
DOGFISH ELECTRORECEPTION AT UTRECHT UNIVERSITY … · DOGFISH ELECTRORECEPTION AT UTRECHT...
12
SDT Reference Series International 1, 54-65 title , Peters-RC, Dogfish Electroreception 1966 ______________________________________________________________________________________________________________________________________________________________________________________ DOGFISH ELECTRORECEPTION AT UTRECHT UNIVERSITY 1966, A RETROSPECTIVE VIEW Peters-RC* 2016 SDT Reference Series International 1, 54-65 Half a century ago, Dijkgraaf and Kalmijn published a paper on dogfish electroreception, a contribution to the 'Festschrift' at the occasion of Karl von Frisch's 80th birthday. As was explained to the author at the time – and as is supported by Dijkgraaf's correspondence – the paper was a rush job, which did not fully meet the standards of a peer-reviewed experimental publication. The following should be considered a footnote to Dijkgraaf & Kalmijn's publication, and as a stage for serendipity and search images and their parts played in the discovery of the biological significance of the ampullae of Lorenzini. Published 23 March 2016 (with corrections April, September 2016) *Corresponding author at http://www.deTraditie.nl © 2016 R.C.Peters ____ Karl von Frisch , Sven Dijkgraaf , Hans W. Lissmann , Peter E.P. Görner, Ad J. Kalmijn , Rob C. Peters , Arie Schuijf , Sir John Eccles , Yuri N. Andrianov , references , timeline 50 years ago – Karl von Frisch's 80th Birthday (1966) 'Karl v. Frisch zum 80. Geburtstag in Verehrung gewidmet' . In their paper Versuche zur biologischen Bedeutung der Lorenzinische Ampullen bei den Elasmobranchiern (Dijkgraaf & Kalmijn 1966), the authors summarize a number of characteristics of the ampullae of Lorenzini, found during their researches. These characteristics were: 1) the detection threshold of the ampullae, 2) the electrical sources to which they respond; at that time presumed to be muscle potentials, 3) the ability to learn to detect electrical dipoles hidden in the sandy bottom, and 4) the biological significance as derived from their being specifically sensitive to electrical stimuli. At the time I worked as a student tutored by Ad Kalmijn at Dijkgraaf's lab. My task was to perform in situ electrophysiological recordings from the nerves innervating the ampullae of Lorenzini. And although I happened to make some publishable recordings, I was rather disappointed that my results were considered valuable enough to be presented in the 'Festschrift', but that acknowledgements were lacking. I had to understand that writing the paper had been a rush job because Professor Dijkgraaf received the invitation to write a contribution rather unexpectedly; he had two months only to write the paper and to have it accepted. Also I noticed that none of the contributions of the other students tutored by Ad Kalmijn were acknowledged. The net result was that the success of my implantation experiment was balanced by the omission in the acknowledgements, allowing me to continue my existence in modesty. During my major internship at Dijkgraaf's lab, I had the impression that the study on electroreception in dogfish knew a long tradition. To my surprise I noticed recently in the card-tray of Dijkgraaf's slide collection, that the ampullae of Lorenzini were filed under 'Mechanoreceptors' (fig. 1). This incited me to have a closer look at Dijkgraaf's involvement in ampullary research by digging in his legacy, conserved by Stichting De Traditie. The following is an elaboration of item 4 in the summary of the aforementioned publication of Dijkgraaf & Kalmijn (1966) on the biological significance of the ampullae of Lorenzini in dogfish. 54
Transcript of DOGFISH ELECTRORECEPTION AT UTRECHT UNIVERSITY … · DOGFISH ELECTRORECEPTION AT UTRECHT...

SDT Reference Series International 1, 54-65 title, Peters-RC, Dogfish Electroreception 1966______________________________________________________________________________________________________________________________________________________________________________________
DOGFISH ELECTRORECEPTION AT UTRECHT UNIVERSITY 1966,
A RETROSPECTIVE VIEW
Peters-RC* 2016
SDT Reference Series International 1, 54-65
Half a century ago, Dijkgraaf and Kalmijn published a paper on dogfish electroreception, a contribution to the 'Festschrift' at the occasion of Karl von Frisch's
80th birthday. As was explained to the author at the time – and as is supported by Dijkgraaf's correspondence – the paper was a rush job, which did not fully meet the
standards of a peer-reviewed experimental publication. The following should be considered a footnote to Dijkgraaf & Kalmijn's publication, and as a stage for serendipity and search images and their parts played in the discovery of the
biological significance of the ampullae of Lorenzini.
Published 23 March 2016 (with corrections April, September 2016)
*Corresponding author at http://www.deTraditie.nl
© 2016 R.C.Peters
____
Karl von Frisch, Sven Dijkgraaf, Hans W. Lissmann, Peter E.P. Görner, Ad J. Kalmijn,
Rob C. Peters, Arie Schuijf, Sir John Eccles, Yuri N. Andrianov, references, timeline
50 years ago – Karl von Frisch's 80th Birthday (1966)
'Karl v. Frisch zum 80. Geburtstag in Verehrung gewidmet'. In their paper Versuche
zur biologischen Bedeutung der Lorenzinische Ampullen bei den Elasmobranchiern
(Dijkgraaf & Kalmijn 1966), the authors summarize a number of characteristics of the
ampullae of Lorenzini, found during their researches. These characteristics were: 1)
the detection threshold of the ampullae, 2) the electrical sources to which they
respond; at that time presumed to be muscle potentials, 3) the ability to learn to
detect electrical dipoles hidden in the sandy bottom, and 4) the biological
significance as derived from their being specifically sensitive to electrical stimuli.
At the time I worked as a student tutored by Ad Kalmijn at Dijkgraaf's lab. My task
was to perform in situ electrophysiological recordings from the nerves innervating
the ampullae of Lorenzini. And although I happened to make some publishable
recordings, I was rather disappointed that my results were considered valuable
enough to be presented in the 'Festschrift', but that acknowledgements were
lacking. I had to understand that writing the paper had been a rush job because
Professor Dijkgraaf received the invitation to write a contribution rather
unexpectedly; he had two months only to write the paper and to have it accepted.
Also I noticed that none of the contributions of the other students tutored by Ad
Kalmijn were acknowledged. The net result was that the success of my implantation
experiment was balanced by the omission in the acknowledgements, allowing me to
continue my existence in modesty.
During my major internship at Dijkgraaf's lab, I had the impression that the study on
electroreception in dogfish knew a long tradition. To my surprise I noticed recently in
the card-tray of Dijkgraaf's slide collection, that the ampullae of Lorenzini were filed
under 'Mechanoreceptors' (fig. 1). This incited me to have a closer look at Dijkgraaf's
involvement in ampullary research by digging in his legacy, conserved by Stichting
De Traditie. The following is an elaboration of item 4 in the summary of the
aforementioned publication of Dijkgraaf & Kalmijn (1966) on the biological
significance of the ampullae of Lorenzini in dogfish.
54

SDT Reference Series International 1, 54-65 title, Peters-RC, Dogfish Electroreception 1966______________________________________________________________________________________________________________________________________________________________________________________
Figure 1. Two of Dijkgraaf's index cards of the collection of transparencies used for teaching. Both deal with electroreception and ampullae of Lorenzini, but are marked 'Mechanoreception'. M46 shows how a silver wire electrode is mounted around ampullary nerves in dogfish. M47 shows the response of an ampullary nerve bundle to an iron rod. Both illustrations are copies of figures of Dijkgraaf & Kalmijn 1966.
Enter Sven Dijkgraaf (1932), Lorenzinian ampullae
According to Görner c.s. (1996), Dijkgraaf's first contact with the lateral line system
took place in Vienna, at professor Abel's lab. However, rather soon Dijkgraaf moved
to Karl von Frisch's in München, where he presumably had more freedom to choose
the experimental approach. There he began to work on rheotaxis and the lateral line
in minnows. The first evidence of his involvement with the ampullae of Lorenzini is a
Referat on 24 February 1932, entitled Bau und Funktion der Lorenzinischen
Ampullen (Dotterweich-H 1932. Zool. Jahrb. Abt. Phys. 50, 347-418).
After finishing his PhD thesis (1933), he follows a course on Comparative Physiology
at Hermann Jordan's lab at Utrecht. Jordan was a proponent of conduction with
decrement in invertebrate nerve fibers. As a consequence, we retrace Dijkgraaf not
much later in the Zoological Station at Naples, fighting with a string galvanometer,
trying to, but unable to, confirm Jordan's view on conduction with decrement. A
manuscript on these findings was submitted to Pflügers Archiv, but rejected by
editor Bethe. Stripped of some expletives, Dijkgraaf's reaction is given by the
drawing on the rejection letter: Aplysia Arbeit, Ruhe sanft (fig. 2).
Figure 2. The drawing at the back of the letter in which Bethe explains why he cannot accept the manuscript for publication. The drawing is self evident: 'Ruhe sanft', and 'Aplysia Arbeit' makes citation of certain expletives unnecessary.
In the same notebook in which he made notes on the galvanometer experiment,
experiments with Scyllium are described; lateral line experiments on the detection of
water current. However, a note dated 14 June 1935 describes his observation of the
reaction of Scyllium to iron wires. As he later wrote me, when I asked permission to
publish his notes, he attributed the reaction to leakage currents from the mains. It is
very likely that this period causes his undeniable future aversion of the string
galvanometer. In 1936 he receives a teaching post at Groningen University at Engel
Hazelhoff's lab to teach biology to medical students. His experimental notes at
55

SDT Reference Series International 1, 54-65 title, Peters-RC, Dogfish Electroreception 1966______________________________________________________________________________________________________________________________________________________________________________________
Groningen reveal among other things the assistance of A.H.J. Vendrik in recording
nerve impulses of the lateral line fibers in Xenopus. Incidentally this seems to be
successful. Then comes WW2. In the mean time there are no involvements with
ampullae of Lorenzini. After the war – 1948 - he is nominated professor of
Comparative Physiology at Utrecht, Jordan's successor! B.J. Krijgsman (1940, 1947)
is the local electrophysiologist who can handle the string galvanometer, who favours
the oscilloscope, but who leaves to South Africa in 1950. The ampullae of Lorenzini
remain unexplored at Utrecht, but the urgent need of an electrophysiologist who can
study the lateral line system can no longer be denied.
Enter Hans W. Lissmann (1948), electric fish
In 1948, shortly after Dijkgraaf's nomination, Hans Lissmann from Cambridge seeks
contact with Dijkgraaf in order to visit the sea aquarium of the laboratory at Utrecht.
A rather intensive correspondence takes place. In 1951, Lissmann's paper appears
on the electrical signals from the tail of a fish, after which Dijkgraaf writes to
Lissmann about his peculiar observation of dogfish responding to iron rods in Naples,
and about the unexplained electrical sensitivity of catfish to weak electrical currents
described by Parker and van Heusen (1917). In this letter to Lissmann, Dijkgraaf
dates his famous observation in 1931, but Dijkgraaf's 'Versuchsprotokollen' show a
different year: 1935. He also writes that it would be very interesting to test the
sensitivity of the ampullae of Lorenzini to electrical stimuli, because they seem to be
very insensitive to mechanical stimuli. (fig. 3. Dijkgraaf to Lissmann: 17 December
1951). In 1952, student Peter F. Elbers, later professor of Electron Microscopy, starts
with electrophysiological recordings on the lateral line of Xenopus. It would take a
couple of years extra, and the efforts of two more students – A.C.J. Burgers and A.M.
Stadhouders – to gain publishable results (Dijkgraaf 1956).Figure 3. Sections of a carbon copy of the letter by Sven Dijkgraaf to Hans Lissmann. The lower section is the back of the letter (verso), the top two sections are the front (recto), minus a few personal lines. Apparently Dijkgraaf realizes the electrosensitive potential of the Lorenzinian ampullae, but somehow he does not seem to have plans to tackle the problem himself electrophysiologically.
56

SDT Reference Series International 1, 54-65 title, Peters-RC, Dogfish Electroreception 1966______________________________________________________________________________________________________________________________________________________________________________________
Enter Peter E.P. Görner & Ad J. Kalmijn (1959), electrophysiology
The need of an experienced electrophysiologist is high. Dijkgraaf falls back on his
Münchener background, and finds in Peter Görner the man who can continue the
electrophysiological experiments on Xenopus, The Utrecht Philips-Voigtlander
oscilloscope camera is transported from Utrecht to München, and when Görner sends
his first scuccessful recordings of the lateral line organs of Xenopus, a position in
Utrecht is created. Görner begins at Utrecht in 1959. All efforts are focused on he
mechanoreceptive function of the lateral line system, for which Görner receives
assistance from a student, Ad Kalmijn. He further creates a sound basis for the
instruction of electrophysiological recording techniques. In 1959 we see, finally, for
the first time continuation of electrophysiological research by students. In the
internal annual of Utrecht University of 1959/1960, students are listed who work on
the electrophysiology of the lateral line (W. Berendsen), and on the ampullae of
Lorenzini (A.J. Kalmijn). Students working on the electrophysiology of the ampullae of
Lorenzini are also mentioned in the annual of 1961/1962, but are absent in that of
1962/1963. The latter annual mentions students who are going to work on the
electrophysiology of the Crustacean eye (R.C. Peters), frog labyrinth (J. Atema),
chemoreception in flies (L. Schouten), and Crustaean statocysts (H.G. Gallé). In the
mean time H. Hensel (1955, 1956) has been investigating electrophysiologically the
effects of mechanical and thermal stimulation on the ampullae of Lorenzini, and R.W.
Murray is performing electrophysiology on Xenopus lateral line organs and also
ampullae of Lorenzini in dogfish (1957, 1959, 1960, 1962). It is Murray (1962) who
finds moreover an extreme sensitivity to electrical stimulation. This must have been
an unpleasant surprise for the Utrecht student whose task it was to perform
electrophysiology on the ampullae of Lorenzini. On the other hand, the biological
significance of the ampullae remained still an enigma, because of the sensitivity to
both thermal, mechanical, and electrical stimuli. It is Lissmann again who suggests
tentatively that muscle potentials of other fish might be the relevant stimuli to the
ampullae (Lissmann 1958, p. 183). Muscle potentials, because dogfish have no
electric organ like Lissmann's Gymnarchus.
Enter Rob C. Peters, in situ recordings (1966)
My internship at Comparative Physiology began in 1963 as assistant of the 3rd year
practical. I was scouted by Jelle Atema and more or less pressed to sign a working
contract. Supervisor of the muscle and nerve physiology part was Ad Kalmijn, who
succeeded Peter Görner as teacher of electrophysiological techniques.
My first recollection of the work at the lab was the battery operated electrical
stimulator, specifically designed by Ad Kalmijn for field work on the
electrosensitivity of dogfish. Ad's proposal for my internship was to spend the first
weeks of my internship at the lab in order to get a feeling of what was needed to
perform electrophysiological experiments successfully. Then I should do a 6 months
minor in physics with, among other things, electronics and theoretical electricity,
after which I could return to Comparative Physiology to finish my 12 months major.
This turned out to be a very profitable study program, which later helped me to
survive in the academic environment. During the last part of my major, I was
redirected to the electric sense in dogfish. The question of the biological significance
mentioned in Dijkgraaf & Kalmijn (1962) was still not answered. Electrophysiological
experiments had demonstrated sensitivity of the ampullae of Lorenzini to
mechanical, thermal, and electrical stimuli, but behavioural evidence for a specific
natural biological use was still missing. I got the task to record spikes from the
nerves innervating the ampullae of Lorenzini in situ, without damaging the ampullary
capsules, to investigate the ampullary responses under conditions as natural as
possible. The procedure outlined by Kalmijn was straightforward. Anesthetize the
dogfish with MS222 – which he introduced as a central anesthetic – expose the
ampullary nerves which would not be affected by the anesthetic, implant a silver
wire electrode after having established a provisional response, and have the animal
recover to swim around freely. There were, of course, some minor difficulties to
overcome. Kalmijn's idea was to make a T-piece of perspex in which the electrode
and nerve could be mounted. The mechanics workshop considered this task to
cumbersome, so that I had to spend many hours behind the lathe myself. After
having tested these T-pieces and other structures on nerve fibers of frogs, eventually
57

SDT Reference Series International 1, 54-65 title, Peters-RC, Dogfish Electroreception 1966______________________________________________________________________________________________________________________________________________________________________________________
< Figure 4. Drawings made in 1966 by the author to be copied into the typewritten report (1966). Remember that at the time the Lab did not have a Xerox machine; the figures had to be photographed manually first. The figures show how the silver electrode wire was attached finally to the nerve bundle (top left), and the drawing of the custom made waterproof connector (top right) The middle row shows the position and size of the adhesive patch on the dogfish's head. The bottom row shows a drawing of the custom made pespex T-piece (bottom left), and a simple polythene cannulae (bottom right), both intended as insulation compartments for the recording site. The legends of the figures are omitted here. Copies of these originals were pasted into the carbon copies of the report. See also figure 11.
Figure 5. Photograph of the anesthetized dogfish with two insulation stockings glued on its head. Only after hardening of the glue, the skin in front of the stockings was opened in order to expose the underlying nerve bundles of the ampullae of Lorenzini. Size of the dogfish about 50 cm TL.
58

SDT Reference Series International 1, 54-65 title, Peters-RC, Dogfish Electroreception 1966______________________________________________________________________________________________________________________________________________________________________________________
Figure 6. Impression of the equipment in use at the Laboratory of Comparative Physiology at Utrecht in 1966. Demonstrator is enthusiastic and expert Jelle Atema. Close to Jelle the Tektronix 502A Oscilloscope, and behind him the Tektronix 122/125 Pre-amplifier. The tape recorder to the right is a Revox G36 for the recording of action potentials and gross activity. Next to the Revox stands the Hellige Helgoscript He86 paper chart recorder. Above the Hellige recorder, its two pre-amplifier stages. In front, part of the plastic wading pool in use for dogfish and rays.
a simple piece of insulation stocking proved the best solution, more or less as
described in Delgado (1964, p. 122). Another problem was the mechanical fixation of
the electrode wire onto the skin of the dogfish. At the time we choose for a rather
new two component adhesive that seemed non toxic. Hardening however took more
than an hour. For the implantation procedure the dogfish was put on the table top
(fig. 5), and provided the fish was kept moist with seawater, its oxygen exchange
Figure 7. Recording of the approach of an iron rod to the ampullae of Lorenzini. Before writing my final report on the research period at Ad Kalmijn's, I was asked to make some pictures that could be used for the Karl von Frisch Festschrift. Also I should write a preliminary report with my results briefly summarized, for the same purpose. Inset: picture of the dogfish in the plastic pool. The recordings were filmed with a Philips-Voigtlander oscilloscope camera type FE106-5500 ; for the picture of the dogfish with its floating connector attached I used my Exa Ia camera
through gills and skin proved sufficient to have the animal survive the 3 hours
needed for the operation. After having anesthetized the dogfish, I tried to record
table top wise from the nerves, but the only responses I got were trains of spikes
that might be related with respiration, as well as 'contact' responses on touching
with a metal pin. On the whole a very disappointing result. I made two or three of
such uncuccessful operations.
59
SDT Reference Series International 1, 54-65 title, Peters-RC, Dogfish Electroreception 1966______________________________________________________________________________________________________________________________________________________________________________________
Enters: Arie Schuijf (1966), slide show of Arie on Safari
The last time I tried to implant an electrode was on March 23, 1966. At the end of
another apparent failure, I put the dogfish back into its tank, with the intention to
have a quick meal, and then to return to the lab in order to check the treated dogfish
before attending the slide show by Arie Schuijf. Arie had made a beautiful series of
transparencies during a holiday trip through Africa, which was referred to as the
Tombuctu and Kilimanjaro trip: Arie on Safari! I did not want to miss it. When I
returned to the lab, I was happy to see that the dogfish had recovered fully from its
anesthesia and was swimming around actively. So I decided to skip the first part of
Arie's talk, and check the implanted electrode, just in case a miracle would have
happened. To my surprise, a miracle did have happened. When I connected the
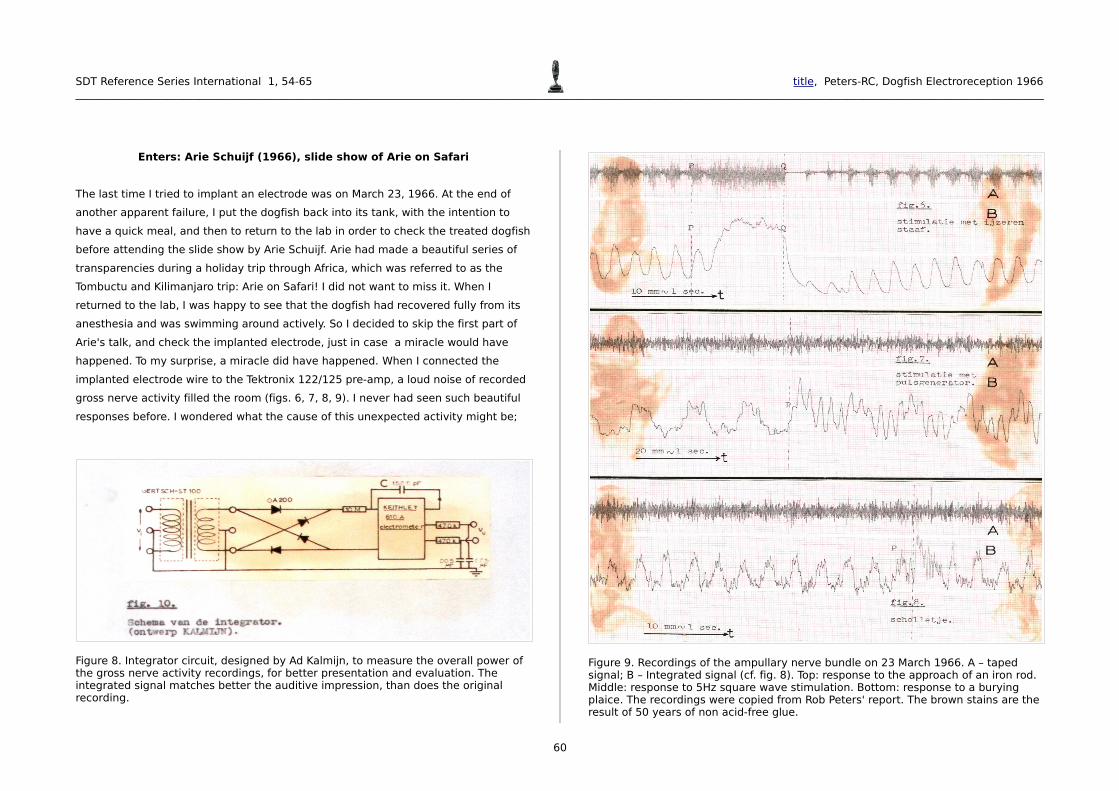
implanted electrode wire to the Tektronix 122/125 pre-amp, a loud noise of recorded
gross nerve activity filled the room (figs. 6, 7, 8, 9). I never had seen such beautiful
responses before. I wondered what the cause of this unexpected activity might be;
Figure 8. Integrator circuit, designed by Ad Kalmijn, to measure the overall power of the gross nerve activity recordings, for better presentation and evaluation. The integrated signal matches better the auditive impression, than does the original recording.
Figure 9. Recordings of the ampullary nerve bundle on 23 March 1966. A – taped signal; B – Integrated signal (cf. fig. 8). Top: response to the approach of an iron rod. Middle: response to 5Hz square wave stimulation. Bottom: response to a burying plaice. The recordings were copied from Rob Peters' report. The brown stains are the result of 50 years of non acid-free glue.
60

SDT Reference Series International 1, 54-65 title, Peters-RC, Dogfish Electroreception 1966______________________________________________________________________________________________________________________________________________________________________________________
most likely the electrode had been displaced and made better contact with the
nerve. At least that is what I thought at that moment. So without any further delay I
took a piece of iron and brought it in the vicinity of the dogfish's snout. The
response was enormous (fig. 7). A further conspicuous feature was that the
responses were heavily modulated in the rhythm of the gill movements. The only
other thing I could do at the time was to stimulate the ampullae electrically with
Kalmijn's battery operated square wave generator; square waves of 5 Hz. Then I
realized that a plaice was lying in the same pool, and since the objective of my
internship was to find out to what kind of stimuli the ampullae of Lorenzini
responded under 'normal' circumstances, I spent about half an hour to manipulate
the dogfish above the plaice just in time to have it respond to the burying
movements of the plaice (fig. 9). Remember that Lissmann (1958) had suggested
that muscle potentials might be a biologically relevant stimulus source for the
ampullae of Lorenzini. To see if under these conditions the ampullae also responded
to temperature changes, salt concentrations, and mechanical stimulation, I poured
warm seawater into the pool, tap water, and I tapped the snout of the dogfish with a
plastic rod. All to no avail, i.e. I did not recognize audible responses, whereas, the
square waves, the iron rod, the respiration movements, and the burying place
yielded clearly audible modulations in the gross activity. All this took about one hour,
so in the break of Arie's talk I invited professor Dijkgraaf, Ad Kalmijn, and of course
my fiancée Tonny Gest, downstairs in the basement to enjoy these unexpected
findings. I demonstrated the iron rod trick again, and we all returned happily
afterwards to Arie's slides.
On Kalmijn's request I wrote a quick report to be used by Dijkgraaf for the von Frisch
paper, and a more elaborate one as the final report of my internship. In both I tried
to describe dutifully the procedure followed, but did not dwell on the discrepancy
between my unsuccessful experiment the afternoon, and the successful recording
that same evening. I just took the success for granted. This was – my view - partly
due to Kalmijn's rather imperative and detailed way of editing my reports.
At the time nobody asked how it came that the recording of such beautiful responses
had been impossible in the afternoon. Nobody realized that MS222 blocked the
synapses of the ampullary system, and that the procedure I had described could
never have yielded proper responses during anesthesia. It was my guess then, as
unexperienced electrophysiologist, that the electrode had been moved somehow,
with better recording conditions as the result. When Kalmijn went to Banyuls in the
fall of 1969 to repeat my experiments more in depth himself, he discovered to his
chagrin, that it was impossible to get decent responses under MS222 anesthesia. He
told me that it took him at least a couple of weeks before he realized that MS222
also blocks the ampullary synapses. I have tried to recover in vain why he thought
MS222 to sedate the central nervous system selectively; in the literature MS222 was
simply described as a general anesthetic. I was further very disappointed that
Kalmijn did not accept the responses to the burying plaice as responses to electrical
stimuli. According to him, they were mechanosensory lateral line responses. I
reluctantly adapted my report according to his instructions, but I have been always
convinced that these responses were electrosensory, mainly because I could not
evoke responses by tapping the ventral skin with a plastic rod (cf. Dijkgraaf &
Kalmijn 1966, summary nr. 4). And last but not least, neither Dijkgraaf nor Kalmijn
were there when I made this recording.
Enter Sir John Eccles (1969), Direct Current stimuli
Sir John Eccles does not need an introduction. However, what is not generally known,
is his contribution to the understanding of the biological significance in
electrosensitive fish without electric organs. Somewhere in 1969 he visited Utrecht
to give a lecture on what I remember as the evolution of the visual arts1; at the same
occasion he visited Dijkgraaf's lab, and was introduced to Kalmijn's dogfish work
(personal communication by Kalmijn, and later by Dijkgraaf). At the time it was still
not known to what electrical stimuli the ampullae of Lorenzini responded under
natural conditions. The most promising sources were muscle potentials of other fish,
as suggested by Lissmann. It was Sir John Eccles who provided an alternative: strong
1 Up till now I was unable to pinpoint the exact date of his visit.
61
SDT Reference Series International 1, 54-65 title, Peters-RC, Dogfish Electroreception 1966______________________________________________________________________________________________________________________________________________________________________________________
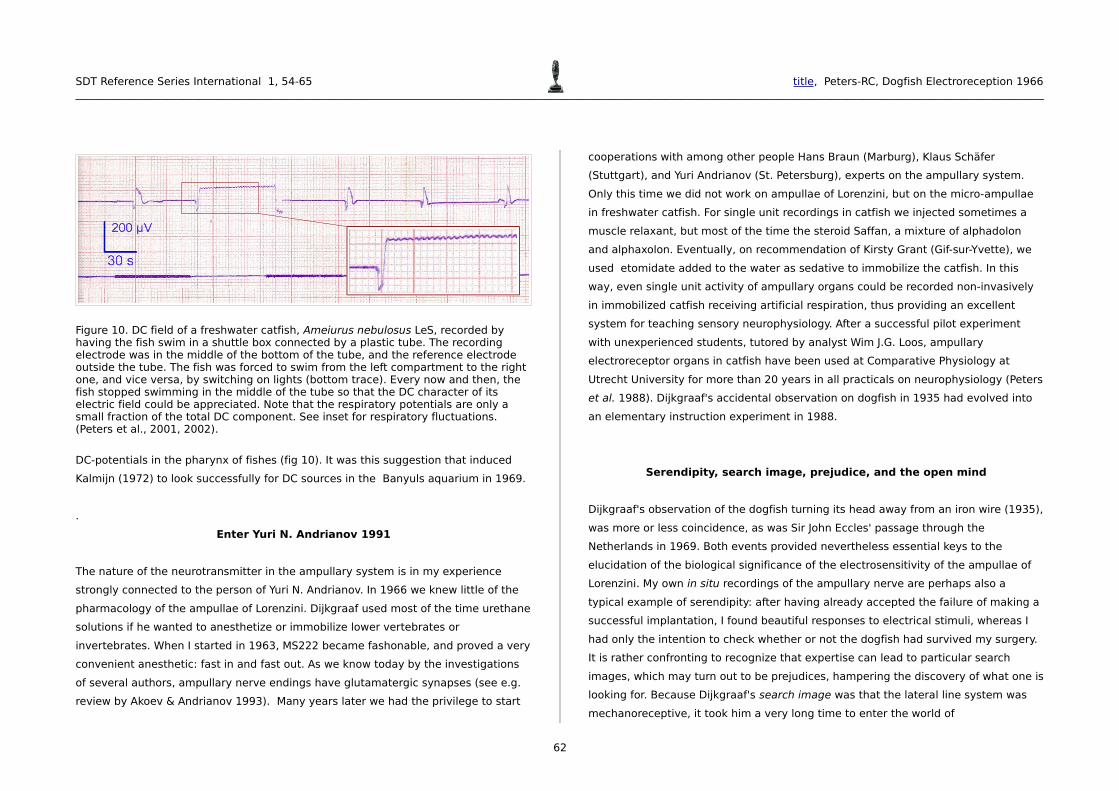
Figure 10. DC field of a freshwater catfish, Ameiurus nebulosus LeS, recorded by having the fish swim in a shuttle box connected by a plastic tube. The recording electrode was in the middle of the bottom of the tube, and the reference electrode outside the tube. The fish was forced to swim from the left compartment to the right one, and vice versa, by switching on lights (bottom trace). Every now and then, the fish stopped swimming in the middle of the tube so that the DC character of its electric field could be appreciated. Note that the respiratory potentials are only a small fraction of the total DC component. See inset for respiratory fluctuations. (Peters et al., 2001, 2002).
DC-potentials in the pharynx of fishes (fig 10). It was this suggestion that induced
Kalmijn (1972) to look successfully for DC sources in the Banyuls aquarium in 1969.
.
Enter Yuri N. Andrianov 1991
The nature of the neurotransmitter in the ampullary system is in my experience
strongly connected to the person of Yuri N. Andrianov. In 1966 we knew little of the
pharmacology of the ampullae of Lorenzini. Dijkgraaf used most of the time urethane
solutions if he wanted to anesthetize or immobilize lower vertebrates or
invertebrates. When I started in 1963, MS222 became fashonable, and proved a very
convenient anesthetic: fast in and fast out. As we know today by the investigations
of several authors, ampullary nerve endings have glutamatergic synapses (see e.g.
review by Akoev & Andrianov 1993). Many years later we had the privilege to start
cooperations with among other people Hans Braun (Marburg), Klaus Schäfer
(Stuttgart), and Yuri Andrianov (St. Petersburg), experts on the ampullary system.
Only this time we did not work on ampullae of Lorenzini, but on the micro-ampullae
in freshwater catfish. For single unit recordings in catfish we injected sometimes a
muscle relaxant, but most of the time the steroid Saffan, a mixture of alphadolon
and alphaxolon. Eventually, on recommendation of Kirsty Grant (Gif-sur-Yvette), we
used etomidate added to the water as sedative to immobilize the catfish. In this
way, even single unit activity of ampullary organs could be recorded non-invasively
in immobilized catfish receiving artificial respiration, thus providing an excellent
system for teaching sensory neurophysiology. After a successful pilot experiment
with unexperienced students, tutored by analyst Wim J.G. Loos, ampullary
electroreceptor organs in catfish have been used at Comparative Physiology at
Utrecht University for more than 20 years in all practicals on neurophysiology (Peters
et al. 1988). Dijkgraaf's accidental observation on dogfish in 1935 had evolved into
an elementary instruction experiment in 1988.
Serendipity, search image, prejudice, and the open mind
Dijkgraaf's observation of the dogfish turning its head away from an iron wire (1935),
was more or less coincidence, as was Sir John Eccles' passage through the
Netherlands in 1969. Both events provided nevertheless essential keys to the
elucidation of the biological significance of the electrosensitivity of the ampullae of
Lorenzini. My own in situ recordings of the ampullary nerve are perhaps also a
typical example of serendipity: after having already accepted the failure of making a
successful implantation, I found beautiful responses to electrical stimuli, whereas I
had only the intention to check whether or not the dogfish had survived my surgery.
It is rather confronting to recognize that expertise can lead to particular search
images, which may turn out to be prejudices, hampering the discovery of what one is
looking for. Because Dijkgraaf's search image was that the lateral line system was
mechanoreceptive, it took him a very long time to enter the world of
62

SDT Reference Series International 1, 54-65 title, Peters-RC, Dogfish Electroreception 1966______________________________________________________________________________________________________________________________________________________________________________________
electroreception. Also Kalmijn's expectation that the general anesthetic MS222 was a
central anesthetic only, which would leave the synapses of the ampullary system
untouched, took time that could have been used to investigate the responses of the
Lorenzinian ampullae to pieces of food and prey under 'free-swimming' conditions.
It is also revealing that in the world of sensory physiologists, electrical potential
differences belong to the world of nerve and muscle fibers, whereas the standing DC
potentials, mentioned by Sir John Eccles, belong to the expertise of chemical
physiological transport scientists. Apparently roaming and scavenging through the
sciences is a 'must' for innovation.
Halfway the 19th century, the eminent physiologist Johannes Müller wrote about the
senses: The manifestation of different objects to each other cannot express the
nature of light ; that it renders objects visible to us depends merely on our having an
organ of vision with vital properties. And in this way many other agents have the
same power of rendering objects manifest: were we endowed with as delicate an
organic re-agent for electricity as for light, electricity would have the same influence
as light in rendering manifest the corporeal world (Müller 1842, p. 1068).
A whole century later, the search image for the ampullae of Lorenzini was still
traditional. The simple idea that there could exist sensory organs typically adapted
to very weak electrical stimuli was too far fetched. Even after successful
electrophysiologcal recordings (e.g. Hensel 1955, 1956; Murray 1957, 1959, 1960,
1962), the biological significance remained enigmatic. Sir John Eccles' advice to have
a look at standing DC potentials in the pharynx of fishes, marks in my view the
turning point in the understanding of the biological significance of the ampullae of
Lorenzini. After Sir John Eccles' suggestion, von Uexküll's trininy (1909) could be
completed: 1) the physiology of the sensory organ was pretty well understood, 2)
appropriate electrical stimuli had been recognized, and 3) behaviour contributing to
survival had been described. Looking back: It took some time, but it was fun!
References
Akoev-GN, Andrianov-GN 1993. Sensory hair cells, synaptic transmission. Springer,
Berlin.
Delgado-JMR 1964. Electrodes for extracellular recording and stimulation,
pp. 88-143. in: Nastuk-WL 1964, Physical techniques in biological research
V. Academic Press, New York and London. Blz 122, fig 12.
Dijkgraaf-S 1956. Elektrophysiologische Untersuchungen an der Seitenlinie von
Xenopus laevis. Experientia 12, 276-278.
Dijkgraaf-S, Kalmijn-AJ 1962. Verhaltensversuche zur Funktion der Lorenzinischen
Ampullen. Naturwiss. 49, 400.
Dijkgraaf-S, Kalmijn-AJ 1966. Versuche zur biologischen Bedeutug der Lorenzinischen
Ampullen bei den Elasmobranchoern. Z. vergl Physiol. 53, 187-194.
Dotterweich-H 1932. Bau und Funktion der Lorenzinischen Ampullen. Zool. Jahrb.
Abt. Phys. 50, 347-418.
Görner-P, Coombs-S, Atema-J 1996. Sven Dijkgraaf 1908-1998. Brain Behav. Evol.
48, 350-358.
Hensel-H 1955. Quantitative Beziehugen zwischen Temperaturreiz und Aktions-
potentialen der Lorenzinischen Ampullen. Z. vergl Physiol. 37, 509-526.
Hensel-H 1956. Die Wirkung thermischer und mechanischer Reize auf die
Lorenzinischen Ampullen der Selachiern. Pflügers Archiv ges. Physiol. 263,
48-53.
Kalmijn-AJ 1972. Bioelectric fields in sea water and the function of the ampullae of
Lorenzini in elasmobranch fishes. SIO Reference Series 72-83, 1-21.
Krijgsman-BJ 1940. Untersuchungen über die Erregungsleitung in
Gastropodennerven. Proc. Kon. Ned. Akad. Wetensch. 43 (8), 1065-1077.
Krijgsman-BJ 1947. Physiologisch onderzoek met den electronenstraaloscillograaf.
N.V. Philips, Eindhoven.
Lissmann-HW 1951. Continuous electric signals from the tail of a fish, Gymnarchus
niloticus Cuv. Nature (London) 167, 201.
63

SDT Reference Series International 1, 54-65 title, Peters-RC, Dogfish Electroreception 1966______________________________________________________________________________________________________________________________________________________________________________________
Lissmann-HW 1958. On the function and evolution of electric organs in fish. J. Exp.
Biol. 35, 156-191.
Müller-J 1842. Elements of Physiology II. Taylor and Walton, London; page 1068.
Murray-RW 1957. Evidence for a mechanoreceptive function of the ampullae of
Lorenzini. Nature 179(4550), 106-107.
Murray-RW 1959. The response of the ampullae of Lorenzini to combined stimulation
by temperature change and weak direct currents. J. Physiol (London) 146(1),
1-13.
Murray-RW 1960. The response of the ampullae of Lorenzini to mechanical
stimulation. J. Exp. Biol. 37(2), 417-424.
Murray-RW 1962. Response of ampullae of Lorenzini of Elasmobranchs to electrical
stimulation. J. Exp. Biol. 39(1), 119-128.
Parker-GH, Heusen-AP-van 1917. The responses of the catfish, Amiurus nebulosus, to
metallic and non-metallic rods. Amer. J. Physiol. 44, 405-420.
Peters-RC, Teunis-PFM, Bretschneider-F, Weerden-R-van 1988. Ampullary
electroreceptors in neurophysiological instruction. J. Biol. Education 22(1),
61-66.
Peters-RC, Van-Honk-C, Bretschneider-F 2001. Variability of the bioelectric field of the
catfish Ictalurus nebulosus (Le Sueur, 1819; Pisces, Teleostei, Siluriformes).
Belgian J. Zool. 131(supplement 2), 123-126.
Peters-RC, Wessel, T-van, Wollenberg-JW-van-den, Bretschneider-F, Olijslager-AE
2002. The bioelectric field of the catfish Ictalurus nebulosus. J. Physiol.
(Paris). 96(5-6), 397-404.
Uexküll-J-von 1909. Umwelt und Innenwelt der Tiere. Julius Springer, Berlin.
Acknowledgments
Perhaps this is a good place to recognize how much I owe to my very demanding,
but excellent tutor Ad J. Kalmijn, but also how much I owe to my colleague Franklin
Bretschneider, and to all our students. I should not forget Jelle Atema, who
kidnapped me on the back of his motorbike in 1963, in order to sign a student-
assistant contract for the 3rd year practical of Comparative Physiology. I still feel his
enthusiastic aura. I should also thank the remembrance of my promotor, professor
Sven Dijkgraaf, who after a slight hesitation, retired with the knowledge that
teaching electrophysiology was finally in good hands: those of Franklin Bretschneider
and myself. This paper would not have been possible without access to the archive
of Stichting De Traditie.
64

SDT Reference Series International 1, 54-65 title, Peters-RC, Dogfish Electroreception 1966______________________________________________________________________________________________________________________________________________________________________________________
Appendix
Figure 11. About two decennia later another type of waterproof connector was used: a piece of an Amphenol strip connector and vaseline turned out to be both a waterproof, and an easy and good connection (Coen Ballintijn, Groningen, personal communication). The adhesive used to glue the connector to the skin hardened in about 10 minutes. Reference: Peters-RC, Evers-HP 1985. Frequency selectivity in the ampullary system of an Elasmobranch fish (Scyliorhinus canicula). J. Exp. Biol. 118, 99-109.
Ampullary essentials – abbreviated timeline
yyyymmdd
19320223 Dijkgraaf presents 'Referat' on the Lorenzinian ampullae in München
19350614 Dijkgraaf observes the sensitivity of dogfish to corrosion currents
19511217 Dijkgraaf writes to Lissmann about his observation and about the electric sensitivity of catfish, as discovered by Parker & van Heusen.
19530924 Dijkgraaf correponds with Hensel about the biological significance of thermo- and mechanosensitivity of dogfish
19600712 Dijkgraaf asks Murray about plans re electrophysiology and behaviour
19600714 Murray writes that he already found electrosensitivity for the ampullae
19611016 Murray-RW 1962. The response of the ampullae of Lorenzini of elasmobranchs to electrical stimulation. J. Exp. Biol. 39, 119-128
19620525 Dijkgraaf-S, Kalmijn-AJ 1962. Verhaltensversuche zur Funktion der Lorenzinischen Ampullen. Naturwiss. 49, 400.
19630425 Dijkgraaf determines electrical detection threshold for dogfish in Villefranche
19630704 Dijkgraaf to Lissmann: found in Villefranche that ampullae are electroreceptors
19640201 Dijkgraaf-S 1964. Electroreception and the ampullae of Lorenzini in Elasmobranchs. Nature (London) 201, 523.
19661210 Kalmijn-AJ 1966. Electro-perception in sharks and rays. Nature (London) 212, 1232-1233.
19660323 Rob Peters records ampullary activity in freely swimming dogfish
19660414 Lindauer invites Dijkgraaf to write a paper for 'Festschrift von Frisch'
19660606 Manuscript for Festschrift accepted: Dijkgraaf-S, Kalmijn-AJ 1966. Versuche zur biologischen Bedeutug der Lorenzinischen Ampullen bei den Elasmobranchoern. Z. vergl Physiol. 53, 187-194.
19680520 Dijkgraaf to Lissmann on inadequacy of ampullae as electroreceptors, as suggested in J. Exp. Biol. 135, p 183.
19680522 Lissmann to Dijkgraaf about reasons to consider muscle potentials as putative biologically adequate stimuli
1969 Sir John Eccles suggests pharynx DC potentials as adequate stimuli
65


















