
Ecological response to collapse of the biological pump ... · lub, Yucatan, Mexico. ... 1991). In...
16
Biogeosciences, 14, 885–900, 2017 www.biogeosciences.net/14/885/2017/ doi:10.5194/bg-14-885-2017 © Author(s) 2017. CC Attribution 3.0 License. Ecological response to collapse of the biological pump following the mass extinction at the Cretaceous–Paleogene boundary Johan Vellekoop 1,2,* , Lineke Woelders 2,* , Sanem Açikalin 3 , Jan Smit 4 , Bas van de Schootbrugge 1 , Ismail Ö. Yilmaz 5,6 , Henk Brinkhuis 1,7 , and Robert P. Speijer 2 1 Marine Palynology, Laboratory of Palaeobotany and Palynology, Faculty of Geosciences, Utrecht University, Utrecht, 3584 CD, the Netherlands 2 Division of Geology, Department of Earth and Environmental Sciences, KU Leuven, Leuven-Heverlee, 3001, Belgium 3 School of Civil Engineering and Geosciences, Newcastle University, NE1 7RU, Newcastle upon Tyne, UK 4 Department of Sedimentology and Marine Geology, Faculty of Earth and Life Science, Vrije Universiteit Amsterdam, Amsterdam, 1018HV, the Netherlands 5 Department of Geological Engineering, Middle East Technical University, Ankara, Turkey 6 Department of Geological Sciences, University of Texas at Austin, Austin, TX 78712, USA 7 Royal Netherlands Institute for Sea Research (NIOZ), Landsdiep 4, ’t Horntje, Texel, 1797 SZ, the Netherlands * These authors contributed equally to this work. Correspondence to: Johan Vellekoop ([email protected]) Received: 30 June 2016 – Discussion started: 3 August 2016 Revised: 16 December 2016 – Accepted: 26 January 2017 – Published: 27 February 2017 Abstract. It is commonly accepted that the mass extinction associated with the Cretaceous–Paleogene (K–Pg) bound- ary ( ∼ 66 Ma) is related to the environmental effects of a large extraterrestrial impact. The biological and oceano- graphic consequences of the mass extinction are, however, still poorly understood. According to the “Living Ocean” model, the biological crisis at the K–Pg boundary resulted in a long-term reduction of export productivity in the early Paleocene. Here, we combine organic-walled dinoflagellate cyst (dinocyst) and benthic foraminiferal analyses to pro- vide new insights into changes in the coupling of pelagic and benthic ecosystems. To this end, we perform dinocyst and benthic foraminiferal analyses on the recently discov- ered Tethyan K–Pg boundary section at Okçular, Turkey, and compare the results with other K–Pg boundary sites in the Tethys. The post-impact dominance of epibenthic morpho- types and an increase of inferred heterotrophic dinocysts in the early Paleocene at Okçular are consistent with published records from other western Tethyan sites. Together, these records indicate that during the early Paleocene more nutri- ents remained available for the Tethyan planktonic commu- nity, whereas benthic communities were deprived of food. Hence, in the post-impact phase the reduction of export pro- ductivity likely resulted in enhanced recycling of nutrients in the upper part of the water column, all along the western Tethyan margins. 1 Introduction It is now commonly accepted that the Cretaceous–Paleogene (K–Pg) boundary (∼ 66 Ma) mass extinction was associated with the impact of a large extraterrestrial body at Chicxu- lub, Yucatan, Mexico. The short- and long-term environmen- tal implications of this impact resulted in the extinction of a large number of biological clades (Sepkoski, 1996). Pale- ontological records indicate that approximately 50 % of ma- rine genera became extinct across the K–Pg boundary. This episode thus represents one of the largest mass-extinction events in Earth history (Sepkoski, 1996; D’Hondt, 2005). Apart from short-term global environmental consequences, such as an initial “impact winter” phase (Alvarez et al., 1980; Vellekoop et al., 2014, 2016), the event also had major long- term biological consequences. The large-scale extinctions Published by Copernicus Publications on behalf of the European Geosciences Union.
Transcript of Ecological response to collapse of the biological pump ... · lub, Yucatan, Mexico. ... 1991). In...

Biogeosciences 14 885ndash900 2017wwwbiogeosciencesnet148852017doi105194bg-14-885-2017copy Author(s) 2017 CC Attribution 30 License
Ecological response to collapse of the biological pump following themass extinction at the CretaceousndashPaleogene boundaryJohan Vellekoop12 Lineke Woelders2 Sanem Accedilikalin3 Jan Smit4 Bas van de Schootbrugge1 Ismail Ouml Yilmaz56Henk Brinkhuis17 and Robert P Speijer2
1Marine Palynology Laboratory of Palaeobotany and Palynology Faculty of Geosciences Utrecht UniversityUtrecht 3584 CD the Netherlands2Division of Geology Department of Earth and Environmental Sciences KU Leuven Leuven-Heverlee3001 Belgium3School of Civil Engineering and Geosciences Newcastle University NE1 7RU Newcastle upon Tyne UK4Department of Sedimentology and Marine Geology Faculty of Earth and Life Science Vrije Universiteit AmsterdamAmsterdam 1018HV the Netherlands5Department of Geological Engineering Middle East Technical University Ankara Turkey6Department of Geological Sciences University of Texas at Austin Austin TX 78712 USA7Royal Netherlands Institute for Sea Research (NIOZ) Landsdiep 4 rsquot Horntje Texel 1797 SZ the NetherlandsThese authors contributed equally to this work
Correspondence to Johan Vellekoop (johanvellekoopkuleuvenbe)
Received 30 June 2016 ndash Discussion started 3 August 2016Revised 16 December 2016 ndash Accepted 26 January 2017 ndash Published 27 February 2017
Abstract It is commonly accepted that the mass extinctionassociated with the CretaceousndashPaleogene (KndashPg) bound-ary (sim 66 Ma) is related to the environmental effects ofa large extraterrestrial impact The biological and oceano-graphic consequences of the mass extinction are howeverstill poorly understood According to the ldquoLiving Oceanrdquomodel the biological crisis at the KndashPg boundary resultedin a long-term reduction of export productivity in the earlyPaleocene Here we combine organic-walled dinoflagellatecyst (dinocyst) and benthic foraminiferal analyses to pro-vide new insights into changes in the coupling of pelagicand benthic ecosystems To this end we perform dinocystand benthic foraminiferal analyses on the recently discov-ered Tethyan KndashPg boundary section at Okccedilular Turkey andcompare the results with other KndashPg boundary sites in theTethys The post-impact dominance of epibenthic morpho-types and an increase of inferred heterotrophic dinocysts inthe early Paleocene at Okccedilular are consistent with publishedrecords from other western Tethyan sites Together theserecords indicate that during the early Paleocene more nutri-ents remained available for the Tethyan planktonic commu-nity whereas benthic communities were deprived of food
Hence in the post-impact phase the reduction of export pro-ductivity likely resulted in enhanced recycling of nutrientsin the upper part of the water column all along the westernTethyan margins
1 Introduction
It is now commonly accepted that the CretaceousndashPaleogene(KndashPg) boundary (sim 66 Ma) mass extinction was associatedwith the impact of a large extraterrestrial body at Chicxu-lub Yucatan Mexico The short- and long-term environmen-tal implications of this impact resulted in the extinction ofa large number of biological clades (Sepkoski 1996) Pale-ontological records indicate that approximately 50 of ma-rine genera became extinct across the KndashPg boundary Thisepisode thus represents one of the largest mass-extinctionevents in Earth history (Sepkoski 1996 DrsquoHondt 2005)Apart from short-term global environmental consequencessuch as an initial ldquoimpact winterrdquo phase (Alvarez et al 1980Vellekoop et al 2014 2016) the event also had major long-term biological consequences The large-scale extinctions
Published by Copernicus Publications on behalf of the European Geosciences Union
886 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
amongst primary producers caused a major restructuring ofglobal food webs and carbon cycling (DrsquoHondt 2005 Cox-all et al 2006) Moreover a collapse in the oceanic stablecarbon isotope gradient between surface and bottom waterspersisted for up to a few million years (Hsuuml and McKenzie1985 Zachos et al 1989 DrsquoHondt et al 1998) Initiallythe ldquoStrangelove Oceanrdquo hypothesis was invoked to explainthis collapse suggesting that primary productivity sharplydecreased or ceased immediately after the KndashPg boundary asa consequence of the extinction of primary producers (egHsuuml and McKenzie 1985)
However modeling the carbon isotope gradient responseto the extinctions suggests that carbon burial had to continuenearly unabated to prevent the carbon isotopic signature ofthe global ocean from drifting towards that of the weath-ering input (Kump 1991) In addition both the persistenceof certain primary producers such as dinoflagellates (egBrinkhuis and Zachariasse 1988) as well as the survivalof benthic foraminifera (eg Culver 2003) argue againstprolonged cessation of primary productivity in the globaloceans Integration of neritic and deep-sea planktic and ben-thic foraminiferal carbon isotopic records suggests that thebreakdown of this gradient reflects a global collapse of exportproductivity ie the fraction of organic carbon that sinksfrom the photic zone to the deep ocean via the biologicalpump (Kump 1991 DrsquoHondt et al 1998 DrsquoHondt 2005Hilting et al 2008 Hain et al 2014 Esmeray-Senlet etal 2015) rather than the shutdown of primary productivityThis conceptual model is generally referred to as the ldquoLiv-ing Oceanrdquo model (DrsquoHondt and Zachos 1998 DrsquoHondt etal 1998 DrsquoHondt 2005) According to the Living Oceanmodel total biological productivity recovered rapidly afterthe extinction event but the total global export productivityfrom the photic zone was reduced for hundreds of thousandsof years (DrsquoHondt et al 1998 DrsquoHondt 2005 Birch et al2016) Although there has been considerable debate on thispossible collapse of the biological pump following the KndashPgboundary mass extinction (eg Alegret and Thomas 2009Hull et al 2011) most recent studies are in agreement thatglobal post-KndashPg export productivity was reduced to someextent (eg Esmerey-Senlet et al 2015 Birch et al 2016)Current discussions are mostly related to the severity of thereduction of export productivity or the geographical differ-ences (ie the ldquoheterogeneityrdquo) of the oceans In particularthe records from the Pacific open ocean could potentiallybe explained as showing no reduction of the biological pump(eg Alegret et al 2012 Hull et al 2011) while sites fromthe Tethys Ocean Atlantic Ocean Southern Ocean and In-dian Ocean show convincing evidence for a collapse of a bi-ological pump at the KndashPg boundary (eg Thomas 1990Olsson et al 1996 Hull et al 2011 Alegret et al 2012)
The inferred reduction in the organic flux to deep watersrecorded at most sites might be a consequence of the ecosys-tem reorganization that resulted from the mass extinctionVariations in species assemblages of pelagic communities
can lead to large changes in the rates of particulate export(eg Buesseler 1998 Stemmann and Boss 2012) A generalreduction of the number of pelagic grazers (such as macro-zooplankton) or a shift in dominance from grazers that cre-ate fecal pellets (eg fish) to grazers that do not produce fe-cal pellets (eg dinoflagellates) could have greatly reducedthe packaging of biomass into large particles sinking to thedeep ocean (DrsquoHondt et al 1998 DrsquoHondt 2005) This isexemplified by the record at Gubbio Italy which indicatesan abrupt decline of fish stocks at the KndashPg boundary re-maining depressed for millions of years in the western Tethys(Sibert et al 2014)
Additionally the replacement of large tests of Creta-ceous planktic foraminifera and calcareous nanoplankton bysmaller early Paleocene forms (eg Bernaola and Monechi2007 Molina 2015) might have reduced biomineral ballast-ing (Armstrong et al 2002) during the early Paleocene re-sulting in a further reduction of the carbon flux to the oceanfloor (Coxall et al 2006)
Although numerous studies have been performed to seekphysical evidence for the KndashPg boundary impact the result-ing millennial-scale biotic responses are still poorly under-stood Some of the most intensely studied microfossil groupsused in paleoenvironmental reconstructions such as plankticforaminifera and calcareous nanoplankton experienced ma-jor extinctions and subsequent radiations (eg Smit 1982Huber et al 2002 Coxall et al 2006 Molina 2015 Schuethet al 2015) hampering biota-based environmental recon-structions across the KndashPg boundary interval In contrastbenthic foraminifera and organic-cyst-producing dinoflagel-lates are much better suited as they show no significant ex-tinction above background levels at the end of the Cretaceous(Brinkhuis and Zachariasse 1988 Culver 2003)
A few high-resolution KndashPg boundary benthicforaminiferal and organic-walled dinoflagellate cysts(dinocyst) records have been published particularly fromthe southern and western shallow margins of the TethysOcean for example from Tunisia Israel Egypt Spain andMorocco (Brinkhuis and Zachariasse 1988 Eshet et al1992 Coccioni and Galeotti 1994 Speijer and van derZwaan 1996 Brinkhuis et al 1998 Peryt et al 2002Alegret et al 2003 Slimani et al 2010 Vellekoop et al2015) (Fig 1) These records thus potentially provide acomprehensive Tethys-wide portrayal of the changes inpelagic-benthic-coupling across the KndashPg boundary Thebenthic foraminiferal and dinocyst records from the south-ern margin of the Tethys do reveal indications for majorshort-term oceanographic changes in eg temperatureredox and trophic conditions across the KndashPg boundary(eg Brinkhuis and Zachariasse 1988 Speijer and van derZwaan 1996 Brinkhuis et al 1998) Especially quantitativebenthic foraminiferal records show a strong response to theimpact generally portraying an abrupt benthic communityimpoverishment across the boundary At many of theseKndashPg boundary sites after an initial short-lived prolifer-
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
J Vellekoop et al Ecological response to KndashPg boundary mass extinction 887
Figure 1 Reconstruction of the late Cretaceousndashearly Paleogene paleogeography of the North Atlantic and Mediterranean regions afterScotese (2004) and Scotese and Dreher (2012) The four sites discussed are indicated Okccedilular section Turkey (Accedilikalin et al 2015 thisstudy) El Kef Tunisia (Brinkhuis and Zachariasse 1988 Speijer and van der Zwaan 1996 Brinkhuis et al 1998) Ouled Haddou Morocco(Slimani et al 2010) and ODP Hole 1049C (Blake Nose Alegret and Thomas 2004) The red dot marks the Chicxulub impact crater
ation of endobenthic forms (eg Coccioni and Galeotti1994 Alegret et al 2015) epibenthic forms dominatethe post-impact ldquodisasterrdquo phase (Culver 2003) Since ingeneral endobenthic forms are considered indicative of ahigh flux of organic matter to the sea floor andor relativelylow oxygen conditions and epibenthic forms are generallymore common in oligotrophic open marine environments(eg Corliss 1985 Peryt et al 2002 Jorissen et al 2007Woelders and Speijer 2015) the post-impact abundance ofepibenthic forms is often explained as food starvation atthe sea floor (Culver 2003) Following this disaster phasemost benthic foraminiferal records show a relatively longrecovery phase with endobenthic forms gradually returningas diversity starts to increase again (Alegret et al 2003Culver 2003) and endobenthic microhabitats diversify(Speijer and van der Zwaan 1996)
Although combining quantitative dinocyst and benthicforaminifera analyses could provide crucial insight intochanges in pelagic-benthic-coupling (eg Guasti et al2005) perhaps surprisingly no such attempts have beenmade for the KndashPg boundary interval so far Earlier stud-ies discussed either benthic foraminiferal or dinoflagellateresponse to the KndashPg boundary each without making an at-tempt to combine the results of these studies into a compre-hensive integrated ocean-wide characterization of ecologi-cal patterns observed across the boundary In addition al-though the southern margins of the western Tethys provideda fair number of high-resolution records spanning the KndashPgboundary no such high-resolution records yet exist from thenortheastern margins of the western Tethys To be able toprovide a comprehensive (western) Tethys-wide portrayal ofthe surface and bottom water ecological changes across theKndashPg boundary and the coupling between pelagic and ben-thic systems additional dinocyst and benthic foraminiferal
records need to be generated from the northeastern margin ofthe western Tethys
The MudurnundashGoumlynuumlk basin in the central Sakarya re-gion northwestern Turkey provides new opportunities forhigh-resolution KndashPg boundary benthic foraminiferal anddinocyst records from the northern margin of the Tethys Re-cently well-preserved outcrops of ancient continental mar-gin deposits spanning the KndashPg boundary have been discov-ered in this basin (Accedilikalin et al 2015) These outcrops in-clude amongst others the Okccedilular section Here an inte-gration of dinocyst and benthic foraminifera records of thisbiostratigraphically well-constrained KndashPg boundary transi-tion is used to provide new insights into changes in and therelationship between planktic and benthic communities
Linking these records to both the previously gener-ated bulk-carbonate carbon isotope record of this section(Accedilikalin et al 2015) and to other benthic foraminiferal andpalynological records in the Tethys enables a supra-regionalportrayal of the KndashPg boundary pelagic crisis and the bio-logical changes caused by the collapse of export productivityThis integrated approach enables an evaluation of the paleoe-cological and paleoceanographic consequences of the earlyDanian Living Ocean condition
2 Geological setting and age assessment
The Okccedilular section is located in the MudurnundashGoumlynuumlkbasin (northwestern Turkey Fig 1) In this basin the KndashPg boundary interval is represented by the Tarakli Forma-tion (Saner 1980 Altiner et al 1991 Accedilikalin et al 2015)In the eastern part of the basin the upper Maastrichtianis characterized by an intercalation of mudstones and tur-bidites whereas in the western part of the basin the tur-bidites are absent The KndashPg boundary is marked by a red-dish ejecta layer at the base of a 15ndash20 cm thick boundary
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
888 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
clay layer Throughout the basin the lower 30ndash50 m of theDanian is characterized by a rhythmic alternation of fine-grained limestones and carbonate-rich mudstones (Accedilikalinet al 2015) During the latest Cretaceous to early Paleocenethis basin was characterized by mixed siliciclasticndashcarbonatesedimentation in an outer neritic to upper bathyal environ-ment (Accedilikalin et al 2015)
The Okccedilular section has been analyzed for siderophiletrace elements including Ir and other platinum groupelements (PGEs) bulk stable carbon isotopes plankticforaminifera calcareous nannofossils and dinocysts Basedon these results a detailed stratigraphic framework was out-lined (Accedilikalin et al 2015) allowing a confident age assess-ment of the boundary interval The age model shows thatthe section contains a chronostratigraphically complete KndashPg boundary interval The studied interval ranges from thetop part of the Maastrichtian A mayaroensis Zone up to thebasal part of the Danian planktic foraminiferal Zone P1b andcovers globally occurring first occurrences (FO) of dinocystmarker taxa such as Senoniasphaera inornata Damassa-dinium californicum and Carpatela cornuta
3 Materials and methods
31 Sampling
High-resolution (centimeter-scale) sample sets were ac-quired during two field campaigns in 2010 and in 2011 Formore detail on these sampling campaigns see Accedilikalin etal (2015) The samples were split for micropaleontologicaland palynological analyses
32 Foraminiferal analysis
Twenty-seven samples were processed at KU Leuven forforaminiferal studies following standard micropaleontolog-ical procedures Of these 20 samples were used for quan-titative benthic foraminiferal analyses (Fig 2) Rock sam-ples were dried in an oven at 60 C for at least 24 h De-pending on sample size 4 to 60 g of dry rock was soakedin a soda (Na2CO3) solution (50 g Lminus1) After disintegrationsamples were washed over 2 mm and 63 microm sieves If neces-sary the tenside Rewoquat was used to clean the residues andthe procedure was repeated Clean residues were dry-sievedinto three fractions 63ndash125 125ndash630 and gt 630 microm In orderto enable a direct comparison with the benthic foraminiferalrecord of the KndashPg boundary GSSP (Global Boundary Stra-totype Section and Point) of El Kef (Tunisia) we used thesame size fraction (gt 125 microm) for Okccedilular as for El Kef(Speijer and van der Zwaan 1996) Note that El Kef holdsthe GSSP of the base of the Danian (Molina et al 2006)and is thus of particular importance with respect to corre-lation and comparison of the KndashPg boundary Representativealiquots of the gt 125 microm fraction were obtained containing atleast 300 benthic foraminiferal specimens Picked specimens
from this size fraction were permanently stored in Plummerslides Benthic foraminifera were identified using the tax-onomy of Cushman (1946 1951) Kellough (1965) Aubertand Berggren (1976) Berggren and Aubert (1975) and Spei-jer (1994)
Benthic foraminifera are commonly used as indicators forbottom water oxygenation and trophic conditions (eg Joris-sen et al 2007) Here the benthic foraminiferal accumula-tion rate (BFAR number of foraminifera per cm2 per kyrsee Text S2 in the Supplement for details on the estimationof BFAR) was calculated as a semi-quantitative proxy for pa-leoproductivity (Jorissen et al 2007 and references therein)In addition the percentage of endobenthic morphotypes wascalculated using the assumed microhabitat preferences in-ferred from benthic foraminiferal morphotype analysis (egCorliss 1985 Corliss and Chen 1988 Alegret et al 2003Woelders and Speijer 2015) In general endobenthic formsare considered indicative of a high flux of organic matter tothe sea floor andor relatively low oxygen conditions whileabundance of epibenthic forms is considered characteristicfor more oligotrophic open marine environments (Jorissen etal 1995 Peryt 2004 Jorissen et al 2007) Furthermore thebi- and triserial endobenthic forms are particularly indica-tive of high food supply and low oxygenation (eg Bern-hard 1986 Corliss and Chen 1988 Jorissen et al 2007)Therefore in this study the percentage of endobenthic formswas calculated for each sample as well as the percentage bi-triserial benthic taxa to unravel food supply and oxygena-tion patterns
It should however be noted that assuming such an ana-logue with modern fauna has limitations and shortcomingsFor instance the percentage of endobenthics based on mor-photypes and using bi-triserials as an indicator for hypoxiaand high food supply is not universally applicable (Buzas etal 1993 Jorissen et al 2007 and references therein)
33 Palynological analysis
In this study the palynological data from Accedilikalin etal (2015) were analyzed and interpreted In addition twoadditional samples were analyzed (OK 15 and OK 6) toincrease the resolution of the dataset and two samples ofthe Accedilikalin et al (2015) dataset were re-counted (OK 25and OK 250 see Dataset 2 in the Supplement) Quantitativeslides of the 15ndash250 microm fraction were used All slides arestored in the collection of the Laboratory of Palaeobotanyand Palynology Utrecht University the Netherlands
To identify major changes in the dinocyst record morpho-logically closely related taxa were grouped into complexesusing a similar approach to Schioslashler et al (1997) Sluijsand Brinkhuis (2009) and Machalski et al (2016) In ourstudy the following morphological complexes were estab-lished (1) the Spiniferites complex combining all species ofSpiniferites and the morphologically similar genus Achomop-shaera (2) Manumiella spp grouping all species of Man-
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
J Vellekoop et al Ecological response to KndashPg boundary mass extinction 889
Figure 2 The benthic foraminiferal record (gt 125 microm) of the Okccedilular section The biostratigraphy is from Accedilikalin et al (2015) The clusteranalysis on the benthic foraminiferal data using paired group (UPGMA) correlation distance allows the identification of four main clustersAndashD of benthic foraminiferal taxa
umiella (3) hexaperidinioids lumping all other peridinioidcysts with a hexaform archeopyle and (4) other dinocystswhich include all other dinocyst taxa and unidentifiabledinocysts (Fig 3)
Of these different dinocyst groups previous studies haveshown that in the Tethys the hexaperidinioids in particularshow strong variations across the KndashPg boundary (Brinkhuiset al 1998 Vellekoop et al 2015) Based on statistical cor-relations between palynological records and other paleoprox-ies it has been suggested that this inferred heterotrophicgroup flourished best under high-nutrient conditions in thephotic zone (Eshet et al 1994 Brinkhuis et al 1998 Sluijsand Brinkhuis 2009) Therefore in this study abundances ofhexaperidinioids are considered indicative of nutrient avail-ability in the photic zone (see Fig S1 and Text S1 in theSupplement for a more detailed discussion on this matter)
34 Statistical analysis
To assess changes in diversity of benthic foraminifera anddinocysts across the studied interval the Shannon diversityindex (H ) the species richness per sample (S) the number ofspecimens observed per sample (N ) and the BergerndashParkerindex were calculated for both biological groups followingHayek and Buzas (2013) (Fig 4) In addition in order torecognize the main faunal associations within the benthicforaminiferal data a cluster analysis was performed usingpaired group (UPGMA) correlation distance
Q-mode non-metric multidimensional scaling (NM-MDS)was performed on the benthic foraminiferal sample compo-sitions to assess patterns in assemblage response to KndashPgboundary perturbations Since taxa may not have a linear re-sponse to environmental changes across the KndashPg boundaryQ-mode NM-MDS is preferred over PCA and CCA (Ram-ette 2007)
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
890 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
Figure 3 The organic-walled dinoflagellate cyst record of theOkccedilular section The biostratigraphy and bulk carbonate stable car-bon isotope records are from Accedilikalin et al (2015) The four maindinocyst complexes are indicated
4 Results
41 Benthic foraminifera
The benthic foraminiferal record of the Okccedilular section ischaracterized by a distinct turnover across the KndashPg bound-ary (Figs 2 4 5 6) Of the common taxa 8 out of 30(sim 27 ) disappear across the KndashPg boundary After the KndashPg boundary crisis three new taxa appear The estimatedBFAR shows a major decrease across the boundary (Fig 5)For the foraminiferal counts and illustrations of commonforms see Dataset S1 and Figs S2 S3
The cluster analysis allows the identification of fourmain clusters of benthic foraminiferal taxa clusters A toD (Fig 2) Cluster C is relatively large and can be subdi-vided into five subclusters Based on the succession of ben-thic faunal assemblages characterized by strong changes inthe Shannon diversity index (H ) four intervals can be rec-ognized in the benthic foraminiferal record of the Okccedilularsection (Figs 4 6)
The first interval comprises the uppermost Maastrichtianand is characterized by relatively high diversity dominatedby the taxa in cluster D In this assemblage the bi-triserialbenthic taxa are relatively abundant (22ndash28 ) with char-acteristic taxa such as Bulimina arkadelphiana Eouvigerinasubsculptura and Praebulimina reussi About 50ndash60 of theassemblage consists of inferred endobenthic taxa The KndashPg boundary marks an abrupt benthic community impover-
ishment and the decimation of the taxa in cluster D Abovethe boundary the bi-triserial benthic taxa virtually disappearfrom the record (mostly below lt 05 )
The second interval characterized by low diversity com-prises the lowermost Danian approximately correspondingto planktic foraminiferal Zone P0 In this interval epibenthicforms are most abundant (70ndash90 ) It is dominated by thetaxa of cluster A encompassing successive peak occurrencesof the taxa Anomalinoides praeacutus Trochammina sppand Cibicidoides pseudoacutus
The third interval covers the part of the succession approx-imately correlative to planktic foraminiferal Zone Pα Thisinterval is characterized by a recovery of diversity althoughthe diversity is still lower than that of the top Maastrichtianassemblage The abundance of taxa in cluster A slowly de-creases and taxa in cluster B mainly represented by Osan-gularia plummerae and Cibicidoides sp become abundantEndobenthic forms recover and make up 30ndash40 of the totalassemblage
The fourth interval starts in the interval correlative to ZoneP1a Here the benthic community has recovered as the di-versity has stabilized and is almost similar to pre-impact val-ues This interval is characterized by taxa in cluster C a typ-ical Paleocene Midway-type fauna (Berggren and Aubert1975) with representatives such as Anomalinoides praea-cutus Coryphostoma midwayensis Cibicidoides alleni andOsangularia plummerae
42 Palynology
Palynological samples from the Okccedilular site yield abundantpalynomorphs dominated by dinocysts and with minor con-tributions of acritarchs prasinophytes organic foraminiferallinings and terrestrial palynomorphs (Dataset S2) Thedinocyst associations of the MudurnundashGoumlynuumlk basin arequite diverse including components characteristic of boththe Tethyan and Boreal realms (Accedilikalin et al 2015) As ex-pected the dinocyst record does not show major changes indiversity across the KndashPg boundary (Fig 4) There is a steadydecrease in dinocyst diversity from planktic foraminiferalZone P1a upwards probably a long-term change not re-lated to the impact The estimated dinocyst accumulationrate (DAR number of preserved cysts per cm2 per kyr seeText S2 in the Supplement for details on the estimation of theDAR) shows no major changes across the KndashPg boundary
Throughout the palynological record the Spiniferites com-plex is consistently the most dominant morphogroup in gen-eral comprising 40ndash50 of the total assemblage (Fig 3)Similar to other KndashPg boundary sites worldwide (Habib andSaeedi 2007) Manumiella spp show an episode of higherrelative abundance near the top of the Maastrichtian some30ndash40 cm below the KndashPg boundary Hexaperidinioids gen-erally make up a relatively small component of the Maas-trichtian assemblage (3ndash17 ) but show a strong increaseacross the KndashPg boundary In the boundary clay layer cor-
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
J Vellekoop et al Ecological response to KndashPg boundary mass extinction 891
Figure 4 Diversity indices on the benthic foraminiferal and dinocyst records of the Okccedilular section with the Shannon diversity index (H ) thespecies richness per sample (S) the number of specimen observed per sample (N ) and the BergerndashParker index As the bulk stable carbonisotope record and the Shannon diversity index of the benthic foraminiferal record show similar trends the combination of these records canbe used to subdivide the KndashPg boundary transition into four intervals I the Maastrichtian ie ldquopre-impactrdquo interval II the post-impactinterval representing the ldquodisasterrdquo phase III a recovery interval and IV the early Paleocene post-recovery interval The division criteriaare explained in the text
relative to planktic foraminiferal Zone P0 this group in-creases to 35 of the assemblage In the peak intervals thehexaperidinioids are mostly represented by representativesof the genera Senegalinium and Cerodinium After an ini-tial drop in relative abundance in the upper half of Zone P0this group reaches a second maximum (sim 35 ) at the baseof Zone Pα and remains relatively abundant (15ndash30 ) up toZone P1a After this the hexaperidinioids gradually decreasetowards the top of the studied interval The ldquoother dinocystsrdquogroup ndash with representatives such as Areoligera spp Im-pagidinium spp Hystrichosphaeridium tubiferum Opercu-lodinium centrocarpum and Palynodinium grallator ndash gen-erally makes up 25ndash50 of the assemblage
5 Discussion
51 Tethyan benthic foraminiferal turnover sequence
The major turnover in the benthic community at Okccedilularis largely comparable with the previously published ben-thic foraminiferal record (size fraction gt 125 microm) of the KndashPg boundary GSSP of El Kef (Figs 7 8 Speijer and Vander Zwaan 1996) In addition other benthic foraminiferalrecords from the margins of the Tethys also show a strong
similarity to the Okccedilular and El Kef records including stud-ies that use the gt 63 microm size fraction (Alegret et al 2003Culver 2003 and references therein)
Although it is sometimes suggested that different sizefractions cannot be directly compared because the differ-ent size fractions sometimes harbor different ecological in-formation (Giusberti et al 2016) intercomparison studieshave shown that when comparing the gt 125 and gt 63 microm sizefractions in benthic foraminiferal assemblages the overallpatterns are generally robust (Ernst et al 2006) The factthat gt 63 and gt 125 microm KndashPg boundary benthic foraminiferalrecords from different Tethys-margin sites (eg Speijer andVan der Zwaan 1996 Peryt et al 2002 Culver et al 2003)show striking similarities even though both the size fractionsused (gt 63 vs gt 125 microm) and specific taxa making up theforaminiferal assemblages differ among sites confirms theobservations by Ernst et al (2006) Furthermore even whenit would be assumed that different size fractions are diffi-cult to compare directly the striking similarities between theTethyan gt 125 and gt 63 microm size fraction KndashPg boundary ben-thic foraminiferal records provide compelling evidence for astrong disruption of the entire benthic community
Tethyan middle neritic to middle bathyal KndashPg boundarysuccessions generally show a sequence of (1) a typical high
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
892 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
Figure 5 Estimated benthic foraminiferal accumulation rate (BFAR number of foraminifera of the gt 63 microm size fraction per cm2 sea floorper kyr) and estimated dinocyst accumulation rate (DAR number of preserved cysts per cm2 sea floor per kyr) of the Okccedilular sectionUncertainty in estimated accumulation rates resulting from uncertainties in sediment densities and sedimentation rates are indicated in greyproviding a range of estimated accumulation rates For more detailed info on the calculation of the BFAR and DAR see Text S2 of theSupplement
diversity assemblage in the Maastrichtian (2) a low diver-sity ldquodisasterrdquo assemblage directly after the KndashPg boundaryimpact characterized by a short-lived bloom of endobenthicforms observed in several high-resolution records (eg Coc-cioni and Galeotti 1994 Speijer and van der Zwaan 1996)followed by a dominance of epibenthic forms (3) a ldquore-coveryrdquo assemblage characterized by an increasing diversityand returning endobenthic forms and (4) a new high diver-sity assemblage dominated by a Paleocene fauna with bothepi- and endobenthic forms equally represented
Hence based on this succession of benthic assemblageswhich are characterized by strong changes in diversity (H )the KndashPg boundary benthic foraminiferal records from theTethys can be subdivided into four intervals (Figs 2 4ndash6)The intervals I to IV approximately correspond to the up-permost Maastrichtian planktic foraminiferal Zone P0 sub-zones PαndashP1a and subzones P1andashP1b respectively There-fore this succession roughly follows the 5-fold sedimen-tological and planktic foraminiferal sequence of Smit andRomein (1985) in which unit 2 represents the ejecta layerand units 1 3 4 and 5 correspond to intervals I to IV in ourstudy
The sequence of benthic foraminiferal turnover in theTethys is also well illustrated by a Q-mode NM-MDS
(Fig 7) The Q-mode NM-MDS of the benthic foraminiferalsample compositions of the combined data from Okccedilular(this study) and El Kef (Speijer and van der Zwaan 1996)demonstrates that although the assemblages differ betweenthese localities on opposite margins of the Tethys the re-sponses of the benthic foraminiferal assemblages to the KndashPgboundary perturbations express very similar overall patternsacross the KndashPg boundary At both Okccedilular and El Kef thebenthic foraminiferal records show a rapid transition from astable Maastrichtian assemblage to the earliest Danian dis-aster phase in Zone P0 followed by a gradual change backtowards conditions similar to the Maastrichtian (marked bythe return of Lazarus taxa) Therefore both the successionof benthic faunal assemblages including diversity indicesand Q-mode NM-MDS analysis provide powerful tools to al-low the comparison of KndashPg boundary benthic foraminiferalrecords from different localities
52 Ecological responses to reduced export productivity
521 Okccedilular section
The palynological benthic foraminiferal and bulk stable iso-tope records of the Okccedilular section show major changesacross the KndashPg boundary interval (Figs 2ndash7) portraying
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
J Vellekoop et al Ecological response to KndashPg boundary mass extinction 893
Figure 6 Relative abundances of the four main clusters of benthic foraminiferal taxa (gt 125 microm) relative abundances of epibenthic andendobenthic forms and relative abundance of bi-triserial benthic taxa
Figure 7 Q-mode non-metric multidimensional scaling (on all sam-ples in the gt 125 microm size fraction benthic foraminiferal datasets ofOkccedilular (this study) and El Kef (Speijer and van der Zwaan 1996)After an abrupt transition from latest Maastrichtian pre-impact as-semblages to disaster assemblages in Zone P0 and recovery assem-blages in Zone Pα a gradual transition towards a new equilibriumpost-recovery assemblage can be observed in subzones P1a and P1b
the biological crisis following the impact Following the KndashPg boundary mass extinction endobenthic forms stronglydecrease with bi-triserial benthic taxa almost disappearingfrom the benthic community simultaneously the estimatedBFAR shows a major fall (Fig 5) This interval approxi-mately correlative to foraminiferal Zone P0 represents thedisaster phase The drop in bi-triserial benthic taxa (Fig 6)and decrease in BFAR (Fig 5) point to a reduction in foodsupply to the benthic community indicating that the trans-port of organic matter to the sea floor must have decreasedIn the dinocyst community however hexaperidinioids showa strong increase in relative abundance during this phase(Figs 3 6 7) suggesting an increase in nutrient availabilityin the photic zone while the DAR remains relatively stableacross the KndashPg boundary (Fig 5)
The decrease in benthic diversity and the high relativeabundances of opportunistic Trochammina spp and Cibici-doides pseudoacutus suggest that the benthic community ex-perienced additional stress during Zone P0 besides food lim-itation (see Jorissen et al 2007) If food limitation were theonly factor involved upper bathyal assemblages as observedat Okccedilular would be expected to become more similar tooligotrophic middle to lower bathyal assemblages Insteadthe opposite is true the assemblage composition of the dis-aster phase is closer to neritic Midway-type assemblages
It was previously suggested that the epibenthic taxon Cibi-cidoides pseudoacutus could potentially be hypoxia resistant
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
894 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
(Speijer and van der Zwaan 1996) As a collapse of the bi-ological pump should lead to enhanced remineralization oforganic matter in the photic zone it is indeed likely thatthe oxygen minimum zones expanded and shoaled leadingto oxygen stress on the sea floor Although early Paleocenedysoxia at El Kef is supported by the ostracod record (Donzeet al 1982) and ichnofacies analyses (Rodriguez-Tovaret al 2016) supporting (eg geochemical) evidence fordysoxia during the disaster phase at Okccedilular record is absentFurthermore an additional stressor for the benthic commu-nity during Zone P0 may be present In contrast to calcare-ous nanoplankton organic-walled cyst-producing dinoflagel-lates did not suffer extinctions and may have become a moreimportant component in the early Paleocene phytoplanktoncommunity Hence besides the reduction of the amount oforganic matter transported to the sea floor the compositionof food supplied by the photic zone likely changed signifi-cantly across the KndashPg boundary as well (DrsquoHondt 2005)possibly presenting an additional stress factor for the benthiccommunity (Speijer and van der Zwaan 1996 Alegret andThomas 2009)
522 Western Tethys
The patterns in benthic foraminiferal and dinoflagellate re-sponse to the KndashPg boundary perturbations at Okccedilular appearto be characteristic for the continental margins of the westernTethyan realm The benthic foraminiferal patterns are similaracross different size fractions suggesting that they are repre-sentative of the overall benthic communities In Tunisia andSpain where both benthic foraminiferal and dinoflagellaterecords are available the earliest Danian is also characterizedby a decrease in endobenthic foraminifera at the sea floor(Speijer and Van der Zwaan 1996 Peryt et al 2002 Ale-gret et al 2003) and simultaneously blooms of hexaperidin-ioids in the water column (Brinkhuis et al 1998 Vellekoopet al 2015 Fig 8) In the Ouled Haddou section Moroccoin the westernmost Tethys (Fig 1) the lowermost Danian isalso characterized by a strong increase of the hexaperidin-ioid Senegalinium group up to 30 of the assemblage (Sli-mani et al 2010) that is very similar to the Okccedilular record(Fig 8)
The KndashPg boundary dinocyst record of El Kef is strikinglysimilar to the one from the Okccedilular section showing two dis-tinct peaks in hexaperidinioids one in Zone P0 and one atthe basal part of Zone Pα At El Kef the initial post-impactdominance of hexaperidinioids is however less pronounced(Fig 8) This difference in expression might be related tosmall differences in paleogeographic and paleoceanographicsettings between sites
In addition some high-resolution benthic KndashPg records inthe Tethyan realm show a short-lived bloom of specific en-dobenthic foraminiferal taxa in the few centimeter directlyabove the KndashPg boundary sometimes accompanied by otherindicators of low oxygen levels (eg Coccioni and Gale-
otti 1994 Speijer and Van der Zwaan 1996 Kaiho et al1999) This suggests that at least locally the sea floor wastemporarily characterized by hypoxic conditions directly fol-lowing the impact This short-lived deoxygenation is possi-bly related to the mass mortality at the KndashPg boundary re-sulting in a large short-lived flux of organic matter to thesea floor (Coccioni and Galeotti 1994) However since ex-port productivity was greatly reduced after the KndashPg bound-ary mass extinction very little ldquonewrdquo organic matter reachedthe sea floor after the initial post-impact influx Hence afteraccumulated organic matter was remineralized the benthiccommunity starved resulting in a transition to an epibenthic-dominated benthic fauna similar to the post-impact fauna ofthe Okccedilular record
The combined dinocyst records suggest that the early Pa-leocene margins of the Tethys were characterized by an in-crease in nutrient availability in the photic zone whereascoeval benthic foraminiferal records indicate a major de-crease in nutrient supply to the sea floor These changesin the benthic foraminiferal and dinocyst communities arelikely caused by the major reduction of both the efficiencyand strength of the biological pump (ie the fraction ofprimary production exported from the photic zone and theamount of organic matter transported down) in accordancewith the Living Ocean model (DrsquoHondt and Zachos 1998DrsquoHondt et al 1998 DrsquoHondt 2005 Coxall et al 2006)For a detailed explanation of the difference between biologi-cal pump efficiency and biological pump strength see Hiltinget al (2008)
As bulk carbonate δ13C reflects the isotopic compositionof the surface ocean which is set by burial fractions and bythe photosynthetic isotope effect (Kump 1991 Hain et al2014) the excursion recorded in bulk δ13C records world-wide and the rapid collapse in surface to deep-ocean carbonisotope gradients likely reflect the reduction of the global in-tensity (ie efficiency) of the biological pump (Hain et al2014) The recorded inverse change in nutrient availabilitysuggests a causal link A reduced efficiency of the biologi-cal pump and associated decrease of the fraction of biomasstransported out of the photic zone could have resulted inhigh rates of nutrient recycling in the upper part of the wa-ter column (DrsquoHondt 2005) This suggests that the reduc-tion of the biological pump strength recorded by the benthicforaminiferal record is a consequence of the decreased effi-ciency of the biological pump recorded by the carbon iso-tope and dinoflagellate cyst records The strong correlationbetween the bulk carbon isotope curves and the Shannondiversity index (H ) of the benthic foraminiferal records atboth Okccedilular (R2
= 073 p lt 0001) and El Kef (R2= 074
p lt 0001) suggest a clear link between the changes in ben-thic foraminiferal assemblages and the collapse and recoveryof biological pump efficiency
As a result of the reduced biological pump efficiency morenutrients remained available for the early Paleocene phyto-plankton community At Tethyan margin sites this is indi-
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
J Vellekoop et al Ecological response to KndashPg boundary mass extinction 895
Figure 8 The benthic foraminiferal (gt 125 microm) dinocyst and bulk stable carbon isotope records of Okccedilular (Turkey) and El Kef (Tunisia)the dinocyst record of Ouled Haddou (Morocco) and the benthic foraminiferal record (gt 63 microm) of ODP Hole 1049C Blake Nose (NorthAtlantic) Note that the definition of ldquohexaperidinioidsrdquo used is provided in the text The four phases identified based on the bulk stablecarbon isotopes and the Shannon diversity index (H ) of the benthic foraminiferal records are indicated (IndashIV) The gt 63 microm size fractionbenthic foraminiferal record of ODP Hole 1049C is from Alegret and Thomas (2004) who argued that the lowermost Paleocene of thissite comprised reworked foraminifera This interval is indicated by a dashed bar in the figure The biostratigraphy of Ouled Haddou is fromSlimani and Toufiq (2013) whereas the dinocyst record of this site is from Slimani et al (2010) The biostratigraphy of El Kef is based onBrinkhuis et al (1998) Speijer and van der Zwaan (1996) and Molina et al (2006) The fine-fraction stable carbon isotope record (blackdots) of El Kef is from Keller and Lindinger (1989) The Shannon diversity index (red dots) relative abundances of bi-triserial benthic taxa(dark purple) and all endobenthic foraminifera (light purple) are based on the gt 125 microm size fraction foraminiferal data of Speijer and vander Zwaan (1996) The palynological data of El Kef are from Brinkhuis et al (1998) (lower part) and Guasti et al (2005) (upper part) TheShannon diversity index of the gt 125 microm size fraction benthic foraminiferal record (red dots) the relative contribution of bi-triserial benthictaxa (dark purple) all endobenthic foraminifera (light purple) and hexaperidinioid dinocysts from Okccedilular are from this study The bulkstable carbon isotope record of Okccedilular (black dots) is from Accedilikalin et al (2015) Mbsf indicates meters below sea floor
cated by the higher abundance of hexaperidinioids Similarlyblooms of eutrophic survivor taxa of calcareous nanoplank-ton at open ocean sites have been suggested to be relatedto the buildup of nutrients in the open ocean photic zone(Schueth et al 2015) Although the carbon isotope gradientsbetween surface and deep waters indicate that the recoveryof the biological pump took hundreds of thousands of years(Zachos et al 1989 Kump 1991 DrsquoHondt et al 1998 Cox-all et al 2006 Birch et al 2016) our records suggest thatthe enhanced recycling of nutrients in the Tethys was partic-ularly intense during two phases in the first tens of thousandsyears after the impact (Fig 8)
523 Global responses
The decrease in export productivity following the KndashPgboundary is not only recognized in the Tethyan realm In theAtlantic Ocean Southern Ocean and the Indian Ocean a de-crease of the biological pump strength after the KndashPg bound-ary was also observed in neritic to abyssal environments(eg Thomas 1990 Olsson et al 1996 Hull et al 2011Alegret et al 2012) As an example the gt 63 microm size frac-tion benthic foraminiferal KndashPg boundary record of BlakeNose (ODP Hole 1049C northwestern Atlantic Alegret andThomas 2004) shows a pattern that is strikingly similar tothe gt 125 microm size fraction records in the Tethyan realm witha strong decrease of endobenthics in Zone P0 followed by a
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
896 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
gradual recovery across zones Pα and P1a (Fig 8) The simi-larities between the benthic foraminiferal patterns of Tethyanouter neritic to middle bathyal (200ndash1500 m) sites and thoseof the middle to lower bathyal (1500ndash1600 m) record ofBlake Nose (Alegret and Thomas 2004) suggest that theserecords register a global decrease in export productivity fol-lowing the KndashPg boundary
However several open ocean sites mostly in the Pacificshow an opposite trend with several lines of evidence sug-gesting an increased biological pump strength at these sites(eg Hull et al 2011 Alegret et al 2012) This led Ale-gret et al (2012) to conclude that the decrease in biologicalpump strength after the KndashPg boundary was most likely aregional instead of a global effect arguing against the Liv-ing Ocean hypothesis We would argue that low-productivityopen ocean sites like the central Pacific are characterized byvery different ecosystem structures and higher productivityas those along the Tethyan margins (eg Dortch and Pack-erd 1989) Therefore the consequence of ecosystem reorga-nization resulting from the mass extinction was likely verydifferent in the open ocean Pacific (eg Sibert et al 2014)Esmeray-Senlet et al (2015) proposed the term ldquoHeteroge-neous Oceanrdquo for this conceptual model as an alternativeto the Living Ocean model Supposedly the HeterogeneousOcean was characterized by a strong geographic heterogene-ity in the extinction patterns and food supply to the sea floor
However as the primary food source for open ocean deep-sea benthic communities generally consists of refractory or-ganic matter (eg Jiao et al 2010) it remains uncertainwhether changes in the intensity of recycling of the gener-ally more labile organic matter in the surface ocean will re-sult in changes in the amount of organic matter that benthiccommunities receive as they will likely continue to receiverecalcitrant organic matter at barely diminished rates despitethe collapse of the biological pump
In addition it should be noted that although the bio-logical pump efficiency is generally relatively high in openocean realms such as the central Pacific the biological pumpstrength is generally very low at these sites (Honjo et al2008 Henson et al 2011) Small changes in absolute bio-logical pump strength at such a locality could therefore havehad a large effect on the local oligotrophic benthic commu-nity (Alegret and Thomas 2009) but they likely representedonly a small fraction of the net amount of organic matterglobally transported to the deep Hence even though localbiological pump strength might have increased at some low-productivity deep-sea sites the large decrease in biologi-cal pump strength recorded at many other sites means thatthe net amount of organic matter globally transported out ofthe surface ocean was likely still reduced in the post-impactworld (Birch et al 2016) Therefore we argue that such Pa-cific records do not represent the general pattern The globalocean response can be characterized as heterogeneous how-ever the KndashPg boundary mass extinction did lead to a strongreduction of the net amount of organic matter globally trans-
ported from the surface to the sea floor while biological pro-ductivity recovered rapidly after the extinction event in ac-cordance with the Living Ocean hypothesis
53 Long-term recovery
Following the disaster phase the hexaperidinioid cysts atOkccedilular and El Kef decreased in abundance and the abun-dance of endobenthic forms increased again as the recoveryof the benthic community was initiated This recovery phaseis approximately correlative to Zone Pα and the basal part ofZone P1a which according to the Paleogene age constraintsof Vandenberghe et al (2012) represents at least 300 kyrThis duration is in agreement with the estimations providedby Birch et al (2016) During this phase the carbon iso-tope records remain well below pre-impact values and thediversity of the benthic foraminiferal community has not yetfully recovered (Figs 4ndash8) This indicates that the rapid andshort-lived KndashPg boundary disaster was followed by a rela-tively long recovery phase in line with previous estimatesof a multimillion-year biological recovery (eg Coxall etal 2006) While the impact-related environmental perturba-tions were short lived (Kring 2007 Vellekoop et al 20142016) the extinctions amongst important biological groupsled to a reduction of the organic flux from the photic zone todeep water resulting in major long-term biological and pa-leoceanographic reorganizations Only with the evolutionaryrecovery of the pelagic community governing the biologicalcarbon pump did export productivity start to increase again(eg Coxall et al 2006 Birch et al 2016)
6 Conclusions
The marine palynological benthic foraminiferal and bulkstable isotope records of the Okccedilular and El Kef sections re-veal major changes across the KndashPg boundary interval por-traying the biological crisis at the KndashPg boundary and subse-quent recovery in the early Paleocene Based on the succes-sion of benthic faunal assemblages at a number of Tethyanmargin sites four phases can be recognized across the KndashPgboundary interval the Maastrichtian or pre-impact phase (I)a disaster phase (II) a recovery phase (III) and an early Pa-leocene post-recovery phase (IV)
Following the KndashPg boundary impact some localitiesshow a large short-lived flux of food to the sea floor likelyrelated to the mass mortality at the KndashPg boundary Howeversince export productivity was greatly reduced after the KndashPgboundary mass extinction it is likely that after the initialpost-impact influx very little fresh organic matter reachedthe sea floor This reduction of export productivity in thepost-extinction disaster phase eventually resulted in a lowerfood supply to the sea floor This presented a major stressfactor for benthic organisms in the western Tethys The re-duced food supply resulted in an abrupt impoverishment of
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
J Vellekoop et al Ecological response to KndashPg boundary mass extinction 897
benthic communities As the downward transport of nutri-ents was slowed down recycling in upper layers increasedAs a result more nutrients remained available for the earlyPaleocene phytoplankton community leading to blooms ofdinoflagellates along the Tethyan margins and blooms of cal-careous nanoplankton taxa in more open ocean sites Ourrecords show that the enhanced recycling of nutrients in theTethys was particularly intense in the tens of thousands ofyears after the impact Following this the slow evolution-ary recovery of the pelagic community governing the bio-logical carbon pump resulted in a gradually increasing ex-port productivity in the hundreds of thousands of years afterthe impact Hence our integration of dinocyst and benthicforaminiferal records across the KndashPg boundary provides im-portant new insights into the ecological responses to the re-duction of export productivity following the mass extinctionat the CretaceousndashPaleogene boundary highlighting the di-rect ecological consequences of the Living Ocean conditionsin the post-impact world
7 Data availability
All data related to publication are available in the Supple-ment
The Supplement related to this article is available onlineat doi105194bg-14-885-2017-supplement
Author contributions Johan Vellekoop Lineke Woelders andHenk Brinkhuis designed the research Samples were collectedin the field by Johan Vellekoop Sanem Accedilikalin Ismail Ouml Yil-maz and Jan Smit Palynological analyses were carried out by Jo-han Vellekoop Benthic foraminiferal analyses were carried out byLineke Woelders and Robert P Speijer Johan Vellekoop LinekeWoelders and Bas van de Schootbrugge prepared the manuscriptwith input from all authors
Competing interests The authors declare that they have no conflictof interest
Acknowledgements This work was supported by the NetherlandsOrganization for Scientific Research (NWO) Open CompetitionGrant ALWPJ09047 to H Brinkhuis and the Research FoundationFlanders (FWO) grant G0B8513 to R P Speijer We thank LKump F Boscolo Galazzo P Roopnarine and one anonymousreferee for their suggestions
Edited by D GillikinReviewed by L Kump F Boscolo Galazzo and P Roopnarine
References
Accedilikalin S Vellekoop J Ocakoglu F Yılmaz I Ouml Smit J Al-tiner S O Goderis S Vonhof H Speijer R P WoeldersL Fornaciari E and Brinkhuis H Geochemical and pale-ontological characterization of a new KndashPg Boundary local-ity from the Northern branch of the Neo-Tethys MudurnundashGoumlynuumlk Basin NW Turkey Cretaceous Res 52 251ndash267doi101016jcretres201407011 2015
Alegret L and Thomas E Food supply to the seafloor in the Pa-cific Ocean after the CretaceousPaleogene boundary event MarMicropaleontol 73 105ndash116 2009
Alegret L Molina E and Thomas E Benthic foraminiferalturnover across the CretaceousPaleogene boundary at Agost(southeastern Spain) paleoenvironmental inferences Mar Mi-cropaleontol 48 251ndash279 2003
Alegret L Thomas E and Lohmann K C End-Cretaceous ma-rine mass extinction not caused by productivity collapse P NatlAcad Sci USA 109 728ndash732 doi101073pnas11106011092012
Alegret L Rodriacuteguez-Tovar F J and Uchman A How bioturba-tion obscured the CretaceousndashPaleogene boundary record TerraNova 27 225ndash230 doi101111ter12151 2015
Altiner D Koccedilyigit A Farinacci A Nicosia U and ContiM A Jurassic-Lower Cretaceous stratigraphy and paleogeo-graphic evolution of the southern part of north-western Anatolia(Turkey) Geologica Romana 27 13ndash80 1991
Alvarez LW Alvarez W Asaro F and Michel H V Extrater-restrial cause for the Cretaceous-Tertiary extinction Science208 1095ndash1108 1980
Alegret L and Thomas E Benthic foraminifera and environmen-tal turnover across the CretaceousPaleogene boundary at BlakeNose (ODP Hole 1049C Northwestern Atlantic) PalaeogeogrPalaeocl 208 59ndash83 doi101016jpalaeo200402028 2004
Armstrong R A Lee C Hedges J I Honjo S and WakehamS G A new mechanistic model for organic carbon fluxes in theocean based on the quantitative association of POC with ballastmaterials Deep-Sea Res Pt II 49 219ndash236 2002
Aubert J and Berggren W A Paleocene benthic foraminiferalbiostratigraphy and paleoecology of Tunisia Bull du Cent deRech de Pau ndash SNPA 10 379ndash469 1976
Berggren W A and Aubert J Paleocene benthonic foraminiferalbiostratigraphy paleobiogeography and paleoecology ofAtlanticndashTethyan regions midway-type fauna PalaeogeogrPalaeocl 18 73ndash192 1975
Bernaola G and Monechi S Calcareous nannofossil ex-tinction and survivorship across the CretaceousndashPaleogeneboundary at Walvis Ridge (ODP Hole 1262C SouthAtlantic Ocean) Palaeogeogr Palaeocl 255 132ndash156doi101016jpalaeo200702045 2007
Bernhard J M Characteristic assemblages and morphologies ofbenthic foraminifera from anoxic organic-rich deposits Jurassicthrough Holocene J Foramin Res 16 207ndash215 1986
Birch H S Coxall H K Pearson P N Kroon D andSchmidt D N Partial collapse of the marine carbon pump af-ter the CretaceousndashPaleogene boundary Geology 44 287ndash290doi101130G375811 2016
Brinkhuis H and Zachariasse W J Dinoflagellate cysts sealevel changes and planktonic foraminifers across the Cretaceous-
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
898 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
Tertiary boundary at El Haria northwest Tunisia Mar Micropa-leontol 13 153ndash191 1988
Brinkhuis H Bujak J P Smit J Versteegh G J M and Viss-cher H Dinoflagellate-based sea surface temperature recon-structions across the Cretaceous-Tertiary boundary PalaeogeogrPalaeocl 141 67ndash83 1998
Buesseler K O The decoupling of production and particulate ex-port in the surface ocean Global Biogeochem Cy 12 297ndash3101998
Buzas M A Culver S J and Jorissen F J A statisti-cal evaluation of the microhabitats of living (stained) infau-nal benthic foraminifera Mar Micropaleontol 20 311ndash320doi1010160377-8398(93)90040-5 1993
Coccioni R and Galeotti S KndashT boundary extinction geolog-ically instantaneous or gradual event Evidence from deep-seabenthic foraminifera Geology 22 779ndash782 1994
Corliss B H Microhabitats of benthic foraminifera within deep-sea sediments Nature 314 435ndash438 doi101038314435a01985
Corliss B H and Chen C Morphotype patterns of Nor-wegian Sea deep-sea benthic foraminifera and ecologi-cal implications Geology 16 716ndash719 doi1011300091-7613(1988)016lt0716MPONSDgt23CO2 1988
Coxall H K DrsquoHondt S and Zachos J C Pelagic evolution andenvironmental recovery after the CretaceousndashPaleogene mass ex-tinction Geology 34 297ndash300 doi101130G217021 2006
Culver S J Benthic foraminifera across the CretaceousndashTertiary(KndashT) boundary a review Mar Micropaleontol 47 177ndash2262003
Cushman J A Upper Cretaceous foraminifera of the Gulf coastalregion of the United States and adjacent areas US Geolog SurvProf Pap 206 241 pp 1946
Cushman J A Paleocene foraminifera of the Gulf Coastal Regionof the United States and adjacent regions US Geol Surv ProfPap 232 75 pp 1951
DrsquoHondt S Consequences of the CretaceousPaleogene Mass Ex-tinction for Marine Ecosystems Annu Rev Ecol Evol S36 295ndash317 doi101146annurevecolsys350211031057152005
DrsquoHondt S and Zachos J C Cretaceous foraminifera and the evo-lutionary history of planktic photosymbiosis Paleobiology 24512ndash523 1998
DrsquoHondt S Donaghay P Zachos J C Luttenberg D andLindinger M Organic Carbon Fluxes and Ecological Recov-ery from the Cretaceous-Tertiary Mass Extinction Science 282276ndash279 1998
Donze P Colin J-P Damotte R Oertli HJ Peypouquet J-Pand Said R Les ostracodes du Campanien terminal agrave lrsquoEocegraveneinfeacuterieur de la coupe du Kef Tuneacutesie nord-occidentale Bull desCentr Recher Expl-Prod Elf Aquit 6 273ndash335 1982
Dortch Q and Packard T T Differences in biomass structure be-tween oligotrophic and eutrophic marine ecosystems Deep-SeaRes 36 223ndash240 1989
Ernst S R Guasti E Dupuis C and Speijer R P En-vironmental perturbation in the southern Tethys across thePaleoceneEocene boundary (Dababiya Egypt) Foraminiferaland clay mineral records Mar Micropaleontol 60 89ndash111doi101016jmarmicro200603002 2006
Eshet Y Moshkovitz S Habib D Benjamini C and Maga-ritz M Calcareous nannofossil and dinoflagellate stratigraphyacross the CretaceousTertiary boundary at Hor Hahar IsraelMar Micropaleontol 18 199ndash228 1992
Eshet Y Almogi-Labin A and Bein A Dinoflagellate cysts pa-leoproductivity and upwelling systems A Late Cretaceous ex-ample from Israel Mar Micropaleontol 23 231ndash240 1994
Esmeray-Senlet S Wright J D Olsson R K MillerK G Browning J V and Quan T M Evidencefor reduced export productivity following the Creta-ceousPaleogene mass extinction Paleoceanography 30718ndash738 doi1010022014PA002724 2015
Giusberti L Boscolo Galazzo F and Thomas E Variability inclimate and productivity during the Paleocene-Eocene ThermalMaximum in the western Tethys (Forada section) Clim Past12 213ndash240 doi105194cp-12-213-2016 2016
Guasti E Kouwenhoven T J Brinkhuis H and Speijer R PPaleocene sea-level and productivity changes at the southernTethyan margin (El Kef Tunisia) Mar Micropaleontol 55 1ndash17 2005
Habib D and Saeedi F The Manumiella seelandica global spikeCooling during regression at the close of the MaastrichtianPalaeogeogr Palaeocl 255 87ndash97 2007
Hain M P Sigmal D M and Haug G H 818 ndash The biolog-ical Pump in the Past in Reference Module in Earth Systemsand Environmental Sciences Treatise on Geochemistry (SecondEdition) The Oceans and Marine Geochemistry 8 485ndash517doi101016B978-0-08-095975-700618-5 2014
Hayek L C and Buzas M A On the proper and efficient useof diversity measures with individual field samples J ForaminRes 43 305ndash313 2013
Henson S A Sanders R Madsen E Morris P J Le MoigneF and Quartly G D A reduced estimate of the strength ofthe oceanrsquos biological carbon pump Geophys Res Lett 38L04606 doi1010292011GL046735 2011
Hilting A K Kump L R and Bralower T J Variations in theoceanic vertical carbon isotope gradient and their implicationsfor the Paleocene-Eocene biological pump Paleoceanography23 PA3222 doi1010292007PA001458 2008
Honjo S Manganini S J Krishfield R A and Francois RParticulate organic carbon fluxes to the ocean interior and fac-tors controlling the biological pump A synthesis of global sed-iment trap progams since 1983 Prog Oceanogr 76 217ndash285doi101016jpocean200711003 2008
Huber B T MacLeod K G and Norris R D Abrupt extinctionand subsequent reworking of Cretaceous planktonic foraminiferaacross the Cretaceous-Tertiary boundary Evidence from the sub-tropical North Atlantic in Catastrophic Events and Mass Extinc-tions Impacts and Beyond edited by Koeberl C and MacLeodK G Geol S Am S 356 277ndash289 Boulder Colorado USA2002
Hull P M Norris R D Bralower T J and Schueth J D A rolefor chance in marine recovery from the end-Cretaceous extinc-tion Nat Geosci 4 856ndash860 doi101038ngeo1302 2011
Hsuuml K J and McKenzie J A A ldquoStrangeloverdquo ocean in the earli-est Tertiary in Natural variations Archean to present edited bySundquist E T and Broecker W S The carbon cycle and atmo-spheric CO2 American Geophysical Union Washington DCUSA 487ndash492 1985
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
J Vellekoop et al Ecological response to KndashPg boundary mass extinction 899
Jiao N Herndl G J Hansell D A Benner R Kattner G Wil-helm S W Kirchman D L Weinbauer M G Luo T ChenF and Azam F Microbial production of recalcitrant dissolvedorganic matter long-term carbon storage in the global oceanNat Rev 8 593ndash599 doi101038nrmicro2386 2010
Jorissen F J de Stigter H C and Widmark J G V A concep-tual model explaining benthic foraminiferal microhabitats MarMicropaleontol 26 3ndash15 doi1010160377-8398(95)00047-X1995
Jorissen F J Fontanier C and Thomas E Paleoceanographi-cal proxies based on deep-sea benthic foraminiferal assemblagecharacteristics in Proxies in Late Cenozoic PaleoceanographyPt 2 Biological tracers and biomarkers edited by Hillaire-Marcel C and de Vernal A 263ndash326 Elsevier Amsterdamthe Netherlands 2007
Kaiho K Kajiwara Y Tazaki K Ueshima M Takeda NKawahata H Arinobu T Ishiwatari R Hirai A and Lam-olda M A Oceanic primary productivity and dissolved oxygenlevels at the CretaceousTertiary boundary their decrease sub-sequent warming and recovery Paleoceanography 14 511ndash5241999
Keller G and Lindinger M Stable isotope TOC and CaCO3record across the CretaceousTertiary boundary at El KefTunisia Palaeogeogr Palaeocl 73 243ndash265 1989
Kellough G R Paleoecology of the Foraminiferida of the WillsPoint Formation (Midway Group) in northeast Texas Trans GulfCoast Assoc Geol Soc 15 73ndash153 1965
Kring D A The Chicxulub impact event and its environmentalconsequences at the CretaceousndashTertiary boundary PalaeogeogrPalaeocl 255 4ndash21 2007
Kump L R Interpreting carbon-isotope excursions Strangeloveoceans Geology 19 299ndash302 1991
Machalski M Vellekoop J Dubicka Z Peryt D and Ha-rasimiuk M Late Maastrichtian cephalopods dinoflagellatecysts and foraminifera from the CretaceousndashPaleogene suc-cession at Lechoacutewka southeast Poland Stratigraphic andenvironmental implications Cretaceous Res 57 208ndash227doi101016jcretres201508012 2016
Molina E Evidence and causes of the main extinctionevents in the Paleogene based on extinction and survivalpatterns of foraminifera Earth-Sci Rev 140 166ndash181doi101016jearscirev201411008 2015
Molina E Alegret L Arenillas I and Arz J The GlobalBoundary Stratotype Section and Point for the base of the Da-nian Stage (Paleocene Paleogeneldquo Tertiaryrdquo Cenozoic) at ElKef Tunisia-Original definition and revision Episodes 29 263ndash273 2006
Olsson R K Liu C and van Fossen M The Cretaceous-Tertiarycatastrophic event at Millers Ferry Alabama in The Cretaceous-Tertiary Event and Other Catastrophes in Earth History editedby Ryder G Fastovsky D and Gartner S Geol S Am S307 263ndash277 1996
Peryt D Benthic foraminiferal response to the Cenomanian-Turonian and CretaceousndashPaleogene boundary events PrzegladGeol 52 827ndash832 2004
Peryt D Alegret L and Molina E The CretaceousPalaeogene(KP) boundary at Aiumln Settara Tunisia restructuring of benthicforaminiferal assemblages Terra Nova 14 101ndash107 2002
Ramette A Multivariate analyses in microbial ecology FEMS Mi-crobiol Ecol 62 142ndash160 2007
Rodriacuteguez-Tovar F J Uchman A MrsquoHamdi A Riahi Sand Ismail-Lattrache K B Ichnological record of palaeoen-vironment from the CretaceousndashPaleogene boundary intervalat El Kef Tunisia The first study of old and new sec-tions at the stratotype area J Afr Earth Sci 120 23ndash30doi101016jjafrearsci201604017 2016
Saner S Mudurnu-Goumlynuumlk Havzasının Jura ve Sonrasıccediloumlkelimnitelikleriyle paleocografya yorumlaması TJK Buumllteni 23 39ndash52 1980
Schioslashler P Brinkhuis H Roncaglia L and Wilson G J Di-noflagellate biostratigraphy and sequence stratigraphy of theType Maastrichtian (Upper Cretaceous) ENCI Quarry TheNetherlands Mar Micropaleontol 31 65ndash95 1997
Schueth J D Bralower T J Jiang S and Patzkowsky ME The role of regional survivor incumbency in the evolu-tionary recovery of calcareous nannoplankton from the Creta-ceousPaleogene (KPg) mass extinction Paleobiology 41 661ndash679 doi101017pab201528 2015
Scotese C R A continental drift flipbook J Geology 112 729ndash741 2004
Scotese C R and Dreher C GlobalGeology available at httpwwwGlobalGeologycom (last access 11 March 2014) 2012
Sepkoski J J Patterns of Phanerozoic extinction a perspectivefrom global data bases in Global Events and Event Stratigra-phy edited by Walliser O H Springer Berlin Germany 35ndash52 2016
Sibert E C Hull P M and Norris R D Resilience of Pacificpelagic fish across the CretaceousPalaeogene mass extinctionNat Geosci 7 667ndash670 doi101038NGEO2227 2014
Slimani H and Toufiq A A CretaceousndashPaleogene boundary geo-logical site revealed by planktic foraminifera and dinoflagellatecysts at Ouled Haddou eastern external Rif Chain Morocco JAfr Earth Sci 88 38ndash52 doi101016jjafrearsci2013080082013
Slimani H Louwye S and Toufiq A Dinoflagellate cystsform the CretaceousndashPaleogene boundary at Ouled Had-dou southeastern Rif Morocco biostratigraphy paleoenvi-ronments and paleobiogeography Palynology 34 90ndash124doi10108001916121003629933 2010
Sluijs A and Brinkhuis H A dynamic climate and ecosystemstate during the Paleocene-Eocene Thermal Maximum infer-ences from dinoflagellate cyst assemblages on the New JerseyShelf Biogeosciences 6 1755ndash1781 doi105194bg-6-1755-2009 2009
Smit J Extinction and evolution of planktonic foraminifera after amajor impact at the CretaceousTertiary boundary Geol S AmS 190 329ndash352 1982
Smit J and Romein A J T A sequence of events across theCretaceous-Tertiary boundary Earth Planet Sci Lett 74 155ndash170 1985
Speijer R P Extinction and recovery patterns in ben-thic foraminiferal paleocommunities across the Creta-ceousPaleogene and PaleoceneEocene boundaries GeologicaUltraiectina 124 Faculteit Aardwetenschappen PhD ThesisUniversiteit Utrecht Utrecht the Netherlands 1994
Speijer R P and van Der Zwaan G J Extinction and survivor-ship of southern Tethyan benthic foraminifera across the Cre-
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
900 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
taceousPaleogene boundary in Biotic recovery from mass ex-tinction events edited by Hart M B Geolog Soc Spec PublLondon UK 102 343ndash371 1996
Stemmann L and Boss E Plankton and Particle Size andPackaging From Determining Optical Properties to Drivingthe Biological Pump Annu Rev Mar Sci 4 263ndash290doi101146annurev-marine-120710-100853 2012
Thomas E Late Cretaceous-early Eocene mass extinctions in thedeep sea Geol S Am S 247 481ndash495 1990
Vandenberghe N Hilgen F J and Speijer R P The PaleogenePeriod in The Geologic Time Scale 2012 edited by GradsteinF M Ogg J G Schmitz M D and Ogg G M 2 855ndash921Elsevier Boston USA 2012
Vellekoop J Sluijs A Smit J Schouten S Weijers J WH Sinninghe Damsteacute J S and Brinkhuis H Rapid short-term cooling following the Chicxulub impact at the CretaceousndashPaleogene boundary P Natl Acad Sci USA 111 7537ndash7541doi101073pnas1319253111 2014
Vellekoop J Smit J van de Schootbrugge B Weijers J W HGaleotti S Sinninghe Damsteacute J S and Brinkhuis H Palyno-logical evidence for prolonged cooling along the Tunisian conti-nental shelf following the KndashPg boundary impact PalaeogeogrPalaeocl 426 216ndash228 doi101016jpalaeo2015030212015
Vellekoop J Esmeray-Senlet S Miller K G Browning J VSluijs A van de Schootbrugge B Sinninghe Damsteacute J Sand Brinkhuis H Evidence for CretaceousndashPaleogene bound-ary bolide ldquoimpact winterrdquo conditions from New Jersey USAGeology 44 619ndash622 doi101130G379611 2016
Woelders L and Speijer R Stable seafloor conditions sealevel and food supply during the latest Maastrichtian atBrazos River Texas Mar Micropaleontol 121 41ndash51doi101016jmarmicro201510002 2015
Zachos J C Arthur M A and Dean W E Geochemical evi-dence for suppression of pelagic marine productivity at the Cre-taceousTertiary boundary Nature 337 61ndash64 1989
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
- Abstract
- Introduction
- Geological setting and age assessment
- Materials and methods
-
- Sampling
- Foraminiferal analysis
- Palynological analysis
- Statistical analysis
-
- Results
-
- Benthic foraminifera
- Palynology
-
- Discussion
-
- Tethyan benthic foraminiferal turnover sequence
- Ecological responses to reduced export productivity
-
- Okccedilular section
- Western Tethys
- Global responses
-
- Long-term recovery
-
- Conclusions
- Data availability
- Author contributions
- Competing interests
- Acknowledgements
- References
-

886 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
amongst primary producers caused a major restructuring ofglobal food webs and carbon cycling (DrsquoHondt 2005 Cox-all et al 2006) Moreover a collapse in the oceanic stablecarbon isotope gradient between surface and bottom waterspersisted for up to a few million years (Hsuuml and McKenzie1985 Zachos et al 1989 DrsquoHondt et al 1998) Initiallythe ldquoStrangelove Oceanrdquo hypothesis was invoked to explainthis collapse suggesting that primary productivity sharplydecreased or ceased immediately after the KndashPg boundary asa consequence of the extinction of primary producers (egHsuuml and McKenzie 1985)
However modeling the carbon isotope gradient responseto the extinctions suggests that carbon burial had to continuenearly unabated to prevent the carbon isotopic signature ofthe global ocean from drifting towards that of the weath-ering input (Kump 1991) In addition both the persistenceof certain primary producers such as dinoflagellates (egBrinkhuis and Zachariasse 1988) as well as the survivalof benthic foraminifera (eg Culver 2003) argue againstprolonged cessation of primary productivity in the globaloceans Integration of neritic and deep-sea planktic and ben-thic foraminiferal carbon isotopic records suggests that thebreakdown of this gradient reflects a global collapse of exportproductivity ie the fraction of organic carbon that sinksfrom the photic zone to the deep ocean via the biologicalpump (Kump 1991 DrsquoHondt et al 1998 DrsquoHondt 2005Hilting et al 2008 Hain et al 2014 Esmeray-Senlet etal 2015) rather than the shutdown of primary productivityThis conceptual model is generally referred to as the ldquoLiv-ing Oceanrdquo model (DrsquoHondt and Zachos 1998 DrsquoHondt etal 1998 DrsquoHondt 2005) According to the Living Oceanmodel total biological productivity recovered rapidly afterthe extinction event but the total global export productivityfrom the photic zone was reduced for hundreds of thousandsof years (DrsquoHondt et al 1998 DrsquoHondt 2005 Birch et al2016) Although there has been considerable debate on thispossible collapse of the biological pump following the KndashPgboundary mass extinction (eg Alegret and Thomas 2009Hull et al 2011) most recent studies are in agreement thatglobal post-KndashPg export productivity was reduced to someextent (eg Esmerey-Senlet et al 2015 Birch et al 2016)Current discussions are mostly related to the severity of thereduction of export productivity or the geographical differ-ences (ie the ldquoheterogeneityrdquo) of the oceans In particularthe records from the Pacific open ocean could potentiallybe explained as showing no reduction of the biological pump(eg Alegret et al 2012 Hull et al 2011) while sites fromthe Tethys Ocean Atlantic Ocean Southern Ocean and In-dian Ocean show convincing evidence for a collapse of a bi-ological pump at the KndashPg boundary (eg Thomas 1990Olsson et al 1996 Hull et al 2011 Alegret et al 2012)
The inferred reduction in the organic flux to deep watersrecorded at most sites might be a consequence of the ecosys-tem reorganization that resulted from the mass extinctionVariations in species assemblages of pelagic communities
can lead to large changes in the rates of particulate export(eg Buesseler 1998 Stemmann and Boss 2012) A generalreduction of the number of pelagic grazers (such as macro-zooplankton) or a shift in dominance from grazers that cre-ate fecal pellets (eg fish) to grazers that do not produce fe-cal pellets (eg dinoflagellates) could have greatly reducedthe packaging of biomass into large particles sinking to thedeep ocean (DrsquoHondt et al 1998 DrsquoHondt 2005) This isexemplified by the record at Gubbio Italy which indicatesan abrupt decline of fish stocks at the KndashPg boundary re-maining depressed for millions of years in the western Tethys(Sibert et al 2014)
Additionally the replacement of large tests of Creta-ceous planktic foraminifera and calcareous nanoplankton bysmaller early Paleocene forms (eg Bernaola and Monechi2007 Molina 2015) might have reduced biomineral ballast-ing (Armstrong et al 2002) during the early Paleocene re-sulting in a further reduction of the carbon flux to the oceanfloor (Coxall et al 2006)
Although numerous studies have been performed to seekphysical evidence for the KndashPg boundary impact the result-ing millennial-scale biotic responses are still poorly under-stood Some of the most intensely studied microfossil groupsused in paleoenvironmental reconstructions such as plankticforaminifera and calcareous nanoplankton experienced ma-jor extinctions and subsequent radiations (eg Smit 1982Huber et al 2002 Coxall et al 2006 Molina 2015 Schuethet al 2015) hampering biota-based environmental recon-structions across the KndashPg boundary interval In contrastbenthic foraminifera and organic-cyst-producing dinoflagel-lates are much better suited as they show no significant ex-tinction above background levels at the end of the Cretaceous(Brinkhuis and Zachariasse 1988 Culver 2003)
A few high-resolution KndashPg boundary benthicforaminiferal and organic-walled dinoflagellate cysts(dinocyst) records have been published particularly fromthe southern and western shallow margins of the TethysOcean for example from Tunisia Israel Egypt Spain andMorocco (Brinkhuis and Zachariasse 1988 Eshet et al1992 Coccioni and Galeotti 1994 Speijer and van derZwaan 1996 Brinkhuis et al 1998 Peryt et al 2002Alegret et al 2003 Slimani et al 2010 Vellekoop et al2015) (Fig 1) These records thus potentially provide acomprehensive Tethys-wide portrayal of the changes inpelagic-benthic-coupling across the KndashPg boundary Thebenthic foraminiferal and dinocyst records from the south-ern margin of the Tethys do reveal indications for majorshort-term oceanographic changes in eg temperatureredox and trophic conditions across the KndashPg boundary(eg Brinkhuis and Zachariasse 1988 Speijer and van derZwaan 1996 Brinkhuis et al 1998) Especially quantitativebenthic foraminiferal records show a strong response to theimpact generally portraying an abrupt benthic communityimpoverishment across the boundary At many of theseKndashPg boundary sites after an initial short-lived prolifer-
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
J Vellekoop et al Ecological response to KndashPg boundary mass extinction 887
Figure 1 Reconstruction of the late Cretaceousndashearly Paleogene paleogeography of the North Atlantic and Mediterranean regions afterScotese (2004) and Scotese and Dreher (2012) The four sites discussed are indicated Okccedilular section Turkey (Accedilikalin et al 2015 thisstudy) El Kef Tunisia (Brinkhuis and Zachariasse 1988 Speijer and van der Zwaan 1996 Brinkhuis et al 1998) Ouled Haddou Morocco(Slimani et al 2010) and ODP Hole 1049C (Blake Nose Alegret and Thomas 2004) The red dot marks the Chicxulub impact crater
ation of endobenthic forms (eg Coccioni and Galeotti1994 Alegret et al 2015) epibenthic forms dominatethe post-impact ldquodisasterrdquo phase (Culver 2003) Since ingeneral endobenthic forms are considered indicative of ahigh flux of organic matter to the sea floor andor relativelylow oxygen conditions and epibenthic forms are generallymore common in oligotrophic open marine environments(eg Corliss 1985 Peryt et al 2002 Jorissen et al 2007Woelders and Speijer 2015) the post-impact abundance ofepibenthic forms is often explained as food starvation atthe sea floor (Culver 2003) Following this disaster phasemost benthic foraminiferal records show a relatively longrecovery phase with endobenthic forms gradually returningas diversity starts to increase again (Alegret et al 2003Culver 2003) and endobenthic microhabitats diversify(Speijer and van der Zwaan 1996)
Although combining quantitative dinocyst and benthicforaminifera analyses could provide crucial insight intochanges in pelagic-benthic-coupling (eg Guasti et al2005) perhaps surprisingly no such attempts have beenmade for the KndashPg boundary interval so far Earlier stud-ies discussed either benthic foraminiferal or dinoflagellateresponse to the KndashPg boundary each without making an at-tempt to combine the results of these studies into a compre-hensive integrated ocean-wide characterization of ecologi-cal patterns observed across the boundary In addition al-though the southern margins of the western Tethys provideda fair number of high-resolution records spanning the KndashPgboundary no such high-resolution records yet exist from thenortheastern margins of the western Tethys To be able toprovide a comprehensive (western) Tethys-wide portrayal ofthe surface and bottom water ecological changes across theKndashPg boundary and the coupling between pelagic and ben-thic systems additional dinocyst and benthic foraminiferal
records need to be generated from the northeastern margin ofthe western Tethys
The MudurnundashGoumlynuumlk basin in the central Sakarya re-gion northwestern Turkey provides new opportunities forhigh-resolution KndashPg boundary benthic foraminiferal anddinocyst records from the northern margin of the Tethys Re-cently well-preserved outcrops of ancient continental mar-gin deposits spanning the KndashPg boundary have been discov-ered in this basin (Accedilikalin et al 2015) These outcrops in-clude amongst others the Okccedilular section Here an inte-gration of dinocyst and benthic foraminifera records of thisbiostratigraphically well-constrained KndashPg boundary transi-tion is used to provide new insights into changes in and therelationship between planktic and benthic communities
Linking these records to both the previously gener-ated bulk-carbonate carbon isotope record of this section(Accedilikalin et al 2015) and to other benthic foraminiferal andpalynological records in the Tethys enables a supra-regionalportrayal of the KndashPg boundary pelagic crisis and the bio-logical changes caused by the collapse of export productivityThis integrated approach enables an evaluation of the paleoe-cological and paleoceanographic consequences of the earlyDanian Living Ocean condition
2 Geological setting and age assessment
The Okccedilular section is located in the MudurnundashGoumlynuumlkbasin (northwestern Turkey Fig 1) In this basin the KndashPg boundary interval is represented by the Tarakli Forma-tion (Saner 1980 Altiner et al 1991 Accedilikalin et al 2015)In the eastern part of the basin the upper Maastrichtianis characterized by an intercalation of mudstones and tur-bidites whereas in the western part of the basin the tur-bidites are absent The KndashPg boundary is marked by a red-dish ejecta layer at the base of a 15ndash20 cm thick boundary
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
888 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
clay layer Throughout the basin the lower 30ndash50 m of theDanian is characterized by a rhythmic alternation of fine-grained limestones and carbonate-rich mudstones (Accedilikalinet al 2015) During the latest Cretaceous to early Paleocenethis basin was characterized by mixed siliciclasticndashcarbonatesedimentation in an outer neritic to upper bathyal environ-ment (Accedilikalin et al 2015)
The Okccedilular section has been analyzed for siderophiletrace elements including Ir and other platinum groupelements (PGEs) bulk stable carbon isotopes plankticforaminifera calcareous nannofossils and dinocysts Basedon these results a detailed stratigraphic framework was out-lined (Accedilikalin et al 2015) allowing a confident age assess-ment of the boundary interval The age model shows thatthe section contains a chronostratigraphically complete KndashPg boundary interval The studied interval ranges from thetop part of the Maastrichtian A mayaroensis Zone up to thebasal part of the Danian planktic foraminiferal Zone P1b andcovers globally occurring first occurrences (FO) of dinocystmarker taxa such as Senoniasphaera inornata Damassa-dinium californicum and Carpatela cornuta
3 Materials and methods
31 Sampling
High-resolution (centimeter-scale) sample sets were ac-quired during two field campaigns in 2010 and in 2011 Formore detail on these sampling campaigns see Accedilikalin etal (2015) The samples were split for micropaleontologicaland palynological analyses
32 Foraminiferal analysis
Twenty-seven samples were processed at KU Leuven forforaminiferal studies following standard micropaleontolog-ical procedures Of these 20 samples were used for quan-titative benthic foraminiferal analyses (Fig 2) Rock sam-ples were dried in an oven at 60 C for at least 24 h De-pending on sample size 4 to 60 g of dry rock was soakedin a soda (Na2CO3) solution (50 g Lminus1) After disintegrationsamples were washed over 2 mm and 63 microm sieves If neces-sary the tenside Rewoquat was used to clean the residues andthe procedure was repeated Clean residues were dry-sievedinto three fractions 63ndash125 125ndash630 and gt 630 microm In orderto enable a direct comparison with the benthic foraminiferalrecord of the KndashPg boundary GSSP (Global Boundary Stra-totype Section and Point) of El Kef (Tunisia) we used thesame size fraction (gt 125 microm) for Okccedilular as for El Kef(Speijer and van der Zwaan 1996) Note that El Kef holdsthe GSSP of the base of the Danian (Molina et al 2006)and is thus of particular importance with respect to corre-lation and comparison of the KndashPg boundary Representativealiquots of the gt 125 microm fraction were obtained containing atleast 300 benthic foraminiferal specimens Picked specimens
from this size fraction were permanently stored in Plummerslides Benthic foraminifera were identified using the tax-onomy of Cushman (1946 1951) Kellough (1965) Aubertand Berggren (1976) Berggren and Aubert (1975) and Spei-jer (1994)
Benthic foraminifera are commonly used as indicators forbottom water oxygenation and trophic conditions (eg Joris-sen et al 2007) Here the benthic foraminiferal accumula-tion rate (BFAR number of foraminifera per cm2 per kyrsee Text S2 in the Supplement for details on the estimationof BFAR) was calculated as a semi-quantitative proxy for pa-leoproductivity (Jorissen et al 2007 and references therein)In addition the percentage of endobenthic morphotypes wascalculated using the assumed microhabitat preferences in-ferred from benthic foraminiferal morphotype analysis (egCorliss 1985 Corliss and Chen 1988 Alegret et al 2003Woelders and Speijer 2015) In general endobenthic formsare considered indicative of a high flux of organic matter tothe sea floor andor relatively low oxygen conditions whileabundance of epibenthic forms is considered characteristicfor more oligotrophic open marine environments (Jorissen etal 1995 Peryt 2004 Jorissen et al 2007) Furthermore thebi- and triserial endobenthic forms are particularly indica-tive of high food supply and low oxygenation (eg Bern-hard 1986 Corliss and Chen 1988 Jorissen et al 2007)Therefore in this study the percentage of endobenthic formswas calculated for each sample as well as the percentage bi-triserial benthic taxa to unravel food supply and oxygena-tion patterns
It should however be noted that assuming such an ana-logue with modern fauna has limitations and shortcomingsFor instance the percentage of endobenthics based on mor-photypes and using bi-triserials as an indicator for hypoxiaand high food supply is not universally applicable (Buzas etal 1993 Jorissen et al 2007 and references therein)
33 Palynological analysis
In this study the palynological data from Accedilikalin etal (2015) were analyzed and interpreted In addition twoadditional samples were analyzed (OK 15 and OK 6) toincrease the resolution of the dataset and two samples ofthe Accedilikalin et al (2015) dataset were re-counted (OK 25and OK 250 see Dataset 2 in the Supplement) Quantitativeslides of the 15ndash250 microm fraction were used All slides arestored in the collection of the Laboratory of Palaeobotanyand Palynology Utrecht University the Netherlands
To identify major changes in the dinocyst record morpho-logically closely related taxa were grouped into complexesusing a similar approach to Schioslashler et al (1997) Sluijsand Brinkhuis (2009) and Machalski et al (2016) In ourstudy the following morphological complexes were estab-lished (1) the Spiniferites complex combining all species ofSpiniferites and the morphologically similar genus Achomop-shaera (2) Manumiella spp grouping all species of Man-
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
J Vellekoop et al Ecological response to KndashPg boundary mass extinction 889
Figure 2 The benthic foraminiferal record (gt 125 microm) of the Okccedilular section The biostratigraphy is from Accedilikalin et al (2015) The clusteranalysis on the benthic foraminiferal data using paired group (UPGMA) correlation distance allows the identification of four main clustersAndashD of benthic foraminiferal taxa
umiella (3) hexaperidinioids lumping all other peridinioidcysts with a hexaform archeopyle and (4) other dinocystswhich include all other dinocyst taxa and unidentifiabledinocysts (Fig 3)
Of these different dinocyst groups previous studies haveshown that in the Tethys the hexaperidinioids in particularshow strong variations across the KndashPg boundary (Brinkhuiset al 1998 Vellekoop et al 2015) Based on statistical cor-relations between palynological records and other paleoprox-ies it has been suggested that this inferred heterotrophicgroup flourished best under high-nutrient conditions in thephotic zone (Eshet et al 1994 Brinkhuis et al 1998 Sluijsand Brinkhuis 2009) Therefore in this study abundances ofhexaperidinioids are considered indicative of nutrient avail-ability in the photic zone (see Fig S1 and Text S1 in theSupplement for a more detailed discussion on this matter)
34 Statistical analysis
To assess changes in diversity of benthic foraminifera anddinocysts across the studied interval the Shannon diversityindex (H ) the species richness per sample (S) the number ofspecimens observed per sample (N ) and the BergerndashParkerindex were calculated for both biological groups followingHayek and Buzas (2013) (Fig 4) In addition in order torecognize the main faunal associations within the benthicforaminiferal data a cluster analysis was performed usingpaired group (UPGMA) correlation distance
Q-mode non-metric multidimensional scaling (NM-MDS)was performed on the benthic foraminiferal sample compo-sitions to assess patterns in assemblage response to KndashPgboundary perturbations Since taxa may not have a linear re-sponse to environmental changes across the KndashPg boundaryQ-mode NM-MDS is preferred over PCA and CCA (Ram-ette 2007)
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
890 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
Figure 3 The organic-walled dinoflagellate cyst record of theOkccedilular section The biostratigraphy and bulk carbonate stable car-bon isotope records are from Accedilikalin et al (2015) The four maindinocyst complexes are indicated
4 Results
41 Benthic foraminifera
The benthic foraminiferal record of the Okccedilular section ischaracterized by a distinct turnover across the KndashPg bound-ary (Figs 2 4 5 6) Of the common taxa 8 out of 30(sim 27 ) disappear across the KndashPg boundary After the KndashPg boundary crisis three new taxa appear The estimatedBFAR shows a major decrease across the boundary (Fig 5)For the foraminiferal counts and illustrations of commonforms see Dataset S1 and Figs S2 S3
The cluster analysis allows the identification of fourmain clusters of benthic foraminiferal taxa clusters A toD (Fig 2) Cluster C is relatively large and can be subdi-vided into five subclusters Based on the succession of ben-thic faunal assemblages characterized by strong changes inthe Shannon diversity index (H ) four intervals can be rec-ognized in the benthic foraminiferal record of the Okccedilularsection (Figs 4 6)
The first interval comprises the uppermost Maastrichtianand is characterized by relatively high diversity dominatedby the taxa in cluster D In this assemblage the bi-triserialbenthic taxa are relatively abundant (22ndash28 ) with char-acteristic taxa such as Bulimina arkadelphiana Eouvigerinasubsculptura and Praebulimina reussi About 50ndash60 of theassemblage consists of inferred endobenthic taxa The KndashPg boundary marks an abrupt benthic community impover-
ishment and the decimation of the taxa in cluster D Abovethe boundary the bi-triserial benthic taxa virtually disappearfrom the record (mostly below lt 05 )
The second interval characterized by low diversity com-prises the lowermost Danian approximately correspondingto planktic foraminiferal Zone P0 In this interval epibenthicforms are most abundant (70ndash90 ) It is dominated by thetaxa of cluster A encompassing successive peak occurrencesof the taxa Anomalinoides praeacutus Trochammina sppand Cibicidoides pseudoacutus
The third interval covers the part of the succession approx-imately correlative to planktic foraminiferal Zone Pα Thisinterval is characterized by a recovery of diversity althoughthe diversity is still lower than that of the top Maastrichtianassemblage The abundance of taxa in cluster A slowly de-creases and taxa in cluster B mainly represented by Osan-gularia plummerae and Cibicidoides sp become abundantEndobenthic forms recover and make up 30ndash40 of the totalassemblage
The fourth interval starts in the interval correlative to ZoneP1a Here the benthic community has recovered as the di-versity has stabilized and is almost similar to pre-impact val-ues This interval is characterized by taxa in cluster C a typ-ical Paleocene Midway-type fauna (Berggren and Aubert1975) with representatives such as Anomalinoides praea-cutus Coryphostoma midwayensis Cibicidoides alleni andOsangularia plummerae
42 Palynology
Palynological samples from the Okccedilular site yield abundantpalynomorphs dominated by dinocysts and with minor con-tributions of acritarchs prasinophytes organic foraminiferallinings and terrestrial palynomorphs (Dataset S2) Thedinocyst associations of the MudurnundashGoumlynuumlk basin arequite diverse including components characteristic of boththe Tethyan and Boreal realms (Accedilikalin et al 2015) As ex-pected the dinocyst record does not show major changes indiversity across the KndashPg boundary (Fig 4) There is a steadydecrease in dinocyst diversity from planktic foraminiferalZone P1a upwards probably a long-term change not re-lated to the impact The estimated dinocyst accumulationrate (DAR number of preserved cysts per cm2 per kyr seeText S2 in the Supplement for details on the estimation of theDAR) shows no major changes across the KndashPg boundary
Throughout the palynological record the Spiniferites com-plex is consistently the most dominant morphogroup in gen-eral comprising 40ndash50 of the total assemblage (Fig 3)Similar to other KndashPg boundary sites worldwide (Habib andSaeedi 2007) Manumiella spp show an episode of higherrelative abundance near the top of the Maastrichtian some30ndash40 cm below the KndashPg boundary Hexaperidinioids gen-erally make up a relatively small component of the Maas-trichtian assemblage (3ndash17 ) but show a strong increaseacross the KndashPg boundary In the boundary clay layer cor-
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
J Vellekoop et al Ecological response to KndashPg boundary mass extinction 891
Figure 4 Diversity indices on the benthic foraminiferal and dinocyst records of the Okccedilular section with the Shannon diversity index (H ) thespecies richness per sample (S) the number of specimen observed per sample (N ) and the BergerndashParker index As the bulk stable carbonisotope record and the Shannon diversity index of the benthic foraminiferal record show similar trends the combination of these records canbe used to subdivide the KndashPg boundary transition into four intervals I the Maastrichtian ie ldquopre-impactrdquo interval II the post-impactinterval representing the ldquodisasterrdquo phase III a recovery interval and IV the early Paleocene post-recovery interval The division criteriaare explained in the text
relative to planktic foraminiferal Zone P0 this group in-creases to 35 of the assemblage In the peak intervals thehexaperidinioids are mostly represented by representativesof the genera Senegalinium and Cerodinium After an ini-tial drop in relative abundance in the upper half of Zone P0this group reaches a second maximum (sim 35 ) at the baseof Zone Pα and remains relatively abundant (15ndash30 ) up toZone P1a After this the hexaperidinioids gradually decreasetowards the top of the studied interval The ldquoother dinocystsrdquogroup ndash with representatives such as Areoligera spp Im-pagidinium spp Hystrichosphaeridium tubiferum Opercu-lodinium centrocarpum and Palynodinium grallator ndash gen-erally makes up 25ndash50 of the assemblage
5 Discussion
51 Tethyan benthic foraminiferal turnover sequence
The major turnover in the benthic community at Okccedilularis largely comparable with the previously published ben-thic foraminiferal record (size fraction gt 125 microm) of the KndashPg boundary GSSP of El Kef (Figs 7 8 Speijer and Vander Zwaan 1996) In addition other benthic foraminiferalrecords from the margins of the Tethys also show a strong
similarity to the Okccedilular and El Kef records including stud-ies that use the gt 63 microm size fraction (Alegret et al 2003Culver 2003 and references therein)
Although it is sometimes suggested that different sizefractions cannot be directly compared because the differ-ent size fractions sometimes harbor different ecological in-formation (Giusberti et al 2016) intercomparison studieshave shown that when comparing the gt 125 and gt 63 microm sizefractions in benthic foraminiferal assemblages the overallpatterns are generally robust (Ernst et al 2006) The factthat gt 63 and gt 125 microm KndashPg boundary benthic foraminiferalrecords from different Tethys-margin sites (eg Speijer andVan der Zwaan 1996 Peryt et al 2002 Culver et al 2003)show striking similarities even though both the size fractionsused (gt 63 vs gt 125 microm) and specific taxa making up theforaminiferal assemblages differ among sites confirms theobservations by Ernst et al (2006) Furthermore even whenit would be assumed that different size fractions are diffi-cult to compare directly the striking similarities between theTethyan gt 125 and gt 63 microm size fraction KndashPg boundary ben-thic foraminiferal records provide compelling evidence for astrong disruption of the entire benthic community
Tethyan middle neritic to middle bathyal KndashPg boundarysuccessions generally show a sequence of (1) a typical high
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
892 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
Figure 5 Estimated benthic foraminiferal accumulation rate (BFAR number of foraminifera of the gt 63 microm size fraction per cm2 sea floorper kyr) and estimated dinocyst accumulation rate (DAR number of preserved cysts per cm2 sea floor per kyr) of the Okccedilular sectionUncertainty in estimated accumulation rates resulting from uncertainties in sediment densities and sedimentation rates are indicated in greyproviding a range of estimated accumulation rates For more detailed info on the calculation of the BFAR and DAR see Text S2 of theSupplement
diversity assemblage in the Maastrichtian (2) a low diver-sity ldquodisasterrdquo assemblage directly after the KndashPg boundaryimpact characterized by a short-lived bloom of endobenthicforms observed in several high-resolution records (eg Coc-cioni and Galeotti 1994 Speijer and van der Zwaan 1996)followed by a dominance of epibenthic forms (3) a ldquore-coveryrdquo assemblage characterized by an increasing diversityand returning endobenthic forms and (4) a new high diver-sity assemblage dominated by a Paleocene fauna with bothepi- and endobenthic forms equally represented
Hence based on this succession of benthic assemblageswhich are characterized by strong changes in diversity (H )the KndashPg boundary benthic foraminiferal records from theTethys can be subdivided into four intervals (Figs 2 4ndash6)The intervals I to IV approximately correspond to the up-permost Maastrichtian planktic foraminiferal Zone P0 sub-zones PαndashP1a and subzones P1andashP1b respectively There-fore this succession roughly follows the 5-fold sedimen-tological and planktic foraminiferal sequence of Smit andRomein (1985) in which unit 2 represents the ejecta layerand units 1 3 4 and 5 correspond to intervals I to IV in ourstudy
The sequence of benthic foraminiferal turnover in theTethys is also well illustrated by a Q-mode NM-MDS
(Fig 7) The Q-mode NM-MDS of the benthic foraminiferalsample compositions of the combined data from Okccedilular(this study) and El Kef (Speijer and van der Zwaan 1996)demonstrates that although the assemblages differ betweenthese localities on opposite margins of the Tethys the re-sponses of the benthic foraminiferal assemblages to the KndashPgboundary perturbations express very similar overall patternsacross the KndashPg boundary At both Okccedilular and El Kef thebenthic foraminiferal records show a rapid transition from astable Maastrichtian assemblage to the earliest Danian dis-aster phase in Zone P0 followed by a gradual change backtowards conditions similar to the Maastrichtian (marked bythe return of Lazarus taxa) Therefore both the successionof benthic faunal assemblages including diversity indicesand Q-mode NM-MDS analysis provide powerful tools to al-low the comparison of KndashPg boundary benthic foraminiferalrecords from different localities
52 Ecological responses to reduced export productivity
521 Okccedilular section
The palynological benthic foraminiferal and bulk stable iso-tope records of the Okccedilular section show major changesacross the KndashPg boundary interval (Figs 2ndash7) portraying
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
J Vellekoop et al Ecological response to KndashPg boundary mass extinction 893
Figure 6 Relative abundances of the four main clusters of benthic foraminiferal taxa (gt 125 microm) relative abundances of epibenthic andendobenthic forms and relative abundance of bi-triserial benthic taxa
Figure 7 Q-mode non-metric multidimensional scaling (on all sam-ples in the gt 125 microm size fraction benthic foraminiferal datasets ofOkccedilular (this study) and El Kef (Speijer and van der Zwaan 1996)After an abrupt transition from latest Maastrichtian pre-impact as-semblages to disaster assemblages in Zone P0 and recovery assem-blages in Zone Pα a gradual transition towards a new equilibriumpost-recovery assemblage can be observed in subzones P1a and P1b
the biological crisis following the impact Following the KndashPg boundary mass extinction endobenthic forms stronglydecrease with bi-triserial benthic taxa almost disappearingfrom the benthic community simultaneously the estimatedBFAR shows a major fall (Fig 5) This interval approxi-mately correlative to foraminiferal Zone P0 represents thedisaster phase The drop in bi-triserial benthic taxa (Fig 6)and decrease in BFAR (Fig 5) point to a reduction in foodsupply to the benthic community indicating that the trans-port of organic matter to the sea floor must have decreasedIn the dinocyst community however hexaperidinioids showa strong increase in relative abundance during this phase(Figs 3 6 7) suggesting an increase in nutrient availabilityin the photic zone while the DAR remains relatively stableacross the KndashPg boundary (Fig 5)
The decrease in benthic diversity and the high relativeabundances of opportunistic Trochammina spp and Cibici-doides pseudoacutus suggest that the benthic community ex-perienced additional stress during Zone P0 besides food lim-itation (see Jorissen et al 2007) If food limitation were theonly factor involved upper bathyal assemblages as observedat Okccedilular would be expected to become more similar tooligotrophic middle to lower bathyal assemblages Insteadthe opposite is true the assemblage composition of the dis-aster phase is closer to neritic Midway-type assemblages
It was previously suggested that the epibenthic taxon Cibi-cidoides pseudoacutus could potentially be hypoxia resistant
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
894 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
(Speijer and van der Zwaan 1996) As a collapse of the bi-ological pump should lead to enhanced remineralization oforganic matter in the photic zone it is indeed likely thatthe oxygen minimum zones expanded and shoaled leadingto oxygen stress on the sea floor Although early Paleocenedysoxia at El Kef is supported by the ostracod record (Donzeet al 1982) and ichnofacies analyses (Rodriguez-Tovaret al 2016) supporting (eg geochemical) evidence fordysoxia during the disaster phase at Okccedilular record is absentFurthermore an additional stressor for the benthic commu-nity during Zone P0 may be present In contrast to calcare-ous nanoplankton organic-walled cyst-producing dinoflagel-lates did not suffer extinctions and may have become a moreimportant component in the early Paleocene phytoplanktoncommunity Hence besides the reduction of the amount oforganic matter transported to the sea floor the compositionof food supplied by the photic zone likely changed signifi-cantly across the KndashPg boundary as well (DrsquoHondt 2005)possibly presenting an additional stress factor for the benthiccommunity (Speijer and van der Zwaan 1996 Alegret andThomas 2009)
522 Western Tethys
The patterns in benthic foraminiferal and dinoflagellate re-sponse to the KndashPg boundary perturbations at Okccedilular appearto be characteristic for the continental margins of the westernTethyan realm The benthic foraminiferal patterns are similaracross different size fractions suggesting that they are repre-sentative of the overall benthic communities In Tunisia andSpain where both benthic foraminiferal and dinoflagellaterecords are available the earliest Danian is also characterizedby a decrease in endobenthic foraminifera at the sea floor(Speijer and Van der Zwaan 1996 Peryt et al 2002 Ale-gret et al 2003) and simultaneously blooms of hexaperidin-ioids in the water column (Brinkhuis et al 1998 Vellekoopet al 2015 Fig 8) In the Ouled Haddou section Moroccoin the westernmost Tethys (Fig 1) the lowermost Danian isalso characterized by a strong increase of the hexaperidin-ioid Senegalinium group up to 30 of the assemblage (Sli-mani et al 2010) that is very similar to the Okccedilular record(Fig 8)
The KndashPg boundary dinocyst record of El Kef is strikinglysimilar to the one from the Okccedilular section showing two dis-tinct peaks in hexaperidinioids one in Zone P0 and one atthe basal part of Zone Pα At El Kef the initial post-impactdominance of hexaperidinioids is however less pronounced(Fig 8) This difference in expression might be related tosmall differences in paleogeographic and paleoceanographicsettings between sites
In addition some high-resolution benthic KndashPg records inthe Tethyan realm show a short-lived bloom of specific en-dobenthic foraminiferal taxa in the few centimeter directlyabove the KndashPg boundary sometimes accompanied by otherindicators of low oxygen levels (eg Coccioni and Gale-
otti 1994 Speijer and Van der Zwaan 1996 Kaiho et al1999) This suggests that at least locally the sea floor wastemporarily characterized by hypoxic conditions directly fol-lowing the impact This short-lived deoxygenation is possi-bly related to the mass mortality at the KndashPg boundary re-sulting in a large short-lived flux of organic matter to thesea floor (Coccioni and Galeotti 1994) However since ex-port productivity was greatly reduced after the KndashPg bound-ary mass extinction very little ldquonewrdquo organic matter reachedthe sea floor after the initial post-impact influx Hence afteraccumulated organic matter was remineralized the benthiccommunity starved resulting in a transition to an epibenthic-dominated benthic fauna similar to the post-impact fauna ofthe Okccedilular record
The combined dinocyst records suggest that the early Pa-leocene margins of the Tethys were characterized by an in-crease in nutrient availability in the photic zone whereascoeval benthic foraminiferal records indicate a major de-crease in nutrient supply to the sea floor These changesin the benthic foraminiferal and dinocyst communities arelikely caused by the major reduction of both the efficiencyand strength of the biological pump (ie the fraction ofprimary production exported from the photic zone and theamount of organic matter transported down) in accordancewith the Living Ocean model (DrsquoHondt and Zachos 1998DrsquoHondt et al 1998 DrsquoHondt 2005 Coxall et al 2006)For a detailed explanation of the difference between biologi-cal pump efficiency and biological pump strength see Hiltinget al (2008)
As bulk carbonate δ13C reflects the isotopic compositionof the surface ocean which is set by burial fractions and bythe photosynthetic isotope effect (Kump 1991 Hain et al2014) the excursion recorded in bulk δ13C records world-wide and the rapid collapse in surface to deep-ocean carbonisotope gradients likely reflect the reduction of the global in-tensity (ie efficiency) of the biological pump (Hain et al2014) The recorded inverse change in nutrient availabilitysuggests a causal link A reduced efficiency of the biologi-cal pump and associated decrease of the fraction of biomasstransported out of the photic zone could have resulted inhigh rates of nutrient recycling in the upper part of the wa-ter column (DrsquoHondt 2005) This suggests that the reduc-tion of the biological pump strength recorded by the benthicforaminiferal record is a consequence of the decreased effi-ciency of the biological pump recorded by the carbon iso-tope and dinoflagellate cyst records The strong correlationbetween the bulk carbon isotope curves and the Shannondiversity index (H ) of the benthic foraminiferal records atboth Okccedilular (R2
= 073 p lt 0001) and El Kef (R2= 074
p lt 0001) suggest a clear link between the changes in ben-thic foraminiferal assemblages and the collapse and recoveryof biological pump efficiency
As a result of the reduced biological pump efficiency morenutrients remained available for the early Paleocene phyto-plankton community At Tethyan margin sites this is indi-
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
J Vellekoop et al Ecological response to KndashPg boundary mass extinction 895
Figure 8 The benthic foraminiferal (gt 125 microm) dinocyst and bulk stable carbon isotope records of Okccedilular (Turkey) and El Kef (Tunisia)the dinocyst record of Ouled Haddou (Morocco) and the benthic foraminiferal record (gt 63 microm) of ODP Hole 1049C Blake Nose (NorthAtlantic) Note that the definition of ldquohexaperidinioidsrdquo used is provided in the text The four phases identified based on the bulk stablecarbon isotopes and the Shannon diversity index (H ) of the benthic foraminiferal records are indicated (IndashIV) The gt 63 microm size fractionbenthic foraminiferal record of ODP Hole 1049C is from Alegret and Thomas (2004) who argued that the lowermost Paleocene of thissite comprised reworked foraminifera This interval is indicated by a dashed bar in the figure The biostratigraphy of Ouled Haddou is fromSlimani and Toufiq (2013) whereas the dinocyst record of this site is from Slimani et al (2010) The biostratigraphy of El Kef is based onBrinkhuis et al (1998) Speijer and van der Zwaan (1996) and Molina et al (2006) The fine-fraction stable carbon isotope record (blackdots) of El Kef is from Keller and Lindinger (1989) The Shannon diversity index (red dots) relative abundances of bi-triserial benthic taxa(dark purple) and all endobenthic foraminifera (light purple) are based on the gt 125 microm size fraction foraminiferal data of Speijer and vander Zwaan (1996) The palynological data of El Kef are from Brinkhuis et al (1998) (lower part) and Guasti et al (2005) (upper part) TheShannon diversity index of the gt 125 microm size fraction benthic foraminiferal record (red dots) the relative contribution of bi-triserial benthictaxa (dark purple) all endobenthic foraminifera (light purple) and hexaperidinioid dinocysts from Okccedilular are from this study The bulkstable carbon isotope record of Okccedilular (black dots) is from Accedilikalin et al (2015) Mbsf indicates meters below sea floor
cated by the higher abundance of hexaperidinioids Similarlyblooms of eutrophic survivor taxa of calcareous nanoplank-ton at open ocean sites have been suggested to be relatedto the buildup of nutrients in the open ocean photic zone(Schueth et al 2015) Although the carbon isotope gradientsbetween surface and deep waters indicate that the recoveryof the biological pump took hundreds of thousands of years(Zachos et al 1989 Kump 1991 DrsquoHondt et al 1998 Cox-all et al 2006 Birch et al 2016) our records suggest thatthe enhanced recycling of nutrients in the Tethys was partic-ularly intense during two phases in the first tens of thousandsyears after the impact (Fig 8)
523 Global responses
The decrease in export productivity following the KndashPgboundary is not only recognized in the Tethyan realm In theAtlantic Ocean Southern Ocean and the Indian Ocean a de-crease of the biological pump strength after the KndashPg bound-ary was also observed in neritic to abyssal environments(eg Thomas 1990 Olsson et al 1996 Hull et al 2011Alegret et al 2012) As an example the gt 63 microm size frac-tion benthic foraminiferal KndashPg boundary record of BlakeNose (ODP Hole 1049C northwestern Atlantic Alegret andThomas 2004) shows a pattern that is strikingly similar tothe gt 125 microm size fraction records in the Tethyan realm witha strong decrease of endobenthics in Zone P0 followed by a
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
896 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
gradual recovery across zones Pα and P1a (Fig 8) The simi-larities between the benthic foraminiferal patterns of Tethyanouter neritic to middle bathyal (200ndash1500 m) sites and thoseof the middle to lower bathyal (1500ndash1600 m) record ofBlake Nose (Alegret and Thomas 2004) suggest that theserecords register a global decrease in export productivity fol-lowing the KndashPg boundary
However several open ocean sites mostly in the Pacificshow an opposite trend with several lines of evidence sug-gesting an increased biological pump strength at these sites(eg Hull et al 2011 Alegret et al 2012) This led Ale-gret et al (2012) to conclude that the decrease in biologicalpump strength after the KndashPg boundary was most likely aregional instead of a global effect arguing against the Liv-ing Ocean hypothesis We would argue that low-productivityopen ocean sites like the central Pacific are characterized byvery different ecosystem structures and higher productivityas those along the Tethyan margins (eg Dortch and Pack-erd 1989) Therefore the consequence of ecosystem reorga-nization resulting from the mass extinction was likely verydifferent in the open ocean Pacific (eg Sibert et al 2014)Esmeray-Senlet et al (2015) proposed the term ldquoHeteroge-neous Oceanrdquo for this conceptual model as an alternativeto the Living Ocean model Supposedly the HeterogeneousOcean was characterized by a strong geographic heterogene-ity in the extinction patterns and food supply to the sea floor
However as the primary food source for open ocean deep-sea benthic communities generally consists of refractory or-ganic matter (eg Jiao et al 2010) it remains uncertainwhether changes in the intensity of recycling of the gener-ally more labile organic matter in the surface ocean will re-sult in changes in the amount of organic matter that benthiccommunities receive as they will likely continue to receiverecalcitrant organic matter at barely diminished rates despitethe collapse of the biological pump
In addition it should be noted that although the bio-logical pump efficiency is generally relatively high in openocean realms such as the central Pacific the biological pumpstrength is generally very low at these sites (Honjo et al2008 Henson et al 2011) Small changes in absolute bio-logical pump strength at such a locality could therefore havehad a large effect on the local oligotrophic benthic commu-nity (Alegret and Thomas 2009) but they likely representedonly a small fraction of the net amount of organic matterglobally transported to the deep Hence even though localbiological pump strength might have increased at some low-productivity deep-sea sites the large decrease in biologi-cal pump strength recorded at many other sites means thatthe net amount of organic matter globally transported out ofthe surface ocean was likely still reduced in the post-impactworld (Birch et al 2016) Therefore we argue that such Pa-cific records do not represent the general pattern The globalocean response can be characterized as heterogeneous how-ever the KndashPg boundary mass extinction did lead to a strongreduction of the net amount of organic matter globally trans-
ported from the surface to the sea floor while biological pro-ductivity recovered rapidly after the extinction event in ac-cordance with the Living Ocean hypothesis
53 Long-term recovery
Following the disaster phase the hexaperidinioid cysts atOkccedilular and El Kef decreased in abundance and the abun-dance of endobenthic forms increased again as the recoveryof the benthic community was initiated This recovery phaseis approximately correlative to Zone Pα and the basal part ofZone P1a which according to the Paleogene age constraintsof Vandenberghe et al (2012) represents at least 300 kyrThis duration is in agreement with the estimations providedby Birch et al (2016) During this phase the carbon iso-tope records remain well below pre-impact values and thediversity of the benthic foraminiferal community has not yetfully recovered (Figs 4ndash8) This indicates that the rapid andshort-lived KndashPg boundary disaster was followed by a rela-tively long recovery phase in line with previous estimatesof a multimillion-year biological recovery (eg Coxall etal 2006) While the impact-related environmental perturba-tions were short lived (Kring 2007 Vellekoop et al 20142016) the extinctions amongst important biological groupsled to a reduction of the organic flux from the photic zone todeep water resulting in major long-term biological and pa-leoceanographic reorganizations Only with the evolutionaryrecovery of the pelagic community governing the biologicalcarbon pump did export productivity start to increase again(eg Coxall et al 2006 Birch et al 2016)
6 Conclusions
The marine palynological benthic foraminiferal and bulkstable isotope records of the Okccedilular and El Kef sections re-veal major changes across the KndashPg boundary interval por-traying the biological crisis at the KndashPg boundary and subse-quent recovery in the early Paleocene Based on the succes-sion of benthic faunal assemblages at a number of Tethyanmargin sites four phases can be recognized across the KndashPgboundary interval the Maastrichtian or pre-impact phase (I)a disaster phase (II) a recovery phase (III) and an early Pa-leocene post-recovery phase (IV)
Following the KndashPg boundary impact some localitiesshow a large short-lived flux of food to the sea floor likelyrelated to the mass mortality at the KndashPg boundary Howeversince export productivity was greatly reduced after the KndashPgboundary mass extinction it is likely that after the initialpost-impact influx very little fresh organic matter reachedthe sea floor This reduction of export productivity in thepost-extinction disaster phase eventually resulted in a lowerfood supply to the sea floor This presented a major stressfactor for benthic organisms in the western Tethys The re-duced food supply resulted in an abrupt impoverishment of
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
J Vellekoop et al Ecological response to KndashPg boundary mass extinction 897
benthic communities As the downward transport of nutri-ents was slowed down recycling in upper layers increasedAs a result more nutrients remained available for the earlyPaleocene phytoplankton community leading to blooms ofdinoflagellates along the Tethyan margins and blooms of cal-careous nanoplankton taxa in more open ocean sites Ourrecords show that the enhanced recycling of nutrients in theTethys was particularly intense in the tens of thousands ofyears after the impact Following this the slow evolution-ary recovery of the pelagic community governing the bio-logical carbon pump resulted in a gradually increasing ex-port productivity in the hundreds of thousands of years afterthe impact Hence our integration of dinocyst and benthicforaminiferal records across the KndashPg boundary provides im-portant new insights into the ecological responses to the re-duction of export productivity following the mass extinctionat the CretaceousndashPaleogene boundary highlighting the di-rect ecological consequences of the Living Ocean conditionsin the post-impact world
7 Data availability
All data related to publication are available in the Supple-ment
The Supplement related to this article is available onlineat doi105194bg-14-885-2017-supplement
Author contributions Johan Vellekoop Lineke Woelders andHenk Brinkhuis designed the research Samples were collectedin the field by Johan Vellekoop Sanem Accedilikalin Ismail Ouml Yil-maz and Jan Smit Palynological analyses were carried out by Jo-han Vellekoop Benthic foraminiferal analyses were carried out byLineke Woelders and Robert P Speijer Johan Vellekoop LinekeWoelders and Bas van de Schootbrugge prepared the manuscriptwith input from all authors
Competing interests The authors declare that they have no conflictof interest
Acknowledgements This work was supported by the NetherlandsOrganization for Scientific Research (NWO) Open CompetitionGrant ALWPJ09047 to H Brinkhuis and the Research FoundationFlanders (FWO) grant G0B8513 to R P Speijer We thank LKump F Boscolo Galazzo P Roopnarine and one anonymousreferee for their suggestions
Edited by D GillikinReviewed by L Kump F Boscolo Galazzo and P Roopnarine
References
Accedilikalin S Vellekoop J Ocakoglu F Yılmaz I Ouml Smit J Al-tiner S O Goderis S Vonhof H Speijer R P WoeldersL Fornaciari E and Brinkhuis H Geochemical and pale-ontological characterization of a new KndashPg Boundary local-ity from the Northern branch of the Neo-Tethys MudurnundashGoumlynuumlk Basin NW Turkey Cretaceous Res 52 251ndash267doi101016jcretres201407011 2015
Alegret L and Thomas E Food supply to the seafloor in the Pa-cific Ocean after the CretaceousPaleogene boundary event MarMicropaleontol 73 105ndash116 2009
Alegret L Molina E and Thomas E Benthic foraminiferalturnover across the CretaceousPaleogene boundary at Agost(southeastern Spain) paleoenvironmental inferences Mar Mi-cropaleontol 48 251ndash279 2003
Alegret L Thomas E and Lohmann K C End-Cretaceous ma-rine mass extinction not caused by productivity collapse P NatlAcad Sci USA 109 728ndash732 doi101073pnas11106011092012
Alegret L Rodriacuteguez-Tovar F J and Uchman A How bioturba-tion obscured the CretaceousndashPaleogene boundary record TerraNova 27 225ndash230 doi101111ter12151 2015
Altiner D Koccedilyigit A Farinacci A Nicosia U and ContiM A Jurassic-Lower Cretaceous stratigraphy and paleogeo-graphic evolution of the southern part of north-western Anatolia(Turkey) Geologica Romana 27 13ndash80 1991
Alvarez LW Alvarez W Asaro F and Michel H V Extrater-restrial cause for the Cretaceous-Tertiary extinction Science208 1095ndash1108 1980
Alegret L and Thomas E Benthic foraminifera and environmen-tal turnover across the CretaceousPaleogene boundary at BlakeNose (ODP Hole 1049C Northwestern Atlantic) PalaeogeogrPalaeocl 208 59ndash83 doi101016jpalaeo200402028 2004
Armstrong R A Lee C Hedges J I Honjo S and WakehamS G A new mechanistic model for organic carbon fluxes in theocean based on the quantitative association of POC with ballastmaterials Deep-Sea Res Pt II 49 219ndash236 2002
Aubert J and Berggren W A Paleocene benthic foraminiferalbiostratigraphy and paleoecology of Tunisia Bull du Cent deRech de Pau ndash SNPA 10 379ndash469 1976
Berggren W A and Aubert J Paleocene benthonic foraminiferalbiostratigraphy paleobiogeography and paleoecology ofAtlanticndashTethyan regions midway-type fauna PalaeogeogrPalaeocl 18 73ndash192 1975
Bernaola G and Monechi S Calcareous nannofossil ex-tinction and survivorship across the CretaceousndashPaleogeneboundary at Walvis Ridge (ODP Hole 1262C SouthAtlantic Ocean) Palaeogeogr Palaeocl 255 132ndash156doi101016jpalaeo200702045 2007
Bernhard J M Characteristic assemblages and morphologies ofbenthic foraminifera from anoxic organic-rich deposits Jurassicthrough Holocene J Foramin Res 16 207ndash215 1986
Birch H S Coxall H K Pearson P N Kroon D andSchmidt D N Partial collapse of the marine carbon pump af-ter the CretaceousndashPaleogene boundary Geology 44 287ndash290doi101130G375811 2016
Brinkhuis H and Zachariasse W J Dinoflagellate cysts sealevel changes and planktonic foraminifers across the Cretaceous-
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
898 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
Tertiary boundary at El Haria northwest Tunisia Mar Micropa-leontol 13 153ndash191 1988
Brinkhuis H Bujak J P Smit J Versteegh G J M and Viss-cher H Dinoflagellate-based sea surface temperature recon-structions across the Cretaceous-Tertiary boundary PalaeogeogrPalaeocl 141 67ndash83 1998
Buesseler K O The decoupling of production and particulate ex-port in the surface ocean Global Biogeochem Cy 12 297ndash3101998
Buzas M A Culver S J and Jorissen F J A statisti-cal evaluation of the microhabitats of living (stained) infau-nal benthic foraminifera Mar Micropaleontol 20 311ndash320doi1010160377-8398(93)90040-5 1993
Coccioni R and Galeotti S KndashT boundary extinction geolog-ically instantaneous or gradual event Evidence from deep-seabenthic foraminifera Geology 22 779ndash782 1994
Corliss B H Microhabitats of benthic foraminifera within deep-sea sediments Nature 314 435ndash438 doi101038314435a01985
Corliss B H and Chen C Morphotype patterns of Nor-wegian Sea deep-sea benthic foraminifera and ecologi-cal implications Geology 16 716ndash719 doi1011300091-7613(1988)016lt0716MPONSDgt23CO2 1988
Coxall H K DrsquoHondt S and Zachos J C Pelagic evolution andenvironmental recovery after the CretaceousndashPaleogene mass ex-tinction Geology 34 297ndash300 doi101130G217021 2006
Culver S J Benthic foraminifera across the CretaceousndashTertiary(KndashT) boundary a review Mar Micropaleontol 47 177ndash2262003
Cushman J A Upper Cretaceous foraminifera of the Gulf coastalregion of the United States and adjacent areas US Geolog SurvProf Pap 206 241 pp 1946
Cushman J A Paleocene foraminifera of the Gulf Coastal Regionof the United States and adjacent regions US Geol Surv ProfPap 232 75 pp 1951
DrsquoHondt S Consequences of the CretaceousPaleogene Mass Ex-tinction for Marine Ecosystems Annu Rev Ecol Evol S36 295ndash317 doi101146annurevecolsys350211031057152005
DrsquoHondt S and Zachos J C Cretaceous foraminifera and the evo-lutionary history of planktic photosymbiosis Paleobiology 24512ndash523 1998
DrsquoHondt S Donaghay P Zachos J C Luttenberg D andLindinger M Organic Carbon Fluxes and Ecological Recov-ery from the Cretaceous-Tertiary Mass Extinction Science 282276ndash279 1998
Donze P Colin J-P Damotte R Oertli HJ Peypouquet J-Pand Said R Les ostracodes du Campanien terminal agrave lrsquoEocegraveneinfeacuterieur de la coupe du Kef Tuneacutesie nord-occidentale Bull desCentr Recher Expl-Prod Elf Aquit 6 273ndash335 1982
Dortch Q and Packard T T Differences in biomass structure be-tween oligotrophic and eutrophic marine ecosystems Deep-SeaRes 36 223ndash240 1989
Ernst S R Guasti E Dupuis C and Speijer R P En-vironmental perturbation in the southern Tethys across thePaleoceneEocene boundary (Dababiya Egypt) Foraminiferaland clay mineral records Mar Micropaleontol 60 89ndash111doi101016jmarmicro200603002 2006
Eshet Y Moshkovitz S Habib D Benjamini C and Maga-ritz M Calcareous nannofossil and dinoflagellate stratigraphyacross the CretaceousTertiary boundary at Hor Hahar IsraelMar Micropaleontol 18 199ndash228 1992
Eshet Y Almogi-Labin A and Bein A Dinoflagellate cysts pa-leoproductivity and upwelling systems A Late Cretaceous ex-ample from Israel Mar Micropaleontol 23 231ndash240 1994
Esmeray-Senlet S Wright J D Olsson R K MillerK G Browning J V and Quan T M Evidencefor reduced export productivity following the Creta-ceousPaleogene mass extinction Paleoceanography 30718ndash738 doi1010022014PA002724 2015
Giusberti L Boscolo Galazzo F and Thomas E Variability inclimate and productivity during the Paleocene-Eocene ThermalMaximum in the western Tethys (Forada section) Clim Past12 213ndash240 doi105194cp-12-213-2016 2016
Guasti E Kouwenhoven T J Brinkhuis H and Speijer R PPaleocene sea-level and productivity changes at the southernTethyan margin (El Kef Tunisia) Mar Micropaleontol 55 1ndash17 2005
Habib D and Saeedi F The Manumiella seelandica global spikeCooling during regression at the close of the MaastrichtianPalaeogeogr Palaeocl 255 87ndash97 2007
Hain M P Sigmal D M and Haug G H 818 ndash The biolog-ical Pump in the Past in Reference Module in Earth Systemsand Environmental Sciences Treatise on Geochemistry (SecondEdition) The Oceans and Marine Geochemistry 8 485ndash517doi101016B978-0-08-095975-700618-5 2014
Hayek L C and Buzas M A On the proper and efficient useof diversity measures with individual field samples J ForaminRes 43 305ndash313 2013
Henson S A Sanders R Madsen E Morris P J Le MoigneF and Quartly G D A reduced estimate of the strength ofthe oceanrsquos biological carbon pump Geophys Res Lett 38L04606 doi1010292011GL046735 2011
Hilting A K Kump L R and Bralower T J Variations in theoceanic vertical carbon isotope gradient and their implicationsfor the Paleocene-Eocene biological pump Paleoceanography23 PA3222 doi1010292007PA001458 2008
Honjo S Manganini S J Krishfield R A and Francois RParticulate organic carbon fluxes to the ocean interior and fac-tors controlling the biological pump A synthesis of global sed-iment trap progams since 1983 Prog Oceanogr 76 217ndash285doi101016jpocean200711003 2008
Huber B T MacLeod K G and Norris R D Abrupt extinctionand subsequent reworking of Cretaceous planktonic foraminiferaacross the Cretaceous-Tertiary boundary Evidence from the sub-tropical North Atlantic in Catastrophic Events and Mass Extinc-tions Impacts and Beyond edited by Koeberl C and MacLeodK G Geol S Am S 356 277ndash289 Boulder Colorado USA2002
Hull P M Norris R D Bralower T J and Schueth J D A rolefor chance in marine recovery from the end-Cretaceous extinc-tion Nat Geosci 4 856ndash860 doi101038ngeo1302 2011
Hsuuml K J and McKenzie J A A ldquoStrangeloverdquo ocean in the earli-est Tertiary in Natural variations Archean to present edited bySundquist E T and Broecker W S The carbon cycle and atmo-spheric CO2 American Geophysical Union Washington DCUSA 487ndash492 1985
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
J Vellekoop et al Ecological response to KndashPg boundary mass extinction 899
Jiao N Herndl G J Hansell D A Benner R Kattner G Wil-helm S W Kirchman D L Weinbauer M G Luo T ChenF and Azam F Microbial production of recalcitrant dissolvedorganic matter long-term carbon storage in the global oceanNat Rev 8 593ndash599 doi101038nrmicro2386 2010
Jorissen F J de Stigter H C and Widmark J G V A concep-tual model explaining benthic foraminiferal microhabitats MarMicropaleontol 26 3ndash15 doi1010160377-8398(95)00047-X1995
Jorissen F J Fontanier C and Thomas E Paleoceanographi-cal proxies based on deep-sea benthic foraminiferal assemblagecharacteristics in Proxies in Late Cenozoic PaleoceanographyPt 2 Biological tracers and biomarkers edited by Hillaire-Marcel C and de Vernal A 263ndash326 Elsevier Amsterdamthe Netherlands 2007
Kaiho K Kajiwara Y Tazaki K Ueshima M Takeda NKawahata H Arinobu T Ishiwatari R Hirai A and Lam-olda M A Oceanic primary productivity and dissolved oxygenlevels at the CretaceousTertiary boundary their decrease sub-sequent warming and recovery Paleoceanography 14 511ndash5241999
Keller G and Lindinger M Stable isotope TOC and CaCO3record across the CretaceousTertiary boundary at El KefTunisia Palaeogeogr Palaeocl 73 243ndash265 1989
Kellough G R Paleoecology of the Foraminiferida of the WillsPoint Formation (Midway Group) in northeast Texas Trans GulfCoast Assoc Geol Soc 15 73ndash153 1965
Kring D A The Chicxulub impact event and its environmentalconsequences at the CretaceousndashTertiary boundary PalaeogeogrPalaeocl 255 4ndash21 2007
Kump L R Interpreting carbon-isotope excursions Strangeloveoceans Geology 19 299ndash302 1991
Machalski M Vellekoop J Dubicka Z Peryt D and Ha-rasimiuk M Late Maastrichtian cephalopods dinoflagellatecysts and foraminifera from the CretaceousndashPaleogene suc-cession at Lechoacutewka southeast Poland Stratigraphic andenvironmental implications Cretaceous Res 57 208ndash227doi101016jcretres201508012 2016
Molina E Evidence and causes of the main extinctionevents in the Paleogene based on extinction and survivalpatterns of foraminifera Earth-Sci Rev 140 166ndash181doi101016jearscirev201411008 2015
Molina E Alegret L Arenillas I and Arz J The GlobalBoundary Stratotype Section and Point for the base of the Da-nian Stage (Paleocene Paleogeneldquo Tertiaryrdquo Cenozoic) at ElKef Tunisia-Original definition and revision Episodes 29 263ndash273 2006
Olsson R K Liu C and van Fossen M The Cretaceous-Tertiarycatastrophic event at Millers Ferry Alabama in The Cretaceous-Tertiary Event and Other Catastrophes in Earth History editedby Ryder G Fastovsky D and Gartner S Geol S Am S307 263ndash277 1996
Peryt D Benthic foraminiferal response to the Cenomanian-Turonian and CretaceousndashPaleogene boundary events PrzegladGeol 52 827ndash832 2004
Peryt D Alegret L and Molina E The CretaceousPalaeogene(KP) boundary at Aiumln Settara Tunisia restructuring of benthicforaminiferal assemblages Terra Nova 14 101ndash107 2002
Ramette A Multivariate analyses in microbial ecology FEMS Mi-crobiol Ecol 62 142ndash160 2007
Rodriacuteguez-Tovar F J Uchman A MrsquoHamdi A Riahi Sand Ismail-Lattrache K B Ichnological record of palaeoen-vironment from the CretaceousndashPaleogene boundary intervalat El Kef Tunisia The first study of old and new sec-tions at the stratotype area J Afr Earth Sci 120 23ndash30doi101016jjafrearsci201604017 2016
Saner S Mudurnu-Goumlynuumlk Havzasının Jura ve Sonrasıccediloumlkelimnitelikleriyle paleocografya yorumlaması TJK Buumllteni 23 39ndash52 1980
Schioslashler P Brinkhuis H Roncaglia L and Wilson G J Di-noflagellate biostratigraphy and sequence stratigraphy of theType Maastrichtian (Upper Cretaceous) ENCI Quarry TheNetherlands Mar Micropaleontol 31 65ndash95 1997
Schueth J D Bralower T J Jiang S and Patzkowsky ME The role of regional survivor incumbency in the evolu-tionary recovery of calcareous nannoplankton from the Creta-ceousPaleogene (KPg) mass extinction Paleobiology 41 661ndash679 doi101017pab201528 2015
Scotese C R A continental drift flipbook J Geology 112 729ndash741 2004
Scotese C R and Dreher C GlobalGeology available at httpwwwGlobalGeologycom (last access 11 March 2014) 2012
Sepkoski J J Patterns of Phanerozoic extinction a perspectivefrom global data bases in Global Events and Event Stratigra-phy edited by Walliser O H Springer Berlin Germany 35ndash52 2016
Sibert E C Hull P M and Norris R D Resilience of Pacificpelagic fish across the CretaceousPalaeogene mass extinctionNat Geosci 7 667ndash670 doi101038NGEO2227 2014
Slimani H and Toufiq A A CretaceousndashPaleogene boundary geo-logical site revealed by planktic foraminifera and dinoflagellatecysts at Ouled Haddou eastern external Rif Chain Morocco JAfr Earth Sci 88 38ndash52 doi101016jjafrearsci2013080082013
Slimani H Louwye S and Toufiq A Dinoflagellate cystsform the CretaceousndashPaleogene boundary at Ouled Had-dou southeastern Rif Morocco biostratigraphy paleoenvi-ronments and paleobiogeography Palynology 34 90ndash124doi10108001916121003629933 2010
Sluijs A and Brinkhuis H A dynamic climate and ecosystemstate during the Paleocene-Eocene Thermal Maximum infer-ences from dinoflagellate cyst assemblages on the New JerseyShelf Biogeosciences 6 1755ndash1781 doi105194bg-6-1755-2009 2009
Smit J Extinction and evolution of planktonic foraminifera after amajor impact at the CretaceousTertiary boundary Geol S AmS 190 329ndash352 1982
Smit J and Romein A J T A sequence of events across theCretaceous-Tertiary boundary Earth Planet Sci Lett 74 155ndash170 1985
Speijer R P Extinction and recovery patterns in ben-thic foraminiferal paleocommunities across the Creta-ceousPaleogene and PaleoceneEocene boundaries GeologicaUltraiectina 124 Faculteit Aardwetenschappen PhD ThesisUniversiteit Utrecht Utrecht the Netherlands 1994
Speijer R P and van Der Zwaan G J Extinction and survivor-ship of southern Tethyan benthic foraminifera across the Cre-
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
900 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
taceousPaleogene boundary in Biotic recovery from mass ex-tinction events edited by Hart M B Geolog Soc Spec PublLondon UK 102 343ndash371 1996
Stemmann L and Boss E Plankton and Particle Size andPackaging From Determining Optical Properties to Drivingthe Biological Pump Annu Rev Mar Sci 4 263ndash290doi101146annurev-marine-120710-100853 2012
Thomas E Late Cretaceous-early Eocene mass extinctions in thedeep sea Geol S Am S 247 481ndash495 1990
Vandenberghe N Hilgen F J and Speijer R P The PaleogenePeriod in The Geologic Time Scale 2012 edited by GradsteinF M Ogg J G Schmitz M D and Ogg G M 2 855ndash921Elsevier Boston USA 2012
Vellekoop J Sluijs A Smit J Schouten S Weijers J WH Sinninghe Damsteacute J S and Brinkhuis H Rapid short-term cooling following the Chicxulub impact at the CretaceousndashPaleogene boundary P Natl Acad Sci USA 111 7537ndash7541doi101073pnas1319253111 2014
Vellekoop J Smit J van de Schootbrugge B Weijers J W HGaleotti S Sinninghe Damsteacute J S and Brinkhuis H Palyno-logical evidence for prolonged cooling along the Tunisian conti-nental shelf following the KndashPg boundary impact PalaeogeogrPalaeocl 426 216ndash228 doi101016jpalaeo2015030212015
Vellekoop J Esmeray-Senlet S Miller K G Browning J VSluijs A van de Schootbrugge B Sinninghe Damsteacute J Sand Brinkhuis H Evidence for CretaceousndashPaleogene bound-ary bolide ldquoimpact winterrdquo conditions from New Jersey USAGeology 44 619ndash622 doi101130G379611 2016
Woelders L and Speijer R Stable seafloor conditions sealevel and food supply during the latest Maastrichtian atBrazos River Texas Mar Micropaleontol 121 41ndash51doi101016jmarmicro201510002 2015
Zachos J C Arthur M A and Dean W E Geochemical evi-dence for suppression of pelagic marine productivity at the Cre-taceousTertiary boundary Nature 337 61ndash64 1989
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
- Abstract
- Introduction
- Geological setting and age assessment
- Materials and methods
-
- Sampling
- Foraminiferal analysis
- Palynological analysis
- Statistical analysis
-
- Results
-
- Benthic foraminifera
- Palynology
-
- Discussion
-
- Tethyan benthic foraminiferal turnover sequence
- Ecological responses to reduced export productivity
-
- Okccedilular section
- Western Tethys
- Global responses
-
- Long-term recovery
-
- Conclusions
- Data availability
- Author contributions
- Competing interests
- Acknowledgements
- References
-

J Vellekoop et al Ecological response to KndashPg boundary mass extinction 887
Figure 1 Reconstruction of the late Cretaceousndashearly Paleogene paleogeography of the North Atlantic and Mediterranean regions afterScotese (2004) and Scotese and Dreher (2012) The four sites discussed are indicated Okccedilular section Turkey (Accedilikalin et al 2015 thisstudy) El Kef Tunisia (Brinkhuis and Zachariasse 1988 Speijer and van der Zwaan 1996 Brinkhuis et al 1998) Ouled Haddou Morocco(Slimani et al 2010) and ODP Hole 1049C (Blake Nose Alegret and Thomas 2004) The red dot marks the Chicxulub impact crater
ation of endobenthic forms (eg Coccioni and Galeotti1994 Alegret et al 2015) epibenthic forms dominatethe post-impact ldquodisasterrdquo phase (Culver 2003) Since ingeneral endobenthic forms are considered indicative of ahigh flux of organic matter to the sea floor andor relativelylow oxygen conditions and epibenthic forms are generallymore common in oligotrophic open marine environments(eg Corliss 1985 Peryt et al 2002 Jorissen et al 2007Woelders and Speijer 2015) the post-impact abundance ofepibenthic forms is often explained as food starvation atthe sea floor (Culver 2003) Following this disaster phasemost benthic foraminiferal records show a relatively longrecovery phase with endobenthic forms gradually returningas diversity starts to increase again (Alegret et al 2003Culver 2003) and endobenthic microhabitats diversify(Speijer and van der Zwaan 1996)
Although combining quantitative dinocyst and benthicforaminifera analyses could provide crucial insight intochanges in pelagic-benthic-coupling (eg Guasti et al2005) perhaps surprisingly no such attempts have beenmade for the KndashPg boundary interval so far Earlier stud-ies discussed either benthic foraminiferal or dinoflagellateresponse to the KndashPg boundary each without making an at-tempt to combine the results of these studies into a compre-hensive integrated ocean-wide characterization of ecologi-cal patterns observed across the boundary In addition al-though the southern margins of the western Tethys provideda fair number of high-resolution records spanning the KndashPgboundary no such high-resolution records yet exist from thenortheastern margins of the western Tethys To be able toprovide a comprehensive (western) Tethys-wide portrayal ofthe surface and bottom water ecological changes across theKndashPg boundary and the coupling between pelagic and ben-thic systems additional dinocyst and benthic foraminiferal
records need to be generated from the northeastern margin ofthe western Tethys
The MudurnundashGoumlynuumlk basin in the central Sakarya re-gion northwestern Turkey provides new opportunities forhigh-resolution KndashPg boundary benthic foraminiferal anddinocyst records from the northern margin of the Tethys Re-cently well-preserved outcrops of ancient continental mar-gin deposits spanning the KndashPg boundary have been discov-ered in this basin (Accedilikalin et al 2015) These outcrops in-clude amongst others the Okccedilular section Here an inte-gration of dinocyst and benthic foraminifera records of thisbiostratigraphically well-constrained KndashPg boundary transi-tion is used to provide new insights into changes in and therelationship between planktic and benthic communities
Linking these records to both the previously gener-ated bulk-carbonate carbon isotope record of this section(Accedilikalin et al 2015) and to other benthic foraminiferal andpalynological records in the Tethys enables a supra-regionalportrayal of the KndashPg boundary pelagic crisis and the bio-logical changes caused by the collapse of export productivityThis integrated approach enables an evaluation of the paleoe-cological and paleoceanographic consequences of the earlyDanian Living Ocean condition
2 Geological setting and age assessment
The Okccedilular section is located in the MudurnundashGoumlynuumlkbasin (northwestern Turkey Fig 1) In this basin the KndashPg boundary interval is represented by the Tarakli Forma-tion (Saner 1980 Altiner et al 1991 Accedilikalin et al 2015)In the eastern part of the basin the upper Maastrichtianis characterized by an intercalation of mudstones and tur-bidites whereas in the western part of the basin the tur-bidites are absent The KndashPg boundary is marked by a red-dish ejecta layer at the base of a 15ndash20 cm thick boundary
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
888 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
clay layer Throughout the basin the lower 30ndash50 m of theDanian is characterized by a rhythmic alternation of fine-grained limestones and carbonate-rich mudstones (Accedilikalinet al 2015) During the latest Cretaceous to early Paleocenethis basin was characterized by mixed siliciclasticndashcarbonatesedimentation in an outer neritic to upper bathyal environ-ment (Accedilikalin et al 2015)
The Okccedilular section has been analyzed for siderophiletrace elements including Ir and other platinum groupelements (PGEs) bulk stable carbon isotopes plankticforaminifera calcareous nannofossils and dinocysts Basedon these results a detailed stratigraphic framework was out-lined (Accedilikalin et al 2015) allowing a confident age assess-ment of the boundary interval The age model shows thatthe section contains a chronostratigraphically complete KndashPg boundary interval The studied interval ranges from thetop part of the Maastrichtian A mayaroensis Zone up to thebasal part of the Danian planktic foraminiferal Zone P1b andcovers globally occurring first occurrences (FO) of dinocystmarker taxa such as Senoniasphaera inornata Damassa-dinium californicum and Carpatela cornuta
3 Materials and methods
31 Sampling
High-resolution (centimeter-scale) sample sets were ac-quired during two field campaigns in 2010 and in 2011 Formore detail on these sampling campaigns see Accedilikalin etal (2015) The samples were split for micropaleontologicaland palynological analyses
32 Foraminiferal analysis
Twenty-seven samples were processed at KU Leuven forforaminiferal studies following standard micropaleontolog-ical procedures Of these 20 samples were used for quan-titative benthic foraminiferal analyses (Fig 2) Rock sam-ples were dried in an oven at 60 C for at least 24 h De-pending on sample size 4 to 60 g of dry rock was soakedin a soda (Na2CO3) solution (50 g Lminus1) After disintegrationsamples were washed over 2 mm and 63 microm sieves If neces-sary the tenside Rewoquat was used to clean the residues andthe procedure was repeated Clean residues were dry-sievedinto three fractions 63ndash125 125ndash630 and gt 630 microm In orderto enable a direct comparison with the benthic foraminiferalrecord of the KndashPg boundary GSSP (Global Boundary Stra-totype Section and Point) of El Kef (Tunisia) we used thesame size fraction (gt 125 microm) for Okccedilular as for El Kef(Speijer and van der Zwaan 1996) Note that El Kef holdsthe GSSP of the base of the Danian (Molina et al 2006)and is thus of particular importance with respect to corre-lation and comparison of the KndashPg boundary Representativealiquots of the gt 125 microm fraction were obtained containing atleast 300 benthic foraminiferal specimens Picked specimens
from this size fraction were permanently stored in Plummerslides Benthic foraminifera were identified using the tax-onomy of Cushman (1946 1951) Kellough (1965) Aubertand Berggren (1976) Berggren and Aubert (1975) and Spei-jer (1994)
Benthic foraminifera are commonly used as indicators forbottom water oxygenation and trophic conditions (eg Joris-sen et al 2007) Here the benthic foraminiferal accumula-tion rate (BFAR number of foraminifera per cm2 per kyrsee Text S2 in the Supplement for details on the estimationof BFAR) was calculated as a semi-quantitative proxy for pa-leoproductivity (Jorissen et al 2007 and references therein)In addition the percentage of endobenthic morphotypes wascalculated using the assumed microhabitat preferences in-ferred from benthic foraminiferal morphotype analysis (egCorliss 1985 Corliss and Chen 1988 Alegret et al 2003Woelders and Speijer 2015) In general endobenthic formsare considered indicative of a high flux of organic matter tothe sea floor andor relatively low oxygen conditions whileabundance of epibenthic forms is considered characteristicfor more oligotrophic open marine environments (Jorissen etal 1995 Peryt 2004 Jorissen et al 2007) Furthermore thebi- and triserial endobenthic forms are particularly indica-tive of high food supply and low oxygenation (eg Bern-hard 1986 Corliss and Chen 1988 Jorissen et al 2007)Therefore in this study the percentage of endobenthic formswas calculated for each sample as well as the percentage bi-triserial benthic taxa to unravel food supply and oxygena-tion patterns
It should however be noted that assuming such an ana-logue with modern fauna has limitations and shortcomingsFor instance the percentage of endobenthics based on mor-photypes and using bi-triserials as an indicator for hypoxiaand high food supply is not universally applicable (Buzas etal 1993 Jorissen et al 2007 and references therein)
33 Palynological analysis
In this study the palynological data from Accedilikalin etal (2015) were analyzed and interpreted In addition twoadditional samples were analyzed (OK 15 and OK 6) toincrease the resolution of the dataset and two samples ofthe Accedilikalin et al (2015) dataset were re-counted (OK 25and OK 250 see Dataset 2 in the Supplement) Quantitativeslides of the 15ndash250 microm fraction were used All slides arestored in the collection of the Laboratory of Palaeobotanyand Palynology Utrecht University the Netherlands
To identify major changes in the dinocyst record morpho-logically closely related taxa were grouped into complexesusing a similar approach to Schioslashler et al (1997) Sluijsand Brinkhuis (2009) and Machalski et al (2016) In ourstudy the following morphological complexes were estab-lished (1) the Spiniferites complex combining all species ofSpiniferites and the morphologically similar genus Achomop-shaera (2) Manumiella spp grouping all species of Man-
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
J Vellekoop et al Ecological response to KndashPg boundary mass extinction 889
Figure 2 The benthic foraminiferal record (gt 125 microm) of the Okccedilular section The biostratigraphy is from Accedilikalin et al (2015) The clusteranalysis on the benthic foraminiferal data using paired group (UPGMA) correlation distance allows the identification of four main clustersAndashD of benthic foraminiferal taxa
umiella (3) hexaperidinioids lumping all other peridinioidcysts with a hexaform archeopyle and (4) other dinocystswhich include all other dinocyst taxa and unidentifiabledinocysts (Fig 3)
Of these different dinocyst groups previous studies haveshown that in the Tethys the hexaperidinioids in particularshow strong variations across the KndashPg boundary (Brinkhuiset al 1998 Vellekoop et al 2015) Based on statistical cor-relations between palynological records and other paleoprox-ies it has been suggested that this inferred heterotrophicgroup flourished best under high-nutrient conditions in thephotic zone (Eshet et al 1994 Brinkhuis et al 1998 Sluijsand Brinkhuis 2009) Therefore in this study abundances ofhexaperidinioids are considered indicative of nutrient avail-ability in the photic zone (see Fig S1 and Text S1 in theSupplement for a more detailed discussion on this matter)
34 Statistical analysis
To assess changes in diversity of benthic foraminifera anddinocysts across the studied interval the Shannon diversityindex (H ) the species richness per sample (S) the number ofspecimens observed per sample (N ) and the BergerndashParkerindex were calculated for both biological groups followingHayek and Buzas (2013) (Fig 4) In addition in order torecognize the main faunal associations within the benthicforaminiferal data a cluster analysis was performed usingpaired group (UPGMA) correlation distance
Q-mode non-metric multidimensional scaling (NM-MDS)was performed on the benthic foraminiferal sample compo-sitions to assess patterns in assemblage response to KndashPgboundary perturbations Since taxa may not have a linear re-sponse to environmental changes across the KndashPg boundaryQ-mode NM-MDS is preferred over PCA and CCA (Ram-ette 2007)
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
890 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
Figure 3 The organic-walled dinoflagellate cyst record of theOkccedilular section The biostratigraphy and bulk carbonate stable car-bon isotope records are from Accedilikalin et al (2015) The four maindinocyst complexes are indicated
4 Results
41 Benthic foraminifera
The benthic foraminiferal record of the Okccedilular section ischaracterized by a distinct turnover across the KndashPg bound-ary (Figs 2 4 5 6) Of the common taxa 8 out of 30(sim 27 ) disappear across the KndashPg boundary After the KndashPg boundary crisis three new taxa appear The estimatedBFAR shows a major decrease across the boundary (Fig 5)For the foraminiferal counts and illustrations of commonforms see Dataset S1 and Figs S2 S3
The cluster analysis allows the identification of fourmain clusters of benthic foraminiferal taxa clusters A toD (Fig 2) Cluster C is relatively large and can be subdi-vided into five subclusters Based on the succession of ben-thic faunal assemblages characterized by strong changes inthe Shannon diversity index (H ) four intervals can be rec-ognized in the benthic foraminiferal record of the Okccedilularsection (Figs 4 6)
The first interval comprises the uppermost Maastrichtianand is characterized by relatively high diversity dominatedby the taxa in cluster D In this assemblage the bi-triserialbenthic taxa are relatively abundant (22ndash28 ) with char-acteristic taxa such as Bulimina arkadelphiana Eouvigerinasubsculptura and Praebulimina reussi About 50ndash60 of theassemblage consists of inferred endobenthic taxa The KndashPg boundary marks an abrupt benthic community impover-
ishment and the decimation of the taxa in cluster D Abovethe boundary the bi-triserial benthic taxa virtually disappearfrom the record (mostly below lt 05 )
The second interval characterized by low diversity com-prises the lowermost Danian approximately correspondingto planktic foraminiferal Zone P0 In this interval epibenthicforms are most abundant (70ndash90 ) It is dominated by thetaxa of cluster A encompassing successive peak occurrencesof the taxa Anomalinoides praeacutus Trochammina sppand Cibicidoides pseudoacutus
The third interval covers the part of the succession approx-imately correlative to planktic foraminiferal Zone Pα Thisinterval is characterized by a recovery of diversity althoughthe diversity is still lower than that of the top Maastrichtianassemblage The abundance of taxa in cluster A slowly de-creases and taxa in cluster B mainly represented by Osan-gularia plummerae and Cibicidoides sp become abundantEndobenthic forms recover and make up 30ndash40 of the totalassemblage
The fourth interval starts in the interval correlative to ZoneP1a Here the benthic community has recovered as the di-versity has stabilized and is almost similar to pre-impact val-ues This interval is characterized by taxa in cluster C a typ-ical Paleocene Midway-type fauna (Berggren and Aubert1975) with representatives such as Anomalinoides praea-cutus Coryphostoma midwayensis Cibicidoides alleni andOsangularia plummerae
42 Palynology
Palynological samples from the Okccedilular site yield abundantpalynomorphs dominated by dinocysts and with minor con-tributions of acritarchs prasinophytes organic foraminiferallinings and terrestrial palynomorphs (Dataset S2) Thedinocyst associations of the MudurnundashGoumlynuumlk basin arequite diverse including components characteristic of boththe Tethyan and Boreal realms (Accedilikalin et al 2015) As ex-pected the dinocyst record does not show major changes indiversity across the KndashPg boundary (Fig 4) There is a steadydecrease in dinocyst diversity from planktic foraminiferalZone P1a upwards probably a long-term change not re-lated to the impact The estimated dinocyst accumulationrate (DAR number of preserved cysts per cm2 per kyr seeText S2 in the Supplement for details on the estimation of theDAR) shows no major changes across the KndashPg boundary
Throughout the palynological record the Spiniferites com-plex is consistently the most dominant morphogroup in gen-eral comprising 40ndash50 of the total assemblage (Fig 3)Similar to other KndashPg boundary sites worldwide (Habib andSaeedi 2007) Manumiella spp show an episode of higherrelative abundance near the top of the Maastrichtian some30ndash40 cm below the KndashPg boundary Hexaperidinioids gen-erally make up a relatively small component of the Maas-trichtian assemblage (3ndash17 ) but show a strong increaseacross the KndashPg boundary In the boundary clay layer cor-
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
J Vellekoop et al Ecological response to KndashPg boundary mass extinction 891
Figure 4 Diversity indices on the benthic foraminiferal and dinocyst records of the Okccedilular section with the Shannon diversity index (H ) thespecies richness per sample (S) the number of specimen observed per sample (N ) and the BergerndashParker index As the bulk stable carbonisotope record and the Shannon diversity index of the benthic foraminiferal record show similar trends the combination of these records canbe used to subdivide the KndashPg boundary transition into four intervals I the Maastrichtian ie ldquopre-impactrdquo interval II the post-impactinterval representing the ldquodisasterrdquo phase III a recovery interval and IV the early Paleocene post-recovery interval The division criteriaare explained in the text
relative to planktic foraminiferal Zone P0 this group in-creases to 35 of the assemblage In the peak intervals thehexaperidinioids are mostly represented by representativesof the genera Senegalinium and Cerodinium After an ini-tial drop in relative abundance in the upper half of Zone P0this group reaches a second maximum (sim 35 ) at the baseof Zone Pα and remains relatively abundant (15ndash30 ) up toZone P1a After this the hexaperidinioids gradually decreasetowards the top of the studied interval The ldquoother dinocystsrdquogroup ndash with representatives such as Areoligera spp Im-pagidinium spp Hystrichosphaeridium tubiferum Opercu-lodinium centrocarpum and Palynodinium grallator ndash gen-erally makes up 25ndash50 of the assemblage
5 Discussion
51 Tethyan benthic foraminiferal turnover sequence
The major turnover in the benthic community at Okccedilularis largely comparable with the previously published ben-thic foraminiferal record (size fraction gt 125 microm) of the KndashPg boundary GSSP of El Kef (Figs 7 8 Speijer and Vander Zwaan 1996) In addition other benthic foraminiferalrecords from the margins of the Tethys also show a strong
similarity to the Okccedilular and El Kef records including stud-ies that use the gt 63 microm size fraction (Alegret et al 2003Culver 2003 and references therein)
Although it is sometimes suggested that different sizefractions cannot be directly compared because the differ-ent size fractions sometimes harbor different ecological in-formation (Giusberti et al 2016) intercomparison studieshave shown that when comparing the gt 125 and gt 63 microm sizefractions in benthic foraminiferal assemblages the overallpatterns are generally robust (Ernst et al 2006) The factthat gt 63 and gt 125 microm KndashPg boundary benthic foraminiferalrecords from different Tethys-margin sites (eg Speijer andVan der Zwaan 1996 Peryt et al 2002 Culver et al 2003)show striking similarities even though both the size fractionsused (gt 63 vs gt 125 microm) and specific taxa making up theforaminiferal assemblages differ among sites confirms theobservations by Ernst et al (2006) Furthermore even whenit would be assumed that different size fractions are diffi-cult to compare directly the striking similarities between theTethyan gt 125 and gt 63 microm size fraction KndashPg boundary ben-thic foraminiferal records provide compelling evidence for astrong disruption of the entire benthic community
Tethyan middle neritic to middle bathyal KndashPg boundarysuccessions generally show a sequence of (1) a typical high
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
892 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
Figure 5 Estimated benthic foraminiferal accumulation rate (BFAR number of foraminifera of the gt 63 microm size fraction per cm2 sea floorper kyr) and estimated dinocyst accumulation rate (DAR number of preserved cysts per cm2 sea floor per kyr) of the Okccedilular sectionUncertainty in estimated accumulation rates resulting from uncertainties in sediment densities and sedimentation rates are indicated in greyproviding a range of estimated accumulation rates For more detailed info on the calculation of the BFAR and DAR see Text S2 of theSupplement
diversity assemblage in the Maastrichtian (2) a low diver-sity ldquodisasterrdquo assemblage directly after the KndashPg boundaryimpact characterized by a short-lived bloom of endobenthicforms observed in several high-resolution records (eg Coc-cioni and Galeotti 1994 Speijer and van der Zwaan 1996)followed by a dominance of epibenthic forms (3) a ldquore-coveryrdquo assemblage characterized by an increasing diversityand returning endobenthic forms and (4) a new high diver-sity assemblage dominated by a Paleocene fauna with bothepi- and endobenthic forms equally represented
Hence based on this succession of benthic assemblageswhich are characterized by strong changes in diversity (H )the KndashPg boundary benthic foraminiferal records from theTethys can be subdivided into four intervals (Figs 2 4ndash6)The intervals I to IV approximately correspond to the up-permost Maastrichtian planktic foraminiferal Zone P0 sub-zones PαndashP1a and subzones P1andashP1b respectively There-fore this succession roughly follows the 5-fold sedimen-tological and planktic foraminiferal sequence of Smit andRomein (1985) in which unit 2 represents the ejecta layerand units 1 3 4 and 5 correspond to intervals I to IV in ourstudy
The sequence of benthic foraminiferal turnover in theTethys is also well illustrated by a Q-mode NM-MDS
(Fig 7) The Q-mode NM-MDS of the benthic foraminiferalsample compositions of the combined data from Okccedilular(this study) and El Kef (Speijer and van der Zwaan 1996)demonstrates that although the assemblages differ betweenthese localities on opposite margins of the Tethys the re-sponses of the benthic foraminiferal assemblages to the KndashPgboundary perturbations express very similar overall patternsacross the KndashPg boundary At both Okccedilular and El Kef thebenthic foraminiferal records show a rapid transition from astable Maastrichtian assemblage to the earliest Danian dis-aster phase in Zone P0 followed by a gradual change backtowards conditions similar to the Maastrichtian (marked bythe return of Lazarus taxa) Therefore both the successionof benthic faunal assemblages including diversity indicesand Q-mode NM-MDS analysis provide powerful tools to al-low the comparison of KndashPg boundary benthic foraminiferalrecords from different localities
52 Ecological responses to reduced export productivity
521 Okccedilular section
The palynological benthic foraminiferal and bulk stable iso-tope records of the Okccedilular section show major changesacross the KndashPg boundary interval (Figs 2ndash7) portraying
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
J Vellekoop et al Ecological response to KndashPg boundary mass extinction 893
Figure 6 Relative abundances of the four main clusters of benthic foraminiferal taxa (gt 125 microm) relative abundances of epibenthic andendobenthic forms and relative abundance of bi-triserial benthic taxa
Figure 7 Q-mode non-metric multidimensional scaling (on all sam-ples in the gt 125 microm size fraction benthic foraminiferal datasets ofOkccedilular (this study) and El Kef (Speijer and van der Zwaan 1996)After an abrupt transition from latest Maastrichtian pre-impact as-semblages to disaster assemblages in Zone P0 and recovery assem-blages in Zone Pα a gradual transition towards a new equilibriumpost-recovery assemblage can be observed in subzones P1a and P1b
the biological crisis following the impact Following the KndashPg boundary mass extinction endobenthic forms stronglydecrease with bi-triserial benthic taxa almost disappearingfrom the benthic community simultaneously the estimatedBFAR shows a major fall (Fig 5) This interval approxi-mately correlative to foraminiferal Zone P0 represents thedisaster phase The drop in bi-triserial benthic taxa (Fig 6)and decrease in BFAR (Fig 5) point to a reduction in foodsupply to the benthic community indicating that the trans-port of organic matter to the sea floor must have decreasedIn the dinocyst community however hexaperidinioids showa strong increase in relative abundance during this phase(Figs 3 6 7) suggesting an increase in nutrient availabilityin the photic zone while the DAR remains relatively stableacross the KndashPg boundary (Fig 5)
The decrease in benthic diversity and the high relativeabundances of opportunistic Trochammina spp and Cibici-doides pseudoacutus suggest that the benthic community ex-perienced additional stress during Zone P0 besides food lim-itation (see Jorissen et al 2007) If food limitation were theonly factor involved upper bathyal assemblages as observedat Okccedilular would be expected to become more similar tooligotrophic middle to lower bathyal assemblages Insteadthe opposite is true the assemblage composition of the dis-aster phase is closer to neritic Midway-type assemblages
It was previously suggested that the epibenthic taxon Cibi-cidoides pseudoacutus could potentially be hypoxia resistant
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
894 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
(Speijer and van der Zwaan 1996) As a collapse of the bi-ological pump should lead to enhanced remineralization oforganic matter in the photic zone it is indeed likely thatthe oxygen minimum zones expanded and shoaled leadingto oxygen stress on the sea floor Although early Paleocenedysoxia at El Kef is supported by the ostracod record (Donzeet al 1982) and ichnofacies analyses (Rodriguez-Tovaret al 2016) supporting (eg geochemical) evidence fordysoxia during the disaster phase at Okccedilular record is absentFurthermore an additional stressor for the benthic commu-nity during Zone P0 may be present In contrast to calcare-ous nanoplankton organic-walled cyst-producing dinoflagel-lates did not suffer extinctions and may have become a moreimportant component in the early Paleocene phytoplanktoncommunity Hence besides the reduction of the amount oforganic matter transported to the sea floor the compositionof food supplied by the photic zone likely changed signifi-cantly across the KndashPg boundary as well (DrsquoHondt 2005)possibly presenting an additional stress factor for the benthiccommunity (Speijer and van der Zwaan 1996 Alegret andThomas 2009)
522 Western Tethys
The patterns in benthic foraminiferal and dinoflagellate re-sponse to the KndashPg boundary perturbations at Okccedilular appearto be characteristic for the continental margins of the westernTethyan realm The benthic foraminiferal patterns are similaracross different size fractions suggesting that they are repre-sentative of the overall benthic communities In Tunisia andSpain where both benthic foraminiferal and dinoflagellaterecords are available the earliest Danian is also characterizedby a decrease in endobenthic foraminifera at the sea floor(Speijer and Van der Zwaan 1996 Peryt et al 2002 Ale-gret et al 2003) and simultaneously blooms of hexaperidin-ioids in the water column (Brinkhuis et al 1998 Vellekoopet al 2015 Fig 8) In the Ouled Haddou section Moroccoin the westernmost Tethys (Fig 1) the lowermost Danian isalso characterized by a strong increase of the hexaperidin-ioid Senegalinium group up to 30 of the assemblage (Sli-mani et al 2010) that is very similar to the Okccedilular record(Fig 8)
The KndashPg boundary dinocyst record of El Kef is strikinglysimilar to the one from the Okccedilular section showing two dis-tinct peaks in hexaperidinioids one in Zone P0 and one atthe basal part of Zone Pα At El Kef the initial post-impactdominance of hexaperidinioids is however less pronounced(Fig 8) This difference in expression might be related tosmall differences in paleogeographic and paleoceanographicsettings between sites
In addition some high-resolution benthic KndashPg records inthe Tethyan realm show a short-lived bloom of specific en-dobenthic foraminiferal taxa in the few centimeter directlyabove the KndashPg boundary sometimes accompanied by otherindicators of low oxygen levels (eg Coccioni and Gale-
otti 1994 Speijer and Van der Zwaan 1996 Kaiho et al1999) This suggests that at least locally the sea floor wastemporarily characterized by hypoxic conditions directly fol-lowing the impact This short-lived deoxygenation is possi-bly related to the mass mortality at the KndashPg boundary re-sulting in a large short-lived flux of organic matter to thesea floor (Coccioni and Galeotti 1994) However since ex-port productivity was greatly reduced after the KndashPg bound-ary mass extinction very little ldquonewrdquo organic matter reachedthe sea floor after the initial post-impact influx Hence afteraccumulated organic matter was remineralized the benthiccommunity starved resulting in a transition to an epibenthic-dominated benthic fauna similar to the post-impact fauna ofthe Okccedilular record
The combined dinocyst records suggest that the early Pa-leocene margins of the Tethys were characterized by an in-crease in nutrient availability in the photic zone whereascoeval benthic foraminiferal records indicate a major de-crease in nutrient supply to the sea floor These changesin the benthic foraminiferal and dinocyst communities arelikely caused by the major reduction of both the efficiencyand strength of the biological pump (ie the fraction ofprimary production exported from the photic zone and theamount of organic matter transported down) in accordancewith the Living Ocean model (DrsquoHondt and Zachos 1998DrsquoHondt et al 1998 DrsquoHondt 2005 Coxall et al 2006)For a detailed explanation of the difference between biologi-cal pump efficiency and biological pump strength see Hiltinget al (2008)
As bulk carbonate δ13C reflects the isotopic compositionof the surface ocean which is set by burial fractions and bythe photosynthetic isotope effect (Kump 1991 Hain et al2014) the excursion recorded in bulk δ13C records world-wide and the rapid collapse in surface to deep-ocean carbonisotope gradients likely reflect the reduction of the global in-tensity (ie efficiency) of the biological pump (Hain et al2014) The recorded inverse change in nutrient availabilitysuggests a causal link A reduced efficiency of the biologi-cal pump and associated decrease of the fraction of biomasstransported out of the photic zone could have resulted inhigh rates of nutrient recycling in the upper part of the wa-ter column (DrsquoHondt 2005) This suggests that the reduc-tion of the biological pump strength recorded by the benthicforaminiferal record is a consequence of the decreased effi-ciency of the biological pump recorded by the carbon iso-tope and dinoflagellate cyst records The strong correlationbetween the bulk carbon isotope curves and the Shannondiversity index (H ) of the benthic foraminiferal records atboth Okccedilular (R2
= 073 p lt 0001) and El Kef (R2= 074
p lt 0001) suggest a clear link between the changes in ben-thic foraminiferal assemblages and the collapse and recoveryof biological pump efficiency
As a result of the reduced biological pump efficiency morenutrients remained available for the early Paleocene phyto-plankton community At Tethyan margin sites this is indi-
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
J Vellekoop et al Ecological response to KndashPg boundary mass extinction 895
Figure 8 The benthic foraminiferal (gt 125 microm) dinocyst and bulk stable carbon isotope records of Okccedilular (Turkey) and El Kef (Tunisia)the dinocyst record of Ouled Haddou (Morocco) and the benthic foraminiferal record (gt 63 microm) of ODP Hole 1049C Blake Nose (NorthAtlantic) Note that the definition of ldquohexaperidinioidsrdquo used is provided in the text The four phases identified based on the bulk stablecarbon isotopes and the Shannon diversity index (H ) of the benthic foraminiferal records are indicated (IndashIV) The gt 63 microm size fractionbenthic foraminiferal record of ODP Hole 1049C is from Alegret and Thomas (2004) who argued that the lowermost Paleocene of thissite comprised reworked foraminifera This interval is indicated by a dashed bar in the figure The biostratigraphy of Ouled Haddou is fromSlimani and Toufiq (2013) whereas the dinocyst record of this site is from Slimani et al (2010) The biostratigraphy of El Kef is based onBrinkhuis et al (1998) Speijer and van der Zwaan (1996) and Molina et al (2006) The fine-fraction stable carbon isotope record (blackdots) of El Kef is from Keller and Lindinger (1989) The Shannon diversity index (red dots) relative abundances of bi-triserial benthic taxa(dark purple) and all endobenthic foraminifera (light purple) are based on the gt 125 microm size fraction foraminiferal data of Speijer and vander Zwaan (1996) The palynological data of El Kef are from Brinkhuis et al (1998) (lower part) and Guasti et al (2005) (upper part) TheShannon diversity index of the gt 125 microm size fraction benthic foraminiferal record (red dots) the relative contribution of bi-triserial benthictaxa (dark purple) all endobenthic foraminifera (light purple) and hexaperidinioid dinocysts from Okccedilular are from this study The bulkstable carbon isotope record of Okccedilular (black dots) is from Accedilikalin et al (2015) Mbsf indicates meters below sea floor
cated by the higher abundance of hexaperidinioids Similarlyblooms of eutrophic survivor taxa of calcareous nanoplank-ton at open ocean sites have been suggested to be relatedto the buildup of nutrients in the open ocean photic zone(Schueth et al 2015) Although the carbon isotope gradientsbetween surface and deep waters indicate that the recoveryof the biological pump took hundreds of thousands of years(Zachos et al 1989 Kump 1991 DrsquoHondt et al 1998 Cox-all et al 2006 Birch et al 2016) our records suggest thatthe enhanced recycling of nutrients in the Tethys was partic-ularly intense during two phases in the first tens of thousandsyears after the impact (Fig 8)
523 Global responses
The decrease in export productivity following the KndashPgboundary is not only recognized in the Tethyan realm In theAtlantic Ocean Southern Ocean and the Indian Ocean a de-crease of the biological pump strength after the KndashPg bound-ary was also observed in neritic to abyssal environments(eg Thomas 1990 Olsson et al 1996 Hull et al 2011Alegret et al 2012) As an example the gt 63 microm size frac-tion benthic foraminiferal KndashPg boundary record of BlakeNose (ODP Hole 1049C northwestern Atlantic Alegret andThomas 2004) shows a pattern that is strikingly similar tothe gt 125 microm size fraction records in the Tethyan realm witha strong decrease of endobenthics in Zone P0 followed by a
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
896 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
gradual recovery across zones Pα and P1a (Fig 8) The simi-larities between the benthic foraminiferal patterns of Tethyanouter neritic to middle bathyal (200ndash1500 m) sites and thoseof the middle to lower bathyal (1500ndash1600 m) record ofBlake Nose (Alegret and Thomas 2004) suggest that theserecords register a global decrease in export productivity fol-lowing the KndashPg boundary
However several open ocean sites mostly in the Pacificshow an opposite trend with several lines of evidence sug-gesting an increased biological pump strength at these sites(eg Hull et al 2011 Alegret et al 2012) This led Ale-gret et al (2012) to conclude that the decrease in biologicalpump strength after the KndashPg boundary was most likely aregional instead of a global effect arguing against the Liv-ing Ocean hypothesis We would argue that low-productivityopen ocean sites like the central Pacific are characterized byvery different ecosystem structures and higher productivityas those along the Tethyan margins (eg Dortch and Pack-erd 1989) Therefore the consequence of ecosystem reorga-nization resulting from the mass extinction was likely verydifferent in the open ocean Pacific (eg Sibert et al 2014)Esmeray-Senlet et al (2015) proposed the term ldquoHeteroge-neous Oceanrdquo for this conceptual model as an alternativeto the Living Ocean model Supposedly the HeterogeneousOcean was characterized by a strong geographic heterogene-ity in the extinction patterns and food supply to the sea floor
However as the primary food source for open ocean deep-sea benthic communities generally consists of refractory or-ganic matter (eg Jiao et al 2010) it remains uncertainwhether changes in the intensity of recycling of the gener-ally more labile organic matter in the surface ocean will re-sult in changes in the amount of organic matter that benthiccommunities receive as they will likely continue to receiverecalcitrant organic matter at barely diminished rates despitethe collapse of the biological pump
In addition it should be noted that although the bio-logical pump efficiency is generally relatively high in openocean realms such as the central Pacific the biological pumpstrength is generally very low at these sites (Honjo et al2008 Henson et al 2011) Small changes in absolute bio-logical pump strength at such a locality could therefore havehad a large effect on the local oligotrophic benthic commu-nity (Alegret and Thomas 2009) but they likely representedonly a small fraction of the net amount of organic matterglobally transported to the deep Hence even though localbiological pump strength might have increased at some low-productivity deep-sea sites the large decrease in biologi-cal pump strength recorded at many other sites means thatthe net amount of organic matter globally transported out ofthe surface ocean was likely still reduced in the post-impactworld (Birch et al 2016) Therefore we argue that such Pa-cific records do not represent the general pattern The globalocean response can be characterized as heterogeneous how-ever the KndashPg boundary mass extinction did lead to a strongreduction of the net amount of organic matter globally trans-
ported from the surface to the sea floor while biological pro-ductivity recovered rapidly after the extinction event in ac-cordance with the Living Ocean hypothesis
53 Long-term recovery
Following the disaster phase the hexaperidinioid cysts atOkccedilular and El Kef decreased in abundance and the abun-dance of endobenthic forms increased again as the recoveryof the benthic community was initiated This recovery phaseis approximately correlative to Zone Pα and the basal part ofZone P1a which according to the Paleogene age constraintsof Vandenberghe et al (2012) represents at least 300 kyrThis duration is in agreement with the estimations providedby Birch et al (2016) During this phase the carbon iso-tope records remain well below pre-impact values and thediversity of the benthic foraminiferal community has not yetfully recovered (Figs 4ndash8) This indicates that the rapid andshort-lived KndashPg boundary disaster was followed by a rela-tively long recovery phase in line with previous estimatesof a multimillion-year biological recovery (eg Coxall etal 2006) While the impact-related environmental perturba-tions were short lived (Kring 2007 Vellekoop et al 20142016) the extinctions amongst important biological groupsled to a reduction of the organic flux from the photic zone todeep water resulting in major long-term biological and pa-leoceanographic reorganizations Only with the evolutionaryrecovery of the pelagic community governing the biologicalcarbon pump did export productivity start to increase again(eg Coxall et al 2006 Birch et al 2016)
6 Conclusions
The marine palynological benthic foraminiferal and bulkstable isotope records of the Okccedilular and El Kef sections re-veal major changes across the KndashPg boundary interval por-traying the biological crisis at the KndashPg boundary and subse-quent recovery in the early Paleocene Based on the succes-sion of benthic faunal assemblages at a number of Tethyanmargin sites four phases can be recognized across the KndashPgboundary interval the Maastrichtian or pre-impact phase (I)a disaster phase (II) a recovery phase (III) and an early Pa-leocene post-recovery phase (IV)
Following the KndashPg boundary impact some localitiesshow a large short-lived flux of food to the sea floor likelyrelated to the mass mortality at the KndashPg boundary Howeversince export productivity was greatly reduced after the KndashPgboundary mass extinction it is likely that after the initialpost-impact influx very little fresh organic matter reachedthe sea floor This reduction of export productivity in thepost-extinction disaster phase eventually resulted in a lowerfood supply to the sea floor This presented a major stressfactor for benthic organisms in the western Tethys The re-duced food supply resulted in an abrupt impoverishment of
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
J Vellekoop et al Ecological response to KndashPg boundary mass extinction 897
benthic communities As the downward transport of nutri-ents was slowed down recycling in upper layers increasedAs a result more nutrients remained available for the earlyPaleocene phytoplankton community leading to blooms ofdinoflagellates along the Tethyan margins and blooms of cal-careous nanoplankton taxa in more open ocean sites Ourrecords show that the enhanced recycling of nutrients in theTethys was particularly intense in the tens of thousands ofyears after the impact Following this the slow evolution-ary recovery of the pelagic community governing the bio-logical carbon pump resulted in a gradually increasing ex-port productivity in the hundreds of thousands of years afterthe impact Hence our integration of dinocyst and benthicforaminiferal records across the KndashPg boundary provides im-portant new insights into the ecological responses to the re-duction of export productivity following the mass extinctionat the CretaceousndashPaleogene boundary highlighting the di-rect ecological consequences of the Living Ocean conditionsin the post-impact world
7 Data availability
All data related to publication are available in the Supple-ment
The Supplement related to this article is available onlineat doi105194bg-14-885-2017-supplement
Author contributions Johan Vellekoop Lineke Woelders andHenk Brinkhuis designed the research Samples were collectedin the field by Johan Vellekoop Sanem Accedilikalin Ismail Ouml Yil-maz and Jan Smit Palynological analyses were carried out by Jo-han Vellekoop Benthic foraminiferal analyses were carried out byLineke Woelders and Robert P Speijer Johan Vellekoop LinekeWoelders and Bas van de Schootbrugge prepared the manuscriptwith input from all authors
Competing interests The authors declare that they have no conflictof interest
Acknowledgements This work was supported by the NetherlandsOrganization for Scientific Research (NWO) Open CompetitionGrant ALWPJ09047 to H Brinkhuis and the Research FoundationFlanders (FWO) grant G0B8513 to R P Speijer We thank LKump F Boscolo Galazzo P Roopnarine and one anonymousreferee for their suggestions
Edited by D GillikinReviewed by L Kump F Boscolo Galazzo and P Roopnarine
References
Accedilikalin S Vellekoop J Ocakoglu F Yılmaz I Ouml Smit J Al-tiner S O Goderis S Vonhof H Speijer R P WoeldersL Fornaciari E and Brinkhuis H Geochemical and pale-ontological characterization of a new KndashPg Boundary local-ity from the Northern branch of the Neo-Tethys MudurnundashGoumlynuumlk Basin NW Turkey Cretaceous Res 52 251ndash267doi101016jcretres201407011 2015
Alegret L and Thomas E Food supply to the seafloor in the Pa-cific Ocean after the CretaceousPaleogene boundary event MarMicropaleontol 73 105ndash116 2009
Alegret L Molina E and Thomas E Benthic foraminiferalturnover across the CretaceousPaleogene boundary at Agost(southeastern Spain) paleoenvironmental inferences Mar Mi-cropaleontol 48 251ndash279 2003
Alegret L Thomas E and Lohmann K C End-Cretaceous ma-rine mass extinction not caused by productivity collapse P NatlAcad Sci USA 109 728ndash732 doi101073pnas11106011092012
Alegret L Rodriacuteguez-Tovar F J and Uchman A How bioturba-tion obscured the CretaceousndashPaleogene boundary record TerraNova 27 225ndash230 doi101111ter12151 2015
Altiner D Koccedilyigit A Farinacci A Nicosia U and ContiM A Jurassic-Lower Cretaceous stratigraphy and paleogeo-graphic evolution of the southern part of north-western Anatolia(Turkey) Geologica Romana 27 13ndash80 1991
Alvarez LW Alvarez W Asaro F and Michel H V Extrater-restrial cause for the Cretaceous-Tertiary extinction Science208 1095ndash1108 1980
Alegret L and Thomas E Benthic foraminifera and environmen-tal turnover across the CretaceousPaleogene boundary at BlakeNose (ODP Hole 1049C Northwestern Atlantic) PalaeogeogrPalaeocl 208 59ndash83 doi101016jpalaeo200402028 2004
Armstrong R A Lee C Hedges J I Honjo S and WakehamS G A new mechanistic model for organic carbon fluxes in theocean based on the quantitative association of POC with ballastmaterials Deep-Sea Res Pt II 49 219ndash236 2002
Aubert J and Berggren W A Paleocene benthic foraminiferalbiostratigraphy and paleoecology of Tunisia Bull du Cent deRech de Pau ndash SNPA 10 379ndash469 1976
Berggren W A and Aubert J Paleocene benthonic foraminiferalbiostratigraphy paleobiogeography and paleoecology ofAtlanticndashTethyan regions midway-type fauna PalaeogeogrPalaeocl 18 73ndash192 1975
Bernaola G and Monechi S Calcareous nannofossil ex-tinction and survivorship across the CretaceousndashPaleogeneboundary at Walvis Ridge (ODP Hole 1262C SouthAtlantic Ocean) Palaeogeogr Palaeocl 255 132ndash156doi101016jpalaeo200702045 2007
Bernhard J M Characteristic assemblages and morphologies ofbenthic foraminifera from anoxic organic-rich deposits Jurassicthrough Holocene J Foramin Res 16 207ndash215 1986
Birch H S Coxall H K Pearson P N Kroon D andSchmidt D N Partial collapse of the marine carbon pump af-ter the CretaceousndashPaleogene boundary Geology 44 287ndash290doi101130G375811 2016
Brinkhuis H and Zachariasse W J Dinoflagellate cysts sealevel changes and planktonic foraminifers across the Cretaceous-
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
898 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
Tertiary boundary at El Haria northwest Tunisia Mar Micropa-leontol 13 153ndash191 1988
Brinkhuis H Bujak J P Smit J Versteegh G J M and Viss-cher H Dinoflagellate-based sea surface temperature recon-structions across the Cretaceous-Tertiary boundary PalaeogeogrPalaeocl 141 67ndash83 1998
Buesseler K O The decoupling of production and particulate ex-port in the surface ocean Global Biogeochem Cy 12 297ndash3101998
Buzas M A Culver S J and Jorissen F J A statisti-cal evaluation of the microhabitats of living (stained) infau-nal benthic foraminifera Mar Micropaleontol 20 311ndash320doi1010160377-8398(93)90040-5 1993
Coccioni R and Galeotti S KndashT boundary extinction geolog-ically instantaneous or gradual event Evidence from deep-seabenthic foraminifera Geology 22 779ndash782 1994
Corliss B H Microhabitats of benthic foraminifera within deep-sea sediments Nature 314 435ndash438 doi101038314435a01985
Corliss B H and Chen C Morphotype patterns of Nor-wegian Sea deep-sea benthic foraminifera and ecologi-cal implications Geology 16 716ndash719 doi1011300091-7613(1988)016lt0716MPONSDgt23CO2 1988
Coxall H K DrsquoHondt S and Zachos J C Pelagic evolution andenvironmental recovery after the CretaceousndashPaleogene mass ex-tinction Geology 34 297ndash300 doi101130G217021 2006
Culver S J Benthic foraminifera across the CretaceousndashTertiary(KndashT) boundary a review Mar Micropaleontol 47 177ndash2262003
Cushman J A Upper Cretaceous foraminifera of the Gulf coastalregion of the United States and adjacent areas US Geolog SurvProf Pap 206 241 pp 1946
Cushman J A Paleocene foraminifera of the Gulf Coastal Regionof the United States and adjacent regions US Geol Surv ProfPap 232 75 pp 1951
DrsquoHondt S Consequences of the CretaceousPaleogene Mass Ex-tinction for Marine Ecosystems Annu Rev Ecol Evol S36 295ndash317 doi101146annurevecolsys350211031057152005
DrsquoHondt S and Zachos J C Cretaceous foraminifera and the evo-lutionary history of planktic photosymbiosis Paleobiology 24512ndash523 1998
DrsquoHondt S Donaghay P Zachos J C Luttenberg D andLindinger M Organic Carbon Fluxes and Ecological Recov-ery from the Cretaceous-Tertiary Mass Extinction Science 282276ndash279 1998
Donze P Colin J-P Damotte R Oertli HJ Peypouquet J-Pand Said R Les ostracodes du Campanien terminal agrave lrsquoEocegraveneinfeacuterieur de la coupe du Kef Tuneacutesie nord-occidentale Bull desCentr Recher Expl-Prod Elf Aquit 6 273ndash335 1982
Dortch Q and Packard T T Differences in biomass structure be-tween oligotrophic and eutrophic marine ecosystems Deep-SeaRes 36 223ndash240 1989
Ernst S R Guasti E Dupuis C and Speijer R P En-vironmental perturbation in the southern Tethys across thePaleoceneEocene boundary (Dababiya Egypt) Foraminiferaland clay mineral records Mar Micropaleontol 60 89ndash111doi101016jmarmicro200603002 2006
Eshet Y Moshkovitz S Habib D Benjamini C and Maga-ritz M Calcareous nannofossil and dinoflagellate stratigraphyacross the CretaceousTertiary boundary at Hor Hahar IsraelMar Micropaleontol 18 199ndash228 1992
Eshet Y Almogi-Labin A and Bein A Dinoflagellate cysts pa-leoproductivity and upwelling systems A Late Cretaceous ex-ample from Israel Mar Micropaleontol 23 231ndash240 1994
Esmeray-Senlet S Wright J D Olsson R K MillerK G Browning J V and Quan T M Evidencefor reduced export productivity following the Creta-ceousPaleogene mass extinction Paleoceanography 30718ndash738 doi1010022014PA002724 2015
Giusberti L Boscolo Galazzo F and Thomas E Variability inclimate and productivity during the Paleocene-Eocene ThermalMaximum in the western Tethys (Forada section) Clim Past12 213ndash240 doi105194cp-12-213-2016 2016
Guasti E Kouwenhoven T J Brinkhuis H and Speijer R PPaleocene sea-level and productivity changes at the southernTethyan margin (El Kef Tunisia) Mar Micropaleontol 55 1ndash17 2005
Habib D and Saeedi F The Manumiella seelandica global spikeCooling during regression at the close of the MaastrichtianPalaeogeogr Palaeocl 255 87ndash97 2007
Hain M P Sigmal D M and Haug G H 818 ndash The biolog-ical Pump in the Past in Reference Module in Earth Systemsand Environmental Sciences Treatise on Geochemistry (SecondEdition) The Oceans and Marine Geochemistry 8 485ndash517doi101016B978-0-08-095975-700618-5 2014
Hayek L C and Buzas M A On the proper and efficient useof diversity measures with individual field samples J ForaminRes 43 305ndash313 2013
Henson S A Sanders R Madsen E Morris P J Le MoigneF and Quartly G D A reduced estimate of the strength ofthe oceanrsquos biological carbon pump Geophys Res Lett 38L04606 doi1010292011GL046735 2011
Hilting A K Kump L R and Bralower T J Variations in theoceanic vertical carbon isotope gradient and their implicationsfor the Paleocene-Eocene biological pump Paleoceanography23 PA3222 doi1010292007PA001458 2008
Honjo S Manganini S J Krishfield R A and Francois RParticulate organic carbon fluxes to the ocean interior and fac-tors controlling the biological pump A synthesis of global sed-iment trap progams since 1983 Prog Oceanogr 76 217ndash285doi101016jpocean200711003 2008
Huber B T MacLeod K G and Norris R D Abrupt extinctionand subsequent reworking of Cretaceous planktonic foraminiferaacross the Cretaceous-Tertiary boundary Evidence from the sub-tropical North Atlantic in Catastrophic Events and Mass Extinc-tions Impacts and Beyond edited by Koeberl C and MacLeodK G Geol S Am S 356 277ndash289 Boulder Colorado USA2002
Hull P M Norris R D Bralower T J and Schueth J D A rolefor chance in marine recovery from the end-Cretaceous extinc-tion Nat Geosci 4 856ndash860 doi101038ngeo1302 2011
Hsuuml K J and McKenzie J A A ldquoStrangeloverdquo ocean in the earli-est Tertiary in Natural variations Archean to present edited bySundquist E T and Broecker W S The carbon cycle and atmo-spheric CO2 American Geophysical Union Washington DCUSA 487ndash492 1985
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
J Vellekoop et al Ecological response to KndashPg boundary mass extinction 899
Jiao N Herndl G J Hansell D A Benner R Kattner G Wil-helm S W Kirchman D L Weinbauer M G Luo T ChenF and Azam F Microbial production of recalcitrant dissolvedorganic matter long-term carbon storage in the global oceanNat Rev 8 593ndash599 doi101038nrmicro2386 2010
Jorissen F J de Stigter H C and Widmark J G V A concep-tual model explaining benthic foraminiferal microhabitats MarMicropaleontol 26 3ndash15 doi1010160377-8398(95)00047-X1995
Jorissen F J Fontanier C and Thomas E Paleoceanographi-cal proxies based on deep-sea benthic foraminiferal assemblagecharacteristics in Proxies in Late Cenozoic PaleoceanographyPt 2 Biological tracers and biomarkers edited by Hillaire-Marcel C and de Vernal A 263ndash326 Elsevier Amsterdamthe Netherlands 2007
Kaiho K Kajiwara Y Tazaki K Ueshima M Takeda NKawahata H Arinobu T Ishiwatari R Hirai A and Lam-olda M A Oceanic primary productivity and dissolved oxygenlevels at the CretaceousTertiary boundary their decrease sub-sequent warming and recovery Paleoceanography 14 511ndash5241999
Keller G and Lindinger M Stable isotope TOC and CaCO3record across the CretaceousTertiary boundary at El KefTunisia Palaeogeogr Palaeocl 73 243ndash265 1989
Kellough G R Paleoecology of the Foraminiferida of the WillsPoint Formation (Midway Group) in northeast Texas Trans GulfCoast Assoc Geol Soc 15 73ndash153 1965
Kring D A The Chicxulub impact event and its environmentalconsequences at the CretaceousndashTertiary boundary PalaeogeogrPalaeocl 255 4ndash21 2007
Kump L R Interpreting carbon-isotope excursions Strangeloveoceans Geology 19 299ndash302 1991
Machalski M Vellekoop J Dubicka Z Peryt D and Ha-rasimiuk M Late Maastrichtian cephalopods dinoflagellatecysts and foraminifera from the CretaceousndashPaleogene suc-cession at Lechoacutewka southeast Poland Stratigraphic andenvironmental implications Cretaceous Res 57 208ndash227doi101016jcretres201508012 2016
Molina E Evidence and causes of the main extinctionevents in the Paleogene based on extinction and survivalpatterns of foraminifera Earth-Sci Rev 140 166ndash181doi101016jearscirev201411008 2015
Molina E Alegret L Arenillas I and Arz J The GlobalBoundary Stratotype Section and Point for the base of the Da-nian Stage (Paleocene Paleogeneldquo Tertiaryrdquo Cenozoic) at ElKef Tunisia-Original definition and revision Episodes 29 263ndash273 2006
Olsson R K Liu C and van Fossen M The Cretaceous-Tertiarycatastrophic event at Millers Ferry Alabama in The Cretaceous-Tertiary Event and Other Catastrophes in Earth History editedby Ryder G Fastovsky D and Gartner S Geol S Am S307 263ndash277 1996
Peryt D Benthic foraminiferal response to the Cenomanian-Turonian and CretaceousndashPaleogene boundary events PrzegladGeol 52 827ndash832 2004
Peryt D Alegret L and Molina E The CretaceousPalaeogene(KP) boundary at Aiumln Settara Tunisia restructuring of benthicforaminiferal assemblages Terra Nova 14 101ndash107 2002
Ramette A Multivariate analyses in microbial ecology FEMS Mi-crobiol Ecol 62 142ndash160 2007
Rodriacuteguez-Tovar F J Uchman A MrsquoHamdi A Riahi Sand Ismail-Lattrache K B Ichnological record of palaeoen-vironment from the CretaceousndashPaleogene boundary intervalat El Kef Tunisia The first study of old and new sec-tions at the stratotype area J Afr Earth Sci 120 23ndash30doi101016jjafrearsci201604017 2016
Saner S Mudurnu-Goumlynuumlk Havzasının Jura ve Sonrasıccediloumlkelimnitelikleriyle paleocografya yorumlaması TJK Buumllteni 23 39ndash52 1980
Schioslashler P Brinkhuis H Roncaglia L and Wilson G J Di-noflagellate biostratigraphy and sequence stratigraphy of theType Maastrichtian (Upper Cretaceous) ENCI Quarry TheNetherlands Mar Micropaleontol 31 65ndash95 1997
Schueth J D Bralower T J Jiang S and Patzkowsky ME The role of regional survivor incumbency in the evolu-tionary recovery of calcareous nannoplankton from the Creta-ceousPaleogene (KPg) mass extinction Paleobiology 41 661ndash679 doi101017pab201528 2015
Scotese C R A continental drift flipbook J Geology 112 729ndash741 2004
Scotese C R and Dreher C GlobalGeology available at httpwwwGlobalGeologycom (last access 11 March 2014) 2012
Sepkoski J J Patterns of Phanerozoic extinction a perspectivefrom global data bases in Global Events and Event Stratigra-phy edited by Walliser O H Springer Berlin Germany 35ndash52 2016
Sibert E C Hull P M and Norris R D Resilience of Pacificpelagic fish across the CretaceousPalaeogene mass extinctionNat Geosci 7 667ndash670 doi101038NGEO2227 2014
Slimani H and Toufiq A A CretaceousndashPaleogene boundary geo-logical site revealed by planktic foraminifera and dinoflagellatecysts at Ouled Haddou eastern external Rif Chain Morocco JAfr Earth Sci 88 38ndash52 doi101016jjafrearsci2013080082013
Slimani H Louwye S and Toufiq A Dinoflagellate cystsform the CretaceousndashPaleogene boundary at Ouled Had-dou southeastern Rif Morocco biostratigraphy paleoenvi-ronments and paleobiogeography Palynology 34 90ndash124doi10108001916121003629933 2010
Sluijs A and Brinkhuis H A dynamic climate and ecosystemstate during the Paleocene-Eocene Thermal Maximum infer-ences from dinoflagellate cyst assemblages on the New JerseyShelf Biogeosciences 6 1755ndash1781 doi105194bg-6-1755-2009 2009
Smit J Extinction and evolution of planktonic foraminifera after amajor impact at the CretaceousTertiary boundary Geol S AmS 190 329ndash352 1982
Smit J and Romein A J T A sequence of events across theCretaceous-Tertiary boundary Earth Planet Sci Lett 74 155ndash170 1985
Speijer R P Extinction and recovery patterns in ben-thic foraminiferal paleocommunities across the Creta-ceousPaleogene and PaleoceneEocene boundaries GeologicaUltraiectina 124 Faculteit Aardwetenschappen PhD ThesisUniversiteit Utrecht Utrecht the Netherlands 1994
Speijer R P and van Der Zwaan G J Extinction and survivor-ship of southern Tethyan benthic foraminifera across the Cre-
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
900 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
taceousPaleogene boundary in Biotic recovery from mass ex-tinction events edited by Hart M B Geolog Soc Spec PublLondon UK 102 343ndash371 1996
Stemmann L and Boss E Plankton and Particle Size andPackaging From Determining Optical Properties to Drivingthe Biological Pump Annu Rev Mar Sci 4 263ndash290doi101146annurev-marine-120710-100853 2012
Thomas E Late Cretaceous-early Eocene mass extinctions in thedeep sea Geol S Am S 247 481ndash495 1990
Vandenberghe N Hilgen F J and Speijer R P The PaleogenePeriod in The Geologic Time Scale 2012 edited by GradsteinF M Ogg J G Schmitz M D and Ogg G M 2 855ndash921Elsevier Boston USA 2012
Vellekoop J Sluijs A Smit J Schouten S Weijers J WH Sinninghe Damsteacute J S and Brinkhuis H Rapid short-term cooling following the Chicxulub impact at the CretaceousndashPaleogene boundary P Natl Acad Sci USA 111 7537ndash7541doi101073pnas1319253111 2014
Vellekoop J Smit J van de Schootbrugge B Weijers J W HGaleotti S Sinninghe Damsteacute J S and Brinkhuis H Palyno-logical evidence for prolonged cooling along the Tunisian conti-nental shelf following the KndashPg boundary impact PalaeogeogrPalaeocl 426 216ndash228 doi101016jpalaeo2015030212015
Vellekoop J Esmeray-Senlet S Miller K G Browning J VSluijs A van de Schootbrugge B Sinninghe Damsteacute J Sand Brinkhuis H Evidence for CretaceousndashPaleogene bound-ary bolide ldquoimpact winterrdquo conditions from New Jersey USAGeology 44 619ndash622 doi101130G379611 2016
Woelders L and Speijer R Stable seafloor conditions sealevel and food supply during the latest Maastrichtian atBrazos River Texas Mar Micropaleontol 121 41ndash51doi101016jmarmicro201510002 2015
Zachos J C Arthur M A and Dean W E Geochemical evi-dence for suppression of pelagic marine productivity at the Cre-taceousTertiary boundary Nature 337 61ndash64 1989
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
- Abstract
- Introduction
- Geological setting and age assessment
- Materials and methods
-
- Sampling
- Foraminiferal analysis
- Palynological analysis
- Statistical analysis
-
- Results
-
- Benthic foraminifera
- Palynology
-
- Discussion
-
- Tethyan benthic foraminiferal turnover sequence
- Ecological responses to reduced export productivity
-
- Okccedilular section
- Western Tethys
- Global responses
-
- Long-term recovery
-
- Conclusions
- Data availability
- Author contributions
- Competing interests
- Acknowledgements
- References
-

888 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
clay layer Throughout the basin the lower 30ndash50 m of theDanian is characterized by a rhythmic alternation of fine-grained limestones and carbonate-rich mudstones (Accedilikalinet al 2015) During the latest Cretaceous to early Paleocenethis basin was characterized by mixed siliciclasticndashcarbonatesedimentation in an outer neritic to upper bathyal environ-ment (Accedilikalin et al 2015)
The Okccedilular section has been analyzed for siderophiletrace elements including Ir and other platinum groupelements (PGEs) bulk stable carbon isotopes plankticforaminifera calcareous nannofossils and dinocysts Basedon these results a detailed stratigraphic framework was out-lined (Accedilikalin et al 2015) allowing a confident age assess-ment of the boundary interval The age model shows thatthe section contains a chronostratigraphically complete KndashPg boundary interval The studied interval ranges from thetop part of the Maastrichtian A mayaroensis Zone up to thebasal part of the Danian planktic foraminiferal Zone P1b andcovers globally occurring first occurrences (FO) of dinocystmarker taxa such as Senoniasphaera inornata Damassa-dinium californicum and Carpatela cornuta
3 Materials and methods
31 Sampling
High-resolution (centimeter-scale) sample sets were ac-quired during two field campaigns in 2010 and in 2011 Formore detail on these sampling campaigns see Accedilikalin etal (2015) The samples were split for micropaleontologicaland palynological analyses
32 Foraminiferal analysis
Twenty-seven samples were processed at KU Leuven forforaminiferal studies following standard micropaleontolog-ical procedures Of these 20 samples were used for quan-titative benthic foraminiferal analyses (Fig 2) Rock sam-ples were dried in an oven at 60 C for at least 24 h De-pending on sample size 4 to 60 g of dry rock was soakedin a soda (Na2CO3) solution (50 g Lminus1) After disintegrationsamples were washed over 2 mm and 63 microm sieves If neces-sary the tenside Rewoquat was used to clean the residues andthe procedure was repeated Clean residues were dry-sievedinto three fractions 63ndash125 125ndash630 and gt 630 microm In orderto enable a direct comparison with the benthic foraminiferalrecord of the KndashPg boundary GSSP (Global Boundary Stra-totype Section and Point) of El Kef (Tunisia) we used thesame size fraction (gt 125 microm) for Okccedilular as for El Kef(Speijer and van der Zwaan 1996) Note that El Kef holdsthe GSSP of the base of the Danian (Molina et al 2006)and is thus of particular importance with respect to corre-lation and comparison of the KndashPg boundary Representativealiquots of the gt 125 microm fraction were obtained containing atleast 300 benthic foraminiferal specimens Picked specimens
from this size fraction were permanently stored in Plummerslides Benthic foraminifera were identified using the tax-onomy of Cushman (1946 1951) Kellough (1965) Aubertand Berggren (1976) Berggren and Aubert (1975) and Spei-jer (1994)
Benthic foraminifera are commonly used as indicators forbottom water oxygenation and trophic conditions (eg Joris-sen et al 2007) Here the benthic foraminiferal accumula-tion rate (BFAR number of foraminifera per cm2 per kyrsee Text S2 in the Supplement for details on the estimationof BFAR) was calculated as a semi-quantitative proxy for pa-leoproductivity (Jorissen et al 2007 and references therein)In addition the percentage of endobenthic morphotypes wascalculated using the assumed microhabitat preferences in-ferred from benthic foraminiferal morphotype analysis (egCorliss 1985 Corliss and Chen 1988 Alegret et al 2003Woelders and Speijer 2015) In general endobenthic formsare considered indicative of a high flux of organic matter tothe sea floor andor relatively low oxygen conditions whileabundance of epibenthic forms is considered characteristicfor more oligotrophic open marine environments (Jorissen etal 1995 Peryt 2004 Jorissen et al 2007) Furthermore thebi- and triserial endobenthic forms are particularly indica-tive of high food supply and low oxygenation (eg Bern-hard 1986 Corliss and Chen 1988 Jorissen et al 2007)Therefore in this study the percentage of endobenthic formswas calculated for each sample as well as the percentage bi-triserial benthic taxa to unravel food supply and oxygena-tion patterns
It should however be noted that assuming such an ana-logue with modern fauna has limitations and shortcomingsFor instance the percentage of endobenthics based on mor-photypes and using bi-triserials as an indicator for hypoxiaand high food supply is not universally applicable (Buzas etal 1993 Jorissen et al 2007 and references therein)
33 Palynological analysis
In this study the palynological data from Accedilikalin etal (2015) were analyzed and interpreted In addition twoadditional samples were analyzed (OK 15 and OK 6) toincrease the resolution of the dataset and two samples ofthe Accedilikalin et al (2015) dataset were re-counted (OK 25and OK 250 see Dataset 2 in the Supplement) Quantitativeslides of the 15ndash250 microm fraction were used All slides arestored in the collection of the Laboratory of Palaeobotanyand Palynology Utrecht University the Netherlands
To identify major changes in the dinocyst record morpho-logically closely related taxa were grouped into complexesusing a similar approach to Schioslashler et al (1997) Sluijsand Brinkhuis (2009) and Machalski et al (2016) In ourstudy the following morphological complexes were estab-lished (1) the Spiniferites complex combining all species ofSpiniferites and the morphologically similar genus Achomop-shaera (2) Manumiella spp grouping all species of Man-
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
J Vellekoop et al Ecological response to KndashPg boundary mass extinction 889
Figure 2 The benthic foraminiferal record (gt 125 microm) of the Okccedilular section The biostratigraphy is from Accedilikalin et al (2015) The clusteranalysis on the benthic foraminiferal data using paired group (UPGMA) correlation distance allows the identification of four main clustersAndashD of benthic foraminiferal taxa
umiella (3) hexaperidinioids lumping all other peridinioidcysts with a hexaform archeopyle and (4) other dinocystswhich include all other dinocyst taxa and unidentifiabledinocysts (Fig 3)
Of these different dinocyst groups previous studies haveshown that in the Tethys the hexaperidinioids in particularshow strong variations across the KndashPg boundary (Brinkhuiset al 1998 Vellekoop et al 2015) Based on statistical cor-relations between palynological records and other paleoprox-ies it has been suggested that this inferred heterotrophicgroup flourished best under high-nutrient conditions in thephotic zone (Eshet et al 1994 Brinkhuis et al 1998 Sluijsand Brinkhuis 2009) Therefore in this study abundances ofhexaperidinioids are considered indicative of nutrient avail-ability in the photic zone (see Fig S1 and Text S1 in theSupplement for a more detailed discussion on this matter)
34 Statistical analysis
To assess changes in diversity of benthic foraminifera anddinocysts across the studied interval the Shannon diversityindex (H ) the species richness per sample (S) the number ofspecimens observed per sample (N ) and the BergerndashParkerindex were calculated for both biological groups followingHayek and Buzas (2013) (Fig 4) In addition in order torecognize the main faunal associations within the benthicforaminiferal data a cluster analysis was performed usingpaired group (UPGMA) correlation distance
Q-mode non-metric multidimensional scaling (NM-MDS)was performed on the benthic foraminiferal sample compo-sitions to assess patterns in assemblage response to KndashPgboundary perturbations Since taxa may not have a linear re-sponse to environmental changes across the KndashPg boundaryQ-mode NM-MDS is preferred over PCA and CCA (Ram-ette 2007)
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
890 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
Figure 3 The organic-walled dinoflagellate cyst record of theOkccedilular section The biostratigraphy and bulk carbonate stable car-bon isotope records are from Accedilikalin et al (2015) The four maindinocyst complexes are indicated
4 Results
41 Benthic foraminifera
The benthic foraminiferal record of the Okccedilular section ischaracterized by a distinct turnover across the KndashPg bound-ary (Figs 2 4 5 6) Of the common taxa 8 out of 30(sim 27 ) disappear across the KndashPg boundary After the KndashPg boundary crisis three new taxa appear The estimatedBFAR shows a major decrease across the boundary (Fig 5)For the foraminiferal counts and illustrations of commonforms see Dataset S1 and Figs S2 S3
The cluster analysis allows the identification of fourmain clusters of benthic foraminiferal taxa clusters A toD (Fig 2) Cluster C is relatively large and can be subdi-vided into five subclusters Based on the succession of ben-thic faunal assemblages characterized by strong changes inthe Shannon diversity index (H ) four intervals can be rec-ognized in the benthic foraminiferal record of the Okccedilularsection (Figs 4 6)
The first interval comprises the uppermost Maastrichtianand is characterized by relatively high diversity dominatedby the taxa in cluster D In this assemblage the bi-triserialbenthic taxa are relatively abundant (22ndash28 ) with char-acteristic taxa such as Bulimina arkadelphiana Eouvigerinasubsculptura and Praebulimina reussi About 50ndash60 of theassemblage consists of inferred endobenthic taxa The KndashPg boundary marks an abrupt benthic community impover-
ishment and the decimation of the taxa in cluster D Abovethe boundary the bi-triserial benthic taxa virtually disappearfrom the record (mostly below lt 05 )
The second interval characterized by low diversity com-prises the lowermost Danian approximately correspondingto planktic foraminiferal Zone P0 In this interval epibenthicforms are most abundant (70ndash90 ) It is dominated by thetaxa of cluster A encompassing successive peak occurrencesof the taxa Anomalinoides praeacutus Trochammina sppand Cibicidoides pseudoacutus
The third interval covers the part of the succession approx-imately correlative to planktic foraminiferal Zone Pα Thisinterval is characterized by a recovery of diversity althoughthe diversity is still lower than that of the top Maastrichtianassemblage The abundance of taxa in cluster A slowly de-creases and taxa in cluster B mainly represented by Osan-gularia plummerae and Cibicidoides sp become abundantEndobenthic forms recover and make up 30ndash40 of the totalassemblage
The fourth interval starts in the interval correlative to ZoneP1a Here the benthic community has recovered as the di-versity has stabilized and is almost similar to pre-impact val-ues This interval is characterized by taxa in cluster C a typ-ical Paleocene Midway-type fauna (Berggren and Aubert1975) with representatives such as Anomalinoides praea-cutus Coryphostoma midwayensis Cibicidoides alleni andOsangularia plummerae
42 Palynology
Palynological samples from the Okccedilular site yield abundantpalynomorphs dominated by dinocysts and with minor con-tributions of acritarchs prasinophytes organic foraminiferallinings and terrestrial palynomorphs (Dataset S2) Thedinocyst associations of the MudurnundashGoumlynuumlk basin arequite diverse including components characteristic of boththe Tethyan and Boreal realms (Accedilikalin et al 2015) As ex-pected the dinocyst record does not show major changes indiversity across the KndashPg boundary (Fig 4) There is a steadydecrease in dinocyst diversity from planktic foraminiferalZone P1a upwards probably a long-term change not re-lated to the impact The estimated dinocyst accumulationrate (DAR number of preserved cysts per cm2 per kyr seeText S2 in the Supplement for details on the estimation of theDAR) shows no major changes across the KndashPg boundary
Throughout the palynological record the Spiniferites com-plex is consistently the most dominant morphogroup in gen-eral comprising 40ndash50 of the total assemblage (Fig 3)Similar to other KndashPg boundary sites worldwide (Habib andSaeedi 2007) Manumiella spp show an episode of higherrelative abundance near the top of the Maastrichtian some30ndash40 cm below the KndashPg boundary Hexaperidinioids gen-erally make up a relatively small component of the Maas-trichtian assemblage (3ndash17 ) but show a strong increaseacross the KndashPg boundary In the boundary clay layer cor-
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
J Vellekoop et al Ecological response to KndashPg boundary mass extinction 891
Figure 4 Diversity indices on the benthic foraminiferal and dinocyst records of the Okccedilular section with the Shannon diversity index (H ) thespecies richness per sample (S) the number of specimen observed per sample (N ) and the BergerndashParker index As the bulk stable carbonisotope record and the Shannon diversity index of the benthic foraminiferal record show similar trends the combination of these records canbe used to subdivide the KndashPg boundary transition into four intervals I the Maastrichtian ie ldquopre-impactrdquo interval II the post-impactinterval representing the ldquodisasterrdquo phase III a recovery interval and IV the early Paleocene post-recovery interval The division criteriaare explained in the text
relative to planktic foraminiferal Zone P0 this group in-creases to 35 of the assemblage In the peak intervals thehexaperidinioids are mostly represented by representativesof the genera Senegalinium and Cerodinium After an ini-tial drop in relative abundance in the upper half of Zone P0this group reaches a second maximum (sim 35 ) at the baseof Zone Pα and remains relatively abundant (15ndash30 ) up toZone P1a After this the hexaperidinioids gradually decreasetowards the top of the studied interval The ldquoother dinocystsrdquogroup ndash with representatives such as Areoligera spp Im-pagidinium spp Hystrichosphaeridium tubiferum Opercu-lodinium centrocarpum and Palynodinium grallator ndash gen-erally makes up 25ndash50 of the assemblage
5 Discussion
51 Tethyan benthic foraminiferal turnover sequence
The major turnover in the benthic community at Okccedilularis largely comparable with the previously published ben-thic foraminiferal record (size fraction gt 125 microm) of the KndashPg boundary GSSP of El Kef (Figs 7 8 Speijer and Vander Zwaan 1996) In addition other benthic foraminiferalrecords from the margins of the Tethys also show a strong
similarity to the Okccedilular and El Kef records including stud-ies that use the gt 63 microm size fraction (Alegret et al 2003Culver 2003 and references therein)
Although it is sometimes suggested that different sizefractions cannot be directly compared because the differ-ent size fractions sometimes harbor different ecological in-formation (Giusberti et al 2016) intercomparison studieshave shown that when comparing the gt 125 and gt 63 microm sizefractions in benthic foraminiferal assemblages the overallpatterns are generally robust (Ernst et al 2006) The factthat gt 63 and gt 125 microm KndashPg boundary benthic foraminiferalrecords from different Tethys-margin sites (eg Speijer andVan der Zwaan 1996 Peryt et al 2002 Culver et al 2003)show striking similarities even though both the size fractionsused (gt 63 vs gt 125 microm) and specific taxa making up theforaminiferal assemblages differ among sites confirms theobservations by Ernst et al (2006) Furthermore even whenit would be assumed that different size fractions are diffi-cult to compare directly the striking similarities between theTethyan gt 125 and gt 63 microm size fraction KndashPg boundary ben-thic foraminiferal records provide compelling evidence for astrong disruption of the entire benthic community
Tethyan middle neritic to middle bathyal KndashPg boundarysuccessions generally show a sequence of (1) a typical high
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
892 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
Figure 5 Estimated benthic foraminiferal accumulation rate (BFAR number of foraminifera of the gt 63 microm size fraction per cm2 sea floorper kyr) and estimated dinocyst accumulation rate (DAR number of preserved cysts per cm2 sea floor per kyr) of the Okccedilular sectionUncertainty in estimated accumulation rates resulting from uncertainties in sediment densities and sedimentation rates are indicated in greyproviding a range of estimated accumulation rates For more detailed info on the calculation of the BFAR and DAR see Text S2 of theSupplement
diversity assemblage in the Maastrichtian (2) a low diver-sity ldquodisasterrdquo assemblage directly after the KndashPg boundaryimpact characterized by a short-lived bloom of endobenthicforms observed in several high-resolution records (eg Coc-cioni and Galeotti 1994 Speijer and van der Zwaan 1996)followed by a dominance of epibenthic forms (3) a ldquore-coveryrdquo assemblage characterized by an increasing diversityand returning endobenthic forms and (4) a new high diver-sity assemblage dominated by a Paleocene fauna with bothepi- and endobenthic forms equally represented
Hence based on this succession of benthic assemblageswhich are characterized by strong changes in diversity (H )the KndashPg boundary benthic foraminiferal records from theTethys can be subdivided into four intervals (Figs 2 4ndash6)The intervals I to IV approximately correspond to the up-permost Maastrichtian planktic foraminiferal Zone P0 sub-zones PαndashP1a and subzones P1andashP1b respectively There-fore this succession roughly follows the 5-fold sedimen-tological and planktic foraminiferal sequence of Smit andRomein (1985) in which unit 2 represents the ejecta layerand units 1 3 4 and 5 correspond to intervals I to IV in ourstudy
The sequence of benthic foraminiferal turnover in theTethys is also well illustrated by a Q-mode NM-MDS
(Fig 7) The Q-mode NM-MDS of the benthic foraminiferalsample compositions of the combined data from Okccedilular(this study) and El Kef (Speijer and van der Zwaan 1996)demonstrates that although the assemblages differ betweenthese localities on opposite margins of the Tethys the re-sponses of the benthic foraminiferal assemblages to the KndashPgboundary perturbations express very similar overall patternsacross the KndashPg boundary At both Okccedilular and El Kef thebenthic foraminiferal records show a rapid transition from astable Maastrichtian assemblage to the earliest Danian dis-aster phase in Zone P0 followed by a gradual change backtowards conditions similar to the Maastrichtian (marked bythe return of Lazarus taxa) Therefore both the successionof benthic faunal assemblages including diversity indicesand Q-mode NM-MDS analysis provide powerful tools to al-low the comparison of KndashPg boundary benthic foraminiferalrecords from different localities
52 Ecological responses to reduced export productivity
521 Okccedilular section
The palynological benthic foraminiferal and bulk stable iso-tope records of the Okccedilular section show major changesacross the KndashPg boundary interval (Figs 2ndash7) portraying
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
J Vellekoop et al Ecological response to KndashPg boundary mass extinction 893
Figure 6 Relative abundances of the four main clusters of benthic foraminiferal taxa (gt 125 microm) relative abundances of epibenthic andendobenthic forms and relative abundance of bi-triserial benthic taxa
Figure 7 Q-mode non-metric multidimensional scaling (on all sam-ples in the gt 125 microm size fraction benthic foraminiferal datasets ofOkccedilular (this study) and El Kef (Speijer and van der Zwaan 1996)After an abrupt transition from latest Maastrichtian pre-impact as-semblages to disaster assemblages in Zone P0 and recovery assem-blages in Zone Pα a gradual transition towards a new equilibriumpost-recovery assemblage can be observed in subzones P1a and P1b
the biological crisis following the impact Following the KndashPg boundary mass extinction endobenthic forms stronglydecrease with bi-triserial benthic taxa almost disappearingfrom the benthic community simultaneously the estimatedBFAR shows a major fall (Fig 5) This interval approxi-mately correlative to foraminiferal Zone P0 represents thedisaster phase The drop in bi-triserial benthic taxa (Fig 6)and decrease in BFAR (Fig 5) point to a reduction in foodsupply to the benthic community indicating that the trans-port of organic matter to the sea floor must have decreasedIn the dinocyst community however hexaperidinioids showa strong increase in relative abundance during this phase(Figs 3 6 7) suggesting an increase in nutrient availabilityin the photic zone while the DAR remains relatively stableacross the KndashPg boundary (Fig 5)
The decrease in benthic diversity and the high relativeabundances of opportunistic Trochammina spp and Cibici-doides pseudoacutus suggest that the benthic community ex-perienced additional stress during Zone P0 besides food lim-itation (see Jorissen et al 2007) If food limitation were theonly factor involved upper bathyal assemblages as observedat Okccedilular would be expected to become more similar tooligotrophic middle to lower bathyal assemblages Insteadthe opposite is true the assemblage composition of the dis-aster phase is closer to neritic Midway-type assemblages
It was previously suggested that the epibenthic taxon Cibi-cidoides pseudoacutus could potentially be hypoxia resistant
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
894 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
(Speijer and van der Zwaan 1996) As a collapse of the bi-ological pump should lead to enhanced remineralization oforganic matter in the photic zone it is indeed likely thatthe oxygen minimum zones expanded and shoaled leadingto oxygen stress on the sea floor Although early Paleocenedysoxia at El Kef is supported by the ostracod record (Donzeet al 1982) and ichnofacies analyses (Rodriguez-Tovaret al 2016) supporting (eg geochemical) evidence fordysoxia during the disaster phase at Okccedilular record is absentFurthermore an additional stressor for the benthic commu-nity during Zone P0 may be present In contrast to calcare-ous nanoplankton organic-walled cyst-producing dinoflagel-lates did not suffer extinctions and may have become a moreimportant component in the early Paleocene phytoplanktoncommunity Hence besides the reduction of the amount oforganic matter transported to the sea floor the compositionof food supplied by the photic zone likely changed signifi-cantly across the KndashPg boundary as well (DrsquoHondt 2005)possibly presenting an additional stress factor for the benthiccommunity (Speijer and van der Zwaan 1996 Alegret andThomas 2009)
522 Western Tethys
The patterns in benthic foraminiferal and dinoflagellate re-sponse to the KndashPg boundary perturbations at Okccedilular appearto be characteristic for the continental margins of the westernTethyan realm The benthic foraminiferal patterns are similaracross different size fractions suggesting that they are repre-sentative of the overall benthic communities In Tunisia andSpain where both benthic foraminiferal and dinoflagellaterecords are available the earliest Danian is also characterizedby a decrease in endobenthic foraminifera at the sea floor(Speijer and Van der Zwaan 1996 Peryt et al 2002 Ale-gret et al 2003) and simultaneously blooms of hexaperidin-ioids in the water column (Brinkhuis et al 1998 Vellekoopet al 2015 Fig 8) In the Ouled Haddou section Moroccoin the westernmost Tethys (Fig 1) the lowermost Danian isalso characterized by a strong increase of the hexaperidin-ioid Senegalinium group up to 30 of the assemblage (Sli-mani et al 2010) that is very similar to the Okccedilular record(Fig 8)
The KndashPg boundary dinocyst record of El Kef is strikinglysimilar to the one from the Okccedilular section showing two dis-tinct peaks in hexaperidinioids one in Zone P0 and one atthe basal part of Zone Pα At El Kef the initial post-impactdominance of hexaperidinioids is however less pronounced(Fig 8) This difference in expression might be related tosmall differences in paleogeographic and paleoceanographicsettings between sites
In addition some high-resolution benthic KndashPg records inthe Tethyan realm show a short-lived bloom of specific en-dobenthic foraminiferal taxa in the few centimeter directlyabove the KndashPg boundary sometimes accompanied by otherindicators of low oxygen levels (eg Coccioni and Gale-
otti 1994 Speijer and Van der Zwaan 1996 Kaiho et al1999) This suggests that at least locally the sea floor wastemporarily characterized by hypoxic conditions directly fol-lowing the impact This short-lived deoxygenation is possi-bly related to the mass mortality at the KndashPg boundary re-sulting in a large short-lived flux of organic matter to thesea floor (Coccioni and Galeotti 1994) However since ex-port productivity was greatly reduced after the KndashPg bound-ary mass extinction very little ldquonewrdquo organic matter reachedthe sea floor after the initial post-impact influx Hence afteraccumulated organic matter was remineralized the benthiccommunity starved resulting in a transition to an epibenthic-dominated benthic fauna similar to the post-impact fauna ofthe Okccedilular record
The combined dinocyst records suggest that the early Pa-leocene margins of the Tethys were characterized by an in-crease in nutrient availability in the photic zone whereascoeval benthic foraminiferal records indicate a major de-crease in nutrient supply to the sea floor These changesin the benthic foraminiferal and dinocyst communities arelikely caused by the major reduction of both the efficiencyand strength of the biological pump (ie the fraction ofprimary production exported from the photic zone and theamount of organic matter transported down) in accordancewith the Living Ocean model (DrsquoHondt and Zachos 1998DrsquoHondt et al 1998 DrsquoHondt 2005 Coxall et al 2006)For a detailed explanation of the difference between biologi-cal pump efficiency and biological pump strength see Hiltinget al (2008)
As bulk carbonate δ13C reflects the isotopic compositionof the surface ocean which is set by burial fractions and bythe photosynthetic isotope effect (Kump 1991 Hain et al2014) the excursion recorded in bulk δ13C records world-wide and the rapid collapse in surface to deep-ocean carbonisotope gradients likely reflect the reduction of the global in-tensity (ie efficiency) of the biological pump (Hain et al2014) The recorded inverse change in nutrient availabilitysuggests a causal link A reduced efficiency of the biologi-cal pump and associated decrease of the fraction of biomasstransported out of the photic zone could have resulted inhigh rates of nutrient recycling in the upper part of the wa-ter column (DrsquoHondt 2005) This suggests that the reduc-tion of the biological pump strength recorded by the benthicforaminiferal record is a consequence of the decreased effi-ciency of the biological pump recorded by the carbon iso-tope and dinoflagellate cyst records The strong correlationbetween the bulk carbon isotope curves and the Shannondiversity index (H ) of the benthic foraminiferal records atboth Okccedilular (R2
= 073 p lt 0001) and El Kef (R2= 074
p lt 0001) suggest a clear link between the changes in ben-thic foraminiferal assemblages and the collapse and recoveryof biological pump efficiency
As a result of the reduced biological pump efficiency morenutrients remained available for the early Paleocene phyto-plankton community At Tethyan margin sites this is indi-
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
J Vellekoop et al Ecological response to KndashPg boundary mass extinction 895
Figure 8 The benthic foraminiferal (gt 125 microm) dinocyst and bulk stable carbon isotope records of Okccedilular (Turkey) and El Kef (Tunisia)the dinocyst record of Ouled Haddou (Morocco) and the benthic foraminiferal record (gt 63 microm) of ODP Hole 1049C Blake Nose (NorthAtlantic) Note that the definition of ldquohexaperidinioidsrdquo used is provided in the text The four phases identified based on the bulk stablecarbon isotopes and the Shannon diversity index (H ) of the benthic foraminiferal records are indicated (IndashIV) The gt 63 microm size fractionbenthic foraminiferal record of ODP Hole 1049C is from Alegret and Thomas (2004) who argued that the lowermost Paleocene of thissite comprised reworked foraminifera This interval is indicated by a dashed bar in the figure The biostratigraphy of Ouled Haddou is fromSlimani and Toufiq (2013) whereas the dinocyst record of this site is from Slimani et al (2010) The biostratigraphy of El Kef is based onBrinkhuis et al (1998) Speijer and van der Zwaan (1996) and Molina et al (2006) The fine-fraction stable carbon isotope record (blackdots) of El Kef is from Keller and Lindinger (1989) The Shannon diversity index (red dots) relative abundances of bi-triserial benthic taxa(dark purple) and all endobenthic foraminifera (light purple) are based on the gt 125 microm size fraction foraminiferal data of Speijer and vander Zwaan (1996) The palynological data of El Kef are from Brinkhuis et al (1998) (lower part) and Guasti et al (2005) (upper part) TheShannon diversity index of the gt 125 microm size fraction benthic foraminiferal record (red dots) the relative contribution of bi-triserial benthictaxa (dark purple) all endobenthic foraminifera (light purple) and hexaperidinioid dinocysts from Okccedilular are from this study The bulkstable carbon isotope record of Okccedilular (black dots) is from Accedilikalin et al (2015) Mbsf indicates meters below sea floor
cated by the higher abundance of hexaperidinioids Similarlyblooms of eutrophic survivor taxa of calcareous nanoplank-ton at open ocean sites have been suggested to be relatedto the buildup of nutrients in the open ocean photic zone(Schueth et al 2015) Although the carbon isotope gradientsbetween surface and deep waters indicate that the recoveryof the biological pump took hundreds of thousands of years(Zachos et al 1989 Kump 1991 DrsquoHondt et al 1998 Cox-all et al 2006 Birch et al 2016) our records suggest thatthe enhanced recycling of nutrients in the Tethys was partic-ularly intense during two phases in the first tens of thousandsyears after the impact (Fig 8)
523 Global responses
The decrease in export productivity following the KndashPgboundary is not only recognized in the Tethyan realm In theAtlantic Ocean Southern Ocean and the Indian Ocean a de-crease of the biological pump strength after the KndashPg bound-ary was also observed in neritic to abyssal environments(eg Thomas 1990 Olsson et al 1996 Hull et al 2011Alegret et al 2012) As an example the gt 63 microm size frac-tion benthic foraminiferal KndashPg boundary record of BlakeNose (ODP Hole 1049C northwestern Atlantic Alegret andThomas 2004) shows a pattern that is strikingly similar tothe gt 125 microm size fraction records in the Tethyan realm witha strong decrease of endobenthics in Zone P0 followed by a
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
896 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
gradual recovery across zones Pα and P1a (Fig 8) The simi-larities between the benthic foraminiferal patterns of Tethyanouter neritic to middle bathyal (200ndash1500 m) sites and thoseof the middle to lower bathyal (1500ndash1600 m) record ofBlake Nose (Alegret and Thomas 2004) suggest that theserecords register a global decrease in export productivity fol-lowing the KndashPg boundary
However several open ocean sites mostly in the Pacificshow an opposite trend with several lines of evidence sug-gesting an increased biological pump strength at these sites(eg Hull et al 2011 Alegret et al 2012) This led Ale-gret et al (2012) to conclude that the decrease in biologicalpump strength after the KndashPg boundary was most likely aregional instead of a global effect arguing against the Liv-ing Ocean hypothesis We would argue that low-productivityopen ocean sites like the central Pacific are characterized byvery different ecosystem structures and higher productivityas those along the Tethyan margins (eg Dortch and Pack-erd 1989) Therefore the consequence of ecosystem reorga-nization resulting from the mass extinction was likely verydifferent in the open ocean Pacific (eg Sibert et al 2014)Esmeray-Senlet et al (2015) proposed the term ldquoHeteroge-neous Oceanrdquo for this conceptual model as an alternativeto the Living Ocean model Supposedly the HeterogeneousOcean was characterized by a strong geographic heterogene-ity in the extinction patterns and food supply to the sea floor
However as the primary food source for open ocean deep-sea benthic communities generally consists of refractory or-ganic matter (eg Jiao et al 2010) it remains uncertainwhether changes in the intensity of recycling of the gener-ally more labile organic matter in the surface ocean will re-sult in changes in the amount of organic matter that benthiccommunities receive as they will likely continue to receiverecalcitrant organic matter at barely diminished rates despitethe collapse of the biological pump
In addition it should be noted that although the bio-logical pump efficiency is generally relatively high in openocean realms such as the central Pacific the biological pumpstrength is generally very low at these sites (Honjo et al2008 Henson et al 2011) Small changes in absolute bio-logical pump strength at such a locality could therefore havehad a large effect on the local oligotrophic benthic commu-nity (Alegret and Thomas 2009) but they likely representedonly a small fraction of the net amount of organic matterglobally transported to the deep Hence even though localbiological pump strength might have increased at some low-productivity deep-sea sites the large decrease in biologi-cal pump strength recorded at many other sites means thatthe net amount of organic matter globally transported out ofthe surface ocean was likely still reduced in the post-impactworld (Birch et al 2016) Therefore we argue that such Pa-cific records do not represent the general pattern The globalocean response can be characterized as heterogeneous how-ever the KndashPg boundary mass extinction did lead to a strongreduction of the net amount of organic matter globally trans-
ported from the surface to the sea floor while biological pro-ductivity recovered rapidly after the extinction event in ac-cordance with the Living Ocean hypothesis
53 Long-term recovery
Following the disaster phase the hexaperidinioid cysts atOkccedilular and El Kef decreased in abundance and the abun-dance of endobenthic forms increased again as the recoveryof the benthic community was initiated This recovery phaseis approximately correlative to Zone Pα and the basal part ofZone P1a which according to the Paleogene age constraintsof Vandenberghe et al (2012) represents at least 300 kyrThis duration is in agreement with the estimations providedby Birch et al (2016) During this phase the carbon iso-tope records remain well below pre-impact values and thediversity of the benthic foraminiferal community has not yetfully recovered (Figs 4ndash8) This indicates that the rapid andshort-lived KndashPg boundary disaster was followed by a rela-tively long recovery phase in line with previous estimatesof a multimillion-year biological recovery (eg Coxall etal 2006) While the impact-related environmental perturba-tions were short lived (Kring 2007 Vellekoop et al 20142016) the extinctions amongst important biological groupsled to a reduction of the organic flux from the photic zone todeep water resulting in major long-term biological and pa-leoceanographic reorganizations Only with the evolutionaryrecovery of the pelagic community governing the biologicalcarbon pump did export productivity start to increase again(eg Coxall et al 2006 Birch et al 2016)
6 Conclusions
The marine palynological benthic foraminiferal and bulkstable isotope records of the Okccedilular and El Kef sections re-veal major changes across the KndashPg boundary interval por-traying the biological crisis at the KndashPg boundary and subse-quent recovery in the early Paleocene Based on the succes-sion of benthic faunal assemblages at a number of Tethyanmargin sites four phases can be recognized across the KndashPgboundary interval the Maastrichtian or pre-impact phase (I)a disaster phase (II) a recovery phase (III) and an early Pa-leocene post-recovery phase (IV)
Following the KndashPg boundary impact some localitiesshow a large short-lived flux of food to the sea floor likelyrelated to the mass mortality at the KndashPg boundary Howeversince export productivity was greatly reduced after the KndashPgboundary mass extinction it is likely that after the initialpost-impact influx very little fresh organic matter reachedthe sea floor This reduction of export productivity in thepost-extinction disaster phase eventually resulted in a lowerfood supply to the sea floor This presented a major stressfactor for benthic organisms in the western Tethys The re-duced food supply resulted in an abrupt impoverishment of
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
J Vellekoop et al Ecological response to KndashPg boundary mass extinction 897
benthic communities As the downward transport of nutri-ents was slowed down recycling in upper layers increasedAs a result more nutrients remained available for the earlyPaleocene phytoplankton community leading to blooms ofdinoflagellates along the Tethyan margins and blooms of cal-careous nanoplankton taxa in more open ocean sites Ourrecords show that the enhanced recycling of nutrients in theTethys was particularly intense in the tens of thousands ofyears after the impact Following this the slow evolution-ary recovery of the pelagic community governing the bio-logical carbon pump resulted in a gradually increasing ex-port productivity in the hundreds of thousands of years afterthe impact Hence our integration of dinocyst and benthicforaminiferal records across the KndashPg boundary provides im-portant new insights into the ecological responses to the re-duction of export productivity following the mass extinctionat the CretaceousndashPaleogene boundary highlighting the di-rect ecological consequences of the Living Ocean conditionsin the post-impact world
7 Data availability
All data related to publication are available in the Supple-ment
The Supplement related to this article is available onlineat doi105194bg-14-885-2017-supplement
Author contributions Johan Vellekoop Lineke Woelders andHenk Brinkhuis designed the research Samples were collectedin the field by Johan Vellekoop Sanem Accedilikalin Ismail Ouml Yil-maz and Jan Smit Palynological analyses were carried out by Jo-han Vellekoop Benthic foraminiferal analyses were carried out byLineke Woelders and Robert P Speijer Johan Vellekoop LinekeWoelders and Bas van de Schootbrugge prepared the manuscriptwith input from all authors
Competing interests The authors declare that they have no conflictof interest
Acknowledgements This work was supported by the NetherlandsOrganization for Scientific Research (NWO) Open CompetitionGrant ALWPJ09047 to H Brinkhuis and the Research FoundationFlanders (FWO) grant G0B8513 to R P Speijer We thank LKump F Boscolo Galazzo P Roopnarine and one anonymousreferee for their suggestions
Edited by D GillikinReviewed by L Kump F Boscolo Galazzo and P Roopnarine
References
Accedilikalin S Vellekoop J Ocakoglu F Yılmaz I Ouml Smit J Al-tiner S O Goderis S Vonhof H Speijer R P WoeldersL Fornaciari E and Brinkhuis H Geochemical and pale-ontological characterization of a new KndashPg Boundary local-ity from the Northern branch of the Neo-Tethys MudurnundashGoumlynuumlk Basin NW Turkey Cretaceous Res 52 251ndash267doi101016jcretres201407011 2015
Alegret L and Thomas E Food supply to the seafloor in the Pa-cific Ocean after the CretaceousPaleogene boundary event MarMicropaleontol 73 105ndash116 2009
Alegret L Molina E and Thomas E Benthic foraminiferalturnover across the CretaceousPaleogene boundary at Agost(southeastern Spain) paleoenvironmental inferences Mar Mi-cropaleontol 48 251ndash279 2003
Alegret L Thomas E and Lohmann K C End-Cretaceous ma-rine mass extinction not caused by productivity collapse P NatlAcad Sci USA 109 728ndash732 doi101073pnas11106011092012
Alegret L Rodriacuteguez-Tovar F J and Uchman A How bioturba-tion obscured the CretaceousndashPaleogene boundary record TerraNova 27 225ndash230 doi101111ter12151 2015
Altiner D Koccedilyigit A Farinacci A Nicosia U and ContiM A Jurassic-Lower Cretaceous stratigraphy and paleogeo-graphic evolution of the southern part of north-western Anatolia(Turkey) Geologica Romana 27 13ndash80 1991
Alvarez LW Alvarez W Asaro F and Michel H V Extrater-restrial cause for the Cretaceous-Tertiary extinction Science208 1095ndash1108 1980
Alegret L and Thomas E Benthic foraminifera and environmen-tal turnover across the CretaceousPaleogene boundary at BlakeNose (ODP Hole 1049C Northwestern Atlantic) PalaeogeogrPalaeocl 208 59ndash83 doi101016jpalaeo200402028 2004
Armstrong R A Lee C Hedges J I Honjo S and WakehamS G A new mechanistic model for organic carbon fluxes in theocean based on the quantitative association of POC with ballastmaterials Deep-Sea Res Pt II 49 219ndash236 2002
Aubert J and Berggren W A Paleocene benthic foraminiferalbiostratigraphy and paleoecology of Tunisia Bull du Cent deRech de Pau ndash SNPA 10 379ndash469 1976
Berggren W A and Aubert J Paleocene benthonic foraminiferalbiostratigraphy paleobiogeography and paleoecology ofAtlanticndashTethyan regions midway-type fauna PalaeogeogrPalaeocl 18 73ndash192 1975
Bernaola G and Monechi S Calcareous nannofossil ex-tinction and survivorship across the CretaceousndashPaleogeneboundary at Walvis Ridge (ODP Hole 1262C SouthAtlantic Ocean) Palaeogeogr Palaeocl 255 132ndash156doi101016jpalaeo200702045 2007
Bernhard J M Characteristic assemblages and morphologies ofbenthic foraminifera from anoxic organic-rich deposits Jurassicthrough Holocene J Foramin Res 16 207ndash215 1986
Birch H S Coxall H K Pearson P N Kroon D andSchmidt D N Partial collapse of the marine carbon pump af-ter the CretaceousndashPaleogene boundary Geology 44 287ndash290doi101130G375811 2016
Brinkhuis H and Zachariasse W J Dinoflagellate cysts sealevel changes and planktonic foraminifers across the Cretaceous-
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
898 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
Tertiary boundary at El Haria northwest Tunisia Mar Micropa-leontol 13 153ndash191 1988
Brinkhuis H Bujak J P Smit J Versteegh G J M and Viss-cher H Dinoflagellate-based sea surface temperature recon-structions across the Cretaceous-Tertiary boundary PalaeogeogrPalaeocl 141 67ndash83 1998
Buesseler K O The decoupling of production and particulate ex-port in the surface ocean Global Biogeochem Cy 12 297ndash3101998
Buzas M A Culver S J and Jorissen F J A statisti-cal evaluation of the microhabitats of living (stained) infau-nal benthic foraminifera Mar Micropaleontol 20 311ndash320doi1010160377-8398(93)90040-5 1993
Coccioni R and Galeotti S KndashT boundary extinction geolog-ically instantaneous or gradual event Evidence from deep-seabenthic foraminifera Geology 22 779ndash782 1994
Corliss B H Microhabitats of benthic foraminifera within deep-sea sediments Nature 314 435ndash438 doi101038314435a01985
Corliss B H and Chen C Morphotype patterns of Nor-wegian Sea deep-sea benthic foraminifera and ecologi-cal implications Geology 16 716ndash719 doi1011300091-7613(1988)016lt0716MPONSDgt23CO2 1988
Coxall H K DrsquoHondt S and Zachos J C Pelagic evolution andenvironmental recovery after the CretaceousndashPaleogene mass ex-tinction Geology 34 297ndash300 doi101130G217021 2006
Culver S J Benthic foraminifera across the CretaceousndashTertiary(KndashT) boundary a review Mar Micropaleontol 47 177ndash2262003
Cushman J A Upper Cretaceous foraminifera of the Gulf coastalregion of the United States and adjacent areas US Geolog SurvProf Pap 206 241 pp 1946
Cushman J A Paleocene foraminifera of the Gulf Coastal Regionof the United States and adjacent regions US Geol Surv ProfPap 232 75 pp 1951
DrsquoHondt S Consequences of the CretaceousPaleogene Mass Ex-tinction for Marine Ecosystems Annu Rev Ecol Evol S36 295ndash317 doi101146annurevecolsys350211031057152005
DrsquoHondt S and Zachos J C Cretaceous foraminifera and the evo-lutionary history of planktic photosymbiosis Paleobiology 24512ndash523 1998
DrsquoHondt S Donaghay P Zachos J C Luttenberg D andLindinger M Organic Carbon Fluxes and Ecological Recov-ery from the Cretaceous-Tertiary Mass Extinction Science 282276ndash279 1998
Donze P Colin J-P Damotte R Oertli HJ Peypouquet J-Pand Said R Les ostracodes du Campanien terminal agrave lrsquoEocegraveneinfeacuterieur de la coupe du Kef Tuneacutesie nord-occidentale Bull desCentr Recher Expl-Prod Elf Aquit 6 273ndash335 1982
Dortch Q and Packard T T Differences in biomass structure be-tween oligotrophic and eutrophic marine ecosystems Deep-SeaRes 36 223ndash240 1989
Ernst S R Guasti E Dupuis C and Speijer R P En-vironmental perturbation in the southern Tethys across thePaleoceneEocene boundary (Dababiya Egypt) Foraminiferaland clay mineral records Mar Micropaleontol 60 89ndash111doi101016jmarmicro200603002 2006
Eshet Y Moshkovitz S Habib D Benjamini C and Maga-ritz M Calcareous nannofossil and dinoflagellate stratigraphyacross the CretaceousTertiary boundary at Hor Hahar IsraelMar Micropaleontol 18 199ndash228 1992
Eshet Y Almogi-Labin A and Bein A Dinoflagellate cysts pa-leoproductivity and upwelling systems A Late Cretaceous ex-ample from Israel Mar Micropaleontol 23 231ndash240 1994
Esmeray-Senlet S Wright J D Olsson R K MillerK G Browning J V and Quan T M Evidencefor reduced export productivity following the Creta-ceousPaleogene mass extinction Paleoceanography 30718ndash738 doi1010022014PA002724 2015
Giusberti L Boscolo Galazzo F and Thomas E Variability inclimate and productivity during the Paleocene-Eocene ThermalMaximum in the western Tethys (Forada section) Clim Past12 213ndash240 doi105194cp-12-213-2016 2016
Guasti E Kouwenhoven T J Brinkhuis H and Speijer R PPaleocene sea-level and productivity changes at the southernTethyan margin (El Kef Tunisia) Mar Micropaleontol 55 1ndash17 2005
Habib D and Saeedi F The Manumiella seelandica global spikeCooling during regression at the close of the MaastrichtianPalaeogeogr Palaeocl 255 87ndash97 2007
Hain M P Sigmal D M and Haug G H 818 ndash The biolog-ical Pump in the Past in Reference Module in Earth Systemsand Environmental Sciences Treatise on Geochemistry (SecondEdition) The Oceans and Marine Geochemistry 8 485ndash517doi101016B978-0-08-095975-700618-5 2014
Hayek L C and Buzas M A On the proper and efficient useof diversity measures with individual field samples J ForaminRes 43 305ndash313 2013
Henson S A Sanders R Madsen E Morris P J Le MoigneF and Quartly G D A reduced estimate of the strength ofthe oceanrsquos biological carbon pump Geophys Res Lett 38L04606 doi1010292011GL046735 2011
Hilting A K Kump L R and Bralower T J Variations in theoceanic vertical carbon isotope gradient and their implicationsfor the Paleocene-Eocene biological pump Paleoceanography23 PA3222 doi1010292007PA001458 2008
Honjo S Manganini S J Krishfield R A and Francois RParticulate organic carbon fluxes to the ocean interior and fac-tors controlling the biological pump A synthesis of global sed-iment trap progams since 1983 Prog Oceanogr 76 217ndash285doi101016jpocean200711003 2008
Huber B T MacLeod K G and Norris R D Abrupt extinctionand subsequent reworking of Cretaceous planktonic foraminiferaacross the Cretaceous-Tertiary boundary Evidence from the sub-tropical North Atlantic in Catastrophic Events and Mass Extinc-tions Impacts and Beyond edited by Koeberl C and MacLeodK G Geol S Am S 356 277ndash289 Boulder Colorado USA2002
Hull P M Norris R D Bralower T J and Schueth J D A rolefor chance in marine recovery from the end-Cretaceous extinc-tion Nat Geosci 4 856ndash860 doi101038ngeo1302 2011
Hsuuml K J and McKenzie J A A ldquoStrangeloverdquo ocean in the earli-est Tertiary in Natural variations Archean to present edited bySundquist E T and Broecker W S The carbon cycle and atmo-spheric CO2 American Geophysical Union Washington DCUSA 487ndash492 1985
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
J Vellekoop et al Ecological response to KndashPg boundary mass extinction 899
Jiao N Herndl G J Hansell D A Benner R Kattner G Wil-helm S W Kirchman D L Weinbauer M G Luo T ChenF and Azam F Microbial production of recalcitrant dissolvedorganic matter long-term carbon storage in the global oceanNat Rev 8 593ndash599 doi101038nrmicro2386 2010
Jorissen F J de Stigter H C and Widmark J G V A concep-tual model explaining benthic foraminiferal microhabitats MarMicropaleontol 26 3ndash15 doi1010160377-8398(95)00047-X1995
Jorissen F J Fontanier C and Thomas E Paleoceanographi-cal proxies based on deep-sea benthic foraminiferal assemblagecharacteristics in Proxies in Late Cenozoic PaleoceanographyPt 2 Biological tracers and biomarkers edited by Hillaire-Marcel C and de Vernal A 263ndash326 Elsevier Amsterdamthe Netherlands 2007
Kaiho K Kajiwara Y Tazaki K Ueshima M Takeda NKawahata H Arinobu T Ishiwatari R Hirai A and Lam-olda M A Oceanic primary productivity and dissolved oxygenlevels at the CretaceousTertiary boundary their decrease sub-sequent warming and recovery Paleoceanography 14 511ndash5241999
Keller G and Lindinger M Stable isotope TOC and CaCO3record across the CretaceousTertiary boundary at El KefTunisia Palaeogeogr Palaeocl 73 243ndash265 1989
Kellough G R Paleoecology of the Foraminiferida of the WillsPoint Formation (Midway Group) in northeast Texas Trans GulfCoast Assoc Geol Soc 15 73ndash153 1965
Kring D A The Chicxulub impact event and its environmentalconsequences at the CretaceousndashTertiary boundary PalaeogeogrPalaeocl 255 4ndash21 2007
Kump L R Interpreting carbon-isotope excursions Strangeloveoceans Geology 19 299ndash302 1991
Machalski M Vellekoop J Dubicka Z Peryt D and Ha-rasimiuk M Late Maastrichtian cephalopods dinoflagellatecysts and foraminifera from the CretaceousndashPaleogene suc-cession at Lechoacutewka southeast Poland Stratigraphic andenvironmental implications Cretaceous Res 57 208ndash227doi101016jcretres201508012 2016
Molina E Evidence and causes of the main extinctionevents in the Paleogene based on extinction and survivalpatterns of foraminifera Earth-Sci Rev 140 166ndash181doi101016jearscirev201411008 2015
Molina E Alegret L Arenillas I and Arz J The GlobalBoundary Stratotype Section and Point for the base of the Da-nian Stage (Paleocene Paleogeneldquo Tertiaryrdquo Cenozoic) at ElKef Tunisia-Original definition and revision Episodes 29 263ndash273 2006
Olsson R K Liu C and van Fossen M The Cretaceous-Tertiarycatastrophic event at Millers Ferry Alabama in The Cretaceous-Tertiary Event and Other Catastrophes in Earth History editedby Ryder G Fastovsky D and Gartner S Geol S Am S307 263ndash277 1996
Peryt D Benthic foraminiferal response to the Cenomanian-Turonian and CretaceousndashPaleogene boundary events PrzegladGeol 52 827ndash832 2004
Peryt D Alegret L and Molina E The CretaceousPalaeogene(KP) boundary at Aiumln Settara Tunisia restructuring of benthicforaminiferal assemblages Terra Nova 14 101ndash107 2002
Ramette A Multivariate analyses in microbial ecology FEMS Mi-crobiol Ecol 62 142ndash160 2007
Rodriacuteguez-Tovar F J Uchman A MrsquoHamdi A Riahi Sand Ismail-Lattrache K B Ichnological record of palaeoen-vironment from the CretaceousndashPaleogene boundary intervalat El Kef Tunisia The first study of old and new sec-tions at the stratotype area J Afr Earth Sci 120 23ndash30doi101016jjafrearsci201604017 2016
Saner S Mudurnu-Goumlynuumlk Havzasının Jura ve Sonrasıccediloumlkelimnitelikleriyle paleocografya yorumlaması TJK Buumllteni 23 39ndash52 1980
Schioslashler P Brinkhuis H Roncaglia L and Wilson G J Di-noflagellate biostratigraphy and sequence stratigraphy of theType Maastrichtian (Upper Cretaceous) ENCI Quarry TheNetherlands Mar Micropaleontol 31 65ndash95 1997
Schueth J D Bralower T J Jiang S and Patzkowsky ME The role of regional survivor incumbency in the evolu-tionary recovery of calcareous nannoplankton from the Creta-ceousPaleogene (KPg) mass extinction Paleobiology 41 661ndash679 doi101017pab201528 2015
Scotese C R A continental drift flipbook J Geology 112 729ndash741 2004
Scotese C R and Dreher C GlobalGeology available at httpwwwGlobalGeologycom (last access 11 March 2014) 2012
Sepkoski J J Patterns of Phanerozoic extinction a perspectivefrom global data bases in Global Events and Event Stratigra-phy edited by Walliser O H Springer Berlin Germany 35ndash52 2016
Sibert E C Hull P M and Norris R D Resilience of Pacificpelagic fish across the CretaceousPalaeogene mass extinctionNat Geosci 7 667ndash670 doi101038NGEO2227 2014
Slimani H and Toufiq A A CretaceousndashPaleogene boundary geo-logical site revealed by planktic foraminifera and dinoflagellatecysts at Ouled Haddou eastern external Rif Chain Morocco JAfr Earth Sci 88 38ndash52 doi101016jjafrearsci2013080082013
Slimani H Louwye S and Toufiq A Dinoflagellate cystsform the CretaceousndashPaleogene boundary at Ouled Had-dou southeastern Rif Morocco biostratigraphy paleoenvi-ronments and paleobiogeography Palynology 34 90ndash124doi10108001916121003629933 2010
Sluijs A and Brinkhuis H A dynamic climate and ecosystemstate during the Paleocene-Eocene Thermal Maximum infer-ences from dinoflagellate cyst assemblages on the New JerseyShelf Biogeosciences 6 1755ndash1781 doi105194bg-6-1755-2009 2009
Smit J Extinction and evolution of planktonic foraminifera after amajor impact at the CretaceousTertiary boundary Geol S AmS 190 329ndash352 1982
Smit J and Romein A J T A sequence of events across theCretaceous-Tertiary boundary Earth Planet Sci Lett 74 155ndash170 1985
Speijer R P Extinction and recovery patterns in ben-thic foraminiferal paleocommunities across the Creta-ceousPaleogene and PaleoceneEocene boundaries GeologicaUltraiectina 124 Faculteit Aardwetenschappen PhD ThesisUniversiteit Utrecht Utrecht the Netherlands 1994
Speijer R P and van Der Zwaan G J Extinction and survivor-ship of southern Tethyan benthic foraminifera across the Cre-
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
900 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
taceousPaleogene boundary in Biotic recovery from mass ex-tinction events edited by Hart M B Geolog Soc Spec PublLondon UK 102 343ndash371 1996
Stemmann L and Boss E Plankton and Particle Size andPackaging From Determining Optical Properties to Drivingthe Biological Pump Annu Rev Mar Sci 4 263ndash290doi101146annurev-marine-120710-100853 2012
Thomas E Late Cretaceous-early Eocene mass extinctions in thedeep sea Geol S Am S 247 481ndash495 1990
Vandenberghe N Hilgen F J and Speijer R P The PaleogenePeriod in The Geologic Time Scale 2012 edited by GradsteinF M Ogg J G Schmitz M D and Ogg G M 2 855ndash921Elsevier Boston USA 2012
Vellekoop J Sluijs A Smit J Schouten S Weijers J WH Sinninghe Damsteacute J S and Brinkhuis H Rapid short-term cooling following the Chicxulub impact at the CretaceousndashPaleogene boundary P Natl Acad Sci USA 111 7537ndash7541doi101073pnas1319253111 2014
Vellekoop J Smit J van de Schootbrugge B Weijers J W HGaleotti S Sinninghe Damsteacute J S and Brinkhuis H Palyno-logical evidence for prolonged cooling along the Tunisian conti-nental shelf following the KndashPg boundary impact PalaeogeogrPalaeocl 426 216ndash228 doi101016jpalaeo2015030212015
Vellekoop J Esmeray-Senlet S Miller K G Browning J VSluijs A van de Schootbrugge B Sinninghe Damsteacute J Sand Brinkhuis H Evidence for CretaceousndashPaleogene bound-ary bolide ldquoimpact winterrdquo conditions from New Jersey USAGeology 44 619ndash622 doi101130G379611 2016
Woelders L and Speijer R Stable seafloor conditions sealevel and food supply during the latest Maastrichtian atBrazos River Texas Mar Micropaleontol 121 41ndash51doi101016jmarmicro201510002 2015
Zachos J C Arthur M A and Dean W E Geochemical evi-dence for suppression of pelagic marine productivity at the Cre-taceousTertiary boundary Nature 337 61ndash64 1989
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
- Abstract
- Introduction
- Geological setting and age assessment
- Materials and methods
-
- Sampling
- Foraminiferal analysis
- Palynological analysis
- Statistical analysis
-
- Results
-
- Benthic foraminifera
- Palynology
-
- Discussion
-
- Tethyan benthic foraminiferal turnover sequence
- Ecological responses to reduced export productivity
-
- Okccedilular section
- Western Tethys
- Global responses
-
- Long-term recovery
-
- Conclusions
- Data availability
- Author contributions
- Competing interests
- Acknowledgements
- References
-

J Vellekoop et al Ecological response to KndashPg boundary mass extinction 889
Figure 2 The benthic foraminiferal record (gt 125 microm) of the Okccedilular section The biostratigraphy is from Accedilikalin et al (2015) The clusteranalysis on the benthic foraminiferal data using paired group (UPGMA) correlation distance allows the identification of four main clustersAndashD of benthic foraminiferal taxa
umiella (3) hexaperidinioids lumping all other peridinioidcysts with a hexaform archeopyle and (4) other dinocystswhich include all other dinocyst taxa and unidentifiabledinocysts (Fig 3)
Of these different dinocyst groups previous studies haveshown that in the Tethys the hexaperidinioids in particularshow strong variations across the KndashPg boundary (Brinkhuiset al 1998 Vellekoop et al 2015) Based on statistical cor-relations between palynological records and other paleoprox-ies it has been suggested that this inferred heterotrophicgroup flourished best under high-nutrient conditions in thephotic zone (Eshet et al 1994 Brinkhuis et al 1998 Sluijsand Brinkhuis 2009) Therefore in this study abundances ofhexaperidinioids are considered indicative of nutrient avail-ability in the photic zone (see Fig S1 and Text S1 in theSupplement for a more detailed discussion on this matter)
34 Statistical analysis
To assess changes in diversity of benthic foraminifera anddinocysts across the studied interval the Shannon diversityindex (H ) the species richness per sample (S) the number ofspecimens observed per sample (N ) and the BergerndashParkerindex were calculated for both biological groups followingHayek and Buzas (2013) (Fig 4) In addition in order torecognize the main faunal associations within the benthicforaminiferal data a cluster analysis was performed usingpaired group (UPGMA) correlation distance
Q-mode non-metric multidimensional scaling (NM-MDS)was performed on the benthic foraminiferal sample compo-sitions to assess patterns in assemblage response to KndashPgboundary perturbations Since taxa may not have a linear re-sponse to environmental changes across the KndashPg boundaryQ-mode NM-MDS is preferred over PCA and CCA (Ram-ette 2007)
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
890 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
Figure 3 The organic-walled dinoflagellate cyst record of theOkccedilular section The biostratigraphy and bulk carbonate stable car-bon isotope records are from Accedilikalin et al (2015) The four maindinocyst complexes are indicated
4 Results
41 Benthic foraminifera
The benthic foraminiferal record of the Okccedilular section ischaracterized by a distinct turnover across the KndashPg bound-ary (Figs 2 4 5 6) Of the common taxa 8 out of 30(sim 27 ) disappear across the KndashPg boundary After the KndashPg boundary crisis three new taxa appear The estimatedBFAR shows a major decrease across the boundary (Fig 5)For the foraminiferal counts and illustrations of commonforms see Dataset S1 and Figs S2 S3
The cluster analysis allows the identification of fourmain clusters of benthic foraminiferal taxa clusters A toD (Fig 2) Cluster C is relatively large and can be subdi-vided into five subclusters Based on the succession of ben-thic faunal assemblages characterized by strong changes inthe Shannon diversity index (H ) four intervals can be rec-ognized in the benthic foraminiferal record of the Okccedilularsection (Figs 4 6)
The first interval comprises the uppermost Maastrichtianand is characterized by relatively high diversity dominatedby the taxa in cluster D In this assemblage the bi-triserialbenthic taxa are relatively abundant (22ndash28 ) with char-acteristic taxa such as Bulimina arkadelphiana Eouvigerinasubsculptura and Praebulimina reussi About 50ndash60 of theassemblage consists of inferred endobenthic taxa The KndashPg boundary marks an abrupt benthic community impover-
ishment and the decimation of the taxa in cluster D Abovethe boundary the bi-triserial benthic taxa virtually disappearfrom the record (mostly below lt 05 )
The second interval characterized by low diversity com-prises the lowermost Danian approximately correspondingto planktic foraminiferal Zone P0 In this interval epibenthicforms are most abundant (70ndash90 ) It is dominated by thetaxa of cluster A encompassing successive peak occurrencesof the taxa Anomalinoides praeacutus Trochammina sppand Cibicidoides pseudoacutus
The third interval covers the part of the succession approx-imately correlative to planktic foraminiferal Zone Pα Thisinterval is characterized by a recovery of diversity althoughthe diversity is still lower than that of the top Maastrichtianassemblage The abundance of taxa in cluster A slowly de-creases and taxa in cluster B mainly represented by Osan-gularia plummerae and Cibicidoides sp become abundantEndobenthic forms recover and make up 30ndash40 of the totalassemblage
The fourth interval starts in the interval correlative to ZoneP1a Here the benthic community has recovered as the di-versity has stabilized and is almost similar to pre-impact val-ues This interval is characterized by taxa in cluster C a typ-ical Paleocene Midway-type fauna (Berggren and Aubert1975) with representatives such as Anomalinoides praea-cutus Coryphostoma midwayensis Cibicidoides alleni andOsangularia plummerae
42 Palynology
Palynological samples from the Okccedilular site yield abundantpalynomorphs dominated by dinocysts and with minor con-tributions of acritarchs prasinophytes organic foraminiferallinings and terrestrial palynomorphs (Dataset S2) Thedinocyst associations of the MudurnundashGoumlynuumlk basin arequite diverse including components characteristic of boththe Tethyan and Boreal realms (Accedilikalin et al 2015) As ex-pected the dinocyst record does not show major changes indiversity across the KndashPg boundary (Fig 4) There is a steadydecrease in dinocyst diversity from planktic foraminiferalZone P1a upwards probably a long-term change not re-lated to the impact The estimated dinocyst accumulationrate (DAR number of preserved cysts per cm2 per kyr seeText S2 in the Supplement for details on the estimation of theDAR) shows no major changes across the KndashPg boundary
Throughout the palynological record the Spiniferites com-plex is consistently the most dominant morphogroup in gen-eral comprising 40ndash50 of the total assemblage (Fig 3)Similar to other KndashPg boundary sites worldwide (Habib andSaeedi 2007) Manumiella spp show an episode of higherrelative abundance near the top of the Maastrichtian some30ndash40 cm below the KndashPg boundary Hexaperidinioids gen-erally make up a relatively small component of the Maas-trichtian assemblage (3ndash17 ) but show a strong increaseacross the KndashPg boundary In the boundary clay layer cor-
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
J Vellekoop et al Ecological response to KndashPg boundary mass extinction 891
Figure 4 Diversity indices on the benthic foraminiferal and dinocyst records of the Okccedilular section with the Shannon diversity index (H ) thespecies richness per sample (S) the number of specimen observed per sample (N ) and the BergerndashParker index As the bulk stable carbonisotope record and the Shannon diversity index of the benthic foraminiferal record show similar trends the combination of these records canbe used to subdivide the KndashPg boundary transition into four intervals I the Maastrichtian ie ldquopre-impactrdquo interval II the post-impactinterval representing the ldquodisasterrdquo phase III a recovery interval and IV the early Paleocene post-recovery interval The division criteriaare explained in the text
relative to planktic foraminiferal Zone P0 this group in-creases to 35 of the assemblage In the peak intervals thehexaperidinioids are mostly represented by representativesof the genera Senegalinium and Cerodinium After an ini-tial drop in relative abundance in the upper half of Zone P0this group reaches a second maximum (sim 35 ) at the baseof Zone Pα and remains relatively abundant (15ndash30 ) up toZone P1a After this the hexaperidinioids gradually decreasetowards the top of the studied interval The ldquoother dinocystsrdquogroup ndash with representatives such as Areoligera spp Im-pagidinium spp Hystrichosphaeridium tubiferum Opercu-lodinium centrocarpum and Palynodinium grallator ndash gen-erally makes up 25ndash50 of the assemblage
5 Discussion
51 Tethyan benthic foraminiferal turnover sequence
The major turnover in the benthic community at Okccedilularis largely comparable with the previously published ben-thic foraminiferal record (size fraction gt 125 microm) of the KndashPg boundary GSSP of El Kef (Figs 7 8 Speijer and Vander Zwaan 1996) In addition other benthic foraminiferalrecords from the margins of the Tethys also show a strong
similarity to the Okccedilular and El Kef records including stud-ies that use the gt 63 microm size fraction (Alegret et al 2003Culver 2003 and references therein)
Although it is sometimes suggested that different sizefractions cannot be directly compared because the differ-ent size fractions sometimes harbor different ecological in-formation (Giusberti et al 2016) intercomparison studieshave shown that when comparing the gt 125 and gt 63 microm sizefractions in benthic foraminiferal assemblages the overallpatterns are generally robust (Ernst et al 2006) The factthat gt 63 and gt 125 microm KndashPg boundary benthic foraminiferalrecords from different Tethys-margin sites (eg Speijer andVan der Zwaan 1996 Peryt et al 2002 Culver et al 2003)show striking similarities even though both the size fractionsused (gt 63 vs gt 125 microm) and specific taxa making up theforaminiferal assemblages differ among sites confirms theobservations by Ernst et al (2006) Furthermore even whenit would be assumed that different size fractions are diffi-cult to compare directly the striking similarities between theTethyan gt 125 and gt 63 microm size fraction KndashPg boundary ben-thic foraminiferal records provide compelling evidence for astrong disruption of the entire benthic community
Tethyan middle neritic to middle bathyal KndashPg boundarysuccessions generally show a sequence of (1) a typical high
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
892 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
Figure 5 Estimated benthic foraminiferal accumulation rate (BFAR number of foraminifera of the gt 63 microm size fraction per cm2 sea floorper kyr) and estimated dinocyst accumulation rate (DAR number of preserved cysts per cm2 sea floor per kyr) of the Okccedilular sectionUncertainty in estimated accumulation rates resulting from uncertainties in sediment densities and sedimentation rates are indicated in greyproviding a range of estimated accumulation rates For more detailed info on the calculation of the BFAR and DAR see Text S2 of theSupplement
diversity assemblage in the Maastrichtian (2) a low diver-sity ldquodisasterrdquo assemblage directly after the KndashPg boundaryimpact characterized by a short-lived bloom of endobenthicforms observed in several high-resolution records (eg Coc-cioni and Galeotti 1994 Speijer and van der Zwaan 1996)followed by a dominance of epibenthic forms (3) a ldquore-coveryrdquo assemblage characterized by an increasing diversityand returning endobenthic forms and (4) a new high diver-sity assemblage dominated by a Paleocene fauna with bothepi- and endobenthic forms equally represented
Hence based on this succession of benthic assemblageswhich are characterized by strong changes in diversity (H )the KndashPg boundary benthic foraminiferal records from theTethys can be subdivided into four intervals (Figs 2 4ndash6)The intervals I to IV approximately correspond to the up-permost Maastrichtian planktic foraminiferal Zone P0 sub-zones PαndashP1a and subzones P1andashP1b respectively There-fore this succession roughly follows the 5-fold sedimen-tological and planktic foraminiferal sequence of Smit andRomein (1985) in which unit 2 represents the ejecta layerand units 1 3 4 and 5 correspond to intervals I to IV in ourstudy
The sequence of benthic foraminiferal turnover in theTethys is also well illustrated by a Q-mode NM-MDS
(Fig 7) The Q-mode NM-MDS of the benthic foraminiferalsample compositions of the combined data from Okccedilular(this study) and El Kef (Speijer and van der Zwaan 1996)demonstrates that although the assemblages differ betweenthese localities on opposite margins of the Tethys the re-sponses of the benthic foraminiferal assemblages to the KndashPgboundary perturbations express very similar overall patternsacross the KndashPg boundary At both Okccedilular and El Kef thebenthic foraminiferal records show a rapid transition from astable Maastrichtian assemblage to the earliest Danian dis-aster phase in Zone P0 followed by a gradual change backtowards conditions similar to the Maastrichtian (marked bythe return of Lazarus taxa) Therefore both the successionof benthic faunal assemblages including diversity indicesand Q-mode NM-MDS analysis provide powerful tools to al-low the comparison of KndashPg boundary benthic foraminiferalrecords from different localities
52 Ecological responses to reduced export productivity
521 Okccedilular section
The palynological benthic foraminiferal and bulk stable iso-tope records of the Okccedilular section show major changesacross the KndashPg boundary interval (Figs 2ndash7) portraying
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
J Vellekoop et al Ecological response to KndashPg boundary mass extinction 893
Figure 6 Relative abundances of the four main clusters of benthic foraminiferal taxa (gt 125 microm) relative abundances of epibenthic andendobenthic forms and relative abundance of bi-triserial benthic taxa
Figure 7 Q-mode non-metric multidimensional scaling (on all sam-ples in the gt 125 microm size fraction benthic foraminiferal datasets ofOkccedilular (this study) and El Kef (Speijer and van der Zwaan 1996)After an abrupt transition from latest Maastrichtian pre-impact as-semblages to disaster assemblages in Zone P0 and recovery assem-blages in Zone Pα a gradual transition towards a new equilibriumpost-recovery assemblage can be observed in subzones P1a and P1b
the biological crisis following the impact Following the KndashPg boundary mass extinction endobenthic forms stronglydecrease with bi-triserial benthic taxa almost disappearingfrom the benthic community simultaneously the estimatedBFAR shows a major fall (Fig 5) This interval approxi-mately correlative to foraminiferal Zone P0 represents thedisaster phase The drop in bi-triserial benthic taxa (Fig 6)and decrease in BFAR (Fig 5) point to a reduction in foodsupply to the benthic community indicating that the trans-port of organic matter to the sea floor must have decreasedIn the dinocyst community however hexaperidinioids showa strong increase in relative abundance during this phase(Figs 3 6 7) suggesting an increase in nutrient availabilityin the photic zone while the DAR remains relatively stableacross the KndashPg boundary (Fig 5)
The decrease in benthic diversity and the high relativeabundances of opportunistic Trochammina spp and Cibici-doides pseudoacutus suggest that the benthic community ex-perienced additional stress during Zone P0 besides food lim-itation (see Jorissen et al 2007) If food limitation were theonly factor involved upper bathyal assemblages as observedat Okccedilular would be expected to become more similar tooligotrophic middle to lower bathyal assemblages Insteadthe opposite is true the assemblage composition of the dis-aster phase is closer to neritic Midway-type assemblages
It was previously suggested that the epibenthic taxon Cibi-cidoides pseudoacutus could potentially be hypoxia resistant
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
894 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
(Speijer and van der Zwaan 1996) As a collapse of the bi-ological pump should lead to enhanced remineralization oforganic matter in the photic zone it is indeed likely thatthe oxygen minimum zones expanded and shoaled leadingto oxygen stress on the sea floor Although early Paleocenedysoxia at El Kef is supported by the ostracod record (Donzeet al 1982) and ichnofacies analyses (Rodriguez-Tovaret al 2016) supporting (eg geochemical) evidence fordysoxia during the disaster phase at Okccedilular record is absentFurthermore an additional stressor for the benthic commu-nity during Zone P0 may be present In contrast to calcare-ous nanoplankton organic-walled cyst-producing dinoflagel-lates did not suffer extinctions and may have become a moreimportant component in the early Paleocene phytoplanktoncommunity Hence besides the reduction of the amount oforganic matter transported to the sea floor the compositionof food supplied by the photic zone likely changed signifi-cantly across the KndashPg boundary as well (DrsquoHondt 2005)possibly presenting an additional stress factor for the benthiccommunity (Speijer and van der Zwaan 1996 Alegret andThomas 2009)
522 Western Tethys
The patterns in benthic foraminiferal and dinoflagellate re-sponse to the KndashPg boundary perturbations at Okccedilular appearto be characteristic for the continental margins of the westernTethyan realm The benthic foraminiferal patterns are similaracross different size fractions suggesting that they are repre-sentative of the overall benthic communities In Tunisia andSpain where both benthic foraminiferal and dinoflagellaterecords are available the earliest Danian is also characterizedby a decrease in endobenthic foraminifera at the sea floor(Speijer and Van der Zwaan 1996 Peryt et al 2002 Ale-gret et al 2003) and simultaneously blooms of hexaperidin-ioids in the water column (Brinkhuis et al 1998 Vellekoopet al 2015 Fig 8) In the Ouled Haddou section Moroccoin the westernmost Tethys (Fig 1) the lowermost Danian isalso characterized by a strong increase of the hexaperidin-ioid Senegalinium group up to 30 of the assemblage (Sli-mani et al 2010) that is very similar to the Okccedilular record(Fig 8)
The KndashPg boundary dinocyst record of El Kef is strikinglysimilar to the one from the Okccedilular section showing two dis-tinct peaks in hexaperidinioids one in Zone P0 and one atthe basal part of Zone Pα At El Kef the initial post-impactdominance of hexaperidinioids is however less pronounced(Fig 8) This difference in expression might be related tosmall differences in paleogeographic and paleoceanographicsettings between sites
In addition some high-resolution benthic KndashPg records inthe Tethyan realm show a short-lived bloom of specific en-dobenthic foraminiferal taxa in the few centimeter directlyabove the KndashPg boundary sometimes accompanied by otherindicators of low oxygen levels (eg Coccioni and Gale-
otti 1994 Speijer and Van der Zwaan 1996 Kaiho et al1999) This suggests that at least locally the sea floor wastemporarily characterized by hypoxic conditions directly fol-lowing the impact This short-lived deoxygenation is possi-bly related to the mass mortality at the KndashPg boundary re-sulting in a large short-lived flux of organic matter to thesea floor (Coccioni and Galeotti 1994) However since ex-port productivity was greatly reduced after the KndashPg bound-ary mass extinction very little ldquonewrdquo organic matter reachedthe sea floor after the initial post-impact influx Hence afteraccumulated organic matter was remineralized the benthiccommunity starved resulting in a transition to an epibenthic-dominated benthic fauna similar to the post-impact fauna ofthe Okccedilular record
The combined dinocyst records suggest that the early Pa-leocene margins of the Tethys were characterized by an in-crease in nutrient availability in the photic zone whereascoeval benthic foraminiferal records indicate a major de-crease in nutrient supply to the sea floor These changesin the benthic foraminiferal and dinocyst communities arelikely caused by the major reduction of both the efficiencyand strength of the biological pump (ie the fraction ofprimary production exported from the photic zone and theamount of organic matter transported down) in accordancewith the Living Ocean model (DrsquoHondt and Zachos 1998DrsquoHondt et al 1998 DrsquoHondt 2005 Coxall et al 2006)For a detailed explanation of the difference between biologi-cal pump efficiency and biological pump strength see Hiltinget al (2008)
As bulk carbonate δ13C reflects the isotopic compositionof the surface ocean which is set by burial fractions and bythe photosynthetic isotope effect (Kump 1991 Hain et al2014) the excursion recorded in bulk δ13C records world-wide and the rapid collapse in surface to deep-ocean carbonisotope gradients likely reflect the reduction of the global in-tensity (ie efficiency) of the biological pump (Hain et al2014) The recorded inverse change in nutrient availabilitysuggests a causal link A reduced efficiency of the biologi-cal pump and associated decrease of the fraction of biomasstransported out of the photic zone could have resulted inhigh rates of nutrient recycling in the upper part of the wa-ter column (DrsquoHondt 2005) This suggests that the reduc-tion of the biological pump strength recorded by the benthicforaminiferal record is a consequence of the decreased effi-ciency of the biological pump recorded by the carbon iso-tope and dinoflagellate cyst records The strong correlationbetween the bulk carbon isotope curves and the Shannondiversity index (H ) of the benthic foraminiferal records atboth Okccedilular (R2
= 073 p lt 0001) and El Kef (R2= 074
p lt 0001) suggest a clear link between the changes in ben-thic foraminiferal assemblages and the collapse and recoveryof biological pump efficiency
As a result of the reduced biological pump efficiency morenutrients remained available for the early Paleocene phyto-plankton community At Tethyan margin sites this is indi-
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
J Vellekoop et al Ecological response to KndashPg boundary mass extinction 895
Figure 8 The benthic foraminiferal (gt 125 microm) dinocyst and bulk stable carbon isotope records of Okccedilular (Turkey) and El Kef (Tunisia)the dinocyst record of Ouled Haddou (Morocco) and the benthic foraminiferal record (gt 63 microm) of ODP Hole 1049C Blake Nose (NorthAtlantic) Note that the definition of ldquohexaperidinioidsrdquo used is provided in the text The four phases identified based on the bulk stablecarbon isotopes and the Shannon diversity index (H ) of the benthic foraminiferal records are indicated (IndashIV) The gt 63 microm size fractionbenthic foraminiferal record of ODP Hole 1049C is from Alegret and Thomas (2004) who argued that the lowermost Paleocene of thissite comprised reworked foraminifera This interval is indicated by a dashed bar in the figure The biostratigraphy of Ouled Haddou is fromSlimani and Toufiq (2013) whereas the dinocyst record of this site is from Slimani et al (2010) The biostratigraphy of El Kef is based onBrinkhuis et al (1998) Speijer and van der Zwaan (1996) and Molina et al (2006) The fine-fraction stable carbon isotope record (blackdots) of El Kef is from Keller and Lindinger (1989) The Shannon diversity index (red dots) relative abundances of bi-triserial benthic taxa(dark purple) and all endobenthic foraminifera (light purple) are based on the gt 125 microm size fraction foraminiferal data of Speijer and vander Zwaan (1996) The palynological data of El Kef are from Brinkhuis et al (1998) (lower part) and Guasti et al (2005) (upper part) TheShannon diversity index of the gt 125 microm size fraction benthic foraminiferal record (red dots) the relative contribution of bi-triserial benthictaxa (dark purple) all endobenthic foraminifera (light purple) and hexaperidinioid dinocysts from Okccedilular are from this study The bulkstable carbon isotope record of Okccedilular (black dots) is from Accedilikalin et al (2015) Mbsf indicates meters below sea floor
cated by the higher abundance of hexaperidinioids Similarlyblooms of eutrophic survivor taxa of calcareous nanoplank-ton at open ocean sites have been suggested to be relatedto the buildup of nutrients in the open ocean photic zone(Schueth et al 2015) Although the carbon isotope gradientsbetween surface and deep waters indicate that the recoveryof the biological pump took hundreds of thousands of years(Zachos et al 1989 Kump 1991 DrsquoHondt et al 1998 Cox-all et al 2006 Birch et al 2016) our records suggest thatthe enhanced recycling of nutrients in the Tethys was partic-ularly intense during two phases in the first tens of thousandsyears after the impact (Fig 8)
523 Global responses
The decrease in export productivity following the KndashPgboundary is not only recognized in the Tethyan realm In theAtlantic Ocean Southern Ocean and the Indian Ocean a de-crease of the biological pump strength after the KndashPg bound-ary was also observed in neritic to abyssal environments(eg Thomas 1990 Olsson et al 1996 Hull et al 2011Alegret et al 2012) As an example the gt 63 microm size frac-tion benthic foraminiferal KndashPg boundary record of BlakeNose (ODP Hole 1049C northwestern Atlantic Alegret andThomas 2004) shows a pattern that is strikingly similar tothe gt 125 microm size fraction records in the Tethyan realm witha strong decrease of endobenthics in Zone P0 followed by a
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
896 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
gradual recovery across zones Pα and P1a (Fig 8) The simi-larities between the benthic foraminiferal patterns of Tethyanouter neritic to middle bathyal (200ndash1500 m) sites and thoseof the middle to lower bathyal (1500ndash1600 m) record ofBlake Nose (Alegret and Thomas 2004) suggest that theserecords register a global decrease in export productivity fol-lowing the KndashPg boundary
However several open ocean sites mostly in the Pacificshow an opposite trend with several lines of evidence sug-gesting an increased biological pump strength at these sites(eg Hull et al 2011 Alegret et al 2012) This led Ale-gret et al (2012) to conclude that the decrease in biologicalpump strength after the KndashPg boundary was most likely aregional instead of a global effect arguing against the Liv-ing Ocean hypothesis We would argue that low-productivityopen ocean sites like the central Pacific are characterized byvery different ecosystem structures and higher productivityas those along the Tethyan margins (eg Dortch and Pack-erd 1989) Therefore the consequence of ecosystem reorga-nization resulting from the mass extinction was likely verydifferent in the open ocean Pacific (eg Sibert et al 2014)Esmeray-Senlet et al (2015) proposed the term ldquoHeteroge-neous Oceanrdquo for this conceptual model as an alternativeto the Living Ocean model Supposedly the HeterogeneousOcean was characterized by a strong geographic heterogene-ity in the extinction patterns and food supply to the sea floor
However as the primary food source for open ocean deep-sea benthic communities generally consists of refractory or-ganic matter (eg Jiao et al 2010) it remains uncertainwhether changes in the intensity of recycling of the gener-ally more labile organic matter in the surface ocean will re-sult in changes in the amount of organic matter that benthiccommunities receive as they will likely continue to receiverecalcitrant organic matter at barely diminished rates despitethe collapse of the biological pump
In addition it should be noted that although the bio-logical pump efficiency is generally relatively high in openocean realms such as the central Pacific the biological pumpstrength is generally very low at these sites (Honjo et al2008 Henson et al 2011) Small changes in absolute bio-logical pump strength at such a locality could therefore havehad a large effect on the local oligotrophic benthic commu-nity (Alegret and Thomas 2009) but they likely representedonly a small fraction of the net amount of organic matterglobally transported to the deep Hence even though localbiological pump strength might have increased at some low-productivity deep-sea sites the large decrease in biologi-cal pump strength recorded at many other sites means thatthe net amount of organic matter globally transported out ofthe surface ocean was likely still reduced in the post-impactworld (Birch et al 2016) Therefore we argue that such Pa-cific records do not represent the general pattern The globalocean response can be characterized as heterogeneous how-ever the KndashPg boundary mass extinction did lead to a strongreduction of the net amount of organic matter globally trans-
ported from the surface to the sea floor while biological pro-ductivity recovered rapidly after the extinction event in ac-cordance with the Living Ocean hypothesis
53 Long-term recovery
Following the disaster phase the hexaperidinioid cysts atOkccedilular and El Kef decreased in abundance and the abun-dance of endobenthic forms increased again as the recoveryof the benthic community was initiated This recovery phaseis approximately correlative to Zone Pα and the basal part ofZone P1a which according to the Paleogene age constraintsof Vandenberghe et al (2012) represents at least 300 kyrThis duration is in agreement with the estimations providedby Birch et al (2016) During this phase the carbon iso-tope records remain well below pre-impact values and thediversity of the benthic foraminiferal community has not yetfully recovered (Figs 4ndash8) This indicates that the rapid andshort-lived KndashPg boundary disaster was followed by a rela-tively long recovery phase in line with previous estimatesof a multimillion-year biological recovery (eg Coxall etal 2006) While the impact-related environmental perturba-tions were short lived (Kring 2007 Vellekoop et al 20142016) the extinctions amongst important biological groupsled to a reduction of the organic flux from the photic zone todeep water resulting in major long-term biological and pa-leoceanographic reorganizations Only with the evolutionaryrecovery of the pelagic community governing the biologicalcarbon pump did export productivity start to increase again(eg Coxall et al 2006 Birch et al 2016)
6 Conclusions
The marine palynological benthic foraminiferal and bulkstable isotope records of the Okccedilular and El Kef sections re-veal major changes across the KndashPg boundary interval por-traying the biological crisis at the KndashPg boundary and subse-quent recovery in the early Paleocene Based on the succes-sion of benthic faunal assemblages at a number of Tethyanmargin sites four phases can be recognized across the KndashPgboundary interval the Maastrichtian or pre-impact phase (I)a disaster phase (II) a recovery phase (III) and an early Pa-leocene post-recovery phase (IV)
Following the KndashPg boundary impact some localitiesshow a large short-lived flux of food to the sea floor likelyrelated to the mass mortality at the KndashPg boundary Howeversince export productivity was greatly reduced after the KndashPgboundary mass extinction it is likely that after the initialpost-impact influx very little fresh organic matter reachedthe sea floor This reduction of export productivity in thepost-extinction disaster phase eventually resulted in a lowerfood supply to the sea floor This presented a major stressfactor for benthic organisms in the western Tethys The re-duced food supply resulted in an abrupt impoverishment of
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
J Vellekoop et al Ecological response to KndashPg boundary mass extinction 897
benthic communities As the downward transport of nutri-ents was slowed down recycling in upper layers increasedAs a result more nutrients remained available for the earlyPaleocene phytoplankton community leading to blooms ofdinoflagellates along the Tethyan margins and blooms of cal-careous nanoplankton taxa in more open ocean sites Ourrecords show that the enhanced recycling of nutrients in theTethys was particularly intense in the tens of thousands ofyears after the impact Following this the slow evolution-ary recovery of the pelagic community governing the bio-logical carbon pump resulted in a gradually increasing ex-port productivity in the hundreds of thousands of years afterthe impact Hence our integration of dinocyst and benthicforaminiferal records across the KndashPg boundary provides im-portant new insights into the ecological responses to the re-duction of export productivity following the mass extinctionat the CretaceousndashPaleogene boundary highlighting the di-rect ecological consequences of the Living Ocean conditionsin the post-impact world
7 Data availability
All data related to publication are available in the Supple-ment
The Supplement related to this article is available onlineat doi105194bg-14-885-2017-supplement
Author contributions Johan Vellekoop Lineke Woelders andHenk Brinkhuis designed the research Samples were collectedin the field by Johan Vellekoop Sanem Accedilikalin Ismail Ouml Yil-maz and Jan Smit Palynological analyses were carried out by Jo-han Vellekoop Benthic foraminiferal analyses were carried out byLineke Woelders and Robert P Speijer Johan Vellekoop LinekeWoelders and Bas van de Schootbrugge prepared the manuscriptwith input from all authors
Competing interests The authors declare that they have no conflictof interest
Acknowledgements This work was supported by the NetherlandsOrganization for Scientific Research (NWO) Open CompetitionGrant ALWPJ09047 to H Brinkhuis and the Research FoundationFlanders (FWO) grant G0B8513 to R P Speijer We thank LKump F Boscolo Galazzo P Roopnarine and one anonymousreferee for their suggestions
Edited by D GillikinReviewed by L Kump F Boscolo Galazzo and P Roopnarine
References
Accedilikalin S Vellekoop J Ocakoglu F Yılmaz I Ouml Smit J Al-tiner S O Goderis S Vonhof H Speijer R P WoeldersL Fornaciari E and Brinkhuis H Geochemical and pale-ontological characterization of a new KndashPg Boundary local-ity from the Northern branch of the Neo-Tethys MudurnundashGoumlynuumlk Basin NW Turkey Cretaceous Res 52 251ndash267doi101016jcretres201407011 2015
Alegret L and Thomas E Food supply to the seafloor in the Pa-cific Ocean after the CretaceousPaleogene boundary event MarMicropaleontol 73 105ndash116 2009
Alegret L Molina E and Thomas E Benthic foraminiferalturnover across the CretaceousPaleogene boundary at Agost(southeastern Spain) paleoenvironmental inferences Mar Mi-cropaleontol 48 251ndash279 2003
Alegret L Thomas E and Lohmann K C End-Cretaceous ma-rine mass extinction not caused by productivity collapse P NatlAcad Sci USA 109 728ndash732 doi101073pnas11106011092012
Alegret L Rodriacuteguez-Tovar F J and Uchman A How bioturba-tion obscured the CretaceousndashPaleogene boundary record TerraNova 27 225ndash230 doi101111ter12151 2015
Altiner D Koccedilyigit A Farinacci A Nicosia U and ContiM A Jurassic-Lower Cretaceous stratigraphy and paleogeo-graphic evolution of the southern part of north-western Anatolia(Turkey) Geologica Romana 27 13ndash80 1991
Alvarez LW Alvarez W Asaro F and Michel H V Extrater-restrial cause for the Cretaceous-Tertiary extinction Science208 1095ndash1108 1980
Alegret L and Thomas E Benthic foraminifera and environmen-tal turnover across the CretaceousPaleogene boundary at BlakeNose (ODP Hole 1049C Northwestern Atlantic) PalaeogeogrPalaeocl 208 59ndash83 doi101016jpalaeo200402028 2004
Armstrong R A Lee C Hedges J I Honjo S and WakehamS G A new mechanistic model for organic carbon fluxes in theocean based on the quantitative association of POC with ballastmaterials Deep-Sea Res Pt II 49 219ndash236 2002
Aubert J and Berggren W A Paleocene benthic foraminiferalbiostratigraphy and paleoecology of Tunisia Bull du Cent deRech de Pau ndash SNPA 10 379ndash469 1976
Berggren W A and Aubert J Paleocene benthonic foraminiferalbiostratigraphy paleobiogeography and paleoecology ofAtlanticndashTethyan regions midway-type fauna PalaeogeogrPalaeocl 18 73ndash192 1975
Bernaola G and Monechi S Calcareous nannofossil ex-tinction and survivorship across the CretaceousndashPaleogeneboundary at Walvis Ridge (ODP Hole 1262C SouthAtlantic Ocean) Palaeogeogr Palaeocl 255 132ndash156doi101016jpalaeo200702045 2007
Bernhard J M Characteristic assemblages and morphologies ofbenthic foraminifera from anoxic organic-rich deposits Jurassicthrough Holocene J Foramin Res 16 207ndash215 1986
Birch H S Coxall H K Pearson P N Kroon D andSchmidt D N Partial collapse of the marine carbon pump af-ter the CretaceousndashPaleogene boundary Geology 44 287ndash290doi101130G375811 2016
Brinkhuis H and Zachariasse W J Dinoflagellate cysts sealevel changes and planktonic foraminifers across the Cretaceous-
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
898 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
Tertiary boundary at El Haria northwest Tunisia Mar Micropa-leontol 13 153ndash191 1988
Brinkhuis H Bujak J P Smit J Versteegh G J M and Viss-cher H Dinoflagellate-based sea surface temperature recon-structions across the Cretaceous-Tertiary boundary PalaeogeogrPalaeocl 141 67ndash83 1998
Buesseler K O The decoupling of production and particulate ex-port in the surface ocean Global Biogeochem Cy 12 297ndash3101998
Buzas M A Culver S J and Jorissen F J A statisti-cal evaluation of the microhabitats of living (stained) infau-nal benthic foraminifera Mar Micropaleontol 20 311ndash320doi1010160377-8398(93)90040-5 1993
Coccioni R and Galeotti S KndashT boundary extinction geolog-ically instantaneous or gradual event Evidence from deep-seabenthic foraminifera Geology 22 779ndash782 1994
Corliss B H Microhabitats of benthic foraminifera within deep-sea sediments Nature 314 435ndash438 doi101038314435a01985
Corliss B H and Chen C Morphotype patterns of Nor-wegian Sea deep-sea benthic foraminifera and ecologi-cal implications Geology 16 716ndash719 doi1011300091-7613(1988)016lt0716MPONSDgt23CO2 1988
Coxall H K DrsquoHondt S and Zachos J C Pelagic evolution andenvironmental recovery after the CretaceousndashPaleogene mass ex-tinction Geology 34 297ndash300 doi101130G217021 2006
Culver S J Benthic foraminifera across the CretaceousndashTertiary(KndashT) boundary a review Mar Micropaleontol 47 177ndash2262003
Cushman J A Upper Cretaceous foraminifera of the Gulf coastalregion of the United States and adjacent areas US Geolog SurvProf Pap 206 241 pp 1946
Cushman J A Paleocene foraminifera of the Gulf Coastal Regionof the United States and adjacent regions US Geol Surv ProfPap 232 75 pp 1951
DrsquoHondt S Consequences of the CretaceousPaleogene Mass Ex-tinction for Marine Ecosystems Annu Rev Ecol Evol S36 295ndash317 doi101146annurevecolsys350211031057152005
DrsquoHondt S and Zachos J C Cretaceous foraminifera and the evo-lutionary history of planktic photosymbiosis Paleobiology 24512ndash523 1998
DrsquoHondt S Donaghay P Zachos J C Luttenberg D andLindinger M Organic Carbon Fluxes and Ecological Recov-ery from the Cretaceous-Tertiary Mass Extinction Science 282276ndash279 1998
Donze P Colin J-P Damotte R Oertli HJ Peypouquet J-Pand Said R Les ostracodes du Campanien terminal agrave lrsquoEocegraveneinfeacuterieur de la coupe du Kef Tuneacutesie nord-occidentale Bull desCentr Recher Expl-Prod Elf Aquit 6 273ndash335 1982
Dortch Q and Packard T T Differences in biomass structure be-tween oligotrophic and eutrophic marine ecosystems Deep-SeaRes 36 223ndash240 1989
Ernst S R Guasti E Dupuis C and Speijer R P En-vironmental perturbation in the southern Tethys across thePaleoceneEocene boundary (Dababiya Egypt) Foraminiferaland clay mineral records Mar Micropaleontol 60 89ndash111doi101016jmarmicro200603002 2006
Eshet Y Moshkovitz S Habib D Benjamini C and Maga-ritz M Calcareous nannofossil and dinoflagellate stratigraphyacross the CretaceousTertiary boundary at Hor Hahar IsraelMar Micropaleontol 18 199ndash228 1992
Eshet Y Almogi-Labin A and Bein A Dinoflagellate cysts pa-leoproductivity and upwelling systems A Late Cretaceous ex-ample from Israel Mar Micropaleontol 23 231ndash240 1994
Esmeray-Senlet S Wright J D Olsson R K MillerK G Browning J V and Quan T M Evidencefor reduced export productivity following the Creta-ceousPaleogene mass extinction Paleoceanography 30718ndash738 doi1010022014PA002724 2015
Giusberti L Boscolo Galazzo F and Thomas E Variability inclimate and productivity during the Paleocene-Eocene ThermalMaximum in the western Tethys (Forada section) Clim Past12 213ndash240 doi105194cp-12-213-2016 2016
Guasti E Kouwenhoven T J Brinkhuis H and Speijer R PPaleocene sea-level and productivity changes at the southernTethyan margin (El Kef Tunisia) Mar Micropaleontol 55 1ndash17 2005
Habib D and Saeedi F The Manumiella seelandica global spikeCooling during regression at the close of the MaastrichtianPalaeogeogr Palaeocl 255 87ndash97 2007
Hain M P Sigmal D M and Haug G H 818 ndash The biolog-ical Pump in the Past in Reference Module in Earth Systemsand Environmental Sciences Treatise on Geochemistry (SecondEdition) The Oceans and Marine Geochemistry 8 485ndash517doi101016B978-0-08-095975-700618-5 2014
Hayek L C and Buzas M A On the proper and efficient useof diversity measures with individual field samples J ForaminRes 43 305ndash313 2013
Henson S A Sanders R Madsen E Morris P J Le MoigneF and Quartly G D A reduced estimate of the strength ofthe oceanrsquos biological carbon pump Geophys Res Lett 38L04606 doi1010292011GL046735 2011
Hilting A K Kump L R and Bralower T J Variations in theoceanic vertical carbon isotope gradient and their implicationsfor the Paleocene-Eocene biological pump Paleoceanography23 PA3222 doi1010292007PA001458 2008
Honjo S Manganini S J Krishfield R A and Francois RParticulate organic carbon fluxes to the ocean interior and fac-tors controlling the biological pump A synthesis of global sed-iment trap progams since 1983 Prog Oceanogr 76 217ndash285doi101016jpocean200711003 2008
Huber B T MacLeod K G and Norris R D Abrupt extinctionand subsequent reworking of Cretaceous planktonic foraminiferaacross the Cretaceous-Tertiary boundary Evidence from the sub-tropical North Atlantic in Catastrophic Events and Mass Extinc-tions Impacts and Beyond edited by Koeberl C and MacLeodK G Geol S Am S 356 277ndash289 Boulder Colorado USA2002
Hull P M Norris R D Bralower T J and Schueth J D A rolefor chance in marine recovery from the end-Cretaceous extinc-tion Nat Geosci 4 856ndash860 doi101038ngeo1302 2011
Hsuuml K J and McKenzie J A A ldquoStrangeloverdquo ocean in the earli-est Tertiary in Natural variations Archean to present edited bySundquist E T and Broecker W S The carbon cycle and atmo-spheric CO2 American Geophysical Union Washington DCUSA 487ndash492 1985
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
J Vellekoop et al Ecological response to KndashPg boundary mass extinction 899
Jiao N Herndl G J Hansell D A Benner R Kattner G Wil-helm S W Kirchman D L Weinbauer M G Luo T ChenF and Azam F Microbial production of recalcitrant dissolvedorganic matter long-term carbon storage in the global oceanNat Rev 8 593ndash599 doi101038nrmicro2386 2010
Jorissen F J de Stigter H C and Widmark J G V A concep-tual model explaining benthic foraminiferal microhabitats MarMicropaleontol 26 3ndash15 doi1010160377-8398(95)00047-X1995
Jorissen F J Fontanier C and Thomas E Paleoceanographi-cal proxies based on deep-sea benthic foraminiferal assemblagecharacteristics in Proxies in Late Cenozoic PaleoceanographyPt 2 Biological tracers and biomarkers edited by Hillaire-Marcel C and de Vernal A 263ndash326 Elsevier Amsterdamthe Netherlands 2007
Kaiho K Kajiwara Y Tazaki K Ueshima M Takeda NKawahata H Arinobu T Ishiwatari R Hirai A and Lam-olda M A Oceanic primary productivity and dissolved oxygenlevels at the CretaceousTertiary boundary their decrease sub-sequent warming and recovery Paleoceanography 14 511ndash5241999
Keller G and Lindinger M Stable isotope TOC and CaCO3record across the CretaceousTertiary boundary at El KefTunisia Palaeogeogr Palaeocl 73 243ndash265 1989
Kellough G R Paleoecology of the Foraminiferida of the WillsPoint Formation (Midway Group) in northeast Texas Trans GulfCoast Assoc Geol Soc 15 73ndash153 1965
Kring D A The Chicxulub impact event and its environmentalconsequences at the CretaceousndashTertiary boundary PalaeogeogrPalaeocl 255 4ndash21 2007
Kump L R Interpreting carbon-isotope excursions Strangeloveoceans Geology 19 299ndash302 1991
Machalski M Vellekoop J Dubicka Z Peryt D and Ha-rasimiuk M Late Maastrichtian cephalopods dinoflagellatecysts and foraminifera from the CretaceousndashPaleogene suc-cession at Lechoacutewka southeast Poland Stratigraphic andenvironmental implications Cretaceous Res 57 208ndash227doi101016jcretres201508012 2016
Molina E Evidence and causes of the main extinctionevents in the Paleogene based on extinction and survivalpatterns of foraminifera Earth-Sci Rev 140 166ndash181doi101016jearscirev201411008 2015
Molina E Alegret L Arenillas I and Arz J The GlobalBoundary Stratotype Section and Point for the base of the Da-nian Stage (Paleocene Paleogeneldquo Tertiaryrdquo Cenozoic) at ElKef Tunisia-Original definition and revision Episodes 29 263ndash273 2006
Olsson R K Liu C and van Fossen M The Cretaceous-Tertiarycatastrophic event at Millers Ferry Alabama in The Cretaceous-Tertiary Event and Other Catastrophes in Earth History editedby Ryder G Fastovsky D and Gartner S Geol S Am S307 263ndash277 1996
Peryt D Benthic foraminiferal response to the Cenomanian-Turonian and CretaceousndashPaleogene boundary events PrzegladGeol 52 827ndash832 2004
Peryt D Alegret L and Molina E The CretaceousPalaeogene(KP) boundary at Aiumln Settara Tunisia restructuring of benthicforaminiferal assemblages Terra Nova 14 101ndash107 2002
Ramette A Multivariate analyses in microbial ecology FEMS Mi-crobiol Ecol 62 142ndash160 2007
Rodriacuteguez-Tovar F J Uchman A MrsquoHamdi A Riahi Sand Ismail-Lattrache K B Ichnological record of palaeoen-vironment from the CretaceousndashPaleogene boundary intervalat El Kef Tunisia The first study of old and new sec-tions at the stratotype area J Afr Earth Sci 120 23ndash30doi101016jjafrearsci201604017 2016
Saner S Mudurnu-Goumlynuumlk Havzasının Jura ve Sonrasıccediloumlkelimnitelikleriyle paleocografya yorumlaması TJK Buumllteni 23 39ndash52 1980
Schioslashler P Brinkhuis H Roncaglia L and Wilson G J Di-noflagellate biostratigraphy and sequence stratigraphy of theType Maastrichtian (Upper Cretaceous) ENCI Quarry TheNetherlands Mar Micropaleontol 31 65ndash95 1997
Schueth J D Bralower T J Jiang S and Patzkowsky ME The role of regional survivor incumbency in the evolu-tionary recovery of calcareous nannoplankton from the Creta-ceousPaleogene (KPg) mass extinction Paleobiology 41 661ndash679 doi101017pab201528 2015
Scotese C R A continental drift flipbook J Geology 112 729ndash741 2004
Scotese C R and Dreher C GlobalGeology available at httpwwwGlobalGeologycom (last access 11 March 2014) 2012
Sepkoski J J Patterns of Phanerozoic extinction a perspectivefrom global data bases in Global Events and Event Stratigra-phy edited by Walliser O H Springer Berlin Germany 35ndash52 2016
Sibert E C Hull P M and Norris R D Resilience of Pacificpelagic fish across the CretaceousPalaeogene mass extinctionNat Geosci 7 667ndash670 doi101038NGEO2227 2014
Slimani H and Toufiq A A CretaceousndashPaleogene boundary geo-logical site revealed by planktic foraminifera and dinoflagellatecysts at Ouled Haddou eastern external Rif Chain Morocco JAfr Earth Sci 88 38ndash52 doi101016jjafrearsci2013080082013
Slimani H Louwye S and Toufiq A Dinoflagellate cystsform the CretaceousndashPaleogene boundary at Ouled Had-dou southeastern Rif Morocco biostratigraphy paleoenvi-ronments and paleobiogeography Palynology 34 90ndash124doi10108001916121003629933 2010
Sluijs A and Brinkhuis H A dynamic climate and ecosystemstate during the Paleocene-Eocene Thermal Maximum infer-ences from dinoflagellate cyst assemblages on the New JerseyShelf Biogeosciences 6 1755ndash1781 doi105194bg-6-1755-2009 2009
Smit J Extinction and evolution of planktonic foraminifera after amajor impact at the CretaceousTertiary boundary Geol S AmS 190 329ndash352 1982
Smit J and Romein A J T A sequence of events across theCretaceous-Tertiary boundary Earth Planet Sci Lett 74 155ndash170 1985
Speijer R P Extinction and recovery patterns in ben-thic foraminiferal paleocommunities across the Creta-ceousPaleogene and PaleoceneEocene boundaries GeologicaUltraiectina 124 Faculteit Aardwetenschappen PhD ThesisUniversiteit Utrecht Utrecht the Netherlands 1994
Speijer R P and van Der Zwaan G J Extinction and survivor-ship of southern Tethyan benthic foraminifera across the Cre-
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
900 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
taceousPaleogene boundary in Biotic recovery from mass ex-tinction events edited by Hart M B Geolog Soc Spec PublLondon UK 102 343ndash371 1996
Stemmann L and Boss E Plankton and Particle Size andPackaging From Determining Optical Properties to Drivingthe Biological Pump Annu Rev Mar Sci 4 263ndash290doi101146annurev-marine-120710-100853 2012
Thomas E Late Cretaceous-early Eocene mass extinctions in thedeep sea Geol S Am S 247 481ndash495 1990
Vandenberghe N Hilgen F J and Speijer R P The PaleogenePeriod in The Geologic Time Scale 2012 edited by GradsteinF M Ogg J G Schmitz M D and Ogg G M 2 855ndash921Elsevier Boston USA 2012
Vellekoop J Sluijs A Smit J Schouten S Weijers J WH Sinninghe Damsteacute J S and Brinkhuis H Rapid short-term cooling following the Chicxulub impact at the CretaceousndashPaleogene boundary P Natl Acad Sci USA 111 7537ndash7541doi101073pnas1319253111 2014
Vellekoop J Smit J van de Schootbrugge B Weijers J W HGaleotti S Sinninghe Damsteacute J S and Brinkhuis H Palyno-logical evidence for prolonged cooling along the Tunisian conti-nental shelf following the KndashPg boundary impact PalaeogeogrPalaeocl 426 216ndash228 doi101016jpalaeo2015030212015
Vellekoop J Esmeray-Senlet S Miller K G Browning J VSluijs A van de Schootbrugge B Sinninghe Damsteacute J Sand Brinkhuis H Evidence for CretaceousndashPaleogene bound-ary bolide ldquoimpact winterrdquo conditions from New Jersey USAGeology 44 619ndash622 doi101130G379611 2016
Woelders L and Speijer R Stable seafloor conditions sealevel and food supply during the latest Maastrichtian atBrazos River Texas Mar Micropaleontol 121 41ndash51doi101016jmarmicro201510002 2015
Zachos J C Arthur M A and Dean W E Geochemical evi-dence for suppression of pelagic marine productivity at the Cre-taceousTertiary boundary Nature 337 61ndash64 1989
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
- Abstract
- Introduction
- Geological setting and age assessment
- Materials and methods
-
- Sampling
- Foraminiferal analysis
- Palynological analysis
- Statistical analysis
-
- Results
-
- Benthic foraminifera
- Palynology
-
- Discussion
-
- Tethyan benthic foraminiferal turnover sequence
- Ecological responses to reduced export productivity
-
- Okccedilular section
- Western Tethys
- Global responses
-
- Long-term recovery
-
- Conclusions
- Data availability
- Author contributions
- Competing interests
- Acknowledgements
- References
-

890 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
Figure 3 The organic-walled dinoflagellate cyst record of theOkccedilular section The biostratigraphy and bulk carbonate stable car-bon isotope records are from Accedilikalin et al (2015) The four maindinocyst complexes are indicated
4 Results
41 Benthic foraminifera
The benthic foraminiferal record of the Okccedilular section ischaracterized by a distinct turnover across the KndashPg bound-ary (Figs 2 4 5 6) Of the common taxa 8 out of 30(sim 27 ) disappear across the KndashPg boundary After the KndashPg boundary crisis three new taxa appear The estimatedBFAR shows a major decrease across the boundary (Fig 5)For the foraminiferal counts and illustrations of commonforms see Dataset S1 and Figs S2 S3
The cluster analysis allows the identification of fourmain clusters of benthic foraminiferal taxa clusters A toD (Fig 2) Cluster C is relatively large and can be subdi-vided into five subclusters Based on the succession of ben-thic faunal assemblages characterized by strong changes inthe Shannon diversity index (H ) four intervals can be rec-ognized in the benthic foraminiferal record of the Okccedilularsection (Figs 4 6)
The first interval comprises the uppermost Maastrichtianand is characterized by relatively high diversity dominatedby the taxa in cluster D In this assemblage the bi-triserialbenthic taxa are relatively abundant (22ndash28 ) with char-acteristic taxa such as Bulimina arkadelphiana Eouvigerinasubsculptura and Praebulimina reussi About 50ndash60 of theassemblage consists of inferred endobenthic taxa The KndashPg boundary marks an abrupt benthic community impover-
ishment and the decimation of the taxa in cluster D Abovethe boundary the bi-triserial benthic taxa virtually disappearfrom the record (mostly below lt 05 )
The second interval characterized by low diversity com-prises the lowermost Danian approximately correspondingto planktic foraminiferal Zone P0 In this interval epibenthicforms are most abundant (70ndash90 ) It is dominated by thetaxa of cluster A encompassing successive peak occurrencesof the taxa Anomalinoides praeacutus Trochammina sppand Cibicidoides pseudoacutus
The third interval covers the part of the succession approx-imately correlative to planktic foraminiferal Zone Pα Thisinterval is characterized by a recovery of diversity althoughthe diversity is still lower than that of the top Maastrichtianassemblage The abundance of taxa in cluster A slowly de-creases and taxa in cluster B mainly represented by Osan-gularia plummerae and Cibicidoides sp become abundantEndobenthic forms recover and make up 30ndash40 of the totalassemblage
The fourth interval starts in the interval correlative to ZoneP1a Here the benthic community has recovered as the di-versity has stabilized and is almost similar to pre-impact val-ues This interval is characterized by taxa in cluster C a typ-ical Paleocene Midway-type fauna (Berggren and Aubert1975) with representatives such as Anomalinoides praea-cutus Coryphostoma midwayensis Cibicidoides alleni andOsangularia plummerae
42 Palynology
Palynological samples from the Okccedilular site yield abundantpalynomorphs dominated by dinocysts and with minor con-tributions of acritarchs prasinophytes organic foraminiferallinings and terrestrial palynomorphs (Dataset S2) Thedinocyst associations of the MudurnundashGoumlynuumlk basin arequite diverse including components characteristic of boththe Tethyan and Boreal realms (Accedilikalin et al 2015) As ex-pected the dinocyst record does not show major changes indiversity across the KndashPg boundary (Fig 4) There is a steadydecrease in dinocyst diversity from planktic foraminiferalZone P1a upwards probably a long-term change not re-lated to the impact The estimated dinocyst accumulationrate (DAR number of preserved cysts per cm2 per kyr seeText S2 in the Supplement for details on the estimation of theDAR) shows no major changes across the KndashPg boundary
Throughout the palynological record the Spiniferites com-plex is consistently the most dominant morphogroup in gen-eral comprising 40ndash50 of the total assemblage (Fig 3)Similar to other KndashPg boundary sites worldwide (Habib andSaeedi 2007) Manumiella spp show an episode of higherrelative abundance near the top of the Maastrichtian some30ndash40 cm below the KndashPg boundary Hexaperidinioids gen-erally make up a relatively small component of the Maas-trichtian assemblage (3ndash17 ) but show a strong increaseacross the KndashPg boundary In the boundary clay layer cor-
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
J Vellekoop et al Ecological response to KndashPg boundary mass extinction 891
Figure 4 Diversity indices on the benthic foraminiferal and dinocyst records of the Okccedilular section with the Shannon diversity index (H ) thespecies richness per sample (S) the number of specimen observed per sample (N ) and the BergerndashParker index As the bulk stable carbonisotope record and the Shannon diversity index of the benthic foraminiferal record show similar trends the combination of these records canbe used to subdivide the KndashPg boundary transition into four intervals I the Maastrichtian ie ldquopre-impactrdquo interval II the post-impactinterval representing the ldquodisasterrdquo phase III a recovery interval and IV the early Paleocene post-recovery interval The division criteriaare explained in the text
relative to planktic foraminiferal Zone P0 this group in-creases to 35 of the assemblage In the peak intervals thehexaperidinioids are mostly represented by representativesof the genera Senegalinium and Cerodinium After an ini-tial drop in relative abundance in the upper half of Zone P0this group reaches a second maximum (sim 35 ) at the baseof Zone Pα and remains relatively abundant (15ndash30 ) up toZone P1a After this the hexaperidinioids gradually decreasetowards the top of the studied interval The ldquoother dinocystsrdquogroup ndash with representatives such as Areoligera spp Im-pagidinium spp Hystrichosphaeridium tubiferum Opercu-lodinium centrocarpum and Palynodinium grallator ndash gen-erally makes up 25ndash50 of the assemblage
5 Discussion
51 Tethyan benthic foraminiferal turnover sequence
The major turnover in the benthic community at Okccedilularis largely comparable with the previously published ben-thic foraminiferal record (size fraction gt 125 microm) of the KndashPg boundary GSSP of El Kef (Figs 7 8 Speijer and Vander Zwaan 1996) In addition other benthic foraminiferalrecords from the margins of the Tethys also show a strong
similarity to the Okccedilular and El Kef records including stud-ies that use the gt 63 microm size fraction (Alegret et al 2003Culver 2003 and references therein)
Although it is sometimes suggested that different sizefractions cannot be directly compared because the differ-ent size fractions sometimes harbor different ecological in-formation (Giusberti et al 2016) intercomparison studieshave shown that when comparing the gt 125 and gt 63 microm sizefractions in benthic foraminiferal assemblages the overallpatterns are generally robust (Ernst et al 2006) The factthat gt 63 and gt 125 microm KndashPg boundary benthic foraminiferalrecords from different Tethys-margin sites (eg Speijer andVan der Zwaan 1996 Peryt et al 2002 Culver et al 2003)show striking similarities even though both the size fractionsused (gt 63 vs gt 125 microm) and specific taxa making up theforaminiferal assemblages differ among sites confirms theobservations by Ernst et al (2006) Furthermore even whenit would be assumed that different size fractions are diffi-cult to compare directly the striking similarities between theTethyan gt 125 and gt 63 microm size fraction KndashPg boundary ben-thic foraminiferal records provide compelling evidence for astrong disruption of the entire benthic community
Tethyan middle neritic to middle bathyal KndashPg boundarysuccessions generally show a sequence of (1) a typical high
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
892 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
Figure 5 Estimated benthic foraminiferal accumulation rate (BFAR number of foraminifera of the gt 63 microm size fraction per cm2 sea floorper kyr) and estimated dinocyst accumulation rate (DAR number of preserved cysts per cm2 sea floor per kyr) of the Okccedilular sectionUncertainty in estimated accumulation rates resulting from uncertainties in sediment densities and sedimentation rates are indicated in greyproviding a range of estimated accumulation rates For more detailed info on the calculation of the BFAR and DAR see Text S2 of theSupplement
diversity assemblage in the Maastrichtian (2) a low diver-sity ldquodisasterrdquo assemblage directly after the KndashPg boundaryimpact characterized by a short-lived bloom of endobenthicforms observed in several high-resolution records (eg Coc-cioni and Galeotti 1994 Speijer and van der Zwaan 1996)followed by a dominance of epibenthic forms (3) a ldquore-coveryrdquo assemblage characterized by an increasing diversityand returning endobenthic forms and (4) a new high diver-sity assemblage dominated by a Paleocene fauna with bothepi- and endobenthic forms equally represented
Hence based on this succession of benthic assemblageswhich are characterized by strong changes in diversity (H )the KndashPg boundary benthic foraminiferal records from theTethys can be subdivided into four intervals (Figs 2 4ndash6)The intervals I to IV approximately correspond to the up-permost Maastrichtian planktic foraminiferal Zone P0 sub-zones PαndashP1a and subzones P1andashP1b respectively There-fore this succession roughly follows the 5-fold sedimen-tological and planktic foraminiferal sequence of Smit andRomein (1985) in which unit 2 represents the ejecta layerand units 1 3 4 and 5 correspond to intervals I to IV in ourstudy
The sequence of benthic foraminiferal turnover in theTethys is also well illustrated by a Q-mode NM-MDS
(Fig 7) The Q-mode NM-MDS of the benthic foraminiferalsample compositions of the combined data from Okccedilular(this study) and El Kef (Speijer and van der Zwaan 1996)demonstrates that although the assemblages differ betweenthese localities on opposite margins of the Tethys the re-sponses of the benthic foraminiferal assemblages to the KndashPgboundary perturbations express very similar overall patternsacross the KndashPg boundary At both Okccedilular and El Kef thebenthic foraminiferal records show a rapid transition from astable Maastrichtian assemblage to the earliest Danian dis-aster phase in Zone P0 followed by a gradual change backtowards conditions similar to the Maastrichtian (marked bythe return of Lazarus taxa) Therefore both the successionof benthic faunal assemblages including diversity indicesand Q-mode NM-MDS analysis provide powerful tools to al-low the comparison of KndashPg boundary benthic foraminiferalrecords from different localities
52 Ecological responses to reduced export productivity
521 Okccedilular section
The palynological benthic foraminiferal and bulk stable iso-tope records of the Okccedilular section show major changesacross the KndashPg boundary interval (Figs 2ndash7) portraying
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
J Vellekoop et al Ecological response to KndashPg boundary mass extinction 893
Figure 6 Relative abundances of the four main clusters of benthic foraminiferal taxa (gt 125 microm) relative abundances of epibenthic andendobenthic forms and relative abundance of bi-triserial benthic taxa
Figure 7 Q-mode non-metric multidimensional scaling (on all sam-ples in the gt 125 microm size fraction benthic foraminiferal datasets ofOkccedilular (this study) and El Kef (Speijer and van der Zwaan 1996)After an abrupt transition from latest Maastrichtian pre-impact as-semblages to disaster assemblages in Zone P0 and recovery assem-blages in Zone Pα a gradual transition towards a new equilibriumpost-recovery assemblage can be observed in subzones P1a and P1b
the biological crisis following the impact Following the KndashPg boundary mass extinction endobenthic forms stronglydecrease with bi-triserial benthic taxa almost disappearingfrom the benthic community simultaneously the estimatedBFAR shows a major fall (Fig 5) This interval approxi-mately correlative to foraminiferal Zone P0 represents thedisaster phase The drop in bi-triserial benthic taxa (Fig 6)and decrease in BFAR (Fig 5) point to a reduction in foodsupply to the benthic community indicating that the trans-port of organic matter to the sea floor must have decreasedIn the dinocyst community however hexaperidinioids showa strong increase in relative abundance during this phase(Figs 3 6 7) suggesting an increase in nutrient availabilityin the photic zone while the DAR remains relatively stableacross the KndashPg boundary (Fig 5)
The decrease in benthic diversity and the high relativeabundances of opportunistic Trochammina spp and Cibici-doides pseudoacutus suggest that the benthic community ex-perienced additional stress during Zone P0 besides food lim-itation (see Jorissen et al 2007) If food limitation were theonly factor involved upper bathyal assemblages as observedat Okccedilular would be expected to become more similar tooligotrophic middle to lower bathyal assemblages Insteadthe opposite is true the assemblage composition of the dis-aster phase is closer to neritic Midway-type assemblages
It was previously suggested that the epibenthic taxon Cibi-cidoides pseudoacutus could potentially be hypoxia resistant
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
894 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
(Speijer and van der Zwaan 1996) As a collapse of the bi-ological pump should lead to enhanced remineralization oforganic matter in the photic zone it is indeed likely thatthe oxygen minimum zones expanded and shoaled leadingto oxygen stress on the sea floor Although early Paleocenedysoxia at El Kef is supported by the ostracod record (Donzeet al 1982) and ichnofacies analyses (Rodriguez-Tovaret al 2016) supporting (eg geochemical) evidence fordysoxia during the disaster phase at Okccedilular record is absentFurthermore an additional stressor for the benthic commu-nity during Zone P0 may be present In contrast to calcare-ous nanoplankton organic-walled cyst-producing dinoflagel-lates did not suffer extinctions and may have become a moreimportant component in the early Paleocene phytoplanktoncommunity Hence besides the reduction of the amount oforganic matter transported to the sea floor the compositionof food supplied by the photic zone likely changed signifi-cantly across the KndashPg boundary as well (DrsquoHondt 2005)possibly presenting an additional stress factor for the benthiccommunity (Speijer and van der Zwaan 1996 Alegret andThomas 2009)
522 Western Tethys
The patterns in benthic foraminiferal and dinoflagellate re-sponse to the KndashPg boundary perturbations at Okccedilular appearto be characteristic for the continental margins of the westernTethyan realm The benthic foraminiferal patterns are similaracross different size fractions suggesting that they are repre-sentative of the overall benthic communities In Tunisia andSpain where both benthic foraminiferal and dinoflagellaterecords are available the earliest Danian is also characterizedby a decrease in endobenthic foraminifera at the sea floor(Speijer and Van der Zwaan 1996 Peryt et al 2002 Ale-gret et al 2003) and simultaneously blooms of hexaperidin-ioids in the water column (Brinkhuis et al 1998 Vellekoopet al 2015 Fig 8) In the Ouled Haddou section Moroccoin the westernmost Tethys (Fig 1) the lowermost Danian isalso characterized by a strong increase of the hexaperidin-ioid Senegalinium group up to 30 of the assemblage (Sli-mani et al 2010) that is very similar to the Okccedilular record(Fig 8)
The KndashPg boundary dinocyst record of El Kef is strikinglysimilar to the one from the Okccedilular section showing two dis-tinct peaks in hexaperidinioids one in Zone P0 and one atthe basal part of Zone Pα At El Kef the initial post-impactdominance of hexaperidinioids is however less pronounced(Fig 8) This difference in expression might be related tosmall differences in paleogeographic and paleoceanographicsettings between sites
In addition some high-resolution benthic KndashPg records inthe Tethyan realm show a short-lived bloom of specific en-dobenthic foraminiferal taxa in the few centimeter directlyabove the KndashPg boundary sometimes accompanied by otherindicators of low oxygen levels (eg Coccioni and Gale-
otti 1994 Speijer and Van der Zwaan 1996 Kaiho et al1999) This suggests that at least locally the sea floor wastemporarily characterized by hypoxic conditions directly fol-lowing the impact This short-lived deoxygenation is possi-bly related to the mass mortality at the KndashPg boundary re-sulting in a large short-lived flux of organic matter to thesea floor (Coccioni and Galeotti 1994) However since ex-port productivity was greatly reduced after the KndashPg bound-ary mass extinction very little ldquonewrdquo organic matter reachedthe sea floor after the initial post-impact influx Hence afteraccumulated organic matter was remineralized the benthiccommunity starved resulting in a transition to an epibenthic-dominated benthic fauna similar to the post-impact fauna ofthe Okccedilular record
The combined dinocyst records suggest that the early Pa-leocene margins of the Tethys were characterized by an in-crease in nutrient availability in the photic zone whereascoeval benthic foraminiferal records indicate a major de-crease in nutrient supply to the sea floor These changesin the benthic foraminiferal and dinocyst communities arelikely caused by the major reduction of both the efficiencyand strength of the biological pump (ie the fraction ofprimary production exported from the photic zone and theamount of organic matter transported down) in accordancewith the Living Ocean model (DrsquoHondt and Zachos 1998DrsquoHondt et al 1998 DrsquoHondt 2005 Coxall et al 2006)For a detailed explanation of the difference between biologi-cal pump efficiency and biological pump strength see Hiltinget al (2008)
As bulk carbonate δ13C reflects the isotopic compositionof the surface ocean which is set by burial fractions and bythe photosynthetic isotope effect (Kump 1991 Hain et al2014) the excursion recorded in bulk δ13C records world-wide and the rapid collapse in surface to deep-ocean carbonisotope gradients likely reflect the reduction of the global in-tensity (ie efficiency) of the biological pump (Hain et al2014) The recorded inverse change in nutrient availabilitysuggests a causal link A reduced efficiency of the biologi-cal pump and associated decrease of the fraction of biomasstransported out of the photic zone could have resulted inhigh rates of nutrient recycling in the upper part of the wa-ter column (DrsquoHondt 2005) This suggests that the reduc-tion of the biological pump strength recorded by the benthicforaminiferal record is a consequence of the decreased effi-ciency of the biological pump recorded by the carbon iso-tope and dinoflagellate cyst records The strong correlationbetween the bulk carbon isotope curves and the Shannondiversity index (H ) of the benthic foraminiferal records atboth Okccedilular (R2
= 073 p lt 0001) and El Kef (R2= 074
p lt 0001) suggest a clear link between the changes in ben-thic foraminiferal assemblages and the collapse and recoveryof biological pump efficiency
As a result of the reduced biological pump efficiency morenutrients remained available for the early Paleocene phyto-plankton community At Tethyan margin sites this is indi-
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
J Vellekoop et al Ecological response to KndashPg boundary mass extinction 895
Figure 8 The benthic foraminiferal (gt 125 microm) dinocyst and bulk stable carbon isotope records of Okccedilular (Turkey) and El Kef (Tunisia)the dinocyst record of Ouled Haddou (Morocco) and the benthic foraminiferal record (gt 63 microm) of ODP Hole 1049C Blake Nose (NorthAtlantic) Note that the definition of ldquohexaperidinioidsrdquo used is provided in the text The four phases identified based on the bulk stablecarbon isotopes and the Shannon diversity index (H ) of the benthic foraminiferal records are indicated (IndashIV) The gt 63 microm size fractionbenthic foraminiferal record of ODP Hole 1049C is from Alegret and Thomas (2004) who argued that the lowermost Paleocene of thissite comprised reworked foraminifera This interval is indicated by a dashed bar in the figure The biostratigraphy of Ouled Haddou is fromSlimani and Toufiq (2013) whereas the dinocyst record of this site is from Slimani et al (2010) The biostratigraphy of El Kef is based onBrinkhuis et al (1998) Speijer and van der Zwaan (1996) and Molina et al (2006) The fine-fraction stable carbon isotope record (blackdots) of El Kef is from Keller and Lindinger (1989) The Shannon diversity index (red dots) relative abundances of bi-triserial benthic taxa(dark purple) and all endobenthic foraminifera (light purple) are based on the gt 125 microm size fraction foraminiferal data of Speijer and vander Zwaan (1996) The palynological data of El Kef are from Brinkhuis et al (1998) (lower part) and Guasti et al (2005) (upper part) TheShannon diversity index of the gt 125 microm size fraction benthic foraminiferal record (red dots) the relative contribution of bi-triserial benthictaxa (dark purple) all endobenthic foraminifera (light purple) and hexaperidinioid dinocysts from Okccedilular are from this study The bulkstable carbon isotope record of Okccedilular (black dots) is from Accedilikalin et al (2015) Mbsf indicates meters below sea floor
cated by the higher abundance of hexaperidinioids Similarlyblooms of eutrophic survivor taxa of calcareous nanoplank-ton at open ocean sites have been suggested to be relatedto the buildup of nutrients in the open ocean photic zone(Schueth et al 2015) Although the carbon isotope gradientsbetween surface and deep waters indicate that the recoveryof the biological pump took hundreds of thousands of years(Zachos et al 1989 Kump 1991 DrsquoHondt et al 1998 Cox-all et al 2006 Birch et al 2016) our records suggest thatthe enhanced recycling of nutrients in the Tethys was partic-ularly intense during two phases in the first tens of thousandsyears after the impact (Fig 8)
523 Global responses
The decrease in export productivity following the KndashPgboundary is not only recognized in the Tethyan realm In theAtlantic Ocean Southern Ocean and the Indian Ocean a de-crease of the biological pump strength after the KndashPg bound-ary was also observed in neritic to abyssal environments(eg Thomas 1990 Olsson et al 1996 Hull et al 2011Alegret et al 2012) As an example the gt 63 microm size frac-tion benthic foraminiferal KndashPg boundary record of BlakeNose (ODP Hole 1049C northwestern Atlantic Alegret andThomas 2004) shows a pattern that is strikingly similar tothe gt 125 microm size fraction records in the Tethyan realm witha strong decrease of endobenthics in Zone P0 followed by a
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
896 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
gradual recovery across zones Pα and P1a (Fig 8) The simi-larities between the benthic foraminiferal patterns of Tethyanouter neritic to middle bathyal (200ndash1500 m) sites and thoseof the middle to lower bathyal (1500ndash1600 m) record ofBlake Nose (Alegret and Thomas 2004) suggest that theserecords register a global decrease in export productivity fol-lowing the KndashPg boundary
However several open ocean sites mostly in the Pacificshow an opposite trend with several lines of evidence sug-gesting an increased biological pump strength at these sites(eg Hull et al 2011 Alegret et al 2012) This led Ale-gret et al (2012) to conclude that the decrease in biologicalpump strength after the KndashPg boundary was most likely aregional instead of a global effect arguing against the Liv-ing Ocean hypothesis We would argue that low-productivityopen ocean sites like the central Pacific are characterized byvery different ecosystem structures and higher productivityas those along the Tethyan margins (eg Dortch and Pack-erd 1989) Therefore the consequence of ecosystem reorga-nization resulting from the mass extinction was likely verydifferent in the open ocean Pacific (eg Sibert et al 2014)Esmeray-Senlet et al (2015) proposed the term ldquoHeteroge-neous Oceanrdquo for this conceptual model as an alternativeto the Living Ocean model Supposedly the HeterogeneousOcean was characterized by a strong geographic heterogene-ity in the extinction patterns and food supply to the sea floor
However as the primary food source for open ocean deep-sea benthic communities generally consists of refractory or-ganic matter (eg Jiao et al 2010) it remains uncertainwhether changes in the intensity of recycling of the gener-ally more labile organic matter in the surface ocean will re-sult in changes in the amount of organic matter that benthiccommunities receive as they will likely continue to receiverecalcitrant organic matter at barely diminished rates despitethe collapse of the biological pump
In addition it should be noted that although the bio-logical pump efficiency is generally relatively high in openocean realms such as the central Pacific the biological pumpstrength is generally very low at these sites (Honjo et al2008 Henson et al 2011) Small changes in absolute bio-logical pump strength at such a locality could therefore havehad a large effect on the local oligotrophic benthic commu-nity (Alegret and Thomas 2009) but they likely representedonly a small fraction of the net amount of organic matterglobally transported to the deep Hence even though localbiological pump strength might have increased at some low-productivity deep-sea sites the large decrease in biologi-cal pump strength recorded at many other sites means thatthe net amount of organic matter globally transported out ofthe surface ocean was likely still reduced in the post-impactworld (Birch et al 2016) Therefore we argue that such Pa-cific records do not represent the general pattern The globalocean response can be characterized as heterogeneous how-ever the KndashPg boundary mass extinction did lead to a strongreduction of the net amount of organic matter globally trans-
ported from the surface to the sea floor while biological pro-ductivity recovered rapidly after the extinction event in ac-cordance with the Living Ocean hypothesis
53 Long-term recovery
Following the disaster phase the hexaperidinioid cysts atOkccedilular and El Kef decreased in abundance and the abun-dance of endobenthic forms increased again as the recoveryof the benthic community was initiated This recovery phaseis approximately correlative to Zone Pα and the basal part ofZone P1a which according to the Paleogene age constraintsof Vandenberghe et al (2012) represents at least 300 kyrThis duration is in agreement with the estimations providedby Birch et al (2016) During this phase the carbon iso-tope records remain well below pre-impact values and thediversity of the benthic foraminiferal community has not yetfully recovered (Figs 4ndash8) This indicates that the rapid andshort-lived KndashPg boundary disaster was followed by a rela-tively long recovery phase in line with previous estimatesof a multimillion-year biological recovery (eg Coxall etal 2006) While the impact-related environmental perturba-tions were short lived (Kring 2007 Vellekoop et al 20142016) the extinctions amongst important biological groupsled to a reduction of the organic flux from the photic zone todeep water resulting in major long-term biological and pa-leoceanographic reorganizations Only with the evolutionaryrecovery of the pelagic community governing the biologicalcarbon pump did export productivity start to increase again(eg Coxall et al 2006 Birch et al 2016)
6 Conclusions
The marine palynological benthic foraminiferal and bulkstable isotope records of the Okccedilular and El Kef sections re-veal major changes across the KndashPg boundary interval por-traying the biological crisis at the KndashPg boundary and subse-quent recovery in the early Paleocene Based on the succes-sion of benthic faunal assemblages at a number of Tethyanmargin sites four phases can be recognized across the KndashPgboundary interval the Maastrichtian or pre-impact phase (I)a disaster phase (II) a recovery phase (III) and an early Pa-leocene post-recovery phase (IV)
Following the KndashPg boundary impact some localitiesshow a large short-lived flux of food to the sea floor likelyrelated to the mass mortality at the KndashPg boundary Howeversince export productivity was greatly reduced after the KndashPgboundary mass extinction it is likely that after the initialpost-impact influx very little fresh organic matter reachedthe sea floor This reduction of export productivity in thepost-extinction disaster phase eventually resulted in a lowerfood supply to the sea floor This presented a major stressfactor for benthic organisms in the western Tethys The re-duced food supply resulted in an abrupt impoverishment of
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
J Vellekoop et al Ecological response to KndashPg boundary mass extinction 897
benthic communities As the downward transport of nutri-ents was slowed down recycling in upper layers increasedAs a result more nutrients remained available for the earlyPaleocene phytoplankton community leading to blooms ofdinoflagellates along the Tethyan margins and blooms of cal-careous nanoplankton taxa in more open ocean sites Ourrecords show that the enhanced recycling of nutrients in theTethys was particularly intense in the tens of thousands ofyears after the impact Following this the slow evolution-ary recovery of the pelagic community governing the bio-logical carbon pump resulted in a gradually increasing ex-port productivity in the hundreds of thousands of years afterthe impact Hence our integration of dinocyst and benthicforaminiferal records across the KndashPg boundary provides im-portant new insights into the ecological responses to the re-duction of export productivity following the mass extinctionat the CretaceousndashPaleogene boundary highlighting the di-rect ecological consequences of the Living Ocean conditionsin the post-impact world
7 Data availability
All data related to publication are available in the Supple-ment
The Supplement related to this article is available onlineat doi105194bg-14-885-2017-supplement
Author contributions Johan Vellekoop Lineke Woelders andHenk Brinkhuis designed the research Samples were collectedin the field by Johan Vellekoop Sanem Accedilikalin Ismail Ouml Yil-maz and Jan Smit Palynological analyses were carried out by Jo-han Vellekoop Benthic foraminiferal analyses were carried out byLineke Woelders and Robert P Speijer Johan Vellekoop LinekeWoelders and Bas van de Schootbrugge prepared the manuscriptwith input from all authors
Competing interests The authors declare that they have no conflictof interest
Acknowledgements This work was supported by the NetherlandsOrganization for Scientific Research (NWO) Open CompetitionGrant ALWPJ09047 to H Brinkhuis and the Research FoundationFlanders (FWO) grant G0B8513 to R P Speijer We thank LKump F Boscolo Galazzo P Roopnarine and one anonymousreferee for their suggestions
Edited by D GillikinReviewed by L Kump F Boscolo Galazzo and P Roopnarine
References
Accedilikalin S Vellekoop J Ocakoglu F Yılmaz I Ouml Smit J Al-tiner S O Goderis S Vonhof H Speijer R P WoeldersL Fornaciari E and Brinkhuis H Geochemical and pale-ontological characterization of a new KndashPg Boundary local-ity from the Northern branch of the Neo-Tethys MudurnundashGoumlynuumlk Basin NW Turkey Cretaceous Res 52 251ndash267doi101016jcretres201407011 2015
Alegret L and Thomas E Food supply to the seafloor in the Pa-cific Ocean after the CretaceousPaleogene boundary event MarMicropaleontol 73 105ndash116 2009
Alegret L Molina E and Thomas E Benthic foraminiferalturnover across the CretaceousPaleogene boundary at Agost(southeastern Spain) paleoenvironmental inferences Mar Mi-cropaleontol 48 251ndash279 2003
Alegret L Thomas E and Lohmann K C End-Cretaceous ma-rine mass extinction not caused by productivity collapse P NatlAcad Sci USA 109 728ndash732 doi101073pnas11106011092012
Alegret L Rodriacuteguez-Tovar F J and Uchman A How bioturba-tion obscured the CretaceousndashPaleogene boundary record TerraNova 27 225ndash230 doi101111ter12151 2015
Altiner D Koccedilyigit A Farinacci A Nicosia U and ContiM A Jurassic-Lower Cretaceous stratigraphy and paleogeo-graphic evolution of the southern part of north-western Anatolia(Turkey) Geologica Romana 27 13ndash80 1991
Alvarez LW Alvarez W Asaro F and Michel H V Extrater-restrial cause for the Cretaceous-Tertiary extinction Science208 1095ndash1108 1980
Alegret L and Thomas E Benthic foraminifera and environmen-tal turnover across the CretaceousPaleogene boundary at BlakeNose (ODP Hole 1049C Northwestern Atlantic) PalaeogeogrPalaeocl 208 59ndash83 doi101016jpalaeo200402028 2004
Armstrong R A Lee C Hedges J I Honjo S and WakehamS G A new mechanistic model for organic carbon fluxes in theocean based on the quantitative association of POC with ballastmaterials Deep-Sea Res Pt II 49 219ndash236 2002
Aubert J and Berggren W A Paleocene benthic foraminiferalbiostratigraphy and paleoecology of Tunisia Bull du Cent deRech de Pau ndash SNPA 10 379ndash469 1976
Berggren W A and Aubert J Paleocene benthonic foraminiferalbiostratigraphy paleobiogeography and paleoecology ofAtlanticndashTethyan regions midway-type fauna PalaeogeogrPalaeocl 18 73ndash192 1975
Bernaola G and Monechi S Calcareous nannofossil ex-tinction and survivorship across the CretaceousndashPaleogeneboundary at Walvis Ridge (ODP Hole 1262C SouthAtlantic Ocean) Palaeogeogr Palaeocl 255 132ndash156doi101016jpalaeo200702045 2007
Bernhard J M Characteristic assemblages and morphologies ofbenthic foraminifera from anoxic organic-rich deposits Jurassicthrough Holocene J Foramin Res 16 207ndash215 1986
Birch H S Coxall H K Pearson P N Kroon D andSchmidt D N Partial collapse of the marine carbon pump af-ter the CretaceousndashPaleogene boundary Geology 44 287ndash290doi101130G375811 2016
Brinkhuis H and Zachariasse W J Dinoflagellate cysts sealevel changes and planktonic foraminifers across the Cretaceous-
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
898 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
Tertiary boundary at El Haria northwest Tunisia Mar Micropa-leontol 13 153ndash191 1988
Brinkhuis H Bujak J P Smit J Versteegh G J M and Viss-cher H Dinoflagellate-based sea surface temperature recon-structions across the Cretaceous-Tertiary boundary PalaeogeogrPalaeocl 141 67ndash83 1998
Buesseler K O The decoupling of production and particulate ex-port in the surface ocean Global Biogeochem Cy 12 297ndash3101998
Buzas M A Culver S J and Jorissen F J A statisti-cal evaluation of the microhabitats of living (stained) infau-nal benthic foraminifera Mar Micropaleontol 20 311ndash320doi1010160377-8398(93)90040-5 1993
Coccioni R and Galeotti S KndashT boundary extinction geolog-ically instantaneous or gradual event Evidence from deep-seabenthic foraminifera Geology 22 779ndash782 1994
Corliss B H Microhabitats of benthic foraminifera within deep-sea sediments Nature 314 435ndash438 doi101038314435a01985
Corliss B H and Chen C Morphotype patterns of Nor-wegian Sea deep-sea benthic foraminifera and ecologi-cal implications Geology 16 716ndash719 doi1011300091-7613(1988)016lt0716MPONSDgt23CO2 1988
Coxall H K DrsquoHondt S and Zachos J C Pelagic evolution andenvironmental recovery after the CretaceousndashPaleogene mass ex-tinction Geology 34 297ndash300 doi101130G217021 2006
Culver S J Benthic foraminifera across the CretaceousndashTertiary(KndashT) boundary a review Mar Micropaleontol 47 177ndash2262003
Cushman J A Upper Cretaceous foraminifera of the Gulf coastalregion of the United States and adjacent areas US Geolog SurvProf Pap 206 241 pp 1946
Cushman J A Paleocene foraminifera of the Gulf Coastal Regionof the United States and adjacent regions US Geol Surv ProfPap 232 75 pp 1951
DrsquoHondt S Consequences of the CretaceousPaleogene Mass Ex-tinction for Marine Ecosystems Annu Rev Ecol Evol S36 295ndash317 doi101146annurevecolsys350211031057152005
DrsquoHondt S and Zachos J C Cretaceous foraminifera and the evo-lutionary history of planktic photosymbiosis Paleobiology 24512ndash523 1998
DrsquoHondt S Donaghay P Zachos J C Luttenberg D andLindinger M Organic Carbon Fluxes and Ecological Recov-ery from the Cretaceous-Tertiary Mass Extinction Science 282276ndash279 1998
Donze P Colin J-P Damotte R Oertli HJ Peypouquet J-Pand Said R Les ostracodes du Campanien terminal agrave lrsquoEocegraveneinfeacuterieur de la coupe du Kef Tuneacutesie nord-occidentale Bull desCentr Recher Expl-Prod Elf Aquit 6 273ndash335 1982
Dortch Q and Packard T T Differences in biomass structure be-tween oligotrophic and eutrophic marine ecosystems Deep-SeaRes 36 223ndash240 1989
Ernst S R Guasti E Dupuis C and Speijer R P En-vironmental perturbation in the southern Tethys across thePaleoceneEocene boundary (Dababiya Egypt) Foraminiferaland clay mineral records Mar Micropaleontol 60 89ndash111doi101016jmarmicro200603002 2006
Eshet Y Moshkovitz S Habib D Benjamini C and Maga-ritz M Calcareous nannofossil and dinoflagellate stratigraphyacross the CretaceousTertiary boundary at Hor Hahar IsraelMar Micropaleontol 18 199ndash228 1992
Eshet Y Almogi-Labin A and Bein A Dinoflagellate cysts pa-leoproductivity and upwelling systems A Late Cretaceous ex-ample from Israel Mar Micropaleontol 23 231ndash240 1994
Esmeray-Senlet S Wright J D Olsson R K MillerK G Browning J V and Quan T M Evidencefor reduced export productivity following the Creta-ceousPaleogene mass extinction Paleoceanography 30718ndash738 doi1010022014PA002724 2015
Giusberti L Boscolo Galazzo F and Thomas E Variability inclimate and productivity during the Paleocene-Eocene ThermalMaximum in the western Tethys (Forada section) Clim Past12 213ndash240 doi105194cp-12-213-2016 2016
Guasti E Kouwenhoven T J Brinkhuis H and Speijer R PPaleocene sea-level and productivity changes at the southernTethyan margin (El Kef Tunisia) Mar Micropaleontol 55 1ndash17 2005
Habib D and Saeedi F The Manumiella seelandica global spikeCooling during regression at the close of the MaastrichtianPalaeogeogr Palaeocl 255 87ndash97 2007
Hain M P Sigmal D M and Haug G H 818 ndash The biolog-ical Pump in the Past in Reference Module in Earth Systemsand Environmental Sciences Treatise on Geochemistry (SecondEdition) The Oceans and Marine Geochemistry 8 485ndash517doi101016B978-0-08-095975-700618-5 2014
Hayek L C and Buzas M A On the proper and efficient useof diversity measures with individual field samples J ForaminRes 43 305ndash313 2013
Henson S A Sanders R Madsen E Morris P J Le MoigneF and Quartly G D A reduced estimate of the strength ofthe oceanrsquos biological carbon pump Geophys Res Lett 38L04606 doi1010292011GL046735 2011
Hilting A K Kump L R and Bralower T J Variations in theoceanic vertical carbon isotope gradient and their implicationsfor the Paleocene-Eocene biological pump Paleoceanography23 PA3222 doi1010292007PA001458 2008
Honjo S Manganini S J Krishfield R A and Francois RParticulate organic carbon fluxes to the ocean interior and fac-tors controlling the biological pump A synthesis of global sed-iment trap progams since 1983 Prog Oceanogr 76 217ndash285doi101016jpocean200711003 2008
Huber B T MacLeod K G and Norris R D Abrupt extinctionand subsequent reworking of Cretaceous planktonic foraminiferaacross the Cretaceous-Tertiary boundary Evidence from the sub-tropical North Atlantic in Catastrophic Events and Mass Extinc-tions Impacts and Beyond edited by Koeberl C and MacLeodK G Geol S Am S 356 277ndash289 Boulder Colorado USA2002
Hull P M Norris R D Bralower T J and Schueth J D A rolefor chance in marine recovery from the end-Cretaceous extinc-tion Nat Geosci 4 856ndash860 doi101038ngeo1302 2011
Hsuuml K J and McKenzie J A A ldquoStrangeloverdquo ocean in the earli-est Tertiary in Natural variations Archean to present edited bySundquist E T and Broecker W S The carbon cycle and atmo-spheric CO2 American Geophysical Union Washington DCUSA 487ndash492 1985
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
J Vellekoop et al Ecological response to KndashPg boundary mass extinction 899
Jiao N Herndl G J Hansell D A Benner R Kattner G Wil-helm S W Kirchman D L Weinbauer M G Luo T ChenF and Azam F Microbial production of recalcitrant dissolvedorganic matter long-term carbon storage in the global oceanNat Rev 8 593ndash599 doi101038nrmicro2386 2010
Jorissen F J de Stigter H C and Widmark J G V A concep-tual model explaining benthic foraminiferal microhabitats MarMicropaleontol 26 3ndash15 doi1010160377-8398(95)00047-X1995
Jorissen F J Fontanier C and Thomas E Paleoceanographi-cal proxies based on deep-sea benthic foraminiferal assemblagecharacteristics in Proxies in Late Cenozoic PaleoceanographyPt 2 Biological tracers and biomarkers edited by Hillaire-Marcel C and de Vernal A 263ndash326 Elsevier Amsterdamthe Netherlands 2007
Kaiho K Kajiwara Y Tazaki K Ueshima M Takeda NKawahata H Arinobu T Ishiwatari R Hirai A and Lam-olda M A Oceanic primary productivity and dissolved oxygenlevels at the CretaceousTertiary boundary their decrease sub-sequent warming and recovery Paleoceanography 14 511ndash5241999
Keller G and Lindinger M Stable isotope TOC and CaCO3record across the CretaceousTertiary boundary at El KefTunisia Palaeogeogr Palaeocl 73 243ndash265 1989
Kellough G R Paleoecology of the Foraminiferida of the WillsPoint Formation (Midway Group) in northeast Texas Trans GulfCoast Assoc Geol Soc 15 73ndash153 1965
Kring D A The Chicxulub impact event and its environmentalconsequences at the CretaceousndashTertiary boundary PalaeogeogrPalaeocl 255 4ndash21 2007
Kump L R Interpreting carbon-isotope excursions Strangeloveoceans Geology 19 299ndash302 1991
Machalski M Vellekoop J Dubicka Z Peryt D and Ha-rasimiuk M Late Maastrichtian cephalopods dinoflagellatecysts and foraminifera from the CretaceousndashPaleogene suc-cession at Lechoacutewka southeast Poland Stratigraphic andenvironmental implications Cretaceous Res 57 208ndash227doi101016jcretres201508012 2016
Molina E Evidence and causes of the main extinctionevents in the Paleogene based on extinction and survivalpatterns of foraminifera Earth-Sci Rev 140 166ndash181doi101016jearscirev201411008 2015
Molina E Alegret L Arenillas I and Arz J The GlobalBoundary Stratotype Section and Point for the base of the Da-nian Stage (Paleocene Paleogeneldquo Tertiaryrdquo Cenozoic) at ElKef Tunisia-Original definition and revision Episodes 29 263ndash273 2006
Olsson R K Liu C and van Fossen M The Cretaceous-Tertiarycatastrophic event at Millers Ferry Alabama in The Cretaceous-Tertiary Event and Other Catastrophes in Earth History editedby Ryder G Fastovsky D and Gartner S Geol S Am S307 263ndash277 1996
Peryt D Benthic foraminiferal response to the Cenomanian-Turonian and CretaceousndashPaleogene boundary events PrzegladGeol 52 827ndash832 2004
Peryt D Alegret L and Molina E The CretaceousPalaeogene(KP) boundary at Aiumln Settara Tunisia restructuring of benthicforaminiferal assemblages Terra Nova 14 101ndash107 2002
Ramette A Multivariate analyses in microbial ecology FEMS Mi-crobiol Ecol 62 142ndash160 2007
Rodriacuteguez-Tovar F J Uchman A MrsquoHamdi A Riahi Sand Ismail-Lattrache K B Ichnological record of palaeoen-vironment from the CretaceousndashPaleogene boundary intervalat El Kef Tunisia The first study of old and new sec-tions at the stratotype area J Afr Earth Sci 120 23ndash30doi101016jjafrearsci201604017 2016
Saner S Mudurnu-Goumlynuumlk Havzasının Jura ve Sonrasıccediloumlkelimnitelikleriyle paleocografya yorumlaması TJK Buumllteni 23 39ndash52 1980
Schioslashler P Brinkhuis H Roncaglia L and Wilson G J Di-noflagellate biostratigraphy and sequence stratigraphy of theType Maastrichtian (Upper Cretaceous) ENCI Quarry TheNetherlands Mar Micropaleontol 31 65ndash95 1997
Schueth J D Bralower T J Jiang S and Patzkowsky ME The role of regional survivor incumbency in the evolu-tionary recovery of calcareous nannoplankton from the Creta-ceousPaleogene (KPg) mass extinction Paleobiology 41 661ndash679 doi101017pab201528 2015
Scotese C R A continental drift flipbook J Geology 112 729ndash741 2004
Scotese C R and Dreher C GlobalGeology available at httpwwwGlobalGeologycom (last access 11 March 2014) 2012
Sepkoski J J Patterns of Phanerozoic extinction a perspectivefrom global data bases in Global Events and Event Stratigra-phy edited by Walliser O H Springer Berlin Germany 35ndash52 2016
Sibert E C Hull P M and Norris R D Resilience of Pacificpelagic fish across the CretaceousPalaeogene mass extinctionNat Geosci 7 667ndash670 doi101038NGEO2227 2014
Slimani H and Toufiq A A CretaceousndashPaleogene boundary geo-logical site revealed by planktic foraminifera and dinoflagellatecysts at Ouled Haddou eastern external Rif Chain Morocco JAfr Earth Sci 88 38ndash52 doi101016jjafrearsci2013080082013
Slimani H Louwye S and Toufiq A Dinoflagellate cystsform the CretaceousndashPaleogene boundary at Ouled Had-dou southeastern Rif Morocco biostratigraphy paleoenvi-ronments and paleobiogeography Palynology 34 90ndash124doi10108001916121003629933 2010
Sluijs A and Brinkhuis H A dynamic climate and ecosystemstate during the Paleocene-Eocene Thermal Maximum infer-ences from dinoflagellate cyst assemblages on the New JerseyShelf Biogeosciences 6 1755ndash1781 doi105194bg-6-1755-2009 2009
Smit J Extinction and evolution of planktonic foraminifera after amajor impact at the CretaceousTertiary boundary Geol S AmS 190 329ndash352 1982
Smit J and Romein A J T A sequence of events across theCretaceous-Tertiary boundary Earth Planet Sci Lett 74 155ndash170 1985
Speijer R P Extinction and recovery patterns in ben-thic foraminiferal paleocommunities across the Creta-ceousPaleogene and PaleoceneEocene boundaries GeologicaUltraiectina 124 Faculteit Aardwetenschappen PhD ThesisUniversiteit Utrecht Utrecht the Netherlands 1994
Speijer R P and van Der Zwaan G J Extinction and survivor-ship of southern Tethyan benthic foraminifera across the Cre-
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
900 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
taceousPaleogene boundary in Biotic recovery from mass ex-tinction events edited by Hart M B Geolog Soc Spec PublLondon UK 102 343ndash371 1996
Stemmann L and Boss E Plankton and Particle Size andPackaging From Determining Optical Properties to Drivingthe Biological Pump Annu Rev Mar Sci 4 263ndash290doi101146annurev-marine-120710-100853 2012
Thomas E Late Cretaceous-early Eocene mass extinctions in thedeep sea Geol S Am S 247 481ndash495 1990
Vandenberghe N Hilgen F J and Speijer R P The PaleogenePeriod in The Geologic Time Scale 2012 edited by GradsteinF M Ogg J G Schmitz M D and Ogg G M 2 855ndash921Elsevier Boston USA 2012
Vellekoop J Sluijs A Smit J Schouten S Weijers J WH Sinninghe Damsteacute J S and Brinkhuis H Rapid short-term cooling following the Chicxulub impact at the CretaceousndashPaleogene boundary P Natl Acad Sci USA 111 7537ndash7541doi101073pnas1319253111 2014
Vellekoop J Smit J van de Schootbrugge B Weijers J W HGaleotti S Sinninghe Damsteacute J S and Brinkhuis H Palyno-logical evidence for prolonged cooling along the Tunisian conti-nental shelf following the KndashPg boundary impact PalaeogeogrPalaeocl 426 216ndash228 doi101016jpalaeo2015030212015
Vellekoop J Esmeray-Senlet S Miller K G Browning J VSluijs A van de Schootbrugge B Sinninghe Damsteacute J Sand Brinkhuis H Evidence for CretaceousndashPaleogene bound-ary bolide ldquoimpact winterrdquo conditions from New Jersey USAGeology 44 619ndash622 doi101130G379611 2016
Woelders L and Speijer R Stable seafloor conditions sealevel and food supply during the latest Maastrichtian atBrazos River Texas Mar Micropaleontol 121 41ndash51doi101016jmarmicro201510002 2015
Zachos J C Arthur M A and Dean W E Geochemical evi-dence for suppression of pelagic marine productivity at the Cre-taceousTertiary boundary Nature 337 61ndash64 1989
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
- Abstract
- Introduction
- Geological setting and age assessment
- Materials and methods
-
- Sampling
- Foraminiferal analysis
- Palynological analysis
- Statistical analysis
-
- Results
-
- Benthic foraminifera
- Palynology
-
- Discussion
-
- Tethyan benthic foraminiferal turnover sequence
- Ecological responses to reduced export productivity
-
- Okccedilular section
- Western Tethys
- Global responses
-
- Long-term recovery
-
- Conclusions
- Data availability
- Author contributions
- Competing interests
- Acknowledgements
- References
-

J Vellekoop et al Ecological response to KndashPg boundary mass extinction 891
Figure 4 Diversity indices on the benthic foraminiferal and dinocyst records of the Okccedilular section with the Shannon diversity index (H ) thespecies richness per sample (S) the number of specimen observed per sample (N ) and the BergerndashParker index As the bulk stable carbonisotope record and the Shannon diversity index of the benthic foraminiferal record show similar trends the combination of these records canbe used to subdivide the KndashPg boundary transition into four intervals I the Maastrichtian ie ldquopre-impactrdquo interval II the post-impactinterval representing the ldquodisasterrdquo phase III a recovery interval and IV the early Paleocene post-recovery interval The division criteriaare explained in the text
relative to planktic foraminiferal Zone P0 this group in-creases to 35 of the assemblage In the peak intervals thehexaperidinioids are mostly represented by representativesof the genera Senegalinium and Cerodinium After an ini-tial drop in relative abundance in the upper half of Zone P0this group reaches a second maximum (sim 35 ) at the baseof Zone Pα and remains relatively abundant (15ndash30 ) up toZone P1a After this the hexaperidinioids gradually decreasetowards the top of the studied interval The ldquoother dinocystsrdquogroup ndash with representatives such as Areoligera spp Im-pagidinium spp Hystrichosphaeridium tubiferum Opercu-lodinium centrocarpum and Palynodinium grallator ndash gen-erally makes up 25ndash50 of the assemblage
5 Discussion
51 Tethyan benthic foraminiferal turnover sequence
The major turnover in the benthic community at Okccedilularis largely comparable with the previously published ben-thic foraminiferal record (size fraction gt 125 microm) of the KndashPg boundary GSSP of El Kef (Figs 7 8 Speijer and Vander Zwaan 1996) In addition other benthic foraminiferalrecords from the margins of the Tethys also show a strong
similarity to the Okccedilular and El Kef records including stud-ies that use the gt 63 microm size fraction (Alegret et al 2003Culver 2003 and references therein)
Although it is sometimes suggested that different sizefractions cannot be directly compared because the differ-ent size fractions sometimes harbor different ecological in-formation (Giusberti et al 2016) intercomparison studieshave shown that when comparing the gt 125 and gt 63 microm sizefractions in benthic foraminiferal assemblages the overallpatterns are generally robust (Ernst et al 2006) The factthat gt 63 and gt 125 microm KndashPg boundary benthic foraminiferalrecords from different Tethys-margin sites (eg Speijer andVan der Zwaan 1996 Peryt et al 2002 Culver et al 2003)show striking similarities even though both the size fractionsused (gt 63 vs gt 125 microm) and specific taxa making up theforaminiferal assemblages differ among sites confirms theobservations by Ernst et al (2006) Furthermore even whenit would be assumed that different size fractions are diffi-cult to compare directly the striking similarities between theTethyan gt 125 and gt 63 microm size fraction KndashPg boundary ben-thic foraminiferal records provide compelling evidence for astrong disruption of the entire benthic community
Tethyan middle neritic to middle bathyal KndashPg boundarysuccessions generally show a sequence of (1) a typical high
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
892 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
Figure 5 Estimated benthic foraminiferal accumulation rate (BFAR number of foraminifera of the gt 63 microm size fraction per cm2 sea floorper kyr) and estimated dinocyst accumulation rate (DAR number of preserved cysts per cm2 sea floor per kyr) of the Okccedilular sectionUncertainty in estimated accumulation rates resulting from uncertainties in sediment densities and sedimentation rates are indicated in greyproviding a range of estimated accumulation rates For more detailed info on the calculation of the BFAR and DAR see Text S2 of theSupplement
diversity assemblage in the Maastrichtian (2) a low diver-sity ldquodisasterrdquo assemblage directly after the KndashPg boundaryimpact characterized by a short-lived bloom of endobenthicforms observed in several high-resolution records (eg Coc-cioni and Galeotti 1994 Speijer and van der Zwaan 1996)followed by a dominance of epibenthic forms (3) a ldquore-coveryrdquo assemblage characterized by an increasing diversityand returning endobenthic forms and (4) a new high diver-sity assemblage dominated by a Paleocene fauna with bothepi- and endobenthic forms equally represented
Hence based on this succession of benthic assemblageswhich are characterized by strong changes in diversity (H )the KndashPg boundary benthic foraminiferal records from theTethys can be subdivided into four intervals (Figs 2 4ndash6)The intervals I to IV approximately correspond to the up-permost Maastrichtian planktic foraminiferal Zone P0 sub-zones PαndashP1a and subzones P1andashP1b respectively There-fore this succession roughly follows the 5-fold sedimen-tological and planktic foraminiferal sequence of Smit andRomein (1985) in which unit 2 represents the ejecta layerand units 1 3 4 and 5 correspond to intervals I to IV in ourstudy
The sequence of benthic foraminiferal turnover in theTethys is also well illustrated by a Q-mode NM-MDS
(Fig 7) The Q-mode NM-MDS of the benthic foraminiferalsample compositions of the combined data from Okccedilular(this study) and El Kef (Speijer and van der Zwaan 1996)demonstrates that although the assemblages differ betweenthese localities on opposite margins of the Tethys the re-sponses of the benthic foraminiferal assemblages to the KndashPgboundary perturbations express very similar overall patternsacross the KndashPg boundary At both Okccedilular and El Kef thebenthic foraminiferal records show a rapid transition from astable Maastrichtian assemblage to the earliest Danian dis-aster phase in Zone P0 followed by a gradual change backtowards conditions similar to the Maastrichtian (marked bythe return of Lazarus taxa) Therefore both the successionof benthic faunal assemblages including diversity indicesand Q-mode NM-MDS analysis provide powerful tools to al-low the comparison of KndashPg boundary benthic foraminiferalrecords from different localities
52 Ecological responses to reduced export productivity
521 Okccedilular section
The palynological benthic foraminiferal and bulk stable iso-tope records of the Okccedilular section show major changesacross the KndashPg boundary interval (Figs 2ndash7) portraying
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
J Vellekoop et al Ecological response to KndashPg boundary mass extinction 893
Figure 6 Relative abundances of the four main clusters of benthic foraminiferal taxa (gt 125 microm) relative abundances of epibenthic andendobenthic forms and relative abundance of bi-triserial benthic taxa
Figure 7 Q-mode non-metric multidimensional scaling (on all sam-ples in the gt 125 microm size fraction benthic foraminiferal datasets ofOkccedilular (this study) and El Kef (Speijer and van der Zwaan 1996)After an abrupt transition from latest Maastrichtian pre-impact as-semblages to disaster assemblages in Zone P0 and recovery assem-blages in Zone Pα a gradual transition towards a new equilibriumpost-recovery assemblage can be observed in subzones P1a and P1b
the biological crisis following the impact Following the KndashPg boundary mass extinction endobenthic forms stronglydecrease with bi-triserial benthic taxa almost disappearingfrom the benthic community simultaneously the estimatedBFAR shows a major fall (Fig 5) This interval approxi-mately correlative to foraminiferal Zone P0 represents thedisaster phase The drop in bi-triserial benthic taxa (Fig 6)and decrease in BFAR (Fig 5) point to a reduction in foodsupply to the benthic community indicating that the trans-port of organic matter to the sea floor must have decreasedIn the dinocyst community however hexaperidinioids showa strong increase in relative abundance during this phase(Figs 3 6 7) suggesting an increase in nutrient availabilityin the photic zone while the DAR remains relatively stableacross the KndashPg boundary (Fig 5)
The decrease in benthic diversity and the high relativeabundances of opportunistic Trochammina spp and Cibici-doides pseudoacutus suggest that the benthic community ex-perienced additional stress during Zone P0 besides food lim-itation (see Jorissen et al 2007) If food limitation were theonly factor involved upper bathyal assemblages as observedat Okccedilular would be expected to become more similar tooligotrophic middle to lower bathyal assemblages Insteadthe opposite is true the assemblage composition of the dis-aster phase is closer to neritic Midway-type assemblages
It was previously suggested that the epibenthic taxon Cibi-cidoides pseudoacutus could potentially be hypoxia resistant
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
894 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
(Speijer and van der Zwaan 1996) As a collapse of the bi-ological pump should lead to enhanced remineralization oforganic matter in the photic zone it is indeed likely thatthe oxygen minimum zones expanded and shoaled leadingto oxygen stress on the sea floor Although early Paleocenedysoxia at El Kef is supported by the ostracod record (Donzeet al 1982) and ichnofacies analyses (Rodriguez-Tovaret al 2016) supporting (eg geochemical) evidence fordysoxia during the disaster phase at Okccedilular record is absentFurthermore an additional stressor for the benthic commu-nity during Zone P0 may be present In contrast to calcare-ous nanoplankton organic-walled cyst-producing dinoflagel-lates did not suffer extinctions and may have become a moreimportant component in the early Paleocene phytoplanktoncommunity Hence besides the reduction of the amount oforganic matter transported to the sea floor the compositionof food supplied by the photic zone likely changed signifi-cantly across the KndashPg boundary as well (DrsquoHondt 2005)possibly presenting an additional stress factor for the benthiccommunity (Speijer and van der Zwaan 1996 Alegret andThomas 2009)
522 Western Tethys
The patterns in benthic foraminiferal and dinoflagellate re-sponse to the KndashPg boundary perturbations at Okccedilular appearto be characteristic for the continental margins of the westernTethyan realm The benthic foraminiferal patterns are similaracross different size fractions suggesting that they are repre-sentative of the overall benthic communities In Tunisia andSpain where both benthic foraminiferal and dinoflagellaterecords are available the earliest Danian is also characterizedby a decrease in endobenthic foraminifera at the sea floor(Speijer and Van der Zwaan 1996 Peryt et al 2002 Ale-gret et al 2003) and simultaneously blooms of hexaperidin-ioids in the water column (Brinkhuis et al 1998 Vellekoopet al 2015 Fig 8) In the Ouled Haddou section Moroccoin the westernmost Tethys (Fig 1) the lowermost Danian isalso characterized by a strong increase of the hexaperidin-ioid Senegalinium group up to 30 of the assemblage (Sli-mani et al 2010) that is very similar to the Okccedilular record(Fig 8)
The KndashPg boundary dinocyst record of El Kef is strikinglysimilar to the one from the Okccedilular section showing two dis-tinct peaks in hexaperidinioids one in Zone P0 and one atthe basal part of Zone Pα At El Kef the initial post-impactdominance of hexaperidinioids is however less pronounced(Fig 8) This difference in expression might be related tosmall differences in paleogeographic and paleoceanographicsettings between sites
In addition some high-resolution benthic KndashPg records inthe Tethyan realm show a short-lived bloom of specific en-dobenthic foraminiferal taxa in the few centimeter directlyabove the KndashPg boundary sometimes accompanied by otherindicators of low oxygen levels (eg Coccioni and Gale-
otti 1994 Speijer and Van der Zwaan 1996 Kaiho et al1999) This suggests that at least locally the sea floor wastemporarily characterized by hypoxic conditions directly fol-lowing the impact This short-lived deoxygenation is possi-bly related to the mass mortality at the KndashPg boundary re-sulting in a large short-lived flux of organic matter to thesea floor (Coccioni and Galeotti 1994) However since ex-port productivity was greatly reduced after the KndashPg bound-ary mass extinction very little ldquonewrdquo organic matter reachedthe sea floor after the initial post-impact influx Hence afteraccumulated organic matter was remineralized the benthiccommunity starved resulting in a transition to an epibenthic-dominated benthic fauna similar to the post-impact fauna ofthe Okccedilular record
The combined dinocyst records suggest that the early Pa-leocene margins of the Tethys were characterized by an in-crease in nutrient availability in the photic zone whereascoeval benthic foraminiferal records indicate a major de-crease in nutrient supply to the sea floor These changesin the benthic foraminiferal and dinocyst communities arelikely caused by the major reduction of both the efficiencyand strength of the biological pump (ie the fraction ofprimary production exported from the photic zone and theamount of organic matter transported down) in accordancewith the Living Ocean model (DrsquoHondt and Zachos 1998DrsquoHondt et al 1998 DrsquoHondt 2005 Coxall et al 2006)For a detailed explanation of the difference between biologi-cal pump efficiency and biological pump strength see Hiltinget al (2008)
As bulk carbonate δ13C reflects the isotopic compositionof the surface ocean which is set by burial fractions and bythe photosynthetic isotope effect (Kump 1991 Hain et al2014) the excursion recorded in bulk δ13C records world-wide and the rapid collapse in surface to deep-ocean carbonisotope gradients likely reflect the reduction of the global in-tensity (ie efficiency) of the biological pump (Hain et al2014) The recorded inverse change in nutrient availabilitysuggests a causal link A reduced efficiency of the biologi-cal pump and associated decrease of the fraction of biomasstransported out of the photic zone could have resulted inhigh rates of nutrient recycling in the upper part of the wa-ter column (DrsquoHondt 2005) This suggests that the reduc-tion of the biological pump strength recorded by the benthicforaminiferal record is a consequence of the decreased effi-ciency of the biological pump recorded by the carbon iso-tope and dinoflagellate cyst records The strong correlationbetween the bulk carbon isotope curves and the Shannondiversity index (H ) of the benthic foraminiferal records atboth Okccedilular (R2
= 073 p lt 0001) and El Kef (R2= 074
p lt 0001) suggest a clear link between the changes in ben-thic foraminiferal assemblages and the collapse and recoveryof biological pump efficiency
As a result of the reduced biological pump efficiency morenutrients remained available for the early Paleocene phyto-plankton community At Tethyan margin sites this is indi-
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
J Vellekoop et al Ecological response to KndashPg boundary mass extinction 895
Figure 8 The benthic foraminiferal (gt 125 microm) dinocyst and bulk stable carbon isotope records of Okccedilular (Turkey) and El Kef (Tunisia)the dinocyst record of Ouled Haddou (Morocco) and the benthic foraminiferal record (gt 63 microm) of ODP Hole 1049C Blake Nose (NorthAtlantic) Note that the definition of ldquohexaperidinioidsrdquo used is provided in the text The four phases identified based on the bulk stablecarbon isotopes and the Shannon diversity index (H ) of the benthic foraminiferal records are indicated (IndashIV) The gt 63 microm size fractionbenthic foraminiferal record of ODP Hole 1049C is from Alegret and Thomas (2004) who argued that the lowermost Paleocene of thissite comprised reworked foraminifera This interval is indicated by a dashed bar in the figure The biostratigraphy of Ouled Haddou is fromSlimani and Toufiq (2013) whereas the dinocyst record of this site is from Slimani et al (2010) The biostratigraphy of El Kef is based onBrinkhuis et al (1998) Speijer and van der Zwaan (1996) and Molina et al (2006) The fine-fraction stable carbon isotope record (blackdots) of El Kef is from Keller and Lindinger (1989) The Shannon diversity index (red dots) relative abundances of bi-triserial benthic taxa(dark purple) and all endobenthic foraminifera (light purple) are based on the gt 125 microm size fraction foraminiferal data of Speijer and vander Zwaan (1996) The palynological data of El Kef are from Brinkhuis et al (1998) (lower part) and Guasti et al (2005) (upper part) TheShannon diversity index of the gt 125 microm size fraction benthic foraminiferal record (red dots) the relative contribution of bi-triserial benthictaxa (dark purple) all endobenthic foraminifera (light purple) and hexaperidinioid dinocysts from Okccedilular are from this study The bulkstable carbon isotope record of Okccedilular (black dots) is from Accedilikalin et al (2015) Mbsf indicates meters below sea floor
cated by the higher abundance of hexaperidinioids Similarlyblooms of eutrophic survivor taxa of calcareous nanoplank-ton at open ocean sites have been suggested to be relatedto the buildup of nutrients in the open ocean photic zone(Schueth et al 2015) Although the carbon isotope gradientsbetween surface and deep waters indicate that the recoveryof the biological pump took hundreds of thousands of years(Zachos et al 1989 Kump 1991 DrsquoHondt et al 1998 Cox-all et al 2006 Birch et al 2016) our records suggest thatthe enhanced recycling of nutrients in the Tethys was partic-ularly intense during two phases in the first tens of thousandsyears after the impact (Fig 8)
523 Global responses
The decrease in export productivity following the KndashPgboundary is not only recognized in the Tethyan realm In theAtlantic Ocean Southern Ocean and the Indian Ocean a de-crease of the biological pump strength after the KndashPg bound-ary was also observed in neritic to abyssal environments(eg Thomas 1990 Olsson et al 1996 Hull et al 2011Alegret et al 2012) As an example the gt 63 microm size frac-tion benthic foraminiferal KndashPg boundary record of BlakeNose (ODP Hole 1049C northwestern Atlantic Alegret andThomas 2004) shows a pattern that is strikingly similar tothe gt 125 microm size fraction records in the Tethyan realm witha strong decrease of endobenthics in Zone P0 followed by a
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
896 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
gradual recovery across zones Pα and P1a (Fig 8) The simi-larities between the benthic foraminiferal patterns of Tethyanouter neritic to middle bathyal (200ndash1500 m) sites and thoseof the middle to lower bathyal (1500ndash1600 m) record ofBlake Nose (Alegret and Thomas 2004) suggest that theserecords register a global decrease in export productivity fol-lowing the KndashPg boundary
However several open ocean sites mostly in the Pacificshow an opposite trend with several lines of evidence sug-gesting an increased biological pump strength at these sites(eg Hull et al 2011 Alegret et al 2012) This led Ale-gret et al (2012) to conclude that the decrease in biologicalpump strength after the KndashPg boundary was most likely aregional instead of a global effect arguing against the Liv-ing Ocean hypothesis We would argue that low-productivityopen ocean sites like the central Pacific are characterized byvery different ecosystem structures and higher productivityas those along the Tethyan margins (eg Dortch and Pack-erd 1989) Therefore the consequence of ecosystem reorga-nization resulting from the mass extinction was likely verydifferent in the open ocean Pacific (eg Sibert et al 2014)Esmeray-Senlet et al (2015) proposed the term ldquoHeteroge-neous Oceanrdquo for this conceptual model as an alternativeto the Living Ocean model Supposedly the HeterogeneousOcean was characterized by a strong geographic heterogene-ity in the extinction patterns and food supply to the sea floor
However as the primary food source for open ocean deep-sea benthic communities generally consists of refractory or-ganic matter (eg Jiao et al 2010) it remains uncertainwhether changes in the intensity of recycling of the gener-ally more labile organic matter in the surface ocean will re-sult in changes in the amount of organic matter that benthiccommunities receive as they will likely continue to receiverecalcitrant organic matter at barely diminished rates despitethe collapse of the biological pump
In addition it should be noted that although the bio-logical pump efficiency is generally relatively high in openocean realms such as the central Pacific the biological pumpstrength is generally very low at these sites (Honjo et al2008 Henson et al 2011) Small changes in absolute bio-logical pump strength at such a locality could therefore havehad a large effect on the local oligotrophic benthic commu-nity (Alegret and Thomas 2009) but they likely representedonly a small fraction of the net amount of organic matterglobally transported to the deep Hence even though localbiological pump strength might have increased at some low-productivity deep-sea sites the large decrease in biologi-cal pump strength recorded at many other sites means thatthe net amount of organic matter globally transported out ofthe surface ocean was likely still reduced in the post-impactworld (Birch et al 2016) Therefore we argue that such Pa-cific records do not represent the general pattern The globalocean response can be characterized as heterogeneous how-ever the KndashPg boundary mass extinction did lead to a strongreduction of the net amount of organic matter globally trans-
ported from the surface to the sea floor while biological pro-ductivity recovered rapidly after the extinction event in ac-cordance with the Living Ocean hypothesis
53 Long-term recovery
Following the disaster phase the hexaperidinioid cysts atOkccedilular and El Kef decreased in abundance and the abun-dance of endobenthic forms increased again as the recoveryof the benthic community was initiated This recovery phaseis approximately correlative to Zone Pα and the basal part ofZone P1a which according to the Paleogene age constraintsof Vandenberghe et al (2012) represents at least 300 kyrThis duration is in agreement with the estimations providedby Birch et al (2016) During this phase the carbon iso-tope records remain well below pre-impact values and thediversity of the benthic foraminiferal community has not yetfully recovered (Figs 4ndash8) This indicates that the rapid andshort-lived KndashPg boundary disaster was followed by a rela-tively long recovery phase in line with previous estimatesof a multimillion-year biological recovery (eg Coxall etal 2006) While the impact-related environmental perturba-tions were short lived (Kring 2007 Vellekoop et al 20142016) the extinctions amongst important biological groupsled to a reduction of the organic flux from the photic zone todeep water resulting in major long-term biological and pa-leoceanographic reorganizations Only with the evolutionaryrecovery of the pelagic community governing the biologicalcarbon pump did export productivity start to increase again(eg Coxall et al 2006 Birch et al 2016)
6 Conclusions
The marine palynological benthic foraminiferal and bulkstable isotope records of the Okccedilular and El Kef sections re-veal major changes across the KndashPg boundary interval por-traying the biological crisis at the KndashPg boundary and subse-quent recovery in the early Paleocene Based on the succes-sion of benthic faunal assemblages at a number of Tethyanmargin sites four phases can be recognized across the KndashPgboundary interval the Maastrichtian or pre-impact phase (I)a disaster phase (II) a recovery phase (III) and an early Pa-leocene post-recovery phase (IV)
Following the KndashPg boundary impact some localitiesshow a large short-lived flux of food to the sea floor likelyrelated to the mass mortality at the KndashPg boundary Howeversince export productivity was greatly reduced after the KndashPgboundary mass extinction it is likely that after the initialpost-impact influx very little fresh organic matter reachedthe sea floor This reduction of export productivity in thepost-extinction disaster phase eventually resulted in a lowerfood supply to the sea floor This presented a major stressfactor for benthic organisms in the western Tethys The re-duced food supply resulted in an abrupt impoverishment of
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
J Vellekoop et al Ecological response to KndashPg boundary mass extinction 897
benthic communities As the downward transport of nutri-ents was slowed down recycling in upper layers increasedAs a result more nutrients remained available for the earlyPaleocene phytoplankton community leading to blooms ofdinoflagellates along the Tethyan margins and blooms of cal-careous nanoplankton taxa in more open ocean sites Ourrecords show that the enhanced recycling of nutrients in theTethys was particularly intense in the tens of thousands ofyears after the impact Following this the slow evolution-ary recovery of the pelagic community governing the bio-logical carbon pump resulted in a gradually increasing ex-port productivity in the hundreds of thousands of years afterthe impact Hence our integration of dinocyst and benthicforaminiferal records across the KndashPg boundary provides im-portant new insights into the ecological responses to the re-duction of export productivity following the mass extinctionat the CretaceousndashPaleogene boundary highlighting the di-rect ecological consequences of the Living Ocean conditionsin the post-impact world
7 Data availability
All data related to publication are available in the Supple-ment
The Supplement related to this article is available onlineat doi105194bg-14-885-2017-supplement
Author contributions Johan Vellekoop Lineke Woelders andHenk Brinkhuis designed the research Samples were collectedin the field by Johan Vellekoop Sanem Accedilikalin Ismail Ouml Yil-maz and Jan Smit Palynological analyses were carried out by Jo-han Vellekoop Benthic foraminiferal analyses were carried out byLineke Woelders and Robert P Speijer Johan Vellekoop LinekeWoelders and Bas van de Schootbrugge prepared the manuscriptwith input from all authors
Competing interests The authors declare that they have no conflictof interest
Acknowledgements This work was supported by the NetherlandsOrganization for Scientific Research (NWO) Open CompetitionGrant ALWPJ09047 to H Brinkhuis and the Research FoundationFlanders (FWO) grant G0B8513 to R P Speijer We thank LKump F Boscolo Galazzo P Roopnarine and one anonymousreferee for their suggestions
Edited by D GillikinReviewed by L Kump F Boscolo Galazzo and P Roopnarine
References
Accedilikalin S Vellekoop J Ocakoglu F Yılmaz I Ouml Smit J Al-tiner S O Goderis S Vonhof H Speijer R P WoeldersL Fornaciari E and Brinkhuis H Geochemical and pale-ontological characterization of a new KndashPg Boundary local-ity from the Northern branch of the Neo-Tethys MudurnundashGoumlynuumlk Basin NW Turkey Cretaceous Res 52 251ndash267doi101016jcretres201407011 2015
Alegret L and Thomas E Food supply to the seafloor in the Pa-cific Ocean after the CretaceousPaleogene boundary event MarMicropaleontol 73 105ndash116 2009
Alegret L Molina E and Thomas E Benthic foraminiferalturnover across the CretaceousPaleogene boundary at Agost(southeastern Spain) paleoenvironmental inferences Mar Mi-cropaleontol 48 251ndash279 2003
Alegret L Thomas E and Lohmann K C End-Cretaceous ma-rine mass extinction not caused by productivity collapse P NatlAcad Sci USA 109 728ndash732 doi101073pnas11106011092012
Alegret L Rodriacuteguez-Tovar F J and Uchman A How bioturba-tion obscured the CretaceousndashPaleogene boundary record TerraNova 27 225ndash230 doi101111ter12151 2015
Altiner D Koccedilyigit A Farinacci A Nicosia U and ContiM A Jurassic-Lower Cretaceous stratigraphy and paleogeo-graphic evolution of the southern part of north-western Anatolia(Turkey) Geologica Romana 27 13ndash80 1991
Alvarez LW Alvarez W Asaro F and Michel H V Extrater-restrial cause for the Cretaceous-Tertiary extinction Science208 1095ndash1108 1980
Alegret L and Thomas E Benthic foraminifera and environmen-tal turnover across the CretaceousPaleogene boundary at BlakeNose (ODP Hole 1049C Northwestern Atlantic) PalaeogeogrPalaeocl 208 59ndash83 doi101016jpalaeo200402028 2004
Armstrong R A Lee C Hedges J I Honjo S and WakehamS G A new mechanistic model for organic carbon fluxes in theocean based on the quantitative association of POC with ballastmaterials Deep-Sea Res Pt II 49 219ndash236 2002
Aubert J and Berggren W A Paleocene benthic foraminiferalbiostratigraphy and paleoecology of Tunisia Bull du Cent deRech de Pau ndash SNPA 10 379ndash469 1976
Berggren W A and Aubert J Paleocene benthonic foraminiferalbiostratigraphy paleobiogeography and paleoecology ofAtlanticndashTethyan regions midway-type fauna PalaeogeogrPalaeocl 18 73ndash192 1975
Bernaola G and Monechi S Calcareous nannofossil ex-tinction and survivorship across the CretaceousndashPaleogeneboundary at Walvis Ridge (ODP Hole 1262C SouthAtlantic Ocean) Palaeogeogr Palaeocl 255 132ndash156doi101016jpalaeo200702045 2007
Bernhard J M Characteristic assemblages and morphologies ofbenthic foraminifera from anoxic organic-rich deposits Jurassicthrough Holocene J Foramin Res 16 207ndash215 1986
Birch H S Coxall H K Pearson P N Kroon D andSchmidt D N Partial collapse of the marine carbon pump af-ter the CretaceousndashPaleogene boundary Geology 44 287ndash290doi101130G375811 2016
Brinkhuis H and Zachariasse W J Dinoflagellate cysts sealevel changes and planktonic foraminifers across the Cretaceous-
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
898 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
Tertiary boundary at El Haria northwest Tunisia Mar Micropa-leontol 13 153ndash191 1988
Brinkhuis H Bujak J P Smit J Versteegh G J M and Viss-cher H Dinoflagellate-based sea surface temperature recon-structions across the Cretaceous-Tertiary boundary PalaeogeogrPalaeocl 141 67ndash83 1998
Buesseler K O The decoupling of production and particulate ex-port in the surface ocean Global Biogeochem Cy 12 297ndash3101998
Buzas M A Culver S J and Jorissen F J A statisti-cal evaluation of the microhabitats of living (stained) infau-nal benthic foraminifera Mar Micropaleontol 20 311ndash320doi1010160377-8398(93)90040-5 1993
Coccioni R and Galeotti S KndashT boundary extinction geolog-ically instantaneous or gradual event Evidence from deep-seabenthic foraminifera Geology 22 779ndash782 1994
Corliss B H Microhabitats of benthic foraminifera within deep-sea sediments Nature 314 435ndash438 doi101038314435a01985
Corliss B H and Chen C Morphotype patterns of Nor-wegian Sea deep-sea benthic foraminifera and ecologi-cal implications Geology 16 716ndash719 doi1011300091-7613(1988)016lt0716MPONSDgt23CO2 1988
Coxall H K DrsquoHondt S and Zachos J C Pelagic evolution andenvironmental recovery after the CretaceousndashPaleogene mass ex-tinction Geology 34 297ndash300 doi101130G217021 2006
Culver S J Benthic foraminifera across the CretaceousndashTertiary(KndashT) boundary a review Mar Micropaleontol 47 177ndash2262003
Cushman J A Upper Cretaceous foraminifera of the Gulf coastalregion of the United States and adjacent areas US Geolog SurvProf Pap 206 241 pp 1946
Cushman J A Paleocene foraminifera of the Gulf Coastal Regionof the United States and adjacent regions US Geol Surv ProfPap 232 75 pp 1951
DrsquoHondt S Consequences of the CretaceousPaleogene Mass Ex-tinction for Marine Ecosystems Annu Rev Ecol Evol S36 295ndash317 doi101146annurevecolsys350211031057152005
DrsquoHondt S and Zachos J C Cretaceous foraminifera and the evo-lutionary history of planktic photosymbiosis Paleobiology 24512ndash523 1998
DrsquoHondt S Donaghay P Zachos J C Luttenberg D andLindinger M Organic Carbon Fluxes and Ecological Recov-ery from the Cretaceous-Tertiary Mass Extinction Science 282276ndash279 1998
Donze P Colin J-P Damotte R Oertli HJ Peypouquet J-Pand Said R Les ostracodes du Campanien terminal agrave lrsquoEocegraveneinfeacuterieur de la coupe du Kef Tuneacutesie nord-occidentale Bull desCentr Recher Expl-Prod Elf Aquit 6 273ndash335 1982
Dortch Q and Packard T T Differences in biomass structure be-tween oligotrophic and eutrophic marine ecosystems Deep-SeaRes 36 223ndash240 1989
Ernst S R Guasti E Dupuis C and Speijer R P En-vironmental perturbation in the southern Tethys across thePaleoceneEocene boundary (Dababiya Egypt) Foraminiferaland clay mineral records Mar Micropaleontol 60 89ndash111doi101016jmarmicro200603002 2006
Eshet Y Moshkovitz S Habib D Benjamini C and Maga-ritz M Calcareous nannofossil and dinoflagellate stratigraphyacross the CretaceousTertiary boundary at Hor Hahar IsraelMar Micropaleontol 18 199ndash228 1992
Eshet Y Almogi-Labin A and Bein A Dinoflagellate cysts pa-leoproductivity and upwelling systems A Late Cretaceous ex-ample from Israel Mar Micropaleontol 23 231ndash240 1994
Esmeray-Senlet S Wright J D Olsson R K MillerK G Browning J V and Quan T M Evidencefor reduced export productivity following the Creta-ceousPaleogene mass extinction Paleoceanography 30718ndash738 doi1010022014PA002724 2015
Giusberti L Boscolo Galazzo F and Thomas E Variability inclimate and productivity during the Paleocene-Eocene ThermalMaximum in the western Tethys (Forada section) Clim Past12 213ndash240 doi105194cp-12-213-2016 2016
Guasti E Kouwenhoven T J Brinkhuis H and Speijer R PPaleocene sea-level and productivity changes at the southernTethyan margin (El Kef Tunisia) Mar Micropaleontol 55 1ndash17 2005
Habib D and Saeedi F The Manumiella seelandica global spikeCooling during regression at the close of the MaastrichtianPalaeogeogr Palaeocl 255 87ndash97 2007
Hain M P Sigmal D M and Haug G H 818 ndash The biolog-ical Pump in the Past in Reference Module in Earth Systemsand Environmental Sciences Treatise on Geochemistry (SecondEdition) The Oceans and Marine Geochemistry 8 485ndash517doi101016B978-0-08-095975-700618-5 2014
Hayek L C and Buzas M A On the proper and efficient useof diversity measures with individual field samples J ForaminRes 43 305ndash313 2013
Henson S A Sanders R Madsen E Morris P J Le MoigneF and Quartly G D A reduced estimate of the strength ofthe oceanrsquos biological carbon pump Geophys Res Lett 38L04606 doi1010292011GL046735 2011
Hilting A K Kump L R and Bralower T J Variations in theoceanic vertical carbon isotope gradient and their implicationsfor the Paleocene-Eocene biological pump Paleoceanography23 PA3222 doi1010292007PA001458 2008
Honjo S Manganini S J Krishfield R A and Francois RParticulate organic carbon fluxes to the ocean interior and fac-tors controlling the biological pump A synthesis of global sed-iment trap progams since 1983 Prog Oceanogr 76 217ndash285doi101016jpocean200711003 2008
Huber B T MacLeod K G and Norris R D Abrupt extinctionand subsequent reworking of Cretaceous planktonic foraminiferaacross the Cretaceous-Tertiary boundary Evidence from the sub-tropical North Atlantic in Catastrophic Events and Mass Extinc-tions Impacts and Beyond edited by Koeberl C and MacLeodK G Geol S Am S 356 277ndash289 Boulder Colorado USA2002
Hull P M Norris R D Bralower T J and Schueth J D A rolefor chance in marine recovery from the end-Cretaceous extinc-tion Nat Geosci 4 856ndash860 doi101038ngeo1302 2011
Hsuuml K J and McKenzie J A A ldquoStrangeloverdquo ocean in the earli-est Tertiary in Natural variations Archean to present edited bySundquist E T and Broecker W S The carbon cycle and atmo-spheric CO2 American Geophysical Union Washington DCUSA 487ndash492 1985
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
J Vellekoop et al Ecological response to KndashPg boundary mass extinction 899
Jiao N Herndl G J Hansell D A Benner R Kattner G Wil-helm S W Kirchman D L Weinbauer M G Luo T ChenF and Azam F Microbial production of recalcitrant dissolvedorganic matter long-term carbon storage in the global oceanNat Rev 8 593ndash599 doi101038nrmicro2386 2010
Jorissen F J de Stigter H C and Widmark J G V A concep-tual model explaining benthic foraminiferal microhabitats MarMicropaleontol 26 3ndash15 doi1010160377-8398(95)00047-X1995
Jorissen F J Fontanier C and Thomas E Paleoceanographi-cal proxies based on deep-sea benthic foraminiferal assemblagecharacteristics in Proxies in Late Cenozoic PaleoceanographyPt 2 Biological tracers and biomarkers edited by Hillaire-Marcel C and de Vernal A 263ndash326 Elsevier Amsterdamthe Netherlands 2007
Kaiho K Kajiwara Y Tazaki K Ueshima M Takeda NKawahata H Arinobu T Ishiwatari R Hirai A and Lam-olda M A Oceanic primary productivity and dissolved oxygenlevels at the CretaceousTertiary boundary their decrease sub-sequent warming and recovery Paleoceanography 14 511ndash5241999
Keller G and Lindinger M Stable isotope TOC and CaCO3record across the CretaceousTertiary boundary at El KefTunisia Palaeogeogr Palaeocl 73 243ndash265 1989
Kellough G R Paleoecology of the Foraminiferida of the WillsPoint Formation (Midway Group) in northeast Texas Trans GulfCoast Assoc Geol Soc 15 73ndash153 1965
Kring D A The Chicxulub impact event and its environmentalconsequences at the CretaceousndashTertiary boundary PalaeogeogrPalaeocl 255 4ndash21 2007
Kump L R Interpreting carbon-isotope excursions Strangeloveoceans Geology 19 299ndash302 1991
Machalski M Vellekoop J Dubicka Z Peryt D and Ha-rasimiuk M Late Maastrichtian cephalopods dinoflagellatecysts and foraminifera from the CretaceousndashPaleogene suc-cession at Lechoacutewka southeast Poland Stratigraphic andenvironmental implications Cretaceous Res 57 208ndash227doi101016jcretres201508012 2016
Molina E Evidence and causes of the main extinctionevents in the Paleogene based on extinction and survivalpatterns of foraminifera Earth-Sci Rev 140 166ndash181doi101016jearscirev201411008 2015
Molina E Alegret L Arenillas I and Arz J The GlobalBoundary Stratotype Section and Point for the base of the Da-nian Stage (Paleocene Paleogeneldquo Tertiaryrdquo Cenozoic) at ElKef Tunisia-Original definition and revision Episodes 29 263ndash273 2006
Olsson R K Liu C and van Fossen M The Cretaceous-Tertiarycatastrophic event at Millers Ferry Alabama in The Cretaceous-Tertiary Event and Other Catastrophes in Earth History editedby Ryder G Fastovsky D and Gartner S Geol S Am S307 263ndash277 1996
Peryt D Benthic foraminiferal response to the Cenomanian-Turonian and CretaceousndashPaleogene boundary events PrzegladGeol 52 827ndash832 2004
Peryt D Alegret L and Molina E The CretaceousPalaeogene(KP) boundary at Aiumln Settara Tunisia restructuring of benthicforaminiferal assemblages Terra Nova 14 101ndash107 2002
Ramette A Multivariate analyses in microbial ecology FEMS Mi-crobiol Ecol 62 142ndash160 2007
Rodriacuteguez-Tovar F J Uchman A MrsquoHamdi A Riahi Sand Ismail-Lattrache K B Ichnological record of palaeoen-vironment from the CretaceousndashPaleogene boundary intervalat El Kef Tunisia The first study of old and new sec-tions at the stratotype area J Afr Earth Sci 120 23ndash30doi101016jjafrearsci201604017 2016
Saner S Mudurnu-Goumlynuumlk Havzasının Jura ve Sonrasıccediloumlkelimnitelikleriyle paleocografya yorumlaması TJK Buumllteni 23 39ndash52 1980
Schioslashler P Brinkhuis H Roncaglia L and Wilson G J Di-noflagellate biostratigraphy and sequence stratigraphy of theType Maastrichtian (Upper Cretaceous) ENCI Quarry TheNetherlands Mar Micropaleontol 31 65ndash95 1997
Schueth J D Bralower T J Jiang S and Patzkowsky ME The role of regional survivor incumbency in the evolu-tionary recovery of calcareous nannoplankton from the Creta-ceousPaleogene (KPg) mass extinction Paleobiology 41 661ndash679 doi101017pab201528 2015
Scotese C R A continental drift flipbook J Geology 112 729ndash741 2004
Scotese C R and Dreher C GlobalGeology available at httpwwwGlobalGeologycom (last access 11 March 2014) 2012
Sepkoski J J Patterns of Phanerozoic extinction a perspectivefrom global data bases in Global Events and Event Stratigra-phy edited by Walliser O H Springer Berlin Germany 35ndash52 2016
Sibert E C Hull P M and Norris R D Resilience of Pacificpelagic fish across the CretaceousPalaeogene mass extinctionNat Geosci 7 667ndash670 doi101038NGEO2227 2014
Slimani H and Toufiq A A CretaceousndashPaleogene boundary geo-logical site revealed by planktic foraminifera and dinoflagellatecysts at Ouled Haddou eastern external Rif Chain Morocco JAfr Earth Sci 88 38ndash52 doi101016jjafrearsci2013080082013
Slimani H Louwye S and Toufiq A Dinoflagellate cystsform the CretaceousndashPaleogene boundary at Ouled Had-dou southeastern Rif Morocco biostratigraphy paleoenvi-ronments and paleobiogeography Palynology 34 90ndash124doi10108001916121003629933 2010
Sluijs A and Brinkhuis H A dynamic climate and ecosystemstate during the Paleocene-Eocene Thermal Maximum infer-ences from dinoflagellate cyst assemblages on the New JerseyShelf Biogeosciences 6 1755ndash1781 doi105194bg-6-1755-2009 2009
Smit J Extinction and evolution of planktonic foraminifera after amajor impact at the CretaceousTertiary boundary Geol S AmS 190 329ndash352 1982
Smit J and Romein A J T A sequence of events across theCretaceous-Tertiary boundary Earth Planet Sci Lett 74 155ndash170 1985
Speijer R P Extinction and recovery patterns in ben-thic foraminiferal paleocommunities across the Creta-ceousPaleogene and PaleoceneEocene boundaries GeologicaUltraiectina 124 Faculteit Aardwetenschappen PhD ThesisUniversiteit Utrecht Utrecht the Netherlands 1994
Speijer R P and van Der Zwaan G J Extinction and survivor-ship of southern Tethyan benthic foraminifera across the Cre-
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
900 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
taceousPaleogene boundary in Biotic recovery from mass ex-tinction events edited by Hart M B Geolog Soc Spec PublLondon UK 102 343ndash371 1996
Stemmann L and Boss E Plankton and Particle Size andPackaging From Determining Optical Properties to Drivingthe Biological Pump Annu Rev Mar Sci 4 263ndash290doi101146annurev-marine-120710-100853 2012
Thomas E Late Cretaceous-early Eocene mass extinctions in thedeep sea Geol S Am S 247 481ndash495 1990
Vandenberghe N Hilgen F J and Speijer R P The PaleogenePeriod in The Geologic Time Scale 2012 edited by GradsteinF M Ogg J G Schmitz M D and Ogg G M 2 855ndash921Elsevier Boston USA 2012
Vellekoop J Sluijs A Smit J Schouten S Weijers J WH Sinninghe Damsteacute J S and Brinkhuis H Rapid short-term cooling following the Chicxulub impact at the CretaceousndashPaleogene boundary P Natl Acad Sci USA 111 7537ndash7541doi101073pnas1319253111 2014
Vellekoop J Smit J van de Schootbrugge B Weijers J W HGaleotti S Sinninghe Damsteacute J S and Brinkhuis H Palyno-logical evidence for prolonged cooling along the Tunisian conti-nental shelf following the KndashPg boundary impact PalaeogeogrPalaeocl 426 216ndash228 doi101016jpalaeo2015030212015
Vellekoop J Esmeray-Senlet S Miller K G Browning J VSluijs A van de Schootbrugge B Sinninghe Damsteacute J Sand Brinkhuis H Evidence for CretaceousndashPaleogene bound-ary bolide ldquoimpact winterrdquo conditions from New Jersey USAGeology 44 619ndash622 doi101130G379611 2016
Woelders L and Speijer R Stable seafloor conditions sealevel and food supply during the latest Maastrichtian atBrazos River Texas Mar Micropaleontol 121 41ndash51doi101016jmarmicro201510002 2015
Zachos J C Arthur M A and Dean W E Geochemical evi-dence for suppression of pelagic marine productivity at the Cre-taceousTertiary boundary Nature 337 61ndash64 1989
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
- Abstract
- Introduction
- Geological setting and age assessment
- Materials and methods
-
- Sampling
- Foraminiferal analysis
- Palynological analysis
- Statistical analysis
-
- Results
-
- Benthic foraminifera
- Palynology
-
- Discussion
-
- Tethyan benthic foraminiferal turnover sequence
- Ecological responses to reduced export productivity
-
- Okccedilular section
- Western Tethys
- Global responses
-
- Long-term recovery
-
- Conclusions
- Data availability
- Author contributions
- Competing interests
- Acknowledgements
- References
-
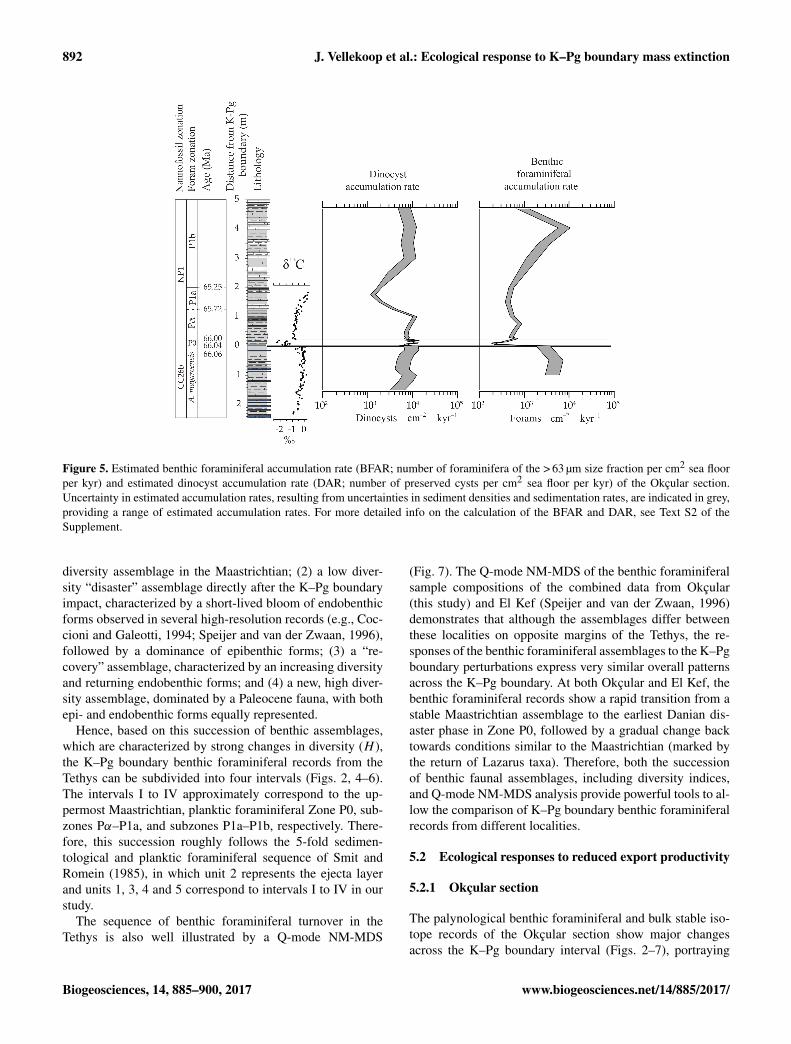
892 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
Figure 5 Estimated benthic foraminiferal accumulation rate (BFAR number of foraminifera of the gt 63 microm size fraction per cm2 sea floorper kyr) and estimated dinocyst accumulation rate (DAR number of preserved cysts per cm2 sea floor per kyr) of the Okccedilular sectionUncertainty in estimated accumulation rates resulting from uncertainties in sediment densities and sedimentation rates are indicated in greyproviding a range of estimated accumulation rates For more detailed info on the calculation of the BFAR and DAR see Text S2 of theSupplement
diversity assemblage in the Maastrichtian (2) a low diver-sity ldquodisasterrdquo assemblage directly after the KndashPg boundaryimpact characterized by a short-lived bloom of endobenthicforms observed in several high-resolution records (eg Coc-cioni and Galeotti 1994 Speijer and van der Zwaan 1996)followed by a dominance of epibenthic forms (3) a ldquore-coveryrdquo assemblage characterized by an increasing diversityand returning endobenthic forms and (4) a new high diver-sity assemblage dominated by a Paleocene fauna with bothepi- and endobenthic forms equally represented
Hence based on this succession of benthic assemblageswhich are characterized by strong changes in diversity (H )the KndashPg boundary benthic foraminiferal records from theTethys can be subdivided into four intervals (Figs 2 4ndash6)The intervals I to IV approximately correspond to the up-permost Maastrichtian planktic foraminiferal Zone P0 sub-zones PαndashP1a and subzones P1andashP1b respectively There-fore this succession roughly follows the 5-fold sedimen-tological and planktic foraminiferal sequence of Smit andRomein (1985) in which unit 2 represents the ejecta layerand units 1 3 4 and 5 correspond to intervals I to IV in ourstudy
The sequence of benthic foraminiferal turnover in theTethys is also well illustrated by a Q-mode NM-MDS
(Fig 7) The Q-mode NM-MDS of the benthic foraminiferalsample compositions of the combined data from Okccedilular(this study) and El Kef (Speijer and van der Zwaan 1996)demonstrates that although the assemblages differ betweenthese localities on opposite margins of the Tethys the re-sponses of the benthic foraminiferal assemblages to the KndashPgboundary perturbations express very similar overall patternsacross the KndashPg boundary At both Okccedilular and El Kef thebenthic foraminiferal records show a rapid transition from astable Maastrichtian assemblage to the earliest Danian dis-aster phase in Zone P0 followed by a gradual change backtowards conditions similar to the Maastrichtian (marked bythe return of Lazarus taxa) Therefore both the successionof benthic faunal assemblages including diversity indicesand Q-mode NM-MDS analysis provide powerful tools to al-low the comparison of KndashPg boundary benthic foraminiferalrecords from different localities
52 Ecological responses to reduced export productivity
521 Okccedilular section
The palynological benthic foraminiferal and bulk stable iso-tope records of the Okccedilular section show major changesacross the KndashPg boundary interval (Figs 2ndash7) portraying
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
J Vellekoop et al Ecological response to KndashPg boundary mass extinction 893
Figure 6 Relative abundances of the four main clusters of benthic foraminiferal taxa (gt 125 microm) relative abundances of epibenthic andendobenthic forms and relative abundance of bi-triserial benthic taxa
Figure 7 Q-mode non-metric multidimensional scaling (on all sam-ples in the gt 125 microm size fraction benthic foraminiferal datasets ofOkccedilular (this study) and El Kef (Speijer and van der Zwaan 1996)After an abrupt transition from latest Maastrichtian pre-impact as-semblages to disaster assemblages in Zone P0 and recovery assem-blages in Zone Pα a gradual transition towards a new equilibriumpost-recovery assemblage can be observed in subzones P1a and P1b
the biological crisis following the impact Following the KndashPg boundary mass extinction endobenthic forms stronglydecrease with bi-triserial benthic taxa almost disappearingfrom the benthic community simultaneously the estimatedBFAR shows a major fall (Fig 5) This interval approxi-mately correlative to foraminiferal Zone P0 represents thedisaster phase The drop in bi-triserial benthic taxa (Fig 6)and decrease in BFAR (Fig 5) point to a reduction in foodsupply to the benthic community indicating that the trans-port of organic matter to the sea floor must have decreasedIn the dinocyst community however hexaperidinioids showa strong increase in relative abundance during this phase(Figs 3 6 7) suggesting an increase in nutrient availabilityin the photic zone while the DAR remains relatively stableacross the KndashPg boundary (Fig 5)
The decrease in benthic diversity and the high relativeabundances of opportunistic Trochammina spp and Cibici-doides pseudoacutus suggest that the benthic community ex-perienced additional stress during Zone P0 besides food lim-itation (see Jorissen et al 2007) If food limitation were theonly factor involved upper bathyal assemblages as observedat Okccedilular would be expected to become more similar tooligotrophic middle to lower bathyal assemblages Insteadthe opposite is true the assemblage composition of the dis-aster phase is closer to neritic Midway-type assemblages
It was previously suggested that the epibenthic taxon Cibi-cidoides pseudoacutus could potentially be hypoxia resistant
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
894 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
(Speijer and van der Zwaan 1996) As a collapse of the bi-ological pump should lead to enhanced remineralization oforganic matter in the photic zone it is indeed likely thatthe oxygen minimum zones expanded and shoaled leadingto oxygen stress on the sea floor Although early Paleocenedysoxia at El Kef is supported by the ostracod record (Donzeet al 1982) and ichnofacies analyses (Rodriguez-Tovaret al 2016) supporting (eg geochemical) evidence fordysoxia during the disaster phase at Okccedilular record is absentFurthermore an additional stressor for the benthic commu-nity during Zone P0 may be present In contrast to calcare-ous nanoplankton organic-walled cyst-producing dinoflagel-lates did not suffer extinctions and may have become a moreimportant component in the early Paleocene phytoplanktoncommunity Hence besides the reduction of the amount oforganic matter transported to the sea floor the compositionof food supplied by the photic zone likely changed signifi-cantly across the KndashPg boundary as well (DrsquoHondt 2005)possibly presenting an additional stress factor for the benthiccommunity (Speijer and van der Zwaan 1996 Alegret andThomas 2009)
522 Western Tethys
The patterns in benthic foraminiferal and dinoflagellate re-sponse to the KndashPg boundary perturbations at Okccedilular appearto be characteristic for the continental margins of the westernTethyan realm The benthic foraminiferal patterns are similaracross different size fractions suggesting that they are repre-sentative of the overall benthic communities In Tunisia andSpain where both benthic foraminiferal and dinoflagellaterecords are available the earliest Danian is also characterizedby a decrease in endobenthic foraminifera at the sea floor(Speijer and Van der Zwaan 1996 Peryt et al 2002 Ale-gret et al 2003) and simultaneously blooms of hexaperidin-ioids in the water column (Brinkhuis et al 1998 Vellekoopet al 2015 Fig 8) In the Ouled Haddou section Moroccoin the westernmost Tethys (Fig 1) the lowermost Danian isalso characterized by a strong increase of the hexaperidin-ioid Senegalinium group up to 30 of the assemblage (Sli-mani et al 2010) that is very similar to the Okccedilular record(Fig 8)
The KndashPg boundary dinocyst record of El Kef is strikinglysimilar to the one from the Okccedilular section showing two dis-tinct peaks in hexaperidinioids one in Zone P0 and one atthe basal part of Zone Pα At El Kef the initial post-impactdominance of hexaperidinioids is however less pronounced(Fig 8) This difference in expression might be related tosmall differences in paleogeographic and paleoceanographicsettings between sites
In addition some high-resolution benthic KndashPg records inthe Tethyan realm show a short-lived bloom of specific en-dobenthic foraminiferal taxa in the few centimeter directlyabove the KndashPg boundary sometimes accompanied by otherindicators of low oxygen levels (eg Coccioni and Gale-
otti 1994 Speijer and Van der Zwaan 1996 Kaiho et al1999) This suggests that at least locally the sea floor wastemporarily characterized by hypoxic conditions directly fol-lowing the impact This short-lived deoxygenation is possi-bly related to the mass mortality at the KndashPg boundary re-sulting in a large short-lived flux of organic matter to thesea floor (Coccioni and Galeotti 1994) However since ex-port productivity was greatly reduced after the KndashPg bound-ary mass extinction very little ldquonewrdquo organic matter reachedthe sea floor after the initial post-impact influx Hence afteraccumulated organic matter was remineralized the benthiccommunity starved resulting in a transition to an epibenthic-dominated benthic fauna similar to the post-impact fauna ofthe Okccedilular record
The combined dinocyst records suggest that the early Pa-leocene margins of the Tethys were characterized by an in-crease in nutrient availability in the photic zone whereascoeval benthic foraminiferal records indicate a major de-crease in nutrient supply to the sea floor These changesin the benthic foraminiferal and dinocyst communities arelikely caused by the major reduction of both the efficiencyand strength of the biological pump (ie the fraction ofprimary production exported from the photic zone and theamount of organic matter transported down) in accordancewith the Living Ocean model (DrsquoHondt and Zachos 1998DrsquoHondt et al 1998 DrsquoHondt 2005 Coxall et al 2006)For a detailed explanation of the difference between biologi-cal pump efficiency and biological pump strength see Hiltinget al (2008)
As bulk carbonate δ13C reflects the isotopic compositionof the surface ocean which is set by burial fractions and bythe photosynthetic isotope effect (Kump 1991 Hain et al2014) the excursion recorded in bulk δ13C records world-wide and the rapid collapse in surface to deep-ocean carbonisotope gradients likely reflect the reduction of the global in-tensity (ie efficiency) of the biological pump (Hain et al2014) The recorded inverse change in nutrient availabilitysuggests a causal link A reduced efficiency of the biologi-cal pump and associated decrease of the fraction of biomasstransported out of the photic zone could have resulted inhigh rates of nutrient recycling in the upper part of the wa-ter column (DrsquoHondt 2005) This suggests that the reduc-tion of the biological pump strength recorded by the benthicforaminiferal record is a consequence of the decreased effi-ciency of the biological pump recorded by the carbon iso-tope and dinoflagellate cyst records The strong correlationbetween the bulk carbon isotope curves and the Shannondiversity index (H ) of the benthic foraminiferal records atboth Okccedilular (R2
= 073 p lt 0001) and El Kef (R2= 074
p lt 0001) suggest a clear link between the changes in ben-thic foraminiferal assemblages and the collapse and recoveryof biological pump efficiency
As a result of the reduced biological pump efficiency morenutrients remained available for the early Paleocene phyto-plankton community At Tethyan margin sites this is indi-
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
J Vellekoop et al Ecological response to KndashPg boundary mass extinction 895
Figure 8 The benthic foraminiferal (gt 125 microm) dinocyst and bulk stable carbon isotope records of Okccedilular (Turkey) and El Kef (Tunisia)the dinocyst record of Ouled Haddou (Morocco) and the benthic foraminiferal record (gt 63 microm) of ODP Hole 1049C Blake Nose (NorthAtlantic) Note that the definition of ldquohexaperidinioidsrdquo used is provided in the text The four phases identified based on the bulk stablecarbon isotopes and the Shannon diversity index (H ) of the benthic foraminiferal records are indicated (IndashIV) The gt 63 microm size fractionbenthic foraminiferal record of ODP Hole 1049C is from Alegret and Thomas (2004) who argued that the lowermost Paleocene of thissite comprised reworked foraminifera This interval is indicated by a dashed bar in the figure The biostratigraphy of Ouled Haddou is fromSlimani and Toufiq (2013) whereas the dinocyst record of this site is from Slimani et al (2010) The biostratigraphy of El Kef is based onBrinkhuis et al (1998) Speijer and van der Zwaan (1996) and Molina et al (2006) The fine-fraction stable carbon isotope record (blackdots) of El Kef is from Keller and Lindinger (1989) The Shannon diversity index (red dots) relative abundances of bi-triserial benthic taxa(dark purple) and all endobenthic foraminifera (light purple) are based on the gt 125 microm size fraction foraminiferal data of Speijer and vander Zwaan (1996) The palynological data of El Kef are from Brinkhuis et al (1998) (lower part) and Guasti et al (2005) (upper part) TheShannon diversity index of the gt 125 microm size fraction benthic foraminiferal record (red dots) the relative contribution of bi-triserial benthictaxa (dark purple) all endobenthic foraminifera (light purple) and hexaperidinioid dinocysts from Okccedilular are from this study The bulkstable carbon isotope record of Okccedilular (black dots) is from Accedilikalin et al (2015) Mbsf indicates meters below sea floor
cated by the higher abundance of hexaperidinioids Similarlyblooms of eutrophic survivor taxa of calcareous nanoplank-ton at open ocean sites have been suggested to be relatedto the buildup of nutrients in the open ocean photic zone(Schueth et al 2015) Although the carbon isotope gradientsbetween surface and deep waters indicate that the recoveryof the biological pump took hundreds of thousands of years(Zachos et al 1989 Kump 1991 DrsquoHondt et al 1998 Cox-all et al 2006 Birch et al 2016) our records suggest thatthe enhanced recycling of nutrients in the Tethys was partic-ularly intense during two phases in the first tens of thousandsyears after the impact (Fig 8)
523 Global responses
The decrease in export productivity following the KndashPgboundary is not only recognized in the Tethyan realm In theAtlantic Ocean Southern Ocean and the Indian Ocean a de-crease of the biological pump strength after the KndashPg bound-ary was also observed in neritic to abyssal environments(eg Thomas 1990 Olsson et al 1996 Hull et al 2011Alegret et al 2012) As an example the gt 63 microm size frac-tion benthic foraminiferal KndashPg boundary record of BlakeNose (ODP Hole 1049C northwestern Atlantic Alegret andThomas 2004) shows a pattern that is strikingly similar tothe gt 125 microm size fraction records in the Tethyan realm witha strong decrease of endobenthics in Zone P0 followed by a
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
896 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
gradual recovery across zones Pα and P1a (Fig 8) The simi-larities between the benthic foraminiferal patterns of Tethyanouter neritic to middle bathyal (200ndash1500 m) sites and thoseof the middle to lower bathyal (1500ndash1600 m) record ofBlake Nose (Alegret and Thomas 2004) suggest that theserecords register a global decrease in export productivity fol-lowing the KndashPg boundary
However several open ocean sites mostly in the Pacificshow an opposite trend with several lines of evidence sug-gesting an increased biological pump strength at these sites(eg Hull et al 2011 Alegret et al 2012) This led Ale-gret et al (2012) to conclude that the decrease in biologicalpump strength after the KndashPg boundary was most likely aregional instead of a global effect arguing against the Liv-ing Ocean hypothesis We would argue that low-productivityopen ocean sites like the central Pacific are characterized byvery different ecosystem structures and higher productivityas those along the Tethyan margins (eg Dortch and Pack-erd 1989) Therefore the consequence of ecosystem reorga-nization resulting from the mass extinction was likely verydifferent in the open ocean Pacific (eg Sibert et al 2014)Esmeray-Senlet et al (2015) proposed the term ldquoHeteroge-neous Oceanrdquo for this conceptual model as an alternativeto the Living Ocean model Supposedly the HeterogeneousOcean was characterized by a strong geographic heterogene-ity in the extinction patterns and food supply to the sea floor
However as the primary food source for open ocean deep-sea benthic communities generally consists of refractory or-ganic matter (eg Jiao et al 2010) it remains uncertainwhether changes in the intensity of recycling of the gener-ally more labile organic matter in the surface ocean will re-sult in changes in the amount of organic matter that benthiccommunities receive as they will likely continue to receiverecalcitrant organic matter at barely diminished rates despitethe collapse of the biological pump
In addition it should be noted that although the bio-logical pump efficiency is generally relatively high in openocean realms such as the central Pacific the biological pumpstrength is generally very low at these sites (Honjo et al2008 Henson et al 2011) Small changes in absolute bio-logical pump strength at such a locality could therefore havehad a large effect on the local oligotrophic benthic commu-nity (Alegret and Thomas 2009) but they likely representedonly a small fraction of the net amount of organic matterglobally transported to the deep Hence even though localbiological pump strength might have increased at some low-productivity deep-sea sites the large decrease in biologi-cal pump strength recorded at many other sites means thatthe net amount of organic matter globally transported out ofthe surface ocean was likely still reduced in the post-impactworld (Birch et al 2016) Therefore we argue that such Pa-cific records do not represent the general pattern The globalocean response can be characterized as heterogeneous how-ever the KndashPg boundary mass extinction did lead to a strongreduction of the net amount of organic matter globally trans-
ported from the surface to the sea floor while biological pro-ductivity recovered rapidly after the extinction event in ac-cordance with the Living Ocean hypothesis
53 Long-term recovery
Following the disaster phase the hexaperidinioid cysts atOkccedilular and El Kef decreased in abundance and the abun-dance of endobenthic forms increased again as the recoveryof the benthic community was initiated This recovery phaseis approximately correlative to Zone Pα and the basal part ofZone P1a which according to the Paleogene age constraintsof Vandenberghe et al (2012) represents at least 300 kyrThis duration is in agreement with the estimations providedby Birch et al (2016) During this phase the carbon iso-tope records remain well below pre-impact values and thediversity of the benthic foraminiferal community has not yetfully recovered (Figs 4ndash8) This indicates that the rapid andshort-lived KndashPg boundary disaster was followed by a rela-tively long recovery phase in line with previous estimatesof a multimillion-year biological recovery (eg Coxall etal 2006) While the impact-related environmental perturba-tions were short lived (Kring 2007 Vellekoop et al 20142016) the extinctions amongst important biological groupsled to a reduction of the organic flux from the photic zone todeep water resulting in major long-term biological and pa-leoceanographic reorganizations Only with the evolutionaryrecovery of the pelagic community governing the biologicalcarbon pump did export productivity start to increase again(eg Coxall et al 2006 Birch et al 2016)
6 Conclusions
The marine palynological benthic foraminiferal and bulkstable isotope records of the Okccedilular and El Kef sections re-veal major changes across the KndashPg boundary interval por-traying the biological crisis at the KndashPg boundary and subse-quent recovery in the early Paleocene Based on the succes-sion of benthic faunal assemblages at a number of Tethyanmargin sites four phases can be recognized across the KndashPgboundary interval the Maastrichtian or pre-impact phase (I)a disaster phase (II) a recovery phase (III) and an early Pa-leocene post-recovery phase (IV)
Following the KndashPg boundary impact some localitiesshow a large short-lived flux of food to the sea floor likelyrelated to the mass mortality at the KndashPg boundary Howeversince export productivity was greatly reduced after the KndashPgboundary mass extinction it is likely that after the initialpost-impact influx very little fresh organic matter reachedthe sea floor This reduction of export productivity in thepost-extinction disaster phase eventually resulted in a lowerfood supply to the sea floor This presented a major stressfactor for benthic organisms in the western Tethys The re-duced food supply resulted in an abrupt impoverishment of
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
J Vellekoop et al Ecological response to KndashPg boundary mass extinction 897
benthic communities As the downward transport of nutri-ents was slowed down recycling in upper layers increasedAs a result more nutrients remained available for the earlyPaleocene phytoplankton community leading to blooms ofdinoflagellates along the Tethyan margins and blooms of cal-careous nanoplankton taxa in more open ocean sites Ourrecords show that the enhanced recycling of nutrients in theTethys was particularly intense in the tens of thousands ofyears after the impact Following this the slow evolution-ary recovery of the pelagic community governing the bio-logical carbon pump resulted in a gradually increasing ex-port productivity in the hundreds of thousands of years afterthe impact Hence our integration of dinocyst and benthicforaminiferal records across the KndashPg boundary provides im-portant new insights into the ecological responses to the re-duction of export productivity following the mass extinctionat the CretaceousndashPaleogene boundary highlighting the di-rect ecological consequences of the Living Ocean conditionsin the post-impact world
7 Data availability
All data related to publication are available in the Supple-ment
The Supplement related to this article is available onlineat doi105194bg-14-885-2017-supplement
Author contributions Johan Vellekoop Lineke Woelders andHenk Brinkhuis designed the research Samples were collectedin the field by Johan Vellekoop Sanem Accedilikalin Ismail Ouml Yil-maz and Jan Smit Palynological analyses were carried out by Jo-han Vellekoop Benthic foraminiferal analyses were carried out byLineke Woelders and Robert P Speijer Johan Vellekoop LinekeWoelders and Bas van de Schootbrugge prepared the manuscriptwith input from all authors
Competing interests The authors declare that they have no conflictof interest
Acknowledgements This work was supported by the NetherlandsOrganization for Scientific Research (NWO) Open CompetitionGrant ALWPJ09047 to H Brinkhuis and the Research FoundationFlanders (FWO) grant G0B8513 to R P Speijer We thank LKump F Boscolo Galazzo P Roopnarine and one anonymousreferee for their suggestions
Edited by D GillikinReviewed by L Kump F Boscolo Galazzo and P Roopnarine
References
Accedilikalin S Vellekoop J Ocakoglu F Yılmaz I Ouml Smit J Al-tiner S O Goderis S Vonhof H Speijer R P WoeldersL Fornaciari E and Brinkhuis H Geochemical and pale-ontological characterization of a new KndashPg Boundary local-ity from the Northern branch of the Neo-Tethys MudurnundashGoumlynuumlk Basin NW Turkey Cretaceous Res 52 251ndash267doi101016jcretres201407011 2015
Alegret L and Thomas E Food supply to the seafloor in the Pa-cific Ocean after the CretaceousPaleogene boundary event MarMicropaleontol 73 105ndash116 2009
Alegret L Molina E and Thomas E Benthic foraminiferalturnover across the CretaceousPaleogene boundary at Agost(southeastern Spain) paleoenvironmental inferences Mar Mi-cropaleontol 48 251ndash279 2003
Alegret L Thomas E and Lohmann K C End-Cretaceous ma-rine mass extinction not caused by productivity collapse P NatlAcad Sci USA 109 728ndash732 doi101073pnas11106011092012
Alegret L Rodriacuteguez-Tovar F J and Uchman A How bioturba-tion obscured the CretaceousndashPaleogene boundary record TerraNova 27 225ndash230 doi101111ter12151 2015
Altiner D Koccedilyigit A Farinacci A Nicosia U and ContiM A Jurassic-Lower Cretaceous stratigraphy and paleogeo-graphic evolution of the southern part of north-western Anatolia(Turkey) Geologica Romana 27 13ndash80 1991
Alvarez LW Alvarez W Asaro F and Michel H V Extrater-restrial cause for the Cretaceous-Tertiary extinction Science208 1095ndash1108 1980
Alegret L and Thomas E Benthic foraminifera and environmen-tal turnover across the CretaceousPaleogene boundary at BlakeNose (ODP Hole 1049C Northwestern Atlantic) PalaeogeogrPalaeocl 208 59ndash83 doi101016jpalaeo200402028 2004
Armstrong R A Lee C Hedges J I Honjo S and WakehamS G A new mechanistic model for organic carbon fluxes in theocean based on the quantitative association of POC with ballastmaterials Deep-Sea Res Pt II 49 219ndash236 2002
Aubert J and Berggren W A Paleocene benthic foraminiferalbiostratigraphy and paleoecology of Tunisia Bull du Cent deRech de Pau ndash SNPA 10 379ndash469 1976
Berggren W A and Aubert J Paleocene benthonic foraminiferalbiostratigraphy paleobiogeography and paleoecology ofAtlanticndashTethyan regions midway-type fauna PalaeogeogrPalaeocl 18 73ndash192 1975
Bernaola G and Monechi S Calcareous nannofossil ex-tinction and survivorship across the CretaceousndashPaleogeneboundary at Walvis Ridge (ODP Hole 1262C SouthAtlantic Ocean) Palaeogeogr Palaeocl 255 132ndash156doi101016jpalaeo200702045 2007
Bernhard J M Characteristic assemblages and morphologies ofbenthic foraminifera from anoxic organic-rich deposits Jurassicthrough Holocene J Foramin Res 16 207ndash215 1986
Birch H S Coxall H K Pearson P N Kroon D andSchmidt D N Partial collapse of the marine carbon pump af-ter the CretaceousndashPaleogene boundary Geology 44 287ndash290doi101130G375811 2016
Brinkhuis H and Zachariasse W J Dinoflagellate cysts sealevel changes and planktonic foraminifers across the Cretaceous-
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
898 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
Tertiary boundary at El Haria northwest Tunisia Mar Micropa-leontol 13 153ndash191 1988
Brinkhuis H Bujak J P Smit J Versteegh G J M and Viss-cher H Dinoflagellate-based sea surface temperature recon-structions across the Cretaceous-Tertiary boundary PalaeogeogrPalaeocl 141 67ndash83 1998
Buesseler K O The decoupling of production and particulate ex-port in the surface ocean Global Biogeochem Cy 12 297ndash3101998
Buzas M A Culver S J and Jorissen F J A statisti-cal evaluation of the microhabitats of living (stained) infau-nal benthic foraminifera Mar Micropaleontol 20 311ndash320doi1010160377-8398(93)90040-5 1993
Coccioni R and Galeotti S KndashT boundary extinction geolog-ically instantaneous or gradual event Evidence from deep-seabenthic foraminifera Geology 22 779ndash782 1994
Corliss B H Microhabitats of benthic foraminifera within deep-sea sediments Nature 314 435ndash438 doi101038314435a01985
Corliss B H and Chen C Morphotype patterns of Nor-wegian Sea deep-sea benthic foraminifera and ecologi-cal implications Geology 16 716ndash719 doi1011300091-7613(1988)016lt0716MPONSDgt23CO2 1988
Coxall H K DrsquoHondt S and Zachos J C Pelagic evolution andenvironmental recovery after the CretaceousndashPaleogene mass ex-tinction Geology 34 297ndash300 doi101130G217021 2006
Culver S J Benthic foraminifera across the CretaceousndashTertiary(KndashT) boundary a review Mar Micropaleontol 47 177ndash2262003
Cushman J A Upper Cretaceous foraminifera of the Gulf coastalregion of the United States and adjacent areas US Geolog SurvProf Pap 206 241 pp 1946
Cushman J A Paleocene foraminifera of the Gulf Coastal Regionof the United States and adjacent regions US Geol Surv ProfPap 232 75 pp 1951
DrsquoHondt S Consequences of the CretaceousPaleogene Mass Ex-tinction for Marine Ecosystems Annu Rev Ecol Evol S36 295ndash317 doi101146annurevecolsys350211031057152005
DrsquoHondt S and Zachos J C Cretaceous foraminifera and the evo-lutionary history of planktic photosymbiosis Paleobiology 24512ndash523 1998
DrsquoHondt S Donaghay P Zachos J C Luttenberg D andLindinger M Organic Carbon Fluxes and Ecological Recov-ery from the Cretaceous-Tertiary Mass Extinction Science 282276ndash279 1998
Donze P Colin J-P Damotte R Oertli HJ Peypouquet J-Pand Said R Les ostracodes du Campanien terminal agrave lrsquoEocegraveneinfeacuterieur de la coupe du Kef Tuneacutesie nord-occidentale Bull desCentr Recher Expl-Prod Elf Aquit 6 273ndash335 1982
Dortch Q and Packard T T Differences in biomass structure be-tween oligotrophic and eutrophic marine ecosystems Deep-SeaRes 36 223ndash240 1989
Ernst S R Guasti E Dupuis C and Speijer R P En-vironmental perturbation in the southern Tethys across thePaleoceneEocene boundary (Dababiya Egypt) Foraminiferaland clay mineral records Mar Micropaleontol 60 89ndash111doi101016jmarmicro200603002 2006
Eshet Y Moshkovitz S Habib D Benjamini C and Maga-ritz M Calcareous nannofossil and dinoflagellate stratigraphyacross the CretaceousTertiary boundary at Hor Hahar IsraelMar Micropaleontol 18 199ndash228 1992
Eshet Y Almogi-Labin A and Bein A Dinoflagellate cysts pa-leoproductivity and upwelling systems A Late Cretaceous ex-ample from Israel Mar Micropaleontol 23 231ndash240 1994
Esmeray-Senlet S Wright J D Olsson R K MillerK G Browning J V and Quan T M Evidencefor reduced export productivity following the Creta-ceousPaleogene mass extinction Paleoceanography 30718ndash738 doi1010022014PA002724 2015
Giusberti L Boscolo Galazzo F and Thomas E Variability inclimate and productivity during the Paleocene-Eocene ThermalMaximum in the western Tethys (Forada section) Clim Past12 213ndash240 doi105194cp-12-213-2016 2016
Guasti E Kouwenhoven T J Brinkhuis H and Speijer R PPaleocene sea-level and productivity changes at the southernTethyan margin (El Kef Tunisia) Mar Micropaleontol 55 1ndash17 2005
Habib D and Saeedi F The Manumiella seelandica global spikeCooling during regression at the close of the MaastrichtianPalaeogeogr Palaeocl 255 87ndash97 2007
Hain M P Sigmal D M and Haug G H 818 ndash The biolog-ical Pump in the Past in Reference Module in Earth Systemsand Environmental Sciences Treatise on Geochemistry (SecondEdition) The Oceans and Marine Geochemistry 8 485ndash517doi101016B978-0-08-095975-700618-5 2014
Hayek L C and Buzas M A On the proper and efficient useof diversity measures with individual field samples J ForaminRes 43 305ndash313 2013
Henson S A Sanders R Madsen E Morris P J Le MoigneF and Quartly G D A reduced estimate of the strength ofthe oceanrsquos biological carbon pump Geophys Res Lett 38L04606 doi1010292011GL046735 2011
Hilting A K Kump L R and Bralower T J Variations in theoceanic vertical carbon isotope gradient and their implicationsfor the Paleocene-Eocene biological pump Paleoceanography23 PA3222 doi1010292007PA001458 2008
Honjo S Manganini S J Krishfield R A and Francois RParticulate organic carbon fluxes to the ocean interior and fac-tors controlling the biological pump A synthesis of global sed-iment trap progams since 1983 Prog Oceanogr 76 217ndash285doi101016jpocean200711003 2008
Huber B T MacLeod K G and Norris R D Abrupt extinctionand subsequent reworking of Cretaceous planktonic foraminiferaacross the Cretaceous-Tertiary boundary Evidence from the sub-tropical North Atlantic in Catastrophic Events and Mass Extinc-tions Impacts and Beyond edited by Koeberl C and MacLeodK G Geol S Am S 356 277ndash289 Boulder Colorado USA2002
Hull P M Norris R D Bralower T J and Schueth J D A rolefor chance in marine recovery from the end-Cretaceous extinc-tion Nat Geosci 4 856ndash860 doi101038ngeo1302 2011
Hsuuml K J and McKenzie J A A ldquoStrangeloverdquo ocean in the earli-est Tertiary in Natural variations Archean to present edited bySundquist E T and Broecker W S The carbon cycle and atmo-spheric CO2 American Geophysical Union Washington DCUSA 487ndash492 1985
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
J Vellekoop et al Ecological response to KndashPg boundary mass extinction 899
Jiao N Herndl G J Hansell D A Benner R Kattner G Wil-helm S W Kirchman D L Weinbauer M G Luo T ChenF and Azam F Microbial production of recalcitrant dissolvedorganic matter long-term carbon storage in the global oceanNat Rev 8 593ndash599 doi101038nrmicro2386 2010
Jorissen F J de Stigter H C and Widmark J G V A concep-tual model explaining benthic foraminiferal microhabitats MarMicropaleontol 26 3ndash15 doi1010160377-8398(95)00047-X1995
Jorissen F J Fontanier C and Thomas E Paleoceanographi-cal proxies based on deep-sea benthic foraminiferal assemblagecharacteristics in Proxies in Late Cenozoic PaleoceanographyPt 2 Biological tracers and biomarkers edited by Hillaire-Marcel C and de Vernal A 263ndash326 Elsevier Amsterdamthe Netherlands 2007
Kaiho K Kajiwara Y Tazaki K Ueshima M Takeda NKawahata H Arinobu T Ishiwatari R Hirai A and Lam-olda M A Oceanic primary productivity and dissolved oxygenlevels at the CretaceousTertiary boundary their decrease sub-sequent warming and recovery Paleoceanography 14 511ndash5241999
Keller G and Lindinger M Stable isotope TOC and CaCO3record across the CretaceousTertiary boundary at El KefTunisia Palaeogeogr Palaeocl 73 243ndash265 1989
Kellough G R Paleoecology of the Foraminiferida of the WillsPoint Formation (Midway Group) in northeast Texas Trans GulfCoast Assoc Geol Soc 15 73ndash153 1965
Kring D A The Chicxulub impact event and its environmentalconsequences at the CretaceousndashTertiary boundary PalaeogeogrPalaeocl 255 4ndash21 2007
Kump L R Interpreting carbon-isotope excursions Strangeloveoceans Geology 19 299ndash302 1991
Machalski M Vellekoop J Dubicka Z Peryt D and Ha-rasimiuk M Late Maastrichtian cephalopods dinoflagellatecysts and foraminifera from the CretaceousndashPaleogene suc-cession at Lechoacutewka southeast Poland Stratigraphic andenvironmental implications Cretaceous Res 57 208ndash227doi101016jcretres201508012 2016
Molina E Evidence and causes of the main extinctionevents in the Paleogene based on extinction and survivalpatterns of foraminifera Earth-Sci Rev 140 166ndash181doi101016jearscirev201411008 2015
Molina E Alegret L Arenillas I and Arz J The GlobalBoundary Stratotype Section and Point for the base of the Da-nian Stage (Paleocene Paleogeneldquo Tertiaryrdquo Cenozoic) at ElKef Tunisia-Original definition and revision Episodes 29 263ndash273 2006
Olsson R K Liu C and van Fossen M The Cretaceous-Tertiarycatastrophic event at Millers Ferry Alabama in The Cretaceous-Tertiary Event and Other Catastrophes in Earth History editedby Ryder G Fastovsky D and Gartner S Geol S Am S307 263ndash277 1996
Peryt D Benthic foraminiferal response to the Cenomanian-Turonian and CretaceousndashPaleogene boundary events PrzegladGeol 52 827ndash832 2004
Peryt D Alegret L and Molina E The CretaceousPalaeogene(KP) boundary at Aiumln Settara Tunisia restructuring of benthicforaminiferal assemblages Terra Nova 14 101ndash107 2002
Ramette A Multivariate analyses in microbial ecology FEMS Mi-crobiol Ecol 62 142ndash160 2007
Rodriacuteguez-Tovar F J Uchman A MrsquoHamdi A Riahi Sand Ismail-Lattrache K B Ichnological record of palaeoen-vironment from the CretaceousndashPaleogene boundary intervalat El Kef Tunisia The first study of old and new sec-tions at the stratotype area J Afr Earth Sci 120 23ndash30doi101016jjafrearsci201604017 2016
Saner S Mudurnu-Goumlynuumlk Havzasının Jura ve Sonrasıccediloumlkelimnitelikleriyle paleocografya yorumlaması TJK Buumllteni 23 39ndash52 1980
Schioslashler P Brinkhuis H Roncaglia L and Wilson G J Di-noflagellate biostratigraphy and sequence stratigraphy of theType Maastrichtian (Upper Cretaceous) ENCI Quarry TheNetherlands Mar Micropaleontol 31 65ndash95 1997
Schueth J D Bralower T J Jiang S and Patzkowsky ME The role of regional survivor incumbency in the evolu-tionary recovery of calcareous nannoplankton from the Creta-ceousPaleogene (KPg) mass extinction Paleobiology 41 661ndash679 doi101017pab201528 2015
Scotese C R A continental drift flipbook J Geology 112 729ndash741 2004
Scotese C R and Dreher C GlobalGeology available at httpwwwGlobalGeologycom (last access 11 March 2014) 2012
Sepkoski J J Patterns of Phanerozoic extinction a perspectivefrom global data bases in Global Events and Event Stratigra-phy edited by Walliser O H Springer Berlin Germany 35ndash52 2016
Sibert E C Hull P M and Norris R D Resilience of Pacificpelagic fish across the CretaceousPalaeogene mass extinctionNat Geosci 7 667ndash670 doi101038NGEO2227 2014
Slimani H and Toufiq A A CretaceousndashPaleogene boundary geo-logical site revealed by planktic foraminifera and dinoflagellatecysts at Ouled Haddou eastern external Rif Chain Morocco JAfr Earth Sci 88 38ndash52 doi101016jjafrearsci2013080082013
Slimani H Louwye S and Toufiq A Dinoflagellate cystsform the CretaceousndashPaleogene boundary at Ouled Had-dou southeastern Rif Morocco biostratigraphy paleoenvi-ronments and paleobiogeography Palynology 34 90ndash124doi10108001916121003629933 2010
Sluijs A and Brinkhuis H A dynamic climate and ecosystemstate during the Paleocene-Eocene Thermal Maximum infer-ences from dinoflagellate cyst assemblages on the New JerseyShelf Biogeosciences 6 1755ndash1781 doi105194bg-6-1755-2009 2009
Smit J Extinction and evolution of planktonic foraminifera after amajor impact at the CretaceousTertiary boundary Geol S AmS 190 329ndash352 1982
Smit J and Romein A J T A sequence of events across theCretaceous-Tertiary boundary Earth Planet Sci Lett 74 155ndash170 1985
Speijer R P Extinction and recovery patterns in ben-thic foraminiferal paleocommunities across the Creta-ceousPaleogene and PaleoceneEocene boundaries GeologicaUltraiectina 124 Faculteit Aardwetenschappen PhD ThesisUniversiteit Utrecht Utrecht the Netherlands 1994
Speijer R P and van Der Zwaan G J Extinction and survivor-ship of southern Tethyan benthic foraminifera across the Cre-
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
900 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
taceousPaleogene boundary in Biotic recovery from mass ex-tinction events edited by Hart M B Geolog Soc Spec PublLondon UK 102 343ndash371 1996
Stemmann L and Boss E Plankton and Particle Size andPackaging From Determining Optical Properties to Drivingthe Biological Pump Annu Rev Mar Sci 4 263ndash290doi101146annurev-marine-120710-100853 2012
Thomas E Late Cretaceous-early Eocene mass extinctions in thedeep sea Geol S Am S 247 481ndash495 1990
Vandenberghe N Hilgen F J and Speijer R P The PaleogenePeriod in The Geologic Time Scale 2012 edited by GradsteinF M Ogg J G Schmitz M D and Ogg G M 2 855ndash921Elsevier Boston USA 2012
Vellekoop J Sluijs A Smit J Schouten S Weijers J WH Sinninghe Damsteacute J S and Brinkhuis H Rapid short-term cooling following the Chicxulub impact at the CretaceousndashPaleogene boundary P Natl Acad Sci USA 111 7537ndash7541doi101073pnas1319253111 2014
Vellekoop J Smit J van de Schootbrugge B Weijers J W HGaleotti S Sinninghe Damsteacute J S and Brinkhuis H Palyno-logical evidence for prolonged cooling along the Tunisian conti-nental shelf following the KndashPg boundary impact PalaeogeogrPalaeocl 426 216ndash228 doi101016jpalaeo2015030212015
Vellekoop J Esmeray-Senlet S Miller K G Browning J VSluijs A van de Schootbrugge B Sinninghe Damsteacute J Sand Brinkhuis H Evidence for CretaceousndashPaleogene bound-ary bolide ldquoimpact winterrdquo conditions from New Jersey USAGeology 44 619ndash622 doi101130G379611 2016
Woelders L and Speijer R Stable seafloor conditions sealevel and food supply during the latest Maastrichtian atBrazos River Texas Mar Micropaleontol 121 41ndash51doi101016jmarmicro201510002 2015
Zachos J C Arthur M A and Dean W E Geochemical evi-dence for suppression of pelagic marine productivity at the Cre-taceousTertiary boundary Nature 337 61ndash64 1989
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
- Abstract
- Introduction
- Geological setting and age assessment
- Materials and methods
-
- Sampling
- Foraminiferal analysis
- Palynological analysis
- Statistical analysis
-
- Results
-
- Benthic foraminifera
- Palynology
-
- Discussion
-
- Tethyan benthic foraminiferal turnover sequence
- Ecological responses to reduced export productivity
-
- Okccedilular section
- Western Tethys
- Global responses
-
- Long-term recovery
-
- Conclusions
- Data availability
- Author contributions
- Competing interests
- Acknowledgements
- References
-

J Vellekoop et al Ecological response to KndashPg boundary mass extinction 893
Figure 6 Relative abundances of the four main clusters of benthic foraminiferal taxa (gt 125 microm) relative abundances of epibenthic andendobenthic forms and relative abundance of bi-triserial benthic taxa
Figure 7 Q-mode non-metric multidimensional scaling (on all sam-ples in the gt 125 microm size fraction benthic foraminiferal datasets ofOkccedilular (this study) and El Kef (Speijer and van der Zwaan 1996)After an abrupt transition from latest Maastrichtian pre-impact as-semblages to disaster assemblages in Zone P0 and recovery assem-blages in Zone Pα a gradual transition towards a new equilibriumpost-recovery assemblage can be observed in subzones P1a and P1b
the biological crisis following the impact Following the KndashPg boundary mass extinction endobenthic forms stronglydecrease with bi-triserial benthic taxa almost disappearingfrom the benthic community simultaneously the estimatedBFAR shows a major fall (Fig 5) This interval approxi-mately correlative to foraminiferal Zone P0 represents thedisaster phase The drop in bi-triserial benthic taxa (Fig 6)and decrease in BFAR (Fig 5) point to a reduction in foodsupply to the benthic community indicating that the trans-port of organic matter to the sea floor must have decreasedIn the dinocyst community however hexaperidinioids showa strong increase in relative abundance during this phase(Figs 3 6 7) suggesting an increase in nutrient availabilityin the photic zone while the DAR remains relatively stableacross the KndashPg boundary (Fig 5)
The decrease in benthic diversity and the high relativeabundances of opportunistic Trochammina spp and Cibici-doides pseudoacutus suggest that the benthic community ex-perienced additional stress during Zone P0 besides food lim-itation (see Jorissen et al 2007) If food limitation were theonly factor involved upper bathyal assemblages as observedat Okccedilular would be expected to become more similar tooligotrophic middle to lower bathyal assemblages Insteadthe opposite is true the assemblage composition of the dis-aster phase is closer to neritic Midway-type assemblages
It was previously suggested that the epibenthic taxon Cibi-cidoides pseudoacutus could potentially be hypoxia resistant
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
894 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
(Speijer and van der Zwaan 1996) As a collapse of the bi-ological pump should lead to enhanced remineralization oforganic matter in the photic zone it is indeed likely thatthe oxygen minimum zones expanded and shoaled leadingto oxygen stress on the sea floor Although early Paleocenedysoxia at El Kef is supported by the ostracod record (Donzeet al 1982) and ichnofacies analyses (Rodriguez-Tovaret al 2016) supporting (eg geochemical) evidence fordysoxia during the disaster phase at Okccedilular record is absentFurthermore an additional stressor for the benthic commu-nity during Zone P0 may be present In contrast to calcare-ous nanoplankton organic-walled cyst-producing dinoflagel-lates did not suffer extinctions and may have become a moreimportant component in the early Paleocene phytoplanktoncommunity Hence besides the reduction of the amount oforganic matter transported to the sea floor the compositionof food supplied by the photic zone likely changed signifi-cantly across the KndashPg boundary as well (DrsquoHondt 2005)possibly presenting an additional stress factor for the benthiccommunity (Speijer and van der Zwaan 1996 Alegret andThomas 2009)
522 Western Tethys
The patterns in benthic foraminiferal and dinoflagellate re-sponse to the KndashPg boundary perturbations at Okccedilular appearto be characteristic for the continental margins of the westernTethyan realm The benthic foraminiferal patterns are similaracross different size fractions suggesting that they are repre-sentative of the overall benthic communities In Tunisia andSpain where both benthic foraminiferal and dinoflagellaterecords are available the earliest Danian is also characterizedby a decrease in endobenthic foraminifera at the sea floor(Speijer and Van der Zwaan 1996 Peryt et al 2002 Ale-gret et al 2003) and simultaneously blooms of hexaperidin-ioids in the water column (Brinkhuis et al 1998 Vellekoopet al 2015 Fig 8) In the Ouled Haddou section Moroccoin the westernmost Tethys (Fig 1) the lowermost Danian isalso characterized by a strong increase of the hexaperidin-ioid Senegalinium group up to 30 of the assemblage (Sli-mani et al 2010) that is very similar to the Okccedilular record(Fig 8)
The KndashPg boundary dinocyst record of El Kef is strikinglysimilar to the one from the Okccedilular section showing two dis-tinct peaks in hexaperidinioids one in Zone P0 and one atthe basal part of Zone Pα At El Kef the initial post-impactdominance of hexaperidinioids is however less pronounced(Fig 8) This difference in expression might be related tosmall differences in paleogeographic and paleoceanographicsettings between sites
In addition some high-resolution benthic KndashPg records inthe Tethyan realm show a short-lived bloom of specific en-dobenthic foraminiferal taxa in the few centimeter directlyabove the KndashPg boundary sometimes accompanied by otherindicators of low oxygen levels (eg Coccioni and Gale-
otti 1994 Speijer and Van der Zwaan 1996 Kaiho et al1999) This suggests that at least locally the sea floor wastemporarily characterized by hypoxic conditions directly fol-lowing the impact This short-lived deoxygenation is possi-bly related to the mass mortality at the KndashPg boundary re-sulting in a large short-lived flux of organic matter to thesea floor (Coccioni and Galeotti 1994) However since ex-port productivity was greatly reduced after the KndashPg bound-ary mass extinction very little ldquonewrdquo organic matter reachedthe sea floor after the initial post-impact influx Hence afteraccumulated organic matter was remineralized the benthiccommunity starved resulting in a transition to an epibenthic-dominated benthic fauna similar to the post-impact fauna ofthe Okccedilular record
The combined dinocyst records suggest that the early Pa-leocene margins of the Tethys were characterized by an in-crease in nutrient availability in the photic zone whereascoeval benthic foraminiferal records indicate a major de-crease in nutrient supply to the sea floor These changesin the benthic foraminiferal and dinocyst communities arelikely caused by the major reduction of both the efficiencyand strength of the biological pump (ie the fraction ofprimary production exported from the photic zone and theamount of organic matter transported down) in accordancewith the Living Ocean model (DrsquoHondt and Zachos 1998DrsquoHondt et al 1998 DrsquoHondt 2005 Coxall et al 2006)For a detailed explanation of the difference between biologi-cal pump efficiency and biological pump strength see Hiltinget al (2008)
As bulk carbonate δ13C reflects the isotopic compositionof the surface ocean which is set by burial fractions and bythe photosynthetic isotope effect (Kump 1991 Hain et al2014) the excursion recorded in bulk δ13C records world-wide and the rapid collapse in surface to deep-ocean carbonisotope gradients likely reflect the reduction of the global in-tensity (ie efficiency) of the biological pump (Hain et al2014) The recorded inverse change in nutrient availabilitysuggests a causal link A reduced efficiency of the biologi-cal pump and associated decrease of the fraction of biomasstransported out of the photic zone could have resulted inhigh rates of nutrient recycling in the upper part of the wa-ter column (DrsquoHondt 2005) This suggests that the reduc-tion of the biological pump strength recorded by the benthicforaminiferal record is a consequence of the decreased effi-ciency of the biological pump recorded by the carbon iso-tope and dinoflagellate cyst records The strong correlationbetween the bulk carbon isotope curves and the Shannondiversity index (H ) of the benthic foraminiferal records atboth Okccedilular (R2
= 073 p lt 0001) and El Kef (R2= 074
p lt 0001) suggest a clear link between the changes in ben-thic foraminiferal assemblages and the collapse and recoveryof biological pump efficiency
As a result of the reduced biological pump efficiency morenutrients remained available for the early Paleocene phyto-plankton community At Tethyan margin sites this is indi-
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
J Vellekoop et al Ecological response to KndashPg boundary mass extinction 895
Figure 8 The benthic foraminiferal (gt 125 microm) dinocyst and bulk stable carbon isotope records of Okccedilular (Turkey) and El Kef (Tunisia)the dinocyst record of Ouled Haddou (Morocco) and the benthic foraminiferal record (gt 63 microm) of ODP Hole 1049C Blake Nose (NorthAtlantic) Note that the definition of ldquohexaperidinioidsrdquo used is provided in the text The four phases identified based on the bulk stablecarbon isotopes and the Shannon diversity index (H ) of the benthic foraminiferal records are indicated (IndashIV) The gt 63 microm size fractionbenthic foraminiferal record of ODP Hole 1049C is from Alegret and Thomas (2004) who argued that the lowermost Paleocene of thissite comprised reworked foraminifera This interval is indicated by a dashed bar in the figure The biostratigraphy of Ouled Haddou is fromSlimani and Toufiq (2013) whereas the dinocyst record of this site is from Slimani et al (2010) The biostratigraphy of El Kef is based onBrinkhuis et al (1998) Speijer and van der Zwaan (1996) and Molina et al (2006) The fine-fraction stable carbon isotope record (blackdots) of El Kef is from Keller and Lindinger (1989) The Shannon diversity index (red dots) relative abundances of bi-triserial benthic taxa(dark purple) and all endobenthic foraminifera (light purple) are based on the gt 125 microm size fraction foraminiferal data of Speijer and vander Zwaan (1996) The palynological data of El Kef are from Brinkhuis et al (1998) (lower part) and Guasti et al (2005) (upper part) TheShannon diversity index of the gt 125 microm size fraction benthic foraminiferal record (red dots) the relative contribution of bi-triserial benthictaxa (dark purple) all endobenthic foraminifera (light purple) and hexaperidinioid dinocysts from Okccedilular are from this study The bulkstable carbon isotope record of Okccedilular (black dots) is from Accedilikalin et al (2015) Mbsf indicates meters below sea floor
cated by the higher abundance of hexaperidinioids Similarlyblooms of eutrophic survivor taxa of calcareous nanoplank-ton at open ocean sites have been suggested to be relatedto the buildup of nutrients in the open ocean photic zone(Schueth et al 2015) Although the carbon isotope gradientsbetween surface and deep waters indicate that the recoveryof the biological pump took hundreds of thousands of years(Zachos et al 1989 Kump 1991 DrsquoHondt et al 1998 Cox-all et al 2006 Birch et al 2016) our records suggest thatthe enhanced recycling of nutrients in the Tethys was partic-ularly intense during two phases in the first tens of thousandsyears after the impact (Fig 8)
523 Global responses
The decrease in export productivity following the KndashPgboundary is not only recognized in the Tethyan realm In theAtlantic Ocean Southern Ocean and the Indian Ocean a de-crease of the biological pump strength after the KndashPg bound-ary was also observed in neritic to abyssal environments(eg Thomas 1990 Olsson et al 1996 Hull et al 2011Alegret et al 2012) As an example the gt 63 microm size frac-tion benthic foraminiferal KndashPg boundary record of BlakeNose (ODP Hole 1049C northwestern Atlantic Alegret andThomas 2004) shows a pattern that is strikingly similar tothe gt 125 microm size fraction records in the Tethyan realm witha strong decrease of endobenthics in Zone P0 followed by a
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
896 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
gradual recovery across zones Pα and P1a (Fig 8) The simi-larities between the benthic foraminiferal patterns of Tethyanouter neritic to middle bathyal (200ndash1500 m) sites and thoseof the middle to lower bathyal (1500ndash1600 m) record ofBlake Nose (Alegret and Thomas 2004) suggest that theserecords register a global decrease in export productivity fol-lowing the KndashPg boundary
However several open ocean sites mostly in the Pacificshow an opposite trend with several lines of evidence sug-gesting an increased biological pump strength at these sites(eg Hull et al 2011 Alegret et al 2012) This led Ale-gret et al (2012) to conclude that the decrease in biologicalpump strength after the KndashPg boundary was most likely aregional instead of a global effect arguing against the Liv-ing Ocean hypothesis We would argue that low-productivityopen ocean sites like the central Pacific are characterized byvery different ecosystem structures and higher productivityas those along the Tethyan margins (eg Dortch and Pack-erd 1989) Therefore the consequence of ecosystem reorga-nization resulting from the mass extinction was likely verydifferent in the open ocean Pacific (eg Sibert et al 2014)Esmeray-Senlet et al (2015) proposed the term ldquoHeteroge-neous Oceanrdquo for this conceptual model as an alternativeto the Living Ocean model Supposedly the HeterogeneousOcean was characterized by a strong geographic heterogene-ity in the extinction patterns and food supply to the sea floor
However as the primary food source for open ocean deep-sea benthic communities generally consists of refractory or-ganic matter (eg Jiao et al 2010) it remains uncertainwhether changes in the intensity of recycling of the gener-ally more labile organic matter in the surface ocean will re-sult in changes in the amount of organic matter that benthiccommunities receive as they will likely continue to receiverecalcitrant organic matter at barely diminished rates despitethe collapse of the biological pump
In addition it should be noted that although the bio-logical pump efficiency is generally relatively high in openocean realms such as the central Pacific the biological pumpstrength is generally very low at these sites (Honjo et al2008 Henson et al 2011) Small changes in absolute bio-logical pump strength at such a locality could therefore havehad a large effect on the local oligotrophic benthic commu-nity (Alegret and Thomas 2009) but they likely representedonly a small fraction of the net amount of organic matterglobally transported to the deep Hence even though localbiological pump strength might have increased at some low-productivity deep-sea sites the large decrease in biologi-cal pump strength recorded at many other sites means thatthe net amount of organic matter globally transported out ofthe surface ocean was likely still reduced in the post-impactworld (Birch et al 2016) Therefore we argue that such Pa-cific records do not represent the general pattern The globalocean response can be characterized as heterogeneous how-ever the KndashPg boundary mass extinction did lead to a strongreduction of the net amount of organic matter globally trans-
ported from the surface to the sea floor while biological pro-ductivity recovered rapidly after the extinction event in ac-cordance with the Living Ocean hypothesis
53 Long-term recovery
Following the disaster phase the hexaperidinioid cysts atOkccedilular and El Kef decreased in abundance and the abun-dance of endobenthic forms increased again as the recoveryof the benthic community was initiated This recovery phaseis approximately correlative to Zone Pα and the basal part ofZone P1a which according to the Paleogene age constraintsof Vandenberghe et al (2012) represents at least 300 kyrThis duration is in agreement with the estimations providedby Birch et al (2016) During this phase the carbon iso-tope records remain well below pre-impact values and thediversity of the benthic foraminiferal community has not yetfully recovered (Figs 4ndash8) This indicates that the rapid andshort-lived KndashPg boundary disaster was followed by a rela-tively long recovery phase in line with previous estimatesof a multimillion-year biological recovery (eg Coxall etal 2006) While the impact-related environmental perturba-tions were short lived (Kring 2007 Vellekoop et al 20142016) the extinctions amongst important biological groupsled to a reduction of the organic flux from the photic zone todeep water resulting in major long-term biological and pa-leoceanographic reorganizations Only with the evolutionaryrecovery of the pelagic community governing the biologicalcarbon pump did export productivity start to increase again(eg Coxall et al 2006 Birch et al 2016)
6 Conclusions
The marine palynological benthic foraminiferal and bulkstable isotope records of the Okccedilular and El Kef sections re-veal major changes across the KndashPg boundary interval por-traying the biological crisis at the KndashPg boundary and subse-quent recovery in the early Paleocene Based on the succes-sion of benthic faunal assemblages at a number of Tethyanmargin sites four phases can be recognized across the KndashPgboundary interval the Maastrichtian or pre-impact phase (I)a disaster phase (II) a recovery phase (III) and an early Pa-leocene post-recovery phase (IV)
Following the KndashPg boundary impact some localitiesshow a large short-lived flux of food to the sea floor likelyrelated to the mass mortality at the KndashPg boundary Howeversince export productivity was greatly reduced after the KndashPgboundary mass extinction it is likely that after the initialpost-impact influx very little fresh organic matter reachedthe sea floor This reduction of export productivity in thepost-extinction disaster phase eventually resulted in a lowerfood supply to the sea floor This presented a major stressfactor for benthic organisms in the western Tethys The re-duced food supply resulted in an abrupt impoverishment of
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
J Vellekoop et al Ecological response to KndashPg boundary mass extinction 897
benthic communities As the downward transport of nutri-ents was slowed down recycling in upper layers increasedAs a result more nutrients remained available for the earlyPaleocene phytoplankton community leading to blooms ofdinoflagellates along the Tethyan margins and blooms of cal-careous nanoplankton taxa in more open ocean sites Ourrecords show that the enhanced recycling of nutrients in theTethys was particularly intense in the tens of thousands ofyears after the impact Following this the slow evolution-ary recovery of the pelagic community governing the bio-logical carbon pump resulted in a gradually increasing ex-port productivity in the hundreds of thousands of years afterthe impact Hence our integration of dinocyst and benthicforaminiferal records across the KndashPg boundary provides im-portant new insights into the ecological responses to the re-duction of export productivity following the mass extinctionat the CretaceousndashPaleogene boundary highlighting the di-rect ecological consequences of the Living Ocean conditionsin the post-impact world
7 Data availability
All data related to publication are available in the Supple-ment
The Supplement related to this article is available onlineat doi105194bg-14-885-2017-supplement
Author contributions Johan Vellekoop Lineke Woelders andHenk Brinkhuis designed the research Samples were collectedin the field by Johan Vellekoop Sanem Accedilikalin Ismail Ouml Yil-maz and Jan Smit Palynological analyses were carried out by Jo-han Vellekoop Benthic foraminiferal analyses were carried out byLineke Woelders and Robert P Speijer Johan Vellekoop LinekeWoelders and Bas van de Schootbrugge prepared the manuscriptwith input from all authors
Competing interests The authors declare that they have no conflictof interest
Acknowledgements This work was supported by the NetherlandsOrganization for Scientific Research (NWO) Open CompetitionGrant ALWPJ09047 to H Brinkhuis and the Research FoundationFlanders (FWO) grant G0B8513 to R P Speijer We thank LKump F Boscolo Galazzo P Roopnarine and one anonymousreferee for their suggestions
Edited by D GillikinReviewed by L Kump F Boscolo Galazzo and P Roopnarine
References
Accedilikalin S Vellekoop J Ocakoglu F Yılmaz I Ouml Smit J Al-tiner S O Goderis S Vonhof H Speijer R P WoeldersL Fornaciari E and Brinkhuis H Geochemical and pale-ontological characterization of a new KndashPg Boundary local-ity from the Northern branch of the Neo-Tethys MudurnundashGoumlynuumlk Basin NW Turkey Cretaceous Res 52 251ndash267doi101016jcretres201407011 2015
Alegret L and Thomas E Food supply to the seafloor in the Pa-cific Ocean after the CretaceousPaleogene boundary event MarMicropaleontol 73 105ndash116 2009
Alegret L Molina E and Thomas E Benthic foraminiferalturnover across the CretaceousPaleogene boundary at Agost(southeastern Spain) paleoenvironmental inferences Mar Mi-cropaleontol 48 251ndash279 2003
Alegret L Thomas E and Lohmann K C End-Cretaceous ma-rine mass extinction not caused by productivity collapse P NatlAcad Sci USA 109 728ndash732 doi101073pnas11106011092012
Alegret L Rodriacuteguez-Tovar F J and Uchman A How bioturba-tion obscured the CretaceousndashPaleogene boundary record TerraNova 27 225ndash230 doi101111ter12151 2015
Altiner D Koccedilyigit A Farinacci A Nicosia U and ContiM A Jurassic-Lower Cretaceous stratigraphy and paleogeo-graphic evolution of the southern part of north-western Anatolia(Turkey) Geologica Romana 27 13ndash80 1991
Alvarez LW Alvarez W Asaro F and Michel H V Extrater-restrial cause for the Cretaceous-Tertiary extinction Science208 1095ndash1108 1980
Alegret L and Thomas E Benthic foraminifera and environmen-tal turnover across the CretaceousPaleogene boundary at BlakeNose (ODP Hole 1049C Northwestern Atlantic) PalaeogeogrPalaeocl 208 59ndash83 doi101016jpalaeo200402028 2004
Armstrong R A Lee C Hedges J I Honjo S and WakehamS G A new mechanistic model for organic carbon fluxes in theocean based on the quantitative association of POC with ballastmaterials Deep-Sea Res Pt II 49 219ndash236 2002
Aubert J and Berggren W A Paleocene benthic foraminiferalbiostratigraphy and paleoecology of Tunisia Bull du Cent deRech de Pau ndash SNPA 10 379ndash469 1976
Berggren W A and Aubert J Paleocene benthonic foraminiferalbiostratigraphy paleobiogeography and paleoecology ofAtlanticndashTethyan regions midway-type fauna PalaeogeogrPalaeocl 18 73ndash192 1975
Bernaola G and Monechi S Calcareous nannofossil ex-tinction and survivorship across the CretaceousndashPaleogeneboundary at Walvis Ridge (ODP Hole 1262C SouthAtlantic Ocean) Palaeogeogr Palaeocl 255 132ndash156doi101016jpalaeo200702045 2007
Bernhard J M Characteristic assemblages and morphologies ofbenthic foraminifera from anoxic organic-rich deposits Jurassicthrough Holocene J Foramin Res 16 207ndash215 1986
Birch H S Coxall H K Pearson P N Kroon D andSchmidt D N Partial collapse of the marine carbon pump af-ter the CretaceousndashPaleogene boundary Geology 44 287ndash290doi101130G375811 2016
Brinkhuis H and Zachariasse W J Dinoflagellate cysts sealevel changes and planktonic foraminifers across the Cretaceous-
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
898 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
Tertiary boundary at El Haria northwest Tunisia Mar Micropa-leontol 13 153ndash191 1988
Brinkhuis H Bujak J P Smit J Versteegh G J M and Viss-cher H Dinoflagellate-based sea surface temperature recon-structions across the Cretaceous-Tertiary boundary PalaeogeogrPalaeocl 141 67ndash83 1998
Buesseler K O The decoupling of production and particulate ex-port in the surface ocean Global Biogeochem Cy 12 297ndash3101998
Buzas M A Culver S J and Jorissen F J A statisti-cal evaluation of the microhabitats of living (stained) infau-nal benthic foraminifera Mar Micropaleontol 20 311ndash320doi1010160377-8398(93)90040-5 1993
Coccioni R and Galeotti S KndashT boundary extinction geolog-ically instantaneous or gradual event Evidence from deep-seabenthic foraminifera Geology 22 779ndash782 1994
Corliss B H Microhabitats of benthic foraminifera within deep-sea sediments Nature 314 435ndash438 doi101038314435a01985
Corliss B H and Chen C Morphotype patterns of Nor-wegian Sea deep-sea benthic foraminifera and ecologi-cal implications Geology 16 716ndash719 doi1011300091-7613(1988)016lt0716MPONSDgt23CO2 1988
Coxall H K DrsquoHondt S and Zachos J C Pelagic evolution andenvironmental recovery after the CretaceousndashPaleogene mass ex-tinction Geology 34 297ndash300 doi101130G217021 2006
Culver S J Benthic foraminifera across the CretaceousndashTertiary(KndashT) boundary a review Mar Micropaleontol 47 177ndash2262003
Cushman J A Upper Cretaceous foraminifera of the Gulf coastalregion of the United States and adjacent areas US Geolog SurvProf Pap 206 241 pp 1946
Cushman J A Paleocene foraminifera of the Gulf Coastal Regionof the United States and adjacent regions US Geol Surv ProfPap 232 75 pp 1951
DrsquoHondt S Consequences of the CretaceousPaleogene Mass Ex-tinction for Marine Ecosystems Annu Rev Ecol Evol S36 295ndash317 doi101146annurevecolsys350211031057152005
DrsquoHondt S and Zachos J C Cretaceous foraminifera and the evo-lutionary history of planktic photosymbiosis Paleobiology 24512ndash523 1998
DrsquoHondt S Donaghay P Zachos J C Luttenberg D andLindinger M Organic Carbon Fluxes and Ecological Recov-ery from the Cretaceous-Tertiary Mass Extinction Science 282276ndash279 1998
Donze P Colin J-P Damotte R Oertli HJ Peypouquet J-Pand Said R Les ostracodes du Campanien terminal agrave lrsquoEocegraveneinfeacuterieur de la coupe du Kef Tuneacutesie nord-occidentale Bull desCentr Recher Expl-Prod Elf Aquit 6 273ndash335 1982
Dortch Q and Packard T T Differences in biomass structure be-tween oligotrophic and eutrophic marine ecosystems Deep-SeaRes 36 223ndash240 1989
Ernst S R Guasti E Dupuis C and Speijer R P En-vironmental perturbation in the southern Tethys across thePaleoceneEocene boundary (Dababiya Egypt) Foraminiferaland clay mineral records Mar Micropaleontol 60 89ndash111doi101016jmarmicro200603002 2006
Eshet Y Moshkovitz S Habib D Benjamini C and Maga-ritz M Calcareous nannofossil and dinoflagellate stratigraphyacross the CretaceousTertiary boundary at Hor Hahar IsraelMar Micropaleontol 18 199ndash228 1992
Eshet Y Almogi-Labin A and Bein A Dinoflagellate cysts pa-leoproductivity and upwelling systems A Late Cretaceous ex-ample from Israel Mar Micropaleontol 23 231ndash240 1994
Esmeray-Senlet S Wright J D Olsson R K MillerK G Browning J V and Quan T M Evidencefor reduced export productivity following the Creta-ceousPaleogene mass extinction Paleoceanography 30718ndash738 doi1010022014PA002724 2015
Giusberti L Boscolo Galazzo F and Thomas E Variability inclimate and productivity during the Paleocene-Eocene ThermalMaximum in the western Tethys (Forada section) Clim Past12 213ndash240 doi105194cp-12-213-2016 2016
Guasti E Kouwenhoven T J Brinkhuis H and Speijer R PPaleocene sea-level and productivity changes at the southernTethyan margin (El Kef Tunisia) Mar Micropaleontol 55 1ndash17 2005
Habib D and Saeedi F The Manumiella seelandica global spikeCooling during regression at the close of the MaastrichtianPalaeogeogr Palaeocl 255 87ndash97 2007
Hain M P Sigmal D M and Haug G H 818 ndash The biolog-ical Pump in the Past in Reference Module in Earth Systemsand Environmental Sciences Treatise on Geochemistry (SecondEdition) The Oceans and Marine Geochemistry 8 485ndash517doi101016B978-0-08-095975-700618-5 2014
Hayek L C and Buzas M A On the proper and efficient useof diversity measures with individual field samples J ForaminRes 43 305ndash313 2013
Henson S A Sanders R Madsen E Morris P J Le MoigneF and Quartly G D A reduced estimate of the strength ofthe oceanrsquos biological carbon pump Geophys Res Lett 38L04606 doi1010292011GL046735 2011
Hilting A K Kump L R and Bralower T J Variations in theoceanic vertical carbon isotope gradient and their implicationsfor the Paleocene-Eocene biological pump Paleoceanography23 PA3222 doi1010292007PA001458 2008
Honjo S Manganini S J Krishfield R A and Francois RParticulate organic carbon fluxes to the ocean interior and fac-tors controlling the biological pump A synthesis of global sed-iment trap progams since 1983 Prog Oceanogr 76 217ndash285doi101016jpocean200711003 2008
Huber B T MacLeod K G and Norris R D Abrupt extinctionand subsequent reworking of Cretaceous planktonic foraminiferaacross the Cretaceous-Tertiary boundary Evidence from the sub-tropical North Atlantic in Catastrophic Events and Mass Extinc-tions Impacts and Beyond edited by Koeberl C and MacLeodK G Geol S Am S 356 277ndash289 Boulder Colorado USA2002
Hull P M Norris R D Bralower T J and Schueth J D A rolefor chance in marine recovery from the end-Cretaceous extinc-tion Nat Geosci 4 856ndash860 doi101038ngeo1302 2011
Hsuuml K J and McKenzie J A A ldquoStrangeloverdquo ocean in the earli-est Tertiary in Natural variations Archean to present edited bySundquist E T and Broecker W S The carbon cycle and atmo-spheric CO2 American Geophysical Union Washington DCUSA 487ndash492 1985
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
J Vellekoop et al Ecological response to KndashPg boundary mass extinction 899
Jiao N Herndl G J Hansell D A Benner R Kattner G Wil-helm S W Kirchman D L Weinbauer M G Luo T ChenF and Azam F Microbial production of recalcitrant dissolvedorganic matter long-term carbon storage in the global oceanNat Rev 8 593ndash599 doi101038nrmicro2386 2010
Jorissen F J de Stigter H C and Widmark J G V A concep-tual model explaining benthic foraminiferal microhabitats MarMicropaleontol 26 3ndash15 doi1010160377-8398(95)00047-X1995
Jorissen F J Fontanier C and Thomas E Paleoceanographi-cal proxies based on deep-sea benthic foraminiferal assemblagecharacteristics in Proxies in Late Cenozoic PaleoceanographyPt 2 Biological tracers and biomarkers edited by Hillaire-Marcel C and de Vernal A 263ndash326 Elsevier Amsterdamthe Netherlands 2007
Kaiho K Kajiwara Y Tazaki K Ueshima M Takeda NKawahata H Arinobu T Ishiwatari R Hirai A and Lam-olda M A Oceanic primary productivity and dissolved oxygenlevels at the CretaceousTertiary boundary their decrease sub-sequent warming and recovery Paleoceanography 14 511ndash5241999
Keller G and Lindinger M Stable isotope TOC and CaCO3record across the CretaceousTertiary boundary at El KefTunisia Palaeogeogr Palaeocl 73 243ndash265 1989
Kellough G R Paleoecology of the Foraminiferida of the WillsPoint Formation (Midway Group) in northeast Texas Trans GulfCoast Assoc Geol Soc 15 73ndash153 1965
Kring D A The Chicxulub impact event and its environmentalconsequences at the CretaceousndashTertiary boundary PalaeogeogrPalaeocl 255 4ndash21 2007
Kump L R Interpreting carbon-isotope excursions Strangeloveoceans Geology 19 299ndash302 1991
Machalski M Vellekoop J Dubicka Z Peryt D and Ha-rasimiuk M Late Maastrichtian cephalopods dinoflagellatecysts and foraminifera from the CretaceousndashPaleogene suc-cession at Lechoacutewka southeast Poland Stratigraphic andenvironmental implications Cretaceous Res 57 208ndash227doi101016jcretres201508012 2016
Molina E Evidence and causes of the main extinctionevents in the Paleogene based on extinction and survivalpatterns of foraminifera Earth-Sci Rev 140 166ndash181doi101016jearscirev201411008 2015
Molina E Alegret L Arenillas I and Arz J The GlobalBoundary Stratotype Section and Point for the base of the Da-nian Stage (Paleocene Paleogeneldquo Tertiaryrdquo Cenozoic) at ElKef Tunisia-Original definition and revision Episodes 29 263ndash273 2006
Olsson R K Liu C and van Fossen M The Cretaceous-Tertiarycatastrophic event at Millers Ferry Alabama in The Cretaceous-Tertiary Event and Other Catastrophes in Earth History editedby Ryder G Fastovsky D and Gartner S Geol S Am S307 263ndash277 1996
Peryt D Benthic foraminiferal response to the Cenomanian-Turonian and CretaceousndashPaleogene boundary events PrzegladGeol 52 827ndash832 2004
Peryt D Alegret L and Molina E The CretaceousPalaeogene(KP) boundary at Aiumln Settara Tunisia restructuring of benthicforaminiferal assemblages Terra Nova 14 101ndash107 2002
Ramette A Multivariate analyses in microbial ecology FEMS Mi-crobiol Ecol 62 142ndash160 2007
Rodriacuteguez-Tovar F J Uchman A MrsquoHamdi A Riahi Sand Ismail-Lattrache K B Ichnological record of palaeoen-vironment from the CretaceousndashPaleogene boundary intervalat El Kef Tunisia The first study of old and new sec-tions at the stratotype area J Afr Earth Sci 120 23ndash30doi101016jjafrearsci201604017 2016
Saner S Mudurnu-Goumlynuumlk Havzasının Jura ve Sonrasıccediloumlkelimnitelikleriyle paleocografya yorumlaması TJK Buumllteni 23 39ndash52 1980
Schioslashler P Brinkhuis H Roncaglia L and Wilson G J Di-noflagellate biostratigraphy and sequence stratigraphy of theType Maastrichtian (Upper Cretaceous) ENCI Quarry TheNetherlands Mar Micropaleontol 31 65ndash95 1997
Schueth J D Bralower T J Jiang S and Patzkowsky ME The role of regional survivor incumbency in the evolu-tionary recovery of calcareous nannoplankton from the Creta-ceousPaleogene (KPg) mass extinction Paleobiology 41 661ndash679 doi101017pab201528 2015
Scotese C R A continental drift flipbook J Geology 112 729ndash741 2004
Scotese C R and Dreher C GlobalGeology available at httpwwwGlobalGeologycom (last access 11 March 2014) 2012
Sepkoski J J Patterns of Phanerozoic extinction a perspectivefrom global data bases in Global Events and Event Stratigra-phy edited by Walliser O H Springer Berlin Germany 35ndash52 2016
Sibert E C Hull P M and Norris R D Resilience of Pacificpelagic fish across the CretaceousPalaeogene mass extinctionNat Geosci 7 667ndash670 doi101038NGEO2227 2014
Slimani H and Toufiq A A CretaceousndashPaleogene boundary geo-logical site revealed by planktic foraminifera and dinoflagellatecysts at Ouled Haddou eastern external Rif Chain Morocco JAfr Earth Sci 88 38ndash52 doi101016jjafrearsci2013080082013
Slimani H Louwye S and Toufiq A Dinoflagellate cystsform the CretaceousndashPaleogene boundary at Ouled Had-dou southeastern Rif Morocco biostratigraphy paleoenvi-ronments and paleobiogeography Palynology 34 90ndash124doi10108001916121003629933 2010
Sluijs A and Brinkhuis H A dynamic climate and ecosystemstate during the Paleocene-Eocene Thermal Maximum infer-ences from dinoflagellate cyst assemblages on the New JerseyShelf Biogeosciences 6 1755ndash1781 doi105194bg-6-1755-2009 2009
Smit J Extinction and evolution of planktonic foraminifera after amajor impact at the CretaceousTertiary boundary Geol S AmS 190 329ndash352 1982
Smit J and Romein A J T A sequence of events across theCretaceous-Tertiary boundary Earth Planet Sci Lett 74 155ndash170 1985
Speijer R P Extinction and recovery patterns in ben-thic foraminiferal paleocommunities across the Creta-ceousPaleogene and PaleoceneEocene boundaries GeologicaUltraiectina 124 Faculteit Aardwetenschappen PhD ThesisUniversiteit Utrecht Utrecht the Netherlands 1994
Speijer R P and van Der Zwaan G J Extinction and survivor-ship of southern Tethyan benthic foraminifera across the Cre-
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
900 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
taceousPaleogene boundary in Biotic recovery from mass ex-tinction events edited by Hart M B Geolog Soc Spec PublLondon UK 102 343ndash371 1996
Stemmann L and Boss E Plankton and Particle Size andPackaging From Determining Optical Properties to Drivingthe Biological Pump Annu Rev Mar Sci 4 263ndash290doi101146annurev-marine-120710-100853 2012
Thomas E Late Cretaceous-early Eocene mass extinctions in thedeep sea Geol S Am S 247 481ndash495 1990
Vandenberghe N Hilgen F J and Speijer R P The PaleogenePeriod in The Geologic Time Scale 2012 edited by GradsteinF M Ogg J G Schmitz M D and Ogg G M 2 855ndash921Elsevier Boston USA 2012
Vellekoop J Sluijs A Smit J Schouten S Weijers J WH Sinninghe Damsteacute J S and Brinkhuis H Rapid short-term cooling following the Chicxulub impact at the CretaceousndashPaleogene boundary P Natl Acad Sci USA 111 7537ndash7541doi101073pnas1319253111 2014
Vellekoop J Smit J van de Schootbrugge B Weijers J W HGaleotti S Sinninghe Damsteacute J S and Brinkhuis H Palyno-logical evidence for prolonged cooling along the Tunisian conti-nental shelf following the KndashPg boundary impact PalaeogeogrPalaeocl 426 216ndash228 doi101016jpalaeo2015030212015
Vellekoop J Esmeray-Senlet S Miller K G Browning J VSluijs A van de Schootbrugge B Sinninghe Damsteacute J Sand Brinkhuis H Evidence for CretaceousndashPaleogene bound-ary bolide ldquoimpact winterrdquo conditions from New Jersey USAGeology 44 619ndash622 doi101130G379611 2016
Woelders L and Speijer R Stable seafloor conditions sealevel and food supply during the latest Maastrichtian atBrazos River Texas Mar Micropaleontol 121 41ndash51doi101016jmarmicro201510002 2015
Zachos J C Arthur M A and Dean W E Geochemical evi-dence for suppression of pelagic marine productivity at the Cre-taceousTertiary boundary Nature 337 61ndash64 1989
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
- Abstract
- Introduction
- Geological setting and age assessment
- Materials and methods
-
- Sampling
- Foraminiferal analysis
- Palynological analysis
- Statistical analysis
-
- Results
-
- Benthic foraminifera
- Palynology
-
- Discussion
-
- Tethyan benthic foraminiferal turnover sequence
- Ecological responses to reduced export productivity
-
- Okccedilular section
- Western Tethys
- Global responses
-
- Long-term recovery
-
- Conclusions
- Data availability
- Author contributions
- Competing interests
- Acknowledgements
- References
-

894 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
(Speijer and van der Zwaan 1996) As a collapse of the bi-ological pump should lead to enhanced remineralization oforganic matter in the photic zone it is indeed likely thatthe oxygen minimum zones expanded and shoaled leadingto oxygen stress on the sea floor Although early Paleocenedysoxia at El Kef is supported by the ostracod record (Donzeet al 1982) and ichnofacies analyses (Rodriguez-Tovaret al 2016) supporting (eg geochemical) evidence fordysoxia during the disaster phase at Okccedilular record is absentFurthermore an additional stressor for the benthic commu-nity during Zone P0 may be present In contrast to calcare-ous nanoplankton organic-walled cyst-producing dinoflagel-lates did not suffer extinctions and may have become a moreimportant component in the early Paleocene phytoplanktoncommunity Hence besides the reduction of the amount oforganic matter transported to the sea floor the compositionof food supplied by the photic zone likely changed signifi-cantly across the KndashPg boundary as well (DrsquoHondt 2005)possibly presenting an additional stress factor for the benthiccommunity (Speijer and van der Zwaan 1996 Alegret andThomas 2009)
522 Western Tethys
The patterns in benthic foraminiferal and dinoflagellate re-sponse to the KndashPg boundary perturbations at Okccedilular appearto be characteristic for the continental margins of the westernTethyan realm The benthic foraminiferal patterns are similaracross different size fractions suggesting that they are repre-sentative of the overall benthic communities In Tunisia andSpain where both benthic foraminiferal and dinoflagellaterecords are available the earliest Danian is also characterizedby a decrease in endobenthic foraminifera at the sea floor(Speijer and Van der Zwaan 1996 Peryt et al 2002 Ale-gret et al 2003) and simultaneously blooms of hexaperidin-ioids in the water column (Brinkhuis et al 1998 Vellekoopet al 2015 Fig 8) In the Ouled Haddou section Moroccoin the westernmost Tethys (Fig 1) the lowermost Danian isalso characterized by a strong increase of the hexaperidin-ioid Senegalinium group up to 30 of the assemblage (Sli-mani et al 2010) that is very similar to the Okccedilular record(Fig 8)
The KndashPg boundary dinocyst record of El Kef is strikinglysimilar to the one from the Okccedilular section showing two dis-tinct peaks in hexaperidinioids one in Zone P0 and one atthe basal part of Zone Pα At El Kef the initial post-impactdominance of hexaperidinioids is however less pronounced(Fig 8) This difference in expression might be related tosmall differences in paleogeographic and paleoceanographicsettings between sites
In addition some high-resolution benthic KndashPg records inthe Tethyan realm show a short-lived bloom of specific en-dobenthic foraminiferal taxa in the few centimeter directlyabove the KndashPg boundary sometimes accompanied by otherindicators of low oxygen levels (eg Coccioni and Gale-
otti 1994 Speijer and Van der Zwaan 1996 Kaiho et al1999) This suggests that at least locally the sea floor wastemporarily characterized by hypoxic conditions directly fol-lowing the impact This short-lived deoxygenation is possi-bly related to the mass mortality at the KndashPg boundary re-sulting in a large short-lived flux of organic matter to thesea floor (Coccioni and Galeotti 1994) However since ex-port productivity was greatly reduced after the KndashPg bound-ary mass extinction very little ldquonewrdquo organic matter reachedthe sea floor after the initial post-impact influx Hence afteraccumulated organic matter was remineralized the benthiccommunity starved resulting in a transition to an epibenthic-dominated benthic fauna similar to the post-impact fauna ofthe Okccedilular record
The combined dinocyst records suggest that the early Pa-leocene margins of the Tethys were characterized by an in-crease in nutrient availability in the photic zone whereascoeval benthic foraminiferal records indicate a major de-crease in nutrient supply to the sea floor These changesin the benthic foraminiferal and dinocyst communities arelikely caused by the major reduction of both the efficiencyand strength of the biological pump (ie the fraction ofprimary production exported from the photic zone and theamount of organic matter transported down) in accordancewith the Living Ocean model (DrsquoHondt and Zachos 1998DrsquoHondt et al 1998 DrsquoHondt 2005 Coxall et al 2006)For a detailed explanation of the difference between biologi-cal pump efficiency and biological pump strength see Hiltinget al (2008)
As bulk carbonate δ13C reflects the isotopic compositionof the surface ocean which is set by burial fractions and bythe photosynthetic isotope effect (Kump 1991 Hain et al2014) the excursion recorded in bulk δ13C records world-wide and the rapid collapse in surface to deep-ocean carbonisotope gradients likely reflect the reduction of the global in-tensity (ie efficiency) of the biological pump (Hain et al2014) The recorded inverse change in nutrient availabilitysuggests a causal link A reduced efficiency of the biologi-cal pump and associated decrease of the fraction of biomasstransported out of the photic zone could have resulted inhigh rates of nutrient recycling in the upper part of the wa-ter column (DrsquoHondt 2005) This suggests that the reduc-tion of the biological pump strength recorded by the benthicforaminiferal record is a consequence of the decreased effi-ciency of the biological pump recorded by the carbon iso-tope and dinoflagellate cyst records The strong correlationbetween the bulk carbon isotope curves and the Shannondiversity index (H ) of the benthic foraminiferal records atboth Okccedilular (R2
= 073 p lt 0001) and El Kef (R2= 074
p lt 0001) suggest a clear link between the changes in ben-thic foraminiferal assemblages and the collapse and recoveryof biological pump efficiency
As a result of the reduced biological pump efficiency morenutrients remained available for the early Paleocene phyto-plankton community At Tethyan margin sites this is indi-
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
J Vellekoop et al Ecological response to KndashPg boundary mass extinction 895
Figure 8 The benthic foraminiferal (gt 125 microm) dinocyst and bulk stable carbon isotope records of Okccedilular (Turkey) and El Kef (Tunisia)the dinocyst record of Ouled Haddou (Morocco) and the benthic foraminiferal record (gt 63 microm) of ODP Hole 1049C Blake Nose (NorthAtlantic) Note that the definition of ldquohexaperidinioidsrdquo used is provided in the text The four phases identified based on the bulk stablecarbon isotopes and the Shannon diversity index (H ) of the benthic foraminiferal records are indicated (IndashIV) The gt 63 microm size fractionbenthic foraminiferal record of ODP Hole 1049C is from Alegret and Thomas (2004) who argued that the lowermost Paleocene of thissite comprised reworked foraminifera This interval is indicated by a dashed bar in the figure The biostratigraphy of Ouled Haddou is fromSlimani and Toufiq (2013) whereas the dinocyst record of this site is from Slimani et al (2010) The biostratigraphy of El Kef is based onBrinkhuis et al (1998) Speijer and van der Zwaan (1996) and Molina et al (2006) The fine-fraction stable carbon isotope record (blackdots) of El Kef is from Keller and Lindinger (1989) The Shannon diversity index (red dots) relative abundances of bi-triserial benthic taxa(dark purple) and all endobenthic foraminifera (light purple) are based on the gt 125 microm size fraction foraminiferal data of Speijer and vander Zwaan (1996) The palynological data of El Kef are from Brinkhuis et al (1998) (lower part) and Guasti et al (2005) (upper part) TheShannon diversity index of the gt 125 microm size fraction benthic foraminiferal record (red dots) the relative contribution of bi-triserial benthictaxa (dark purple) all endobenthic foraminifera (light purple) and hexaperidinioid dinocysts from Okccedilular are from this study The bulkstable carbon isotope record of Okccedilular (black dots) is from Accedilikalin et al (2015) Mbsf indicates meters below sea floor
cated by the higher abundance of hexaperidinioids Similarlyblooms of eutrophic survivor taxa of calcareous nanoplank-ton at open ocean sites have been suggested to be relatedto the buildup of nutrients in the open ocean photic zone(Schueth et al 2015) Although the carbon isotope gradientsbetween surface and deep waters indicate that the recoveryof the biological pump took hundreds of thousands of years(Zachos et al 1989 Kump 1991 DrsquoHondt et al 1998 Cox-all et al 2006 Birch et al 2016) our records suggest thatthe enhanced recycling of nutrients in the Tethys was partic-ularly intense during two phases in the first tens of thousandsyears after the impact (Fig 8)
523 Global responses
The decrease in export productivity following the KndashPgboundary is not only recognized in the Tethyan realm In theAtlantic Ocean Southern Ocean and the Indian Ocean a de-crease of the biological pump strength after the KndashPg bound-ary was also observed in neritic to abyssal environments(eg Thomas 1990 Olsson et al 1996 Hull et al 2011Alegret et al 2012) As an example the gt 63 microm size frac-tion benthic foraminiferal KndashPg boundary record of BlakeNose (ODP Hole 1049C northwestern Atlantic Alegret andThomas 2004) shows a pattern that is strikingly similar tothe gt 125 microm size fraction records in the Tethyan realm witha strong decrease of endobenthics in Zone P0 followed by a
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
896 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
gradual recovery across zones Pα and P1a (Fig 8) The simi-larities between the benthic foraminiferal patterns of Tethyanouter neritic to middle bathyal (200ndash1500 m) sites and thoseof the middle to lower bathyal (1500ndash1600 m) record ofBlake Nose (Alegret and Thomas 2004) suggest that theserecords register a global decrease in export productivity fol-lowing the KndashPg boundary
However several open ocean sites mostly in the Pacificshow an opposite trend with several lines of evidence sug-gesting an increased biological pump strength at these sites(eg Hull et al 2011 Alegret et al 2012) This led Ale-gret et al (2012) to conclude that the decrease in biologicalpump strength after the KndashPg boundary was most likely aregional instead of a global effect arguing against the Liv-ing Ocean hypothesis We would argue that low-productivityopen ocean sites like the central Pacific are characterized byvery different ecosystem structures and higher productivityas those along the Tethyan margins (eg Dortch and Pack-erd 1989) Therefore the consequence of ecosystem reorga-nization resulting from the mass extinction was likely verydifferent in the open ocean Pacific (eg Sibert et al 2014)Esmeray-Senlet et al (2015) proposed the term ldquoHeteroge-neous Oceanrdquo for this conceptual model as an alternativeto the Living Ocean model Supposedly the HeterogeneousOcean was characterized by a strong geographic heterogene-ity in the extinction patterns and food supply to the sea floor
However as the primary food source for open ocean deep-sea benthic communities generally consists of refractory or-ganic matter (eg Jiao et al 2010) it remains uncertainwhether changes in the intensity of recycling of the gener-ally more labile organic matter in the surface ocean will re-sult in changes in the amount of organic matter that benthiccommunities receive as they will likely continue to receiverecalcitrant organic matter at barely diminished rates despitethe collapse of the biological pump
In addition it should be noted that although the bio-logical pump efficiency is generally relatively high in openocean realms such as the central Pacific the biological pumpstrength is generally very low at these sites (Honjo et al2008 Henson et al 2011) Small changes in absolute bio-logical pump strength at such a locality could therefore havehad a large effect on the local oligotrophic benthic commu-nity (Alegret and Thomas 2009) but they likely representedonly a small fraction of the net amount of organic matterglobally transported to the deep Hence even though localbiological pump strength might have increased at some low-productivity deep-sea sites the large decrease in biologi-cal pump strength recorded at many other sites means thatthe net amount of organic matter globally transported out ofthe surface ocean was likely still reduced in the post-impactworld (Birch et al 2016) Therefore we argue that such Pa-cific records do not represent the general pattern The globalocean response can be characterized as heterogeneous how-ever the KndashPg boundary mass extinction did lead to a strongreduction of the net amount of organic matter globally trans-
ported from the surface to the sea floor while biological pro-ductivity recovered rapidly after the extinction event in ac-cordance with the Living Ocean hypothesis
53 Long-term recovery
Following the disaster phase the hexaperidinioid cysts atOkccedilular and El Kef decreased in abundance and the abun-dance of endobenthic forms increased again as the recoveryof the benthic community was initiated This recovery phaseis approximately correlative to Zone Pα and the basal part ofZone P1a which according to the Paleogene age constraintsof Vandenberghe et al (2012) represents at least 300 kyrThis duration is in agreement with the estimations providedby Birch et al (2016) During this phase the carbon iso-tope records remain well below pre-impact values and thediversity of the benthic foraminiferal community has not yetfully recovered (Figs 4ndash8) This indicates that the rapid andshort-lived KndashPg boundary disaster was followed by a rela-tively long recovery phase in line with previous estimatesof a multimillion-year biological recovery (eg Coxall etal 2006) While the impact-related environmental perturba-tions were short lived (Kring 2007 Vellekoop et al 20142016) the extinctions amongst important biological groupsled to a reduction of the organic flux from the photic zone todeep water resulting in major long-term biological and pa-leoceanographic reorganizations Only with the evolutionaryrecovery of the pelagic community governing the biologicalcarbon pump did export productivity start to increase again(eg Coxall et al 2006 Birch et al 2016)
6 Conclusions
The marine palynological benthic foraminiferal and bulkstable isotope records of the Okccedilular and El Kef sections re-veal major changes across the KndashPg boundary interval por-traying the biological crisis at the KndashPg boundary and subse-quent recovery in the early Paleocene Based on the succes-sion of benthic faunal assemblages at a number of Tethyanmargin sites four phases can be recognized across the KndashPgboundary interval the Maastrichtian or pre-impact phase (I)a disaster phase (II) a recovery phase (III) and an early Pa-leocene post-recovery phase (IV)
Following the KndashPg boundary impact some localitiesshow a large short-lived flux of food to the sea floor likelyrelated to the mass mortality at the KndashPg boundary Howeversince export productivity was greatly reduced after the KndashPgboundary mass extinction it is likely that after the initialpost-impact influx very little fresh organic matter reachedthe sea floor This reduction of export productivity in thepost-extinction disaster phase eventually resulted in a lowerfood supply to the sea floor This presented a major stressfactor for benthic organisms in the western Tethys The re-duced food supply resulted in an abrupt impoverishment of
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
J Vellekoop et al Ecological response to KndashPg boundary mass extinction 897
benthic communities As the downward transport of nutri-ents was slowed down recycling in upper layers increasedAs a result more nutrients remained available for the earlyPaleocene phytoplankton community leading to blooms ofdinoflagellates along the Tethyan margins and blooms of cal-careous nanoplankton taxa in more open ocean sites Ourrecords show that the enhanced recycling of nutrients in theTethys was particularly intense in the tens of thousands ofyears after the impact Following this the slow evolution-ary recovery of the pelagic community governing the bio-logical carbon pump resulted in a gradually increasing ex-port productivity in the hundreds of thousands of years afterthe impact Hence our integration of dinocyst and benthicforaminiferal records across the KndashPg boundary provides im-portant new insights into the ecological responses to the re-duction of export productivity following the mass extinctionat the CretaceousndashPaleogene boundary highlighting the di-rect ecological consequences of the Living Ocean conditionsin the post-impact world
7 Data availability
All data related to publication are available in the Supple-ment
The Supplement related to this article is available onlineat doi105194bg-14-885-2017-supplement
Author contributions Johan Vellekoop Lineke Woelders andHenk Brinkhuis designed the research Samples were collectedin the field by Johan Vellekoop Sanem Accedilikalin Ismail Ouml Yil-maz and Jan Smit Palynological analyses were carried out by Jo-han Vellekoop Benthic foraminiferal analyses were carried out byLineke Woelders and Robert P Speijer Johan Vellekoop LinekeWoelders and Bas van de Schootbrugge prepared the manuscriptwith input from all authors
Competing interests The authors declare that they have no conflictof interest
Acknowledgements This work was supported by the NetherlandsOrganization for Scientific Research (NWO) Open CompetitionGrant ALWPJ09047 to H Brinkhuis and the Research FoundationFlanders (FWO) grant G0B8513 to R P Speijer We thank LKump F Boscolo Galazzo P Roopnarine and one anonymousreferee for their suggestions
Edited by D GillikinReviewed by L Kump F Boscolo Galazzo and P Roopnarine
References
Accedilikalin S Vellekoop J Ocakoglu F Yılmaz I Ouml Smit J Al-tiner S O Goderis S Vonhof H Speijer R P WoeldersL Fornaciari E and Brinkhuis H Geochemical and pale-ontological characterization of a new KndashPg Boundary local-ity from the Northern branch of the Neo-Tethys MudurnundashGoumlynuumlk Basin NW Turkey Cretaceous Res 52 251ndash267doi101016jcretres201407011 2015
Alegret L and Thomas E Food supply to the seafloor in the Pa-cific Ocean after the CretaceousPaleogene boundary event MarMicropaleontol 73 105ndash116 2009
Alegret L Molina E and Thomas E Benthic foraminiferalturnover across the CretaceousPaleogene boundary at Agost(southeastern Spain) paleoenvironmental inferences Mar Mi-cropaleontol 48 251ndash279 2003
Alegret L Thomas E and Lohmann K C End-Cretaceous ma-rine mass extinction not caused by productivity collapse P NatlAcad Sci USA 109 728ndash732 doi101073pnas11106011092012
Alegret L Rodriacuteguez-Tovar F J and Uchman A How bioturba-tion obscured the CretaceousndashPaleogene boundary record TerraNova 27 225ndash230 doi101111ter12151 2015
Altiner D Koccedilyigit A Farinacci A Nicosia U and ContiM A Jurassic-Lower Cretaceous stratigraphy and paleogeo-graphic evolution of the southern part of north-western Anatolia(Turkey) Geologica Romana 27 13ndash80 1991
Alvarez LW Alvarez W Asaro F and Michel H V Extrater-restrial cause for the Cretaceous-Tertiary extinction Science208 1095ndash1108 1980
Alegret L and Thomas E Benthic foraminifera and environmen-tal turnover across the CretaceousPaleogene boundary at BlakeNose (ODP Hole 1049C Northwestern Atlantic) PalaeogeogrPalaeocl 208 59ndash83 doi101016jpalaeo200402028 2004
Armstrong R A Lee C Hedges J I Honjo S and WakehamS G A new mechanistic model for organic carbon fluxes in theocean based on the quantitative association of POC with ballastmaterials Deep-Sea Res Pt II 49 219ndash236 2002
Aubert J and Berggren W A Paleocene benthic foraminiferalbiostratigraphy and paleoecology of Tunisia Bull du Cent deRech de Pau ndash SNPA 10 379ndash469 1976
Berggren W A and Aubert J Paleocene benthonic foraminiferalbiostratigraphy paleobiogeography and paleoecology ofAtlanticndashTethyan regions midway-type fauna PalaeogeogrPalaeocl 18 73ndash192 1975
Bernaola G and Monechi S Calcareous nannofossil ex-tinction and survivorship across the CretaceousndashPaleogeneboundary at Walvis Ridge (ODP Hole 1262C SouthAtlantic Ocean) Palaeogeogr Palaeocl 255 132ndash156doi101016jpalaeo200702045 2007
Bernhard J M Characteristic assemblages and morphologies ofbenthic foraminifera from anoxic organic-rich deposits Jurassicthrough Holocene J Foramin Res 16 207ndash215 1986
Birch H S Coxall H K Pearson P N Kroon D andSchmidt D N Partial collapse of the marine carbon pump af-ter the CretaceousndashPaleogene boundary Geology 44 287ndash290doi101130G375811 2016
Brinkhuis H and Zachariasse W J Dinoflagellate cysts sealevel changes and planktonic foraminifers across the Cretaceous-
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
898 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
Tertiary boundary at El Haria northwest Tunisia Mar Micropa-leontol 13 153ndash191 1988
Brinkhuis H Bujak J P Smit J Versteegh G J M and Viss-cher H Dinoflagellate-based sea surface temperature recon-structions across the Cretaceous-Tertiary boundary PalaeogeogrPalaeocl 141 67ndash83 1998
Buesseler K O The decoupling of production and particulate ex-port in the surface ocean Global Biogeochem Cy 12 297ndash3101998
Buzas M A Culver S J and Jorissen F J A statisti-cal evaluation of the microhabitats of living (stained) infau-nal benthic foraminifera Mar Micropaleontol 20 311ndash320doi1010160377-8398(93)90040-5 1993
Coccioni R and Galeotti S KndashT boundary extinction geolog-ically instantaneous or gradual event Evidence from deep-seabenthic foraminifera Geology 22 779ndash782 1994
Corliss B H Microhabitats of benthic foraminifera within deep-sea sediments Nature 314 435ndash438 doi101038314435a01985
Corliss B H and Chen C Morphotype patterns of Nor-wegian Sea deep-sea benthic foraminifera and ecologi-cal implications Geology 16 716ndash719 doi1011300091-7613(1988)016lt0716MPONSDgt23CO2 1988
Coxall H K DrsquoHondt S and Zachos J C Pelagic evolution andenvironmental recovery after the CretaceousndashPaleogene mass ex-tinction Geology 34 297ndash300 doi101130G217021 2006
Culver S J Benthic foraminifera across the CretaceousndashTertiary(KndashT) boundary a review Mar Micropaleontol 47 177ndash2262003
Cushman J A Upper Cretaceous foraminifera of the Gulf coastalregion of the United States and adjacent areas US Geolog SurvProf Pap 206 241 pp 1946
Cushman J A Paleocene foraminifera of the Gulf Coastal Regionof the United States and adjacent regions US Geol Surv ProfPap 232 75 pp 1951
DrsquoHondt S Consequences of the CretaceousPaleogene Mass Ex-tinction for Marine Ecosystems Annu Rev Ecol Evol S36 295ndash317 doi101146annurevecolsys350211031057152005
DrsquoHondt S and Zachos J C Cretaceous foraminifera and the evo-lutionary history of planktic photosymbiosis Paleobiology 24512ndash523 1998
DrsquoHondt S Donaghay P Zachos J C Luttenberg D andLindinger M Organic Carbon Fluxes and Ecological Recov-ery from the Cretaceous-Tertiary Mass Extinction Science 282276ndash279 1998
Donze P Colin J-P Damotte R Oertli HJ Peypouquet J-Pand Said R Les ostracodes du Campanien terminal agrave lrsquoEocegraveneinfeacuterieur de la coupe du Kef Tuneacutesie nord-occidentale Bull desCentr Recher Expl-Prod Elf Aquit 6 273ndash335 1982
Dortch Q and Packard T T Differences in biomass structure be-tween oligotrophic and eutrophic marine ecosystems Deep-SeaRes 36 223ndash240 1989
Ernst S R Guasti E Dupuis C and Speijer R P En-vironmental perturbation in the southern Tethys across thePaleoceneEocene boundary (Dababiya Egypt) Foraminiferaland clay mineral records Mar Micropaleontol 60 89ndash111doi101016jmarmicro200603002 2006
Eshet Y Moshkovitz S Habib D Benjamini C and Maga-ritz M Calcareous nannofossil and dinoflagellate stratigraphyacross the CretaceousTertiary boundary at Hor Hahar IsraelMar Micropaleontol 18 199ndash228 1992
Eshet Y Almogi-Labin A and Bein A Dinoflagellate cysts pa-leoproductivity and upwelling systems A Late Cretaceous ex-ample from Israel Mar Micropaleontol 23 231ndash240 1994
Esmeray-Senlet S Wright J D Olsson R K MillerK G Browning J V and Quan T M Evidencefor reduced export productivity following the Creta-ceousPaleogene mass extinction Paleoceanography 30718ndash738 doi1010022014PA002724 2015
Giusberti L Boscolo Galazzo F and Thomas E Variability inclimate and productivity during the Paleocene-Eocene ThermalMaximum in the western Tethys (Forada section) Clim Past12 213ndash240 doi105194cp-12-213-2016 2016
Guasti E Kouwenhoven T J Brinkhuis H and Speijer R PPaleocene sea-level and productivity changes at the southernTethyan margin (El Kef Tunisia) Mar Micropaleontol 55 1ndash17 2005
Habib D and Saeedi F The Manumiella seelandica global spikeCooling during regression at the close of the MaastrichtianPalaeogeogr Palaeocl 255 87ndash97 2007
Hain M P Sigmal D M and Haug G H 818 ndash The biolog-ical Pump in the Past in Reference Module in Earth Systemsand Environmental Sciences Treatise on Geochemistry (SecondEdition) The Oceans and Marine Geochemistry 8 485ndash517doi101016B978-0-08-095975-700618-5 2014
Hayek L C and Buzas M A On the proper and efficient useof diversity measures with individual field samples J ForaminRes 43 305ndash313 2013
Henson S A Sanders R Madsen E Morris P J Le MoigneF and Quartly G D A reduced estimate of the strength ofthe oceanrsquos biological carbon pump Geophys Res Lett 38L04606 doi1010292011GL046735 2011
Hilting A K Kump L R and Bralower T J Variations in theoceanic vertical carbon isotope gradient and their implicationsfor the Paleocene-Eocene biological pump Paleoceanography23 PA3222 doi1010292007PA001458 2008
Honjo S Manganini S J Krishfield R A and Francois RParticulate organic carbon fluxes to the ocean interior and fac-tors controlling the biological pump A synthesis of global sed-iment trap progams since 1983 Prog Oceanogr 76 217ndash285doi101016jpocean200711003 2008
Huber B T MacLeod K G and Norris R D Abrupt extinctionand subsequent reworking of Cretaceous planktonic foraminiferaacross the Cretaceous-Tertiary boundary Evidence from the sub-tropical North Atlantic in Catastrophic Events and Mass Extinc-tions Impacts and Beyond edited by Koeberl C and MacLeodK G Geol S Am S 356 277ndash289 Boulder Colorado USA2002
Hull P M Norris R D Bralower T J and Schueth J D A rolefor chance in marine recovery from the end-Cretaceous extinc-tion Nat Geosci 4 856ndash860 doi101038ngeo1302 2011
Hsuuml K J and McKenzie J A A ldquoStrangeloverdquo ocean in the earli-est Tertiary in Natural variations Archean to present edited bySundquist E T and Broecker W S The carbon cycle and atmo-spheric CO2 American Geophysical Union Washington DCUSA 487ndash492 1985
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
J Vellekoop et al Ecological response to KndashPg boundary mass extinction 899
Jiao N Herndl G J Hansell D A Benner R Kattner G Wil-helm S W Kirchman D L Weinbauer M G Luo T ChenF and Azam F Microbial production of recalcitrant dissolvedorganic matter long-term carbon storage in the global oceanNat Rev 8 593ndash599 doi101038nrmicro2386 2010
Jorissen F J de Stigter H C and Widmark J G V A concep-tual model explaining benthic foraminiferal microhabitats MarMicropaleontol 26 3ndash15 doi1010160377-8398(95)00047-X1995
Jorissen F J Fontanier C and Thomas E Paleoceanographi-cal proxies based on deep-sea benthic foraminiferal assemblagecharacteristics in Proxies in Late Cenozoic PaleoceanographyPt 2 Biological tracers and biomarkers edited by Hillaire-Marcel C and de Vernal A 263ndash326 Elsevier Amsterdamthe Netherlands 2007
Kaiho K Kajiwara Y Tazaki K Ueshima M Takeda NKawahata H Arinobu T Ishiwatari R Hirai A and Lam-olda M A Oceanic primary productivity and dissolved oxygenlevels at the CretaceousTertiary boundary their decrease sub-sequent warming and recovery Paleoceanography 14 511ndash5241999
Keller G and Lindinger M Stable isotope TOC and CaCO3record across the CretaceousTertiary boundary at El KefTunisia Palaeogeogr Palaeocl 73 243ndash265 1989
Kellough G R Paleoecology of the Foraminiferida of the WillsPoint Formation (Midway Group) in northeast Texas Trans GulfCoast Assoc Geol Soc 15 73ndash153 1965
Kring D A The Chicxulub impact event and its environmentalconsequences at the CretaceousndashTertiary boundary PalaeogeogrPalaeocl 255 4ndash21 2007
Kump L R Interpreting carbon-isotope excursions Strangeloveoceans Geology 19 299ndash302 1991
Machalski M Vellekoop J Dubicka Z Peryt D and Ha-rasimiuk M Late Maastrichtian cephalopods dinoflagellatecysts and foraminifera from the CretaceousndashPaleogene suc-cession at Lechoacutewka southeast Poland Stratigraphic andenvironmental implications Cretaceous Res 57 208ndash227doi101016jcretres201508012 2016
Molina E Evidence and causes of the main extinctionevents in the Paleogene based on extinction and survivalpatterns of foraminifera Earth-Sci Rev 140 166ndash181doi101016jearscirev201411008 2015
Molina E Alegret L Arenillas I and Arz J The GlobalBoundary Stratotype Section and Point for the base of the Da-nian Stage (Paleocene Paleogeneldquo Tertiaryrdquo Cenozoic) at ElKef Tunisia-Original definition and revision Episodes 29 263ndash273 2006
Olsson R K Liu C and van Fossen M The Cretaceous-Tertiarycatastrophic event at Millers Ferry Alabama in The Cretaceous-Tertiary Event and Other Catastrophes in Earth History editedby Ryder G Fastovsky D and Gartner S Geol S Am S307 263ndash277 1996
Peryt D Benthic foraminiferal response to the Cenomanian-Turonian and CretaceousndashPaleogene boundary events PrzegladGeol 52 827ndash832 2004
Peryt D Alegret L and Molina E The CretaceousPalaeogene(KP) boundary at Aiumln Settara Tunisia restructuring of benthicforaminiferal assemblages Terra Nova 14 101ndash107 2002
Ramette A Multivariate analyses in microbial ecology FEMS Mi-crobiol Ecol 62 142ndash160 2007
Rodriacuteguez-Tovar F J Uchman A MrsquoHamdi A Riahi Sand Ismail-Lattrache K B Ichnological record of palaeoen-vironment from the CretaceousndashPaleogene boundary intervalat El Kef Tunisia The first study of old and new sec-tions at the stratotype area J Afr Earth Sci 120 23ndash30doi101016jjafrearsci201604017 2016
Saner S Mudurnu-Goumlynuumlk Havzasının Jura ve Sonrasıccediloumlkelimnitelikleriyle paleocografya yorumlaması TJK Buumllteni 23 39ndash52 1980
Schioslashler P Brinkhuis H Roncaglia L and Wilson G J Di-noflagellate biostratigraphy and sequence stratigraphy of theType Maastrichtian (Upper Cretaceous) ENCI Quarry TheNetherlands Mar Micropaleontol 31 65ndash95 1997
Schueth J D Bralower T J Jiang S and Patzkowsky ME The role of regional survivor incumbency in the evolu-tionary recovery of calcareous nannoplankton from the Creta-ceousPaleogene (KPg) mass extinction Paleobiology 41 661ndash679 doi101017pab201528 2015
Scotese C R A continental drift flipbook J Geology 112 729ndash741 2004
Scotese C R and Dreher C GlobalGeology available at httpwwwGlobalGeologycom (last access 11 March 2014) 2012
Sepkoski J J Patterns of Phanerozoic extinction a perspectivefrom global data bases in Global Events and Event Stratigra-phy edited by Walliser O H Springer Berlin Germany 35ndash52 2016
Sibert E C Hull P M and Norris R D Resilience of Pacificpelagic fish across the CretaceousPalaeogene mass extinctionNat Geosci 7 667ndash670 doi101038NGEO2227 2014
Slimani H and Toufiq A A CretaceousndashPaleogene boundary geo-logical site revealed by planktic foraminifera and dinoflagellatecysts at Ouled Haddou eastern external Rif Chain Morocco JAfr Earth Sci 88 38ndash52 doi101016jjafrearsci2013080082013
Slimani H Louwye S and Toufiq A Dinoflagellate cystsform the CretaceousndashPaleogene boundary at Ouled Had-dou southeastern Rif Morocco biostratigraphy paleoenvi-ronments and paleobiogeography Palynology 34 90ndash124doi10108001916121003629933 2010
Sluijs A and Brinkhuis H A dynamic climate and ecosystemstate during the Paleocene-Eocene Thermal Maximum infer-ences from dinoflagellate cyst assemblages on the New JerseyShelf Biogeosciences 6 1755ndash1781 doi105194bg-6-1755-2009 2009
Smit J Extinction and evolution of planktonic foraminifera after amajor impact at the CretaceousTertiary boundary Geol S AmS 190 329ndash352 1982
Smit J and Romein A J T A sequence of events across theCretaceous-Tertiary boundary Earth Planet Sci Lett 74 155ndash170 1985
Speijer R P Extinction and recovery patterns in ben-thic foraminiferal paleocommunities across the Creta-ceousPaleogene and PaleoceneEocene boundaries GeologicaUltraiectina 124 Faculteit Aardwetenschappen PhD ThesisUniversiteit Utrecht Utrecht the Netherlands 1994
Speijer R P and van Der Zwaan G J Extinction and survivor-ship of southern Tethyan benthic foraminifera across the Cre-
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
900 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
taceousPaleogene boundary in Biotic recovery from mass ex-tinction events edited by Hart M B Geolog Soc Spec PublLondon UK 102 343ndash371 1996
Stemmann L and Boss E Plankton and Particle Size andPackaging From Determining Optical Properties to Drivingthe Biological Pump Annu Rev Mar Sci 4 263ndash290doi101146annurev-marine-120710-100853 2012
Thomas E Late Cretaceous-early Eocene mass extinctions in thedeep sea Geol S Am S 247 481ndash495 1990
Vandenberghe N Hilgen F J and Speijer R P The PaleogenePeriod in The Geologic Time Scale 2012 edited by GradsteinF M Ogg J G Schmitz M D and Ogg G M 2 855ndash921Elsevier Boston USA 2012
Vellekoop J Sluijs A Smit J Schouten S Weijers J WH Sinninghe Damsteacute J S and Brinkhuis H Rapid short-term cooling following the Chicxulub impact at the CretaceousndashPaleogene boundary P Natl Acad Sci USA 111 7537ndash7541doi101073pnas1319253111 2014
Vellekoop J Smit J van de Schootbrugge B Weijers J W HGaleotti S Sinninghe Damsteacute J S and Brinkhuis H Palyno-logical evidence for prolonged cooling along the Tunisian conti-nental shelf following the KndashPg boundary impact PalaeogeogrPalaeocl 426 216ndash228 doi101016jpalaeo2015030212015
Vellekoop J Esmeray-Senlet S Miller K G Browning J VSluijs A van de Schootbrugge B Sinninghe Damsteacute J Sand Brinkhuis H Evidence for CretaceousndashPaleogene bound-ary bolide ldquoimpact winterrdquo conditions from New Jersey USAGeology 44 619ndash622 doi101130G379611 2016
Woelders L and Speijer R Stable seafloor conditions sealevel and food supply during the latest Maastrichtian atBrazos River Texas Mar Micropaleontol 121 41ndash51doi101016jmarmicro201510002 2015
Zachos J C Arthur M A and Dean W E Geochemical evi-dence for suppression of pelagic marine productivity at the Cre-taceousTertiary boundary Nature 337 61ndash64 1989
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
- Abstract
- Introduction
- Geological setting and age assessment
- Materials and methods
-
- Sampling
- Foraminiferal analysis
- Palynological analysis
- Statistical analysis
-
- Results
-
- Benthic foraminifera
- Palynology
-
- Discussion
-
- Tethyan benthic foraminiferal turnover sequence
- Ecological responses to reduced export productivity
-
- Okccedilular section
- Western Tethys
- Global responses
-
- Long-term recovery
-
- Conclusions
- Data availability
- Author contributions
- Competing interests
- Acknowledgements
- References
-

J Vellekoop et al Ecological response to KndashPg boundary mass extinction 895
Figure 8 The benthic foraminiferal (gt 125 microm) dinocyst and bulk stable carbon isotope records of Okccedilular (Turkey) and El Kef (Tunisia)the dinocyst record of Ouled Haddou (Morocco) and the benthic foraminiferal record (gt 63 microm) of ODP Hole 1049C Blake Nose (NorthAtlantic) Note that the definition of ldquohexaperidinioidsrdquo used is provided in the text The four phases identified based on the bulk stablecarbon isotopes and the Shannon diversity index (H ) of the benthic foraminiferal records are indicated (IndashIV) The gt 63 microm size fractionbenthic foraminiferal record of ODP Hole 1049C is from Alegret and Thomas (2004) who argued that the lowermost Paleocene of thissite comprised reworked foraminifera This interval is indicated by a dashed bar in the figure The biostratigraphy of Ouled Haddou is fromSlimani and Toufiq (2013) whereas the dinocyst record of this site is from Slimani et al (2010) The biostratigraphy of El Kef is based onBrinkhuis et al (1998) Speijer and van der Zwaan (1996) and Molina et al (2006) The fine-fraction stable carbon isotope record (blackdots) of El Kef is from Keller and Lindinger (1989) The Shannon diversity index (red dots) relative abundances of bi-triserial benthic taxa(dark purple) and all endobenthic foraminifera (light purple) are based on the gt 125 microm size fraction foraminiferal data of Speijer and vander Zwaan (1996) The palynological data of El Kef are from Brinkhuis et al (1998) (lower part) and Guasti et al (2005) (upper part) TheShannon diversity index of the gt 125 microm size fraction benthic foraminiferal record (red dots) the relative contribution of bi-triserial benthictaxa (dark purple) all endobenthic foraminifera (light purple) and hexaperidinioid dinocysts from Okccedilular are from this study The bulkstable carbon isotope record of Okccedilular (black dots) is from Accedilikalin et al (2015) Mbsf indicates meters below sea floor
cated by the higher abundance of hexaperidinioids Similarlyblooms of eutrophic survivor taxa of calcareous nanoplank-ton at open ocean sites have been suggested to be relatedto the buildup of nutrients in the open ocean photic zone(Schueth et al 2015) Although the carbon isotope gradientsbetween surface and deep waters indicate that the recoveryof the biological pump took hundreds of thousands of years(Zachos et al 1989 Kump 1991 DrsquoHondt et al 1998 Cox-all et al 2006 Birch et al 2016) our records suggest thatthe enhanced recycling of nutrients in the Tethys was partic-ularly intense during two phases in the first tens of thousandsyears after the impact (Fig 8)
523 Global responses
The decrease in export productivity following the KndashPgboundary is not only recognized in the Tethyan realm In theAtlantic Ocean Southern Ocean and the Indian Ocean a de-crease of the biological pump strength after the KndashPg bound-ary was also observed in neritic to abyssal environments(eg Thomas 1990 Olsson et al 1996 Hull et al 2011Alegret et al 2012) As an example the gt 63 microm size frac-tion benthic foraminiferal KndashPg boundary record of BlakeNose (ODP Hole 1049C northwestern Atlantic Alegret andThomas 2004) shows a pattern that is strikingly similar tothe gt 125 microm size fraction records in the Tethyan realm witha strong decrease of endobenthics in Zone P0 followed by a
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
896 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
gradual recovery across zones Pα and P1a (Fig 8) The simi-larities between the benthic foraminiferal patterns of Tethyanouter neritic to middle bathyal (200ndash1500 m) sites and thoseof the middle to lower bathyal (1500ndash1600 m) record ofBlake Nose (Alegret and Thomas 2004) suggest that theserecords register a global decrease in export productivity fol-lowing the KndashPg boundary
However several open ocean sites mostly in the Pacificshow an opposite trend with several lines of evidence sug-gesting an increased biological pump strength at these sites(eg Hull et al 2011 Alegret et al 2012) This led Ale-gret et al (2012) to conclude that the decrease in biologicalpump strength after the KndashPg boundary was most likely aregional instead of a global effect arguing against the Liv-ing Ocean hypothesis We would argue that low-productivityopen ocean sites like the central Pacific are characterized byvery different ecosystem structures and higher productivityas those along the Tethyan margins (eg Dortch and Pack-erd 1989) Therefore the consequence of ecosystem reorga-nization resulting from the mass extinction was likely verydifferent in the open ocean Pacific (eg Sibert et al 2014)Esmeray-Senlet et al (2015) proposed the term ldquoHeteroge-neous Oceanrdquo for this conceptual model as an alternativeto the Living Ocean model Supposedly the HeterogeneousOcean was characterized by a strong geographic heterogene-ity in the extinction patterns and food supply to the sea floor
However as the primary food source for open ocean deep-sea benthic communities generally consists of refractory or-ganic matter (eg Jiao et al 2010) it remains uncertainwhether changes in the intensity of recycling of the gener-ally more labile organic matter in the surface ocean will re-sult in changes in the amount of organic matter that benthiccommunities receive as they will likely continue to receiverecalcitrant organic matter at barely diminished rates despitethe collapse of the biological pump
In addition it should be noted that although the bio-logical pump efficiency is generally relatively high in openocean realms such as the central Pacific the biological pumpstrength is generally very low at these sites (Honjo et al2008 Henson et al 2011) Small changes in absolute bio-logical pump strength at such a locality could therefore havehad a large effect on the local oligotrophic benthic commu-nity (Alegret and Thomas 2009) but they likely representedonly a small fraction of the net amount of organic matterglobally transported to the deep Hence even though localbiological pump strength might have increased at some low-productivity deep-sea sites the large decrease in biologi-cal pump strength recorded at many other sites means thatthe net amount of organic matter globally transported out ofthe surface ocean was likely still reduced in the post-impactworld (Birch et al 2016) Therefore we argue that such Pa-cific records do not represent the general pattern The globalocean response can be characterized as heterogeneous how-ever the KndashPg boundary mass extinction did lead to a strongreduction of the net amount of organic matter globally trans-
ported from the surface to the sea floor while biological pro-ductivity recovered rapidly after the extinction event in ac-cordance with the Living Ocean hypothesis
53 Long-term recovery
Following the disaster phase the hexaperidinioid cysts atOkccedilular and El Kef decreased in abundance and the abun-dance of endobenthic forms increased again as the recoveryof the benthic community was initiated This recovery phaseis approximately correlative to Zone Pα and the basal part ofZone P1a which according to the Paleogene age constraintsof Vandenberghe et al (2012) represents at least 300 kyrThis duration is in agreement with the estimations providedby Birch et al (2016) During this phase the carbon iso-tope records remain well below pre-impact values and thediversity of the benthic foraminiferal community has not yetfully recovered (Figs 4ndash8) This indicates that the rapid andshort-lived KndashPg boundary disaster was followed by a rela-tively long recovery phase in line with previous estimatesof a multimillion-year biological recovery (eg Coxall etal 2006) While the impact-related environmental perturba-tions were short lived (Kring 2007 Vellekoop et al 20142016) the extinctions amongst important biological groupsled to a reduction of the organic flux from the photic zone todeep water resulting in major long-term biological and pa-leoceanographic reorganizations Only with the evolutionaryrecovery of the pelagic community governing the biologicalcarbon pump did export productivity start to increase again(eg Coxall et al 2006 Birch et al 2016)
6 Conclusions
The marine palynological benthic foraminiferal and bulkstable isotope records of the Okccedilular and El Kef sections re-veal major changes across the KndashPg boundary interval por-traying the biological crisis at the KndashPg boundary and subse-quent recovery in the early Paleocene Based on the succes-sion of benthic faunal assemblages at a number of Tethyanmargin sites four phases can be recognized across the KndashPgboundary interval the Maastrichtian or pre-impact phase (I)a disaster phase (II) a recovery phase (III) and an early Pa-leocene post-recovery phase (IV)
Following the KndashPg boundary impact some localitiesshow a large short-lived flux of food to the sea floor likelyrelated to the mass mortality at the KndashPg boundary Howeversince export productivity was greatly reduced after the KndashPgboundary mass extinction it is likely that after the initialpost-impact influx very little fresh organic matter reachedthe sea floor This reduction of export productivity in thepost-extinction disaster phase eventually resulted in a lowerfood supply to the sea floor This presented a major stressfactor for benthic organisms in the western Tethys The re-duced food supply resulted in an abrupt impoverishment of
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
J Vellekoop et al Ecological response to KndashPg boundary mass extinction 897
benthic communities As the downward transport of nutri-ents was slowed down recycling in upper layers increasedAs a result more nutrients remained available for the earlyPaleocene phytoplankton community leading to blooms ofdinoflagellates along the Tethyan margins and blooms of cal-careous nanoplankton taxa in more open ocean sites Ourrecords show that the enhanced recycling of nutrients in theTethys was particularly intense in the tens of thousands ofyears after the impact Following this the slow evolution-ary recovery of the pelagic community governing the bio-logical carbon pump resulted in a gradually increasing ex-port productivity in the hundreds of thousands of years afterthe impact Hence our integration of dinocyst and benthicforaminiferal records across the KndashPg boundary provides im-portant new insights into the ecological responses to the re-duction of export productivity following the mass extinctionat the CretaceousndashPaleogene boundary highlighting the di-rect ecological consequences of the Living Ocean conditionsin the post-impact world
7 Data availability
All data related to publication are available in the Supple-ment
The Supplement related to this article is available onlineat doi105194bg-14-885-2017-supplement
Author contributions Johan Vellekoop Lineke Woelders andHenk Brinkhuis designed the research Samples were collectedin the field by Johan Vellekoop Sanem Accedilikalin Ismail Ouml Yil-maz and Jan Smit Palynological analyses were carried out by Jo-han Vellekoop Benthic foraminiferal analyses were carried out byLineke Woelders and Robert P Speijer Johan Vellekoop LinekeWoelders and Bas van de Schootbrugge prepared the manuscriptwith input from all authors
Competing interests The authors declare that they have no conflictof interest
Acknowledgements This work was supported by the NetherlandsOrganization for Scientific Research (NWO) Open CompetitionGrant ALWPJ09047 to H Brinkhuis and the Research FoundationFlanders (FWO) grant G0B8513 to R P Speijer We thank LKump F Boscolo Galazzo P Roopnarine and one anonymousreferee for their suggestions
Edited by D GillikinReviewed by L Kump F Boscolo Galazzo and P Roopnarine
References
Accedilikalin S Vellekoop J Ocakoglu F Yılmaz I Ouml Smit J Al-tiner S O Goderis S Vonhof H Speijer R P WoeldersL Fornaciari E and Brinkhuis H Geochemical and pale-ontological characterization of a new KndashPg Boundary local-ity from the Northern branch of the Neo-Tethys MudurnundashGoumlynuumlk Basin NW Turkey Cretaceous Res 52 251ndash267doi101016jcretres201407011 2015
Alegret L and Thomas E Food supply to the seafloor in the Pa-cific Ocean after the CretaceousPaleogene boundary event MarMicropaleontol 73 105ndash116 2009
Alegret L Molina E and Thomas E Benthic foraminiferalturnover across the CretaceousPaleogene boundary at Agost(southeastern Spain) paleoenvironmental inferences Mar Mi-cropaleontol 48 251ndash279 2003
Alegret L Thomas E and Lohmann K C End-Cretaceous ma-rine mass extinction not caused by productivity collapse P NatlAcad Sci USA 109 728ndash732 doi101073pnas11106011092012
Alegret L Rodriacuteguez-Tovar F J and Uchman A How bioturba-tion obscured the CretaceousndashPaleogene boundary record TerraNova 27 225ndash230 doi101111ter12151 2015
Altiner D Koccedilyigit A Farinacci A Nicosia U and ContiM A Jurassic-Lower Cretaceous stratigraphy and paleogeo-graphic evolution of the southern part of north-western Anatolia(Turkey) Geologica Romana 27 13ndash80 1991
Alvarez LW Alvarez W Asaro F and Michel H V Extrater-restrial cause for the Cretaceous-Tertiary extinction Science208 1095ndash1108 1980
Alegret L and Thomas E Benthic foraminifera and environmen-tal turnover across the CretaceousPaleogene boundary at BlakeNose (ODP Hole 1049C Northwestern Atlantic) PalaeogeogrPalaeocl 208 59ndash83 doi101016jpalaeo200402028 2004
Armstrong R A Lee C Hedges J I Honjo S and WakehamS G A new mechanistic model for organic carbon fluxes in theocean based on the quantitative association of POC with ballastmaterials Deep-Sea Res Pt II 49 219ndash236 2002
Aubert J and Berggren W A Paleocene benthic foraminiferalbiostratigraphy and paleoecology of Tunisia Bull du Cent deRech de Pau ndash SNPA 10 379ndash469 1976
Berggren W A and Aubert J Paleocene benthonic foraminiferalbiostratigraphy paleobiogeography and paleoecology ofAtlanticndashTethyan regions midway-type fauna PalaeogeogrPalaeocl 18 73ndash192 1975
Bernaola G and Monechi S Calcareous nannofossil ex-tinction and survivorship across the CretaceousndashPaleogeneboundary at Walvis Ridge (ODP Hole 1262C SouthAtlantic Ocean) Palaeogeogr Palaeocl 255 132ndash156doi101016jpalaeo200702045 2007
Bernhard J M Characteristic assemblages and morphologies ofbenthic foraminifera from anoxic organic-rich deposits Jurassicthrough Holocene J Foramin Res 16 207ndash215 1986
Birch H S Coxall H K Pearson P N Kroon D andSchmidt D N Partial collapse of the marine carbon pump af-ter the CretaceousndashPaleogene boundary Geology 44 287ndash290doi101130G375811 2016
Brinkhuis H and Zachariasse W J Dinoflagellate cysts sealevel changes and planktonic foraminifers across the Cretaceous-
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
898 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
Tertiary boundary at El Haria northwest Tunisia Mar Micropa-leontol 13 153ndash191 1988
Brinkhuis H Bujak J P Smit J Versteegh G J M and Viss-cher H Dinoflagellate-based sea surface temperature recon-structions across the Cretaceous-Tertiary boundary PalaeogeogrPalaeocl 141 67ndash83 1998
Buesseler K O The decoupling of production and particulate ex-port in the surface ocean Global Biogeochem Cy 12 297ndash3101998
Buzas M A Culver S J and Jorissen F J A statisti-cal evaluation of the microhabitats of living (stained) infau-nal benthic foraminifera Mar Micropaleontol 20 311ndash320doi1010160377-8398(93)90040-5 1993
Coccioni R and Galeotti S KndashT boundary extinction geolog-ically instantaneous or gradual event Evidence from deep-seabenthic foraminifera Geology 22 779ndash782 1994
Corliss B H Microhabitats of benthic foraminifera within deep-sea sediments Nature 314 435ndash438 doi101038314435a01985
Corliss B H and Chen C Morphotype patterns of Nor-wegian Sea deep-sea benthic foraminifera and ecologi-cal implications Geology 16 716ndash719 doi1011300091-7613(1988)016lt0716MPONSDgt23CO2 1988
Coxall H K DrsquoHondt S and Zachos J C Pelagic evolution andenvironmental recovery after the CretaceousndashPaleogene mass ex-tinction Geology 34 297ndash300 doi101130G217021 2006
Culver S J Benthic foraminifera across the CretaceousndashTertiary(KndashT) boundary a review Mar Micropaleontol 47 177ndash2262003
Cushman J A Upper Cretaceous foraminifera of the Gulf coastalregion of the United States and adjacent areas US Geolog SurvProf Pap 206 241 pp 1946
Cushman J A Paleocene foraminifera of the Gulf Coastal Regionof the United States and adjacent regions US Geol Surv ProfPap 232 75 pp 1951
DrsquoHondt S Consequences of the CretaceousPaleogene Mass Ex-tinction for Marine Ecosystems Annu Rev Ecol Evol S36 295ndash317 doi101146annurevecolsys350211031057152005
DrsquoHondt S and Zachos J C Cretaceous foraminifera and the evo-lutionary history of planktic photosymbiosis Paleobiology 24512ndash523 1998
DrsquoHondt S Donaghay P Zachos J C Luttenberg D andLindinger M Organic Carbon Fluxes and Ecological Recov-ery from the Cretaceous-Tertiary Mass Extinction Science 282276ndash279 1998
Donze P Colin J-P Damotte R Oertli HJ Peypouquet J-Pand Said R Les ostracodes du Campanien terminal agrave lrsquoEocegraveneinfeacuterieur de la coupe du Kef Tuneacutesie nord-occidentale Bull desCentr Recher Expl-Prod Elf Aquit 6 273ndash335 1982
Dortch Q and Packard T T Differences in biomass structure be-tween oligotrophic and eutrophic marine ecosystems Deep-SeaRes 36 223ndash240 1989
Ernst S R Guasti E Dupuis C and Speijer R P En-vironmental perturbation in the southern Tethys across thePaleoceneEocene boundary (Dababiya Egypt) Foraminiferaland clay mineral records Mar Micropaleontol 60 89ndash111doi101016jmarmicro200603002 2006
Eshet Y Moshkovitz S Habib D Benjamini C and Maga-ritz M Calcareous nannofossil and dinoflagellate stratigraphyacross the CretaceousTertiary boundary at Hor Hahar IsraelMar Micropaleontol 18 199ndash228 1992
Eshet Y Almogi-Labin A and Bein A Dinoflagellate cysts pa-leoproductivity and upwelling systems A Late Cretaceous ex-ample from Israel Mar Micropaleontol 23 231ndash240 1994
Esmeray-Senlet S Wright J D Olsson R K MillerK G Browning J V and Quan T M Evidencefor reduced export productivity following the Creta-ceousPaleogene mass extinction Paleoceanography 30718ndash738 doi1010022014PA002724 2015
Giusberti L Boscolo Galazzo F and Thomas E Variability inclimate and productivity during the Paleocene-Eocene ThermalMaximum in the western Tethys (Forada section) Clim Past12 213ndash240 doi105194cp-12-213-2016 2016
Guasti E Kouwenhoven T J Brinkhuis H and Speijer R PPaleocene sea-level and productivity changes at the southernTethyan margin (El Kef Tunisia) Mar Micropaleontol 55 1ndash17 2005
Habib D and Saeedi F The Manumiella seelandica global spikeCooling during regression at the close of the MaastrichtianPalaeogeogr Palaeocl 255 87ndash97 2007
Hain M P Sigmal D M and Haug G H 818 ndash The biolog-ical Pump in the Past in Reference Module in Earth Systemsand Environmental Sciences Treatise on Geochemistry (SecondEdition) The Oceans and Marine Geochemistry 8 485ndash517doi101016B978-0-08-095975-700618-5 2014
Hayek L C and Buzas M A On the proper and efficient useof diversity measures with individual field samples J ForaminRes 43 305ndash313 2013
Henson S A Sanders R Madsen E Morris P J Le MoigneF and Quartly G D A reduced estimate of the strength ofthe oceanrsquos biological carbon pump Geophys Res Lett 38L04606 doi1010292011GL046735 2011
Hilting A K Kump L R and Bralower T J Variations in theoceanic vertical carbon isotope gradient and their implicationsfor the Paleocene-Eocene biological pump Paleoceanography23 PA3222 doi1010292007PA001458 2008
Honjo S Manganini S J Krishfield R A and Francois RParticulate organic carbon fluxes to the ocean interior and fac-tors controlling the biological pump A synthesis of global sed-iment trap progams since 1983 Prog Oceanogr 76 217ndash285doi101016jpocean200711003 2008
Huber B T MacLeod K G and Norris R D Abrupt extinctionand subsequent reworking of Cretaceous planktonic foraminiferaacross the Cretaceous-Tertiary boundary Evidence from the sub-tropical North Atlantic in Catastrophic Events and Mass Extinc-tions Impacts and Beyond edited by Koeberl C and MacLeodK G Geol S Am S 356 277ndash289 Boulder Colorado USA2002
Hull P M Norris R D Bralower T J and Schueth J D A rolefor chance in marine recovery from the end-Cretaceous extinc-tion Nat Geosci 4 856ndash860 doi101038ngeo1302 2011
Hsuuml K J and McKenzie J A A ldquoStrangeloverdquo ocean in the earli-est Tertiary in Natural variations Archean to present edited bySundquist E T and Broecker W S The carbon cycle and atmo-spheric CO2 American Geophysical Union Washington DCUSA 487ndash492 1985
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
J Vellekoop et al Ecological response to KndashPg boundary mass extinction 899
Jiao N Herndl G J Hansell D A Benner R Kattner G Wil-helm S W Kirchman D L Weinbauer M G Luo T ChenF and Azam F Microbial production of recalcitrant dissolvedorganic matter long-term carbon storage in the global oceanNat Rev 8 593ndash599 doi101038nrmicro2386 2010
Jorissen F J de Stigter H C and Widmark J G V A concep-tual model explaining benthic foraminiferal microhabitats MarMicropaleontol 26 3ndash15 doi1010160377-8398(95)00047-X1995
Jorissen F J Fontanier C and Thomas E Paleoceanographi-cal proxies based on deep-sea benthic foraminiferal assemblagecharacteristics in Proxies in Late Cenozoic PaleoceanographyPt 2 Biological tracers and biomarkers edited by Hillaire-Marcel C and de Vernal A 263ndash326 Elsevier Amsterdamthe Netherlands 2007
Kaiho K Kajiwara Y Tazaki K Ueshima M Takeda NKawahata H Arinobu T Ishiwatari R Hirai A and Lam-olda M A Oceanic primary productivity and dissolved oxygenlevels at the CretaceousTertiary boundary their decrease sub-sequent warming and recovery Paleoceanography 14 511ndash5241999
Keller G and Lindinger M Stable isotope TOC and CaCO3record across the CretaceousTertiary boundary at El KefTunisia Palaeogeogr Palaeocl 73 243ndash265 1989
Kellough G R Paleoecology of the Foraminiferida of the WillsPoint Formation (Midway Group) in northeast Texas Trans GulfCoast Assoc Geol Soc 15 73ndash153 1965
Kring D A The Chicxulub impact event and its environmentalconsequences at the CretaceousndashTertiary boundary PalaeogeogrPalaeocl 255 4ndash21 2007
Kump L R Interpreting carbon-isotope excursions Strangeloveoceans Geology 19 299ndash302 1991
Machalski M Vellekoop J Dubicka Z Peryt D and Ha-rasimiuk M Late Maastrichtian cephalopods dinoflagellatecysts and foraminifera from the CretaceousndashPaleogene suc-cession at Lechoacutewka southeast Poland Stratigraphic andenvironmental implications Cretaceous Res 57 208ndash227doi101016jcretres201508012 2016
Molina E Evidence and causes of the main extinctionevents in the Paleogene based on extinction and survivalpatterns of foraminifera Earth-Sci Rev 140 166ndash181doi101016jearscirev201411008 2015
Molina E Alegret L Arenillas I and Arz J The GlobalBoundary Stratotype Section and Point for the base of the Da-nian Stage (Paleocene Paleogeneldquo Tertiaryrdquo Cenozoic) at ElKef Tunisia-Original definition and revision Episodes 29 263ndash273 2006
Olsson R K Liu C and van Fossen M The Cretaceous-Tertiarycatastrophic event at Millers Ferry Alabama in The Cretaceous-Tertiary Event and Other Catastrophes in Earth History editedby Ryder G Fastovsky D and Gartner S Geol S Am S307 263ndash277 1996
Peryt D Benthic foraminiferal response to the Cenomanian-Turonian and CretaceousndashPaleogene boundary events PrzegladGeol 52 827ndash832 2004
Peryt D Alegret L and Molina E The CretaceousPalaeogene(KP) boundary at Aiumln Settara Tunisia restructuring of benthicforaminiferal assemblages Terra Nova 14 101ndash107 2002
Ramette A Multivariate analyses in microbial ecology FEMS Mi-crobiol Ecol 62 142ndash160 2007
Rodriacuteguez-Tovar F J Uchman A MrsquoHamdi A Riahi Sand Ismail-Lattrache K B Ichnological record of palaeoen-vironment from the CretaceousndashPaleogene boundary intervalat El Kef Tunisia The first study of old and new sec-tions at the stratotype area J Afr Earth Sci 120 23ndash30doi101016jjafrearsci201604017 2016
Saner S Mudurnu-Goumlynuumlk Havzasının Jura ve Sonrasıccediloumlkelimnitelikleriyle paleocografya yorumlaması TJK Buumllteni 23 39ndash52 1980
Schioslashler P Brinkhuis H Roncaglia L and Wilson G J Di-noflagellate biostratigraphy and sequence stratigraphy of theType Maastrichtian (Upper Cretaceous) ENCI Quarry TheNetherlands Mar Micropaleontol 31 65ndash95 1997
Schueth J D Bralower T J Jiang S and Patzkowsky ME The role of regional survivor incumbency in the evolu-tionary recovery of calcareous nannoplankton from the Creta-ceousPaleogene (KPg) mass extinction Paleobiology 41 661ndash679 doi101017pab201528 2015
Scotese C R A continental drift flipbook J Geology 112 729ndash741 2004
Scotese C R and Dreher C GlobalGeology available at httpwwwGlobalGeologycom (last access 11 March 2014) 2012
Sepkoski J J Patterns of Phanerozoic extinction a perspectivefrom global data bases in Global Events and Event Stratigra-phy edited by Walliser O H Springer Berlin Germany 35ndash52 2016
Sibert E C Hull P M and Norris R D Resilience of Pacificpelagic fish across the CretaceousPalaeogene mass extinctionNat Geosci 7 667ndash670 doi101038NGEO2227 2014
Slimani H and Toufiq A A CretaceousndashPaleogene boundary geo-logical site revealed by planktic foraminifera and dinoflagellatecysts at Ouled Haddou eastern external Rif Chain Morocco JAfr Earth Sci 88 38ndash52 doi101016jjafrearsci2013080082013
Slimani H Louwye S and Toufiq A Dinoflagellate cystsform the CretaceousndashPaleogene boundary at Ouled Had-dou southeastern Rif Morocco biostratigraphy paleoenvi-ronments and paleobiogeography Palynology 34 90ndash124doi10108001916121003629933 2010
Sluijs A and Brinkhuis H A dynamic climate and ecosystemstate during the Paleocene-Eocene Thermal Maximum infer-ences from dinoflagellate cyst assemblages on the New JerseyShelf Biogeosciences 6 1755ndash1781 doi105194bg-6-1755-2009 2009
Smit J Extinction and evolution of planktonic foraminifera after amajor impact at the CretaceousTertiary boundary Geol S AmS 190 329ndash352 1982
Smit J and Romein A J T A sequence of events across theCretaceous-Tertiary boundary Earth Planet Sci Lett 74 155ndash170 1985
Speijer R P Extinction and recovery patterns in ben-thic foraminiferal paleocommunities across the Creta-ceousPaleogene and PaleoceneEocene boundaries GeologicaUltraiectina 124 Faculteit Aardwetenschappen PhD ThesisUniversiteit Utrecht Utrecht the Netherlands 1994
Speijer R P and van Der Zwaan G J Extinction and survivor-ship of southern Tethyan benthic foraminifera across the Cre-
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
900 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
taceousPaleogene boundary in Biotic recovery from mass ex-tinction events edited by Hart M B Geolog Soc Spec PublLondon UK 102 343ndash371 1996
Stemmann L and Boss E Plankton and Particle Size andPackaging From Determining Optical Properties to Drivingthe Biological Pump Annu Rev Mar Sci 4 263ndash290doi101146annurev-marine-120710-100853 2012
Thomas E Late Cretaceous-early Eocene mass extinctions in thedeep sea Geol S Am S 247 481ndash495 1990
Vandenberghe N Hilgen F J and Speijer R P The PaleogenePeriod in The Geologic Time Scale 2012 edited by GradsteinF M Ogg J G Schmitz M D and Ogg G M 2 855ndash921Elsevier Boston USA 2012
Vellekoop J Sluijs A Smit J Schouten S Weijers J WH Sinninghe Damsteacute J S and Brinkhuis H Rapid short-term cooling following the Chicxulub impact at the CretaceousndashPaleogene boundary P Natl Acad Sci USA 111 7537ndash7541doi101073pnas1319253111 2014
Vellekoop J Smit J van de Schootbrugge B Weijers J W HGaleotti S Sinninghe Damsteacute J S and Brinkhuis H Palyno-logical evidence for prolonged cooling along the Tunisian conti-nental shelf following the KndashPg boundary impact PalaeogeogrPalaeocl 426 216ndash228 doi101016jpalaeo2015030212015
Vellekoop J Esmeray-Senlet S Miller K G Browning J VSluijs A van de Schootbrugge B Sinninghe Damsteacute J Sand Brinkhuis H Evidence for CretaceousndashPaleogene bound-ary bolide ldquoimpact winterrdquo conditions from New Jersey USAGeology 44 619ndash622 doi101130G379611 2016
Woelders L and Speijer R Stable seafloor conditions sealevel and food supply during the latest Maastrichtian atBrazos River Texas Mar Micropaleontol 121 41ndash51doi101016jmarmicro201510002 2015
Zachos J C Arthur M A and Dean W E Geochemical evi-dence for suppression of pelagic marine productivity at the Cre-taceousTertiary boundary Nature 337 61ndash64 1989
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
- Abstract
- Introduction
- Geological setting and age assessment
- Materials and methods
-
- Sampling
- Foraminiferal analysis
- Palynological analysis
- Statistical analysis
-
- Results
-
- Benthic foraminifera
- Palynology
-
- Discussion
-
- Tethyan benthic foraminiferal turnover sequence
- Ecological responses to reduced export productivity
-
- Okccedilular section
- Western Tethys
- Global responses
-
- Long-term recovery
-
- Conclusions
- Data availability
- Author contributions
- Competing interests
- Acknowledgements
- References
-

896 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
gradual recovery across zones Pα and P1a (Fig 8) The simi-larities between the benthic foraminiferal patterns of Tethyanouter neritic to middle bathyal (200ndash1500 m) sites and thoseof the middle to lower bathyal (1500ndash1600 m) record ofBlake Nose (Alegret and Thomas 2004) suggest that theserecords register a global decrease in export productivity fol-lowing the KndashPg boundary
However several open ocean sites mostly in the Pacificshow an opposite trend with several lines of evidence sug-gesting an increased biological pump strength at these sites(eg Hull et al 2011 Alegret et al 2012) This led Ale-gret et al (2012) to conclude that the decrease in biologicalpump strength after the KndashPg boundary was most likely aregional instead of a global effect arguing against the Liv-ing Ocean hypothesis We would argue that low-productivityopen ocean sites like the central Pacific are characterized byvery different ecosystem structures and higher productivityas those along the Tethyan margins (eg Dortch and Pack-erd 1989) Therefore the consequence of ecosystem reorga-nization resulting from the mass extinction was likely verydifferent in the open ocean Pacific (eg Sibert et al 2014)Esmeray-Senlet et al (2015) proposed the term ldquoHeteroge-neous Oceanrdquo for this conceptual model as an alternativeto the Living Ocean model Supposedly the HeterogeneousOcean was characterized by a strong geographic heterogene-ity in the extinction patterns and food supply to the sea floor
However as the primary food source for open ocean deep-sea benthic communities generally consists of refractory or-ganic matter (eg Jiao et al 2010) it remains uncertainwhether changes in the intensity of recycling of the gener-ally more labile organic matter in the surface ocean will re-sult in changes in the amount of organic matter that benthiccommunities receive as they will likely continue to receiverecalcitrant organic matter at barely diminished rates despitethe collapse of the biological pump
In addition it should be noted that although the bio-logical pump efficiency is generally relatively high in openocean realms such as the central Pacific the biological pumpstrength is generally very low at these sites (Honjo et al2008 Henson et al 2011) Small changes in absolute bio-logical pump strength at such a locality could therefore havehad a large effect on the local oligotrophic benthic commu-nity (Alegret and Thomas 2009) but they likely representedonly a small fraction of the net amount of organic matterglobally transported to the deep Hence even though localbiological pump strength might have increased at some low-productivity deep-sea sites the large decrease in biologi-cal pump strength recorded at many other sites means thatthe net amount of organic matter globally transported out ofthe surface ocean was likely still reduced in the post-impactworld (Birch et al 2016) Therefore we argue that such Pa-cific records do not represent the general pattern The globalocean response can be characterized as heterogeneous how-ever the KndashPg boundary mass extinction did lead to a strongreduction of the net amount of organic matter globally trans-
ported from the surface to the sea floor while biological pro-ductivity recovered rapidly after the extinction event in ac-cordance with the Living Ocean hypothesis
53 Long-term recovery
Following the disaster phase the hexaperidinioid cysts atOkccedilular and El Kef decreased in abundance and the abun-dance of endobenthic forms increased again as the recoveryof the benthic community was initiated This recovery phaseis approximately correlative to Zone Pα and the basal part ofZone P1a which according to the Paleogene age constraintsof Vandenberghe et al (2012) represents at least 300 kyrThis duration is in agreement with the estimations providedby Birch et al (2016) During this phase the carbon iso-tope records remain well below pre-impact values and thediversity of the benthic foraminiferal community has not yetfully recovered (Figs 4ndash8) This indicates that the rapid andshort-lived KndashPg boundary disaster was followed by a rela-tively long recovery phase in line with previous estimatesof a multimillion-year biological recovery (eg Coxall etal 2006) While the impact-related environmental perturba-tions were short lived (Kring 2007 Vellekoop et al 20142016) the extinctions amongst important biological groupsled to a reduction of the organic flux from the photic zone todeep water resulting in major long-term biological and pa-leoceanographic reorganizations Only with the evolutionaryrecovery of the pelagic community governing the biologicalcarbon pump did export productivity start to increase again(eg Coxall et al 2006 Birch et al 2016)
6 Conclusions
The marine palynological benthic foraminiferal and bulkstable isotope records of the Okccedilular and El Kef sections re-veal major changes across the KndashPg boundary interval por-traying the biological crisis at the KndashPg boundary and subse-quent recovery in the early Paleocene Based on the succes-sion of benthic faunal assemblages at a number of Tethyanmargin sites four phases can be recognized across the KndashPgboundary interval the Maastrichtian or pre-impact phase (I)a disaster phase (II) a recovery phase (III) and an early Pa-leocene post-recovery phase (IV)
Following the KndashPg boundary impact some localitiesshow a large short-lived flux of food to the sea floor likelyrelated to the mass mortality at the KndashPg boundary Howeversince export productivity was greatly reduced after the KndashPgboundary mass extinction it is likely that after the initialpost-impact influx very little fresh organic matter reachedthe sea floor This reduction of export productivity in thepost-extinction disaster phase eventually resulted in a lowerfood supply to the sea floor This presented a major stressfactor for benthic organisms in the western Tethys The re-duced food supply resulted in an abrupt impoverishment of
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
J Vellekoop et al Ecological response to KndashPg boundary mass extinction 897
benthic communities As the downward transport of nutri-ents was slowed down recycling in upper layers increasedAs a result more nutrients remained available for the earlyPaleocene phytoplankton community leading to blooms ofdinoflagellates along the Tethyan margins and blooms of cal-careous nanoplankton taxa in more open ocean sites Ourrecords show that the enhanced recycling of nutrients in theTethys was particularly intense in the tens of thousands ofyears after the impact Following this the slow evolution-ary recovery of the pelagic community governing the bio-logical carbon pump resulted in a gradually increasing ex-port productivity in the hundreds of thousands of years afterthe impact Hence our integration of dinocyst and benthicforaminiferal records across the KndashPg boundary provides im-portant new insights into the ecological responses to the re-duction of export productivity following the mass extinctionat the CretaceousndashPaleogene boundary highlighting the di-rect ecological consequences of the Living Ocean conditionsin the post-impact world
7 Data availability
All data related to publication are available in the Supple-ment
The Supplement related to this article is available onlineat doi105194bg-14-885-2017-supplement
Author contributions Johan Vellekoop Lineke Woelders andHenk Brinkhuis designed the research Samples were collectedin the field by Johan Vellekoop Sanem Accedilikalin Ismail Ouml Yil-maz and Jan Smit Palynological analyses were carried out by Jo-han Vellekoop Benthic foraminiferal analyses were carried out byLineke Woelders and Robert P Speijer Johan Vellekoop LinekeWoelders and Bas van de Schootbrugge prepared the manuscriptwith input from all authors
Competing interests The authors declare that they have no conflictof interest
Acknowledgements This work was supported by the NetherlandsOrganization for Scientific Research (NWO) Open CompetitionGrant ALWPJ09047 to H Brinkhuis and the Research FoundationFlanders (FWO) grant G0B8513 to R P Speijer We thank LKump F Boscolo Galazzo P Roopnarine and one anonymousreferee for their suggestions
Edited by D GillikinReviewed by L Kump F Boscolo Galazzo and P Roopnarine
References
Accedilikalin S Vellekoop J Ocakoglu F Yılmaz I Ouml Smit J Al-tiner S O Goderis S Vonhof H Speijer R P WoeldersL Fornaciari E and Brinkhuis H Geochemical and pale-ontological characterization of a new KndashPg Boundary local-ity from the Northern branch of the Neo-Tethys MudurnundashGoumlynuumlk Basin NW Turkey Cretaceous Res 52 251ndash267doi101016jcretres201407011 2015
Alegret L and Thomas E Food supply to the seafloor in the Pa-cific Ocean after the CretaceousPaleogene boundary event MarMicropaleontol 73 105ndash116 2009
Alegret L Molina E and Thomas E Benthic foraminiferalturnover across the CretaceousPaleogene boundary at Agost(southeastern Spain) paleoenvironmental inferences Mar Mi-cropaleontol 48 251ndash279 2003
Alegret L Thomas E and Lohmann K C End-Cretaceous ma-rine mass extinction not caused by productivity collapse P NatlAcad Sci USA 109 728ndash732 doi101073pnas11106011092012
Alegret L Rodriacuteguez-Tovar F J and Uchman A How bioturba-tion obscured the CretaceousndashPaleogene boundary record TerraNova 27 225ndash230 doi101111ter12151 2015
Altiner D Koccedilyigit A Farinacci A Nicosia U and ContiM A Jurassic-Lower Cretaceous stratigraphy and paleogeo-graphic evolution of the southern part of north-western Anatolia(Turkey) Geologica Romana 27 13ndash80 1991
Alvarez LW Alvarez W Asaro F and Michel H V Extrater-restrial cause for the Cretaceous-Tertiary extinction Science208 1095ndash1108 1980
Alegret L and Thomas E Benthic foraminifera and environmen-tal turnover across the CretaceousPaleogene boundary at BlakeNose (ODP Hole 1049C Northwestern Atlantic) PalaeogeogrPalaeocl 208 59ndash83 doi101016jpalaeo200402028 2004
Armstrong R A Lee C Hedges J I Honjo S and WakehamS G A new mechanistic model for organic carbon fluxes in theocean based on the quantitative association of POC with ballastmaterials Deep-Sea Res Pt II 49 219ndash236 2002
Aubert J and Berggren W A Paleocene benthic foraminiferalbiostratigraphy and paleoecology of Tunisia Bull du Cent deRech de Pau ndash SNPA 10 379ndash469 1976
Berggren W A and Aubert J Paleocene benthonic foraminiferalbiostratigraphy paleobiogeography and paleoecology ofAtlanticndashTethyan regions midway-type fauna PalaeogeogrPalaeocl 18 73ndash192 1975
Bernaola G and Monechi S Calcareous nannofossil ex-tinction and survivorship across the CretaceousndashPaleogeneboundary at Walvis Ridge (ODP Hole 1262C SouthAtlantic Ocean) Palaeogeogr Palaeocl 255 132ndash156doi101016jpalaeo200702045 2007
Bernhard J M Characteristic assemblages and morphologies ofbenthic foraminifera from anoxic organic-rich deposits Jurassicthrough Holocene J Foramin Res 16 207ndash215 1986
Birch H S Coxall H K Pearson P N Kroon D andSchmidt D N Partial collapse of the marine carbon pump af-ter the CretaceousndashPaleogene boundary Geology 44 287ndash290doi101130G375811 2016
Brinkhuis H and Zachariasse W J Dinoflagellate cysts sealevel changes and planktonic foraminifers across the Cretaceous-
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
898 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
Tertiary boundary at El Haria northwest Tunisia Mar Micropa-leontol 13 153ndash191 1988
Brinkhuis H Bujak J P Smit J Versteegh G J M and Viss-cher H Dinoflagellate-based sea surface temperature recon-structions across the Cretaceous-Tertiary boundary PalaeogeogrPalaeocl 141 67ndash83 1998
Buesseler K O The decoupling of production and particulate ex-port in the surface ocean Global Biogeochem Cy 12 297ndash3101998
Buzas M A Culver S J and Jorissen F J A statisti-cal evaluation of the microhabitats of living (stained) infau-nal benthic foraminifera Mar Micropaleontol 20 311ndash320doi1010160377-8398(93)90040-5 1993
Coccioni R and Galeotti S KndashT boundary extinction geolog-ically instantaneous or gradual event Evidence from deep-seabenthic foraminifera Geology 22 779ndash782 1994
Corliss B H Microhabitats of benthic foraminifera within deep-sea sediments Nature 314 435ndash438 doi101038314435a01985
Corliss B H and Chen C Morphotype patterns of Nor-wegian Sea deep-sea benthic foraminifera and ecologi-cal implications Geology 16 716ndash719 doi1011300091-7613(1988)016lt0716MPONSDgt23CO2 1988
Coxall H K DrsquoHondt S and Zachos J C Pelagic evolution andenvironmental recovery after the CretaceousndashPaleogene mass ex-tinction Geology 34 297ndash300 doi101130G217021 2006
Culver S J Benthic foraminifera across the CretaceousndashTertiary(KndashT) boundary a review Mar Micropaleontol 47 177ndash2262003
Cushman J A Upper Cretaceous foraminifera of the Gulf coastalregion of the United States and adjacent areas US Geolog SurvProf Pap 206 241 pp 1946
Cushman J A Paleocene foraminifera of the Gulf Coastal Regionof the United States and adjacent regions US Geol Surv ProfPap 232 75 pp 1951
DrsquoHondt S Consequences of the CretaceousPaleogene Mass Ex-tinction for Marine Ecosystems Annu Rev Ecol Evol S36 295ndash317 doi101146annurevecolsys350211031057152005
DrsquoHondt S and Zachos J C Cretaceous foraminifera and the evo-lutionary history of planktic photosymbiosis Paleobiology 24512ndash523 1998
DrsquoHondt S Donaghay P Zachos J C Luttenberg D andLindinger M Organic Carbon Fluxes and Ecological Recov-ery from the Cretaceous-Tertiary Mass Extinction Science 282276ndash279 1998
Donze P Colin J-P Damotte R Oertli HJ Peypouquet J-Pand Said R Les ostracodes du Campanien terminal agrave lrsquoEocegraveneinfeacuterieur de la coupe du Kef Tuneacutesie nord-occidentale Bull desCentr Recher Expl-Prod Elf Aquit 6 273ndash335 1982
Dortch Q and Packard T T Differences in biomass structure be-tween oligotrophic and eutrophic marine ecosystems Deep-SeaRes 36 223ndash240 1989
Ernst S R Guasti E Dupuis C and Speijer R P En-vironmental perturbation in the southern Tethys across thePaleoceneEocene boundary (Dababiya Egypt) Foraminiferaland clay mineral records Mar Micropaleontol 60 89ndash111doi101016jmarmicro200603002 2006
Eshet Y Moshkovitz S Habib D Benjamini C and Maga-ritz M Calcareous nannofossil and dinoflagellate stratigraphyacross the CretaceousTertiary boundary at Hor Hahar IsraelMar Micropaleontol 18 199ndash228 1992
Eshet Y Almogi-Labin A and Bein A Dinoflagellate cysts pa-leoproductivity and upwelling systems A Late Cretaceous ex-ample from Israel Mar Micropaleontol 23 231ndash240 1994
Esmeray-Senlet S Wright J D Olsson R K MillerK G Browning J V and Quan T M Evidencefor reduced export productivity following the Creta-ceousPaleogene mass extinction Paleoceanography 30718ndash738 doi1010022014PA002724 2015
Giusberti L Boscolo Galazzo F and Thomas E Variability inclimate and productivity during the Paleocene-Eocene ThermalMaximum in the western Tethys (Forada section) Clim Past12 213ndash240 doi105194cp-12-213-2016 2016
Guasti E Kouwenhoven T J Brinkhuis H and Speijer R PPaleocene sea-level and productivity changes at the southernTethyan margin (El Kef Tunisia) Mar Micropaleontol 55 1ndash17 2005
Habib D and Saeedi F The Manumiella seelandica global spikeCooling during regression at the close of the MaastrichtianPalaeogeogr Palaeocl 255 87ndash97 2007
Hain M P Sigmal D M and Haug G H 818 ndash The biolog-ical Pump in the Past in Reference Module in Earth Systemsand Environmental Sciences Treatise on Geochemistry (SecondEdition) The Oceans and Marine Geochemistry 8 485ndash517doi101016B978-0-08-095975-700618-5 2014
Hayek L C and Buzas M A On the proper and efficient useof diversity measures with individual field samples J ForaminRes 43 305ndash313 2013
Henson S A Sanders R Madsen E Morris P J Le MoigneF and Quartly G D A reduced estimate of the strength ofthe oceanrsquos biological carbon pump Geophys Res Lett 38L04606 doi1010292011GL046735 2011
Hilting A K Kump L R and Bralower T J Variations in theoceanic vertical carbon isotope gradient and their implicationsfor the Paleocene-Eocene biological pump Paleoceanography23 PA3222 doi1010292007PA001458 2008
Honjo S Manganini S J Krishfield R A and Francois RParticulate organic carbon fluxes to the ocean interior and fac-tors controlling the biological pump A synthesis of global sed-iment trap progams since 1983 Prog Oceanogr 76 217ndash285doi101016jpocean200711003 2008
Huber B T MacLeod K G and Norris R D Abrupt extinctionand subsequent reworking of Cretaceous planktonic foraminiferaacross the Cretaceous-Tertiary boundary Evidence from the sub-tropical North Atlantic in Catastrophic Events and Mass Extinc-tions Impacts and Beyond edited by Koeberl C and MacLeodK G Geol S Am S 356 277ndash289 Boulder Colorado USA2002
Hull P M Norris R D Bralower T J and Schueth J D A rolefor chance in marine recovery from the end-Cretaceous extinc-tion Nat Geosci 4 856ndash860 doi101038ngeo1302 2011
Hsuuml K J and McKenzie J A A ldquoStrangeloverdquo ocean in the earli-est Tertiary in Natural variations Archean to present edited bySundquist E T and Broecker W S The carbon cycle and atmo-spheric CO2 American Geophysical Union Washington DCUSA 487ndash492 1985
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
J Vellekoop et al Ecological response to KndashPg boundary mass extinction 899
Jiao N Herndl G J Hansell D A Benner R Kattner G Wil-helm S W Kirchman D L Weinbauer M G Luo T ChenF and Azam F Microbial production of recalcitrant dissolvedorganic matter long-term carbon storage in the global oceanNat Rev 8 593ndash599 doi101038nrmicro2386 2010
Jorissen F J de Stigter H C and Widmark J G V A concep-tual model explaining benthic foraminiferal microhabitats MarMicropaleontol 26 3ndash15 doi1010160377-8398(95)00047-X1995
Jorissen F J Fontanier C and Thomas E Paleoceanographi-cal proxies based on deep-sea benthic foraminiferal assemblagecharacteristics in Proxies in Late Cenozoic PaleoceanographyPt 2 Biological tracers and biomarkers edited by Hillaire-Marcel C and de Vernal A 263ndash326 Elsevier Amsterdamthe Netherlands 2007
Kaiho K Kajiwara Y Tazaki K Ueshima M Takeda NKawahata H Arinobu T Ishiwatari R Hirai A and Lam-olda M A Oceanic primary productivity and dissolved oxygenlevels at the CretaceousTertiary boundary their decrease sub-sequent warming and recovery Paleoceanography 14 511ndash5241999
Keller G and Lindinger M Stable isotope TOC and CaCO3record across the CretaceousTertiary boundary at El KefTunisia Palaeogeogr Palaeocl 73 243ndash265 1989
Kellough G R Paleoecology of the Foraminiferida of the WillsPoint Formation (Midway Group) in northeast Texas Trans GulfCoast Assoc Geol Soc 15 73ndash153 1965
Kring D A The Chicxulub impact event and its environmentalconsequences at the CretaceousndashTertiary boundary PalaeogeogrPalaeocl 255 4ndash21 2007
Kump L R Interpreting carbon-isotope excursions Strangeloveoceans Geology 19 299ndash302 1991
Machalski M Vellekoop J Dubicka Z Peryt D and Ha-rasimiuk M Late Maastrichtian cephalopods dinoflagellatecysts and foraminifera from the CretaceousndashPaleogene suc-cession at Lechoacutewka southeast Poland Stratigraphic andenvironmental implications Cretaceous Res 57 208ndash227doi101016jcretres201508012 2016
Molina E Evidence and causes of the main extinctionevents in the Paleogene based on extinction and survivalpatterns of foraminifera Earth-Sci Rev 140 166ndash181doi101016jearscirev201411008 2015
Molina E Alegret L Arenillas I and Arz J The GlobalBoundary Stratotype Section and Point for the base of the Da-nian Stage (Paleocene Paleogeneldquo Tertiaryrdquo Cenozoic) at ElKef Tunisia-Original definition and revision Episodes 29 263ndash273 2006
Olsson R K Liu C and van Fossen M The Cretaceous-Tertiarycatastrophic event at Millers Ferry Alabama in The Cretaceous-Tertiary Event and Other Catastrophes in Earth History editedby Ryder G Fastovsky D and Gartner S Geol S Am S307 263ndash277 1996
Peryt D Benthic foraminiferal response to the Cenomanian-Turonian and CretaceousndashPaleogene boundary events PrzegladGeol 52 827ndash832 2004
Peryt D Alegret L and Molina E The CretaceousPalaeogene(KP) boundary at Aiumln Settara Tunisia restructuring of benthicforaminiferal assemblages Terra Nova 14 101ndash107 2002
Ramette A Multivariate analyses in microbial ecology FEMS Mi-crobiol Ecol 62 142ndash160 2007
Rodriacuteguez-Tovar F J Uchman A MrsquoHamdi A Riahi Sand Ismail-Lattrache K B Ichnological record of palaeoen-vironment from the CretaceousndashPaleogene boundary intervalat El Kef Tunisia The first study of old and new sec-tions at the stratotype area J Afr Earth Sci 120 23ndash30doi101016jjafrearsci201604017 2016
Saner S Mudurnu-Goumlynuumlk Havzasının Jura ve Sonrasıccediloumlkelimnitelikleriyle paleocografya yorumlaması TJK Buumllteni 23 39ndash52 1980
Schioslashler P Brinkhuis H Roncaglia L and Wilson G J Di-noflagellate biostratigraphy and sequence stratigraphy of theType Maastrichtian (Upper Cretaceous) ENCI Quarry TheNetherlands Mar Micropaleontol 31 65ndash95 1997
Schueth J D Bralower T J Jiang S and Patzkowsky ME The role of regional survivor incumbency in the evolu-tionary recovery of calcareous nannoplankton from the Creta-ceousPaleogene (KPg) mass extinction Paleobiology 41 661ndash679 doi101017pab201528 2015
Scotese C R A continental drift flipbook J Geology 112 729ndash741 2004
Scotese C R and Dreher C GlobalGeology available at httpwwwGlobalGeologycom (last access 11 March 2014) 2012
Sepkoski J J Patterns of Phanerozoic extinction a perspectivefrom global data bases in Global Events and Event Stratigra-phy edited by Walliser O H Springer Berlin Germany 35ndash52 2016
Sibert E C Hull P M and Norris R D Resilience of Pacificpelagic fish across the CretaceousPalaeogene mass extinctionNat Geosci 7 667ndash670 doi101038NGEO2227 2014
Slimani H and Toufiq A A CretaceousndashPaleogene boundary geo-logical site revealed by planktic foraminifera and dinoflagellatecysts at Ouled Haddou eastern external Rif Chain Morocco JAfr Earth Sci 88 38ndash52 doi101016jjafrearsci2013080082013
Slimani H Louwye S and Toufiq A Dinoflagellate cystsform the CretaceousndashPaleogene boundary at Ouled Had-dou southeastern Rif Morocco biostratigraphy paleoenvi-ronments and paleobiogeography Palynology 34 90ndash124doi10108001916121003629933 2010
Sluijs A and Brinkhuis H A dynamic climate and ecosystemstate during the Paleocene-Eocene Thermal Maximum infer-ences from dinoflagellate cyst assemblages on the New JerseyShelf Biogeosciences 6 1755ndash1781 doi105194bg-6-1755-2009 2009
Smit J Extinction and evolution of planktonic foraminifera after amajor impact at the CretaceousTertiary boundary Geol S AmS 190 329ndash352 1982
Smit J and Romein A J T A sequence of events across theCretaceous-Tertiary boundary Earth Planet Sci Lett 74 155ndash170 1985
Speijer R P Extinction and recovery patterns in ben-thic foraminiferal paleocommunities across the Creta-ceousPaleogene and PaleoceneEocene boundaries GeologicaUltraiectina 124 Faculteit Aardwetenschappen PhD ThesisUniversiteit Utrecht Utrecht the Netherlands 1994
Speijer R P and van Der Zwaan G J Extinction and survivor-ship of southern Tethyan benthic foraminifera across the Cre-
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
900 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
taceousPaleogene boundary in Biotic recovery from mass ex-tinction events edited by Hart M B Geolog Soc Spec PublLondon UK 102 343ndash371 1996
Stemmann L and Boss E Plankton and Particle Size andPackaging From Determining Optical Properties to Drivingthe Biological Pump Annu Rev Mar Sci 4 263ndash290doi101146annurev-marine-120710-100853 2012
Thomas E Late Cretaceous-early Eocene mass extinctions in thedeep sea Geol S Am S 247 481ndash495 1990
Vandenberghe N Hilgen F J and Speijer R P The PaleogenePeriod in The Geologic Time Scale 2012 edited by GradsteinF M Ogg J G Schmitz M D and Ogg G M 2 855ndash921Elsevier Boston USA 2012
Vellekoop J Sluijs A Smit J Schouten S Weijers J WH Sinninghe Damsteacute J S and Brinkhuis H Rapid short-term cooling following the Chicxulub impact at the CretaceousndashPaleogene boundary P Natl Acad Sci USA 111 7537ndash7541doi101073pnas1319253111 2014
Vellekoop J Smit J van de Schootbrugge B Weijers J W HGaleotti S Sinninghe Damsteacute J S and Brinkhuis H Palyno-logical evidence for prolonged cooling along the Tunisian conti-nental shelf following the KndashPg boundary impact PalaeogeogrPalaeocl 426 216ndash228 doi101016jpalaeo2015030212015
Vellekoop J Esmeray-Senlet S Miller K G Browning J VSluijs A van de Schootbrugge B Sinninghe Damsteacute J Sand Brinkhuis H Evidence for CretaceousndashPaleogene bound-ary bolide ldquoimpact winterrdquo conditions from New Jersey USAGeology 44 619ndash622 doi101130G379611 2016
Woelders L and Speijer R Stable seafloor conditions sealevel and food supply during the latest Maastrichtian atBrazos River Texas Mar Micropaleontol 121 41ndash51doi101016jmarmicro201510002 2015
Zachos J C Arthur M A and Dean W E Geochemical evi-dence for suppression of pelagic marine productivity at the Cre-taceousTertiary boundary Nature 337 61ndash64 1989
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
- Abstract
- Introduction
- Geological setting and age assessment
- Materials and methods
-
- Sampling
- Foraminiferal analysis
- Palynological analysis
- Statistical analysis
-
- Results
-
- Benthic foraminifera
- Palynology
-
- Discussion
-
- Tethyan benthic foraminiferal turnover sequence
- Ecological responses to reduced export productivity
-
- Okccedilular section
- Western Tethys
- Global responses
-
- Long-term recovery
-
- Conclusions
- Data availability
- Author contributions
- Competing interests
- Acknowledgements
- References
-

J Vellekoop et al Ecological response to KndashPg boundary mass extinction 897
benthic communities As the downward transport of nutri-ents was slowed down recycling in upper layers increasedAs a result more nutrients remained available for the earlyPaleocene phytoplankton community leading to blooms ofdinoflagellates along the Tethyan margins and blooms of cal-careous nanoplankton taxa in more open ocean sites Ourrecords show that the enhanced recycling of nutrients in theTethys was particularly intense in the tens of thousands ofyears after the impact Following this the slow evolution-ary recovery of the pelagic community governing the bio-logical carbon pump resulted in a gradually increasing ex-port productivity in the hundreds of thousands of years afterthe impact Hence our integration of dinocyst and benthicforaminiferal records across the KndashPg boundary provides im-portant new insights into the ecological responses to the re-duction of export productivity following the mass extinctionat the CretaceousndashPaleogene boundary highlighting the di-rect ecological consequences of the Living Ocean conditionsin the post-impact world
7 Data availability
All data related to publication are available in the Supple-ment
The Supplement related to this article is available onlineat doi105194bg-14-885-2017-supplement
Author contributions Johan Vellekoop Lineke Woelders andHenk Brinkhuis designed the research Samples were collectedin the field by Johan Vellekoop Sanem Accedilikalin Ismail Ouml Yil-maz and Jan Smit Palynological analyses were carried out by Jo-han Vellekoop Benthic foraminiferal analyses were carried out byLineke Woelders and Robert P Speijer Johan Vellekoop LinekeWoelders and Bas van de Schootbrugge prepared the manuscriptwith input from all authors
Competing interests The authors declare that they have no conflictof interest
Acknowledgements This work was supported by the NetherlandsOrganization for Scientific Research (NWO) Open CompetitionGrant ALWPJ09047 to H Brinkhuis and the Research FoundationFlanders (FWO) grant G0B8513 to R P Speijer We thank LKump F Boscolo Galazzo P Roopnarine and one anonymousreferee for their suggestions
Edited by D GillikinReviewed by L Kump F Boscolo Galazzo and P Roopnarine
References
Accedilikalin S Vellekoop J Ocakoglu F Yılmaz I Ouml Smit J Al-tiner S O Goderis S Vonhof H Speijer R P WoeldersL Fornaciari E and Brinkhuis H Geochemical and pale-ontological characterization of a new KndashPg Boundary local-ity from the Northern branch of the Neo-Tethys MudurnundashGoumlynuumlk Basin NW Turkey Cretaceous Res 52 251ndash267doi101016jcretres201407011 2015
Alegret L and Thomas E Food supply to the seafloor in the Pa-cific Ocean after the CretaceousPaleogene boundary event MarMicropaleontol 73 105ndash116 2009
Alegret L Molina E and Thomas E Benthic foraminiferalturnover across the CretaceousPaleogene boundary at Agost(southeastern Spain) paleoenvironmental inferences Mar Mi-cropaleontol 48 251ndash279 2003
Alegret L Thomas E and Lohmann K C End-Cretaceous ma-rine mass extinction not caused by productivity collapse P NatlAcad Sci USA 109 728ndash732 doi101073pnas11106011092012
Alegret L Rodriacuteguez-Tovar F J and Uchman A How bioturba-tion obscured the CretaceousndashPaleogene boundary record TerraNova 27 225ndash230 doi101111ter12151 2015
Altiner D Koccedilyigit A Farinacci A Nicosia U and ContiM A Jurassic-Lower Cretaceous stratigraphy and paleogeo-graphic evolution of the southern part of north-western Anatolia(Turkey) Geologica Romana 27 13ndash80 1991
Alvarez LW Alvarez W Asaro F and Michel H V Extrater-restrial cause for the Cretaceous-Tertiary extinction Science208 1095ndash1108 1980
Alegret L and Thomas E Benthic foraminifera and environmen-tal turnover across the CretaceousPaleogene boundary at BlakeNose (ODP Hole 1049C Northwestern Atlantic) PalaeogeogrPalaeocl 208 59ndash83 doi101016jpalaeo200402028 2004
Armstrong R A Lee C Hedges J I Honjo S and WakehamS G A new mechanistic model for organic carbon fluxes in theocean based on the quantitative association of POC with ballastmaterials Deep-Sea Res Pt II 49 219ndash236 2002
Aubert J and Berggren W A Paleocene benthic foraminiferalbiostratigraphy and paleoecology of Tunisia Bull du Cent deRech de Pau ndash SNPA 10 379ndash469 1976
Berggren W A and Aubert J Paleocene benthonic foraminiferalbiostratigraphy paleobiogeography and paleoecology ofAtlanticndashTethyan regions midway-type fauna PalaeogeogrPalaeocl 18 73ndash192 1975
Bernaola G and Monechi S Calcareous nannofossil ex-tinction and survivorship across the CretaceousndashPaleogeneboundary at Walvis Ridge (ODP Hole 1262C SouthAtlantic Ocean) Palaeogeogr Palaeocl 255 132ndash156doi101016jpalaeo200702045 2007
Bernhard J M Characteristic assemblages and morphologies ofbenthic foraminifera from anoxic organic-rich deposits Jurassicthrough Holocene J Foramin Res 16 207ndash215 1986
Birch H S Coxall H K Pearson P N Kroon D andSchmidt D N Partial collapse of the marine carbon pump af-ter the CretaceousndashPaleogene boundary Geology 44 287ndash290doi101130G375811 2016
Brinkhuis H and Zachariasse W J Dinoflagellate cysts sealevel changes and planktonic foraminifers across the Cretaceous-
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
898 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
Tertiary boundary at El Haria northwest Tunisia Mar Micropa-leontol 13 153ndash191 1988
Brinkhuis H Bujak J P Smit J Versteegh G J M and Viss-cher H Dinoflagellate-based sea surface temperature recon-structions across the Cretaceous-Tertiary boundary PalaeogeogrPalaeocl 141 67ndash83 1998
Buesseler K O The decoupling of production and particulate ex-port in the surface ocean Global Biogeochem Cy 12 297ndash3101998
Buzas M A Culver S J and Jorissen F J A statisti-cal evaluation of the microhabitats of living (stained) infau-nal benthic foraminifera Mar Micropaleontol 20 311ndash320doi1010160377-8398(93)90040-5 1993
Coccioni R and Galeotti S KndashT boundary extinction geolog-ically instantaneous or gradual event Evidence from deep-seabenthic foraminifera Geology 22 779ndash782 1994
Corliss B H Microhabitats of benthic foraminifera within deep-sea sediments Nature 314 435ndash438 doi101038314435a01985
Corliss B H and Chen C Morphotype patterns of Nor-wegian Sea deep-sea benthic foraminifera and ecologi-cal implications Geology 16 716ndash719 doi1011300091-7613(1988)016lt0716MPONSDgt23CO2 1988
Coxall H K DrsquoHondt S and Zachos J C Pelagic evolution andenvironmental recovery after the CretaceousndashPaleogene mass ex-tinction Geology 34 297ndash300 doi101130G217021 2006
Culver S J Benthic foraminifera across the CretaceousndashTertiary(KndashT) boundary a review Mar Micropaleontol 47 177ndash2262003
Cushman J A Upper Cretaceous foraminifera of the Gulf coastalregion of the United States and adjacent areas US Geolog SurvProf Pap 206 241 pp 1946
Cushman J A Paleocene foraminifera of the Gulf Coastal Regionof the United States and adjacent regions US Geol Surv ProfPap 232 75 pp 1951
DrsquoHondt S Consequences of the CretaceousPaleogene Mass Ex-tinction for Marine Ecosystems Annu Rev Ecol Evol S36 295ndash317 doi101146annurevecolsys350211031057152005
DrsquoHondt S and Zachos J C Cretaceous foraminifera and the evo-lutionary history of planktic photosymbiosis Paleobiology 24512ndash523 1998
DrsquoHondt S Donaghay P Zachos J C Luttenberg D andLindinger M Organic Carbon Fluxes and Ecological Recov-ery from the Cretaceous-Tertiary Mass Extinction Science 282276ndash279 1998
Donze P Colin J-P Damotte R Oertli HJ Peypouquet J-Pand Said R Les ostracodes du Campanien terminal agrave lrsquoEocegraveneinfeacuterieur de la coupe du Kef Tuneacutesie nord-occidentale Bull desCentr Recher Expl-Prod Elf Aquit 6 273ndash335 1982
Dortch Q and Packard T T Differences in biomass structure be-tween oligotrophic and eutrophic marine ecosystems Deep-SeaRes 36 223ndash240 1989
Ernst S R Guasti E Dupuis C and Speijer R P En-vironmental perturbation in the southern Tethys across thePaleoceneEocene boundary (Dababiya Egypt) Foraminiferaland clay mineral records Mar Micropaleontol 60 89ndash111doi101016jmarmicro200603002 2006
Eshet Y Moshkovitz S Habib D Benjamini C and Maga-ritz M Calcareous nannofossil and dinoflagellate stratigraphyacross the CretaceousTertiary boundary at Hor Hahar IsraelMar Micropaleontol 18 199ndash228 1992
Eshet Y Almogi-Labin A and Bein A Dinoflagellate cysts pa-leoproductivity and upwelling systems A Late Cretaceous ex-ample from Israel Mar Micropaleontol 23 231ndash240 1994
Esmeray-Senlet S Wright J D Olsson R K MillerK G Browning J V and Quan T M Evidencefor reduced export productivity following the Creta-ceousPaleogene mass extinction Paleoceanography 30718ndash738 doi1010022014PA002724 2015
Giusberti L Boscolo Galazzo F and Thomas E Variability inclimate and productivity during the Paleocene-Eocene ThermalMaximum in the western Tethys (Forada section) Clim Past12 213ndash240 doi105194cp-12-213-2016 2016
Guasti E Kouwenhoven T J Brinkhuis H and Speijer R PPaleocene sea-level and productivity changes at the southernTethyan margin (El Kef Tunisia) Mar Micropaleontol 55 1ndash17 2005
Habib D and Saeedi F The Manumiella seelandica global spikeCooling during regression at the close of the MaastrichtianPalaeogeogr Palaeocl 255 87ndash97 2007
Hain M P Sigmal D M and Haug G H 818 ndash The biolog-ical Pump in the Past in Reference Module in Earth Systemsand Environmental Sciences Treatise on Geochemistry (SecondEdition) The Oceans and Marine Geochemistry 8 485ndash517doi101016B978-0-08-095975-700618-5 2014
Hayek L C and Buzas M A On the proper and efficient useof diversity measures with individual field samples J ForaminRes 43 305ndash313 2013
Henson S A Sanders R Madsen E Morris P J Le MoigneF and Quartly G D A reduced estimate of the strength ofthe oceanrsquos biological carbon pump Geophys Res Lett 38L04606 doi1010292011GL046735 2011
Hilting A K Kump L R and Bralower T J Variations in theoceanic vertical carbon isotope gradient and their implicationsfor the Paleocene-Eocene biological pump Paleoceanography23 PA3222 doi1010292007PA001458 2008
Honjo S Manganini S J Krishfield R A and Francois RParticulate organic carbon fluxes to the ocean interior and fac-tors controlling the biological pump A synthesis of global sed-iment trap progams since 1983 Prog Oceanogr 76 217ndash285doi101016jpocean200711003 2008
Huber B T MacLeod K G and Norris R D Abrupt extinctionand subsequent reworking of Cretaceous planktonic foraminiferaacross the Cretaceous-Tertiary boundary Evidence from the sub-tropical North Atlantic in Catastrophic Events and Mass Extinc-tions Impacts and Beyond edited by Koeberl C and MacLeodK G Geol S Am S 356 277ndash289 Boulder Colorado USA2002
Hull P M Norris R D Bralower T J and Schueth J D A rolefor chance in marine recovery from the end-Cretaceous extinc-tion Nat Geosci 4 856ndash860 doi101038ngeo1302 2011
Hsuuml K J and McKenzie J A A ldquoStrangeloverdquo ocean in the earli-est Tertiary in Natural variations Archean to present edited bySundquist E T and Broecker W S The carbon cycle and atmo-spheric CO2 American Geophysical Union Washington DCUSA 487ndash492 1985
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
J Vellekoop et al Ecological response to KndashPg boundary mass extinction 899
Jiao N Herndl G J Hansell D A Benner R Kattner G Wil-helm S W Kirchman D L Weinbauer M G Luo T ChenF and Azam F Microbial production of recalcitrant dissolvedorganic matter long-term carbon storage in the global oceanNat Rev 8 593ndash599 doi101038nrmicro2386 2010
Jorissen F J de Stigter H C and Widmark J G V A concep-tual model explaining benthic foraminiferal microhabitats MarMicropaleontol 26 3ndash15 doi1010160377-8398(95)00047-X1995
Jorissen F J Fontanier C and Thomas E Paleoceanographi-cal proxies based on deep-sea benthic foraminiferal assemblagecharacteristics in Proxies in Late Cenozoic PaleoceanographyPt 2 Biological tracers and biomarkers edited by Hillaire-Marcel C and de Vernal A 263ndash326 Elsevier Amsterdamthe Netherlands 2007
Kaiho K Kajiwara Y Tazaki K Ueshima M Takeda NKawahata H Arinobu T Ishiwatari R Hirai A and Lam-olda M A Oceanic primary productivity and dissolved oxygenlevels at the CretaceousTertiary boundary their decrease sub-sequent warming and recovery Paleoceanography 14 511ndash5241999
Keller G and Lindinger M Stable isotope TOC and CaCO3record across the CretaceousTertiary boundary at El KefTunisia Palaeogeogr Palaeocl 73 243ndash265 1989
Kellough G R Paleoecology of the Foraminiferida of the WillsPoint Formation (Midway Group) in northeast Texas Trans GulfCoast Assoc Geol Soc 15 73ndash153 1965
Kring D A The Chicxulub impact event and its environmentalconsequences at the CretaceousndashTertiary boundary PalaeogeogrPalaeocl 255 4ndash21 2007
Kump L R Interpreting carbon-isotope excursions Strangeloveoceans Geology 19 299ndash302 1991
Machalski M Vellekoop J Dubicka Z Peryt D and Ha-rasimiuk M Late Maastrichtian cephalopods dinoflagellatecysts and foraminifera from the CretaceousndashPaleogene suc-cession at Lechoacutewka southeast Poland Stratigraphic andenvironmental implications Cretaceous Res 57 208ndash227doi101016jcretres201508012 2016
Molina E Evidence and causes of the main extinctionevents in the Paleogene based on extinction and survivalpatterns of foraminifera Earth-Sci Rev 140 166ndash181doi101016jearscirev201411008 2015
Molina E Alegret L Arenillas I and Arz J The GlobalBoundary Stratotype Section and Point for the base of the Da-nian Stage (Paleocene Paleogeneldquo Tertiaryrdquo Cenozoic) at ElKef Tunisia-Original definition and revision Episodes 29 263ndash273 2006
Olsson R K Liu C and van Fossen M The Cretaceous-Tertiarycatastrophic event at Millers Ferry Alabama in The Cretaceous-Tertiary Event and Other Catastrophes in Earth History editedby Ryder G Fastovsky D and Gartner S Geol S Am S307 263ndash277 1996
Peryt D Benthic foraminiferal response to the Cenomanian-Turonian and CretaceousndashPaleogene boundary events PrzegladGeol 52 827ndash832 2004
Peryt D Alegret L and Molina E The CretaceousPalaeogene(KP) boundary at Aiumln Settara Tunisia restructuring of benthicforaminiferal assemblages Terra Nova 14 101ndash107 2002
Ramette A Multivariate analyses in microbial ecology FEMS Mi-crobiol Ecol 62 142ndash160 2007
Rodriacuteguez-Tovar F J Uchman A MrsquoHamdi A Riahi Sand Ismail-Lattrache K B Ichnological record of palaeoen-vironment from the CretaceousndashPaleogene boundary intervalat El Kef Tunisia The first study of old and new sec-tions at the stratotype area J Afr Earth Sci 120 23ndash30doi101016jjafrearsci201604017 2016
Saner S Mudurnu-Goumlynuumlk Havzasının Jura ve Sonrasıccediloumlkelimnitelikleriyle paleocografya yorumlaması TJK Buumllteni 23 39ndash52 1980
Schioslashler P Brinkhuis H Roncaglia L and Wilson G J Di-noflagellate biostratigraphy and sequence stratigraphy of theType Maastrichtian (Upper Cretaceous) ENCI Quarry TheNetherlands Mar Micropaleontol 31 65ndash95 1997
Schueth J D Bralower T J Jiang S and Patzkowsky ME The role of regional survivor incumbency in the evolu-tionary recovery of calcareous nannoplankton from the Creta-ceousPaleogene (KPg) mass extinction Paleobiology 41 661ndash679 doi101017pab201528 2015
Scotese C R A continental drift flipbook J Geology 112 729ndash741 2004
Scotese C R and Dreher C GlobalGeology available at httpwwwGlobalGeologycom (last access 11 March 2014) 2012
Sepkoski J J Patterns of Phanerozoic extinction a perspectivefrom global data bases in Global Events and Event Stratigra-phy edited by Walliser O H Springer Berlin Germany 35ndash52 2016
Sibert E C Hull P M and Norris R D Resilience of Pacificpelagic fish across the CretaceousPalaeogene mass extinctionNat Geosci 7 667ndash670 doi101038NGEO2227 2014
Slimani H and Toufiq A A CretaceousndashPaleogene boundary geo-logical site revealed by planktic foraminifera and dinoflagellatecysts at Ouled Haddou eastern external Rif Chain Morocco JAfr Earth Sci 88 38ndash52 doi101016jjafrearsci2013080082013
Slimani H Louwye S and Toufiq A Dinoflagellate cystsform the CretaceousndashPaleogene boundary at Ouled Had-dou southeastern Rif Morocco biostratigraphy paleoenvi-ronments and paleobiogeography Palynology 34 90ndash124doi10108001916121003629933 2010
Sluijs A and Brinkhuis H A dynamic climate and ecosystemstate during the Paleocene-Eocene Thermal Maximum infer-ences from dinoflagellate cyst assemblages on the New JerseyShelf Biogeosciences 6 1755ndash1781 doi105194bg-6-1755-2009 2009
Smit J Extinction and evolution of planktonic foraminifera after amajor impact at the CretaceousTertiary boundary Geol S AmS 190 329ndash352 1982
Smit J and Romein A J T A sequence of events across theCretaceous-Tertiary boundary Earth Planet Sci Lett 74 155ndash170 1985
Speijer R P Extinction and recovery patterns in ben-thic foraminiferal paleocommunities across the Creta-ceousPaleogene and PaleoceneEocene boundaries GeologicaUltraiectina 124 Faculteit Aardwetenschappen PhD ThesisUniversiteit Utrecht Utrecht the Netherlands 1994
Speijer R P and van Der Zwaan G J Extinction and survivor-ship of southern Tethyan benthic foraminifera across the Cre-
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
900 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
taceousPaleogene boundary in Biotic recovery from mass ex-tinction events edited by Hart M B Geolog Soc Spec PublLondon UK 102 343ndash371 1996
Stemmann L and Boss E Plankton and Particle Size andPackaging From Determining Optical Properties to Drivingthe Biological Pump Annu Rev Mar Sci 4 263ndash290doi101146annurev-marine-120710-100853 2012
Thomas E Late Cretaceous-early Eocene mass extinctions in thedeep sea Geol S Am S 247 481ndash495 1990
Vandenberghe N Hilgen F J and Speijer R P The PaleogenePeriod in The Geologic Time Scale 2012 edited by GradsteinF M Ogg J G Schmitz M D and Ogg G M 2 855ndash921Elsevier Boston USA 2012
Vellekoop J Sluijs A Smit J Schouten S Weijers J WH Sinninghe Damsteacute J S and Brinkhuis H Rapid short-term cooling following the Chicxulub impact at the CretaceousndashPaleogene boundary P Natl Acad Sci USA 111 7537ndash7541doi101073pnas1319253111 2014
Vellekoop J Smit J van de Schootbrugge B Weijers J W HGaleotti S Sinninghe Damsteacute J S and Brinkhuis H Palyno-logical evidence for prolonged cooling along the Tunisian conti-nental shelf following the KndashPg boundary impact PalaeogeogrPalaeocl 426 216ndash228 doi101016jpalaeo2015030212015
Vellekoop J Esmeray-Senlet S Miller K G Browning J VSluijs A van de Schootbrugge B Sinninghe Damsteacute J Sand Brinkhuis H Evidence for CretaceousndashPaleogene bound-ary bolide ldquoimpact winterrdquo conditions from New Jersey USAGeology 44 619ndash622 doi101130G379611 2016
Woelders L and Speijer R Stable seafloor conditions sealevel and food supply during the latest Maastrichtian atBrazos River Texas Mar Micropaleontol 121 41ndash51doi101016jmarmicro201510002 2015
Zachos J C Arthur M A and Dean W E Geochemical evi-dence for suppression of pelagic marine productivity at the Cre-taceousTertiary boundary Nature 337 61ndash64 1989
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
- Abstract
- Introduction
- Geological setting and age assessment
- Materials and methods
-
- Sampling
- Foraminiferal analysis
- Palynological analysis
- Statistical analysis
-
- Results
-
- Benthic foraminifera
- Palynology
-
- Discussion
-
- Tethyan benthic foraminiferal turnover sequence
- Ecological responses to reduced export productivity
-
- Okccedilular section
- Western Tethys
- Global responses
-
- Long-term recovery
-
- Conclusions
- Data availability
- Author contributions
- Competing interests
- Acknowledgements
- References
-

898 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
Tertiary boundary at El Haria northwest Tunisia Mar Micropa-leontol 13 153ndash191 1988
Brinkhuis H Bujak J P Smit J Versteegh G J M and Viss-cher H Dinoflagellate-based sea surface temperature recon-structions across the Cretaceous-Tertiary boundary PalaeogeogrPalaeocl 141 67ndash83 1998
Buesseler K O The decoupling of production and particulate ex-port in the surface ocean Global Biogeochem Cy 12 297ndash3101998
Buzas M A Culver S J and Jorissen F J A statisti-cal evaluation of the microhabitats of living (stained) infau-nal benthic foraminifera Mar Micropaleontol 20 311ndash320doi1010160377-8398(93)90040-5 1993
Coccioni R and Galeotti S KndashT boundary extinction geolog-ically instantaneous or gradual event Evidence from deep-seabenthic foraminifera Geology 22 779ndash782 1994
Corliss B H Microhabitats of benthic foraminifera within deep-sea sediments Nature 314 435ndash438 doi101038314435a01985
Corliss B H and Chen C Morphotype patterns of Nor-wegian Sea deep-sea benthic foraminifera and ecologi-cal implications Geology 16 716ndash719 doi1011300091-7613(1988)016lt0716MPONSDgt23CO2 1988
Coxall H K DrsquoHondt S and Zachos J C Pelagic evolution andenvironmental recovery after the CretaceousndashPaleogene mass ex-tinction Geology 34 297ndash300 doi101130G217021 2006
Culver S J Benthic foraminifera across the CretaceousndashTertiary(KndashT) boundary a review Mar Micropaleontol 47 177ndash2262003
Cushman J A Upper Cretaceous foraminifera of the Gulf coastalregion of the United States and adjacent areas US Geolog SurvProf Pap 206 241 pp 1946
Cushman J A Paleocene foraminifera of the Gulf Coastal Regionof the United States and adjacent regions US Geol Surv ProfPap 232 75 pp 1951
DrsquoHondt S Consequences of the CretaceousPaleogene Mass Ex-tinction for Marine Ecosystems Annu Rev Ecol Evol S36 295ndash317 doi101146annurevecolsys350211031057152005
DrsquoHondt S and Zachos J C Cretaceous foraminifera and the evo-lutionary history of planktic photosymbiosis Paleobiology 24512ndash523 1998
DrsquoHondt S Donaghay P Zachos J C Luttenberg D andLindinger M Organic Carbon Fluxes and Ecological Recov-ery from the Cretaceous-Tertiary Mass Extinction Science 282276ndash279 1998
Donze P Colin J-P Damotte R Oertli HJ Peypouquet J-Pand Said R Les ostracodes du Campanien terminal agrave lrsquoEocegraveneinfeacuterieur de la coupe du Kef Tuneacutesie nord-occidentale Bull desCentr Recher Expl-Prod Elf Aquit 6 273ndash335 1982
Dortch Q and Packard T T Differences in biomass structure be-tween oligotrophic and eutrophic marine ecosystems Deep-SeaRes 36 223ndash240 1989
Ernst S R Guasti E Dupuis C and Speijer R P En-vironmental perturbation in the southern Tethys across thePaleoceneEocene boundary (Dababiya Egypt) Foraminiferaland clay mineral records Mar Micropaleontol 60 89ndash111doi101016jmarmicro200603002 2006
Eshet Y Moshkovitz S Habib D Benjamini C and Maga-ritz M Calcareous nannofossil and dinoflagellate stratigraphyacross the CretaceousTertiary boundary at Hor Hahar IsraelMar Micropaleontol 18 199ndash228 1992
Eshet Y Almogi-Labin A and Bein A Dinoflagellate cysts pa-leoproductivity and upwelling systems A Late Cretaceous ex-ample from Israel Mar Micropaleontol 23 231ndash240 1994
Esmeray-Senlet S Wright J D Olsson R K MillerK G Browning J V and Quan T M Evidencefor reduced export productivity following the Creta-ceousPaleogene mass extinction Paleoceanography 30718ndash738 doi1010022014PA002724 2015
Giusberti L Boscolo Galazzo F and Thomas E Variability inclimate and productivity during the Paleocene-Eocene ThermalMaximum in the western Tethys (Forada section) Clim Past12 213ndash240 doi105194cp-12-213-2016 2016
Guasti E Kouwenhoven T J Brinkhuis H and Speijer R PPaleocene sea-level and productivity changes at the southernTethyan margin (El Kef Tunisia) Mar Micropaleontol 55 1ndash17 2005
Habib D and Saeedi F The Manumiella seelandica global spikeCooling during regression at the close of the MaastrichtianPalaeogeogr Palaeocl 255 87ndash97 2007
Hain M P Sigmal D M and Haug G H 818 ndash The biolog-ical Pump in the Past in Reference Module in Earth Systemsand Environmental Sciences Treatise on Geochemistry (SecondEdition) The Oceans and Marine Geochemistry 8 485ndash517doi101016B978-0-08-095975-700618-5 2014
Hayek L C and Buzas M A On the proper and efficient useof diversity measures with individual field samples J ForaminRes 43 305ndash313 2013
Henson S A Sanders R Madsen E Morris P J Le MoigneF and Quartly G D A reduced estimate of the strength ofthe oceanrsquos biological carbon pump Geophys Res Lett 38L04606 doi1010292011GL046735 2011
Hilting A K Kump L R and Bralower T J Variations in theoceanic vertical carbon isotope gradient and their implicationsfor the Paleocene-Eocene biological pump Paleoceanography23 PA3222 doi1010292007PA001458 2008
Honjo S Manganini S J Krishfield R A and Francois RParticulate organic carbon fluxes to the ocean interior and fac-tors controlling the biological pump A synthesis of global sed-iment trap progams since 1983 Prog Oceanogr 76 217ndash285doi101016jpocean200711003 2008
Huber B T MacLeod K G and Norris R D Abrupt extinctionand subsequent reworking of Cretaceous planktonic foraminiferaacross the Cretaceous-Tertiary boundary Evidence from the sub-tropical North Atlantic in Catastrophic Events and Mass Extinc-tions Impacts and Beyond edited by Koeberl C and MacLeodK G Geol S Am S 356 277ndash289 Boulder Colorado USA2002
Hull P M Norris R D Bralower T J and Schueth J D A rolefor chance in marine recovery from the end-Cretaceous extinc-tion Nat Geosci 4 856ndash860 doi101038ngeo1302 2011
Hsuuml K J and McKenzie J A A ldquoStrangeloverdquo ocean in the earli-est Tertiary in Natural variations Archean to present edited bySundquist E T and Broecker W S The carbon cycle and atmo-spheric CO2 American Geophysical Union Washington DCUSA 487ndash492 1985
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
J Vellekoop et al Ecological response to KndashPg boundary mass extinction 899
Jiao N Herndl G J Hansell D A Benner R Kattner G Wil-helm S W Kirchman D L Weinbauer M G Luo T ChenF and Azam F Microbial production of recalcitrant dissolvedorganic matter long-term carbon storage in the global oceanNat Rev 8 593ndash599 doi101038nrmicro2386 2010
Jorissen F J de Stigter H C and Widmark J G V A concep-tual model explaining benthic foraminiferal microhabitats MarMicropaleontol 26 3ndash15 doi1010160377-8398(95)00047-X1995
Jorissen F J Fontanier C and Thomas E Paleoceanographi-cal proxies based on deep-sea benthic foraminiferal assemblagecharacteristics in Proxies in Late Cenozoic PaleoceanographyPt 2 Biological tracers and biomarkers edited by Hillaire-Marcel C and de Vernal A 263ndash326 Elsevier Amsterdamthe Netherlands 2007
Kaiho K Kajiwara Y Tazaki K Ueshima M Takeda NKawahata H Arinobu T Ishiwatari R Hirai A and Lam-olda M A Oceanic primary productivity and dissolved oxygenlevels at the CretaceousTertiary boundary their decrease sub-sequent warming and recovery Paleoceanography 14 511ndash5241999
Keller G and Lindinger M Stable isotope TOC and CaCO3record across the CretaceousTertiary boundary at El KefTunisia Palaeogeogr Palaeocl 73 243ndash265 1989
Kellough G R Paleoecology of the Foraminiferida of the WillsPoint Formation (Midway Group) in northeast Texas Trans GulfCoast Assoc Geol Soc 15 73ndash153 1965
Kring D A The Chicxulub impact event and its environmentalconsequences at the CretaceousndashTertiary boundary PalaeogeogrPalaeocl 255 4ndash21 2007
Kump L R Interpreting carbon-isotope excursions Strangeloveoceans Geology 19 299ndash302 1991
Machalski M Vellekoop J Dubicka Z Peryt D and Ha-rasimiuk M Late Maastrichtian cephalopods dinoflagellatecysts and foraminifera from the CretaceousndashPaleogene suc-cession at Lechoacutewka southeast Poland Stratigraphic andenvironmental implications Cretaceous Res 57 208ndash227doi101016jcretres201508012 2016
Molina E Evidence and causes of the main extinctionevents in the Paleogene based on extinction and survivalpatterns of foraminifera Earth-Sci Rev 140 166ndash181doi101016jearscirev201411008 2015
Molina E Alegret L Arenillas I and Arz J The GlobalBoundary Stratotype Section and Point for the base of the Da-nian Stage (Paleocene Paleogeneldquo Tertiaryrdquo Cenozoic) at ElKef Tunisia-Original definition and revision Episodes 29 263ndash273 2006
Olsson R K Liu C and van Fossen M The Cretaceous-Tertiarycatastrophic event at Millers Ferry Alabama in The Cretaceous-Tertiary Event and Other Catastrophes in Earth History editedby Ryder G Fastovsky D and Gartner S Geol S Am S307 263ndash277 1996
Peryt D Benthic foraminiferal response to the Cenomanian-Turonian and CretaceousndashPaleogene boundary events PrzegladGeol 52 827ndash832 2004
Peryt D Alegret L and Molina E The CretaceousPalaeogene(KP) boundary at Aiumln Settara Tunisia restructuring of benthicforaminiferal assemblages Terra Nova 14 101ndash107 2002
Ramette A Multivariate analyses in microbial ecology FEMS Mi-crobiol Ecol 62 142ndash160 2007
Rodriacuteguez-Tovar F J Uchman A MrsquoHamdi A Riahi Sand Ismail-Lattrache K B Ichnological record of palaeoen-vironment from the CretaceousndashPaleogene boundary intervalat El Kef Tunisia The first study of old and new sec-tions at the stratotype area J Afr Earth Sci 120 23ndash30doi101016jjafrearsci201604017 2016
Saner S Mudurnu-Goumlynuumlk Havzasının Jura ve Sonrasıccediloumlkelimnitelikleriyle paleocografya yorumlaması TJK Buumllteni 23 39ndash52 1980
Schioslashler P Brinkhuis H Roncaglia L and Wilson G J Di-noflagellate biostratigraphy and sequence stratigraphy of theType Maastrichtian (Upper Cretaceous) ENCI Quarry TheNetherlands Mar Micropaleontol 31 65ndash95 1997
Schueth J D Bralower T J Jiang S and Patzkowsky ME The role of regional survivor incumbency in the evolu-tionary recovery of calcareous nannoplankton from the Creta-ceousPaleogene (KPg) mass extinction Paleobiology 41 661ndash679 doi101017pab201528 2015
Scotese C R A continental drift flipbook J Geology 112 729ndash741 2004
Scotese C R and Dreher C GlobalGeology available at httpwwwGlobalGeologycom (last access 11 March 2014) 2012
Sepkoski J J Patterns of Phanerozoic extinction a perspectivefrom global data bases in Global Events and Event Stratigra-phy edited by Walliser O H Springer Berlin Germany 35ndash52 2016
Sibert E C Hull P M and Norris R D Resilience of Pacificpelagic fish across the CretaceousPalaeogene mass extinctionNat Geosci 7 667ndash670 doi101038NGEO2227 2014
Slimani H and Toufiq A A CretaceousndashPaleogene boundary geo-logical site revealed by planktic foraminifera and dinoflagellatecysts at Ouled Haddou eastern external Rif Chain Morocco JAfr Earth Sci 88 38ndash52 doi101016jjafrearsci2013080082013
Slimani H Louwye S and Toufiq A Dinoflagellate cystsform the CretaceousndashPaleogene boundary at Ouled Had-dou southeastern Rif Morocco biostratigraphy paleoenvi-ronments and paleobiogeography Palynology 34 90ndash124doi10108001916121003629933 2010
Sluijs A and Brinkhuis H A dynamic climate and ecosystemstate during the Paleocene-Eocene Thermal Maximum infer-ences from dinoflagellate cyst assemblages on the New JerseyShelf Biogeosciences 6 1755ndash1781 doi105194bg-6-1755-2009 2009
Smit J Extinction and evolution of planktonic foraminifera after amajor impact at the CretaceousTertiary boundary Geol S AmS 190 329ndash352 1982
Smit J and Romein A J T A sequence of events across theCretaceous-Tertiary boundary Earth Planet Sci Lett 74 155ndash170 1985
Speijer R P Extinction and recovery patterns in ben-thic foraminiferal paleocommunities across the Creta-ceousPaleogene and PaleoceneEocene boundaries GeologicaUltraiectina 124 Faculteit Aardwetenschappen PhD ThesisUniversiteit Utrecht Utrecht the Netherlands 1994
Speijer R P and van Der Zwaan G J Extinction and survivor-ship of southern Tethyan benthic foraminifera across the Cre-
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
900 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
taceousPaleogene boundary in Biotic recovery from mass ex-tinction events edited by Hart M B Geolog Soc Spec PublLondon UK 102 343ndash371 1996
Stemmann L and Boss E Plankton and Particle Size andPackaging From Determining Optical Properties to Drivingthe Biological Pump Annu Rev Mar Sci 4 263ndash290doi101146annurev-marine-120710-100853 2012
Thomas E Late Cretaceous-early Eocene mass extinctions in thedeep sea Geol S Am S 247 481ndash495 1990
Vandenberghe N Hilgen F J and Speijer R P The PaleogenePeriod in The Geologic Time Scale 2012 edited by GradsteinF M Ogg J G Schmitz M D and Ogg G M 2 855ndash921Elsevier Boston USA 2012
Vellekoop J Sluijs A Smit J Schouten S Weijers J WH Sinninghe Damsteacute J S and Brinkhuis H Rapid short-term cooling following the Chicxulub impact at the CretaceousndashPaleogene boundary P Natl Acad Sci USA 111 7537ndash7541doi101073pnas1319253111 2014
Vellekoop J Smit J van de Schootbrugge B Weijers J W HGaleotti S Sinninghe Damsteacute J S and Brinkhuis H Palyno-logical evidence for prolonged cooling along the Tunisian conti-nental shelf following the KndashPg boundary impact PalaeogeogrPalaeocl 426 216ndash228 doi101016jpalaeo2015030212015
Vellekoop J Esmeray-Senlet S Miller K G Browning J VSluijs A van de Schootbrugge B Sinninghe Damsteacute J Sand Brinkhuis H Evidence for CretaceousndashPaleogene bound-ary bolide ldquoimpact winterrdquo conditions from New Jersey USAGeology 44 619ndash622 doi101130G379611 2016
Woelders L and Speijer R Stable seafloor conditions sealevel and food supply during the latest Maastrichtian atBrazos River Texas Mar Micropaleontol 121 41ndash51doi101016jmarmicro201510002 2015
Zachos J C Arthur M A and Dean W E Geochemical evi-dence for suppression of pelagic marine productivity at the Cre-taceousTertiary boundary Nature 337 61ndash64 1989
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
- Abstract
- Introduction
- Geological setting and age assessment
- Materials and methods
-
- Sampling
- Foraminiferal analysis
- Palynological analysis
- Statistical analysis
-
- Results
-
- Benthic foraminifera
- Palynology
-
- Discussion
-
- Tethyan benthic foraminiferal turnover sequence
- Ecological responses to reduced export productivity
-
- Okccedilular section
- Western Tethys
- Global responses
-
- Long-term recovery
-
- Conclusions
- Data availability
- Author contributions
- Competing interests
- Acknowledgements
- References
-

J Vellekoop et al Ecological response to KndashPg boundary mass extinction 899
Jiao N Herndl G J Hansell D A Benner R Kattner G Wil-helm S W Kirchman D L Weinbauer M G Luo T ChenF and Azam F Microbial production of recalcitrant dissolvedorganic matter long-term carbon storage in the global oceanNat Rev 8 593ndash599 doi101038nrmicro2386 2010
Jorissen F J de Stigter H C and Widmark J G V A concep-tual model explaining benthic foraminiferal microhabitats MarMicropaleontol 26 3ndash15 doi1010160377-8398(95)00047-X1995
Jorissen F J Fontanier C and Thomas E Paleoceanographi-cal proxies based on deep-sea benthic foraminiferal assemblagecharacteristics in Proxies in Late Cenozoic PaleoceanographyPt 2 Biological tracers and biomarkers edited by Hillaire-Marcel C and de Vernal A 263ndash326 Elsevier Amsterdamthe Netherlands 2007
Kaiho K Kajiwara Y Tazaki K Ueshima M Takeda NKawahata H Arinobu T Ishiwatari R Hirai A and Lam-olda M A Oceanic primary productivity and dissolved oxygenlevels at the CretaceousTertiary boundary their decrease sub-sequent warming and recovery Paleoceanography 14 511ndash5241999
Keller G and Lindinger M Stable isotope TOC and CaCO3record across the CretaceousTertiary boundary at El KefTunisia Palaeogeogr Palaeocl 73 243ndash265 1989
Kellough G R Paleoecology of the Foraminiferida of the WillsPoint Formation (Midway Group) in northeast Texas Trans GulfCoast Assoc Geol Soc 15 73ndash153 1965
Kring D A The Chicxulub impact event and its environmentalconsequences at the CretaceousndashTertiary boundary PalaeogeogrPalaeocl 255 4ndash21 2007
Kump L R Interpreting carbon-isotope excursions Strangeloveoceans Geology 19 299ndash302 1991
Machalski M Vellekoop J Dubicka Z Peryt D and Ha-rasimiuk M Late Maastrichtian cephalopods dinoflagellatecysts and foraminifera from the CretaceousndashPaleogene suc-cession at Lechoacutewka southeast Poland Stratigraphic andenvironmental implications Cretaceous Res 57 208ndash227doi101016jcretres201508012 2016
Molina E Evidence and causes of the main extinctionevents in the Paleogene based on extinction and survivalpatterns of foraminifera Earth-Sci Rev 140 166ndash181doi101016jearscirev201411008 2015
Molina E Alegret L Arenillas I and Arz J The GlobalBoundary Stratotype Section and Point for the base of the Da-nian Stage (Paleocene Paleogeneldquo Tertiaryrdquo Cenozoic) at ElKef Tunisia-Original definition and revision Episodes 29 263ndash273 2006
Olsson R K Liu C and van Fossen M The Cretaceous-Tertiarycatastrophic event at Millers Ferry Alabama in The Cretaceous-Tertiary Event and Other Catastrophes in Earth History editedby Ryder G Fastovsky D and Gartner S Geol S Am S307 263ndash277 1996
Peryt D Benthic foraminiferal response to the Cenomanian-Turonian and CretaceousndashPaleogene boundary events PrzegladGeol 52 827ndash832 2004
Peryt D Alegret L and Molina E The CretaceousPalaeogene(KP) boundary at Aiumln Settara Tunisia restructuring of benthicforaminiferal assemblages Terra Nova 14 101ndash107 2002
Ramette A Multivariate analyses in microbial ecology FEMS Mi-crobiol Ecol 62 142ndash160 2007
Rodriacuteguez-Tovar F J Uchman A MrsquoHamdi A Riahi Sand Ismail-Lattrache K B Ichnological record of palaeoen-vironment from the CretaceousndashPaleogene boundary intervalat El Kef Tunisia The first study of old and new sec-tions at the stratotype area J Afr Earth Sci 120 23ndash30doi101016jjafrearsci201604017 2016
Saner S Mudurnu-Goumlynuumlk Havzasının Jura ve Sonrasıccediloumlkelimnitelikleriyle paleocografya yorumlaması TJK Buumllteni 23 39ndash52 1980
Schioslashler P Brinkhuis H Roncaglia L and Wilson G J Di-noflagellate biostratigraphy and sequence stratigraphy of theType Maastrichtian (Upper Cretaceous) ENCI Quarry TheNetherlands Mar Micropaleontol 31 65ndash95 1997
Schueth J D Bralower T J Jiang S and Patzkowsky ME The role of regional survivor incumbency in the evolu-tionary recovery of calcareous nannoplankton from the Creta-ceousPaleogene (KPg) mass extinction Paleobiology 41 661ndash679 doi101017pab201528 2015
Scotese C R A continental drift flipbook J Geology 112 729ndash741 2004
Scotese C R and Dreher C GlobalGeology available at httpwwwGlobalGeologycom (last access 11 March 2014) 2012
Sepkoski J J Patterns of Phanerozoic extinction a perspectivefrom global data bases in Global Events and Event Stratigra-phy edited by Walliser O H Springer Berlin Germany 35ndash52 2016
Sibert E C Hull P M and Norris R D Resilience of Pacificpelagic fish across the CretaceousPalaeogene mass extinctionNat Geosci 7 667ndash670 doi101038NGEO2227 2014
Slimani H and Toufiq A A CretaceousndashPaleogene boundary geo-logical site revealed by planktic foraminifera and dinoflagellatecysts at Ouled Haddou eastern external Rif Chain Morocco JAfr Earth Sci 88 38ndash52 doi101016jjafrearsci2013080082013
Slimani H Louwye S and Toufiq A Dinoflagellate cystsform the CretaceousndashPaleogene boundary at Ouled Had-dou southeastern Rif Morocco biostratigraphy paleoenvi-ronments and paleobiogeography Palynology 34 90ndash124doi10108001916121003629933 2010
Sluijs A and Brinkhuis H A dynamic climate and ecosystemstate during the Paleocene-Eocene Thermal Maximum infer-ences from dinoflagellate cyst assemblages on the New JerseyShelf Biogeosciences 6 1755ndash1781 doi105194bg-6-1755-2009 2009
Smit J Extinction and evolution of planktonic foraminifera after amajor impact at the CretaceousTertiary boundary Geol S AmS 190 329ndash352 1982
Smit J and Romein A J T A sequence of events across theCretaceous-Tertiary boundary Earth Planet Sci Lett 74 155ndash170 1985
Speijer R P Extinction and recovery patterns in ben-thic foraminiferal paleocommunities across the Creta-ceousPaleogene and PaleoceneEocene boundaries GeologicaUltraiectina 124 Faculteit Aardwetenschappen PhD ThesisUniversiteit Utrecht Utrecht the Netherlands 1994
Speijer R P and van Der Zwaan G J Extinction and survivor-ship of southern Tethyan benthic foraminifera across the Cre-
wwwbiogeosciencesnet148852017 Biogeosciences 14 885ndash900 2017
900 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
taceousPaleogene boundary in Biotic recovery from mass ex-tinction events edited by Hart M B Geolog Soc Spec PublLondon UK 102 343ndash371 1996
Stemmann L and Boss E Plankton and Particle Size andPackaging From Determining Optical Properties to Drivingthe Biological Pump Annu Rev Mar Sci 4 263ndash290doi101146annurev-marine-120710-100853 2012
Thomas E Late Cretaceous-early Eocene mass extinctions in thedeep sea Geol S Am S 247 481ndash495 1990
Vandenberghe N Hilgen F J and Speijer R P The PaleogenePeriod in The Geologic Time Scale 2012 edited by GradsteinF M Ogg J G Schmitz M D and Ogg G M 2 855ndash921Elsevier Boston USA 2012
Vellekoop J Sluijs A Smit J Schouten S Weijers J WH Sinninghe Damsteacute J S and Brinkhuis H Rapid short-term cooling following the Chicxulub impact at the CretaceousndashPaleogene boundary P Natl Acad Sci USA 111 7537ndash7541doi101073pnas1319253111 2014
Vellekoop J Smit J van de Schootbrugge B Weijers J W HGaleotti S Sinninghe Damsteacute J S and Brinkhuis H Palyno-logical evidence for prolonged cooling along the Tunisian conti-nental shelf following the KndashPg boundary impact PalaeogeogrPalaeocl 426 216ndash228 doi101016jpalaeo2015030212015
Vellekoop J Esmeray-Senlet S Miller K G Browning J VSluijs A van de Schootbrugge B Sinninghe Damsteacute J Sand Brinkhuis H Evidence for CretaceousndashPaleogene bound-ary bolide ldquoimpact winterrdquo conditions from New Jersey USAGeology 44 619ndash622 doi101130G379611 2016
Woelders L and Speijer R Stable seafloor conditions sealevel and food supply during the latest Maastrichtian atBrazos River Texas Mar Micropaleontol 121 41ndash51doi101016jmarmicro201510002 2015
Zachos J C Arthur M A and Dean W E Geochemical evi-dence for suppression of pelagic marine productivity at the Cre-taceousTertiary boundary Nature 337 61ndash64 1989
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
- Abstract
- Introduction
- Geological setting and age assessment
- Materials and methods
-
- Sampling
- Foraminiferal analysis
- Palynological analysis
- Statistical analysis
-
- Results
-
- Benthic foraminifera
- Palynology
-
- Discussion
-
- Tethyan benthic foraminiferal turnover sequence
- Ecological responses to reduced export productivity
-
- Okccedilular section
- Western Tethys
- Global responses
-
- Long-term recovery
-
- Conclusions
- Data availability
- Author contributions
- Competing interests
- Acknowledgements
- References
-

900 J Vellekoop et al Ecological response to KndashPg boundary mass extinction
taceousPaleogene boundary in Biotic recovery from mass ex-tinction events edited by Hart M B Geolog Soc Spec PublLondon UK 102 343ndash371 1996
Stemmann L and Boss E Plankton and Particle Size andPackaging From Determining Optical Properties to Drivingthe Biological Pump Annu Rev Mar Sci 4 263ndash290doi101146annurev-marine-120710-100853 2012
Thomas E Late Cretaceous-early Eocene mass extinctions in thedeep sea Geol S Am S 247 481ndash495 1990
Vandenberghe N Hilgen F J and Speijer R P The PaleogenePeriod in The Geologic Time Scale 2012 edited by GradsteinF M Ogg J G Schmitz M D and Ogg G M 2 855ndash921Elsevier Boston USA 2012
Vellekoop J Sluijs A Smit J Schouten S Weijers J WH Sinninghe Damsteacute J S and Brinkhuis H Rapid short-term cooling following the Chicxulub impact at the CretaceousndashPaleogene boundary P Natl Acad Sci USA 111 7537ndash7541doi101073pnas1319253111 2014
Vellekoop J Smit J van de Schootbrugge B Weijers J W HGaleotti S Sinninghe Damsteacute J S and Brinkhuis H Palyno-logical evidence for prolonged cooling along the Tunisian conti-nental shelf following the KndashPg boundary impact PalaeogeogrPalaeocl 426 216ndash228 doi101016jpalaeo2015030212015
Vellekoop J Esmeray-Senlet S Miller K G Browning J VSluijs A van de Schootbrugge B Sinninghe Damsteacute J Sand Brinkhuis H Evidence for CretaceousndashPaleogene bound-ary bolide ldquoimpact winterrdquo conditions from New Jersey USAGeology 44 619ndash622 doi101130G379611 2016
Woelders L and Speijer R Stable seafloor conditions sealevel and food supply during the latest Maastrichtian atBrazos River Texas Mar Micropaleontol 121 41ndash51doi101016jmarmicro201510002 2015
Zachos J C Arthur M A and Dean W E Geochemical evi-dence for suppression of pelagic marine productivity at the Cre-taceousTertiary boundary Nature 337 61ndash64 1989
Biogeosciences 14 885ndash900 2017 wwwbiogeosciencesnet148852017
- Abstract
- Introduction
- Geological setting and age assessment
- Materials and methods
-
- Sampling
- Foraminiferal analysis
- Palynological analysis
- Statistical analysis
-
- Results
-
- Benthic foraminifera
- Palynology
-
- Discussion
-
- Tethyan benthic foraminiferal turnover sequence
- Ecological responses to reduced export productivity
-
- Okccedilular section
- Western Tethys
- Global responses
-
- Long-term recovery
-
- Conclusions
- Data availability
- Author contributions
- Competing interests
- Acknowledgements
- References
-


















