
DNA flow cytometry of human spermatozoa: Consistent stoichiometric staining of sperm DNA using a...
6
DNA Flow Cytometry of Human Spermatozoa: Consistent Stoichiometric Staining of Sperm DNA Using a Novel Decondensation Protocol Tam as Kov acs, 1 Gyo ¨ngyi Be ´ke ´si, 1 A ´ kos F abi an, 2 Zsuzsa R akosy, 3,4 G abor Horv ath, 2 L aszlo ´M atyus, 2 Margit Bal azs, 3,4 * Attila Jenei 2 Abstract Rapid flow cytometric measurement of the frequency of aneuploid human sperms is in increasing demand but development of an exploitable method is hindered by difficul- ties of stoichiometric staining of sperm DNA. An aggressive decondensation protocol is needed after which cell integrity still remains intact. We used flow cytometry to exam- ine the effect of lithium diiodosalicylate (LIS, chaotropic agent) on fluorescence inten- sity of propidium iodide-treated human spermatozoa from 10 normozoospermic men. When flow cytometric identification of diploid spermatozoa was achieved, validation was performed after sorting by three-color FISH. In contrast with the extremely vari- able histograms of nondecondensed sperms, consistent identification of haploid and diploid spermatozoa was possible if samples were decondensed with LIS prior to flow cytometry. A 76-fold enrichment of diploid sperms was observed in the sorted fractions by FISH. A significant correlation was found between the proportion of sorted cells and of diploid sperms by FISH. Application of LIS during the preparation of sperm for flow cytometry appears to ensure the stoichiometric staining of sperm DNA, making quanti- fication of aneuploid sperm percentage possible. To our knowledge this is the first report in terms of separating spermatozoa with confirmedly abnormal chromosomal content. High correlation between the proportion of cells identified as having double DNA content by flow cytometry and diploid sperm by FISH allows rapid calculation of diploidy rate. ' 2008 International Society for Advancement of Cytometry Key terms diploid spermatozoa; aneuploid spermatozoa; propidium iodide; lithium diiodosalicy- late; DNA FACS; FISH EVEN in fertile, healthy men at least 6% of spermatozoa carry an abnormal number of chromosomes (1). After recognizing that routinely used parameters of human sperm quality (2) are not closely related to the occurrence of abnormal DNA content in spermatozoa (3–5), a number of techniques were introduced in both research and clinical practice. Flow cytometry offers a rapid and simple way to determine the sperm count (6) and degree of DNA damage (7,8), but the precise measurement of DNA content encounters difficulties arising from the special flattened shape of human sperm cells and from the tightly packed nature of sperm DNA. Distortion coming from the flatness can be solved by using specially shaped flow cytometric injection tubes (9). Subsequent investigations repeatedly failed to identify haploid, diploid, or aneuploid spermatozoa consistently, but it rather identified groups of patients with different patterns of fluorescence intensity (FI), some of them having a continuous, almost even distribution of the number of cells over a wide range of FI (6,10,11). Although certain cell population’s values on these histograms were related to haploid, diploid, and even tetraploid cells, cell sorting was not performed; there- fore, the above assumptions could not be confirmed directly. 1 Department of Obstetrics and Gynecology, University of Debrecen Medical and Health Science Center, Debrecen, Hungary 2 Department of Biophysics and Cell Biology, University of Debrecen Medical and Health Science Center, Debrecen, Hungary 3 Department of Preventive Medicine, University of Debrecen Medical and Health Science Center, Debrecen, Hungary 4 Hungarian Academy of Sciences, University of Debrecen Public Health Research Group, Debrecen, Hungary Received 21 March 2008; Accepted 20 June 2008 Grant sponsor: Hungarian Ministry of Health; Grant number: ETT 534/2003; Grant sponsor: Hungarian Scientific Research Fund; Grant number: OTKA T 04875, Bolyai Fellowship to A.J.; Agency for Research Fund Management and Research Exploitation (KPI) Genomnanotech-DEBRET 06/2004; Bolyai Fellowship to A.J. *Correspondence to: Margit Bal azs Ph.D., D.Sc., Division of Biomarker Analysis, Department of Preventive Medicine, Faculty of Public Health, Medical and Health Science Center, University of Debrecen, 4028 Debrecen Kassai str.26/b, Hungary Email: [email protected] Published online 31 July 2008 in Wiley InterScience (www.interscience.wiley.com) DOI: 10.1002/cyto.a.20618 © 2008 International Society for Advancement of Cytometry Technical Note Cytometry Part A 73A: 965970, 2008
-
Upload
tamas-kovacs -
Category
Documents
-
view
212 -
download
0
Transcript of DNA flow cytometry of human spermatozoa: Consistent stoichiometric staining of sperm DNA using a...

DNA Flow Cytometry of Human Spermatozoa:
Consistent Stoichiometric Staining of Sperm DNA
Using a Novel Decondensation Protocol
Tam�as Kov�acs,1 Gyongyi Bekesi,1 Akos F�abi�an,2 Zsuzsa R�akosy,3,4 G�abor Horv�ath,2
L�aszlo M�atyus,2 Margit Bal�azs,3,4* Attila Jenei2
� AbstractRapid flow cytometric measurement of the frequency of aneuploid human sperms is inincreasing demand but development of an exploitable method is hindered by difficul-ties of stoichiometric staining of sperm DNA. An aggressive decondensation protocol isneeded after which cell integrity still remains intact. We used flow cytometry to exam-ine the effect of lithium diiodosalicylate (LIS, chaotropic agent) on fluorescence inten-sity of propidium iodide-treated human spermatozoa from 10 normozoospermic men.When flow cytometric identification of diploid spermatozoa was achieved, validationwas performed after sorting by three-color FISH. In contrast with the extremely vari-able histograms of nondecondensed sperms, consistent identification of haploid anddiploid spermatozoa was possible if samples were decondensed with LIS prior to flowcytometry. A 76-fold enrichment of diploid sperms was observed in the sorted fractionsby FISH. A significant correlation was found between the proportion of sorted cells andof diploid sperms by FISH. Application of LIS during the preparation of sperm for flowcytometry appears to ensure the stoichiometric staining of sperm DNA, making quanti-fication of aneuploid sperm percentage possible. To our knowledge this is the firstreport in terms of separating spermatozoa with confirmedly abnormal chromosomalcontent. High correlation between the proportion of cells identified as having doubleDNA content by flow cytometry and diploid sperm by FISH allows rapid calculation ofdiploidy rate. ' 2008 International Society for Advancement of Cytometry
� Key termsdiploid spermatozoa; aneuploid spermatozoa; propidium iodide; lithium diiodosalicy-late; DNA FACS; FISH
EVEN in fertile, healthy men at least 6% of spermatozoa carry an abnormal number
of chromosomes (1). After recognizing that routinely used parameters of human
sperm quality (2) are not closely related to the occurrence of abnormal DNA content
in spermatozoa (3–5), a number of techniques were introduced in both research and
clinical practice. Flow cytometry offers a rapid and simple way to determine the
sperm count (6) and degree of DNA damage (7,8), but the precise measurement of
DNA content encounters difficulties arising from the special flattened shape of
human sperm cells and from the tightly packed nature of sperm DNA. Distortion
coming from the flatness can be solved by using specially shaped flow cytometric
injection tubes (9). Subsequent investigations repeatedly failed to identify haploid,
diploid, or aneuploid spermatozoa consistently, but it rather identified groups of
patients with different patterns of fluorescence intensity (FI), some of them having a
continuous, almost even distribution of the number of cells over a wide range of FI
(6,10,11). Although certain cell population’s values on these histograms were related
to haploid, diploid, and even tetraploid cells, cell sorting was not performed; there-
fore, the above assumptions could not be confirmed directly.
1Department of Obstetrics and Gynecology,University of Debrecen Medical andHealth Science Center, Debrecen, Hungary2Department of Biophysics and Cell Biology,University of Debrecen Medical and HealthScience Center, Debrecen, Hungary3Department of Preventive Medicine,University of Debrecen Medical andHealth Science Center, Debrecen,Hungary4Hungarian Academy of Sciences,University of Debrecen Public HealthResearch Group, Debrecen, Hungary
Received 21 March 2008; Accepted 20June 2008
Grant sponsor: Hungarian Ministry ofHealth; Grant number: ETT 534/2003;Grant sponsor: Hungarian ScientificResearch Fund; Grant number: OTKA T04875, Bolyai Fellowship to A.J.; Agencyfor Research Fund Management andResearch Exploitation (KPI)Genomnanotech-DEBRET 06/2004;Bolyai Fellowship to A.J.
*Correspondence to: Margit Bal�azsPh.D., D.Sc., Division of BiomarkerAnalysis, Department of PreventiveMedicine, Faculty of Public Health,Medical and Health Science Center,University of Debrecen, 4028 DebrecenKassai str.26/b, Hungary
Email: [email protected]
Published online 31 July 2008 in WileyInterScience (www.interscience.wiley.com)
DOI: 10.1002/cyto.a.20618
© 2008 International Society forAdvancement of Cytometry
Technical Note
Cytometry Part A � 73A: 965�970, 2008

For these reasons flow cytometry is considered rather a
tool automating conventional sperm analysis and for assessing
sperm DNA damage in general (12), than a technique to deter-
mine the proportion of sperms with numerical chromosome
abnormalities. However, the higher costs and the long hours
of scoring complicating the most widely used method (fluo-
rescence in situ hybridization, FISH) stimulate researchers to
implement the essentially simple, cheap, and rapid technique
of flow cytometry to achieve satisfactory estimation of sperms
with abnormal chromosome number.
During the initiation of this study we hypothesized that
the troublesome effect of the variable degree of DNA conden-
sation can be eliminated, and so a stoichiometric staining of
sperm DNA can be achieved by making it fully accessible for
fluorescent dyes through an aggressive decondensation of
spermatozoa, as with the technique we successfully used in
FISH experiments earlier (13). Furthermore, swelling of sperm
heads during this procedure is also expected to change their
flattened shape in a way that would reduce problems arising
from orientation during flow cytometry. As soon as consistent
identification of chromosomally abnormal spermatozoa is
possible, fluorescence activated cell sorting (FACS) can be per-
formed and the sorted spermatozoa can be examined directly
for aneuploidy and diploidy by FISH.
MATERIALS AND METHODS
Sample Preparation for Flow Cytometry
Fractions of leftover portions of semen samples provided
by 10 healthy normozoospermic sperm donors (some of them
with proven fertility) for routine semen analysis were washed
and centrifuged at 260g for 5 min twice in 10 mmol/l HEPES
buffer, containing 5% bovine serum albumin, (pH 5 8). DNA
Prep Stain (DNA Prep Reagent Kit, Beckman Coulter, Fuller-
ton, CA) containing propidium iodide was added following
permeabilization according to manufacturer’s protocol. The
samples from Patients 1–4 were then diluted with a triple vol-
ume of either 50 mmol/l lithium diiodosalicylate (LIS, result-
ing in a concentration of 37.5 mmol/l) or distilled water and
incubated at 378C for 15 min then kept on ice for 30–60 min
while transferring to the flow cytometer. Samples from Sub-
jects 2 and 3 were also treated with different concentrations of
LIS (37.5 and 75 mmol/l) at 20 and 378C, for 5–180 min. Ali-
quots diluted and treated similarly with buffer at the same
temperature were used as controls. Samples from Patients 4–
10 were processed using 37.5 mmol/l LIS at 378C for 10 min,
and then FACS was performed on them.
Flow Cytometry
Flow cytometry was performed on a Digital Vantage flow
cytometer (Becton Dickinson, San Jose, CA) equipped with an
argon ion laser emission at 488 nm. FI was recorded using a
575 � 10 nm filter. Reflex software was used for data process-
ing. Histograms of cell count per fluorescence channel (with
256 intensity channels) were used to identify the population
of interest. Data were expressed as percentage of the total
number of cells gated as haploid, possibly hyperhaploid, and
diploid. A total of 2–6 million events were acquired. Lower
limit of the sorting gate was identified as the FI with the mini-
mal cell count per channel between the haploid and diploid
peaks on the histogram, the upper limit was set at the appar-
ent change of steepness (Fig. 1, arrows). Spermatozoa with FI
between these points were separated.
The sorted fractions were centrifuged gently at 260 g for
10 min, and then the pellet was smeared on glass slides and
fixed with methanol-acetic acid mixture (3:1 ratio). The slides
were then dehydrated in a series of 70, 80, and 100% ethanol,
air dried and stored at 2208C until FISH was performed.
Smears of the original semen were also prepared and treated
similarly.
Sperm slides were warmed to room temperature, and
decondensed with 10 mmol/l dithiothreitol (DTT, Sigma Che-
micals Inc. St. Louis, MO) in 0.1 mol/l Tris-HCl, pH 8.0 for
30 min. Slides with the original specimen were also treated
with 10 mmol/l LIS in Tris-HCl for 1–3 h.
FISH
Multicolor FISH was preformed in order to detect the fre-
quency of disomies of the sex and the autosomal chromo-
somes and to differentiate between disomies and diploidies.
Chromosomes X, Y, and 17 centromere-specific DNA probes
were applied simultaneously. DNA probes were isolated from
bacteria clones generously supplied by Molecular Genetic
Resources (University of Bari, Bari, Italy). X and Y chromo-
some-specific probes were labeled by nick translation using bi-
otin- and digoxigenin-labeled dUTP according to the protocol
of the supplier (ROCHE Diagnostics GmbH, Applied Science,
Germany). Probe for the centromeric region of chromosome
17 was labeled with biotin and digoxigenin dUTP, and applied
in a 1:1 ratio during hybridization. For the visualization FITC-
avidin and anti-digoxigenin-TexasRed were used. The fluores-
cent signals for chromosome 17 appeared as a combination of
two colors (in our case red and green resulted in orange).
Sperm samples were simultaneously denatured with the
DNA probe mixture (containing biotin-labeled X, digoxi-
genin-labeled Y, and digoxigenin-biotin-labeled 17) at 788Cfor 7 min in a hybridization oven (HYBriteTM Denaturation/
Hybridization System, Abbott Molecular Inc.) Overnight
hybridizations were performed at 378C. Posthybridization
washings were done as described earlier (13,14). Slides were
counterstained with DAPI (40,6-Diamidino-2-phenylindole,
Vector Laboratoratories, Inc.) in an antifade.
For each individual, two samples (sorted fractions and
the original sperm sample) were hybridized. The overall hybri-
dization efficiency was [98% for each probe. Sperm nuclei
were scored according to published criteria (15). The fluores-
cent signals were scored by two independent investigators on
each slide. Overlapping nuclei and cells without fluorescent
signals were excluded from the analysis. In the case of aneu-
ploidy, the presence of the sperm tail was confirmed. A sper-
matozoon was considered disomic when it showed two fluo-
rescent signals for the same chromosome, comparable in size
and brightness in the approximately same focal plane, and
clearly positioned inside the edge of the sperm head at least
TECHNICAL NOTE
966 DNA Flow Cytometry of Human Spermatozoa
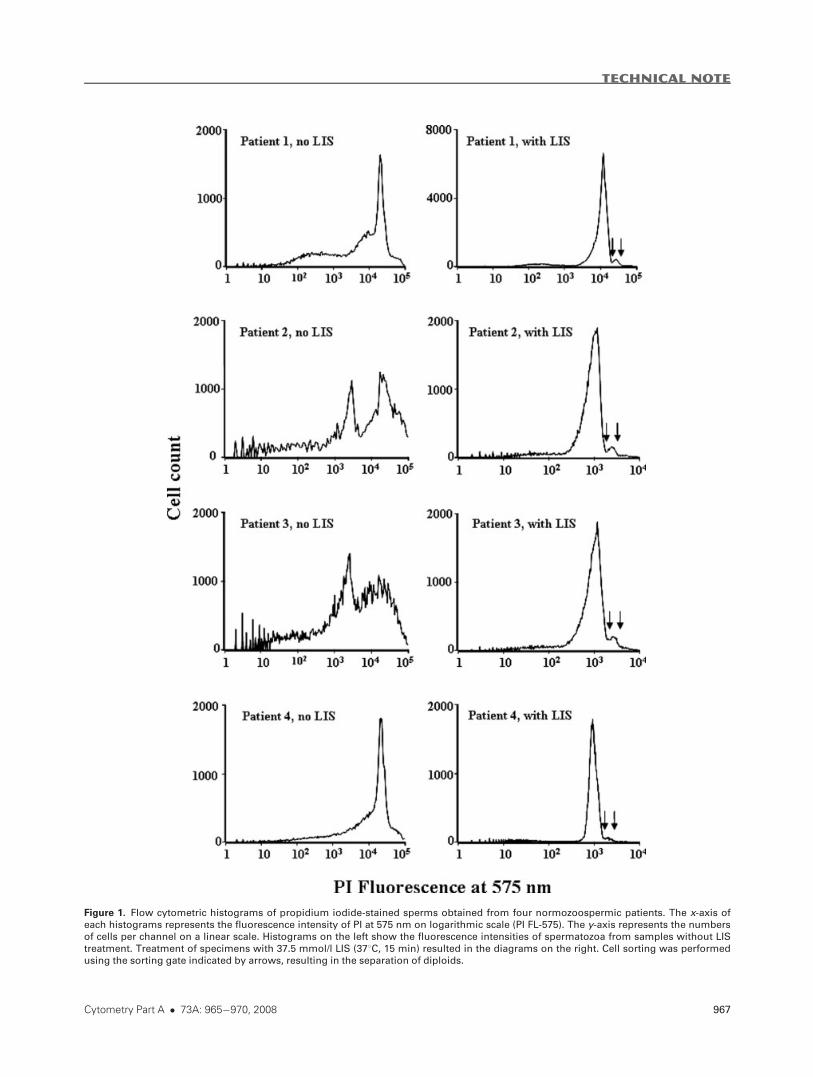
Figure 1. Flow cytometric histograms of propidium iodide-stained sperms obtained from four normozoospermic patients. The x-axis of
each histograms represents the fluorescence intensity of PI at 575 nm on logarithmic scale (PI FL-575). The y-axis represents the numbers
of cells per channel on a linear scale. Histograms on the left show the fluorescence intensities of spermatozoa from samples without LIS
treatment. Treatment of specimens with 37.5 mmol/l LIS (378C, 15 min) resulted in the diagrams on the right. Cell sorting was performedusing the sorting gate indicated by arrows, resulting in the separation of diploids.
TECHNICAL NOTE
Cytometry Part A � 73A: 965�970, 2008 967
one domain apart. Sperms showing disomy both for chromo-
some 17 and sex chromosomes were considered as diploids,
because the frequency of real multiple disomies is negligible if
compared to that of diploids (16).
Scoring was performed on a fluorescent microscope
(OPTON, Oberkochen, Germany) equipped with selective
filters for the detection of FITC, TexasRed, and DAPI. Aneu-
ploid or diploid spermatozoa were always examined with a
phase-contrast objective to verify the presence of the tail and
to exclude apparent diploidy in two spermatozoa in close
proximity.
Statistical Analysis
Proportions of diploid sperms in the original and in the
sorted samples were compared, and correlation between the
proportion of the diploids by FISH in the original samples
and the ratio of sorted cells to the whole number of cells was
calculated using Spearman correlation.
RESULTS
All samples were normozoospermic by World Health Or-
ganization criteria (sperm concentration [20 million/l, pro-
gressive motility[60 %, WHO, 1999). Proportion of diploid
spermatozoa showing 1717XX, 1717YY or 1717XY signals by
FISH (presumed karyotype: 46,XX; 46,YY; 46,XY, respectively)
was in the range of 0.05–0.24%, and was considered normal.
Significant heterogeneity was observed in FI distribution
histograms in those PI-labeled sperm samples that had not
been treated with the chaotropic agent LIS as shown on the
left side diagrams of Figure 1. If LIS was applied as described
earlier, patterns from all the four subjects became similar to
each other as shown on the right side diagrams.
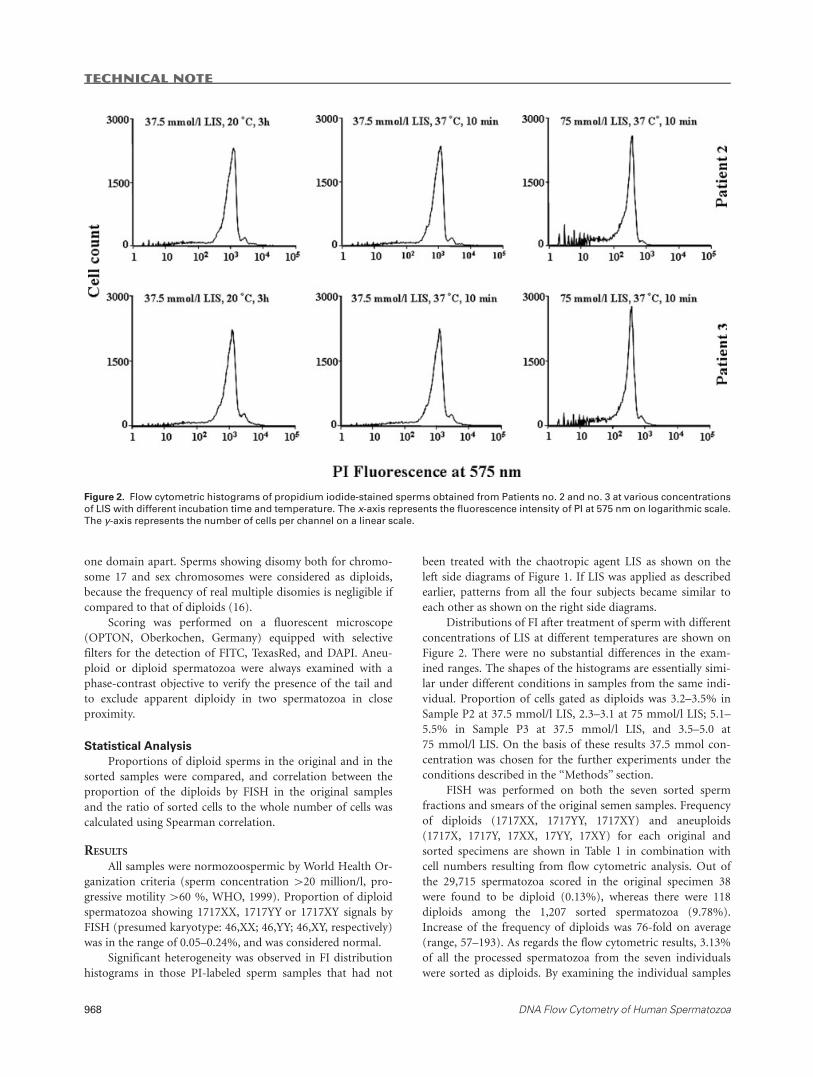
Distributions of FI after treatment of sperm with different
concentrations of LIS at different temperatures are shown on
Figure 2. There were no substantial differences in the exam-
ined ranges. The shapes of the histograms are essentially simi-
lar under different conditions in samples from the same indi-
vidual. Proportion of cells gated as diploids was 3.2–3.5% in
Sample P2 at 37.5 mmol/l LIS, 2.3–3.1 at 75 mmol/l LIS; 5.1–
5.5% in Sample P3 at 37.5 mmol/l LIS, and 3.5–5.0 at
75 mmol/l LIS. On the basis of these results 37.5 mmol con-
centration was chosen for the further experiments under the
conditions described in the ‘‘Methods’’ section.
FISH was performed on both the seven sorted sperm
fractions and smears of the original semen samples. Frequency
of diploids (1717XX, 1717YY, 1717XY) and aneuploids
(1717X, 1717Y, 17XX, 17YY, 17XY) for each original and
sorted specimens are shown in Table 1 in combination with
cell numbers resulting from flow cytometric analysis. Out of
the 29,715 spermatozoa scored in the original specimen 38
were found to be diploid (0.13%), whereas there were 118
diploids among the 1,207 sorted spermatozoa (9.78%).
Increase of the frequency of diploids was 76-fold on average
(range, 57–193). As regards the flow cytometric results, 3.13%
of all the processed spermatozoa from the seven individuals
were sorted as diploids. By examining the individual samples
Figure 2. Flow cytometric histograms of propidium iodide-stained sperms obtained from Patients no. 2 and no. 3 at various concentrations
of LIS with different incubation time and temperature. The x-axis represents the fluorescence intensity of PI at 575 nm on logarithmic scale.
The y-axis represents the number of cells per channel on a linear scale.
TECHNICAL NOTE
968 DNA Flow Cytometry of Human Spermatozoa
it is seen that the higher the proportion of sorted spermatozoa
was, the more diploid spermatozoa were found in the original
semen. The correlation coefficient was R 5 0.857 (P\ 0.01).
There was no correlation with the disomy rate or with the
total numerical abnormality rate.
DISCUSSION
The purpose of our study was to establish a flow cyto-
metric technique for consistent identification and quantifica-
tion of diploid spermatozoa in the human sperm. This is the
first study providing direct evidence for the correct identifica-
tion of diploids by using flow cytometry, which enables us to
estimate the diploidy rate of the original semen sample.
By introducing the chaotropic agent LIS for preparation
of spermatozoa for flow cytometry we achieved a consistent
and reproducible pattern of FI in samples from normozoos-
permic fertile men. This reflects similar DNA content of sper-
matozoa from normal individuals with a similar proportion of
haploid and diploid sperms. These data, however, are in con-
trast with previous reports, where a wide range of variation of
DNA fluorescence was observed. Hacker-Klom et al. (6) cate-
gorized histograms of normozoospermic patients on the basis
of their shape into five groups (alongside three groups of his-
tograms from severely oligozoospermic patients), Levek-
Motola et al. (10) grouped all normozoospermic samples into
one group, showing a single peak of cells with some skewing
toward higher fluorescence intensity. They did not define
diploid sperm in the samples, whereas the previous authors
found diploidy rates of up to 12% in some patients with nor-
mal sperm count; unfortunately no FISH was performed to
confirm the presence of diploids. Although they used different
DNA dyes (DAPI and PI, respectively), it is unlikely that this
would affect the results to such an extent. Simultaneous appli-
cation of FISH and flow cytometry to assess proportion of
aneuploid and diploid cells in one case was reported by Weis-
senberg et al. (10) The shape and position of histograms sug-
gested the predominance of diploids in the sperm specimen,
which was confirmed by FISH in the original sample. How-
ever, it was not obvious how the proportion of diploids could
be correlated to the flow cytometric data, and no FACS was
performed.
High variability of proportion of cells on either side of
the more or less well-defined peak of normal haploid cells is
attributed to the various proportions of cellular debris (6)
and/or noncondensed haploid sperm (11). Others have also
tried to link the heterogeneity between samples to specific
sperm populations (17,18), but the chromosome alterations
were not investigated.
The high variability of staining of sperm DNA is attribu-
ted to the degree of DNA condensation in sperm nuclei (19).
During spermatogenesis, histones—common in somatic cell
nuclei—are replaced by protamine proteins. This process
results in a highly condensed chromatin which is transcrip-
tionally inactive (20,21). The histone-to-protamine transition
seems to be critical in terms of completion of functional devel-
opment of spermatozoa (22,23). Condensation of sperm DNA
results in not only complete inactivation but also in restricted
accessibility of DNA by most fluorochromes; therefore, stain-
ing of sperm nuclei using DNA-specific dyes is not stoichio-
metric during most of the stages of sperm maturation (24).
To achieve stoichiometric labeling, DNA must be thor-
oughly exposed to the fluorescent dye used or DNA must be
effectively decondensed. Agents commonly used for decon-
densation mainly change the protein structure around the
DNA helices (25). During this process some areas still remain
unavailable for fluorochromes, not allowing the DNA dye to
incorporate stoichiometrically. Chaotropic agents as LIS
(26,27) or guanidium thiocyanate (28) are effective to loosen
up the tightly packed DNA by dissolving its binding to nuclear
proteins like histones and, in spermatozoa, protamines. Free
DNA extracted by LIS surrounds the nuclear matrix, forming
a halo-like structure (29), still anchored by specific regions of
DNA to the nuclear matrix, thus maintaining more or less the
original appearance of the cell, but the bare DNA loops are
exposed completely to DNA probes. The membranes also
remain intact, which ensures the proper compartmentalization
of DNA needed for flow cytometry (30) and helps the accu-
racy of automated scoring (31). In our study a wide range of
LIS conditions resulted in uniform, consistent results. It is
noteworthy that increased concentrations of LIS produced
noticeably lower fluorescence intensities. This observation
may result from the inhibiting effect of LIS on PI-DNA bind-
Table 1. Three-color fluorescence in situ hybridization (FISH) for chromosomes 17, X and Y on spermatozoa of 7 normozoospermic men
(Patients No. 4-10) in the original semen and in fractions after fluorescence activated cell sorting (FACS) for diploidy
ALL SPERM
PROCESSED
(3103)
SORTED
(3103)
SORTED
(%)
ANALYZED BY FISH DIPLOIDS (%)
TOTAL NUMERICAL CHROMOSOMAL
ABNORMALITIES
(DIPLOIDS1 ANEUPLOIDS, %)
ORIGINAL SORTED ORIGINAL SORTED ORIGINAL SORTED
P4 7500 200 2.67 5037 49 6 (0.12) 6 (12.24) 14 (0.28) 27 (55.1)
P5 5500 180 3.27 5043 62 5 (0.10) 9 (14.52) 21 (0.42) 14 (22.58)
P6 4000 200 5.00 2642 458 4 (0.15) 41 (8.95) 8 (0.3) 98 (21.4)
P7 2380 39 1.64 4107 178 2 (0.05) 8 (4.49) 26 (0.63) 34 (19.1)
P8 3000 120 4.00 5040 65 12 (0.24) 15 (23.08) 38 (0.75) 20 (30.7)
P9 3500 100 2.86 5034 264 7 (0.14) 21 (7.95) 34 (0.68) 26 (9.85)
P10 4800 120 2.50 2812 131 2 (0.07) 18 (13.74) 10 (0.36) 25 (19.08)
Total 30680 959 3.13 29715 1207 38 (0.13) 118 (9.78) 151 (0.51) 244 (20.22)
TECHNICAL NOTE
Cytometry Part A � 73A: 965�970, 2008 969

ing and suggests that accessibility of DNA for PI cannot be
further increased, meaning that optimal stoichiometric stain-
ing has been achieved.
Earlier we applied LIS successfully for decondensation of
sperm cells for FISH (13). In this study we give evidence that
the highly variable distribution of DNA staining of sperm
nuclei becomes quite uniform after LIS treatment. The peaks
of these histograms are clearly associated with haploid and
diploid sperm populations. To verify that the smaller peak
with double intensity on the right side is really associated with
diploid sperms we sorted the cells by FACS from seven speci-
mens. The sorted fraction was validated by FISH. To our
knowledge this is the first report in terms of separating sper-
matozoa with confirmedly abnormal chromosomal content.
Although the marked increase of diploid cells in the
sorted specimens is obvious, the still high proportions of see-
mingly haploid 17X and 17Y sperm can be unsatisfactory at
first glance. The rate of enhancement of hyperhaploids may
also be surprising as they contain only about 3–8% more
DNA. However, these findings can be easily explained by sup-
posing the enhancement of disomies and even multiple diso-
mies of other chromosomes than chromosomes 17, X, and Y
in the sorted fractions.
In summary, the application of LIS during the prepara-
tion of sperm for flow cytometry appears to ensure the stoi-
chiometric staining of sperm DNA, making quantification of
diploid sperm possible. High correlation between the number
of sorted cells and diploidy rate allows setting up an algorithm
for easy and rapid determination of diploidy rate in sperm.
LITERATURE CITED
1. Egozcue J, Blanco J, Vidal F. Chromosome studies in human sperm nuclei using fluo-rescence in-situ hybridization(FISH). Hum Reprod Update 1997;3:441–452.
2. World Health Organization Laboratory Manual for the Examination of HumanSemen and Sperm-Cervical Mucus Interaction. Cambridge: University Press; 1999.
3. Ryu HM, Lin WW, Lamb DJ, Chuang W, Lipshultz LI, Bischoff FZ. Increased chro-mosome X, Y, and 18 nondisjunction in sperm from infertile patients that were iden-tified as normal by strict morphology: Implication for intracytoplasmic sperm injec-tion. Fertil Steril 2001;76:879–883.
4. Jakab A, Kovacs T, Zavaczki Z, Borsos A, Bray-Ward P, Ward D, Huszar G. Efficacy ofthe swim-up method in eliminating sperm with diminished maturity and aneuploidy.Hum Reprod 2003;18:1481–1488.
5. Egozcue S, Blanco J, Vendrell JM, Garcia F, Veiga A, Aran B, Barri PN, Vidal F, Egoz-cue J. Human male infertility: Chromosome anomalies, meiotic disorders, abnormalspermatozoa and recurrent abortion. Hum Reprod Update 2000;6:93–105.
6. Hacker-Klom UB, Gohde W, Nieschlag E, Behre HM. DNA flow cytometry of humansemen. Hum Reprod 1999;14:2506–2512.
7. Evenson DP, Jost LK, Marshall D, Zinaman MJ, Clegg E, Purvis K, de Angelis P,Claussen OP. Utility of the sperm chromatin structure assay as a diagnostic and prog-nostic tool in the human fertility clinic. Hum Reprod 1999;14:1039–1049.
8. Evenson DP, Wixon R. Clinical aspects of sperm DNA fragmentation detection andmale infertility. Theriogenology 2006;65:979–991.
9. Dean PN, Pinkel D, Mendelsohn ML. Hydrodynamic orientation of sperm heads forflow cytometry. Biophys J 1978;23:7–13.
10. Weissenberg R, Aviram A, Golan R, Lewin LM, Levron J, Madgar I, Dor J, Barkai G,Goldman B. Concurrent use of flow cytometry and fluorescence in-situ hybridizationtechniques for detecting faulty meiosis in a human sperm sample. Mol Hum Reprod1998;4:61–66.
11. Levek-Motola N, Soffer Y, Shochat L, Raziel A, Lewin LM, Golan R. Flow cytometryof human semen: A preliminary study of a non-invasive method for the detection ofspermatogenetic defects. Hum Reprod 2005;20:3469–3475.
12. Cordelli E, Eleuteri P, Leter G, Rescia M, Spano M. Flow cytometry applications inthe evaluation of sperm quality: Semen analysis, sperm function and DNA integrity.Contraception 2005;72:273–279.
13. Kovanci E, Kovacs T, Moretti E, Vigue L, Bray-Ward P, Ward DC, Huszar G. FISHassessment of aneuploidy frequencies in mature and immature human spermatozoaclassified by the absence or presence of cytoplasmic retention. Hum Reprod 2001;16:1209–1217.
14. Juhasz A, Balazs M, Sziklay I, Rakosy Z, Treszl A, Repassy G, Adany R. Chromosomalimbalances in laryngeal and hypopharyngeal cancers detected by comparative geno-mic hybridization. Cytometry A 2005;67A:151–160.
15. Martin RH, Rademaker A. Reliability of aneuploidy estimates in human sperm:Results of fluorescence in situ hybridization studies using two different scoring crite-ria. Mol Reprod Dev 1995;42:89–93.
16. Rademaker A, Spriggs E, Ko E, Martin R. Reliability of estimates of diploid humanspermatozoa using multicolour fluorescence in-situ hybridization. Hum Reprod1997;12:77–79.
17. Fossa SD, Melvik JE, Juul NO, Pettersen EO, Theodorsen L. DNA flow cytometry insperm cells from unilaterally orchiectomized patients with testicular cancer beforefurther treatment. Cytometry 1989;10:345–350.
18. Vicari E, Perdichizzi A, De Palma A, Burrello N, D’Agata R, Calogero AE. Globozoos-permia is associated with chromatin structure abnormalities: Case report. HumReprod 2002;17:2128–2133.
19. Fuentes-Mascorro G, Serrano H, Rosado A. Sperm chromatin. Arch Androl 2000;45:215–225.
20. Oliva R, Dixon GH. Vertebrate protamine genes and the histone-to-protamine repla-cement reaction. Prog Nucleic Acid Res Mol Biol 1991;40:25–94.
21. Aoki VW, Carrell DT. Human protamines and the developing spermatid: Their struc-ture, function, expression and relationship with male infertility. Asian J Androl2003;5:315–324.
22. Zhao M, Shirley CR, Hayashi S, Marcon L, Mohapatra B, Suganuma R, BehringerRR, Boissonneault G, Yanagimachi R, Meistrich ML. Transition nuclear proteins arerequired for normal chromatin condensation and functional sperm development.Genesis 2004;38:200–213.
23. Zhang X, San Gabriel M, Zini A. Sperm nuclear histone to protamine ratio in fertileand infertile men: Evidence of heterogeneous subpopulations of spermatozoa in theejaculate. J Androl 2006;27:414–420.
24. Santisteban MS, Montmasson MP, Giroud F, Ronot X, Brugal G. Fluorescence imagecytometry of nuclear DNA content versus chromatin pattern: A comparative study often fluorochromes. J Histochem Cytochem 1992;40:1789–1797.
25. Molina J, Castilla JA, Gil T, Hortas ML, Vergara F, Herruzo A. Influence of incubationon the chromatin condensation and nuclear stability of human spermatozoa by flowcytometry. Hum Reprod 1995;10:1280–1286.
26. Eberharter A, Grabher A, Gstraunthaler G, Loidl P. Nuclear matrix of the lowereukaryote Physarum polycephalum and the mammalian epithelial LLC-PK1 cell line.A comprehensive investigation of different preparation procedures. Eur J Biochem1993;212:573–580.
27. Mirkovitch J, Mirault M-E, Laemmli UK. Organization of the higher-order chro-matin loop: Specific DNA attachment sites on nuclear scaffold. Cell 1984;39:223–232.
28. Hossain AM, Rizk B, Behzadian A, Thorneycroft IH. Modified guanidinium thio-cyanate method for human sperm DNA isolation. Mol Hum Reprod 1997;3:953–956.
29. Fiorini A, Gouveia Fde S, Fernandez MA. Scaffold/matrix attachment regions andintrinsic DNA curvature. Biochemistry 2006;71:481–488.
30. Smith HC, Ochs RL, Lin D, Chinault AC. Ultrastructural and biochemical compari-sons of nuclear matrices prepared by high salt or LIS extraction. Mol Cell Biochem1987;77:49–61.
31. Perry MJ, Chen X, Lu X. Automated scoring of multiprobe FISH in human spermato-zoa. Cytometry A 2007;71A:968–972.
TECHNICAL NOTE
970 DNA Flow Cytometry of Human Spermatozoa


![DNA Fragmentation in Spermatozoa: A Historical Review · DNA fragmentation exceeds 30% [12,17]. Together, these studies provide important insight into the significance of DNA fragmentation](https://static.fdocuments.in/doc/165x107/5f3a6afb98469b5f937b34fa/dna-fragmentation-in-spermatozoa-a-historical-review-dna-fragmentation-exceeds.jpg)















