
Effect of agar concentration on growth and anatomy of adventitious shoots of Picea abies (L.) Karst
8
Plant Cell Tissue.Organ Culture 3:257-264 (1984). ©Martinus Ni/hoff/Dr W. Junk Publishers, Dordrecht. Printed in the Netherlands. Effect of agar concentration on growth and anatomy of adventitious shoots of Picea abies (L.) Karst. 257 SARA VON ARNOLD and TAGE ERIKSSON Institute of Physiological Botany of the University of Uppsala, Box 540, S-751 21 Uppsala, Sweden (Received 20 January 1984; in revisedform 21 March, 1984; accepted 10 April, 1984) Key words: Adventitious shoot, agar concentration, needle anatomy, propagation rate, vitrification, l~'cea abies Abstract. The development of adventitious shoots of Picea abies was affected by the ag~ concentration in the culture medium. Increasing the agar concentration from 0.5 to 2.0% decreased vitrification, but at the same time reduced shoot growth and roofing potential. Slightiy vitrified plantiets could become acclimatized to greenhouse conditions. The mesophyll of needles developed in vitro was interspersed with large air spaces; the lower the agar concentration, the larger the air spaces. After transfer to the greenhouse, the new needles from the acclimatized plantlets had an anatomy approach- ing that of plants growing in field. Introduction Vitrification of buds and shoots frequently occurs during culture in vitro. This is a great problem since vitrified shoots or plantlets cannot survive transfer to greenhouse conditions; they lose too much water and are very susceptible to infections. The progress of vitrification of Cynara scolymus L. has been described by Debergh et al. [8] as follows: the leafblade and the petiole become humid; very young leaves in the center of the rosette become very turgescent; small turgescent leaves elongate suddenly aftd reach lengths 5 to 6 times greater than those of normal leaves; and finally, the original leaves become almost translucent and later necrotic. Similar phenomena have been observed in Allium porrum L., Brassica oleracea L. var. botrytis L., Dianthus caryophyllus L., Ficus lyrata Warb., Malus spp., Pinus spp., Prunus spp., Raphanus sativus L., and Sequoia sempervirens (Lamb.) Endl. [8]. In cultures of many species, lowered agar concentration promotes the formation of vitrified shoots (8 with ref.). Free water and high humidity in the culture vessels [1] and the type of vessel closure [10], contribute to the formation of these shoots. Furthermore, high cytokinin concentrations increase vitrifi- cation at low agar concentrations [7]. Vitrified and normal plantlets derived in vitro are sensitive to changes in the relative humidity. They often show symptoms of severe water stress when transferred to pots. Excessive water loss from the plantlets often
-
Upload
sara-arnold -
Category
Documents
-
view
216 -
download
1
Transcript of Effect of agar concentration on growth and anatomy of adventitious shoots of Picea abies (L.) Karst

Plant Cell Tissue.Organ Culture 3:257-264 (1984). © Martinus Ni/hoff/Dr W. Junk Publishers, Dordrecht. Printed in the Netherlands.
E f f e c t o f agar c o n c e n t r a t i o n o n g r o w t h a n d a n a t o m y o f
a d v e n t i t i o u s s h o o t s o f Picea abies (L . ) Kars t .
257
SARA VON ARNOLD and TAGE ERIKSSON
Institute of Physiological Botany of the University of Uppsala, Box 540, S-751 21 Uppsala, Sweden
(Received 20 January 1984; in revised form 21 March, 1984; accepted 10 April, 1984)
Key words: Adventitious shoot, agar concentration, needle anatomy, propagation rate, vitrification, l~'cea abies
Abstract. The development of adventitious shoots of Picea abies was affected by the ag~ concentration in the culture medium. Increasing the agar concentration from 0.5 to 2.0% decreased vitrification, but at the same time reduced shoot growth and roofing potential. Slightiy vitrified plantiets could become acclimatized to greenhouse conditions. The mesophyll of needles developed in vitro was interspersed with large air spaces; the lower the agar concentration, the larger the air spaces. After transfer to the greenhouse, the new needles from the acclimatized plantlets had an anatomy approach- ing that of plants growing in field.
Introduction
Vitrification of buds and shoots frequently occurs during culture in vitro. This is a great problem since vitrified shoots or plantlets cannot survive transfer to greenhouse conditions; they lose too much water and are very susceptible to infections. The progress of vitrification o f Cynara scolymus L. has been described by Debergh et al. [8] as follows: the leafblade and the petiole become humid; very young leaves in the center of the rosette become very turgescent; small turgescent leaves elongate suddenly aftd reach lengths 5 to 6 times greater than those of normal leaves; and finally, the original leaves become almost translucent and later necrotic. Similar phenomena have been observed in Allium porrum L., Brassica oleracea L. var. botrytis L., Dianthus caryophyllus L., Ficus lyrata Warb., Malus spp., Pinus spp., Prunus spp., Raphanus sativus L., and Sequoia sempervirens (Lamb.) Endl. [8]. In cultures o f many species, lowered agar concentration promotes the formation of vitrified shoots (8 with ref.). Free water and high humidity in the culture vessels [1] and the type o f vessel closure [10], contribute to the formation of these shoots. Furthermore, high cytokinin concentrations increase vitrifi- cation at low agar concentrations [7].
Vitrified and normal plantlets derived in vitro are sensitive to changes in the relative humidity. They often show symptoms of severe water stress when transferred to pots. Excessive water loss from the plantlets often

258
leads to damage or death. In many cases, survival of plants after transfer to soil requires acclimatization to changes in humidity.
The formation of adventitious shoots on embryos of Picea abies has been described [2]. About 70% of the shoots were vitrified, although most of them only slightly. The percentage of vitrified shoots could be reduced by increasing the agar concentration, but as with Cynara scolymus [8], this lowered the number of adventitious buds and shoots [3, 4]. Therefore, the degree of vitrification obtained on media containing 0.5% agar during the propagation stage had been accepted. The aim of this work was to determine how to manipulate the vitrified adventitious shoots in order to increase the survival of plantlets transferred to pots.
Materials and methods
Plant material
Adventitious shoots were produced from embryos of Picea abies (L.) Karst. according to the method described previously [2]. Newly isolated embryos were cultured on half strength Schenk and Hildebrandt's (SH) medium [13] containing 5 x 10 -4 M of each N6-benzyladenine (BA) and kinetin for one month. Then transferred to half strength SH medium lacking cytokinin. The medium was solidified with 0.5% Bacto agar (Difco). About 1000 slightly vitrified 5 mm long shoots were selected for further studies four to five months after they had been initiated.
Culture procedures
The shoots were transferred to lO0ml Erlenmeyer flasks containing 50ml of half strength SH medium. The medium was gelled with either Bacto agar, (0.5, 1.0, 1.5, 2.0%) or Agarose obtained from Sigma (0.25, 0.50, 0.75, 1.0, 1.5, 2.0%). The Erlenmeyer flasks were closed with cotton wool plugs. If not otherwise stated the shoots were incubated at 200C under 16h light ( 1 - 5 W m -2) with Philips TL 25W 33 white fluorescent tubes. After two months the shoots were given a root-inducing treatment for 24h at 20 *C in the dark in vermiculite which was first autoclaved and then saturated with a sterile filtered (Satorius 0.2/~) water solution containing 10 -4 M IAA and 10 -4 M IBA. After the root-inducing treatment the shoots were washed once in sterile water, and if not otherwise stated, transferred to the same culture conditions as before the treatment. After one to two months the roots were 2 to 5 nun long. Then plantlets were given various acclimatization treatments, as described in the results, before they were transplanted to pots in the greenhouse. The plantlets were planted in pots filled with vermicu- lite and sprayed with water every day. During the first weeks the pots were covered with plastic hoods.

259
Anatomical studies
Leaf samples were fixed in 2.5% glutaraldehyde in 0.1 M cacodylate buffer at 4 °C for at least 24h, then washed once in cacodylate buffer and twice in acetate-veronal buffer. The needles were post fixed with 2% buffered OsO4 for 2 h at 4 °C and washed twice in the buffer. Following dehydration through an ethanol series, samples were embedded in Epon overnight at 20 °C. Sections, 1/am, were stained with basic fuchsin and methylene blue.
Data on percent airspace were based on longitudinal sections of the needles.
Results
Effects o f various agar concentrations
Growth rate. Growth rate of the shoots was greatly influenced by the agar concentration (Table 1 and Figure 1). The shoots grew faster and became
Table 1. Effects of agar concentration in the culture medium on growth of adventitious shoots of Pt'cea abies and their subsequent formation of roots
Agar Degree of Average shoot Rooted shoots Shoots with concentration vitrification length after after four more than (%) after two two months months one root
months (mm) (%) after four months (%)
0.5 +++ 25 87 a 50 a 1.0 ++ 15 27 b 10 b 1.5 + 12 3 e 0 e 2.0 0 7 0 e 0 e
Data are based on averages from 50 to 100 adventitious" shoots from 2 to 4 experiments. Different superscript letters indicate significant differences at the 0.1% level. The cul- tural conditions are described in materials and methods.
more vitrified on lower agar concentrations. Very vitrified shoots on 0.5% agar were dark green and the needles seemed to adhere to each other. While on 2.0% agar the shoots were light green and the needles had a dry appear- ance. Similar results were obtained with agarose (Table 2). Agarose made the medium firmer. Thus, ,concerning the firmness, 0.25% agarose roughly corresponded to 0.5% agar.
Rooting. The rooting ability of the shoots varied depending on the agar concentration used (Table 1). Similar results were obtained irrespective of
whether the shoots after the root.inducing treatment were transferred to the same agar concentration as before or to 1.0% agar. Shoots raised on 1.5 and 2.0% agar would c o n t i n u e t o grow and form roots if they were transferred to lower agar concentrations Within two months; after a longer time, the shoots very rarely started to grow again. The yield of rooted shoots increased
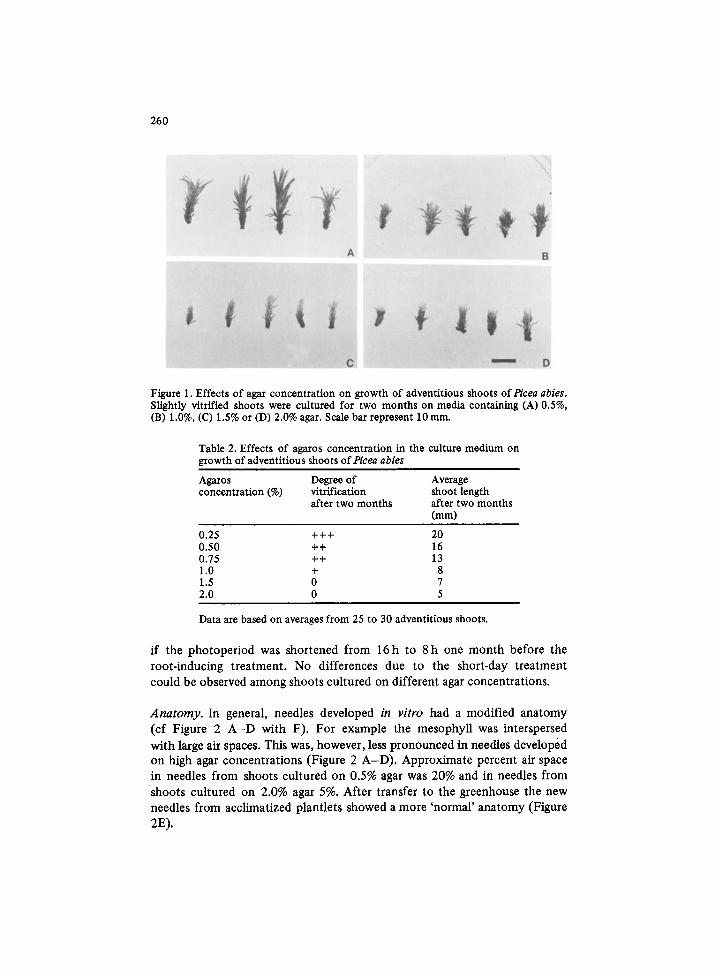
260
Figure 1. Effects of agar concentration on growth of adventitious shoots ofPicea abies. Slightly vitrified shoots were cultured for two months on media containing (A) 0.5%, (B) 1.0%, (C) 1.5% or (D) 2.0% agar. Scale bar represent 10 mm.
Table 2. Effects of agaros concentration in the culture medium on growth of adventitious shoots ofPicea abies
Aga.ros Degree of Average concentration (%) vitrification shoot length
after two months after two months (mm)
0.25 +++ 20 0.50 ++ 16 0.75 ++ 13 1.0 + 8 1.5 0 7 2.0 0 5
Data are based on averages from 25 to 30 adventitious shoots.
if the photoperiod was shortened from 16h to 8 h one month before the root-inducing treatment. No differences due to the short-day treatment could be observed among shoots cultured on different agar concentrations.
Anatomy. In general, needles developed in vitro had a modified anatomy (cf Figure 2 A - D with F). For example the mesophyU was interspersed
with large air spaces. This was, however, less pronounced in needles developed on high agar concentrations (Figure 2 A - D ) . Approximate percent air space in needles from shoots cultured on 0.5% agar was 20% and in needles from shoots cultured on 2.0% agar 5%. After transfer to the greenhouse the new needles from acclimatized plantlets showed a more 'normal ' anatomy (Figure 2E).

261
Figure 2. Microtome section of needles from adventitious shoots and plants of Picea abies, A-D correspond to Figure 1 A-D. E. Needle from plantlets acclimated in the greenhouse. F. Needle f~om a newly flushed plant g~owing in field.
Acclimatization of plantlets produced in vitro
In some preliminary experiments we observed that it was difficult to acclima- tize shoots to nonsterile conditions before the roots had become visible. Furthermore it was easier to obtain root formation on shoots before they were acclimatized than after. Therefore, only rooted shoots have been used in these acclimatization studies.

262
Slightly vitrified plantlets, like the shoots on 1.0% agar in Figure 1, could become acclimatized to survive transplanting to pots under nonsterile con- ditions. This could be carried out either by keeping the plantlets on the same medium for two to three months instead of transferring them to fresh culture medium every month, or by transferring the plantlets to pots contain- ing 1.0% water agar followed by successive lowering the humidity inside the pot by taking off the lid for a longer period each day for about one week. The latter method is time-consuming and care must be taken that the medium does not dry out. Survival by both methods was further improved if the temperature was lowered to about 10 °C.
Very vitrified plantlets, comparable to the shoots on 0.5% agar (Figure 1), never became acclimated unless they were first stimulated to form resting apical buds. When the resting buds resumed growth dry shoots were devel- oped. The older part of the plantlets remained vitrified.
In addition to these acclimatization procedures, the plantlets had to be sprayed with water every day and covered with plastic hoods during the first weeks in the greenhouse. Furthermore, the plantlets withstood the transfer better if they were treated with a commercial evaporation protecting agent (obtained from WeibuU AB, Landskrona, with styren-butadienpolymer as active component) just before the transfer. Under proper cultural con- ditions the plantlets derived in vitro can develop into fast growing plants (Figure 3).
Discussion
These studies have shown that the developmental pattern of adventitious shoots can be manipulated by changing the agar concentration in the culture medium. Increasing the agar concentration from 0.5 to 2.0% decreased vitrification, but at the same time reduced rooting potential and shoot growth. A similar decrease in growth rate with increasing agar concentration has been reported for apical meristems of Picea abies [12] and Dianthus caryophyllus [10], buds of Cynara scolymus [7] and seedlings of Iris [14]. Why higher agar concentrations decrease growth rate is debated. Since agarose at various concentrations gave similar effects to agar, it seems that the effect is not primarily due to impurities in the agar. Romberger and Tabor [12] showed that increasing agar concentration restricted diffusion of macro- molecules. Stoltz [14] found that the decreased growth at higher agar con- centrations was due to reduced water availability. Debergh et al. [8] found that agar is responsible for the matric component of the medium water potential and that changes in the matric potential are responsible for the agar effects. In agreement with their results this study has shown that needles from shoots cultured on low agar concentration (0.5%) had an anatomy resembling helophytic plants (bog plants) while needles developed on higher agar- concentrations were more like that of field-grown Picea plants. Similarly,

263
Figure 3. Adventitious piantlets from an embryo of Picea abies about two yeats after being initiated. Scale bat represent 10 ram.
leaves of tissue-cultured Liquidambar styraciflua L. and Cynara scolymus lacked a differentiated palisade parenchyma and had spongy parenchyma interspersed with large air spaces [16, 8].
The vitrified shoots are genetically similar to the dry shoots because resting buds on vitrified shoots develop into normal dry shoots. Furthermore, the amount of nuclear DNA is the same in dry and vitrified buds [ 11 ].
In order to obtain a high survival, plantlets of Picea abies must first be acclimatized in the culture vessel and then in the greenhouse. Acclimatization has been shown in other species to involve changes in stomatal function [5, 6], the epicuticular wax [ 15] and leaf anatomy [9, 16].
In conclusion it can be postualted that it is not possible to keep the highest propagation rate of embryos of Picea abies, that is on 0.5% agar [3, 4] during the whole culture period, because the shoots became too vitrified. If care is taken to keep the embryos on the surface of the medium, 0.5% agar can be used during the first months in culture. However, when the adventitious shoots are isolated, they have to be transferred to media containing 1% agar. Then they develop into slightly vitrified shoots which

264
can produce roots and later be accl imat ized and t ransplanted to greenhouse
condi t ions.
Acknowledgements
We express our thanks to Karin Bjelke and Ane t te Ax~n for valuable technical
assistance. This research was suppor ted by grants f rom the Swedish Council
for Fores t ry and Agricul tural Research.
References
1. Aitken J, Horgan K, Thorpe T (1981) Influence of explant selection on the shoot- forming capacity of juvenile tissue oft~'nus radiata. Can J For Res 11:112-117
2. Arnold S yon (1982) Factors influencing formation, development and rooting of adventitious shoots from embryos of l~'cea abies (L.) Karst. Plant Sci Lett 27:275-287
3. Arnold S yon, Eriksson T (1979) Induction of adventitious buds of Norway spruce (t~'¢ea abies) gzown in vitro. Physiol Plant 45:29-34
4. Arnold S yon, Eriksson T (1979) Bud induction on isolated needles of Norway spruce (Picea abies L., Karst.) grown in vitro. Plant Sci Lett 15:363-372
5. Brainerd KE, Fuchigami LH (1981) Acclimatization of aseptically cultured apple plants to low relative humidity. J Amer Soc Hort Sci 106:515-518
6. Brainerd KE, Fuchigami LH (1982) Stomatal functioning of in vitro and green- house apple leaves in darkness, mannitol, ABA, and CO2. J Exp Bet 33:388-392
7. Debergh PC (1983) Effects of agar brand and concentration on the tissue culture medium. Physiol Plant 59:270-276
8. Debergh P, Harbaoui Y, Lemeur R (1981) Mass propagation of globe artichoke (Cynara scolymus): Evaluation of different hypotheses to overcome vitrification with special reference to water potential. Physiol Plant 53:181-187
9. Grout B, Aston J (1978) Modified leaf anatomy of cauliflower plantlets regenerated from meristem culture. Ann Bet 42:993-995
10. Hakkaart FA, Versluijs JMA (1983) Some factors affecting glassiness in carnation meristem tip cultures. Neth J Pl Path 89:47-53
11. Hakman I, Arnold S von, Bengtsson A (1984) Cytofluoremettic measurements of nuclear DNA in in vitro regenerated adventitious buds and shoots of Pieea abies (L.) Karst. Physiol Plant 60:321-325
12. Romberger JA, Tabor CA (1971) The l~cea abies shoot apical meristem in culture. I. Agar and autoclaving effects. Amer J Bot 58:131-140
13. Schenk RU, Hildebrandt AC (1972) Medium and techniques for induction and growth of monocotyledonous and dicotyledonous plant cell cultures. Can J Bot 50:199-204
14. Stoltz LP (1971) Agar restriction of the growth of excised mature Iris embryos. J Amer Soc Hort Sci 96:681-682
15. Sutter E, Langhans RW (1982) Formation of epicuticular wax and its effect on water loss in cabbage regenerated from shoot-tip culture. Can J Bot 60:2896- 2902
16. Wetzstein HY, Sommer H (1982) Leaf anatomy of tissue-cultured Liquidambar styraciflua (Hamamelidaceae) during acclimatization. Amer J Bot 69:1576-1586


















