
Draft - University of Toronto T-Space 1 1 Survey of compound microsatellites in multiple...
17
Draft Survey of compound microsatellites in multiple Lactobacillus genomes Journal: Canadian Journal of Microbiology Manuscript ID cjm-2015-0136.R2 Manuscript Type: Article Date Submitted by the Author: 25-Aug-2015 Complete List of Authors: Basharat, Zarrin; Fatima Jinnah Women University, Yasmin, Azra; Fatima Jinnah Women University Keyword: Lactobacillus, compound microsatellite, data mining, in silico https://mc06.manuscriptcentral.com/cjm-pubs Canadian Journal of Microbiology
-
Upload
hoangthuan -
Category
Documents
-
view
218 -
download
5
Transcript of Draft - University of Toronto T-Space 1 1 Survey of compound microsatellites in multiple...

Draft
Survey of compound microsatellites in multiple
Lactobacillus genomes
Journal: Canadian Journal of Microbiology
Manuscript ID cjm-2015-0136.R2
Manuscript Type: Article
Date Submitted by the Author: 25-Aug-2015
Complete List of Authors: Basharat, Zarrin; Fatima Jinnah Women University, Yasmin, Azra; Fatima Jinnah Women University
Keyword: Lactobacillus, compound microsatellite, data mining, in silico
https://mc06.manuscriptcentral.com/cjm-pubs
Canadian Journal of Microbiology

Draft
1
Survey of compound microsatellites in multiple Lactobacillus genomes 1
Zarrin Basharat and Azra Yasmin* 2
Corresponding author email: [email protected] 3
Microbiology & Biotechnology Research Laboratory, Department of Environmental 4
Sciences, Fatima Jinnah Women University, 46000, Pakistan 5
6
7
8
9
10
11
12
13
14
15
Running title: Compound microsatellites in Lactobacillus genomes 16
17
18
Page 1 of 16
https://mc06.manuscriptcentral.com/cjm-pubs
Canadian Journal of Microbiology

Draft
2
Abstract 19
Distinct simple sequence repeats with two or more individual microsatellites joined together 20
or lying adjacent to each other are identified as compound microsatellites. Investigation of 21
such composite microsatellites in the genomes of genus Lactobacillus was the aim of this 22
study. In silico inspection of microsatellite clustering in genomes of fourteen Lactobacillus 23
species revealed a wealth of compound microsatellites. All of the mined compound 24
microsatellites were imperfect, composed of variant motifs and increased in all genomes with 25
increment of dMAX value from 10 to 50. A majority of these repeats were present in the 26
coding regions. Correlation of microsatellite to compound microsatellite density was 27
detected. The difference established in compound microsatellite division between eukaryotes, 28
E. coli and Lactobacilli is suggestive of diverse genomic features and elementary distinction 29
between creation and fixation methods of compound microsatellites among these organisms. . 30
Keywords: Lactobacillus, compound microsatellite, data mining, in silico 31
32
33
34
35
36
37
38
Page 2 of 16
https://mc06.manuscriptcentral.com/cjm-pubs
Canadian Journal of Microbiology

Draft
3
39
Introduction 40
Microsatellite polymorphism has been studied for preliminary typing of lactic acid bacteria 41
(Chebeňová et al. 2010) suggesting presence of compound microsatellites. Compound 42
microsatellites are composed of two or more adjacent individual microsatellites m1-xn-m2 43
termed as ‘2-microsatellite’compound and m1-xn-m2-xt-m3 as ‘3-micro-satellite’ compound. 44
Analysis and characterization of these microsatellites is important for interpretation of the 45
origin, rendering of mutational processes, and comprehending structure of these partly 46
understood sequences. Compound microsatellites have been reported in diverse taxa across 47
viruses, prokaryotes and eukaryotes (Gur-Arie et al. 2000; Chen et al. 2012; George et al. 48
2014), constituting approximately 10% of simple sequence repeats (SSRs) in human genome 49
(Weber 1990), including highly polymorphic compound repeats such as (dC-dA)n(dG-dT)n 50
(Bull et al. 1999). Kofler et al. (2008) carried out in silico examination of microsatellite 51
grouping in genomes of several eukaryotes, with compound microsatellites ranging from 4–52
25% of all microsatellites. 53
Alleles of compound microsatellite loci are capable of variation in end to end distance of one 54
or both repeats (Urquhart et al. 1994; Garza and Freimer, 1996; Brinkmann et al. 1998). 55
Length of the repeat motifs in compound microsatellites are likely to affect the structure of 56
the microsatellite. Knowledge about compound microsatellites may possibly offer valuable 57
information concerning the evolution and imperfection of microsatellites (Kofler et al. 2008). 58
Sequencing can expose disparities between the matching length alleles in compound 59
microsatellites (Bull et al. 1999), as with imperfect ones (Laura et al. 1999). Owing to the 60
sequencing of numerous genomes and the accessibility of sequence data, microsatellites can 61
be explored and analyzed in silico. Only miniscule amount of compound microsatellites in 62
Page 3 of 16
https://mc06.manuscriptcentral.com/cjm-pubs
Canadian Journal of Microbiology

Draft
4
sequenced genomes have been explored and a plethora of genomes need to be explored for 63
this category of microsatellites. Comparative data analysis can thus, be utilized to shed light 64
on microsatellite and genome variation in different organisms. 65
Compound microsatellites have been investigated previously in Escherichia coli whole 66
genomes (Chen et al. 2011) and used as a model for this study. Fourteen bacterial species in 67
the genus Lactobacillus were screened in both coding and non-coding regions for possible 68
occurrence of compound microsatellites. Analysis of compound microsatellite complexity 69
was also carried out. This study can aid in understanding the intricate discrepancies 70
concerning compound simple sequence repeats (cSSRs) in various organisms and contribute 71
to the biological comprehension of this category of microsatellites in bacteria. 72
2 Material and Methods 73
2.1 Genome sequences 74
In the present study, we analyzed compound microsatellites in fourteen whole genomes of 75
Lactobacillus, ranging from 1,827,111 bp in Lactobacillus salivarius UCC118 (Accession 76
ID: NC_007929.1) to 3,308,274 bp in Lactobacillus plantarum WCFS1 (Accession ID: 77
NC_004567.1). All studied genomes are listed in ‘Genome-level Extraction Mode’ of IMEx 78
(Mudunuri and Nagarajaram 2007). The accession numbers are given in Table 1 for explored 79
whole genomes of bacterial species.. 80
2.2 Data mining and analysis 81
Compound microsatellites were mined using IMEx (Mudunuri and Nagarajaram 2007) with 82
the following parametres for Lactobacillus genomes: Include Flanking regions: 10 bp, Type 83
of Repeat: imperfect; Repeat Size: all; Minimum Repeat Number: 12, 6, 4, 3, 3, 3 , 84
Page 4 of 16
https://mc06.manuscriptcentral.com/cjm-pubs
Canadian Journal of Microbiology

Draft
5
Imperfection Limit/repeat unit: 1, 1, 1, 2, 2, 3, Percent Imperfection in Repeat Tract: 10%, 85
Maximum distance allowed between any two adjacent SSRs forming a cSSR (bp) (i-e 86
dMAX): 10 with complete standardization. The obtained results were then analyzed. Previous 87
studies focused on microsatellites with varying lengths such as 15 bp for eukaryotes with 88
mismatch (Kofler et al. 2008), 12 bp for eukaryotes (Toth et al. 2000; Buschiazzo E and 89
Gemmell NJ 2010) and prokaryotes with mismatches (Chen et al. 2011). Comparison to any 90
datasets having varying minimum repeat number is not valid if the value of the minimum 91
repeat number is not similar, so the authors have only used 12 bp for comparison of bacteria 92
with previously studied E. coli genomes. 93
2.3 Statistical analysis 94
All the statistical analysis was performed using IBM SPSS v22. Linear regression (R2) was 95
calculated to evaluate the influence of genome size and GC content on the SSRs and cSSRs 96
and correlation among SSR and cSSR density. A P-value of <0.05 was considered to be 97
significant. 98
3 Results and Discussion 99
Prokaryotes and eukaryote genomes exhibit variability in the composition of compound 100
microsatellites. Compound microsatellites are putatively involved in gene expression 101
regulation and functional dictation of proteins in a number of species (Kashi and King, 2006; 102
Chen et al. 2011). Though their significance in Lactobacillus is yet to be proven 103
experimentally, possible complex regulation at the functional level is suggested. Assessment 104
of cSSRs can prove to be of aid in understanding the incidence, distribution and complexity 105
of compound microsatellites in Lactobacillus microbial genomes. 106
3.1 Number, location and genesis of compound microsatellites 107
Page 5 of 16
https://mc06.manuscriptcentral.com/cjm-pubs
Canadian Journal of Microbiology

Draft
6
Analysis revealed 21,974 simple sequence microsatellites and 285 compound microsatellites 108
in the whole assessed dataset of Lactobacillus species (Table 1). Microsatellite and 109
compound microsatellites were normalized (SSR/cSSR density= number of repeats/Mb of 110
genome) over studied genomes of varying sizes for comparison. Density of microsatellites 111
showed a lot of variation and ranged from 506.7-1040.9 (Mean=747.2, SD=156.4) while 112
density of compound microsatellites ranged from 3.3-16.4 (Mean=10.0, SD=4.7). 113
Lactobacillus salivarius had the highest SSR (1040.9) and cSSR (16.4) density. Lowest SSR 114
density (506.7) was observed in Lactobacillus brevis ATCC 367, while lowest cSSR density 115
was observed in Lactobacillus casei ATCC 334 (3.3). Correlation analysis between GC 116
content and genome size with cSSR density revealed a weak correlation (R2=0.34, P<0.05 117
and R2=0.39, P<0.05). cSSR density was strongly correlated to SSR density (R2=0.79, 118
P<0.01) which varies considerably from the previously analysed E.coli genomes (R2= 0.214, 119
P < 0.01). This suggests that the association of SSR density with the cSSR density may not be 120
dependent on DNA polymerases/ replication method as put forward by Chen et al. (2011), 121
while explaining differences in eukaryotic and prokaryotic cSSR density influenced by SSR 122
distribution in the genomes. Prokaryotes share the same rolling circle replication method, so a 123
difference in our results and those reported by Chen et al. (2011) suggest that association of 124
the densities of SSRs and cSSRs is dependent on species and recombination (presence of 125
certain type of repeat motifs) (Kofler et al. 2008) instead of DNA replication. Remarkably 126
analogous SSR and cSSR distribution was observed among similar species of surveyed 127
Lactobacillus genomes (Lactobacillus reuteri DSM 20016, Lactobacillus reuteri JCM 1112 128
and Lactobacillus delbrueckii subsp. bulgaricus ATCC 11842, Lactobacillus delbrueckii 129
subsp. bulgaricusATCC BAA-365). 130
The majority of compound microsatellites (220/285) existed in coding regions 131
(Supplementary Table 1). It has been demonstrated before that microsatellites are more 132
Page 6 of 16
https://mc06.manuscriptcentral.com/cjm-pubs
Canadian Journal of Microbiology

Draft
7
abundant in coding regions than in non-coding regions of some prokaryotes (Gur-Arie et al. 133
2000; Ellegren 2004; Li et al. 2004) possibly due to an enhanced selection in coding regions. 134
Owing to higher polymorphism, compound microsatellites also have an enhanced ability of 135
altering gene function than a single microsatellite (Chen et al. 2012). However, no specific 136
distribution pattern of cSSRs could be inferred as distribution was not found to be 137
homogeneous within the genomes and also compound microsatellites were not concentrated 138
in specific genes. Numerous SSRs and cSSRs were present in the hypothetical proteins, so it 139
is difficult to remark on the occurrence of cSSRs with reference to specific proteins or their 140
sub units unless annotation information for all proteins in the studied genomes is available. 141
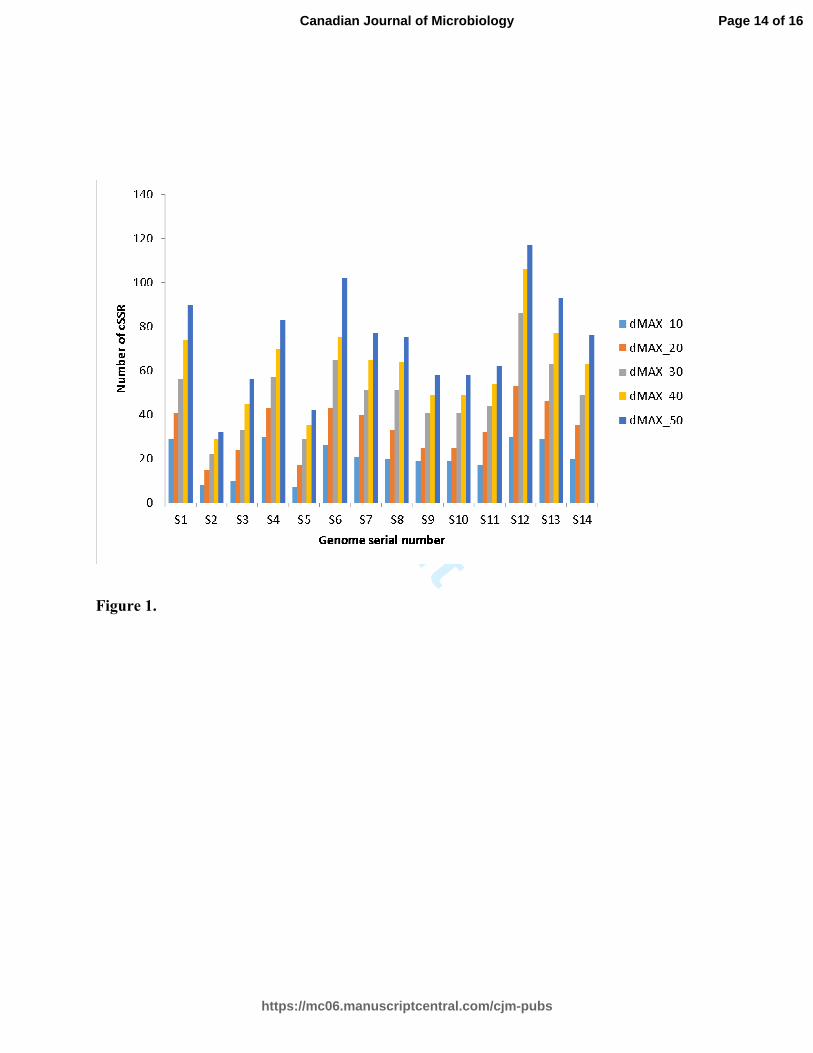
3.2 Effect of dMAX on compound microsatellite occurrence 142
While identifying compound microsatellites in a genome, dMAX is the most powerful 143
parameter. Hence, it is indispensable to verify the impact of dMAX on the detection of 144
compound microsatellites (Kofler et al. 2008; Chen et al. 2011; George et al. 2014). We 145
assessed this impact in all listed Lactobacillus strains (Table 1) and scrutinized the presence 146
of cSSRs with dMAX being varied from default threshold of 10 bp (fit to provide paramount 147
sensitivity for compound microsatellites identification by permitting for mismatches in 148
microsatellite quest) up to 50 bp. Our results indicated that the number of compound 149
microsatellites increased with the increase of dMAX in all Lactobacillus genomes (Figure. 1) 150
and it is, therefore, contemplated that increase in the maximum allowed distance between the 151
two adjoining SSRs can result in increased cSSR formation. 152
3.3 Motifs and complexity of cSSRs in studied genomes 153
Compound microsatellites are supposed to have originated by imperfection in microsatellites. 154
To determine the organization of individual microsatellite motifs and their role in setting up 155
Page 7 of 16
https://mc06.manuscriptcentral.com/cjm-pubs
Canadian Journal of Microbiology

Draft
8
compound microsatellites in Lactobacillus genomes, we probed the arrangement and 156
structural make-up of compound microsatellites too (Supplementary Table 1). Interestingly, 157
all compound microsatellite coupled motifs (e.g AAGGT-CTTGTT) were unique which 158
means they are distinct for each Lactobacillus species. Such motifs could probably have 159
arisen by imperfections in duplication. This observation was consistent with E. coli 160
compound microsatellites, formed of very dissimilar motifs having two or more distinct bases 161
(Chen et al. 2011). In eukaryotes two cSSRs have been reported to have very similar motifs 162
for at least 90% of cases (Kofler et al. 2008). Motif duplication i.e. similar motifs on both 163
ends of spacer sequence existed once in Lactobacillus delbrueckii subsp. bulgaricus ATCC 164
11842 ((AAT)n-(X)y-(AAT)z) and in Lactobacillus delbrueckii subsp. bulgaricus ATCC 165
BAA-365 ((GAA)5-X3-(GAA)4), once in Lactobacillus reuteri DSM 20016 and Lactobacillus 166
reuteri JCM 1112 ((ATT)4-X9-(ATT)4), twice in Lactobacillus gasseri ATCC 33323 ((AT)6-167
X8-(AT)6, (TTTA)3-X8-(TTTA)3)) and four times in Lactobacillus plantarum WCSF1 168
((CAT)4-X9-(CAT)5, (AAT)5-X6-(AAT)5-X6-(AAT)4, (ACT)5-X6-(ACT)4, (TTA)4,-X6-169
(TTA)5)). Occurrence of similar duplicated motifs in the same species (Lactobacillus 170
delbrueckii subsp. bulgaricus and Lactobacillus reuteri) suggests that they follow a similar 171
pattern of duplication in their genomes. Furthermore, a cSSR can either be characterized as a 172
perfect compound [(GT)n(AG)n] microsatellite or regarded as overlapping compound 173
microsatellite (overlap of preceding microsatellite with few bases of next microsatellite 174
(Kumar et al. 2014) e.g (AGG)n(GT)n). Overlapping compound microsatellites motifs existed 175
in all Lactobacillus genomes, ranging from a maximum of seventeen in Lactobacillus 176
johnsoni NCC 533 to a minimum of one in Lactobacillus fermentum IFO 3956 (Table 1). 177
Inspection of cSSR complexity indicated that all of the cSSRs found in surveyed genomes 178
belonged to either ‘2-microsatellite’, ‘3-microsatellite’ or ‘4-microsatellite’ compound 179
microsatellites. This reveals a variation in trend from previously analysed E. coli genomes as 180
Page 8 of 16
https://mc06.manuscriptcentral.com/cjm-pubs
Canadian Journal of Microbiology

Draft
9
they did not comprise any of the ‘4-microsatellite’ compound microsatellites. The complexity 181
of cSSRS increased even more with increased dMAX, with every genome showing at least 182
one ‘3-microsatellite’ cSSR, eight genomes showing ‘4-microsatellite’ cSSR and one genome 183
showing ‘5-microsatellite’ cSSR (Supplementary Table. 2). This confirms more complexity 184
of cSSRs in Lactobacillus genomes as compared to E.coli (Chen et al. 2011) but still less 185
than eukaryotes, that can have a complexity of up to ‘8-microsatellite’ (Kofler et al. 2008). 186
Complexity of compound microsatellites in prokaryotes cannot, thus, be attributed to genome 187
sizes as Lactobacillus genomes range from 1.8 to 3.3 Mb (Kant et al. 2011) as compared to 188
3.5 to 5.5 Mb of E. coli (Bergthorsson and Ochman 1995). We suggest that the complexity is 189
dependent on the abundance of microsatellites, which might result in improved frequency of 190
adjoining microsatellites by chance. 191
4 Conclusion 192
Knowledge about the biological significance of compound microsatellites in bacteria is 193
limited and should be attempted in completely sequenced genomes to systematically reveal 194
the nature and evolutionary dynamics of cSSRs. Comparative analysis of compound 195
microsatellites in various Lactobacillus genomes with previously studied eukaryotic and 196
prokaryotic genomes shows that this category of microsatellites is overrepresented in 197
Lactobacilli. It is assumed that overrepresentation of compound microsatellites in bacteria of 198
genus Lactobacillus is due to either increased duplication of imperfections in microstaellites, 199
or DNA replication slippage. The population size of bacteria may also be a crucial factor 200
governing the abundance of microsatellites and hence, compound microsatellites. The 201
diversity of cSSRs in Lactobacillus genomes may be useful for better understanding of their 202
genetic diversity, evolutionary biology and strain/genotype demarcations. 203
204
Page 9 of 16
https://mc06.manuscriptcentral.com/cjm-pubs
Canadian Journal of Microbiology

Draft
10
References 205
Bergthorsson, U., Ochman, H.1995. Heterogeneity of genome sizes among natural isolates of 206
Escherichia coli. J. Bacteriol., 177(20):5784-5789. 207
Brinkmann, B., Klintschar, M., Neuhuber, F., Hühne, J., Rolf, B. 1998. Mutation rate in 208
human microsatellites: Influence of the structure and length of the tandem repeat. 209
Am. J. Hum. Genet., 62:1408–1415.doi: 10.1086/301869. 210
Bull, L.N., Pabón-Peña, C.R., Freimer, N.B. 1999. Compound microsatellite repeats: 211
practical and theoretical features. Genome Res., 9(9): 830-838. doi: 212
10.1101/gr.9.9.830. 213
Buschiazzo, E., Gemmell, N. J. 2010. Conservation of human microsatellites across 450 214
million years of evolution. Genome Bio. Evol., evq007. doi: 10.1093/gbe/evq007. 215
Chebeňová, V., Berta, G., Kuchta, T., Brežná, B., Pangallo, D. 2010. Randomly-amplified 216
microsatellite polymorphism for preliminary typing of lactic acid bacteria from 217
Bryndza Cheese. Folia Microbiol. (Praha), 55(6):598-602. doi: 10.1007/s12223-218
010-0096-4. 219
Chen, M., Tan, Z., Zeng, G., Zeng, Z. 2012. Differential distribution of compound 220
microsatellites in various human immunodeficiency virus type 1 complete 221
genomes. Infect. Genet. Evol., 12(7): 1452-1457. doi: 222
10.1016/j.meegid.2012.05.006. 223
Chen, M., Zeng, G., Tan, Z., Jiang, M., Zhang, J., Zhang, C., Lu, L., Lin, Y., Peng, J. 2011. 224
Compound microsatellites in complete Escherichia coli genomes. FEBS Letters, 225
585 (7): 1072-1076. doi: 10.1016/j.febslet.2011.03.005. 226
Ellegren, H. 2004. Microsatellites: simple sequences with complex evolution. Nat. Rev. 227
Genet., 5(6): 435-445. doi: 10.1038/nrg1348. 228
Page 10 of 16
https://mc06.manuscriptcentral.com/cjm-pubs
Canadian Journal of Microbiology

Draft
11
Garza, J.C., Freimer, N.B. 1996. Homoplasy for size at microsatellite loci in humans and 229
chimpanzees. Genome Res., 6:211–217. doi: 10.1101/gr.6.3.211. 230
George, B., Gnanasekaran, P., Jain, S.K., Chakraborty, S. 2014. Genome wide survey and 231
analysis of small repetitive sequences in caulimoviruses. Infect. Genet. Evol., 232
27:15-24. doi: 10.1016/j.meegid.2014.06.018. 233
Gur-Arie, R., Cohen, C.J., Eitan, Y., Shelef, L., Hallerman, E.M., Kashi, Y. 2000. Simple 234
sequence repeats in Escherichia coli: abundance, distribution, composition, and 235
polymorphism. Genome Res., 10(1): 62-71. 236
Kant, R., Blom, J., Palva, A., Siezen, R.J., de Vos, W.M. 2011. Comparative genomics of 237
Lactobacillus. Microbial biotechnol., 4(3): 323-332. doi: 10.1111/j.1751-238
7915.2010.00215.x. 239
Kashi, Y., King, D.G. 2006. Simple sequence repeats as advantageous mutators in evolution. 240
Trends Genet., 22(5): 253-259. doi: 10.1016/j.tig.2006.03.005. 241
Kofler, R., Schlotterer, C., Luschutzky, E., Lelley, T. 2008. Survey of microsatellite 242
clustering in eight fully sequenced species sheds light on the origin of compound 243
microsatellites. BMC Genomics, 9(1):612. doi: 10.1186/1471-2164-9-612. 244
Kumar, M., Kapil, A., Shanker, A. 2014. MitoSatPlant: Mitochondrial microsatellites 245
database of viridiplantae. Mitochondrion.19:334-337. doi: 246
10.1016/j.mito.2014.02.002. 247
Laura, N.B., Pabón-Peña, C.R., Freimer, N.B. 1999. Compound microsatellite repeats: 248
practical and theoretical features. Genome Res., 9(9): 830–838. doi: 249
10.1101/gr.9.9.830. 250
Li, Y.C., Korol, A.B., Fahima, T., Nevo, E. 2004. Microsatellites within genes: structure, 251
function, and evolution. Mol. Biol. Evol., 21(6): 991-1007. doi: 252
10.1093/molbev/msh073. 253
Page 11 of 16
https://mc06.manuscriptcentral.com/cjm-pubs
Canadian Journal of Microbiology

Draft
12
Mudunuri, S.B., Nagarajaram, H.A. 2007. IMEx: Imperfect Microsatellite Extractor. 254
Bioinformatics, 23(10):1181-1187. doi: 10.1093/bioinformatics/btm097. 255
Toth, G., Gaspari, Z., Jurka, J. 2000. Microsatellites in different eukaryotic genomes: survey 256
and analysis. Genome Res. 10:967–981. doi: 10.1101/gr.10.7.967. 257
Urquhart, A., Kimpton, C.P., Downes, T.J., Gill, P. 1994. Variation in short tandem repeat 258
sequences—A survey of twelve microsatellite loci for use as forensic identification 259
markers. Int. J. Leg. Med., 107(1):13–20. doi: 10.1007/bf01247268. 260
Weber, J.L. 1990. Informativeness of human (dC-dA)n x (dG-dT)n polymorphisms. Genomics 261
7:524-530. doi: 10.1016/0888-7543(90)90195-z. 262
263
264
265
266
267
268
269
270
271
272
273
274
275
276
277
278
Page 12 of 16
https://mc06.manuscriptcentral.com/cjm-pubs
Canadian Journal of Microbiology

Draft
13
Figure Legend Information 279
Figure 1. Increment of cSSR in Lactobacillus genomes with increased dMAX values. S1, 280
S2.…refer to the genome serial numbers in Table 1. 281
282
Page 13 of 16
https://mc06.manuscriptcentral.com/cjm-pubs
Canadian Journal of Microbiology
Draft
Figure 1.
Page 14 of 16
https://mc06.manuscriptcentral.com/cjm-pubs
Canadian Journal of Microbiology

Draft
Table 1. List of analysed Lactobacillus genomes and compound microsatellites
Serial
#
Accession Organism Genome
size
(bp)
GC
content
(%)
Coding
density
(%)
Total number
of
microsatellites
Total number
of compound
microsatellites
Number of
overlapping
compound
microsatellites
S1 NC_006814.3 Lactobacillus
acidophilus NCFM
1,993,560 34.7 88.2 1819 29 15
S2 NC_008497 Lactobacillus
brevis ATCC 367
2,291,220 46.2 83.4 1161 8 5
S3 NC_008526.1 Lactobacillus casei
ATCC 334
3,079,196 46.6 81.9 1566 10 3
S4 NC_008054.1 Lactobacillus
delbrueckii subsp.
bulgaricus ATCC
11842
1,864,998 49.7 73.46 1370 30 15
S5 NC_010610.1 Lactobacillus
fermentum IFO
3956
2,098,684 51.5 80.2 1304 7 1
S6 NC_008530.1 Lactobacillus
gasseri ATCC
33323
1,894,360 35.3 88.2 1652 26 8
S7 NC_010080.1 Lactobacillus
helveticus DPC
4571
2,080,931 37.1 73.6 1653 21 10
S8 NC_004567.1 Lactobacillus
plantarum WCFS1
3,308,274 44.47 83.4 1944 20 6
S9 NC_009513.1 Lactobacillus
reuteri DSM 20016
1,999,618 38.9 85.0 1544 19 7
S10 NC_010609.1 Lactobacillus
reuteri JCM 1112
2,039,414 38.9 83.3 1575 19 6
S11 NC_007576.1 Lactobacillus sakei
subsp. sakei 23K
1,884,661 41.26 87.0 1313 17 7
Page 15 of 16
https://mc06.manuscriptcentral.com/cjm-pubs
Canadian Journal of Microbiology

Draft
S12 NC_007929.1 Lactobacillus
salivarius UCC118
1,827,111 33.04 82.5 1902 30 12
S13 NC_005362.1 Lactobacillus
johnsonni NCC 533
1,992,676 34.6 89.0 1811 29 17
S14 NC_008529.1 Lactobacillus
delbrueckii subsp.
bulgaricusATCC
BAA-365
1,856,951 49.69 77.65 1360 20 8
Page 16 of 16
https://mc06.manuscriptcentral.com/cjm-pubs
Canadian Journal of Microbiology


















