
Effect of Aluminum Uptake Translocation Phosphorus ... the exception of our preliminary...
7
Effect of Aluminum on Uptake & Translocation of Phosphorus 32 by Perennial Ryegrass ' P. J. Randall 2 & P. B. Vose 3 Welsh Plant Breeding Station, Aberystwyth A major effect of aluminum toxicity in plants is the disturbance of phosphorus assimilation. Wright (15) noted similar symptoms in beet plants grown in phosphorus-deficient media and in culture solu- tion containing aluminum, while work at Aberystwyth (13) has shown that ryegrass lines resistant to alu- minum toxicity require much higher levels of alumi- num to dlevelop aluminunm toxicity/phosphorus de- ficiency symptoms than do control unselected lines. Burgess and Pember (1) suggested that the action of acid phosphate fertilizer in reducing aluminum toxicity effects was due to a phosphate-aluminum in- teraction within the root. Further work by Wright, (16, 17, 18) developed the hypothesis that aluminum was precipitated as the phosphate within the root, while Wallihan (14) thought the precipitation was more likely to occur on the root. With the exception of our preliminary communi- cation (10) little work appears to have been carried out on the possible role of metabolic factors in the aluminum/phosphate interaction, although during the preparation of this paper Ragland and Coleman (9), using excised snap bean roots, have reported results similar to some of ours. This paper describes the influence of aluminum on the uptake and translocation of P32-labeled phos- phorus and the effect on this interaction of certain metabolic inhibitors. Materials & Methods Plant Material. Seeds of perennial ryegrass, Loliumst perenne L. var. Danish were germinated in petri dlishes on filter paper moistened with dleionised water. After 8 days the seedlings were transferred to continuously aerated culture solution in 9.5 liter glazed pots. The solution composition was 0.0025 M Ca(NO3)9 4H.,0; 0.0015 m MgSO4 7HoO; 0.001 MI KCI; 0.0002 M NaH9PO; 5.6 ppm Fe as FeEDTA and micronutrients given as a modified Hoagland A-Z formula. Aluminum, as Al.(SO)3, was supplied to the aluminum treatments. Analytical 1 Received Oct. 29, 1962. 2 Work carried out during the tenure of a postgradu- ate studentship awarded by the Agricultural Research Council. 3 Present address: College of Agriculture, Seoul Na- tional University, Suwon, Korea. grade chemicals anid deionised wvater were usedl throughout. Uptake of Phosphorus. In the initial experimzent plants 8 weeks old were used in a series of uptake studies. Continuously aerated uptake solutions were contained in foil-wrapped liter beakers, and w-ere ad- justed to pH 4.0 to 4.1 using 2 N H2,SO and M Na.,CO. There were two main series of plants: one series was grown without aluminum and alumi- num was added to the uptake solutions; the other series w%as pretreated with 1.85 X 10-4 At or 18.5 X 10-4 I Al during growth and had no aluminum in the uptake solutions. Phosphorus was given as NaH.PO4 at 4.5 x 10 i%f and 22.5 x 10 SI P, labeled respectively with 10 and 50 i'c of p32. Assav of Phosphorus. After each uptake period the roots were washed with five successive changes of deionised water, blotted, dried at 800 and weighed. Shoots and roots were ashed in the muffle on stain- less steel planchets at 4000, the ash being taken up with dilute nitric acid and evaporated to dryness. The activity was counted with a mica end-windowv counter of 1.5 mg/cm2 thickness. Total phosphorus was estimated by extracting the planchets with dilute nitric acid, adjusting the solu- tion to pH 3.0, and determining phosphorus colori- metrically by stannous chloride reduction of the phosphomolybdate. Inhibitors. In the inhibitor experinment plants were grown in liter beakers for 6 weeks, in the same continuously aerated solution as previously. The solution was changed weekly. The procedure for uptake studies was to transfer the plants to deionised water overnight, then place thenm for 1 hour in ap- propriate inhibitor solution prior to transfer to the uptake solution. This comprised the full culture medium with 4.5 X 10-4 AI phosphorus labeled witlh 30 /ic of p32, plus aluminum and inhibitor, and the pH adjusted to 4.0 to 4.1. Inhibitors used were 2,4- dinitrophenol (DNP), KCN and MnSO4. Follow- ing absorption periods, roots were rinsed once in deionised water, followed by one washing in 0.01 3r NH4Cl solution and tw-o more washings in deionised water. Au toradiogra phs. For autoradiographic studieF 9 week old plants were subjected to the same pro- cedure as in the inhibitor experiment. For the time- course studies, roots only were used and tops werc 403 www.plantphysiol.org on June 26, 2018 - Published by Downloaded from Copyright © 1963 American Society of Plant Biologists. All rights reserved.
Transcript of Effect of Aluminum Uptake Translocation Phosphorus ... the exception of our preliminary...

Effect of Aluminum on Uptake & Translocationof Phosphorus 32 by Perennial Ryegrass '
P. J. Randall 2 & P. B. Vose 3
Welsh Plant Breeding Station, Aberystwyth
A major effect of aluminum toxicity in plants isthe disturbance of phosphorus assimilation. Wright(15) noted similar symptoms in beet plants grownin phosphorus-deficient media and in culture solu-tion containing aluminum, while work at Aberystwyth(13) has shown that ryegrass lines resistant to alu-minum toxicity require much higher levels of alumi-num to dlevelop aluminunm toxicity/phosphorus de-ficiency symptoms than do control unselected lines.
Burgess and Pember (1) suggested that the actionof acid phosphate fertilizer in reducing aluminumtoxicity effects was due to a phosphate-aluminum in-teraction within the root. Further work by Wright,(16, 17, 18) developed the hypothesis that aluminumwas precipitated as the phosphate within the root,while Wallihan (14) thought the precipitation wasmore likely to occur on the root.
With the exception of our preliminary communi-cation (10) little work appears to have been carriedout on the possible role of metabolic factors in thealuminum/phosphate interaction, although during thepreparation of this paper Ragland and Coleman (9),using excised snap bean roots, have reported resultssimilar to some of ours.
This paper describes the influence of aluminumon the uptake and translocation of P32-labeled phos-phorus and the effect on this interaction of certainmetabolic inhibitors.
Materials & MethodsPlant Material. Seeds of perennial ryegrass,
Loliumst perenne L. var. Danish were germinated inpetri dlishes on filter paper moistened with dleionisedwater. After 8 days the seedlings were transferredto continuously aerated culture solution in 9.5 literglazed pots. The solution composition was 0.0025 MCa(NO3)9 4H.,0; 0.0015 m MgSO4 7HoO;0.001 MI KCI; 0.0002 M NaH9PO; 5.6 ppm Fe asFeEDTA and micronutrients given as a modifiedHoagland A-Z formula. Aluminum, as Al.(SO)3,was supplied to the aluminum treatments. Analytical
1 Received Oct. 29, 1962.2 Work carried out during the tenure of a postgradu-
ate studentship awarded by the Agricultural ResearchCouncil.
3 Present address: College of Agriculture, Seoul Na-tional University, Suwon, Korea.
grade chemicals anid deionised wvater were usedlthroughout.
Uptake of Phosphorus. In the initial experimzentplants 8 weeks old were used in a series of uptakestudies. Continuously aerated uptake solutions werecontained in foil-wrapped liter beakers, and w-ere ad-justed to pH 4.0 to 4.1 using 2 N H2,SO and MNa.,CO. There were two main series of plants:one series was grown without aluminum and alumi-num was added to the uptake solutions; the otherseries w%as pretreated with 1.85 X 10-4 At or 18.5 X10-4 I Al during growth and had no aluminum inthe uptake solutions. Phosphorus was given asNaH.PO4 at 4.5 x 10 i%f and 22.5 x 10 SI P,labeled respectively with 10 and 50 i'c of p32.
Assav of Phosphorus. After each uptake periodthe roots were washed with five successive changesof deionised water, blotted, dried at 800 and weighed.Shoots and roots were ashed in the muffle on stain-less steel planchets at 4000, the ash being taken upwith dilute nitric acid and evaporated to dryness.The activity was counted with a mica end-windowvcounter of 1.5 mg/cm2 thickness.
Total phosphorus was estimated by extracting theplanchets with dilute nitric acid, adjusting the solu-tion to pH 3.0, and determining phosphorus colori-metrically by stannous chloride reduction of thephosphomolybdate.
Inhibitors. In the inhibitor experinment plantswere grown in liter beakers for 6 weeks, in the samecontinuously aerated solution as previously. Thesolution was changed weekly. The procedure foruptake studies was to transfer the plants to deionisedwater overnight, then place thenm for 1 hour in ap-propriate inhibitor solution prior to transfer to theuptake solution. This comprised the full culturemedium with 4.5 X 10-4 AI phosphorus labeled witlh30 /ic of p32, plus aluminum and inhibitor, and thepH adjusted to 4.0 to 4.1. Inhibitors used were 2,4-dinitrophenol (DNP), KCN and MnSO4. Follow-ing absorption periods, roots were rinsed once indeionised water, followed by one washing in 0.01 3rNH4Cl solution and tw-o more washings in deionisedwater.
Au toradiogra phs. For autoradiographic studieF9 week old plants were subjected to the same pro-cedure as in the inhibitor experiment. For the time-course studies, roots only were used and tops werc
403 www.plantphysiol.orgon June 26, 2018 - Published by Downloaded from
Copyright © 1963 American Society of Plant Biologists. All rights reserved.

P'LANT PHYSIOLOGY
remove(l just prior to placing in the uptake solution.The roots wvere washed with five chlanges of (leionise(l-water. blotted, sprea(l by flotation OIn stiff paper, andblottedI dry. In the case of whole plants the rootsw+ere separatetl frolmi the tops. Autoradiographswere prepare(l by placing the fresh miiaterial in (lirectcontact with Kodirex nio-screen X-ray fillmi. Thegeneiral proce(lure of Yamaguchi and Crafts ( 19)xwas followed, a 1 cm11 rubber hacking sheet being useclto ensure close contact of the pl)ant aln(d filmii. Dur-ing the 16 day exposure period the imiaterial wasmaintained at -10
Results & Discussion
Initial ExperimentYield. WVith only 1.85 x O--' Al. slhoot vield
is substantially reduce(d (table I). Yield of roots,hoxvever. is only re(luce(l xx-itlh 18.5 X 10 4m Al.Root length is greatly reduced, secondary roots fail-inig to formii properly and the initials shox-ilng as pro-tuberanlces on the main root.
Phospho'rius conttenlt. The phosphorus concentra-tion (tahle I) increases in both shoots and roots
with increasing aluminum, the relative inlcrease beinggreater in the roots. The total phosphorus in theshoots increases vith 1.85 X 10-4 -m Al, but with18.5 X 10 m Al the coniteint is beloxv the coIntr-ol.while that in the roots also increases xvith 1 .85 X10-4 m Al. In general, the pIattern of plhl)sphoruisconcentration an(l content follovs the usual tren(d ofhigher- minieral contelnt xvhen an oxerall g,rowtll-limitiig factor is operative.
Shlort-term-l P iupt(ake. Tahle 11 shows the largeinlcrease in P uptake oxer 4 houri-s at the loxver levelsof aluimiinull, both frolmi 4.5 X 10 and 22.5 X10 \I phosphorus solutioins. This increase occursxvhetlher the aluimiilnuI is givein at tlle commliilelnceimlenltof the uptake period or as a pretreatment (itii-ilg tllegrowth periocl.
The increase is generally smaller in the plretreit-ment series, possibly (lue to the highler 1) concentra-tioIn in the aluminiiiumii-treate(d plants at the commiiiienice-mlent of the uptake periodl. In effect, the comparisonbetwveen pretreated and untreated plants may Ilartiallybe a comparison of hiiglh salt and lo\v salt plaints (3).
The (lifferelnce in uptake of P) froml the 4.5 X10-4 1an(l 22.5 X 10 1' levels is ('reatei. Par-
able I)Iflutt1)c' of -lOii,;,,in Lezcl on, Yield, Phosplhoruts C(ontelt, & I)istril)ni10)1
i,n 8 WM,k Old Plants of Lolinti, Perelntne.
Alumilluill Yield Phosphorus cointenlt Total phosphorus Transport index,treaitmllenlt mnig dry Nxvt mg g dry wN-t mig plant
MN X 10 shoot root .shoot root shoot root
0 25. 5.7 5.38 11.85 0.131 0.073 64.21.85 17.5. 8.28 17.57 0.151 0.090 62.6
18.5 10.0 2.4 10.15 44.14 0.086 0.077 52.8
shloot Px 100
total P
Table IIEffect of Ai.nminuni on tfle Short-tcrmn l 'ptaetwc of IPhiosplhorins by Lolinm,, perentn
Treatimienit: Aluminiilumii gixeni at comimleIncemilenit of ul)take period (4 hr ), 11o pretreatment during g-rowth period4.5 X 10-4 -Tr P 22.5 X 10- --\I p
.\lullilluntreatment
I\ x 10-4P uptake
ug g (Iry wxt
Shoots roots
2472
14075
Transport index(V
845451340962395
10.497.03
14.2411.11
P uptakeug/g Idry wvt
shoots roots
113113173169
Transport indlexC,I
3378702169146289
11.146.289.438.86
Treatmenit: Pretreatmiient wxith three levels of alumitnum during 8 week growtth period, no aluminiumii given ldurinig up-take period (4 hr).
P uptakeug g dry xx-t
shoots roots
4.5 10-4r P
Transport index/ C!
10.518.610.1
P uptakeug/'g dry wt
shoots roots
113164
,;
22.5 X 10-4 -r P
Tranisport inidexCl,
33785406
5145
11.118.43.0
404
01.859.25
18.5
A luminutreatment
Mr x 10-4
01.85
18.5
2412472
84519481557
www.plantphysiol.orgon June 26, 2018 - Published by Downloaded from Copyright © 1963 American Society of Plant Biologists. All rights reserved.
RANDALL & VOSE-ALUMINUMI EFFECT ON PHOSPHORUS NUTRITION
V
10
405
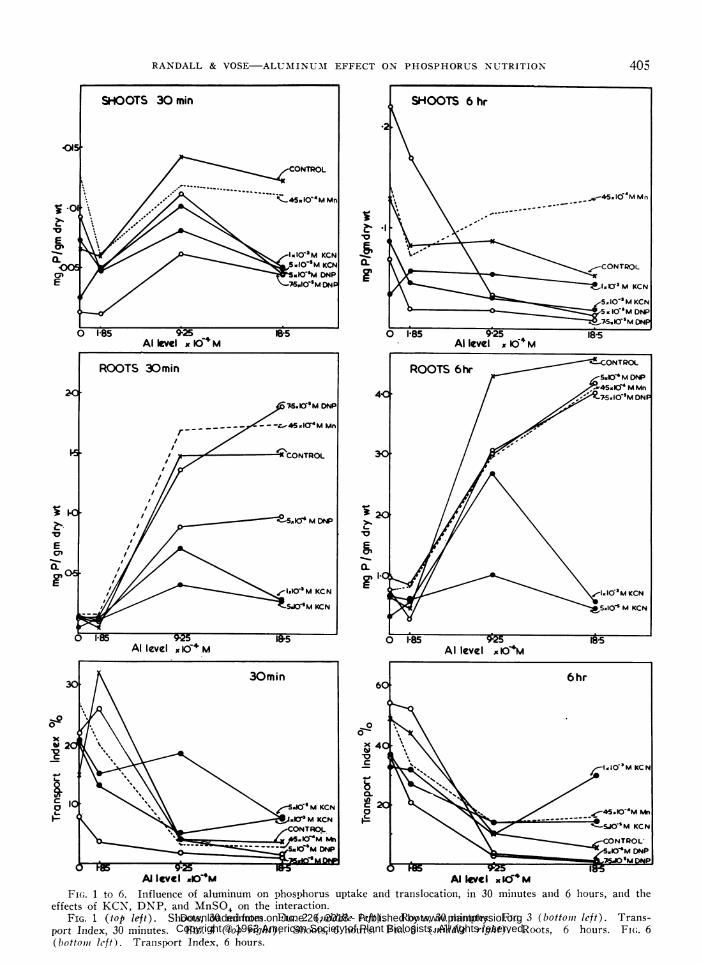
Ailevel 10'4m Al kve IxdmFI(;. 1 to 6. Influence of aluminum on phosphorus uptake and translocatioin, in 30 minutes and 6 hours, and the
effects of KCN, DNP, and MnSO4 on the interaction.FIG. 1 (top left). Shoots, 30 minutes. FIG. 2 (middle left). Roots, 30 minutes. FIG. 3 (bottomI left). Trans-
port Index, 30 minutes. FIG. 4 (top right). Shoots, 6 hours. FIG. 5 (mi1iddle right). Roots, 6 hours. FI(.. 6(bottomii left). Trainsport Index, 6 hours.
www.plantphysiol.orgon June 26, 2018 - Published by Downloaded from Copyright © 1963 American Society of Plant Biologists. All rights reserved.

PLANT PHYSIOLOGY
ticularlv in the case of roots, xvhei aluminum is givenas a pretreatment. This may be due to the increas-ing imilortance of P level xvitlh re(luced root size.
The effect of aluminum in stimulating the uptakeof phosphoruis is in the nature of a Viets effect ( 12).Howev-er- certain (lifferences haxe been poinlted out(9 ) aan(l there seemiis to he no exact comparison.The (questioni arises a5 to \\vhether this stimulationis mletabolic or (lue to precipitation of P by the alumi-illUIl in the free space of tile root. The uptake froml22.5 X 10-- xm P substrate is greater than froml 4.5X 10-4 x\ 1), but tile increase fromii both levels of 1'due to 1.85 X 10-- 4m Al is the same (approx 3.6 mlgper plant ). The (lecrease (lue to 18.5 X 10 ImAlis relatively greater at tile loxver level of l'.
Raglland and Coleman ( 9 ) suggested that the (Irol)in P uptake (lue to high levels of alumiillnulmi in their-experiments wvas (lue to the solubilitv pro(luct ofalulmiinumiphlospllate being exceee(lel, \\ith coIlse(iueletprecipitatioll of }) in tile solution, tilus decreasinogthe P available to the plant. We have foundl (table11) that there is a lower P uptake by tile roots fromltIle 18.5 X 10 4m Al treatillents thlan froml the1.85 X 10-4[ Al treatments, wxhethler the aluilllunuis applied as a plretreatilleilt onlv or (luriilg tile up-take period only. XN bile tile solubility pro(duct argu-Illent could( be applie(l in tile latter case, it seeillsunlikelv that sufficienit aluillinulll could (liffuse outof the pIretreatedl roots to re(luce appreciably substrateP by precipitation. Alternativ,ely since no precipi-tate was observed in the solutioll, it Iligilt lbe thatthe low er aluiiillumi levels stiillulate tile uptake ofP. but ligiher levels block tile mlechlanism. The (if-ference betxveenl P ulptake at the two alunliilunl levelsis greater at tile lox-er lexel of solution phospliorus.Thius raising the P' level in the solution enables by-passing of the blocking mlechanism to soille extent.
Timize couirse. A tille course stud(i in(licates (in-set fig 7 & 8) that xvith 9.25 X 10 4m Al the silOOtssioNx- a stea(dv iilcrease in P) over 24 liours. Theroots shoxv ail iIlitial slharp inlcrease in the first liour,a brief (lecrease at the eil(i of 2 hours, followed by
(al alillost liIlear inlcrease so tilat tile curve aplproachl-es aan asymnlptotic patteril at tile eil(l of 24 hours.Tile general pattern of the curv-e xxwas the saille forbothl substrate levels of P.illilhitor Studlies.
Tile effects oil tile aiuillinluill-pllosphorus iilter-actioil of potassiuilm cyalli(le. DNP anld ilainganesesulfate x-ere stu(lie(d (figs 1-6).
The figures for P uptake witli 1.85 x 10- \i Alslhowv eitlher little cliailge or a decrease coml)aredl tozero Al. as opposeci to the large increase at tile sailleAl level in the initial experimlent. This couldl be
(lue to reactions within the conmplete nutrieint solu-tioin, as use(l in the inhibitor studies, p)recipitatiIlgaluinii1uImi or alunlinulll aIld phosphorus, although io
precipitate xvas observed in solutioil iil eitlier experi-illeint.
Oxver a 30 illilute period aluiliiuili stiillulates I)tranislocatioii to tlle slioots, but after 6 hours it has
an inhibitory effect. There is a large increase inuptake into the roots at tlle higher- le-els of alumillumll.Autoradiographs (not gienehere that in theslhort timles used mullclh of the P in the slhoots ix con-finedl to the junction of roots an(l slhoots and in theslhoot bases. Thlus separation of r )Ots nd slhootsmay be critical hiere.
The effect of manganese in alplparentl\ offsettingaluminum toxicity sy mptom s had been l)revi ouslxnote(d in sand( culture experiments -reese. & Vose.unpublished). In 6 lhours manganese greatly stimllU-lates phosphorus translocation to the slhoots at higherlevels of aluminum ( fig 4). No exlllanation for thiS.can vet le a(Ivaice(l.
D)NP hlas a major effect oni the translocation ofP to the slhoots. The root 1) conitelnt figures are onl\-a little belowx the colntr-ols. \Without (aluminumi, 5 X10-G m, I)NP stimulates 1 translocation at both 30miiniutes and 6 1hi-ours. but with increasing alumillumllthere is an increasing inhibition w itlh DNP. TheDNP-stimulation of P transport with zero alumlmillnum1is in agreenment -with the results of Russell et al. ( 11),who foundl in barley that 5 x 10 -rI)mNP (lecrease(lretenitioin of 1) in the roots anl inlcreatse(d the 1' con-tent of the slhoots. They also fountd that phosphatetransportedl to the shoots asamaiinl\- iln inorganicformls ( 7 ). This sug-ests that the effect of alumi-numii may be to re(luce the inorgan-ic 1) fraction in tlleroot by stimulating the incorlporation of I ill organicformls or to prev,ent the tranisfer- between orgalnic an(linorganic formiis. Alternati velv the phosplhorus maybe precipitate(l by aluminum in lan ins)luble inorganicformii.
KCNlhas mlost effect on root phosphorus- uptake.The shoot values at the highler aluminum lexvels fallbetween the control an(l those for DNP. WVith 18.5X 10-4 m Al there is a very large re(luctioni in 1'content (lue to KCN. It seems unlikely that the cya-ni(le induces leakage of phosphorus into the washinlgsolutions, as this would also be apI)larent at lowerlevels of aluminum. Tlhus at least patrt of the alumli-num-stimulated P uptake appears to be mletabolic innature, and(l cy-anide sensitive salt respiration apl)pearsto be involved in the inlteraction.
The effect of c-anile level is the rev-erse of obvi-ous expectatiolln in both roots and shoots, the higlhcyanide causing less inhlibitioni thalnl the low cyanildelevel wsheni aluminiumii is present. In the absence ofaluminuml, the high cyanide show s greater inhibitionof P uptake and translocation than the lowx cyanidelevel. This phenomenon is also preseint witlh DNP-effect on the roots after 30 mlinutes (fig 2) and thecauses are not obvious. Experimlenital proce(luremlakes it certain that the feature is genuilne and( not(lue to error in halresting or processing the material.
Tim,e courMse. A timle course study (fig 7 & 8)shoxvs clearly that the mlajor effect of DNP in thealulmiinumii-plhosphorus interaction is oni tran-isport ofP to the shoots.
The l) conitenit of the slhoot show s an inicr-easexxith timiie except in the case of the aluminum plus
40
www.plantphysiol.orgon June 26, 2018 - Published by Downloaded from Copyright © 1963 American Society of Plant Biologists. All rights reserved.

RANDALL & VOSE-ALUMINUM EFFECT ON PHOSPHORUS NUTRITION
v
L.
E
TIME hours
4
3
E
0.
F
_ 3 6TIME hours
FIG. 7 (above) & FIG. 8 (below). Time course ofaccumulation of phosphorus as influenced by aluminumand DNP. Expt. I refers to the initial experiment. Theother figures are from the inhibitor experiment.
DNP treatment, which appears to decrease steadilyafter 1 hour. In the case of the roots, in the presenceof aluminunm there is a steep rise in P content inthe first hour followed by a drop, after which it in-creased again steadily. This drop is the same as thatobtained in the initial experiment vith both 4.5 X10-4 Mr and 22.5 x 10-4 \r P substrate levels. Nosuch drop occurred when DNP was given withaluminum. There seems to be some connection be-tween this and the fact that 7.5 X 10- AI DNP in-creased P uptake of the roots ov-er the control at higlhlevels of aluminum at the end of 30 minutes (fig 2).
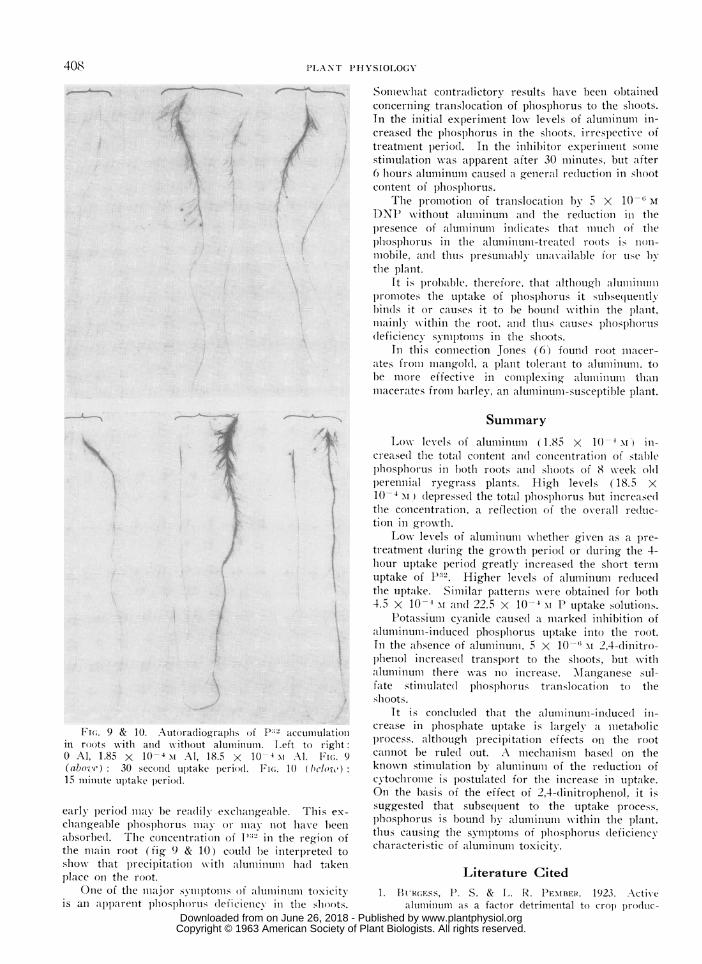
Auttoradiographs. Figures 9 and 10 show thedistribution of P after 30 seconds and 15 minutes inroots excised immediately before being placed in thetest solutions. Other autoradicgraphs run at tinmesup to 30 minutes gave a sinmilar pattern.
Close examination of the lateral roots shows aconcentration of P in the growing points after 30seconds when aluminum is present, but the concen-tration in this region is only just discernible after15 minutes with no aluminum. This would appearas strong evidence for the metabolic nature of thestimulation of P uptake by aluminum.
The other region of high P concentration isaround the upper portion of the main root. Thismay be due to inadequate spreading of the lateralroots which tended to cling to the main root duringthe flotation procedure, to accumulation in the root,or to precipitation on the root. As this region ofconcentration is shown after 15 minutes even in theabsence of aluminum, precipitation by alumlinumii onthe root cannot be the sole cause.
DiscussionMlain interest centers on whether the undoubted
stimulation of phosphorus uptake by aluminunm undercertain conditions is metabolic or merely dlue to pre-cipitation within or on the root. Further, in view ofthe apparent phosphorus deficiency symptoms ofplants suffering from aluminum toxicity, what is theoverall effect of aluminum on phosphorus assinmila-tion and utilization?
That the mechanism of aluminum stinmulation ofphosphate uptake is an internal metabolic process,rather than a precipitation effect is indicated by anumber of factors: A, the similarity of the effect ofaluminum given as a pretreatment or during the up-take period, B, autoradiographs of root tips indicatethat treatments without aluminum require 15 minutesuptake of p32 to show the same activity as that givenby treatments with aluminum in only 30 seconds up-take, C, cyanide causes a marked inhibition of thisaluminum-induced phosphorus uptake.
Jackson et al. (5) have shown that the rate-limit-ing reactions for phosphorus uptake are those whichinvolve the formation of high energy phosphate.Thus aluminum must either stimulate these reac-tions or stimulate phosphorus uptake by some alterna-tive pathway.
The existence of two sites of phosphate uptakehas been suggested (2), one associated with thecytochrome b system, and the other with diphos-phopyridine nucleotide oxidation or reduction in therespiratory chain. It is known (4, 8) that aluminumcan promote the reduction of cytochrome c in a suc-cinic dehydrogenase-cytochrome c system. Themarked inhibition of aluminum-induced phosphateuptake by cyanide, suggests that aluminum stimulatesphosphate uptake at the cytochrome site.
There are some indications that non-metabolicprocesses may also be involved. The sharp dip inthe time course uptake curve for roots (fig 8) be-tween 2 and 4 hours in the presence of aluminum,and the subsequent recovery, is not typical of a pure-ly metabolic process, and indicates that some of thealuminum-stimulated phosphorus absorbed in the
r ~~~~--(75.IWM DN
Q --------''--L
407
www.plantphysiol.orgon June 26, 2018 - Published by Downloaded from Copyright © 1963 American Society of Plant Biologists. All rights reserved.
PLANT PHYSIOLOGY
F(,. 9 & 10. Autoradiographs of Pb2 accumIlulationin roots witi anidl without alumiiiluinu. Left to right:
Al, 1.85 X 10 4 Al, 18.5 X 10-4 m Al. FIw. 9(above) : 30 seconid uptake period. E;Fw. 10 ( bc(el.)ow15 miilniute upitake periodl.
early period may be readilx exchangeable. This ex-
changeable phosphorus milla or may lnot have been
absorbed. The concentration of P1'2 in thle region ofthe main root (fig 9 & 10) coul(d be initerprete(l toshow that precipitation w ith (aluminumiitiii had takenplace on the root.
One of the major vymptoms of aluminum toxicityis ai apl)parent phlosplhorus (leficiency in the shoots.
Somewhat contrad(lictory results have beein obtainedconcerniing translocation of phosphorus to the shoots.In the initial experiment low levels of alumiiinulmi in-crease(I the phosphorus in the slhoots, irrespective oftreatmlenit periodl. In the i nhibitor experimiient somiiestiimiulation was apparent after 30 miiinutes. but after6( hours aluminlumll caused a general reduction in shlootconltenit of phosphorus.
Tlle proniotioni of translocation 1, 5 X 10- -m
DNP without aluniinum and thle re(luctioni in thepresence of aluminum ind(licates that mltucil of tlheplhospshorus in the aluminum-treated roots iS 11011-
mllobile, and tlhuis presumabl uinavalilable for tuse bythe planit.
It is probable. therefore, thlat although1 aluminmlllllp)romotes the uptake of phosphoruis it subsequentlxhinds it or causes it to be bound wlithlinl the plant,mainly within the root, and tlhus causes phosphorusdeficiencv symptoms in the shoots.
In this conniection Jonies (6) found r1oot macer-aites fromi mangold, a plant tolerant to aluminum, tohe miiore effective in complexiug aluminum thanmacerates friomii barlev, an alttllllnum-sutscep)tible plaint.
SummaryLow- levels of aluinlllmllll (1.8s X 10- m ) in-
crease(l the total content and conicenitratioll of stableplospllhoris in both roots and shoots of 8 week 01(lperennial ryegrass plants. High levels ('18.5 X10 \ ) (lepresse(l the total phosphorus but inicrease(lthe concentration, a reflectioln of the overall redutc-tiOln in growth.
Low levels of alumiinium xhwhether given as a pre-treatnmenit (luring the groxx th perio(d or dlurinig the 4-lhour uptake period greatly increasedl the short term-luptake of J32. Higher levels of alumiinulmi re(lucedthe uptake. Simiilar patterns w-ere obtaine(d for both4.5 X 10- r and 22.5 X 10- m P uptake solutions.
Potassiulml cyanide caused a marked inhibition ofatlumiiinulm -incluce(d phosphorus uptake inlto the root.In the absence of aluniinuml, 5 X 10-' a 2,4-dlinitro-plhenol inierease(d transpol-t to the slhoots, but wvithaluminum there xvas nlo increase. Alanganese SUl-fate stimilulate(d phosphorus translocation to thelloots.
It is conclu(ledl that the alumiiniumi-ind(lucedl in-crease in phosphate uptake is largely a metabolicplrocess. although precipitation effects onl the rootcannot be rule(d out. A mechlanismn base(d oni theknown stimiulationl by aluminumll of the redluctioln ofcvtochrome is postulated for the inicrease in uptake.Oln tlle basis of the effect of 2.4-dinitrophenol, it issuggestedl that subse(luent to the uptake process.plhosphorus is bound by aluminum w-ithin the plant.tlhus causing the symptoms of plhosphorus (leficiencxclharacteristic of aluminum toxicitv.
Literature Cited1. Bu<,sss, P. S. & L. R. PENIBER. 1923. ActiVe
altiiinlumil aIs a factor detrimenital to crol) produc-
408
www.plantphysiol.orgon June 26, 2018 - Published by Downloaded from Copyright © 1963 American Society of Plant Biologists. All rights reserved.

RANDALL & VOSE-ALUMINUM EFFECT ON PHOSPHORUS NUTRITION
tion in many acid soils. Rhode Island Agr. Expt.Sta. Bull. 194.
2. HAGEN, C. E., J. E. LEGGETT, & P. C. JACKSON.1957. The sites of orthophosphate uptake by bar-ley roots. Proc. Natl. Acad. Sci. 43: 495-506.
3. HOAGLAND, D. R. & T. C. BROYER. 1936. Generalnature of the process of salt accumulation by roots
with description of experimental methods. PlantPhysiol. 11: 471-507.
4. HORECKER, B. L., E. STOTZ, & T. R. HOGNESS.1939. The promoting effect of aluminum, chromi-um, & the rare earths on succinic dehydrogenase-cytochrome c systems. J. Biol. Chem. 128: 251-56.
5. JACKSON, P. C., S. B. HENDRICKS, & B. M. VASTA.1962. Phosphorylation by barley root mitochon-dria & phosphate absorption by barley roots.Plant Physiol. 37: 8-17.
6. JONES, L. H. 1961. Aluminum uptake & toxicityin plants. Plant & Soil 13: 297-310.
7. LOUGHMAN, B. C. & R. S. RUSSELL. 1957. The ab-sorption & utilization of phosphate by young bar-ley plants. IV. The initial stages of phosphatemetabolism in roots. J. Exptl. Botany 8: 280-93.
8. POTTER, V. R. & W. C. SCHNEIDER. 1942. Studiesin the mechanism of hydrogen transport in animaltissues. V. Dilution effects in the succinic oxi-dase system. J. Biol. Chem. 142: 543-55.
9. RAGLAND, J. L. & N. T. COLE-MAN. 1962. The in-
fluence of aluminum on phosphorus uptake by snapbean roots. Proc. Soil Sci. Soc. Am. 26: 88-90.
10. RANDALL, P. J. & P. B. VoSE. 1961. The effectof aluminum on the short-term uptake of stable& radioactive phosphorus by perennial ryegrass.
Paper presented to the Soc. for Exptl. Biol.Aberystwyth. July, 1961.
11. RUSSELL, R. S., R. P. MARTIN, & 0. N. Bisitop.1953. A study of the absorption & utilization ofphosphate by young barley plants. II. The effectof phosphate status & root metabolism on the dis-tribution of absorbed phosphate between roots &shoots. J. Exptl. Botany 4: 136-56.
12. VIETS, F. G. JR. 1944. Calcium & other poly-valent cations as accelerators of ion accumulationby excised barley roots. Plant Physiol. 19: 466-80.
13. VOSE, P. B. & P. J. RANDALL. 1962. Resistanice toaluminum & manganese toxicities in plants relatedto variety & cation-exchange capacity. Nature196: 85-6.
14. WALLIHAN, E. F. 1948. The influence of alumi-num on the phosphorus nutrition of plants. Am.J. Botany 35: 106-12.
15. WRIGHT, K. E. 1937. Effects of phosphorus &lime in reducing aluminum toxicity of acid soils.Plant Physiol. 12: 173-81.
16. WRIGHT, K. E. 1943. Internal precipitationi ofphosphorus in relation to aluminum toxicity.Plant Physiol. 18: 708-12.
17. WRIGHT, K. E. 1945. Aluminum toxicity. Micro-chemical tests for inorganically & organicallybound phosphorus. Plant Physiol. 20: 310-12.
18. WRIGHT, K. E. & B. A. DONAHUE. 1953. Alumi-num toxicity studies with radioactive phosphorus.Plant Physiol. 28: 674-80.
19. YAMAGUCHI, S. & A. S. CRAFTS. 1958. Auto-radiographic method for studying absorption &translocation of herbicides using C14-labeledcompounds. Hilgardia 28: 161-91.
409
www.plantphysiol.orgon June 26, 2018 - Published by Downloaded from Copyright © 1963 American Society of Plant Biologists. All rights reserved.


















