
Effects of germanium dioxide, an inhibitor of diatom growth, on the microscopic laboratory...
6
J Appl Phycol (2007) 19:27–32 DOI 10.1007/s10811-006-9107-x ORIGINAL ARTICLE Effects of germanium dioxide, an inhibitor of diatom growth, on the microscopic laboratory cultivation stage of the kelp, Laminaria saccharina Ryan Shea · Thierry Chopin Received: 25 May 2006 / Accepted: 1 June 2006 / Published online: 29 November 2006 C Springer Science + Business Media B.V. 2006 Abstract The kelp, Laminaria saccharina, is an eco- nomically and biologically valuable seaweed used in integrated multi-trophic aquaculture. The development of the initial microscopic phase of the life cycle of this species is carried out in the laboratory. A treatment of germanium dioxide (GeO 2 ) can be applied to algal lab- oratory cultures to inhibit the growth of contaminating diatoms. Previous studies reported inhibitory effects also affecting the macroscopic stage of the life cycle of L. saccharina, the sporophyte, but the effects on the microscopic gametophytic life stage are unknown. To determine the effects, cultures of L. saccharina were treated with varying concentrations of GeO 2 and the resulting lengths and surface area of the juvenile sporo- phytes produced were measured. It was determined that GeO 2 follows a typical dose-effect pattern, increasing the growth rate of juvenile sporophytes until a critical point at which an inhibitory effect was observed. To obtain juvenile sporophytes ready for deployment to aquaculture sites in the shortest time and to success- fully reduce diatom contamination, it was determined that a concentration between 0.10 and 0.50 mL of GeO 2 per litre of seawater, applied at day 8, was the most efficient. R. Shea · T. Chopin () Centre for Environmental and Molecular Algal Research, Centre for Coastal Studies and Aquaculture, Department of Biology, University of New Brunswick, Saint John, New Brunswick E2L 4L5, Canada e-mail: [email protected] Keywords Diatom inhibition . Gametophyte . Germanium dioxide . Integrated aquaculture . Laminaria saccharina . Multi-trophic Introduction Laminaria saccharina (Linnaeus) Lamouroux is cur- rently being cultivated as the inorganic extractive com- ponent of an integrated multi-trophic aquaculture sys- tem along with Salmo salar Linnaeus (fed component) and Mytilus edulis Linnaeus (organic extractive compo- nent) in the Bay of Fundy, Canada (Chopin et al., 2004). Laminaria saccharina has an alternation of genera- tion life cycle between two distinct phases: the macro- scopic, diploid sporophyte, and the microscopic, hap- loid gametophyte. In the seedstock production stage, spores extracted from sporophytes are cultured in lab- oratory conditions until the life cycle is completed to produce juvenile kelp sporophytes, at which time they are deployed to the aquaculture sites (Merrill & Gillingham, 1991). During the microscopic stage of production, there is a possibility of contamination, es- pecially by other algae. The most serious effect of foul- ing by algae is the potential overgrowth and elimina- tion of the kelp germlings, most typically by diatoms (Merrill & Gillingham, 1991). A treatment of germa- nium dioxide (GeO 2 ) has been recommended, but its effects have not been clearly documented. Lewin (1966) reported that GeO 2 is a specific in- hibitor of diatom growth and interferes in some way Springer
Transcript of Effects of germanium dioxide, an inhibitor of diatom growth, on the microscopic laboratory...

J Appl Phycol (2007) 19:27–32DOI 10.1007/s10811-006-9107-x
O R I G I N A L A R T I C L E
Effects of germanium dioxide, an inhibitor of diatomgrowth, on the microscopic laboratory cultivation stageof the kelp, Laminaria saccharina
Ryan Shea · Thierry Chopin
Received: 25 May 2006 / Accepted: 1 June 2006 / Published online: 29 November 2006C© Springer Science + Business Media B.V. 2006
Abstract The kelp, Laminaria saccharina, is an eco-nomically and biologically valuable seaweed used inintegrated multi-trophic aquaculture. The developmentof the initial microscopic phase of the life cycle of thisspecies is carried out in the laboratory. A treatment ofgermanium dioxide (GeO2) can be applied to algal lab-oratory cultures to inhibit the growth of contaminatingdiatoms. Previous studies reported inhibitory effectsalso affecting the macroscopic stage of the life cycleof L. saccharina, the sporophyte, but the effects on themicroscopic gametophytic life stage are unknown. Todetermine the effects, cultures of L. saccharina weretreated with varying concentrations of GeO2 and theresulting lengths and surface area of the juvenile sporo-phytes produced were measured. It was determined thatGeO2 follows a typical dose-effect pattern, increasingthe growth rate of juvenile sporophytes until a criticalpoint at which an inhibitory effect was observed. Toobtain juvenile sporophytes ready for deployment toaquaculture sites in the shortest time and to success-fully reduce diatom contamination, it was determinedthat a concentration between 0.10 and 0.50 mL of GeO2
per litre of seawater, applied at day 8, was the mostefficient.
R. Shea · T. Chopin (�)Centre for Environmental and Molecular Algal Research,Centre for Coastal Studies and Aquaculture, Department ofBiology, University of New Brunswick, Saint John, NewBrunswick E2L 4L5, Canadae-mail: [email protected]
Keywords Diatom inhibition . Gametophyte .
Germanium dioxide . Integrated aquaculture .
Laminaria saccharina . Multi-trophic
Introduction
Laminaria saccharina (Linnaeus) Lamouroux is cur-rently being cultivated as the inorganic extractive com-ponent of an integrated multi-trophic aquaculture sys-tem along with Salmo salar Linnaeus (fed component)and Mytilus edulis Linnaeus (organic extractive compo-nent) in the Bay of Fundy, Canada (Chopin et al., 2004).Laminaria saccharina has an alternation of genera-tion life cycle between two distinct phases: the macro-scopic, diploid sporophyte, and the microscopic, hap-loid gametophyte. In the seedstock production stage,spores extracted from sporophytes are cultured in lab-oratory conditions until the life cycle is completedto produce juvenile kelp sporophytes, at which timethey are deployed to the aquaculture sites (Merrill &Gillingham, 1991). During the microscopic stage ofproduction, there is a possibility of contamination, es-pecially by other algae. The most serious effect of foul-ing by algae is the potential overgrowth and elimina-tion of the kelp germlings, most typically by diatoms(Merrill & Gillingham, 1991). A treatment of germa-nium dioxide (GeO2) has been recommended, but itseffects have not been clearly documented.
Lewin (1966) reported that GeO2 is a specific in-hibitor of diatom growth and interferes in some way
Springer

28 J Appl Phycol (2007) 19:27–32
with the processes of silica frustule formation. Theeffect of GeO2 on the main algae in culture hashad very little investigation and is poorly understood.Several early studies on the effect of this compoundconcluded that GeO2 inhibited the growth of matureChlorophyta and Phaeophyta species; however, theRhodophyta species under study showed no growth in-hibition (Chen et al., 1969, 1970; McLachlan et al.,1971; Hopkin & Kain, 1978; Markham & Hagmeier,1982). Markham and Hagmeier (1982) conducted stud-ies on macroscopic L. saccharina sporophytes using the“hole punch” method and concluded that L. saccharinadid show inhibition of growth from GeO2, but sug-gested that GeO2 could still be used without seriousdamage in most cultures of Phaeophyta at concentra-tions not exceeding 0.01–0.04 mL of a saturated solu-tion of GeO2 per litre. Merrill and Gillingham (1991)recommended using a treatment of 1–2 mL of a satu-rated solution of GeO2 per litre during the microscopicgametophyte development of Nereocystis luetkeana(approximately at day 10). However, they did not men-tion any consequences regarding the effects this chemi-cal would have on kelp growth. Since it was known thatgermanium potentially replaced silica atoms in com-pounds, the GeO2 treatment may have been assumedto have no detrimental effects on kelp, since silica isnot a major or essential element for L. saccharina andmost macroalgae (Fritsch, 1971).
Because of these conflicting results between the mi-croscopic and macroscopic stages of different speciesof kelps and the wide range of GeO2 concentrationsused in the studies mentioned above (between 0.045and 10 mg GeO2 per litre), this study was undertakento determine the effect of the diatom inhibitor, GeO2,when applied to the microscopic gametophytic stage ofL. saccharina in culture. This was achieved by compar-ing the effects of different concentrations of GeO2 onthe length and surface area of the juvenile sporophytesproduced.
Materials and methods
Laboratory preparation of cultivation spools
The substrate used for cultivating the kelps was a2.5 mm flat nylon twine, which was cut into 60 separatepieces, each measuring 14 m in length. The pieces ofnylon twine were then soaked in boiling distilled water
to remove the waxy film. The nylon twines were thenspooled onto 60 PVC pipes (18 cm × 7.5 cm diameter),using a mechanical lathe. The spools were then soakedin sodium carbonate, followed by a distilled waterrinse.
Seaweed collection and inoculation
Forty-four sexually mature L. saccharina sporophyteswith large distinct sori were collected at Green’sPoint, Letite, New Brunswick, Canada (45◦02′15′′N;66◦53′30′′W).
The sori were cut from the kelps, cleaned, and placedin a refrigerator overnight. They were then removedfrom the refrigerator and allowed to dry at room temper-ature for approximately 3 h. A spore release followedby re-immersion of the sori into filtered seawater at11 ◦C, and stirring every 30 min for 4 h. The spore solu-tion was then added to the inoculation chamber (200 Ltank with gentle aeration) containing the spools andfilled with seawater filtered to 0.2 μm. The chamberwas covered and kept in darkness for 24 h.
The culture tanks (10 × 60 L white laundry tubs)were supplied with aeration tubes, a transparent Plex-iglas cover, and two cool fluorescent lights (80–100 μE m−2 s−1) set to a 18:6 L:D photoperiod regime.They were filled with 0.2 μm filtered seawater main-tained at 11 ± 1 ◦C, a pH of 8, and a salinity of33 ± 1 ppt. After the 24 h inoculation period, the spoolswere removed from the chamber and randomly placedin the culture tanks with gentle aeration.
Germanium dioxide treatment
A saturated solution of GeO2 was prepared by adding0.894 g of reagent grade, powdered GeO2 (Acros Or-ganics) to a 200 mL volumetric flask with de-ioniseddistilled water. It was then placed on a stirrer and al-lowed to mix overnight. The culture tanks randomlyreceive a treatment of 0.00, 0.02, 0.10, 0.50, or 1.00 mLof GeO2 per litre of seawater on day 8 (two tanks foreach treatment).
Sampling
Samples were taken twice a week once juvenile sporo-phytes were observed on the twines. Three randomlyselected spools from each tank were sampled by pluck-ing a small piece of the nylon twine using a pair
Springer

J Appl Phycol (2007) 19:27–32 29
of forceps. The samples were then placed on threeseparate slides and viewed under a light microscopeequipped with a digital camera. From each slide, fivedigital pictures were captured and transferred to a com-puter for analysis. The photo software program, ImageJ, was used to calculate the sporophyte length and sur-face area.
Statistical analyses
All statistical analyses were preformed with theMinitabTM s14 statistical package. In order to pool thedata from the two tanks with the same treatment, anAnderson – Darling normality test was conducted todetermine if the data points were normally distributed.Once normality was determined, an F-test was con-ducted to observe if there was any difference in vari-ance between the two tanks. All data sets were con-sidered normal (Anderson–Darling test, p > 0.05) andtreatment tanks were homogeneous (F-test, p > 0.05);consequently, the data from the tanks with the sametreatment were pooled.
The effects of the different treatments on the sporo-phyte length and surface area were assessed for eachsampling date by one-way ANOVA, holding sampledate fixed and using either length or surface area asthe response variable. A posteriori comparison amongtreatments was then made using a Tukey post-hoc mul-tiple comparisons test (α = 0.05). The influence of thelength of time on the sporophyte length and surfacearea for each treatment was then assessed by anotherone-way ANOVA, holding the treatment fixed and us-ing the length and surface area at each sample dateas the response. A posteriori comparison among treat-ments was then made using a Tukey post-hoc multiplecomparisons test (α = 0.05).
Results
Diatom contamination was stopped in all the tankstreated with GeO2. Over the length of the study (40days), it was observed that the various GeO2 treatmentsdid not trigger any delay of the time at which the suc-cessive life stages of L. saccharina occurred (based onprevious years’ culture schedules). The effect was onlyobserved on the size of the kelps at each developmentalstage.
Sporophyte length as a function of germaniumdioxide concentration
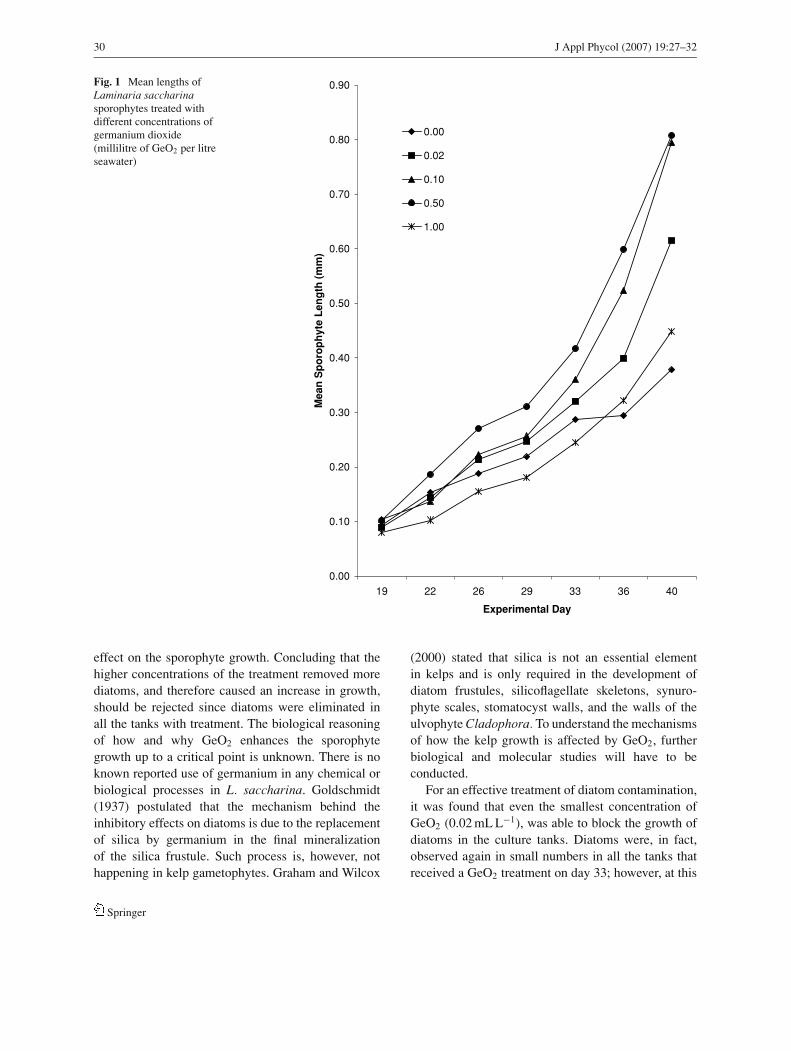
A significant difference was observed in the meanlengths of the sporophytes exposed to different con-centrations of GeO2 (Fig. 1).
On the final experimental day, day 40, the lengths ofthe sporophytes exposed to 0.50 and 0.10 mL GeO2
per litre were not significantly different from eachother but were both significantly longer (p < 0.001)than the sporophytes exposed to all other treatments.The lengths of the sporophytes treated with 1.00 and0.00 mL GeO2 per litre were also not significantly dif-ferent from each other but were significantly shorter(p < 0.001) than those from all the other treatments.
The sporophytes grown under 1.0 mL GeO2 per litretreatment were much more abundant than the ones thatreceived no GeO2 treatment.
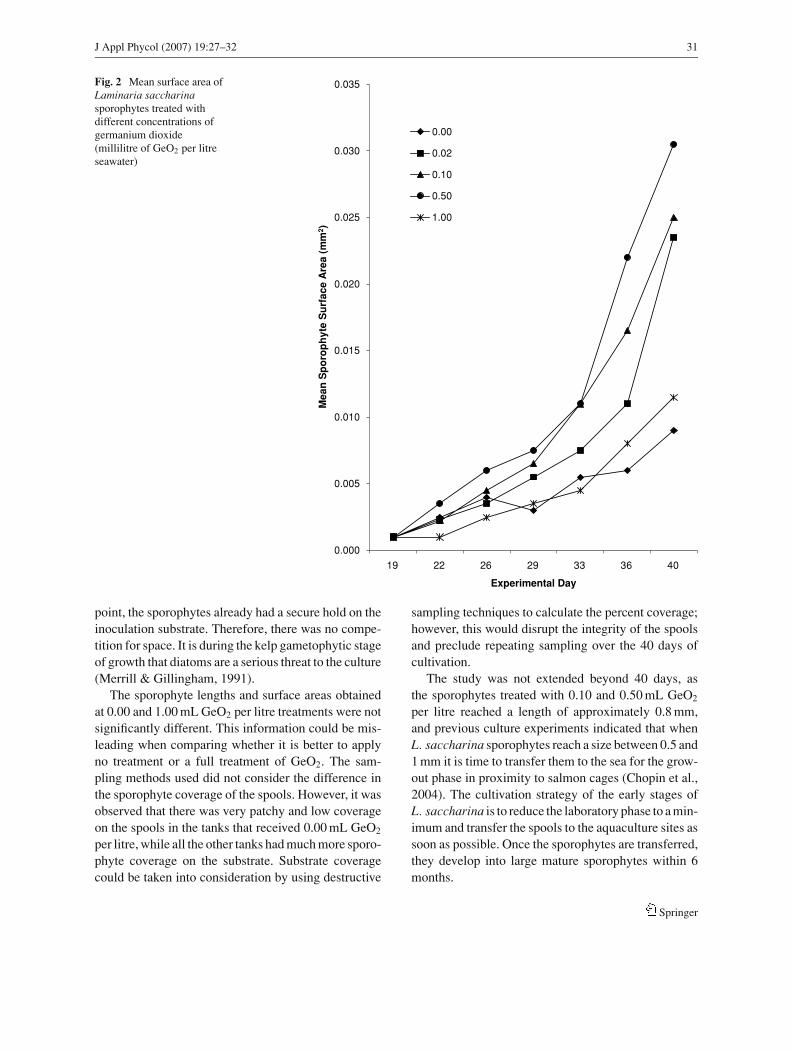
Sporophyte surface area as a function ofgermanium dioxide concentration
On the final sampling date, day 40, the surface areasof the sporophytes receiving 0.02, 0.10, and 0.50 mLGeO2 per litre treatments were not significantly dif-ferent from each other, but were significantly larger(p < 0.001) than those exposed to 0.00 and 1.00 mLGeO2 per litre treatments, for which the surface areaswere not statistically different (Fig. 2).
Discussion
When applied to the gametophytic life stage of L.saccharina, GeO2 was observed to have a significanteffect on the length and surface area of the sporophytesproduced. It, however, did not have an effect on the tim-ing of the occurrence of the successive developmentalstages of the life cycle of L. saccharina comparedto what has been observed in previous years withoutGeO2 treatments. A general dose effect occurred whenan increase in GeO2 concentration increased the growthrate of the sporophytes to a critical point at which GeO2
had a severe inhibitory effect on sporophyte growth.This is observed as an increase in the length andsurface area of the sporophytes receiving a treatmentthat increases from 0.00 to 0.02 to 0.10 and to 0.50 mLGeO2 per litre, respectively. At the highest treatmentof 1.00 mL GeO2 per litre, GeO2 had an inhibitory
Springer
30 J Appl Phycol (2007) 19:27–32
0.00
0.10
0.20
0.30
0.40
0.50
0.60
0.70
0.80
0.90
19 22 26 29 33 36 40
Experimental Day
Mean
Sp
oro
ph
yte
Le
ng
th (
mm
)
0.00
0.02
0.10
0.50
1.00
Fig. 1 Mean lengths ofLaminaria saccharinasporophytes treated withdifferent concentrations ofgermanium dioxide(millilitre of GeO2 per litreseawater)
effect on the sporophyte growth. Concluding that thehigher concentrations of the treatment removed morediatoms, and therefore caused an increase in growth,should be rejected since diatoms were eliminated inall the tanks with treatment. The biological reasoningof how and why GeO2 enhances the sporophytegrowth up to a critical point is unknown. There is noknown reported use of germanium in any chemical orbiological processes in L. saccharina. Goldschmidt(1937) postulated that the mechanism behind theinhibitory effects on diatoms is due to the replacementof silica by germanium in the final mineralizationof the silica frustule. Such process is, however, nothappening in kelp gametophytes. Graham and Wilcox
(2000) stated that silica is not an essential elementin kelps and is only required in the development ofdiatom frustules, silicoflagellate skeletons, synuro-phyte scales, stomatocyst walls, and the walls of theulvophyte Cladophora. To understand the mechanismsof how the kelp growth is affected by GeO2, furtherbiological and molecular studies will have to beconducted.
For an effective treatment of diatom contamination,it was found that even the smallest concentration ofGeO2 (0.02 mL L−1), was able to block the growth ofdiatoms in the culture tanks. Diatoms were, in fact,observed again in small numbers in all the tanks thatreceived a GeO2 treatment on day 33; however, at this
Springer
J Appl Phycol (2007) 19:27–32 31
0.000
0.005
0.010
0.015
0.020
0.025
0.030
0.035
19 22 26 29 33 36 40
Experimental Day
Me
an
Sp
oro
ph
yte
Su
rfa
ce
Are
a (
mm
2)
0.00
0.02
0.10
0.50
1.00
Fig. 2 Mean surface area ofLaminaria saccharinasporophytes treated withdifferent concentrations ofgermanium dioxide(millilitre of GeO2 per litreseawater)
point, the sporophytes already had a secure hold on theinoculation substrate. Therefore, there was no compe-tition for space. It is during the kelp gametophytic stageof growth that diatoms are a serious threat to the culture(Merrill & Gillingham, 1991).
The sporophyte lengths and surface areas obtainedat 0.00 and 1.00 mL GeO2 per litre treatments were notsignificantly different. This information could be mis-leading when comparing whether it is better to applyno treatment or a full treatment of GeO2. The sam-pling methods used did not consider the difference inthe sporophyte coverage of the spools. However, it wasobserved that there was very patchy and low coverageon the spools in the tanks that received 0.00 mL GeO2
per litre, while all the other tanks had much more sporo-phyte coverage on the substrate. Substrate coveragecould be taken into consideration by using destructive
sampling techniques to calculate the percent coverage;however, this would disrupt the integrity of the spoolsand preclude repeating sampling over the 40 days ofcultivation.
The study was not extended beyond 40 days, asthe sporophytes treated with 0.10 and 0.50 mL GeO2
per litre reached a length of approximately 0.8 mm,and previous culture experiments indicated that whenL. saccharina sporophytes reach a size between 0.5 and1 mm it is time to transfer them to the sea for the grow-out phase in proximity to salmon cages (Chopin et al.,2004). The cultivation strategy of the early stages ofL. saccharina is to reduce the laboratory phase to a min-imum and transfer the spools to the aquaculture sites assoon as possible. Once the sporophytes are transferred,they develop into large mature sporophytes within 6months.
Springer

32 J Appl Phycol (2007) 19:27–32
Merrill and Gillingham (1991) reported that ob-taining 0.5 mm long sporophytes may take up to 60days in Nereocystis luetkeana. Results from this studyshow that this length was reached by sporophytes ofL. saccharina treated with 0.50 and 0.10 mL GeO2 perlitre in approximately 34 and 35 days. The sporophytesfrom 0.02 mL GeO2 per litre treatment reached the de-ployment size at about day 38, while the sporophytesexposed to 0.00 and 1.00 mL GeO2 per litre treatmentsdid not reach the length for deployment during thecourse of this study.
In conclusion, GeO2 not only inhibits the diatomcontamination in tanks at a critical time of gameto-phytic development in L. saccharina but also has asignificant effect on the growth of the early stages of de-velopment of the non-targeted species, L. saccharina.This effect is positive until a critical point is reached,between 0.50 and 1.00 mL of GeO2 per litre. At thispoint, GeO2 has a negative inhibitory effect on the labo-ratory growth of L. saccharina. The results of this studysuggest that a treatment of GeO2 between 0.10 and0.50 mL GeO2 per litre should be added to the industrialcultivations of L. saccharina to both inhibit the contam-ination of diatoms and reduce the laboratory cultivationtime by producing sporophytes of appropriate size fordeployment in the shortest time period. Moreover, it isworth noting that our recommended treatment concen-trations are much lower than any previously indicated:between 0.10 and 0.50 mL of a saturated solution ofGeO2 per litre of seawater (or between 0.0004 and0.0022 mg GeO2 per litre of seawater). This treatmentis also economically very reasonable. In the currentconfiguration of our production system, we need 1.07 gof GeO2 to treat the spools, which will produce 60 t ofkelps. The present price of a 10 g bottle of powdered,
reagent grade GeO2 is US$93.40. Consequently, itcosts only US$9.99 for the production of 60 t of kelp,which makes this minimal, and biologically- andcost-effective use of GeO2 a very affordable treatmentin routine commercial scale aquaculture operations.
References
Chen LC, Edelstein T, McLachlan J (1969) Bonnemaisoniahamifera Hariot in nature and in culture. J Phycol 5:211–220
Chen LC, Edelstein T, Ogata E, McLachlan J (1970) The lifehistory of Porphyra miniata. Can J Bot 48:385–389
Chopin T, Robinson S, Sawhney S, Bastarache S, Belyea E,Shea R, Armstrong W, Stewart I, Fitzgerald P (2004)The AquaNet integrated multi-trophic aquaculture project:rationale of the project and development of kelp cultivationas the inorganic extractive component of the system. BullAqua Ass Can 104(3):11–18
Fritsch FE (1971) The structure and reproduction of the algae,vol 1. Cambridge University Press, Cambridge, 791 pp
Goldschmidt VM (1937) The principles of distribution of chem-ical elements in minerals and rocks. J Chem Soc 1937:655–675
Graham LE, Wilcox LW (2000) Algae. Prentice-Hall, Engle-wood Cliffs, 650 pp
Hopkin R, Kain JM (1978) The effects of some pollutants on thesurvival, growth and respiration of Laminaria hyperborea.Est Coast Mar Sci 7:531–553
Lewin J (1966) Silicon metabolism in diatoms. V. Germaniumdioxide, a specific inhibitor of diatom growth. Phycologia6:1–12
Markham JW, Hagmeier E (1982) Observations on the effectsof germanium dioxide on the growth of macro-algae anddiatoms. Phycologia 21:125–130
McLachlan J, Chen LC, Edelstein T (1971) The culture of fourspecies of Fucus under laboratory conditions. Can J Bot49:1463–1469
Merrill JE, Gillingham DM (1991) Bull kelp cultivationhandbook. National Coastal Resources Research andDevelopment Institute, Portland, 69 pp
Springer


















