia a plagas y enfermedades y respon- der mejor a la fertilización, tanto fosforada como nitrogenada. Palabras clave : girasol, Helianthus annuus, ideotipo, rendimiento Hernández, L.F. and G.A. Orioli, 1994. The ideotype in sunflower (Helianthus annuus L ). Agriscientia XI : 87-98. SUMMARY Morphological and physiological characteristics that are important to define the potential of yield of the sunflower plant are described Then an ideal type of plant (ideotype) which should be able to efficiently use the available resources in the field is proposed A description of crop development emphasizing the factors that can affect different stages of its development is made Among the morphological characteristics, it is proposed that plants should have an extended and deep root system and median height shoots with high capacity to store photoassimilates These could then be reallocated to the capitulum during the grain filing period Leaves should have a low senescence rate and the canopy should reach a LAI not lower than 4 in a short time It also proposed that the meristematic surf ace of the receptacle has to reach its maximum size before the floret primordia diff eren- Fecha de recepción: 28/9/94.
Transcript of El ideotipo del girasol (Helianthus annuus L.)€¦ · El ideotipo del girasol (Heilanthus annuus L...

AGRISCIENTIA , 1994, VOL. XI : 87-98
REVISIÓN
El ideotipo del girasol(Helianthus annuus L.)Hernández , L.F. y G.A. Orioli
RESUMEN
Se analizan las características morfológicas y fisiológicas que son importantespara definir el potencial del rendimiento de la planta de girasol En base a ello sepropone un tipo de planta ideal (ideotipo), la cual sea capaz de aprovechar conmáxima eficiencia la disponibilidad de recursos provistos con las actuales prác-ticas agronómicas. Se realiza una descripción del desarrollo del cultivo, ponien-do énfasis en los diferentes factores que pueden afectar cada uno de los esta-dos del desarrolloDentro de las características morfológicas, se propone que las plantas poseanun sistema radical extenso y profundo, tallos de porte mediano con alta capaci-dad de reserva de fotoasimilados, los cuales puedan ser redistribuidos al granodurante el período de su llenado Hojas con un ritmo de senescencia lento y ca-paces de alcanzar un IAF no menor de 4 en corto tiempo Dentro de las caracte-rísticas asociadas con la formación del rendimiento, se propone que la superf i-cie meristemática del receptáculo sea máxima previo al inicio de la diferenciaciónde los primordios florales, y que el capítulo maduro posea un bajo ritmo respira-torio. Se deben lograr capítulos con frutos más grandes y más pesados y con ma-yor contenido porcentual de aceite.Dentro de la respuesta a practicas agronómicas, el cultivo deberá ser capaz deproducir mejor bajo nuevas técnicas de manejo tales como "intercropping" o la-branza "cero", tener una mayor resistencia a plagas y enfermedades y respon-der mejor a la fertilización, tanto fosforada como nitrogenada.
Palabras clave : girasol, Helianthus annuus, ideotipo, rendimiento
Hernández, L.F. and G.A. Orioli, 1994. The ideotype in sunflower (Helianthusannuus L ). Agriscientia XI : 87-98.
SUMMARY
Morphological and physiological characteristics that are important to define thepotential of yield of the sunflower plant are described Then an ideal type of plant(ideotype) which should be able to efficiently use the available resources in thefield is proposed A description of crop development emphasizing the factors thatcan affect different stages of its development is made Among the morphologicalcharacteristics, it is proposed that plants should have an extended and deep rootsystem and median height shoots with high capacity to store photoassimilatesThese could then be reallocated to the capitulum during the grain filing periodLeaves should have a low senescence rate and the canopy should reach a LAInot lower than 4 in a short time It also proposed that the meristematic surf ace ofthe receptacle has to reach its maximum size before the floret primordia diff eren-
Fecha de recepción: 28/9/94.

88 AGRISCIENTIA
tiation begins. The mature capitula should have a low respiratory rate. It also couldbe desirable to get capitula with larger and heavier fruits and with a high oil con-tent.The crop also should be able to respond efficiently to new agronomic practicessuch as "intercropping" or "zero tillage". It should also be more resistant to pla-gues and pests and have a better response to fertilization.
Key words : sunflower, Helianthus annuus, ideotype, yield.
L.F. Hernández y G.A. Orioli. Departamento de Agronomía . UniversidadNacional del Sur. 8000. Bahía Blanca. Argentina.
INTRODUCCIÓNEl desarrollo de nuevos cultivares de plantas de
cultivo es un proceso continuo por el que se buscaaumentar su rendimiento o mejorar la calidad de suproducto ( Evans y Wardiaw , 1976). Estos dos obje-tivos pueden lograrse de diversas maneras, muchasde las cuales han llevado a producir cambios muymarcados en la planta e indirectamente , cambiossutiles o ajustes en sus procesos metabólicos(Bradshaw , A., 1965 ; Bingham, J., 1967 ; Donald yHamblin, 1976; Blocklehurst , P , 1977, Beard yGeng , 1982 ; Connor y Sadras, 1992).
En los últimos años , se ha comenzado a tener encuenta la selección directa de importantes atributosfisiológicos (fotosíntesis , respiración , transporte deminerales y fotoasimilados , etc.) y morfológicos (ro-bustez y longitud del tallo, inclinación del capítulo ala madurez , área foliar , etc.). Sin embargo no se pue-de asegurar que tal tipo de selección será más efec-tiva o más rápida que la selección por productividad.
El curso que siguió la domesticación de los prin-cipales cultivos es bien conocido . Las especies fue-ron sujetas a una selección directa para lograr au-mentos en su producción bajo sistemas agronómicosprogresivamente más eficaces (Bradshaw A., 1965,Donald y Hamblin , 1976). Los cambios fisiológicosque acompañaron y contribuyeron a esa domestica-ción, y quo dioron oligon 11 ouillunto polunclal dulrendimiento , son sin embargo prácticamente desco-nocidos . Conocer cuáles han sido esos cambios, per-mitiría tener una idea sobre qué vías están exhaustasy cuáles pueden ser explotadas para lograr mayoresrendimientos y más estabilidad de los mismos.
En este trabajo se describen algunas caracterís-ticas morfológicas y fisiológicas de la planta de gi-rasol que tienen particular incidencia en la maximi-zación de su rendimiento . El objetivo es proponerque las mismas puedan ser consideradas en planesde mejoramiento genético del cultivo.
Descripción de caracteresposibles de ser mejorados
1. Desarrollo vegetativo y reproductivo
En las plantas silvestres el ciclo de vida está con-trolado por el fotoperiodismo y la vernalización. Res-pondiendo a estos estímulos las plantas se adapta-ron a su medio ambiente. Sin embargo durante sudomesticación y adaptación a nuevas condicionesde crecimiento estos mecanismos de regulación sehan modificado. Es importante entonces conocer lalongitud del ciclo de vida de las especies cultivadascomo así también la longitud relativa de cada unade las cuatro principales etapas de su desarrollo.
En el girasol, la primera etapa del ciclo de vidacomienza con la germinación e Implantación delcultivo. Es importante que la misma se produzca enuna forma rápida y uniforme para lograr un stand pa-rejo de plantas (Anfinrud y Schneiter, 1984, Cardi-nal¡ et al., 1985). Aún cuando en condiciones con-troladas la germinación y emergencia no sonafectadas por el tamaño de los frutos, en condicio-nes de campo sí lo son. Una de las causas puedeser que la velocidad de imbibición de las semillases menor en frutos relativamente más grandes de-bido a una mayor separación entre el pericarpio ylos tegumentos de esta última (Hernández y Oriol¡,1985a). A campo las diferencias observadas, luegode la emergencia, en el número do plantas y su tomaño pueden ser atribuidas a condiciones de siem-bra, humedad y contacto suelo-semilla deficientes,capacidad del hipocotilo para elongarse hasta la su-perficie del suelo, ataque de plagas, disponibilidadde nutrientes y capacidad para absorberlos, etc. Lasdiferencias en el tamaño de las plántulas son impor-tantes. En condiciones de hidroponía las diferenciasentre plántulas provenientes de semillas chicas ygrandes tienden a desaparecer hacia la tercer se-mana de crecimiento mientras que en condicionesde campo las diferencias se mantienen, incidiendoen el rendimiento (Aguirrezábal et al., 1985).

El ideotipo del girasol (Heilanthus annuus L )
300
É 250s
La segunda etapa, la vegetativa o juvenil, se ca-racteriza por el desarrollo de las raíces, tallos y ho-jas. Durante este período influyen la temperatura y lalongitud del día y, en cierto modo, esta fase determi-na la respuesta de la fase siguiente. Los actuales cul-tivares de girasol pueden considerarse como neu-tros en cuanto a los requerimientos de frío yfotoperíodo; de todos modos algunos muestran unaleve tendencia a ser de día corto y se han encontra-do diferencias en el reuno reproductivo con cambiosel ciclo de crecimiento (precoces y tardíos) y con di-ferente fecha de siembra (Marc y Palmer, 1981, Goy-ne y Schneiter, 1987). Esta etapa en el girasol pue-de considerarse corta ya que solamente dura entre20 y 22 días desde la emergencia (DDE) de las plán-tulas. Intervalos de tiempo mayores son posibles, de-pendiendo de las condiciones ambientales y varíe-tales. Durante este período, se desarrollan por lomenos 18 hojas antes de pasar a la siguiente fase(Marc y Palmer, 1981). Se debe destacar que en elgirasol el ritmo dei desarrollo vegetativo durante es-ta etapa es muy bajo por lo que a las densidades co-munes de siembra, (4,5 a 5,5 plantas.m-2), el follajeno alcanza a cubrir totalmente la superficie del sue-lo (Hernández, L., 1983; Hernández y Orioli, 1982;1983; 1992). Esto se traduce en una deficiente inter-cepción de la radiación solar y menor capacidad decompetir con malezas (Pereyra et al., 1977).
Durante la tercera etapa, la reproductiva, quecomprende el intervalo entre la inducción e incia-ción floral hasta la antesis y fertilización (entre los22 y 58 DDE), no solamente se produce el desarro-llo de la inflorescencia sino que se observa un grandesarrollo en altura y en la biomasa foliar y del tallo,que prácticamente cesan cuando comienza la ante-sis (Hernández y Oriol¡, 1982; Hernández, L., 1983).Esta tercera etapa es de suma importancia para de-terminar los componentes del rendimiento: duranteésta no solo se forma el máximo potencial de alma-cenaje de carbohidratos y minerales de la planta si-no que se forma también la inflorescencia. Ambosprocesos dependen de la disponibilidad, en ese mo-mento, de fotoasimilados y nutrientes minerales. Sepuede entonces presentar una competencia entrediferentes destinos: una gran masa vegetativa en ac-tivo crecimiento y un órgano reproductivo (capítulo)en desarrollo. La primera actuará sobre el compo-nente del rendimiento peso de los frutos y el segun-do influenciará fundamentalmente el componente delrendimiento número de frutos. Este período en gira-sol es una etapa altamente dependiente del cultivar(Anderson, W., 1975), de la temperatura (Chapmanet al., 1993), de la capacidad de intercepción e in-tensidad de la radiación (Hernández, L., 1983), de ladisponibilidad de nutrientes entre los cuales el nitró-
ro 200
^ 150^^ 100
d-50
q ^ n
•^
qó
i ii ^
r ^,
^% n ^/^
^/
89
1600
1400
1200
1000
800
600
40020 25 30 35 40 45 50
Días desde la emergencia
^
f0ó
^̂a,^
Z
Figura 1. Evolución del área del receptáculo en su etapa de inci-piente formación y del número de primordios florales en función deltiempo de dos genotipos de girasol, (p) Dekalb G90 y (p) DekalbG100.
geno, el fósforo y el boro juegan un rol importante(Blarney et al., 1979; Hocking y Steer, 1983, Steer yHocking, 1983). La diferenciación de las flores en elcapítulo se produce desde el borde hacia el centrodel receptáculo cubriendo totalmente la superficie li-bre del mismo. El tamaño del ápice reproductivo au-menta unas 100 veces y se diferencian entre 600 y2000 flores (Hernández, L., 1988). En la Fig. 1 se pue-de apreciar la influencia del cultivar
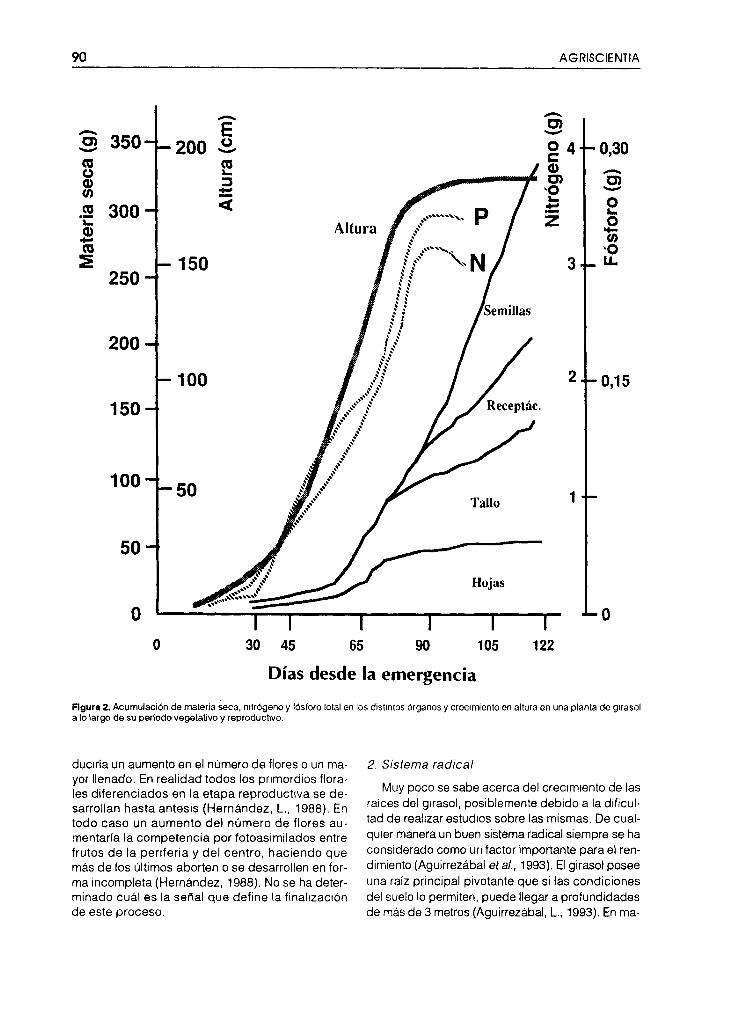
La cuarta etapa es la de llenado de los frutos yposterior maduración . Durante ella no ocurre o haymuy poco crecimiento vegetativo de los demás ór-ganos de la planta (Fig. 2). Dependiendo de la den-sidad de plantas en el cultivo, a comienzos de esteestado puede todavía seguir aumentando el índicede área follar (IAF) por el continuo desarrollo de lashojas superiores (Fig. 3) Existen evidencias que in-dican que el llenado dei grano se produce principal-mente con fotoasimilados actuales pero también conla contribución de lo acumulado durante el períodode preantesis, lo cual puede representar hasta el 20-30 % del total del carbono almacenado en los frutosal momento de cosecha (Hall et al., 1989; 1990). Teó-ricamente convendría no solamente que esa fasefuera más larga sino que durante la misma se con-servara la máxima superficie foliar activa.
Tanto el desarrollo de la inflorescencia como laantesis, fecundación y llenado de los frutos sonprocesos que en el girasol ocurren siempre des-de el borde hacia el centro del capítulo. Se podríasuponer entonces que encontrando condicionesque prolonguen la duración de los mismos se pro-
90 AGRISCIENTIA
rn 350caQa)N
S! 300a)
250
E200 ^
11
200 -
_ 1 (
150 -1-
O
ú)
100 -1
150 -
100
-50-
c4dCn.ó
in-adli
34
- 0,30
(IIIo1.óV)
lo_ U.
2 - 0,15
1
IT- 00 30 45 65 90 105 122
Días desde la emergencia
Figura 2. Acumulación de materia seca, nitrógeno y fósforo total en los distintos órganos y crecimiento en altura en una planta de girasola lo largo de su período vegetativo y reproductivo,
duclría un aumento en el número de flores o un ma-yor llenado. En realidad todos los primordios flora-les diferenciados en la etapa reproductiva se de-sarrollan hasta antesis (Hernández, L., 1988). Entodo caso un aumento del número de flores au-mentaría la competencia por fotoasimilados entrefrutos de la periferia y del centro, haciendo quemás de los últimos aborten o se desarrollen en for-ma incompleta (Hernández, 1988). No se ha deter-minado cuál es la señal que define la finalizaciónde este proceso.
2. Sistema radica!
Muy poco se sabe acerca del crecimiento de lasraíces del girasol, posiblemente debido a la dificul-tad de realizar estudios sobre las mismas. De cual-quier manera un buen sistema radical siempre se haconsiderado como uri factor importante para el ren-dimiento (Aguirrezábal eta!., 1993). El girasol poseeuna raíz principal pivotante que si las condicionesdel suelo lo permiten, puede llegar a profundidadesde más de 3 metros (Aguirrezábal, L., 1993). En ma-

El ideofipo del girasol (Helianthus annuus L )
EBF PA UA6.0
1 it .
5.0i^ jOOO-iO o^ ° s' or
3.0^ °r ^
2.0^
,1.0 ,
J o
0 e-
10 30 50 70 90 110
6.0 o i ,
U-Q
Días desde la emergencia
Figura 3. Evolución del índice de área foliar (IAF) durante el ci-clo de crecimiento de un cultivo de girasol creciendo a dos den-sidades: (0) 5,6 plantas .m-s; (O) 16,6 plantas.m-s. Obsérveseque a menor densidad la antesis ocurre antes de obtener el má-ximo IAF. EBF: emergencia del botón floral; PA: primera antesis;UA: última antesis.
cetas se ha observado que entre la segunda y terce-ra semana desde la emergencia, la raíz principal lle-ga a los 80 cm de profundidad. Sin embargo la elon-gación radical es muy sensible a condiciones delsuelo que opongan resistencia o que sean inhibido-res biológicos. Así el piso de arado, la compactaciónpor tránsito vehicular, la salinidad, producen un efec-to negativo muy marcado en el desarrollo radical ypor lo tanto en el rendimiento. La disminución del de-sarrollo de raíces en suelos y la consecuente reduc-ción del volumen de suelo que las raíces pueden ex-plorar, y por lo tanto del agua y nutrientesdisponibles, será la causa de la disminución en elrendimiento. En la parte superior del sistema radicalse desarrolla una gran cantidad de raíces de distin-ta categoría las que se extienden lateralmente y al-gunas se inclinan hacia abajo hasta llegar a los 60-70 cm de profundidad. El buen desarrollo de labiomasa radical, que en un cultivo puede alcanzar 1t.ha-1, está en relación con el desarrollo de la parteaérea que suministra los recursos energéticos y conuna buena disponibilidad de agua y nutrientes, prin-cipalmente nitrógeno, antes de la iniciación floral(Aguirrezábal et al., 1994). Sin embargo la relaciónparte aérea/sistema radical y el intercambio de men-sajes entre ambos aún está bajo estudio.
Algunas características muy importantes de iden-tificar entre cultivares serian no solamente la dístri-bución de la densidad de raíces sino también la du-
91
ración, velocidad y profundidad de extensión de lasmismas. Es probable que la absorción de nutrientesy agua estén más relacionados con la rapidez deexploración de nuevas superficies de suelo que conel área de la superficie explorada. Además su resis-tencia al flujo de agua (Black, C., 1979; Hernándezy Orioli, 1985d) y nutrientes es también importanteya que se ha observado que es una característicarelacionada directamente con el uso del agua por laplanta de girasol y que varia entre cultivares (Her-nández y Oriol¡, 1985d; Koide, R., 1985).
3. Uso de nutrientes
Los resultados de estudios de fertilización reali-zados en el país y en el mundo son demasiado con-trovertidos por lo que es necesario hacer más tra-bajos sobre este tema. De todos modos se conoceque las curvas de absorción de algunos nutrientesen el girasol, como por ejemplo nitrógeno y fósforo,son similares a la ganancia de altura y peso seco delos diferentes órganos de la planta (Fig. 2) (Oriol¡ etal., 1977). Consecuentemente la concentración deestos nutrientes por unidad de biomasa disminuyeen forma inversamente proporcional al incrementodel peso total de la misma, por lo que si bien duran-te la fase vegetativa hay poca absorción, los nutrlen-tes están presentes en alta concentración. Posterior-mente, durante la elongación del tallo hay una granacumulación, la que cesa cuando ocurre la floración(Fig. 2). Por lo tanto en los frutos se acumularán loselementos minerales móviles, y cuya fuente princi-pal es la cantidad acumulada durante las etapasprevias al llenado (Hocking y Steer, 1983). Es posi-ble que la respuesta a la fertilización no solo esté li-gada al cultivar y a las propiedades físico-químicasdel suelo (Hernández y Oriol¡, 1985b), sino fuerte-mente influenciada por determinadas característi-cas del desarrollo radical. Así el crecimiento en pro-fundidad de las raíces laterales explicaría larespuesta positiva a la fertilización profunda
4. Uso del agua
El girasol tiene un elevado ritmo de transpiración,por lo que su eficiencia en el uso de agua no es muyalta (Rawson, H., 1979; Rawson y Constable, 1980,Hernández y Oriol¡, 1985d). También posee mecanis-mos de regulación osmótica a nivel foliar que en algu-nos casos son importantes para tolerar condicionesde estrés hídrico no muy severos y transitorios (Con-nor y Sadras, 1992). La capacidad de la planta de nocerrar los estomas sino frente a condiciones de estréshídrico realmente severos, hace que en un amplio ran-go de condiciones la transpiración sea principalmen-te controlada por la diferencia de humedad entre lashojas y la atmósfera, y no por el cierre de los estomas(Hernández y Oriol¡, 1985d; Chapman et al., 1993).Como consecuencia de esa característica, bajo con-

92
diciones de estrés hídrico mediano, que modifican lamorfología del follaje (Shell y Lang, 1976; Hernándezy Oriol¡, 1983), la planta puede aún fotosintetizar portener los estomas abiertos. Es posible que la alta den-sidad de pelos en las hojas de girasol sea también unfactor importante para regular la transpiración y por lotanto influya sobre la economía del agua del cultivo(Hernández y Oriol¡, 1985c). A este factor se deberíaagregar el control que las raíces ejercerían sobre elflujo del agua. (ver 2 ) (Hernández y Orioli, 1985d).
5 Indice de cosecha
El rendimiento de un cultivo dependerá de laefectividad de la fotosíntesis en producir la mayorcantidad de materia seca y de la correcta distribu-ción de los fotoasimilados en el momento oportuno(Giff ord y Evans, 1981). En 1956 se introdujeron dostérminos en relación al rendimiento de un cultivo(Evans, L., 1975):
a) Rendimiento Biológico = Rb: el total de mate-ria seca (MS1 producida por las plantas.
b) Rendimiento Económico = Re: el peso secode la parte útil cosechable.
Estos dos parámetros se expresan por otro ori-ginalmente denominado Coeficiente de Efectivi-dad de Formación de la Parte Económica y quees más conocido como índice de Cosecha (IC). Porlo tanto:
IC = Re/Rb , generalmente expresado como por-centaje.
El verdadero Rb debería incluir el peso de raícespero la dificultad en extraerlas hace que en generalno se tenga en cuenta.
Hasta que se pueda analizar la producción en tér-minos fisiológicos y bioquímicos, el IC puede ser uncriterio valioso para la evaluación de cultivares degirasol y de la distribución de fotoasimilados. Parael trigo, por ejemplo, el mejoramiento genético hizovariar el IC sustancialmente a lo largo de los añospasando de 30% a 50% con las variedades enanas(Bingham, J., 1967; Bremner y Rawson, 1978, Mar-tínez-Carrasco y Thorpe, 1979)
El IC que el girasol ha alcanzado en nuestras ex-periencias es definitivamente bajo ya que oscila entre25% y 35% según el material utilizado. Para las mis-mas condiciones de cultivo las variedades presenta-ron un IC menor que el de híbridos. Además el IC pue-de variar con cambios en la densidad y condicionesde cultivo. Así para densidades de 2,8; 5,6; 11,0 y 33,3plantas.m-2 los IC fueron 30%, 37%, 35% y 26% res-pectivamente (Hernández, L., 1983). La variación delIC es un fenómeno complejo en el cual juegan un pa-pel preponderante la fotosíntesis, la respiración y larelación fuente-destino (Gaines et al., 1974, Lloyd yCanvin, 1977, Herzog, H., 1982) (ver 7.)
AGRISCIENTIA
6. Relación entre el rendimiento y el área foliar
El mejoramiento de las plantas de cultivo para ob-tener un aumento del rendimiento económico (pesode semillas, frutos, etc.) trajo como consecuenciaun aumento en el tamaño del otros órganos. Por
ejemplo un aumento de 20 veces en el tamaño de la
semilla de trigo fue acompañado por un aumento
considerable del área foliar (Donald y Hamblin,1976; Evans, L., 1976). Si bien no se puede asegu-rar que esta ha sido la tendencia en el girasol, se co-noce que hay una estrecha relación entre los rendi-mientos biológico y económico y el índice de áreafoliar (IAF) (Fig. 4).
Se han realizado numerosos trabajos en donde seha relacionado el rendimiento económico con variosíndices fisiológicos calculados a partir del área foliary del peso seco acumulado (Evans, L., 1975). Así elíndice de crecimiento relativo, índice de asimilaciónneta, duración del área foliar, etc., calculados en dis-tintos períodos de crecimiento, han producido unagran variedad de datos positivos, negativos o no co-rrelacionados. Esta inconsistencia, de las que seaparta el área foliar, se atribuye principalmente al he-cho de que estos índices no toman en cuenta la in-tercepción de la radiación solar y su utilización.
Es por ello que últimamente también se puso én-fasis en una arquitectura de la planta que pueda ga-rantizar una mayor y más eficiente intercepción dela radiación solar. Eso se puede lograr por cambiosen: la ramificación; longitud de los pecíolos; tama-ño, forma, inclinación y disposición espacial de lashojas; número de hojas; duración del área foliar;densidad de siembra; etc.
600
É 500
400•
300
350
300
A250 q.
W.0
200 °CO
150
IAF
Figura 4 . Relación entre el Indice de área foliar (IAF) y el rendimien-to biológico (Rb, p) y económico (Re, •) obtenido utilizando seisgenotipos de girasol crecidos a una densidad de 5,6 plantas.m-2Las ecuaciones de las curvas ajustadas son-
Rb • 102 = 2,82 - 0,93 IAF; Re • 10-2 = -4,54 + 4,12 IAF - 0,43 IAFY

El ideotipo del girasol (Hellanthus annuus L.)
La evolución del desarrollo del área foliar en gi-rasol es de la forma sigmoide con un máximo IAFque se mantiene por varios días y luego comienza adisminuir rápidamente debido al cese de crecimien-to, aceleración de senescencia y muerte de las ho-jas inferiores ( Fig. 3). Los datos existentes indicaríanque si bien el IAF aumenta con la densidad, un va-lor máximo de 4 se obtiene para ese parámetro adensidades de entre 5,0 y 15,0 plantas.m-2. Ademásse observa una diferencia temporal en cuanto al mo-mento en que se logra el máximo IAF por lo que adensidades bajas la antesis ocurre antes de obte-ner el máximo IAF, mientras que a densidades altasla antesis ocurre después de obtenido el máximoIAF (Hernández y Oriol¡, 1982) (Fig. 3). Así entoncesa alta densidad el desarrollo de la inflorescencia yel comienzo del llenado de los frutos ocurren a va-lores de IAF más altos que a baja densidad. Esto po-dría explicar porqué en condiciones experimentalesel rendimiento económico a altas densidades es su-perior al obtenido en cultivos sembrados a densida-des de entre 3,0 y 7, 0 plantas . m-2 (Hernández yOriol¡, 1982 ; 1992).
Es importante recordar que si bien el porcentajedisminución del IAF por senescencia o daño en lashojas en el momento de cosecha es generalmentedel orden del 50% o mayor (Fig. 3), la disminución delIAF comienza la por pérdida de hojas inferiores. Lue-go las hojas superiores siguen en condiciones fisio-lógicas y ambientales aptas para fotosíntesis por unperíodo más largo ( English et al., 1979 ; Pieters, G.,1985; Rawson , H., 1979 ). En girasol esto es muy im-portante ya que se ha demostrado que el tercio su-perior de hojas del canopeo es el que efectivamentecontribuye al llenado de los frutos.
Una manera de aumentar la capacidad de inter-cepción de la radiación solar incidente sobre el cul-tivo, sería modificar el ángulo de inclinación de lashojas con respecto al tallo. Así por ejemplo, si bienhojas más verticales no interceptan tanta radiacióncomo las horizontales, el follaje en su totalidad pre-sentará un bajo coeficiente de extinción a la radia-ción solar . Esto permitiría que la radiación fuera in-terceptada por más hojas debido a una mayorpenetración de radiación hacia los estratos inferio-res de hojas del cultivo. Es decir que en una plantade girasol con hojas más verticales , el llenado losfrutos se verla beneficiado por el aporte de fotoasi-milados provenientes también de hojas que estánmás abajo de las del tercio superior del canopeo(Hernández y Oriol¡, 1991).
La superficie fotos¡ntetizante y la arquitectura delcanopeo en relación a la intercepción de la radia-ción, son dos características principales que deter-minan la fotosíntesis neta del cultivo. El ángulo de in-serción foliar es el más conveniente estará
93
determinado por el balance entre la radiación inter-ceptada por unidad de suelo y la interceptada por lasuperficie foliar contenida en esa unidad . Una pers-pectiva promisoria se puede presentar al determinaralgunas de estas características en cultivares de por-te mediano o bajo y manejando densidades de siem-bra mayores a las ya establecidas como óptimas.
7. Fotosíntesis y respiración
Una de las áreas más atractivas de estudio paraobtener aumentos de rendimiento es la de procurarun aumento en la tasa de fotosíntesis o reducir la ta-sa de respiración y por lo tanto lograr un aumentoen la fotosíntesis neta de las plantas . La tasa de res-piración de la planta de girasol es alta . En el casode la respiración oscura (escotorrrespiración) su rit-mo es entre cuatro y ocho veces superior a la de loscereales (English et al., 1979, Connor y Sadras,1992). Por lo tanto todo aquello que contribuya a dis-minuir el ritmo de escotorrrespiración contribuirá aun aumento del rendimiento.
La capacidad fotosintética de la planta del gira-sol es conocida . Por sus características anatómicasy metabólicas el girasol es una especie de tipo C3y su tasa fotosintética (55-60 mg CO2 dm-2 h-1) esmenor que en las plantas C4 tales como sorgo omaíz (75 mg CO2 dm-2 h-'). Estas diferencias se re-ducen si se comparan estas especies en condicio-nes de baja luminosidad y baja temperatura. Comola temperatura óptima de fotosíntesis se extiendedesde los 25'C hasta los 40'C y la intensidad de sa-turación luminosa es muy alta, luego el girasol esmuy eficiente tanto en condiciones de baja como dealta radiación y temperatura. Las diferencias entrecultivares de la especie son pequeñas.
La comparación entre la energía solar disponiblepara un cultivo y la energía química acumulada pormedio de la fotosíntesis, es una forma de cuantificarla eficiencia con la cual las plantas aprovechan lamisma. En general solamente alrededor del 2% dela energía solar por unidad de superficie de sueloes transformada en energía química. Esa eficienciase eleva al 7% si se corrige para las ondas de luzcorrespondientes a las fotosintéticamente activas(400-700 nm).
En Balcarce se ha realizado este cálculo en un cul-tivo de girasol teniendo en cuenta la energía recibidaen concepto de radiación solar durante los meses desu desarrollo. La producción potencial fue de 73 y de53 g de materia seca (MS) por m-2.día-' para el pri-mero y segundo período de desarrollo ( meses de No-viembre y Diciembre y de Enero y Febrero respecti-vamente) (Oriol¡ et al., 1977). De las pendientes deacumulación de MS diaria se calculó qué porcentajede esa producción potencial correspondería a cadaperíodo resultando ser 2 % y 57%. Para una produc-

94 AGRISCIENTIA
ción de grano de 100 g por planta, la eficiencia deacumulación de reservas respecto a la producciónpotencial fue de alrededor del 5%.
A modo de comparación en Inglaterra se han ob-tenido récords de producción de MS de 20 t.ha-'con un Re de 10 t.ha-' (Hernández, 1983) en un me-dio ambiente con un Re promedio de 4 t.ha-' paratrigo y cebada (Evans, L., 1976). Estos datos pue-den ser favorablemente comparados con resultadosobtenidos en el valle inferior del Río Colorado endonde un cultivo de girasol bajo riego produjo un ré-cord de MS de 27 t.ha-' con un rendimiento en gra-no de 7 t.ha-1. Como se ve, el IC para los cerealesfue de 50% mientras que para el girasol aún en con-diciones de alta producción se mantiene bajo puesfue apenas de 27%.
Como se expresó más arriba, un mediano estréstemporario de agua debido a altas temperaturas yradiación, producirá un cese de crecimiento peroafectará mucho menos al aparato fotosintético quees más tolerante. Si las hojas de la planta no cierransus estomas luego habrá fotosíntesis y el crecimien-to se reanudará si condiciones nocturnas de bajatemperatura y alta humedad relativa aumentan la tur-gencia de las células y reducen el ritmo de escoto-rrespiración. Esto probablemente explique los resul-tados obtenidos en el valle inferior del Río Colorado.
8. Transporte y distribución de fotoasimilados
Así como todavía no hay claridad acerca del o losmecanismos de transporte de fotoasimilados, pocose sabe acerca de los controles del mismo. Cuálesson los factores y cómo afectan el transporte sonproblemas que están bajo estudio. Por ejemplo nose conoce cuál es la señal ni el mensajero que de-termina el tráfico preferencial de fotoasimilados dehojas superiores a zonas aéreas en activo desarro-llo. Ni tampoco porqué en determinadas circunstan-cias un aporte de nitrógeno puede producir un des-balance de la relación fuente-destino, al magnificarla competencia del destino crecimiento foliar res-pecto al destino crecimiento del grano. En generalse acepta que la capacidad de transporte de fotoa-similados del tejido vascular no es limitante para elrendimiento (Hernández y Oriol¡, 1991).
Mediante defoliaciones selectivas en plantas enel cultivo se ha observado que hay una directa dis-tribución de fotoasimilados hacia los frutos solamen-te desde las hojas superiores que comprenderían elestrato foliar con un IAF de alrededor de la unidad(IAF = 1) (Hernández, L., 1983). Utilizando radioisó-topos se ha ratificado que 'son esas hojas las quecontribuyen al llenado y que además el transporte espreferencial desde hojas hacia porciones del capí-tulo ubicadas directamente por encima de ellas (Her-nández y Orioli, 1991). Así entonces la diferencia de
peso entre los frutos periféricos de porciones supe-riores e inferiores en un capítulo muy inclinado se po-dría explicar por el hecho que al doblarse el cuellose produce una resistencia al flujo de fotoasimiladosdesde las hojas hacia los frutos. Como este flujo esselectivo en cuanto a dirección, el transporte lateralno alcanzaría a compensar la demanda de los frutosubicados en esos sectores del capítulo
Otro ejemplo de la importancia de la posición re-lativa entre la fuente y el destino ocurre en la base ylas brácteas involucrales del capítulo del girasol Lasmismas tienen clorofila y son fotosintéticamente ac-tivas (Rawson y Constable, 1980). Aunque durantealgunas horas del día al menos una fracción del ca-pítulo está a máxima radiación solar, su fotosíntesisneta es levemente negativa debido a pérdidas porrespiración. Ahora bien, como esta fracción verdede la inflorescencia está muy cerca de los frutos endesarrollo, la misma puede contribuir al aporte defotoasimilados durante el período de llenado de losfrutos (Srivastava, et al., 1977; Durrieu et al., 1985).Por otro lado hay evidencias que muestran que du-rante la formación del receptáculo y durante el pe-ríodo de antesis, las hojas inmaduras compiten conla inflorescencia por el suministro de fosoasimiladosprovenientes de hojas inferiores (Hernández y Orio-li, 1991; Hernández, L., 1993).
Factibilidades fisiogenéticas
El conocimiento de las bases fisiológicas del ren-dimiento no va a substituir o reemplazar los proce-dimientos clásicos de mejoramiento vegetal. Por elcontrario, el uso de los datos fisiológicos como uncriterio para la selección dentro de progenies segre-gantes quedará limitada por el instrumental necesa-rio para realizar la experimentación, el tiempo reque-rido para ensayar muchas plantas, herenciacomplicada, dificultad en combinar simultáneamen-te esos componentes fisiológicos con calidad co-mercial, etc.
El rendimiento del cultivo es controlado por mu-chos factores, externos y propios de la especie, ysu expresión depende de la interacción de muchosprocesos fisiológicos. Así por ejemplo cualquier genque afecte la fotosíntesis o el transporte afectará elrendimiento. Si bien no se puede afirmar que no haygenes del rendimiento, pues se conoce que un genpuede generar más rinde (líneas isogénicas), aúnno se sabe cuáles serían las diferencias fisiológicasque lo producen. Por lo tanto si se quieren usar com-ponentes fisiológicos en el mejoramiento vegetal, sedebe tener un completo entendimiento de los me-canismos genéticos que controlan sus cambios (Fe-reres etal., 1986).

El Ideotipo del girasol (Hellanthus annuus L.) 95
La información existente hasta el momento sobreel cultivo de girasol indica que en el mismo existevariabilidad genética para casi todos los diferentescomponentes dei rendimiento. Es difícil que uno so-lo cualquiera de estos caracteres se encuentre alta-mente correlacionado con el rendimiento, siendo lomás probable que el mismo sea la resultante de unbalance entre procesos fisiológicos y bioquímicos.Este balance puede llegar a ser tan delicado que laexcesiva actividad de uno de esos procesos puedelimitar algún otro. Por ejemplo un excesivo desarro-llo foliar podría causar sombreado en las hojas y re-ducir el ritmo fotosintético global de la planta o detransporte de fotoasimilados hacia los frutos
La aplicación de estudios fisiogenéticos en el me-joramiento de variedades de alto rendimiento no esun proceso simple ya que hay un gran número decomponentes fisiológicos del rendimiento, los quea su vez pueden dividirse en subcomponentes. Sinembargo, es posible esperar un rápido progresocuando esos componentes fisiológicos son hereda-dos en forma simple como lo son en trigo y arroz loscaracteres hojas erectas, cortas, gruesas y tallo cor-to. Estas características que están asociadas conun aumento en la distribución de la radiación sobreel canopeo, la respuesta al nitrógeno, resistencia alvuelco de las plantas y aumento del flujo de fotoasi-milados a los frutos, pueden ser combinadas conotros genes que condicionan el ritmo fotosintético,el respiratorio y otros rasgos genéticos y fisiológi-cos, para llegar a que los mismos se expresen enforma óptima.
Como ya se ha mencionado, puede haber algu-na contraparte asociada a un carácter buscado, co-mo por ejemplo la presencia de tallos extremada-mente cortos haría que las hojas estuvieran tancercanas unas de otras que eso dificultaría la distri-bución de luz y CO2 y favorecería el desarrollo deenfermedades. Otra tendencia que está siendo re-vertida es la de usar cultivares de amplia adaptabi-lidad para volver a los localmente adaptados, parade esa manera aprovechar al máximo las condicio-nes locales y por otro lado no quedar sujeto a un po-sible ataque general de una enfermedad al usar so-lo algunos genotipos. Uno de los caminos es laselección de padres para cruzamientos.
En poblaciones relativamente homogéneas lasdiferencias genéticas son más fácilmente identifica-bles, por lo que una variedad con alto rendimientoeconómico o biológico debido a su gran área foliaro mejor distribución de la luz interceptada es másfácilmente identificada. Por lo tanto las líneas paren-tales de los cruzamientos podrían ser selecciona-das sobre la base de su complementación fisiológi-ca potencial, asumiendo que la recombinacióngenética de componentes fisiológicos específicos
dará, en promedio, progenies superiores a los pa-dres, de los cuales nada es conocido acerca de lacapacidad de esos componentes.
El mejoramiento del rinde puede aprovechar unavasta variabilidad genética que está disponible, pe-ro su explotación efectiva y eficiente requiere com-prender la fisiología y la genética asociada por lacual el rendimiento es alcanzado. El índice de cose-cha de muchos cultivos ha sido mejorado a lo largodel tiempo sin mayor atención a la fisiología, pero esposible que si se usan datos fisiológicos para selec-cionar padres, luego usando procedimientos de se-lección y mejoramiento estandards, se podrá selec-cionar directamente por rendimiento Además, elconocimiento de la fisiología genética del rendimien-to aumentará el dominio del fitomejorador respectoal tipo de planta deseada y a los procedimientos deselección y mejoramiento.
Últimamente la biotecnología y la ingeniería gené-tica han comenzado a producir aportes altamente sig-nificativos para la producción agrícola mediante la ob-tención de plantas transgénicas. El girasol no haquedado relegado en este sentido y es factible que elmediano plazo podamos observar también nuevoscultivares mejorados obtenidos por este medio
Desarrollo de modelos
El proceso de selección implica tener un criterioprevio y objetivos definidos. Un fitomejorador tienesiempre un concepto prefijado acerca de la clasede cultivar que va a crear como resultado de su pro-grama de selección genética. Así entonces cual-quier nuevo concepto, que puede ser por ejemplouna variedad más resistente a una enfermedad, alfrío o a la sequía, un mejor balance de aminoácidos,o una más alta producción de carbohidratos, pue-de ser definida como un modelo nuevo de planta
En general la manera de encarar el mejoramien-to de una especie se puede realizar de dos formasLa primera, eliminando un defecto, por lo que el ob-jetivo es mejorar la variedad remediando un carác-ter indeseable particular. Como alternativa se pue-de simplemente seleccionar por altos rendimientossin preocuparse por las razones que hacen a eseaumento del rendimiento. La segunda es aquella enla cual se trata de lograr plantas modelos con carac-terísticas deseables prefijadas. En efecto, con el co-nocimiento de la fisiología, morfología y anatomía delas plantas cultivadas, algunos investigadores creenpoder definir modelos de superior productividad, lacombinación teórica de caracteres constituye el finde sus esfuerzos. Encarar el problema de esa ma-nera presupone tener un adecuado conocimientode los rasgos morfológicos, anatómicos, fisiológicosy bioquímicos que hacen al aumento de productivi-

96 AGRISCIENTIA
dad o de la calidad del producto. Sin embargo a pe-sar de la creciente cantidad de información obteni-da recientemente, en la mayoría de los casos loscomponentes que hacen al nuevo modelo no han si-do definidos con precisión. Como ya se ha dicho loscambios que dieron origen al aumento de producti-vidad no han sido aún definidos y posiblemente sondebidos a una gran combinación de factores.
De esta descripción surge entonces que la dife-rencia entre los dos tipos de selección, eliminacióndel defecto y creación del ideotipo, es una cuestiónde semántica. Sin embargo la planificación de unmodelo nuevo significa actuar en un marco más am-plio y abstracto sobre la base del conocimiento delos componentes más relevantes que hacen a la pro-ducción, y así decidir sobre que conjunción de ras-gos constituirán una variedad mejorada. El resulta-do de tales consideraciones puede ser una plantatotalmente distinta a la de las mejores variedadesactuales, y no solamente una forma comparable ala que solo se le ha eliminado un defecto.
Se puede ser optimista respecto a que las nue-vas formas de plantas pueden proveer otro ejemplode éxito. Así mutantes con una morfología drástica-mente diferente a aquellas que han evolucionadobajo la presión de la selección pueden no solo serpromisiorios económicamente sino que pueden in-fluenciar la actitud del fitomejorador hacia las posi-bilidades de reestructurar genéticamente otros cul-tivos, produciendo en ellos innovaciones exitosas.
Muchas veces se conocen cambios bien marca-dos en los cultivos pero que hasta la fecha no hansido estudiados. Cuando se toma la decisión de exa-minar el potencial valor agronómico de esa muta-ción, se encuentra que la misma está disponible so-lo en un marco genético muy mal adaptado alamoderna producción comercial. Por lo tanto la eva-luación debe ser precedida por un programa de cru-zamientos para introducir los genes mutantes en unmarco genético más adaptado.
CONCLUSIONES
Futuros progresos en el mejoramiento del girasolrequerirán que tanto la fotosíntesis como capacidadde almacenaje aumenten en una manera coordina-da; hemos visto que la fotosíntesis durante la fasede crecimiento del grano es una determinante im-portante del rendimiento, pero la fotosíntesis previacontribuye a la determinación de la capacidad dealmacenamiento y genera reservas que pueden sermovilizadas durante la fase de llenado. Es posibleque alguna vez las reservas fueron el mayor com-ponente del rendimiento, tendencia que se revirtiócon la adaptación y mejoramiento de la especie.
¿Hay un límite para que aumente el tamaño y pe-so de los frutos y su contenido de aceite? ¿Cuálesson los factores que limitan su crecimiento? Estasrespuestas distan de ser respondidas con los cono-cimientos actuales. Se entiende sin embargo queexiste una clara tendencia hacia un control de tipoagronómico. Ese control, traducido en técnicas demanejo, determinará que las plantas puedan con-centrar fotoasimilados en los órganos cosechables(granos) a expensa de otros órganos, indispensa-bles para su sobrevivencia bajo condiciones natu-rales. Así con el uso de riego y fertilizantes se requie-ren menos raíces. Con más herbicidas y controlmecánico de las malezas no se necesita tanta altu-ra en los tallos. Con el control de enfermedades sereduce la necesidad de reservas para que la plan-ta se recobre luego de una infestación. Es decir quecon cada avance agronómico indirectamente se lo-gra que una mayor parte del peso total de la planta(Rb) quede libre para ser invertido en los órganosde cosecha (Re).
A continuación se proponen algunos de los ca-racteres morfológicos y fisiológicos que deberíanconstituir el ideotipo de una planta nueva de girasol:
Raíces :'Bien extendidas en profundidad. Capa-cidad elevada de absorción de agua y nutrientes mi-nerales. Moderada resistencia al flujo de agua
Tallos : De un porte mediano, robusto para evitarel vuelco. Capaz de mantener esas condiciones aaltas densidades Abundante clorofila que se man-tenga durante el período de llenado del grano. Acen-tuada funcionalidad en cuanto a servir de órgano dereserva y con buena capacidad para redistribuir du-rante el período de llenado de los frutos los carbo-hidratos acumulados durante preantesis.
Hojas : Que conserven su capacidad de segui-miento de la luz solar durante todo el ciclo de vidaCon un número de 20 a 25 por planta capaces deasegurar un IAF de al menos 3 para antes del perío-do de floración. Alargamiento de la duración delárea foliar superior. Conservar su elevada capaci-dad, de fotosíntesis, saturación luminosa alta, tem-peratura óptima alta, etc. Capacidad de cubrir elsuelo lo más pronto posible luego de la emergencia.Retardo de senescencia foliar a altas densidades.
Capítulo : Diámetro grande desde el inicio de ladiferenciación de los primordios florales. Rápido rit-mo de expansión para permitir ubicar más flores porunidad de superficie meristemática. Elevada auto-compatibilidad. Muchas brácteas para aumentar sucapacidad fotosintética. Menores pérdidas por res-piración.
Frutos : Aumentar el número. Aumento de pesoen condiciones de óptimo crecimiento. Mayor tama-ño. Alto contenido de aceite y alto porcentaje de áci-

El ldeotipo del girasol (Hellanthus annuus L ) 97
do oleico. Alargamiento del período del llenado. Ma- Anfinrud, M.N y A. A. Schneiter, 1984. Relationship of sun-yor lapso entre el comienzo antesis y el comienzo flower germination and vigor test to field performancede su llenado. Crop Sci. 24:341-344.
Índice de cosecha : Aumentarlo por disminucióndel peso del tallo a cosecha.
En general : Disminuir la adaptación amplia pa-ra volver a la adaptación local. Capacidad de pro-ducir a muy altas densidades. Capacidad de pro-ducir bajo nuevas prácticas agronómicas como"intercropping" y labranza "cero". Mejor utilizaciónde los recursos agua y nutrientes minerales. Mejorrespuesta ala fertilización. Resistente a plagas y en-fermedades.
AGRADECIMIENTOSL.F.H. es Miembro de la Carrera del Investigador
Científico de la Comisión de Investigaciones Cientí-ficas de la Provincia de Buenos Aires (CIC). G.A.O.es Miembro de la Carrera del Investigador Científi-co del Consejo Nacional de Investigaciones Cientí-ficas y Técnicas (CONICET).
Los autores agradecen a las Empresas DekalbArgentina S.A. y Cargill SACI por el suministro dematerial para experimentación. Las sugerenciasaportadas por los Ings. Agrs. A. Fernández (DekalbArgentina S.A.) y F. Mockel (Dept. de Agronomía -UNS) son muy apreciadas. Los trabajos de los au-tores que han sido citados en este artículo han sidorealizados con subsidios otorgados por la CIC de laProvincia de Buenos Aires, el CONICET, la Funda-ción Antorchas y la UNS.
BIBLIOGRAFÍAAguirrezábal, L.A.N., 1993 Modelización de la ramifica-
ción y el alargamiento del sistema radical del girasol(Helianthus annuus L ). Efecto de la radiación intercep-tada y de la temperatura. Tesis de Doctorado. Univer-sidad Blas Pascal, Francia
Aguirrezábal. L.A.N., S. Pellerin y F. Tardieu, 1993 Carbonnutrition, root branching and elongation. Can the pre-sent state of knoeledge allow a predictive approach ata whole plant level? Environ. Exp. Bot. 33. 121-130.
Aguirrezábal. L.A.N., E. Deleens y F. Tardieu, 1994. Rootelongation rate is accounted for by intercepted PPFD andsource-sink relationships in field and laboratory grownsunflower. Plant Cell and Env. 17: 443-450.
Aguirrezábal. L.A.N., F.J. Cardinali, G.A. Orioli y L.S. Tes-ton¡, 1985. Influencia del tamaño de aquenio sobre elcrecimiento y rendimiento de la planta de girasol(Helianthus annuus L.). Actas de la XI Cont. Internac.de Girasol. Mar del Plata, Argentina, pp. 39-43.
Anderson, W.K , 1975 Maturation of sunflower Aust. J.Exp Anim Husb 15 833-838
Beard , B.H. y S. Geng, 1982. Interrrelationships of morp-hological and economic characters of sunflower. CropSci. 22: 817-822.
Bingham, J., 1967. Investigation on the physiology of yieldin winter wheat by comparison of varieties and by arti-ficial variation in grain number per ear J Agric Sci 6841 1-422
Black, C.R., 1979 The relative magnitude of the partial re-sistances to transpirational water movement in sunflo-wer (Helianthus annuus L.) J. Exp. Bot. 30 245-253
Blarney, F.P.C., D. Mould y J. Chapman, 1979 Critical bo-ron concentrations in plant tissues of two sunflower cul-tivars in relation to seed yield. Agron. J 71. 243-247
Bradshaw, A.D., 1965. Evolutionary significance of pheno-tipic plasticity in plants. Adv. Genet. 13 115-155
Bremner, P.M. y H.M. Rawson, 1978. The weight of indivi-dual grains of the wheat ear in relation to their growthpotential, the supply of assimilates and interaction bet-ween grains. Aust. J. Plant Physiol. 5 61-72.
Brocklehurst, P.A., 1977. Factors controlling grain weightin wheat. Nature, (Lond.) 266: 348-349
Cardinali, F.J., G.A. Orioli y V.R. Pereyra, 1985. Comporta-miento de dos híbridos de girasol a bajas densidadesde siembra. Rev. Fac. deAgron. (Bs. As.), 6: 131-139.
Connor, D,J, y V.0 Sadras, 1992. Physiology of yield ex-pression in sunflower. Field Crops Res. 30. 333-389
Chapman, S.C., G L. Hammer y M. Meinke, 1993 A sun-flower simulation model. I Model development AgronJ. 85: 725-735.
Donald, C M y J Hamblin, 1976 The biological yield andharvest index of cereals as agronomic and plant bree-ding criteria. Adv. Agron 28 361-405
Durrieu, G , C P du Sort y A. Merrien, 1985 Anatomie ducapitule de tournesol. Consequences sur la nutrition desakenes. Actas de la XI Cont. Internac. de Girasol, Ar-
gentina, pp. 7-12.
English, S.D., J.R McWilliam, R.C.G Smith y J.L David-son, 1979. Photosynthesis and partitioning of dry mat-ter in sunflower. Aust. J. Plant Physiol 6 149-164
Evans, L.T., 1975. The physiology basis of crop yield EnCrop Physiology. (L.T Evans Ed.) Cambridge Univer-sity Press, London. pp 327-355,
Evans, L.T, 1976. Physiological adaptation to perfomanceas crop plants. Phil. Trans. R. Soc. Lond. B. 275: 71-83.
Evans, L.T. y IF. Wardlaw, 1976. Aspects of the compara-tive physiology of grain yield in cereals. Adv. Agron. 28.301-359
Fereres, E., C. Giménez y J.M. Fernández, 1986 Geneticvariability in sunflower cultivars under drought. I. Yieldrelationships. Aust. J. Agric. Res. 37: 573-582
Gaines, M.S., K.J. Vogt, J.L Hamrick y J Caldwéll, 1974Reproductive strategies and growth patterns in'sunflo-wers (Helianthus) Amer Natur 108. 889-894

98
Gifford, M. y L.T. Evans, 1981. Photosynthesis, carbon par-titioning and yield. Annu. Rev. Plant Physiol. 32: 485-509
Goyne, P.J y A.A. Scheneiter, 1987. Photoperiod influen-ce on development in sunflower genotypes. Agron J.79: 704-709
Hall, A J., D.J Connor y D.M. Whitfield, 1989 Contributionof pre-anthesis assimilates to grain filling in irigated andwater-stressed sunflower crops. I. Estimates using la-belled carbon. Field Crops Res. 20: 95-112.
Hall, A.J., D.J Connor y D.M Whitfield, 1990. Contributionof pre-anthesis assimilates to grain filling in irigated andwater-stressed sunflower crops. 11, Estimates from a car-bon budget. Field Crops Res. 24 273-294,
Hernández, L.F., 1983, Estudios sobre el desarrollo de losfrutos de girasol (Helianthus annuus L ) y su respuestaa factores de estrés. Tesis de Magister Scientiae. De-partamento de Agronomía. Universidad Nacional delSur, Argentina, 178 pp.
Hernández, L.F., 1988. Organ Morphogenesis and Phyllo-taxis in the Capitulum of Sunflower (Helianthus annuusL.) Ph D. Thesis. Department of Botany, Univ. of NewSouth Wales, Australia. 217 pp.
Hernández. L.F., 1993. Grain yield of the sunflower capitu-tum promoted py surgical removal of the involucral bractpririnordia Trends in Agricultural Sciences, 1' 127-135.
Hernández, L.F. y G.A. Orioli, 1982 Growth analysis of irri-gated sunflower at two plant populations. Actas de la XConf Internac. de Girasol, Australia, pp. 18-21
Hernández, L F y G A Orioli, 1983. Estudio comparativode la estructura foliar, intercepción de luz y rendimien-to en girasol Anales de Edafol yAgrobiol (Madrid) 42-2137-2148
Hernández, L F y G A Orioli, 1985 a Imbibition and germi-nation rates of sunflower seed according to fruit sizeField Crops Res, 10: 355-360
Hernández, L.F. y G.A Orioli, 1985 b Edaphic variationsin irrigated soils and its influence on growth and yield ofsunflower. Actas de la Xl Conf. Internac. de Girasol, Ar-gentina, pp. 227-233.
Hernández, L.F. y G.A. Oriol¡, 1985 c. Pubescencia foliaren girasol y su influencia sobre la disipación de calordesde la hola. Actas de la XI Conf. Internac. de Girasol.Mar del Plata, Argentina, pp.13-19
Hernández, L.F. y G A Orioli, 1985 d Relationships bet-ween root permeability to water, leaf conductance andtranspiration rate in sunflower (Helianthus annuus Lcultivars. Plant and Soil 85: 229-235.
AGRISCIENTIA
Hernández, L.F. y GA. Oriol¡, 1991. Participación de la di-ferentes hojas de la planta de girasol durante el llena-do de los frutos. Turnalba 41: 330-334.
Hernández, L.F. y G.A Oriol¡, 1992. Incidencia del númerode frutos cosechables en la determinación del poten-cial de rendimiento del cultivo de girasol ProcederAgrotecnológico 4: 56-63.
Herzog, H , 1982 Relation of source and sink during grainfilling period in wheat and some aspects of its regula-tion. Physiol. Plant. 56: 155-160
Hocking, P.J. and B.T. Steer, 1983. Uptake and partitioningof selected mineral elements in sunflower (Helianthusannuus L) during growth. Field Crops Res 6 93-107
Koide, R., 1985 The nature and location of variable hydrau-lic resistance in Helianthus annuus L. (sunflower). JExp. Bot. 36. 1430-1440.
Lloyd, N D H. y D T. Canvin, 1977 Phosynthesis and pho-torespiration in sunflower selections. Can. J. Bot. 55.3006-3012.
Marc, J y J.H Palmer, 1981. Photoperiodic sensitivity ofinflotrescence initiation and development in sunflowerField Crops Res. 4: 155-164.
Martinez-Carrasco, R. y G.N Thorpe, 1979 Phisiological fac-tors limiting grain size in wheat J. Exp. Bot. 30, 669-679.
Orioli, G.A., V.R. Pereyra, J. Beltrano y F. Cardinali, 1977.Acumulación de materia seca, nitrógeno, fósforo y usode energía en un cultivo de girasol. IADO. Actas de la 111Reunión Nacional de Girasol. Buenos Aires. pp. 137-41.
Pereyra, V.R., J. Beltrano y G.A. Orioli, 1977. Densidad desiembra y producción en cultivos de girasol ADO Ac-tas de la Ill Reunión Nacional de Girasol. Buenos Aires.pp. 127-131.
Pieters, G A, 1985. Effects of irradiation level on leaf growth.of sunflower. Physiol Plant 65 263-268
Rawson, H M , 1979 Vertical wilting and photosynthesis,transpioration and water use efficiency of sunflower lea-ves Aust J Plant Physiol 6' 109-120
Rawson, H M. y G.A Constable, 1980. Carbon productionof sunflower cultivars in field and controlled environ-ments. I Photosynthesis and respiration of leaves, stemand heads, Aust. J. Plant Physiol. 7. 555-573
Shell. G.S.G. y A R.G. Lang, 1976. Movements of sunflowerleaves over a 24-h period, Agric. Meteorol 16. 161-17b
Srivastava, G.C., P.S. Deshmukh y D P.S. Tomar, 1977 Sig-nificance of leaf orientation and bract in seed yield insunflower. Indian J. Plant Physiol 20 151-156.
Steer, B.T y P J. Hocking, 1983 Leaf and floret productionin sunflower (Helianthus annuus L.) as affected by nitro-gen supply. Ann. Bot. 52: 267-277.


















