
Effects of Peach Cultivar on Enzymatic Browning Following ... · Effects of Peach Cultivar on...
9
Effects of Peach Cultivar on Enzymatic Browning Following Cell Damage from High-Pressure Processing Chukwan Techakanon, †,‡ Thomas M. Gradziel, § and Diane M. Barrett* ,† † Department of Food Science and Technology and § Department of Pomology, University of California, Davis, One Shields Avenue, Davis, California 95616, United States ‡ Faculty of Science and Industrial Technology, Prince of Songkla University, Surat Thani Campus, 31 Makham Tia, Muang Surat Thani, Suratthani 84000, Thailand ABSTRACT: Peach cultivars contribute to unique product characteristics and may affect the degree of browning after high- pressure processing (HPP). Nine peach cultivars were subjected to HPP at 0, 100, and 400 MPa for 10 min. Proton nuclear magnetic resonance ( 1 H NMR) relaxometry, light microscopy, color, polyphenol oxidase (PPO) activity, and total phenols were evaluated. The development of enzymatic browning during refrigerated storage occurred because of damage during HPP that triggered loss of cell integrity, allowing substrates to interact with enzymes. Increasing pressure levels resulted in greater damage, as determined by shifts in transverse relaxation time (T 2 ) and by light micrographs. Discoloration was triggered by membrane decompartmentalization but limited by PPO activity, which was found to correlate to cultivar harvest time (early, mid, and late season). Outcomes from the microstructure, 1 H NMR ,and PPO activity evaluation were an effective means of determining membrane decompartmentalization and allowed for prediction of browning scenarios. KEYWORDS: high pressure, peach cultivar, 1 H-NMR, cell integrity, enzymatic browning 1. INTRODUCTION Peaches are one of the most popular summer fruits consumed in the U.S. and are well-adapted to California production environments. Because they have a relatively short available season, high-pressure processing (HPP) can be an alternative method for preservation of these high-quality fruits that are rich in nutrients, have a desirable texture, and a natural flavor. HPP is a novel advanced process being extensively studied because of its ability to create products that retain their fresh-like attributes, while inducing destruction of microorganisms and inactivating the enzymes that cause deterioration of quality. 1-3 A recent study reported that the market value of high-pressure preserved products had reached more than $3 billion, and among these, fruit and vegetable products ranked as the largest product group. 4 Peach cultivar plays a crucial role in final product characteristics, e.g. color, taste, texture, and flavor, especially after undergoing high-pressure treatment. During HPP treat- ment, elevated pressures have been reported to induce loss of membrane permeability in plant-based materials. 5 Discoloration in HPP-treated peaches occurs as a consequence of the process, and the hypothesis is that cell integrity is the primary determining factor affecting enzymatic browning reactions in refrigerated and stored high-pressure processed peaches. After loss of fruit cell integrity, the enzyme polyphenol oxidase (PPO), initially located in the plastids, and the phenolic substrates, initially located in the vacuole, are allowed to interact, creating brown products. The discoloration of fruits following HPP has been explored extensively in several commodities, e.g., apple puré e, 2 avocado pure ́ e, 6 banana puré e, 7 green beans, 8 guacamole, 9 and pears. 10 This is the first study to correlate loss of cell integrity to enzymatic browning reactions in different peach cultivars after high- pressure treatment. To quantitatively follow changes in cell integrity resulting from HPP application, nuclear magnetic resonance (NMR) and light microscopy were employed. NMR water proton relaxometry is a tool that can be used to detect physiological changes in the various water compartments in a plant tissue. This method has undergone continuous development and has now been applied in a wide range of plant studies, in particular for quantification of cell membrane damage. 5,11 The degree of membrane damage can be determined by observing changes in water proton relaxation behavior (T 2 ) of the vacuolar, cytoplasm, and cell wall compartments. The proton spin- spin (T 2 ) relaxation time is related to properties of water in different locations in the tissue, water content, and the interaction of water with macromolecules. 12 Because the browning reaction is a consequence of membrane rupture, T 2 relaxation time of the vacuolar compartment can be used in predicting the browning scenario of a product. Observation under a light microscope is a useful complement to 1 H NMR for determining cell integrity of peach samples following HPP. In this experiment, cell integrity was observed under a light microscope using the vacuole staining method developed by Admon et al. (1980) 13 Neutral red (NR) dye diffuses across the tonoplast membrane and ionizes in the acidic environment of the vacuole, appearing as an intense red color. Once cell membrane damage occurs, the vacuoles lose their integrity, thus resulting in the dye spreading throughout the Received: April 25, 2016 Revised: September 9, 2016 Accepted: September 14, 2016 Published: September 14, 2016 Article pubs.acs.org/JAFC © 2016 American Chemical Society 7606 DOI: 10.1021/acs.jafc.6b01879 J. Agric. Food Chem. 2016, 64, 7606-7614
Transcript of Effects of Peach Cultivar on Enzymatic Browning Following ... · Effects of Peach Cultivar on...

Effects of Peach Cultivar on Enzymatic Browning Following CellDamage from High-Pressure ProcessingChukwan Techakanon,†,‡ Thomas M. Gradziel,§ and Diane M. Barrett*,†
†Department of Food Science and Technology and §Department of Pomology, University of California, Davis, One Shields Avenue,Davis, California 95616, United States‡Faculty of Science and Industrial Technology, Prince of Songkla University, Surat Thani Campus, 31 Makham Tia, Muang SuratThani, Suratthani 84000, Thailand
ABSTRACT: Peach cultivars contribute to unique product characteristics and may affect the degree of browning after high-pressure processing (HPP). Nine peach cultivars were subjected to HPP at 0, 100, and 400 MPa for 10 min. Proton nuclearmagnetic resonance (1H NMR) relaxometry, light microscopy, color, polyphenol oxidase (PPO) activity, and total phenols wereevaluated. The development of enzymatic browning during refrigerated storage occurred because of damage during HPP thattriggered loss of cell integrity, allowing substrates to interact with enzymes. Increasing pressure levels resulted in greater damage,as determined by shifts in transverse relaxation time (T2) and by light micrographs. Discoloration was triggered by membranedecompartmentalization but limited by PPO activity, which was found to correlate to cultivar harvest time (early, mid, and lateseason). Outcomes from the microstructure, 1H NMR ,and PPO activity evaluation were an effective means of determiningmembrane decompartmentalization and allowed for prediction of browning scenarios.
KEYWORDS: high pressure, peach cultivar, 1H-NMR, cell integrity, enzymatic browning
1. INTRODUCTION
Peaches are one of the most popular summer fruits consumedin the U.S. and are well-adapted to California productionenvironments. Because they have a relatively short availableseason, high-pressure processing (HPP) can be an alternativemethod for preservation of these high-quality fruits that are richin nutrients, have a desirable texture, and a natural flavor. HPPis a novel advanced process being extensively studied because ofits ability to create products that retain their fresh-likeattributes, while inducing destruction of microorganisms andinactivating the enzymes that cause deterioration of quality.1−3
A recent study reported that the market value of high-pressurepreserved products had reached more than $3 billion, andamong these, fruit and vegetable products ranked as the largestproduct group.4
Peach cultivar plays a crucial role in final productcharacteristics, e.g. color, taste, texture, and flavor, especiallyafter undergoing high-pressure treatment. During HPP treat-ment, elevated pressures have been reported to induce loss ofmembrane permeability in plant-based materials.5 Discolorationin HPP-treated peaches occurs as a consequence of the process,and the hypothesis is that cell integrity is the primarydetermining factor affecting enzymatic browning reactions inrefrigerated and stored high-pressure processed peaches. Afterloss of fruit cell integrity, the enzyme polyphenol oxidase(PPO), initially located in the plastids, and the phenolicsubstrates, initially located in the vacuole, are allowed tointeract, creating brown products. The discoloration of fruitsfollowing HPP has been explored extensively in severalcommodities, e.g., apple puree,2 avocado puree,6 bananapuree,7 green beans,8 guacamole,9 and pears.10 This is thefirst study to correlate loss of cell integrity to enzymatic
browning reactions in different peach cultivars after high-pressure treatment.To quantitatively follow changes in cell integrity resulting
from HPP application, nuclear magnetic resonance (NMR) andlight microscopy were employed. NMR water protonrelaxometry is a tool that can be used to detect physiologicalchanges in the various water compartments in a plant tissue.This method has undergone continuous development and hasnow been applied in a wide range of plant studies, in particularfor quantification of cell membrane damage.5,11 The degree ofmembrane damage can be determined by observing changes inwater proton relaxation behavior (T2) of the vacuolar,cytoplasm, and cell wall compartments. The proton spin−spin (T2) relaxation time is related to properties of water indifferent locations in the tissue, water content, and theinteraction of water with macromolecules.12 Because thebrowning reaction is a consequence of membrane rupture, T2relaxation time of the vacuolar compartment can be used inpredicting the browning scenario of a product.Observation under a light microscope is a useful complement
to 1H NMR for determining cell integrity of peach samplesfollowing HPP. In this experiment, cell integrity was observedunder a light microscope using the vacuole staining methoddeveloped by Admon et al. (1980)13 Neutral red (NR) dyediffuses across the tonoplast membrane and ionizes in the acidicenvironment of the vacuole, appearing as an intense red color.Once cell membrane damage occurs, the vacuoles lose theirintegrity, thus resulting in the dye spreading throughout the
Received: April 25, 2016Revised: September 9, 2016Accepted: September 14, 2016Published: September 14, 2016
Article
pubs.acs.org/JAFC
© 2016 American Chemical Society 7606 DOI: 10.1021/acs.jafc.6b01879J. Agric. Food Chem. 2016, 64, 7606−7614

cell; therefore, it appears less intensely red. The use of this dyeallows for discriminating intact vacuoles from damaged,nonintact ones. The number and color intensity of intactvacuoles represents the level of damage to the cells. Thus, thecomparison of micrographs of samples from different peachcultivars processed using different pressure treatments can becorrelated to the degree of decompartmentalization.The objectives of this study were to determine the effect of
peach cultivar on cell integrity after high-pressure processing atdifferent pressure levels (0, 100, and 400), using NMR andmicroscopic studies, and to correlate that to the development ofbrown color, as affected by PPO activity and total phenolscontent. Phenotypic differences in peach cultivars, such astexture and browning susceptibility, may be useful in definingwhich cultivars are best suited for preservation by HPP.
2. MATERIALS AND METHODS2.1. Raw Materials. The clingstone type peach cultivars, Carson,
Andross, Ross, Evert, Dr. Davis, Late Ross, Halford, and Lilleland, andthe freestone type, Summerset, were harvested by hand fromFoundation Plant Services, at the University of California, Davis,CA. All cultivars were of the same variant (clingstone nonmeltingvariant) except for Summerset, which was a freestone melting type.Peaches were harvested at 157−164 days after bloom and then sortedto a firmness of 35−40 N using a TA-XT2 Texture Analyzer (StableMicro Systems Ltd., Surrey, U.K.) and stored at 4 °C RH 84% forapproximately 2 days until processing. In each replicate, three fruits ofthe same cultivar were hand peeled, sliced into approximately 3 cmthickness and placed into polyethylene bags (4 mil vacuum pouches,Ultrasource, MO, USA). Each bag contained 3 peach slices of the samecultivar, one from each of the three different fruits, and separate bagswere created for each analysis method. In addition, approximately 2mL of peach extracts (see description below) were vacuum packed inpolyethylene bags. The samples were kept at ambient temperature (22± 2 °C) for 30 min after packaging prior to high-pressure treatment.On each of three replicate days of processing, each peach cultivar wasprocessed at each of two pressures (100 and 400 MPa). The controltreatment was an unprocessed, sliced peach sample in a vacuumpackage (approximately 0 MPa). The same fruit was analyzed for allparameters, i.e., difference in lightness, the T2 relaxation time usingNMR, PPO activity, total phenols, and determination of viable cellsusing light microscopy.2.2. Experimental Design. Nine peach cultivars were used in this
study, and each was high-pressure treated at 100 and 400 MPa. Non-HPP treated samples (0 MPa) were used as controls. The HPPprocessing for this entire experiment was three distinct times, whichoccurred on three separate days. On each replicate processing day,three fruit per cultivar were used. Each fruit was sliced into 3 cm thickslices, on the two sides of the pit, and then subdivided into five parts.Each of the five parts was used for a different analytical measurement,i.e., color, NMR (paramagnetic analysis of protons), PPO activity, totalphenols, and light microscopy. Prior to HPP treatment, slices werepackaged according to analytical method, so only one package neededto be opened prior to each analytical method. The entire processdesign was repeated three separate times; therefore, in total, ninesamples per cultivar were analyzed for each of the five analyticalparameters.2.3. High-Pressure Processing. The packaged samples were
processed at 100 and 400 MPa for 10 min in a high-pressureprocessing unit model QFP 2L-700 (Avure Technologies Inc., Kent,WA). The high-pressure unit uses water as a pressurizing medium andhas a maximum pressure level of 600 MPa. The initial temperature inthe chamber (Ti) was around 23 °C. The maximum temperature in thehigh-pressure chamber was dependent on the set pressure, which for100 and 400 MPa was 25 and 33 °C, respectively.2.4. Nuclear Magnetic Resonance Relaxometry. Following
HPP, each sample slice was cut into a cylindrical piece using a corkborer with a 15 mm diameter and 15 mm height. The cylindrical piece
was blotted dry then placed into a covered NMR tube, which wasplaced in a plastic sample holder of the NMR unit. Measurements ofNMR relaxometry were performed using a NMR spectrometer(Aspect AI, Industrial Area Havel Modi’in, Shoham, Israel) with amagnetic field of 1.02 T and frequency of 43.5 MHz. The Carr−Purcell−Meiboom−Gill (CPMG) sequence with an echo time of 0.5ms and 15 000 echoes was used to obtain T2 relaxation decay curve. T2spectrum inversion was performed by non-negative least squares usingProspa (Magritek, Wellington, New Zealand).
2.5. Light Microscopy Study. 2.5.1. Section Preparation. Afterhigh-pressure treatment, two pieces with a rectangular shape ofapproximately 1.0 × 0.5 × 0.3 cm3 were obtained from each slicedsample and placed in a sample holder. Peach specimens approximately200 μm thick were obtained using a Vibratome 1000 Plus (TheVibratome Co., St. Louis, MO) before being submerged for 2 h in thestaining solution. The solution was prepared using 0.5% neutral red inan acetone stock solution filtered twice with Whatman paper # 1 anddiluted to 0.04% in 0.55 M mannitol−0.01 M N-(2-hydroxyethyl)-piperazine-N′-2-ethane-sulfonic acid (HEPES) buffer, pH 7.8. Peachsections were rinsed for 0.5 h in the 0.55 M mannitol−0.01 M HEPESbuffer solution before being mounted on a microscope slide andcovered with a coverslip.
2.5.2. Microscopic Observation. Stained specimens were observedat 40× magnification using a light microscope (Olympus SystemMicroscope, Model BHS, Shinjuku-Ku, Tokyo, Japan). Colorphotomicrographs were captured by a digital color camera (OlympusMicroFire, Olympus, Tokyo, Japan) attached to the microscope usingOlympus software (Olympus America, Melville, NY).
2.5.3. Image Processing and Analysis. Micrographs were analyzedusing ImageJ (NIH, U.S.), an image processing software. Fifteenmicrographs were randomly selected from three replicates for eachpeach cultivar following different treatments: control (0 MPa) andHPP at 100 and 400 MPa. A cell counter plug-in developed by de Vos(2008)14 was used to semiquantify the number of viable cells, whichwere distinguished by smooth pink to red stained cells, while inviablecells appear in a rough membrane without retained dye.
2.6. Degree of Browning (Difference in Lightness). Thedegree of browning of peach samples was determined as the differencein lightness (DL*), which is the difference between initial lightnessand the lightness of the sample after storage at 4 °C for 2 weeks. Thelightness was monitored using a Minolta CR-400 colorimeter (Minoltacamera Co, Ltd., Japan) with a beam diameter of 11 mm and a viewingangle of 0°. The values were expressed by the CIE L*a*b* system. Awhite tile used for calibration has L* = 96.88, a* = 0.02, and b* = 2.05.
2.7. Partial Purification of Peach Polyphenol OxidaseExtracts. The method for protein partial purification using TritonX-114 by Espin et al. (1995)15 was used with some modifications toobtain peach extracts. A peach sample (200 g) was homogenized with100 mL of cold solution of 0.1 M sodium phosphate dibasic anhydrous(Fisher, Fair Lawn, NJ) (pH 7.3), 6% (w/v) Triton X-114 (Sigma-Aldrich, St. Louis, MO) and 20 mM EDTA (Fisher, Fair Lawn, NJ) for2 min. The mixture was refrigerated for 60 min at 4 °C thencentrifuged with a Sorvall-RC5 (E. I. DuPont Co, Wilmington, DE) at4 °C and 28 373.6g for 45 min. The supernatant was mixed with 8%(w/v) of the surfactant Triton X-114 before it was incubated in a 40°C water bath for 15 min, after which a change to opaque yellowishwas observed. After centrifugation at 578.6g for 10 min at 25 °C, adetergent-rich phase was discarded, and the clear supernatant wascollected. A second phase partitioning step with 8% (w/v) Triton X-114 was performed, then it was incubated again in a 40 °C water bathfor 15 min. The enzyme extract was collected from the supernatantafter being centrifuged at 578.6g for 10 min at 25 °C and was stored at−10 °C until use.
2.8. Polyphenol Oxidase Assay. PPO activity was assayed usingthe spectrophotometric method described by Espin et al. (1995)15
with some modifications. This method uses 3-methyl-2-benzothiazo-linone (MBTH) as a chromogenic coupling agent, which reacts withthe quinone products obtained from the oxidation of diphenol withPPO. Assays began by adding 10 μL of peach polyphenol oxidaseextract to 1.0 mL of a medium containing 0.6 mL of 100 mM acetate
Journal of Agricultural and Food Chemistry Article
DOI: 10.1021/acs.jafc.6b01879J. Agric. Food Chem. 2016, 64, 7606−7614
7607
buffer (pH 5.5), 0.2 mL of 2.5 mM MBTH (Sigma-Aldrich, St.Louis,MO) and 0.2 mL of 25.0 mM dihydroxyhydrocinnamic acid (Sigma-Aldrich, St.Louis, MO). The adduct formed from this reactionappeared to be reddish in color. PPO activity was determined by anincrease in absorbance at 500 nm using ultraviolet (UV)spectrophotometry (UV2101PC, Shimadzu Scientific InstrumentsInc., Columbia, MD). The activity calculation was performed usingthe following equation, where Abs(0) is the initial absorbance andAbs(1) is absorbance at the end of linearity.
= − ·PPO Activity (units/mL) Abs(1) Abs(0)/min mL of juice
2.9. Analysis of Total Phenols. Analysis of total phenols in peachsamples was performed by the Folin−Ciocalteu method described byWaterhouse (2002)16 with some modifications. The analysis measuresthe total concentration of phenolic hydroxyl groups in plant extractsbased on reduction of the reagent (Folin−Ciocalteu reagent). Theformation of a blue complex product can be measured at 760 nm, andgallic acid (Arcos Organics, Geel, Belgium) in a range of 0−500 mg/Lis used for calibration of the standard curve. A 20 g peach sample washomogenized with 30 mL of deionized water, then 6.4 g of thehomogenate was blended with 27.6 mL of 76% (v/v) aqueous acetonefor 2 min. The solution was placed in a shaker for 10 min for furthermixing, after which the cell wall particles were separated using acentrifuge (Centra CL2 tabletop centrifuge, IEC, Needham, MA) at578.6g for 10 min. Then 1 mL of the supernatant was vortexed with0.36 mL of 2N Folin reagent (Sigma-Aldrich, Buchs, Switzerland) andallowed to stand for 5 min. After this, 7.5% (w/v) sodium carbonate (6mL) was added to the solution, which was mixed well before 2.64 mLof deionized water was added. The solution was homogenized againand incubated in a 50 °C water bath for 5 min. After the solution was
cooled to room temperature for 1 h, the absorbance was monitored at760 nm. The results were expressed as gallic acid equivalents (GAE)/fresh weight of peaches (g).
2.10. Statistical Analysis. This experiment was carried out inthree replicate processing runs which occurred on three separate days.The effect of peach cultivar on the T2 relaxation time, percent water,percent viable cells, difference in lightness, PPO activity of the intactfruit, PPO activity of the extract, and total phenols content wereanalyzed using analysis of variance for each cultivar and processingpressure level. Fisher’s least significant difference (LSD) test was usedto compare means of each peach cultivar processed at differentpressure levels at p < 0.05 (SAS version 9.4, Cary, NC). The plotspresent the mean with its standard deviation for each determination.
3. RESULTS AND DISCUSSION
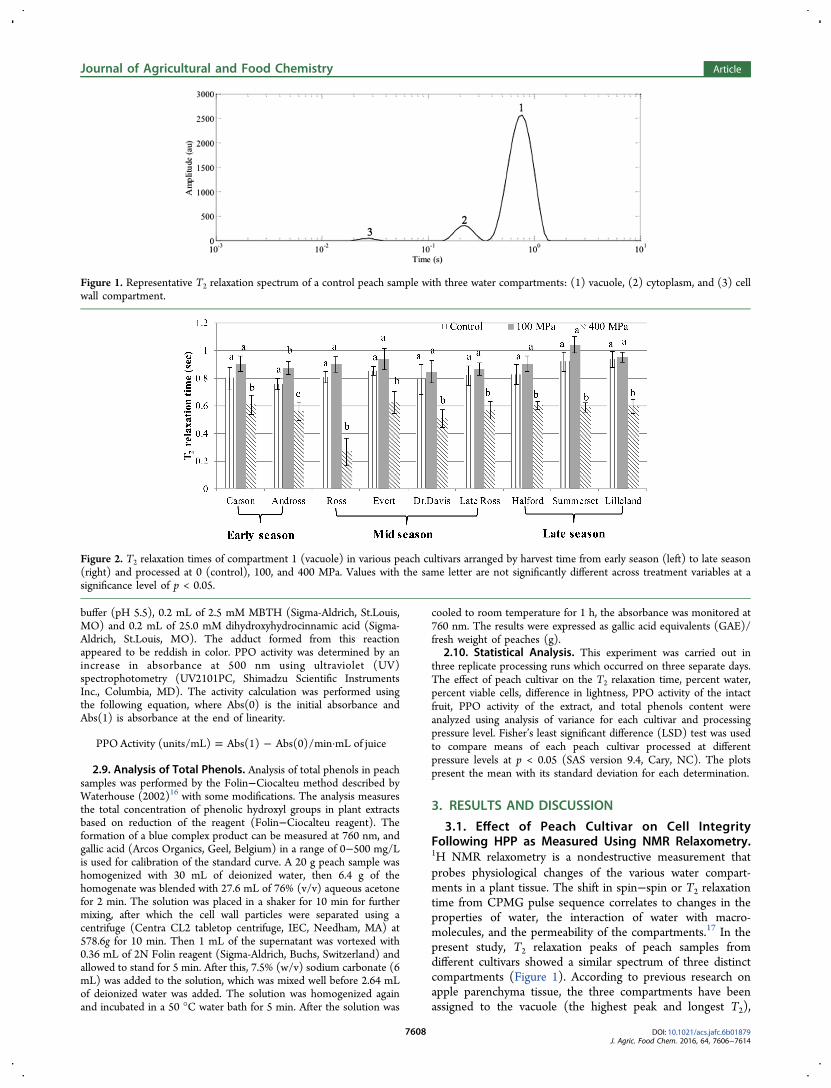
3.1. Effect of Peach Cultivar on Cell IntegrityFollowing HPP as Measured Using NMR Relaxometry.1H NMR relaxometry is a nondestructive measurement thatprobes physiological changes of the various water compart-ments in a plant tissue. The shift in spin−spin or T2 relaxationtime from CPMG pulse sequence correlates to changes in theproperties of water, the interaction of water with macro-molecules, and the permeability of the compartments.17 In thepresent study, T2 relaxation peaks of peach samples fromdifferent cultivars showed a similar spectrum of three distinctcompartments (Figure 1). According to previous research onapple parenchyma tissue, the three compartments have beenassigned to the vacuole (the highest peak and longest T2),
Figure 1. Representative T2 relaxation spectrum of a control peach sample with three water compartments: (1) vacuole, (2) cytoplasm, and (3) cellwall compartment.
Figure 2. T2 relaxation times of compartment 1 (vacuole) in various peach cultivars arranged by harvest time from early season (left) to late season(right) and processed at 0 (control), 100, and 400 MPa. Values with the same letter are not significantly different across treatment variables at asignificance level of p < 0.05.
Journal of Agricultural and Food Chemistry Article
DOI: 10.1021/acs.jafc.6b01879J. Agric. Food Chem. 2016, 64, 7606−7614
7608

cytoplasm (the second compartment), and cell wall (theshortest T2).
17 Figure 2 illustrates that most of the cultivarsshowed the same pattern, with an increasing T2 in the vacuolarcompartment after 100 MPa, which was significant (p < 0.05)in only the Andross cultivar. A previous study by our group onHPP treated onions also found the same increase in T2relaxation time of samples following treatment at 100 MPa.5
A hypothesis for this finding is that a firmer structure resultedfrom a release of the enzyme pectin methylesterase (PME)from the cellular matrix to interact with its pectin substrate. De-esterified pectin created from the interaction can form divalentbonds with either endogenous Ca2+ or Mg2+, allowing the cellto withstand mild pressure treatments.18
A sharp decrease in T2 of the main compartment after the400 MPa treatment was observed in every peach cultivar. Thisdecrease in T2 indicates a change in the proton environment,which may result from a loss in cell integrity, allowing thetransfer of the water between different compartments. TheRoss cultivar had the greatest difference in T2 between thecontrol and 400 MPa. This decrease in T2 relaxation time afterHPP treatment implies a change in the proton environmentwhich may result from a loss of compartmentalization due tomembrane rupture.19 A similar observation of a decrease in T2of the main compartment was also observed in onion tissuesfollowing freeze−thaw (−18 °C), thermal processing (60, 70,and 90 °C), and/or high-pressure processing of onions at 200MPa with an initial temperature of 20 °C.5 These findingsindicate an increased interaction of the water between differentcompartments due to loss of cell integrity as a result of cellmembrane damage.
The relative signal intensity or amplitude of each peakcorrelates to the percent water in each compartment, whichallows for the determination of the change in membraneintegrity. According to the percent water results in Table 1, thechanges in signal intensity of compartment 1 (vacuole)demonstrated the same trend as the T2 relaxation time. Inmost cases, treatment at 100 MPa had no significant effect onsignal intensity or T2 relaxation time (p < 0.05). HPP treatmentat 400 MPa, however, induced a loss in membrane integrity asobserved by the decrease in both percent water (Table 1) andT2 relaxation time (Figure 2). The loss in membrane integrityas a result of pressure treatment caused more water protoninteraction (decreased T2 relaxation time) and water transferfrom the vacuolar compartment to the other water compart-ments (decrease in percent water). A comparison between thedifferent cultivars in terms of the change in amplitude of thevacuolar peak in the 400 MPa treated samples (Figure 2), ascompared to control samples, revealed that they could beclassified into two different groups. The first group had less(10−13%) change in amplitude and included the cultivarsCarson, Andross, Dr. Davis, and Late Ross, while the secondgroup had a greater change, in the range of 15−23%, andconsisted of the cultivars Ross, Evert, Halford, Lilleland, andSummerset.Complementary to the loss of intensity in compartment 1,
the signal intensity of compartments 2 and 3 were found toincrease significantly at a significance level of p < 0.05 afterprocessing at 400 MPa. The reduction in water in compartment1 (vacuole) after the 400 MPa treatment, simultaneous with anincrease in the signal intensity of compartment 2 (cytoplasm)
Table 1. Percent Water in the Vacuolar, Cytoplasmic, and Cell Wall Compartments of Various Peach Cultivars FollowingDifferent Pressure Treatments (0, 100, and 400 MPa)
cultivar pressure (MPa) compartment 1 (vacuole) compartment 2 (cytoplasm) compartment 3 (cell wall)
Carson 0 86.76 ± 0.66 a 12.02 ± 0.66 b 1.22 ± 0.22 c100 87.11 ± 2.54 a 10.10 ± 1.87 b 2.79 ± 0.84 b400 75.16 ± 4.01 b 20.36 ± 2.99 a 4.48 ± 1.38 a
Andross 0 83.57 ± 2.44 a 14.32 ± 2.29 b 2.12 ± 0.48 c100 83.94 ± 1.33 a 12.43 ± 1.26 b 3.63 ± 0.31 b400 71.85 ± 3.05 b 22.52 ± 2.29 a 5.63 ± 1.07 a
Ross 0 82.93 ± 1.60 a 14.85 ± 1.88 b 2.23 ± 0.64 b100 85.16 ± 2.12 a 11.88 ± 1.80 b 2.96 ± 0.40 b400 67.93 ± 4.85 b 26.38 ± 3.89 a 5.69 ± 1.32 a
Evert 0 84.51 ± 1.72 a 13.26 ± 1.29 b 2.23 ± 0.59 c100 82.71 ± 3.05 a 11.70 ± 2.81 b 4.85 ± 1.55 a400 65.37 ± 4.92 b 27.23 ± 3.88 a 7.40 ± 1.47 b
Dr. Davis 0 79.72 ± 3.54 b 18.41 ± 3.16 b 1.87 ± 0.51 c100 85.16 ± 2.40 a 11.25 ± 1.97 c 3.59 ± 0.60 b400 69.15 ± 4.25 c 25.35 ± 3.45 a 5.50 ± 1.30 a
Late Ross 0 80.75 ± 3.18 a 17.46 ± 3.32 b 1.79 ± 0.53 c100 82.08 ± 3.91 a 13.57 ± 2.90 c 4.35 ± 1.11 b400 68.06 ± 3.73 b 25.52 ± 3.09 a 6.42 ± 1.01 c
Halford 0 86.51 ± 1.78 a 11.28 ± 1.60 b 2.21 ± 0.57 c100 86.72 ± 1.30 a 9.64 ± 0.96 b 3.64 ± 0.58 b400 63.26 ± 4.57 b 29.25 ± 4.42 a 7.49 ± 0.82 a
Lilleland 0 83.43 ± 2.08 a 14.98 ± 2.20 b 1.59 ± 0.81 b100 86.14 ± 2.98 a 10.97 ± 1.74 c 2.89 ± 1.53 b400 67.19 ± 3.72 b 27.20 ± 3.11 a 5.60 ± 1.15 a
Summerset 0 86.97 ± 2.00 a 11.12 ± 2.11 b 1.92 ± 0.43 b100 89.38 ± 1.47 a 7.85 ± 0.95 c 2.77 ± 0.62 b400 71.59 ± 5.09 b 22.78 ± 3.46 a 5.63 ± 2.15 a
aValues with the same letter are not significantly different across treatment variables at a significance level of p < 0.05.
Journal of Agricultural and Food Chemistry Article
DOI: 10.1021/acs.jafc.6b01879J. Agric. Food Chem. 2016, 64, 7606−7614
7609

and compartment 3 (cell wall) at the same pressure level,indicates the transfer of water from the vacuole to thecytoplasm and cell wall. Similar findings of water redistributionwere also reported in other plants where damage had occurred,e.g., pomegranate with black heart disease,20 potatoes HPPtreated at 300 MPa,21 and pears with internal browning.22 In allcases, water exchange was found to occur between the vacuolarand cytoplasmic and/or cell wall compartments. This waterredistribution between the compartments may allow phenolicsubstrates initially located in the vacuole,23 and the polyphenoloxidase enzyme, initially located in the plastids,24 to meet in thecytoplasm and produce browning products as a consequence oftheir interaction.25 In our previous study, the measurement ofT2 relaxation time was in agreement with microstructuralchanges determined using light microscopy, and bothcorrelated to the development of brown color following HPP.26
3.2. Evaluation of Number of Viable Cells in VariousPeach Cultivars after HPP Using Light Microscopy. In allpeach cultivars, intact vacuoles appeared only in control andthose samples that were mildly treated at 100 MPa (Table 2).
Of the nine cultivars, Lilleland had the highest initial percentageof viable cells with 71%, while Ross had the lowest count ofviable cells at 30%. Under light microscopy, the stained sections
from all of the nine peach cultivars still showed viable cellsfollowing HPP at 100 MPa; however, the percentage of viablecells decreased significantly as compared to unprocessedcontrol samples in most peach cultivars, except for Evert andRoss. In fact, the cultivars Ross, Evert, Late Ross, and Halfordhad less than a 10% reduction in viable cell count between thecontrol and the 100 MPa samples. There is a strong possibilitythat the cellular structure of these cultivars is relatively strong,such that they can withstand the treatment at 100 MPa. In our400 MPa treated peach samples, however, no viable cells wereobserved in any of the cultivars.A study on Chenopodium rubrum (red goosefoot) cells by
Dornenburg and Knorr (1993)27 indicated that these cellscould resist high-pressure treatments at 100 MPa, but acomplete loss of cell viability occurred at 110 MPa. However,Basak and Ramaswamy (1998)18 reported complete rupture ofthe plant cellular membranes following HPP at pressures of 200MPa and above. The number of viable cells retained after HPPmay correlate to how strong the plant structure is. If themembranes are not ruptured by the treatment, there is lesspossibility for interaction between the enzyme and substrates,and as a consequence, a lower degree of browning. In thecurrent study, as will be discussed below, the Evert, Dr. Davis,and Late Ross peach cultivars also showed less of a difference inlightness, even after processing at 400 MPa.
3.3. Degree of Browning in Peach Cultivars FollowingHPP Treatment. During HPP treatment, damage to cellmembranes can trigger the development of discoloration duringstorage, resulting from enzymatic browning reactions. Aftermild HPP treatment at 100 MPa, the difference in lightness wasminimal and not significantly different (p < 0.05) from thecontrol in most of the peach cultivars except for Late Ross andHalford (Figure 3). In a study on HPP onion treatments byButz et al. (1994),28 they reported that pressures up to 100MPa did not trigger browning reactions because the tonoplastmembrane surrounding the vacuole was not disrupted. In thecurrent study, the difference in color developed morepronouncedly in most cultivars after processing at 400 MPa,in accordance with the hypothesis that interaction ofpolyphenol oxidase and phenolic substrates took place in thecytoplasm after the membranes ruptured. This hypothesis hasalso been supported by findings from previous studies by Knorr
Table 2. Percent Viable Cells in Various Peach CultivarsFollowing Different Pressure Treatments (0 and 100 MPa)
% viable cells
control 100 MPa% decrease after
processing
Carson 32.71 ± 3.70 a 16.37 ± 6.71 b 16.34Andross 51.02 ± 6.09 a 27.85 ± 3.93 b 23.17Ross 29.65 ± 5.37 a 25.65 ± 6.82 a 3.99Evert 37.95 ± 4.48 a 38.59 ± 6.61 a 0.00Dr. Davis 55.38 ± 5.36 a 41.74 ± 6.22 b 13.64Late Ross 46.22 ± 2.37 a 39.04 ± 4.18 b 7.17Halford 38.59 ± 3.09 a 28.36 ± 5.26 b 10.23Lilleland 70.53 ± 6.32 a 35.17 ± 6.37 b 35.35Summerset 46.02 ± 6.52 a 29.02 ± 9.66 b 17.00
aValues with the same letter are not significantly different acrosstreatment variables at a significance level of p < 0.05.
Figure 3. Difference in lightness of various peach cultivars following HPP treatment at 0 (control), 100, and 400 MPa and 2 weeks of refrigeratedstorage. Values with the same letter are not significantly different across treatment variables at a significance level of p < 0.05.
Journal of Agricultural and Food Chemistry Article
DOI: 10.1021/acs.jafc.6b01879J. Agric. Food Chem. 2016, 64, 7606−7614
7610

(1993),29 Basak and Ramaswamy (1998),18 and Gonzalez andBarrett (2010),30 which have been mentioned previously.The ordering of the peach cultivars from left to right, as
presented in Figure 3, is based on the time of harvest. Thecultivars Carson and Andross are in the early season group,which is harvested in July. Ross, Evert, Dr. Davis, and Late Rosspeaches are harvested in the mid season in August, whileHalford, Summerset, and Lilleland are late season fruitharvested in early September. According to our results for thebrowning of different cultivars, the discoloration trend was inagreement with the harvest season. That is, the early seasoncultivars (Carson and Andross) are more susceptible to damagethan peaches from the other season, to a large degree becauseof their early maturity (T. M. Gradziel, personal communica-tion). Because peaches in this group develop so early andrapidly, a weaker endocarp and flesh structure is generallyfound. The midseason group, which includes Ross, Evert, Dr.Davis, and Late Ross, on the other hand, has sufficient time tofully develop; therefore, the fruit retain greater integrity thanthe other groups and thus are normally preferred forprocessing. The brown discoloration in this group was relativelylow compared to that in the early and late season peaches. Thelate season group, which consists of Lilleland and Summerset,
develop during a time of high carbohydrate source-sinkcompetition, when the trees are finalizing flower bud initiationand preparing for dormancy. The Lilleland and Summersetpeaches were thus very susceptible to damage and showed thehighest degree of browning.31
3.4. Effect of Peach Cultivar and HPP Treatment onPPO Activity. In the present study, PPO activity from both theintact fruit (Figure 4) and the extract (Figure 5) variedsignificantly between the different cultivars. The main differ-ence between the PPO activity from the intact fruit and thatfrom the extract is the presence of phenolic compounds in thefruit sample, whereas the phenolic substrates were removedfrom the extracts by a phase partitioning separation prior toHPP. Removal of the phenolic compounds from the extractsprohibited potential product inhibition by quinones andtherefore loss of PPO activity and/or turnover of enzyme.
3.4.1. PPO Activity in Whole Fruit. The trend in PPOactivity in the whole fruit after high-pressure treatment wasdifferent among cultivars (Figure 4). The initial PPO activitywas highest in the cultivar Andross, with a mean value of 47 361units/mL, and the cultivar Halford had the lowest activity witha mean of 7 437 units/mL. At a pressure level of 100 MPa, thecultivars Carson and Ross showed an increase in PPO activity
Figure 4. PPO activity of various cultivars of peach fruit following HPP treatment at 0 (control), 100, and 400 MPa. Values with the same letter arenot significantly different across treatment variables within each cultivar at a significance level of p < 0.05.
Figure 5. PPO activity of various peach cultivar extracts following HPP treatment at 0 (control), 100, and 400 MPa. Values with the same letter arenot significantly different across treatment variables at a significance level of p < 0.05.
Journal of Agricultural and Food Chemistry Article
DOI: 10.1021/acs.jafc.6b01879J. Agric. Food Chem. 2016, 64, 7606−7614
7611

of 48% and 55%, respectively. In contrast, the PPO activity ofDr. Davis and Late Ross decreased significantly at the samepressure level, while the rest of the cultivars had a stable level ofenzyme activity. There was no notable effect of seasonality onPPO activity from fruit.After HPP treatment at 400 MPa, most peach cultivars
showed a reduction in PPO activity, except for the cultivarHalford. In this case, the decrease in activity may have beencaused by either product inhibition due to the presence ofphenolic compounds and/or turnover of the enzyme.Considering the first factor, the hypothesis of productinhibition is supported by the observation of a high degree ofdiscoloration in the HPP treated samples, which at the sametime had relatively low PPO activity. After the interactionbetween PPO and phenolic substrates, o-quinones aregenerated. These quinone products have been reported toaffect PPO activity by their interaction with proteins, afterwhich they may undergo covalent condensation, which wouldnegatively affect protein structure and function.32−36 Inaddition, Felton et al. (1992)37 suggested that quinoneproducts can damage proteins, amino acids, or lipids bygenerating free radicals through redox-recycling.The second hypothesis related to PPO reduction is due to
enzyme turnover. The active site of PPO contains twohomologous subunits, each of which has a copper ioncoordinated with three conserved histidine residues.38,39 Afterloss of cell integrity as a result of HPP, the oxidation processoccurs due to the interaction between PPO and its substrates inthe presence of O2. At this point, PPOs have been reported tolose histidine residues, in particular after free-radical-catalyzedfragmentation occurs at the active site of the enzyme. Theenzyme is therefore irreversibly inactivated as a result of losinghistidine, due to the release of copper at the active site.40−42 Aprevious study reported inactivation of mushroom PPO at arate of approximately 1 in 5000 turnovers of the substrate tothe product.40
3.4.2. PPO Activity in Partially Purified Peach Extracts. ThePPO activity from peach extracts behaved more in agreementwith the observed development of browning and the harvestseason. The early season peaches (Carson and Andross, whichwere harvested in July) had relatively high PPO activity (Figure5), have a weaker endocarp because they develop so early and
rapidly, and exhibited a high degree of enzymatic browning(Figure 3). The midseason peaches (Ross, Evert, Dr. Davis, andLate Ross, which were harvested in August) had the lowestPPO activity and can retain their cell integrity better becausethey have sufficient time to develop; therefore, they showed thelowest difference in lightness. The late season cultivars(Halford, Summerset, and Lilleland, which are harvested inearly September) had relatively high PPO activity and alsodemonstrated a very high difference in lightness.In the present study, at the pressure levels of 100 and 400
MPa, PPO activity in the peach extracts was unaffected in abouthalf of the peach cultivars, i.e., Carson, Evert, Dr. Davis,Summerset, and Lilleland. PPO has been reported to be one ofthe most pressure-resistant enzymes, although its sensitivityvaries in different commodities.43 Several studies on PPO inmushrooms and potatoes have reported that treatment at 800MPa and higher was required for inactivation.44,45
High-pressure treatment has been reported to affect proteinstructure, which in this case is the conformation of PPO.Previous authors have found that the primary structure of theprotein, which is linked together with covalent bonds, is largelyunaffected by high pressure, whereas the secondary structure,which is maintained by H-bonds, is affected at high pressuresabove 200 MPa.43,46 These same authors also observed changesin the tertiary structure, which is dominated by hydrophobicand ionic interactions, at pressures above 200 MPa. In general,the biological activity of enzymes is governed by the active site;therefore, even small changes in this area can trigger eitheractivation or a loss in enzyme activity. Hendricks et al. (1998)43
suggested that high-pressure treatment may affect the intra-molecular arrangement of proteins, leading to the unfolding ordenaturation of the enzyme. However, in the current study,there were also cultivars that showed an increase in PPOactivity after processing extracts at 400 MPa, e.g., in thecultivars Andross and Halford. Conversely, in the Late Rosscultivar, PPO activity actually began to decrease after mildtreatment at 100 MPa, indicating that the PPO extract of thiscultivar was more sensitive to high pressure than the others. Itis possible that the enzyme extracted from this cultivar had aslightly different secondary or tertiary structure, or that theenzyme was more easily released from its cellular matrix afterthe 100 MPa treatment than that in other cultivars.
Figure 6. Total phenols content of various peach cultivars following HPP treatment at 0 (control), 100, and 400 MPa. Values with the same letterare not significantly different across treatment variables at a significance level of p < 0.05.
Journal of Agricultural and Food Chemistry Article
DOI: 10.1021/acs.jafc.6b01879J. Agric. Food Chem. 2016, 64, 7606−7614
7612

Knorr (1993)29 suggested that PPO activity can be eitheractivated or inactivated, depending on the processingconditions used and the type of fruit or vegetable. PPOactivation induced by high-pressure processing at low-pressurelevels (100−400 MPa) has been reported in pear extracts,47
strawberries,48 and onions.28 At the other end of the spectrum,a reduction in PPO activity was observed in guacamole9 andguava49 after exposure to pressure at 600 MPa and higher. Theadvantage of using PPO extracts in the present study was todetermine the effect of HPP treatment on activity of theenzyme alone, after eliminating the effect of phenolic productinhibition. In the nine peach cultivars, three different trends(increasing, stable, and decreasing activity) were observed aftertreatment at 400 MPa. These differences may arise fromvariations in PPO type and physical structure among thevarious peach cultivars, and consequently, the same level ofHPP treatment may have a different effect on them. On thebasis of our results, processors might choose to use onlymidseason peach cultivars, which had desirably low rates ofbrowning after processing.3.5. Total Phenols Content of Different Peach
Cultivars Following HPP. Peaches of the various cultivarshad different amounts of initial total phenols, as illustrated inFigure 6. The cultivars Carson and Evert had the highest levelswith mean values of 9.54 and 10.99 mg/g sample, respectively,while the other cultivars had similarly low levels with meanvalues in the range of 6.00−8.00 mg/g sample. A previous studyof four freestone peach cultivars (Maycrest, Flavorcrest, ElegantLady, and O’Henry) and two nectarine cultivars (Mayglo andFlaming Red) reported total phenols contents in the rangefrom 6.65 to 8.43 mg GAE/g DW,50 which is in the same rangeas most of the cultivars in the current study. There were nosignificant changes (p < 0.05) in phenolic content followingHPP at either 100 or 400 MPa compared to control samples inany of the cultivars. Thus, high-pressure treatment is anexcellent process in terms of the ability to retain these nutrients.In similar studies, no significant difference in total phenolscontent was reported in high-pressure-treated strawberry andblackberry purees after processing at 400 and 500 MPa.51 Thesame observation was also reported in Granny Smith applepuree following HPP treatment at 400 MPa.2 Phenoliccompounds not only act as substrates for PPO but also haveantioxidative, anti-inflamatory, and anticarcinogenic properties.In conclusion, this research demonstrated the importance of
peach cultivar on the final quality of HPP peach products. Afterhigh-pressure treatment, a range in the degree of membranedamage was observed in the nine different peach cultivars, asdetermined by 1H NMR relaxometry and light microscopy. As aresult of the loss of membrane integrity, phenolic substrates inthe vacuole are allowed to interact with PPO from the plastids,creating brown pigments. In this study, the relationshipbetween degree of browning, the activity of PPO in partiallypurified peach extracts, cell integrity using NMR relaxometry,and microscopic techniques showed strong agreement. In acomparison of peach cultivars, discoloration of peach flesh isbetter explained by seasonality and cultivar, as well as the PPOactivity in extracts. The mid season peach cultivars, whichretained greater integrity following HPP and had less PPOactivity, showed less browning compared to the other seasongroups. These cultivars are thus more suitable for high-pressureprocessing.
■ AUTHOR INFORMATIONCorresponding Author*E-mail: [email protected] authors declare no competing financial interest.
■ ACKNOWLEDGMENTSWe thank the Royal Thai Government, Ministry of Science andTechnology and Prince of Songkla University, Surat ThaniCampus for support of this project.
■ REFERENCES(1) Ahmed, J.; Ramaswamy, H. S. High pressure processing of fruitsand vegetables. Stewart Postharvest Rev. 2006, 2, 1−10.(2) Landl, A.; Abadias, M.; Sarraga, C.; Vinas, I.; Picouet, P. Effect ofhigh pressure processing on the quality of acidified Granny Smithapple puree product. Innovative Food Sci. Emerging Technol. 2010, 11,557−564.(3) Kaushik, N.; Kaur, B. P.; Rao, P. S.; Mishra, H. Effect of highpressure processing on color, biochemical and microbiologicalcharacteristics of mango pulp (Mangifera indica cv. Amrapali).Innovative Food Sci. Emerging Technol. 2014, 22, 40−50.(4) Koutchma, T. Adapting High Hydrostatic Pressure (HPP) for FoodProcessing Operations; Academic Press: San Diego, CA, 2014.(5) Gonzalez, M. E.; Barrett, D. M.; McCarthy, M. J.; Vergeldt, F. J.;Gerkema, E.; Matser, A. M.; Van As, H. 1H-NMR Study of the Impactof High Pressure and Thermal Processing on Cell Membrane Integrityof Onions. J. Food Sci. 2010, 75, E417−E425.(6) Lopez-Malo, A.; Palou, E.; Barbosa-Canovas, G.; Welti-Chanes,J.; Swanson, B. Polyphenoloxidase activity and color changes duringstorage of high hydrostatic pressure treated avocado puree. Food Res.Int. 1998, 31, 549−556.(7) Palou, E.; Lopez-Malo, A.; Barbosa-Canovas, G.; Welti-Chanes,J.; Swanson, B. Polyphenoloxidase activity and color of blanched andhigh hydrostatic pressure treated banana puree. J. Food Sci. 1999, 64,42−45.(8) Krebbers, B.; Matser, A.; Koets, M.; Van den Berg, R. Quality andstorage-stability of high-pressure preserved green beans. J. Food Eng.2002, 54, 27−33.(9) Palou, E.; Hernandez-Salgado, C.; Lopez-Malo, A.; Barbosa-Canovas, G.; Swanson, B.; Welti-Chanes, J. High pressure-processedguacamole. Innovative Food Sci. Emerging Technol. 2000, 1, 69−75.(10) Asaka, M.; Hayashi, R. Activation of polyphenoloxidase in pearfruits by high pressure treatment. Agric. Biol. Chem. 1991, 55, 2439−2440.(11) Marigheto, N.; Vial, A.; Wright, K.; Hills, B. A combined NMRand microstructural study of the effect of high-pressure processing onstrawberries. Appl. Magn. Reson. 2004, 26, 521−531.(12) Van As, H. NMR in horticulture: In situ plant water balancestudies with NMR. Acta Hortic. 1991, 304, 103−112.(13) Admon, A.; Jacoby, B.; Goldschmidt, E. E. Assessment ofcytoplasmic contaminations in isolated vacuole preparations. PlantPhysiol. 1980, 65, 85−87.(14) De Vos, K. Cell counter plugin. http://rsb.info.nih.gov/ij/plugins/cell-counter.html (accessed May 2015), 2008.(15) Espín, J. C.; Morales, M.; Varon, R.; Tudela, J.; Garcia-Canovas,F. Monophenolase activity of polyphenol oxidase from Verdedoncellaapple. J. Agric. Food Chem. 1995, 43, 2807−2812.(16) Waterhouse, A. L. Determination of total phenolics. In CurrentProtocols in Food Analytical Chemistry; John Wiley & Son, Inc:Hoboken, NJ, 2002.(17) Snaar, J.; Van As, H. Probing water compartments andmembrane permeability in plant cells by 1H NMR relaxationmeasurements. Biophys. J. 1992, 63, 1654−1658.(18) Basak, S.; Ramaswamy, H. Effect of high pressure processing onthe texture of selected fruits and vegetables. J. Texture Stud. 1998, 29,587−601.
Journal of Agricultural and Food Chemistry Article
DOI: 10.1021/acs.jafc.6b01879J. Agric. Food Chem. 2016, 64, 7606−7614
7613

(19) Hills, B. P.; Remigereau, B. NMR studies of changes insubcellular water compartmentation in parenchyma apple tissue duringdrying and freezing. Int. J. Food Sci. Technol. 1997, 32, 51−61.(20) Zhang, L.; McCarthy, M. J. Black heart characterization anddetection in pomegranate using NMR relaxometry and MR imaging.Postharvest Biol. Technol. 2012, 67, 96−101.(21) Hills, B.; Costa, A.; Marigheto, N.; Wright, K. T 1− T 2 NMRcorrelation studies of high-pressure-processed starch and potato tissue.Appl. Magn. Reson. 2005, 28, 13−27.(22) Hernandez-Sanchez, N.; Hills, B.; Barreiro, P.; Marigheto, N. AnNMR study on internal browning in pears. Postharvest Biol. Technol.2007, 44, 260−270.(23) Hutzler, P.; Fischbach, R.; Heller, W.; Jungblut, T. P.; Reuber,S.; Schmitz, R.; Veit, M.; Weissenbock, G.; Schnitzler, J.-P. Tissuelocalization of phenolic compounds in plants by confocal laserscanning microscopy. J. Exp. Bot. 1998, 49, 953−965.(24) Vaughn, K. C.; Duke, S. O. Function of polyphenol oxidase inhigher plants. Physiol. Plant. 1984, 60, 106−112.(25) Martinez, M. V.; Whitaker, J. R. The biochemistry and controlof enzymatic browning. Trends Food Sci. Technol. 1995, 6, 195−200.(26) Techakanon, C. The Study of Enzymatic Browning Reactions inHigh Hydrostatic Pressure Processed Peach (Prunus persica). Ph.D.Dissertation, University of California, Davis, 2015.(27) Dornenburg, H.; Knorr, D. Cellular permeabilization of culturedplant tissues by high electric field pulses or ultra high pressure for therecovery of secondary metabolites. Food Biotechnol. 1993, 7, 35−48.(28) Butz, P.; Koller, W.; Tauscher, B.; Wolf, S. Ultra-high pressureprocessing of onions: chemical and sensory changes. LWT-Food Sci.and Technol. 1994, 27, 463−467.(29) Knorr, D. Effects of high-hydrostatic-pressure processes on foodsafety and quality. Food Technol. (Chicago, IL, U.S.) 1993, 47, 156−161.(30) Gonzalez, M. E.; Barrett, D. M. Thermal, High Pressure, andElectric Field Processing Effects on Plant Cell Membrane Integrity andRelevance to Fruit and Vegetable Quality. J. Food Sci. 2010, 75, R121−R130.(31) Gradziel, T.; McCaa, J. Processing peach cultivar development.In The Peach: Botany, Production and Uses; Layne, D. R., Bassi, D., Eds.;CAB Int.: Wallingford, CT, 2008.(32) Loomis, W. D.; Battaile, J. Plant phenolic compounds and theisolation of plant enzymes. Phytochemistry 1966, 5, 423−438.(33) Pierpoint, W. o-Quinones formed in plant extracts. Theirreactions with amino acids and peptides. Biochem. J. 1969, 112, 609−616.(34) Mason, H.; Peterson, E. Melanoproteins I. Reactions betweenenzyme-generated quinones and amino acids. Biochim. Biophys. Acta,Gen. Subj. 1965, 111, 134−146.(35) Mathew, A.; Parpia, H. Food browning as a polyphenol reaction.Adv. Food Res. 1971, 19, 75−145.(36) Sanchez-Ferrer, A.; Perez-Gilabert, M.; Nunez, E.; Bru, R.;García-Carmona, F. Triton X-114 phase partitioning in plant proteinpurification. J.Chromatography A 1994, 668, 75−83.(37) Felton, G.; Donato, K.; Broadway, R.; Duffey, S. Impact ofoxidized plant phenolics on the nutritional quality of dietar protein toa noctuid herbivore, Spodoptera exigua. J. Insect Physiol. 1992, 38,277−285.(38) Huber, M.; Hintermann, G.; Lerch, K. Primary structure oftyrosinase from Streptomyces glaucescens. Biochemistry 1985, 24,6038−6044.(39) Lerch, K. Neurospora tyrosinase: structural, spectroscopic andcatalytic properties. Mol. Cell. Biochem. 1983, 52, 125−138.(40) Golan-Goldhirsh, A.; Whitaker, J. R. k CAT Inactivation ofmushroom polyphenol oxidase. J. Mol. Catal. 1985, 32, 141−147.(41) Nelson, J.; Dawson, C. Tyrosinase. Adv. Enzymol. Relat. AreasMol. Biol., Volume 4 2006, 99−152.(42) Dietler, C.; Lerch, K. Reaction inactivation of tyrosinase. InOxidases and related redox systems; Pergamon Press: Oxford, U.K.,1982; pp 305−317.
(43) Hendricks, M.; Ludikhuyze, L.; Van den Broeck, I.; Weemaes,C. Effects of high pressure on enzymes related to food quality. TrendsFood Sci. Technol. 1998, 9, 197−203.(44) Gomes, M.; Ledward, D. Effect of high-pressure treatment onthe activity of some polyphenoloxidases. Food Chem. 1996, 56, 1−5.(45) Yi, J.; Jiang, B.; Zhang, Z.; Liao, X.; Zhang, Y.; Hu, X. Effect ofultrahigh hydrostatic pressure on the activity and structure ofmushroom (Agaricus bisporus) polyphenoloxidase. J. Agric. FoodChem. 2012, 60, 593−599.(46) Tauscher, B. Pasteurization of food by hydrostatic high pressure:chemical aspects. Z. Lebensm.-Unters. Forsch. 1995, 200, 3−13.(47) Asaka, M.; Aoyama, Y.; Nakanishi, R.; Hayashi, R. Purification ofa latent form of polyphenoloxidase from La France pear fruit and itspressure-activation. Biosci., Biotechnol., Biochem. 1994, 58, 1486−1489.(48) Cano, M. P.; Hernandez, A.; Ancos, B. d. High pressure andtemperature effects on enzyme inactivation in strawberry and orangeproducts. J. Food Sci. 1997, 62, 85−88.(49) Yen, G. C.; Lin, H. T. Comparison of high pressure treatmentand thermal pasteurization effects on the quality and shelf life of guavapuree. Int. J. Food Sci. Technol. 1996, 31, 205−213.(50) Cheng, G. W.; Crisosto, C. H. Browning potential, phenoliccomposition, and polyphenoloxidase activity of buffer extracts of peachand nectarine skin tissue. J. Am. Soc. Hortic. Sci. 1995, 120, 835−838.(51) Patras, A.; Brunton, N. P.; Da Pieve, S.; Butler, F. Impact of highpressure processing on total antioxidant activity, phenolic, ascorbicacid, anthocyanin content and colour of strawberry and blackberrypurees. Innovative Food Sci. Emerging Technol. 2009, 10, 308−313.
Journal of Agricultural and Food Chemistry Article
DOI: 10.1021/acs.jafc.6b01879J. Agric. Food Chem. 2016, 64, 7606−7614
7614


















