
Effect of DNA on Thyroid-Hormone Binding by Specific Receptor Proteins from Rat-Liver Nuclei
8
Eur. J. Biochem. 114, 509-516 (1981) 0 FEBS 1981 Effect of DNA on Thyroid-Hormone Binding by Specific Receptor Proteins from Rat-Liver Nuclei Akira INOUE, Koichi NAKAGAWA, and Seiji MORISAWA Department of Biochemistry, Osaka City University Medical School (Received June 27/October 29, 1980) Influence of double-stranded native DNA on the binding of thyroid hormone, 3,5,3'-triiodo-~-thyronine, by the isolated nuclear receptors was studied and the following results were obtained. (1) The receptor-triiodo- thyronine complexes bound to DNA with moderate affinities. (2) DNA enhanced the hormone binding of the receptors. (3) The stimulatory DNA effect on triiodothyronine binding of the receptors was dependent on DNA concentration, showing its maximum at 30 pg/ml. (4) The increase in triiodothyronine binding was observed not only in the initial velocity but also in the plateau level which was attained after sufficient incubation time. (5) There were two types of specific receptors in the rat liver nuclear extract. The dissociation constants and the maximal binding capacities for triiodothyronine, which were determined by Scatchard plot analysis in the presence and absence of DNA, suggested that DNA exerted its effect through increasing binding capacity on one class of the receptors and through enhancing affinity for the hormone on the other class of the receptors. (6) Among various polynucleotides examined, the double-stranded eukaryotic DNA was most effective in enhancing the hormone binding by the receptors. These results indicate that the nuclear thyroid hormone receptors interact with double-stranded DNA in a specific manner and are induced to bind more thyroid hormone. We interpret these results as suggesting that a ternary complex of triiodothyronine, the receptor and DNA is formed in the cell nucleus in vivo, probably representing an intrinsic step in the hormone action. Possible physiological significance of this effect of DNA on the receptors is discussed. It has been shown that thyroid hormone, 3,5,3'-triiodo- thyronine, increases the synthesis of total RNA in liver cells [l], and the levels of specific messenger RNAs of clzU globulin in rat liver [2,3], carbamyl phosphate synthetase I in tadpole liver [4] and growth hormone in cultured pituitary cells [5-71. These effects of the hormone suggest its direct action on the cell nucleus. In fact, specific receptor activity has been shown in the nucleus of target cells [S - lo], characterized by its protein nature with a high affinity for the hormone and a limited number of binding sites [ l l - 151. Recently, the presence of the nuclear receptors has been visualized by autoradiography of the chromatin-associated thyroid hormone [ 161. The receptor proteins belong to the class of non-histone chromosomal proteins [l 1 - 151, and are extracted from the chromatin with moderately high salt concentrations (e.g. 0.4 M KC1) [ll, 121. The isolated receptors are DNA-binding proteins [17- 191 with a molecular weight of approximately Although the mechanism of thyroid hormone action in the cell nucleus is not clear at present, it is likely that ternary complexes between thyroid hormone, the receptor and DNA are formed during the hormone action. In the present study, we examined whether the DNA had any effect on the thyroid hormone binding by the isolated nuclear receptors. The results indicated that double-stranded DNA enhanced the triiodothyronine binding of the receptors. 50000- 70000 [I 5,201. ~ Enzyme. Deoxyribonuclease, DNase I (EC 3.1.21.1). MATERIALS AND METHODS Extraction of Thyroid Hormone Receptor Proteins Thyroid hormone receptor proteins were extracted from rat liver nuclei with a modified method of Samuels et al. [12]. Livers were removed from male Sprague-Dawley rats under anesthesia with ether and rinsed in ice-cold buffer A (0.25 M sucrose, 2 mM MgC12, 20 mM Tris/HCl, pH 7.6). The sub- sequent procedures were carried out at 0-2 "C. The livers (22 g) were finely minced with scissors and homogenized in 220 ml of buffer A by eight strokes with a motor-driven glass/ Teflon homogenizer. After filtration through four layers of cheese-cloth, the homogenate was centrifuged at 1500 x g for 10 min. The pellet was homogenized again with three strokes in 220 ml of the same solution and centrifuged as above. The crude nuclear pellet was successively washed twice with 150-ml portions of buffer A containing 0.5 Triton X-100, and once with buffer B [50 mM NaCI, 10 % (v/v) glycerol, 2 mM EDTA, 5 mM mercaptoethanol, 20 mM Tris/HCl, pH 7.61. Thyroid hormone receptor proteins were then extracted from the washed nuclear pellet by stirring for 30 min in 28 ml of 0.4 M KCI, 1.1 mM MgC12, 20 mM mercaptoethanol, 20 mM Tris/ HC1, pH 7.9. The mixture was centrifuged at 35000 x g for 60 min, and the supernatant dialyzed against 1-1 portions of buffer B with four changes, and centrifuged. The super- natant obtained was used as the nuclear thyroid hormone receptor extract. Protein concentration of the extract was 1.40 mg/ml.
-
Upload
akira-inoue -
Category
Documents
-
view
215 -
download
1
Transcript of Effect of DNA on Thyroid-Hormone Binding by Specific Receptor Proteins from Rat-Liver Nuclei

Eur. J. Biochem. 114, 509-516 (1981) 0 FEBS 1981
Effect of DNA on Thyroid-Hormone Binding by Specific Receptor Proteins from Rat-Liver Nuclei
Akira INOUE, Koichi NAKAGAWA, and Seiji MORISAWA
Department of Biochemistry, Osaka City University Medical School
(Received June 27/October 29, 1980)
Influence of double-stranded native DNA on the binding of thyroid hormone, 3,5,3'-triiodo-~-thyronine, by the isolated nuclear receptors was studied and the following results were obtained. (1) The receptor-triiodo- thyronine complexes bound to DNA with moderate affinities. (2) DNA enhanced the hormone binding of the receptors. (3) The stimulatory DNA effect on triiodothyronine binding of the receptors was dependent on DNA concentration, showing its maximum at 30 pg/ml. (4) The increase in triiodothyronine binding was observed not only in the initial velocity but also in the plateau level which was attained after sufficient incubation time. (5) There were two types of specific receptors in the rat liver nuclear extract. The dissociation constants and the maximal binding capacities for triiodothyronine, which were determined by Scatchard plot analysis in the presence and absence of DNA, suggested that DNA exerted its effect through increasing binding capacity on one class of the receptors and through enhancing affinity for the hormone on the other class of the receptors. (6) Among various polynucleotides examined, the double-stranded eukaryotic DNA was most effective in enhancing the hormone binding by the receptors. These results indicate that the nuclear thyroid hormone receptors interact with double-stranded DNA in a specific manner and are induced to bind more thyroid hormone. We interpret these results as suggesting that a ternary complex of triiodothyronine, the receptor and DNA is formed in the cell nucleus in vivo, probably representing an intrinsic step in the hormone action. Possible physiological significance of this effect of DNA on the receptors is discussed.
It has been shown that thyroid hormone, 3,5,3'-triiodo- thyronine, increases the synthesis of total RNA in liver cells [l], and the levels of specific messenger RNAs of clzU globulin in rat liver [2,3], carbamyl phosphate synthetase I in tadpole liver [4] and growth hormone in cultured pituitary cells [5-71. These effects of the hormone suggest its direct action on the cell nucleus. In fact, specific receptor activity has been shown in the nucleus of target cells [S - lo], characterized by its protein nature with a high affinity for the hormone and a limited number of binding sites [ l l - 151. Recently, the presence of the nuclear receptors has been visualized by autoradiography of the chromatin-associated thyroid hormone [ 161.
The receptor proteins belong to the class of non-histone chromosomal proteins [l 1 - 151, and are extracted from the chromatin with moderately high salt concentrations (e.g. 0.4 M KC1) [ l l , 121. The isolated receptors are DNA-binding proteins [17- 191 with a molecular weight of approximately
Although the mechanism of thyroid hormone action in the cell nucleus is not clear at present, it is likely that ternary complexes between thyroid hormone, the receptor and DNA are formed during the hormone action. In the present study, we examined whether the DNA had any effect on the thyroid hormone binding by the isolated nuclear receptors. The results indicated that double-stranded DNA enhanced the triiodothyronine binding of the receptors.
50000- 70000 [I 5,201.
~
Enzyme. Deoxyribonuclease, DNase I (EC 3.1.21.1).
MATERIALS AND METHODS
Extraction of Thyroid Hormone Receptor Proteins
Thyroid hormone receptor proteins were extracted from rat liver nuclei with a modified method of Samuels et al. [12]. Livers were removed from male Sprague-Dawley rats under anesthesia with ether and rinsed in ice-cold buffer A (0.25 M sucrose, 2 mM MgC12, 20 mM Tris/HCl, pH 7.6). The sub- sequent procedures were carried out at 0-2 "C. The livers (22 g) were finely minced with scissors and homogenized in 220 ml of buffer A by eight strokes with a motor-driven glass/ Teflon homogenizer. After filtration through four layers of cheese-cloth, the homogenate was centrifuged at 1500 x g for 10 min. The pellet was homogenized again with three strokes in 220 ml of the same solution and centrifuged as above. The crude nuclear pellet was successively washed twice with 150-ml portions of buffer A containing 0.5 Triton X-100, and once with buffer B [50 mM NaCI, 10 % (v/v) glycerol, 2 mM EDTA, 5 mM mercaptoethanol, 20 mM Tris/HCl, pH 7.61. Thyroid hormone receptor proteins were then extracted from the washed nuclear pellet by stirring for 30 min in 28 ml of 0.4 M KCI, 1.1 mM MgC12, 20 mM mercaptoethanol, 20 mM Tris/ HC1, pH 7.9. The mixture was centrifuged at 35000 x g for 60 min, and the supernatant dialyzed against 1-1 portions of buffer B with four changes, and centrifuged. The super- natant obtained was used as the nuclear thyroid hormone receptor extract. Protein concentration of the extract was 1.40 mg/ml.

510
Assuy for Thyroid Hormone Receptor Activity
Standard assay procedure for thyroid hormone receptor activity was as follows. The nuclear receptor extract (0- 120 pg protein) and ['2sI]triiodothyronine (0.05-0.10 pmol, Abbott Lab. North Chicago) with or without 200-fold excess of un- labelled triiodothyronine were incubated at 2 "C for 18 - 24 h in a total volume of 0.5 ml of buffer B. When necessary, prior to the incubation the mixtures were warmed at 30 "C for 10 min to permit dissociation of endogenous thyroid hormone from the receptors. At the end of incubation, 3.5 ml of cold buffer B was added to the mixture, and filtered through a cellulose ester membrane (Millipore HAWP 02500, 0.45 pm) at a filtra- tion rate of about 4 m1/20 s under suction at 2°C. The tube was rinsed twice with 3.5 ml portions of buffer B and the contents were poured on the membrane in a similar manner as above. After washing the membrane and the sampling manifold well thoroughly with two 7-ml portions of buffer B, the membrane was placed in a test tube for counting radio- activity with a gamma spectrometer. Correction was made for nonspecific triiodothyronine binding which was deter- mined in the presence of 200-fold excess of unlabelled hor- mone. When necessary, non-specific absorption of free [1251]- triiodothyronine to the cellulose ester membrane was also determined in the absence of the nuclear receptor extract and corrected as blanks.
Digestion of Rut Livcr Nuclei with DNuse I
Rat liver crude nuclear pellet, isolated as described above, was washed three times with 20vol. each of 0.5% (w/w) Triton X-100 in buffer A, three times with 10 mM NaCl in buffer A, and finally suspended in the same 10 mM NaCl solution by a loose Teflon homogenizer. After the aggregates were removed by filtration through a layer of cheese-cloth, 0.8-ml portions of the nuclear suspension (7.7 A260 units) were mixed with 0.2 ml each of DNase I solution ( 5 pg of bovine pancreatic enzyme, Worthington) which was dissolved in the same buffer, and incubated at 30°C for the time periods indicated. The reaction was terminated by chilling the mix- tures to 0 "C followed by addition of 50 p1 of 100 mM EDTA and the mixtures centrifuged at 2000 rev./min for 20 min. The supernatants were saved, and the pellets were washed once with 1 ml of the incubation buffer containing 5 mM EDTA and suspended in 0.2ml of the same solution. The supernatants (0.2 ml) and the pellet suspensions (0.2 ml) were assayed for triiodothyronine receptor activity with [12sI]- triiodothyronine (0.1 pmol) in a total volume of 1 ml.
Dissociation Kinetics qf Receptor-Hormone Complexes
The receptor extract (480 pg protein) was incubated with 0.15 pmol of [12sl]triiodothyronine (final concentration 0.3 nM) in 0.5 ml of buffer B at 3 C for I8 h to allow com- plex formation between the receptor and the hormone. The mixture was then applied to a Sephadex G-25 column (bed volume 2.0 ml, 0.6 x 7.0 cm, fine, Pharmacia Fine Chemicals) at 2 'C which was preequilibrated with buffer B. The column was eluted with the same solution and the effluent of total 1.85 ml, which contained the receptor-triiodothyronine com- plexes, was collected. Under this condition, free hormone was strongly absorbed to the column and not recovered with buffer B [12,21].The solution of the receptor-triiodothyronine complexes was diluted to6.4ml with buffer B, and the triiodo- thyronine concentration was brought to 0 .2pM with un-
labelled hormone. After the mixture was allowed to stand at 0 ° C or 3°C for the time periods indicated, 0.5-ml por- tions were taken and filtered to determine the '"1 radio- activity which remained bound to the receptors.
DNA-Affinity Column Chromutogruphy of the Receptor-Hormone Complexes
DNA-Sephadex G-25 was prepared as described previ- ously [22]. The nuclear receptor extract (1.5 mg protein) and [12sl]triiodothyronine (1.5 pmol) were incubated at 2 "C for 18 h in 3.0 ml of buffer B. The mixture was applied at a flow rate of 1.5 ml/h on a DNA-Sephadex affinity column (bed volume 7.0 ml, 1.6 x 3.5 cm, DNA content 5.2 mg) which was preequilibrated with buffer B. After washing the column with the same buffer solution, the proteins retained on the column were eluted at a flow rate of 7.0 mljh first with a linear concentration gradient of NaCl (50- 300 mM), and then by a stepwise elution with 0.5 M NaCl in lo:/; (vjv) glycerol, 2 mM EDTA, 5 mM mercaptoethanol, 20 mM Tris/ HCI, pH 7.6. Fractions of 2.3 ml were collected, and 0.5 mi of each fraction was filtered through cellulose ester mem- branes to determine the receptor-bound 1251 in a similar manner as described above. Direct counting of the fractions gave an additional '"I peak at the 8th fraction due to 1251-ion which was present as a contaminant in the commercial radio- active thyroid hormone.
RESULTS
Release of Thyroid Hormone Receptor Activity from Rut Liver Nuclei by DNuse I Digestion
The presence of triiodothyronine receptors in the cell nucleus can be shown by DNase 1 digestion of the isolated nuclei. When rat liver nuclei were treated with DNase I , the chromatin was digested to fragments and recovered in the reaction medium. The release of the chromatin fragments proceeded in proportion to incubation time (Fig. 1). Coupled to it, the thyroid hormone receptor activity was also released from the nuclei; in an 8-min digestion, about 72 "/, of the total nuclear activity was recovered in the supernatant of reaction mixture. When hormone binding of the supernatant receptors was roughly examined by Scatchard plot analysis, an equili- brium dissociation constant of about 0.1 nM was estimated. The results indicates that specific triiodothyronine receptors are present in the nuclei constituting the components of chromatin. This observation is a further support of the con- clusions by others derived from several other lines of experi- ment [8 - 151.
Equilibrium Binding oj Hormone to Nuclear Receptors and Dissociation Kinetics of the Receptor-Hormone Complexes
The nuclear receptors were extracted from the washed rat liver nuclei with a buffer solution containing 0.4 M KC1, and subjected to the analyses of their equilibrium hormone- binding and dissociation kinetics of the receptor-hormone complexes.
The equilibrium binding of thyroid hormone to the re- ceptors was examined by Scatchard plot analysis. The re- ceptors were incubated with [12sI]triiodothyronine at 50 mM NaCl for 18 h at 2 C, at which time the receptor-hormone binding reached its plateau level. To assess the receptor-

51 1
0 4 8 I f
0 4 8 I n c u b a t i o n t i m e (min)
Fig. 1 . Release of thyroid hormone receptor activity from rat liver nuclei by DNase I digestion. Rat liver nuclei (7.7 A260 units) in 1 ml of 10 mM NaCI, 0.25 M sucrose, 2 mM MgC12, 20 mM Tris/HCl. pH 7.6 were digested with 5 pg of DNase I at 30°C for the time periods indicated. After termination of the reaction by adding 50p1 of 100mM EDTA, the mixtures were centrifuged. Aliquots of the resultant supernatant (0) and the pellet (0) fractions were assayed for triiodothyronine binding activity as described in Materials and Methods. The digestion of the chromatin was monitored by the DNA which was rendered soluble in 7 % perchloric acid (x-- x ). Incubation in the absence of DNase I was carried out as controls, and the supernatant receptor activity was determined (A). Without the enzyme, the acid-soluble DNA was 3.5% and 4.0% of total at 3 min and 8 min, respectively. Each point is the average of duplicate determinations. Non-specific triiodothyronine bind- ing, measured in the presence of 200-fold molar excess of unlabelled hormone, was corrected at each point
bound triiodothyronine in each assay mixtures, the samples were filtered on cellulose ester membranes and the lZ5I radio- activity retained by the membrane was counted. This filtra- tion method collects proteins on the membrane separating protein-bound triiodothyronine from free hormone. By Scat- chard plot analysis, three classes of thyroid hormone binding activities were detected in the nuclear extract (Fig.2). One of them is considered attributable to non-specific receptors because of its low affinity for the hormone (Kd = 1.03 pM) and an unlimited number of hormone binding sites. The other two sets of the receptors, regarded as specific receptors, had apparent dissociation constants ( K d ) of 0.086 nM and 0.70 nM with limited hormone binding capacities of 0.132 and 0.25 pmol/mg protein, respectively (Fig. 2).
Triiodothyronine binding of the receptors was characterized next by dissociation kinetics where the displacement of recep- tor-bound ['2sI]triiodothyronine by unlabelled hormone was followed (Fig. 3). After incubation of the receptor extract with [1251]triiodothyronine, the receptor-hormone complexes were separated from free hormone by a Sephadex G-25 column. The isolated complexes were then allowed to stand at 0 'C or 3 "C in the presence of a large excess of unlabelled triiodothyronine. Fig. 3 shows the displacement time course at the two temperatures, demonstrating that the complexes dissociate linearly with time, or that the dissociation velocity is not influenced by the concentration of recept~r-['~~I]tri- iodothyronine complexes. Time of half dissociation extra- polated from the curves was 26 h at 3 "C or 36 h at 0 "C. These results indicate that the receptor-thyroid hormone complexes, once formed, dissociate at relatively slow rates at low temperatures, and can be subjected to DNA-affinity column chromatography at 0 - 3 ' C to analyse their DNA- binding property as described below.
Fig. 2. Scatchard plot analysis of triiodothyronine binding by rat liver nuclear receptor. The nuclear extract (120 pg protein) was incubated in 0.5 ml of buffer B with various amounts of ['251]triiodothyronine -
M). After an 18-h incubation at 2 "C, the mixtures were filtered on cellulose ester membranes to determine the receptor-bound ['251]triiodo- thyronine as described in Materials and Methods. Free ['251]triiodo- thyronine non-specifically absorbed to the membrane was also deter- mined at each point in the absence of the receptor extract and corrected for the net protein-bound hormone. The insert represents the same plots with different scales
2
I'L 0 0 10 Time (h) 20 30
Fig. 3. Dissociation kinetics of receptor-hormone comple-yes. After the nuclear receptor extract (480 pg protein) and ['251]triiodothyronine (0.3 nM) were incubated in 0.5 ml of buffer B, the receptor-hormone complexes formed were separated from the unbound hormone on a small Sephadex G-25 column (2-ml bed volume, 0.6 x 7.0 cm) as described in Materials and Methods. The complexes were then incubated at 0 "C (0) or 3 "C (0) in 6.4 ml buffer B in the presence of a large excess of unlabelled hormone (0.2 pM). At the time indicated, aliquots (0.5 ml) of the sample were filtered to determine lZ5I radioactivity which remained bound to the receptors
DNA-Affinity Column Chromatography of Receptor-Hormone Complexes
The nuclear receptors were analysed for their DNA-binding property by use of an affinity column of double-stranded native DNA covalently attached to Sephadex G-25. The
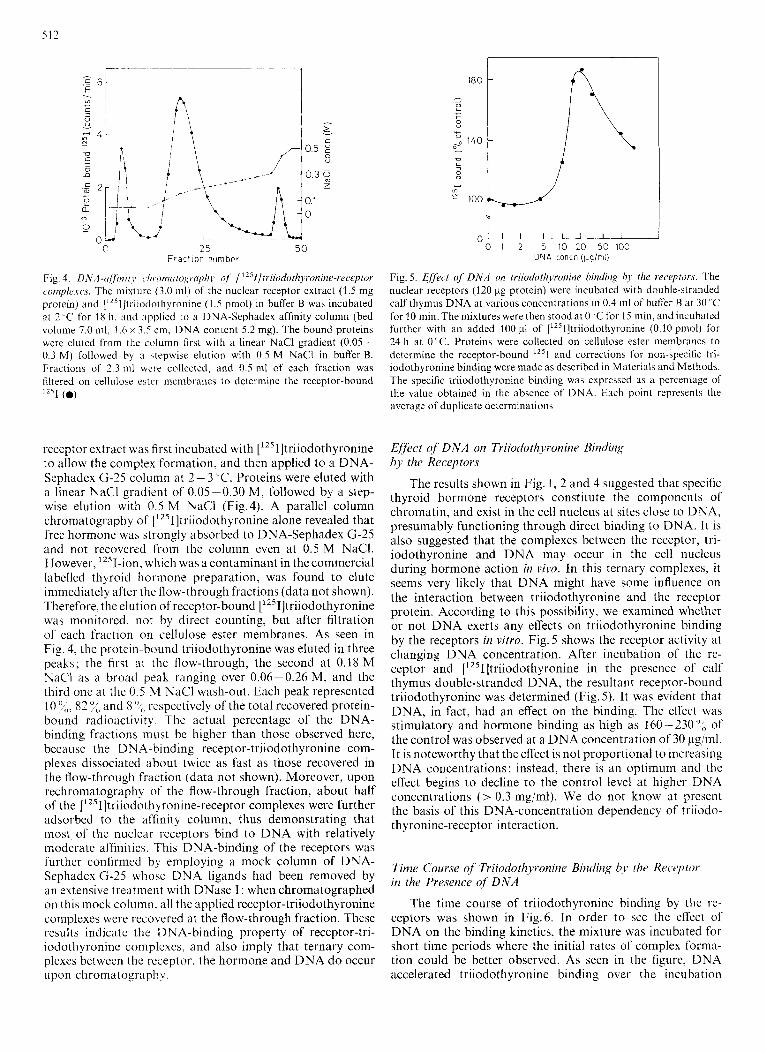
512
- L .; 6 ,
U c 3 0 0
0
0 0 0 25 5 0
Fraction number
Fig, 4. D N A -(iff it7 i r ~ (.I7 ronxi cogrupli)~ of [ " 1 tr iiodothyron ine-receptor comple.ies. The mixturc (3.0 ml) of the nuclear receptor extract (1.5 mg protein) and ["'ll]triiodothyronine (1.5 pmol) in buffer B was incubated at 2 ,C for 18 h. m d applied to a DNA-Sephadex affinity column (bed volume 7.0 ml, I .6 x 3.5 cm, DNA content 5.2 mg). The bound proteins were eluted from thc column first with a linear NaCl gradient (0.05- 0.3 M) followed by a .tepwise elution with 0.5 M NaCl in buffer B. Fractions of 2.3 ml were collected, and 0.5 mi of each fraction was filtered on cellulose cstc'r membranes to determine the receptor-bound 1 ~ ~ 1 (0)
receptor extract was first incubated with ['251]triiodothyronine to allow the complex formation, and then applied to a DNA- Sephadex G-25 column at 2 - 3 "C. Proteins were eluted with a linear NaCl gradient of 0.05-0.30 M, followed by a step- wise elution with 0.5 M NaCl (Fig.4). A parallel column chromatography of [12sI]triiodothyronine alone revealed that free hormone was strongly absorbed to DNA-Sephadex G-25 and not recovered from the column even at 0.5 M NaCI. However, 12sI-ion, which was a contaminant in the commercial labelled thyroid hormone preparation, was found to elute immediately after the flow-through fractions (data not shown). Therefore, the elution of receptor-bound ['251]triiodothyronine was monitored. not by direct counting, but after filtration of each fraction on cellulose ester membranes. As seen in Fig. 4, the protein-bound triiodothyronine was eluted in three peaks; the tirst at the flow-through, the second at 0.18 M NaCl as a broad peak ranging over 0.06-0.26 M, and the third one at the 0.5 M NaCl wash-out. Each peak represented 10 %,, 82 :,< and 8 O ~ o respectively of the total recovered protein- bound radioactivity. The actual percentage of the DNA- binding fractions must be higher than those observed here, because the DNA-binding receptor-triiodothyronine com- plexes dissociated about twice as fast as those recovered in the flow-through fraction (data not shown). Moreover, upon rechromatography of the flow-through fraction, about half of the ['251]triiodothyronine-receptor complexes were further adsorbed to the affinity column, thus demonstrating that most of the nuclear receptors bind to DNA with relatively moderate affinities. This DNA-binding of the receptors was further confirmed by employing a mock column of DNA- Sephadex G-25 whose DNA ligands had been removed by an extensive treatment with DNase I : when chromatographed on this mock column, all the applied receptor-triiodothyronine complexes were recovered at the flow-through fraction. These results indicate the DNA-binding property of receptor-tri- iodothyronine complexes. and also imply that ternary com- plexes between the receptor, the hormone and DNA d o occur upon chromatography.
I 0 1 2 5 10 20 50 100
DNA concn (Kgiml)
Fig. 5. Effect of D N A on triiodothyroninc. binding by tlzc receptors. The nuclear receptors (120 pg protein) were incubated with double-stranded calf thymus DNA at various concentrations in 0.4 ml of buffer B at 30 C for 10 min. The mixtures were then stood at 0 'C for 15 min, and incubated further with an added 100 pl of [1Z51]triiodothyronine (0.10 pinol) for 24 h at 0°C. Proteins were collected on cellulose ester membranes to determine the receptor-bound '*'I and corrections for non-specific tri- iodothyronine binding were made as described in Materials and Methods. The specific triiodothyronine binding was expressed as a percentage of the value obtained in the absence of DNA. Each point reprebents the average of duplicate determinations
Efec t of DNA on Triiodothvronine Bindirig by the Receptors
The results shown in Fig. I , 2 and 4 suggested that specific thyroid hormone receptors constitute the components of chromatin, and exist in the cell nucleus at sites close to DNA, presumably functioning through direct binding to DNA. It is also suggested that the complexes between the receptor, tri- iodothyronine and DNA may occur in the cell nucleus during hormone action in vivo. In this ternary complexes, it seems very likely that DNA might have some influence on the interaction between triiodothyronine and the receptor protein. According to this possibility, we examined whether or not DNA exerts any effects on triiodothyronine binding by the receptors in vitro. Fig. 5 shows the receptor activity at changing DNA concentration. After incubation of the re- ceptor and [1251]triiodothyronine in the presence of calf thymus double-stranded DNA, the resultant receptor-bound triiodothyronine was determined (Fig. 5). It was evident that DNA, in fact, had an effect on the binding. The effect was stimulatory and hormone binding as high as 160-2304" of the control was observed at a DNA concentration of30 pg/ml. It is noteworthy that the effect is not proportional to increasing DNA concentrations; instead, there is an optimum and the effect begins to decline to the control level at higher DNA concentrations (> 0.3 mg/ml). We do not know at present the basis of this DNA-concentration dependency of triiodo- thyronine-receptor interaction.
Time Course of Triiodothyroninc Binding b.y the Receptor in the Presence of DNA
The time course of triiodothyronine binding by the re- ceptors was shown in Fig.6. In order to see the effect of DNA on the binding kinetics, the mixture was incubated for short time periods where the initial rates of complex forma- tion could be better observed. As seen in the figure, DNA accelerated triiodothyronine binding over the incubation
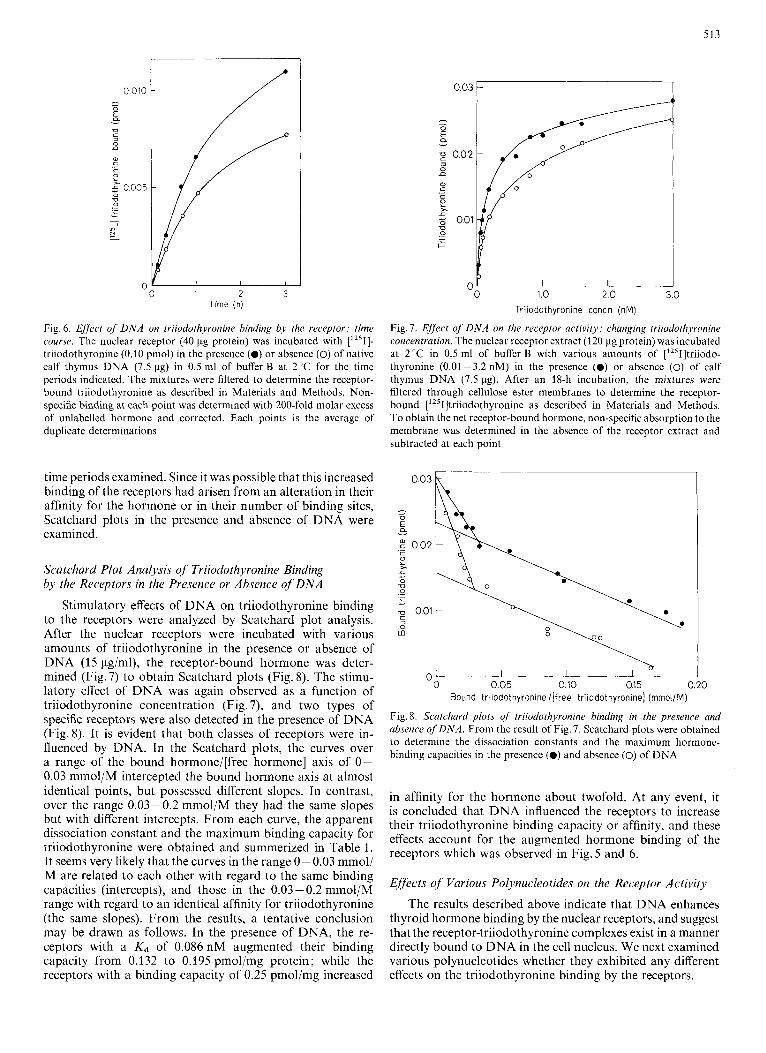
513
Time (h)
Fig. 6. Effect of D N A on triiodothyronine binding by the receptor: time course. The nuclear receptor (40 pg protein) was incubated with [1251]- triiodothyronine (0.10 pmol) in the presence (0) or absence (0) of native calf thymus DNA (7.5 pg) in 0.5 ml of buffer B at 2°C for the time periods indicated. The mixtures were filtered to determine the receptor- bound triiodothyronine as described in Materials and Methods. Non- specific binding at each point was determined with 200-fold molar excess of unlabelled hormone and corrected. Each points is the average of duplicate determinations ~*
time periods examined. Since it was possible that this increased binding of the receptors had arisen from an alteration in their affinity for the hormone or in their number of binding sites, Scatchard plots in the presence and absence of DNA were examined.
Scatchard Plot Analysis of Triiodothyronine Binding by the Receptors in the Presence or Absence of DNA
Stimulatory effects of DNA on triiodothyronine binding to the receptors were analyzed by Scatchard plot analysis. After the nuclear receptors were incubated with various amounts of triiodothyronine in the presence or absence of DNA (15 pg/ml), the receptor-bound hormone was deter- mined (Fig. 7) to obtain Scatchard plots (Fig. 8). The stimu- latory effect of DNA was again observed as a function of triiodothyronine concentration (Fig. 7), and two types of specific receptors were also detected in the presence of DNA (Fig. 8). It is evident that both classes of receptors were in- fluenced by DNA. In the Scatchard plots, the curves over a range of the bound hormone/[free hormone] axis of 0- 0.03 mmol/M intercepted the bound hormone axis at almost identical points, but possessed different slopes. In contrast, over the range 0.03-0.2 mmol/M they had the same slopes but with different intercepts. From each curve, the apparent dissociation constant and the maximum binding capacity for triiodothyronine were obtained and summerized in Table 1. It seems very likely that the curves in the range 0- 0.03 mmol/ M are related to each other with regard to the same binding capacities (intercepts), and those in the 0.03 - 0.2 mmol/M range with regard to an identical affinity for triiodothyronine (the same slopes). From the results, a tentative conclusion may be drawn as follows. In the presence of DNA, the re- ceptors with a Kd of 0.086 nM augmented their binding capacity from 0.132 to 0.195 pmol/mg protein; while the receptors with a binding capacity of 0.25 pmol/mg increased
" 0 1.0 2.0 3.0 Triiodothyronine concn (nM)
Fig. 7 . Effect of D N A on the receptor activity: changing triiodothyronine concentration. The nuclear receptor extract (120 pg protein) was incubated at 2°C in 0.5 ml of buffer B with various amounts of ['Z51]triiodo- thyronine (0.01 -3.2 nM) in the presence (0) or absence (0) of calf thymus DNA (7.5 pg). After an 18-h incubation, the mixtures were filtered through cellulose ester membranes to determine the receptor- bound [1z51]triiodothyronine as described in Materials and Methods. To obtain the net receptor-bound hormone, non-specific absorption to the membrane was determined in the absence of the receptor extract and subtracted at each point
0.03
- - 0
a E - .- E 0.02 c 2 x L
0 73 c
.- .- L c
72 0.01
m 3 0
n "0 0.05 0.10 a15 0.20
Bound triiodothyroninel[free triiodothyronine] (mrnol/M)
Fig. 8. Scatchard plots of triiodothyronine binding in the presence and absence of DNA. From the result of Fig. 7, Scatchard plots were obtained to determine the dissociation constants and the maximum hormone- binding capacities in the presence (0) and absence (0) of DNA
in affinity for the hormone about twofold. At any event, it is concluded that DNA influenced the receptors to increase their triiodothyronine binding capacity or affinity, and these effects account for the augmented hormone binding of the receptors which was observed in Fig. 5 and 6.
Effects of Various Polynucleotides on the Receptor Activity
The results described above indicate that DNA enhances thyroid hormone binding by the nuclear receptors, and suggest that the receptor-triiodothyronine complexes exist in a manner directly bound to DNA in the cell nucleus. We next examined various polynucleotides whether they exhibited any different effects on the triiodothyronine binding by the receptors.

514
Table 1. Diw)cicition constants and maximum capacities ,for triiodo- thyronine binding oj rui liver nuclear receptors in the presence or absence of D N A Scatchard plot analysis was performed for triiodothyronine binding by the nuclear receptors in the presence or absence of DNA (Fig. 8). In Fig. 8, two classes of receptor., are recognized both in the presence and absence of DNA. For each class of receptors, the apparent dissociation constants of triiodothyronine binding and the maximum numbers of hormone binding sites were obtained from the slopes and the bound hormone axis intercepts
Addition Bound hormone, Dissociation Maximum of DNA [free hormone] constant, number of
Kd binding sites
mmol M nM pmo~/mg protein
- DNA 0 - 0 03 0 70 0 258 0 03-0 20 0 086 0 132
+ DNA 0 0 0 3 0 33 0 258 003-020 0 084 0 195
Table 2. 6ffec.i o/ 1;uriou.r polynuclwtides on triiodothyronine hinding of rat liwr nuclrur receptors The nuclear receptor extract (60 pg protein) was incubated with ['2sI]- triiodothyronine (0.05 pmol) at 3 ' C for 20 h in 0.5 ml of buffer B which contained one o f the polynucleotides listed (15 pg). After the incubation, the mixtures were filtered to determine the receptor-bound [12sI]triiodo- thyronine ;is described in Materials and Methods. Each values is the average of duplicate determinations. Heat-denatured DNA was prepared by incubating the native calf thymus DNA at 100'C for 5 min followed by rapid cooling on ice. Polyadenylated RNA was prepared from rat liver according to a modified method of [28]. E. coli D N A and tRNA were purchascd from Sigma; (dT)lo and yeast RNA were from P-L Biochemicals
Polynucleotidc Bound tri- Stimulation iodothyronine of binding
/ o 0
~~~ ~ ~ ~ ~~ ~~~ ~
- Control (minus polynucleotide) I00
Native calf thymu5 DNA 166 66 C. c.oli DNA 132 32 Heat-dcnaturcd c d f Ihymus DNA 120 20 (dT)ro 109 9 Ral liver poly(A)-rich R N A 110 10 Yeast RNA 108 8 E. coli tRNA 108 8
As seen on Table 2, all the polynucleotides examined enhanced the binding. However. DNAs had much larger effects than RNAs. I t was also apparent that native DNA revealed threefold higher stimulation than heat-denatured DNA. Thus, the receptors prefer double-stranded DNA to RNA, or to single-stranded (coil-tangled) DNA. Furthermore, there seems to be a sequence-specific recognition of DNA by the receptors. since DNA from a higher organism (calf) was more effective than that from a prokaryote (Escherichia roli). These results further support the possibility that thyroid hor- mone receptors have an intrinsic property of functioning through direct and specific binding to DNA. Interestingly, DNA sequence-recognition by the receptors is also im- plied by the DNA-affinity chromatography shown in
Fig. 4, where ['251]triiodothyronine-receptor complexes were eluted as a broad peak from the affinity column (by a linear concentration gradient). The broadness of the peak may indicate that there occurred differential inter- actions between DNA and the receptors presumably arising from specific or nonspecific bindings of the receptors to various DNA sequences.
DISCUSSION
In the present work, specific thyroid hormone receptors were demonstrated in rat liver cell nuclei by digestion with DNase I, and the extracted receptors were shown to possess a DNA-binding property by use of a DNA-affinity column. These results support those already reported by others [8 - 10, 17-19], and gave us a basis for assuming that the specific receptors which constitute the components of chromatin, exist in the cell nucleus in vivo at sites close to DNA, presum- ably functioning through direct binding to DNA. On this possibility, double-stranded DNA was examined to see whether it had any effect on the thyroid hormone binding of the receptors, and the DNA was, in fact, found to influence the receptors to increase their hormone binding activity in vitro.
In this study, determination of the receptor-bound thyroid hormone was achieved by a membrane filtration method, because this method was found to give an accurate estimation up to a protein amount of 120 pg/assay tube. In contrast to this, the conventional procedure employing small Sephadex (3-25 columns did not satisfy a linear relationship between the receptor activity and protein concentration particularly in the range of 0-25 pg protein/ml (results will be published elsewhere).
In order to examine the DNA-binding affinity of the recep- tor-hormone complexes, DNA-Sephadex G-25 was used as an affinity adsorbent [22]. Dextran gel, Sephadex (3-25, absorbs thyroid hormone very tightly, and owing to this property it has been used to separate receptor-bound hormone from unbound hormone [12]. This seems to explain why none of the free triiodothyronine was eluted from the DNA-Sephadex column when mixtures of the receptors and the hormone were chromatographed (Fig. 4). DNA column chromatog- raphy revealed that most of the receptor-triiodothyronine complexes (more than 90 7; of the hormone-binding activity) had affinities for DNA. Since DNA binding was observed with the complexes of the receptors and the hormone, it was implied that a ternary complex was formed between triiodo- thyronine, the receptor and DNA in the affinity column.
The present work demonstrated that DNA caused the receptors to bind more hormone; in the presence of DNA, the receptors bound about twofold more hormone. It is not clear at present how DNA increases the receptors' hormone- binding activity. However. it is likely that the receptors bind to DNA and subsequently alter their conformation, 't 1 struc- ture of the triiodothyronine-binding site. If this is the case, this modification of the binding sites may be due to an allosteric-like effect of DNA. The geometrical separation of the binding sites for triiodothyronine and DNA may be able to be conceived from the known three-dimensional structure of prealbumin. Prealbumin, a thyroid hormone-binding pro- tein in plasma, has been shown to possess two hormone- binding sites per molecule [23] and a molecular weight similar to that of the nuclear receptor proteins [15,20,24]. Moreover, at a separate locus from the hormone-binding sites, pre-

515
albumin also has a structure where double-stranded DNA can fit with a structural complementarity (although it has no known function that may involve DNA binding, nor a de- tectable DNA-binding activity) [25]. Hence, the nuclear re- ceptor protein resembles prealbumin in terms of (a) the molecular weight, (b) thyroid hormone-binding activity and (c) the possession of a putative protein-DNA interaction site, and these resemblances have suggested that they may belong to a family whose members have evolved from a common ancestor [25]. Therefore, it is presumed that the hormone- binding and DNA-binding sites of the nuclear receptor are located at separate loci, and that DNA exerts its influence on triiodothyronine binding through an allosteric-like mech- anism.
As mentioned above, most of the nuclear receptor-tri- iodothyronine complexes had affinities for DNA. However, the possibility should be pointed out that the observed DNA- binding might simply be an experimental feature in vitro, not relevant to the physiological hormone action. On the other hand, it is worth emphasizing that, under the same buffer conditions as for DNA-affinity chromatography, the receptor- hormone complexes were also adsorbed to a DEAE-Sephadex column (data not shown): that is, the complexes bind not only to DNA-Sephadex, an anionic biopolymer, but also to DEAE-Sephadex, a synthetic polymer containing cationic groups. Since DNA is the biological substance, and both the receptor proteins and DNA are found in the cell nucleus constituting the chromatin, the observed DNA binding seems to reflect an intrinsic property of the receptors. In support of this, the hormone binding of the receptors was shown to be influenced to various degrees depending on the polynucleo- tides employed (Table 2). This experiment also suggested that the receptors recognize the conformation and the base sequences of DNA (Table 2). Hence, it is very likely that the receptors do exist in association with DNA in the cell nucleus.
We do not know at present the physiological significance of the effect of DNA on triiodothyronine binding of the receptors. However, it seems probable that the biological activity of the receptors can be fully manifested when they are in association with DNA, and gain the ability to bind more thyroid hormone. This process increases the cellular concentration of the ternary complex which is, as mentioned above, formed between triiodothyronine, the receptor and DNA. Provided that thyroid hormone regulates the tran- scriptional process at a level of DNA, the concentration of this ternary complex in the cell nucleus may become a deter- minant for the levels of mRNA productions.
Scatchard plot analysis demonstrated the presence of two sets of specific thyroid hormone receptors in the rat liver nuclear extract (Fig. 2 and 8). Besides the Scatchard analysis, Bio-Gel P-150 filtration, DEAE-Sephadex column chromatog- raphy, and polyacrylamide gel electrophoresis in a neutral pH, all revealed the presence of two sets of receptors in the nuclear extract (unpublished results). However, in either procedure, the fractionated receptor activity was found to be severely reduced and did not allow one to try to correlate with activity demonstrated in the Scatchard plot analysis. Hetero- geneity of the receptors have been reported by others in rat liver nuclear extracts by means of binding analyses, Sephadex G-300 gel filtration [15,26], or ion-exchange chromatography [15,27]. It should also be mentioned that, for an unknown reason, we sometimes observed only one class of the receptors whose Kd was 0.7 nM.
Table 1 revealed that there were 0.39 pmol of triiodo- thyronine-binding sites/mg of the extractable proteins, and
that the number of binding sites whose activity increased in the presence of DNA was 0.32 pmol/mg protein [0.258 and (0.195-0.132) pmol/mg protein, Table I]. A gram of liver tissue yielded 1.32 mg of DNA in the final nuclear pellet and the same amount (1.32 mg) of proteins in the 0.4 M KCI receptor extract. If we take a value of 8 pg for the DNA content in a single rat liver nucleus [29], the number of extractable binding sites can be estimated from Avo- gadro’s number to be 1870 sites/nucleus, and those which were stimulated by DNA to be 1540 sites/nucleus. Since half of the specific sites are already occupied by endogenous tri- iodothyronine [30], and about 35 % of the occupied sites are expected to become free after an 18-h incubation (Fig. 3 and see the incubation condition described the legend of Fig. 7), one can roughly estimate that the extractable triiodothyronine binding capacity is 2800 sites/nucleus. This value is fairly comparable to that (4400 sites/nucleus) observed with the [1251]triiodothyronine injection study in vivo [30,31]. Like- wise, the extractable triiodothyronine-binding capacity which is stimulated by DNA is also calculated to be 2300 sites/ nucleus. This estimated hormone-binding capacity favours the suggestion of the physiological significance : the nuclear receptors seem to interact with specific DNA sequences at limited loci of the genome.
Under the present assay system, the effect of DNA was examined at a protein : DNA ratio (w/w) of 15 : 1 (Fig. 7 and 8, Table 1). This ratio contrasts to that of their amount recovered from the cell nuclei (1 : 1). Hence, the concentration of effective DNA required for the stimulation was less than that expected. This difference may be explained partly by the accessibility of the DNA; DNA employed in the assay was the naked DNA and freely accessible for the receptors while DNA in the cell nucleus is in the form of chromatin with bound histones and non-histone proteins and a considerable portion of DNA may not always be involved in the receptors’ specific asso- ciation.
The present study thus demonstrated an interaction be- tween the nuclear thyroid hormone receptors and the double- stranded DNA, supporting the notion that the receptors are very likely to function through direct binding to DNA in the cell nucleus in vivo.
REFERENCES 1. Towle, H. C., Dillmann, W. H. & Oppenheimer, J. H. (1979) J .
2. Kurtz, D. T., Sippel, A. E. & Feigelson, P. (1976) Biochemistry, 15,
3. Roy, A. K., Schiop, M. J. & Dowbenko, D. J. (1976) FEBS Lett.
4. Mori, M., Morris, S. M., Jr & Cohen, P. P. (1979) Proc. Nut1 Acad.
5. Martial, J. A., Baxter, J. D., Goodman, H. M. & Seeburg, P. H.
6. Seo, H., Vassart, G., Brocas, H. & Refetoff, S. (1977) Proc. Nail
7. Shapiro, L. E., Samuels, H. H. & Yaffe, B. M. (1978) Proc. Nut1
8. Oppenheimer, J. H., Koerner, D., Schwartz, H. L. & Surks, M. I.
9. Samuels, H. H. & Tsai, J. S. (1973) Proc. Nut1 Acad. Sci. USA, 70,
Biol. Chem. 254, 2250-2257.
1031-1036.
64, 396 - 399.
Sci. USA, 76, 3179-3183.
(1977) Proc. Nut1 Acad. Sci. USA, 74, 1816-1820.
Acad Sci. USA, 74, 2054-2058.
Acad. Sci. USA, 75,45 - 49.
(1972) J . Clin. Endocrinol. Metub. 35, 330-333.
3488 - 3492. 10. DeGroot, L. J. & Strausser, J. L. (1974) Endocrinology, 95, 74-83. 11. Surks, M. I., Koerner, D., Dillmann, W. & Oppenheimer, J. H. (1973)
12. Samuels, H. H., Tsai, J . S., Casanova, J. & Stanley, F. (1974) J . J . Biol. Chem. 248,7066-7072.
Clin. Invest. 54, 853-865.

13. Thomopoulos, P., Dastugue, B. & Defer, N. (1974) Biochem. Bio-
14. Torresani, J. & DeGroot, L. J . (1975) Endocrinology, 96, 1201 - 1209. 15. Latham, K. R., Ring, J . C. & Baxter, J. D. (1976) J . Biol. Chem.
16. Torresani, J., Gharbi, J., Lissitzky, S., Mirre, C., Delgrossi, M.-H. & Stahl, A . (1979) Biochern. Biophys. Res. Comrnun. 86, 371 -378.
17. MacLeod, K . M. & Baxter, J. D. (1975) Biochem. Biophys. Res. Commun. 62. 577-583.
18. MacLcod, K . M. Ri Baxtcr, .I. D. (1976) J . B i d Chern. 251, 7380- 7387.
19. Torresdni, J.. Ansclrnet. A . & Wahl, R. (1978) Mol. Cell. Endocrinol.
20. Snrks, M. I., Koerner, I>. & Oppenheimer, J. H. (1975) J . Clin.
21. Green, W. L. (1972) J . Clirornaiogr. 72, 83-91. 22. Allfrey, V. G. Ri lnoue. A. (1978) Meihods Cell B id . 17, 253-270.
phyvs. Res. Comniun. 58. 499 - 506.
251, 7388-7397.
9, 321 - 333.
Invest. 55. 50- 60.
23. Ferguson, R. N., Edelhoch. H., Saroff, H. A. & Robbins, J. (1975)
24. Kanda, Y., Goodmann, Dew. S., Canfield, R. E. & Morgan, F. J.
25. Blake, C. C. F. & Oatley, S. J . (1977) Nature /Lond.j 268, 115- 120. 26. Eberhardt, N. L., Ring, J. C., Johnson, L. K., Lathan, K . R., Apri-
letti, J. W., Kitsis, R. N. & Baxter, J. D. (1979) Proc. Natl Acad. Sci. U S A , 76, 5005 - 5009.
27. Silva, E. S., Astier, H., Thakare, U., Schwartz, H. L. & Oppenheimer, J. H. (1 977) J. Biol. Chem. 252.6799 - 6805.
28. Miller, W. L. & McCarthy, B. J. (1979) J . Biol. Chern. 254, 742-748. 29. Leuchtenberger, C., Vendrely, R. & Vendrely. C. (1951) Proc. Natl
30. Oppenheimer, J. H., Schwartz, H. L. & Surks, M. I. (1974) Endo-
31. Oppenheimer, J. H. (1979) Science (Wash. DC) 203, 971-979.
Biochemistry, 14, 282 - 289.
(1974) J . Biol. Chem. 249, 6796-6805.
Acad. Sci. U S A , 37, 33.
crinology, 95. 897 - 903.
A. Inoue, K . Nakagawa, and S. Morisawa, Department of Biochemistry, Osaka City University Medical School. 1-4-54 Asahi-machi. t\heno-ku, Osaka-shi, Osaka-fu, Japan 545


















