
DMD 40:529–538, 2012 Characterization of Hepatic and...
10
Characterization of Hepatic and Intestinal Glucuronidation of Magnolol: Application of the Relative Activity Factor Approach to Decipher the Contributions of Multiple UDP- Glucuronosyltransferase Isoforms Liangliang Zhu, Guangbo Ge, Hongbo Zhang, Huixin Liu, Guiyuan He, Sicheng Liang, Yanyan Zhang, Zhongze Fang, Peipei Dong, Moshe Finel, and Ling Yang Laboratory of Pharmaceutical Resource Discovery, Dalian Institute of Chemical Physics, Chinese Academy of Sciences, Dalian, China (L.Z., G.G., H.L., G.H., S.L., Y.Z., Z.F., P.D., L.Y.); Graduate University of Chinese Academy of Sciences, Beijing, China (L.Z., G.H.); and Center for Drug Research, Faculty of Pharmacy, University of Helsinki, Helsinki, Finland (H.Z., M.F.) Received August 4, 2011; accepted December 16, 2011 ABSTRACT: Magnolol is a food additive that is often found in mints and gums. Human exposure to this compound can reach a high dose; thus, characterization of magnolol disposition in humans is very impor- tant. Previous studies indicated that magnolol can undergo exten- sive glucuronidation in humans in vivo. In this study, in vitro assays were used to characterize the glucuronidation pathway in human liver and intestine. Assays with recombinant human UDP-glucu- ronosyltransferase enzymes (UGTs) revealed that multiple UGT isoforms were involved in magnolol glucuronidation, including UGT1A1, -1A3, -1A7, -1A8, -1A9, -1A10, and -2B7. Magnolol glucu- ronidation by human liver microsomes (HLM), human intestine microsomes (HIM), and most recombinant UGTs exhibited strong substrate inhibition kinetics. The degree of substrate inhibition was relatively low in the case of UGT1A10, whereas the reaction catalyzed by UGT1A9 followed biphasic kinetics. Chemical inhibi- tion studies and the relative activity factor (RAF) approach were used to identify the individual UGTs that played important roles in magnolol glucuronidation in HLM and HIM. The results indicate that UGT2B7 is mainly responsible for the reaction in HLM, whereas UGT2B7 and UGT1A10 are significant contributors in HIM. In summary, the current study clarifies the glucuronidation path- way of magnolol and demonstrates that the RAF approach can be used as an efficient method for deciphering the roles of individual UGTs in a given glucuronidation pathway in the native tissue that is catalyzed by multiple isoforms with variable and atypical kinetics. Introduction Magnolia officinalis is an herb medicine that has been used to treat many diseases such as nervous disturbance, abdominal disorders, gastrointestinal food stagnancy, abdominal distention, constipation, coughing, and dyspnea in Asian countries for thousands of years (Lee et al., 2011). Magnolol, the most abundant constitute in M. officinalis, is a natural biphenyl compound with a centro-symmetric structure containing two free phenol groups (Fig. 1). Much research has indi- cated that magnolol possesses various desired pharmacological activ- ities, especially as a potent agonist for peroxisome proliferator-acti- vated receptor-, a treatment target for type 2 diabetes and metabolic syndrome (Fakhrudin et al., 2010). Human are often exposed to magnolol in daily life because Mag- nolia bark supercritical carbon dioxide extract (containing 92.5% magnolol) is added to mints and gums to freshen breath (Greenberg et al., 2007). It is estimated that teenage exposure to magnolol can reach 1.64 mg/kg per day (data from http://www.food.gov.uk/multimedia/ pdfs/applicmagbarkextract.pdf). Previous studies indicated that mag- nolol can undergo extensive glucuronidation in human and rats (Hat- tori et al., 1986; Homma et al., 1993; Tsai et al., 1995). Considering the high dose intake of magnolol, characterizing its glucuronidation pathway is very important for understanding its beneficial and poten- tial harmful effects on humans. However, this pathway in humans is not yet well elucidated. The aim of this work was to identify the individual UDP-glucuronosyltransferases (UGTs) that can catalyze This work was supported by the National Natural Science Foundation of China [Grants 30973590, 81001473]; the National Key Technology Research and Development Pro- gram of China [Grant 2009BADB9B02]; the State Key Development Program of Basic Research of China [Grant 2009CB522808]; the National High Technology Research and Development Program of China [Grant 2009AA02Z205]; the Knowledge Innovation Pro- gram of the Chinese Academy of Sciences [Grant KSCX2-YW-G-050]; and the Sigrid Juselius Foundation, Finland. Article, publication date, and citation information can be found at http://dmd.aspetjournals.org. http://dx.doi.org/10.1124/dmd.111.042192. ABBREVIATIONS: UGT, UDP-glucuronosyltransferase; UDPGA, uridine diphosphate glucuronic acid; RAF, relative activity factor; AS-3201, (R)-()-2-(4-bromo-2-fluorobenzyl)-1,2,3,4-tetrahydropyrrolo[1,2-]pyrazine-4-spiro-3-pyrrolidine-1,2,3,5-tetrone; HPLC, high-performance liq- uid chromatography; HLM, human liver microsomes; AZT, 3-azido-3-deoxy-thymidine; HIM, human intestine microsomes; UFLC, ultrafast liquid chromatography; DAD, diode-array detector; MS, mass spectrometry; ESI, electrospray ionization; LC, liquid chromatography; SPE, solid-phase extraction. 1521-009X/12/4003-529–538$25.00 DRUG METABOLISM AND DISPOSITION Vol. 40, No. 3 Copyright © 2012 by The American Society for Pharmacology and Experimental Therapeutics 42192/3752072 DMD 40:529–538, 2012 529 at ASPET Journals on June 30, 2018 dmd.aspetjournals.org Downloaded from
Transcript of DMD 40:529–538, 2012 Characterization of Hepatic and...

Characterization of Hepatic and Intestinal Glucuronidation ofMagnolol: Application of the Relative Activity Factor Approach to
Decipher the Contributions of Multiple UDP-Glucuronosyltransferase Isoforms
Liangliang Zhu, Guangbo Ge, Hongbo Zhang, Huixin Liu, Guiyuan He, Sicheng Liang,Yanyan Zhang, Zhongze Fang, Peipei Dong, Moshe Finel, and Ling Yang
Laboratory of Pharmaceutical Resource Discovery, Dalian Institute of Chemical Physics, Chinese Academy of Sciences, Dalian,China (L.Z., G.G., H.L., G.H., S.L., Y.Z., Z.F., P.D., L.Y.); Graduate University of Chinese Academy of Sciences, Beijing, China
(L.Z., G.H.); and Center for Drug Research, Faculty of Pharmacy, University of Helsinki, Helsinki, Finland (H.Z., M.F.)
Received August 4, 2011; accepted December 16, 2011
ABSTRACT:
Magnolol is a food additive that is often found in mints and gums.Human exposure to this compound can reach a high dose; thus,characterization of magnolol disposition in humans is very impor-tant. Previous studies indicated that magnolol can undergo exten-sive glucuronidation in humans in vivo. In this study, in vitro assayswere used to characterize the glucuronidation pathway in humanliver and intestine. Assays with recombinant human UDP-glucu-ronosyltransferase enzymes (UGTs) revealed that multiple UGTisoforms were involved in magnolol glucuronidation, includingUGT1A1, -1A3, -1A7, -1A8, -1A9, -1A10, and -2B7. Magnolol glucu-ronidation by human liver microsomes (HLM), human intestinemicrosomes (HIM), and most recombinant UGTs exhibited strongsubstrate inhibition kinetics. The degree of substrate inhibition
was relatively low in the case of UGT1A10, whereas the reactioncatalyzed by UGT1A9 followed biphasic kinetics. Chemical inhibi-tion studies and the relative activity factor (RAF) approach wereused to identify the individual UGTs that played important roles inmagnolol glucuronidation in HLM and HIM. The results indicatethat UGT2B7 is mainly responsible for the reaction in HLM,whereas UGT2B7 and UGT1A10 are significant contributors in HIM.In summary, the current study clarifies the glucuronidation path-way of magnolol and demonstrates that the RAF approach can beused as an efficient method for deciphering the roles of individualUGTs in a given glucuronidation pathway in the native tissue that iscatalyzed by multiple isoforms with variable and atypical kinetics.
Introduction
Magnolia officinalis is an herb medicine that has been used to treatmany diseases such as nervous disturbance, abdominal disorders,gastrointestinal food stagnancy, abdominal distention, constipation,coughing, and dyspnea in Asian countries for thousands of years (Leeet al., 2011). Magnolol, the most abundant constitute in M. officinalis,is a natural biphenyl compound with a centro-symmetric structure
containing two free phenol groups (Fig. 1). Much research has indi-cated that magnolol possesses various desired pharmacological activ-ities, especially as a potent agonist for peroxisome proliferator-acti-vated receptor-�, a treatment target for type 2 diabetes and metabolicsyndrome (Fakhrudin et al., 2010).
Human are often exposed to magnolol in daily life because Mag-nolia bark supercritical carbon dioxide extract (containing �92.5%magnolol) is added to mints and gums to freshen breath (Greenberg etal., 2007). It is estimated that teenage exposure to magnolol can reach1.64 mg/kg per day (data from http://www.food.gov.uk/multimedia/pdfs/applicmagbarkextract.pdf). Previous studies indicated that mag-nolol can undergo extensive glucuronidation in human and rats (Hat-tori et al., 1986; Homma et al., 1993; Tsai et al., 1995). Consideringthe high dose intake of magnolol, characterizing its glucuronidationpathway is very important for understanding its beneficial and poten-tial harmful effects on humans. However, this pathway in humans isnot yet well elucidated. The aim of this work was to identify theindividual UDP-glucuronosyltransferases (UGTs) that can catalyze
This work was supported by the National Natural Science Foundation of China [Grants30973590, 81001473]; the National Key Technology Research and Development Pro-gram of China [Grant 2009BADB9B02]; the State Key Development Program of BasicResearch of China [Grant 2009CB522808]; the National High Technology Research andDevelopment Program of China [Grant 2009AA02Z205]; the Knowledge Innovation Pro-gram of the Chinese Academy of Sciences [Grant KSCX2-YW-G-050]; and the SigridJuselius Foundation, Finland.
Article, publication date, and citation information can be found athttp://dmd.aspetjournals.org.
http://dx.doi.org/10.1124/dmd.111.042192.
ABBREVIATIONS: UGT, UDP-glucuronosyltransferase; UDPGA, uridine diphosphate glucuronic acid; RAF, relative activity factor; AS-3201,(R)-(�)-2-(4-bromo-2-fluorobenzyl)-1,2,3,4-tetrahydropyrrolo[1,2-�]pyrazine-4-spiro-3�-pyrrolidine-1,2�,3,5�-tetrone; HPLC, high-performance liq-uid chromatography; HLM, human liver microsomes; AZT, 3�-azido-3�-deoxy-thymidine; HIM, human intestine microsomes; UFLC, ultrafast liquidchromatography; DAD, diode-array detector; MS, mass spectrometry; ESI, electrospray ionization; LC, liquid chromatography; SPE, solid-phaseextraction.
1521-009X/12/4003-529–538$25.00DRUG METABOLISM AND DISPOSITION Vol. 40, No. 3Copyright © 2012 by The American Society for Pharmacology and Experimental Therapeutics 42192/3752072DMD 40:529–538, 2012
529
at ASPE
T Journals on June 30, 2018
dmd.aspetjournals.org
Dow
nloaded from

magnolol glucuronidation and their respective contributions in humanliver and intestine.
The UGTs are membrane enzymes of the endoplasmic reticulumthat catalyze the conjugation of their substrates with glucuronic acidfrom UDP-glucuronic acid (UDPGA), making the substrates moresoluble and readily excreted (Wells et al., 2004). The human UGTisoforms are divided into three subfamilies (1A, 2A, and 2B) based onamino acids sequence identities and gene structure (Mackenzie et al.,2005). They are expressed in various tissues with a tissue-specificmanner. Liver and intestine are recognized as the two most importantglucuronidation sites for orally administrated drugs. Many UGTs areexpressed in both liver and intestine, but some of them exhibittissue-specific expression. For example, UGT1A10 is expressed insmall intestine but not in liver (Ohno and Nakajin, 2009).
UGTs exhibit distinct overlapping substrate specificities, indicatingthat the glucuronidation pathway of interest in native tissue (e.g., liverand intestine) is often catalyzed by several different UGTs. Evenmore, UGTs usually exhibit different affinity toward the “shared”substrate or even different kinetic models. This, in combination witha lack of selective UGT inhibitors, makes it difficult to performglucuronidation phenotyping. At present, glucuronidation phenotyp-ing depends heavily on assays with recombinant UGT isoforms.However, there are significant activity differences between nativetissue microsomes and recombinant enzymes. Results from recombi-nant enzyme assays will not accurately reflect the true contributions ofindividual UGTs in native tissues. One possible way to estimate thecontributions of individual UGTs to a given reaction in tissue micro-somes is to use the relative activity factor (RAF) approach.
The RAF approach was first developed to reflect the differencesbetween recombinant cytochromes P450 and human liver micro-somes, and it has been proved to be a powerful tool in cytochromeP450 reaction phenotyping (Crespi and Miller, 1999). For UGTreaction phenotyping, the RAF approach was seldom used, possi-bly because of the lack of specific substrates. Because glucuroni-dation metabolism is drawing increasing attention, more and morespecific substrates for UGTs are being reported. Although a full setof specific substrates is not available yet (Miners et al., 2010), theRAF approach was used occasionally in UGT phenotyping. To ourknowledge, the RAF approach was once used to clarify the glucu-ronidation pathway of buprenorphine, norbuprenorphine, and (R)-(�)-2-(4-bromo-2-fluorobenzyl)-1,2,3,4-tetrahydropyrrolo[1,2-�]pyrazine-4-spiro-3�-pyrrolidine-1,2�,3,5�-tetrone (AS-3201) inUGT phenotyping (Toide et al., 2004; Rouguieg et al., 2010).However, no validation was conducted in these studies. In thecurrent study, the RAF approach was used to find the isoformshaving a significant contribution to hepatic and intestinal glucu-ronidation of magnolol, and contributions of isoforms were alsotested and verified by selective or partial-selective chemical inhib-
itors. It is hoped that the results presented here will encourage theuse of the RAF approach in UGT phenotyping.
Materials and Methods
Notes. This study was performed, in unequal parts, in two different labo-ratories. Most of the planning and the experimental work was done at theDalian Institute of Chemical Physics of the Chinese Academy of Sciences inDalian, China (DICP laboratory), and additional experiments, mainly fortesting and validating the significant role of UGT1A10 in magnolol glucuroni-dation in the intestine, were performed at the Centre for Drug Research,University of Helsinki, Finland (Helsinki laboratory). Because of difficulties intransportation, the two laboratories used a different set of recombinant humanUGTs and different batches of liver and intestine microsomes. In addition, theHPLC instruments and analytical methods in two laboratories were alsodifferent. In the following, we have indicated in which materials and methodswere used in each laboratory.
Materials: DICP Laboratory. Magnolol (purity �98%) was purchasedfrom the Victory Company (Chengdu, China). UDPGA (trisodium salt), al-amethicin, diclofenac sodium, niflumic acid, phenylbutazone, D-saccharic acid1,4-lactone-�-D-glucuronidase, dopamine, propofol, and 3�-azido-3�-deoxy-thymidine (AZT) were purchased from Sigma-Aldrich (St. Louis, MO). Ethi-nylestradiol and 4-methylumbelliferyl-�-D-glucuronide (�98%) were pur-chased from Alfa Aesar (Beijing, China). Propofol glucuronide and AZTglucuronide were obtained from Toronto Research Chemicals Inc. (NorthYork, ON, Canada). Pooled human liver microsomes (HLM) from 25 donors(lot LSL) and pooled human intestine microsomes (HIM) from 10 donors (lotUGU) were purchased from the Research Institute for Liver Diseases (Shang-hai, China). Pooled Sprague-Dawley rat liver microsomes (n � 10) wereproduced as described previously (Li et al., 2008). A panel of recombinanthuman UGT isoforms (UGT1A1, -1A3, -1A4, -1A6, -1A7, -1A8, -1A9, -1A10,-2B4, -2B7, -2B15, and -2B17) expressed in baculovirus-infected insect cellswere purchased from BD Gentest (Woburn, MA). Catalytic activities of the 12UGTs were measured again in the laboratory before the experiment, and theactivities were equal to those provided by the supplier. All other reagents wereof HPLC grade or of the highest grade commercially available.
Materials: Helsinki Laboratory. AZT, alamethicin, dopamine, and UDPGA(triammonium salt) were purchased from Sigma-Aldrich. Magnolol was ob-tained from the DICP laboratory. HIM, a pool of 6 donors (lot 15031), werefrom BD Biosciences (San Jose, CA). Recombinant UGT1A10 and UGT2B7were produced in our laboratory, as described previously (Kuuranne et al.,2003; Kurkela et al., 2007). To avoid confusion due to the different sources ofrecombinant UGTs, those from the Helsinki laboratory will be indicated in thisarticle by an asterisk (*), namely UGT1A10* or -1A10* and UGT2B7* or-2B7*. This marking (*) is also used to indicate the tissue microsomes thatwere used in Helsinki (HIM*). All the solvents used in the Helsinki laboratorywere of HPLC grade.
Methods and Assays: DCIP Laboratory. Magnolol glucuronidation as-says. Magnolol was incubated with HLM, HIM, or recombinant UGT isoformsin a reaction mixture containing 50 mM Tris-HCl buffer (pH 7.4), 5 mMMgCl2, and 10 mM D-saccharic acid 1,4-lactone in a final volume of 200 �l.When HLM or HIM were assayed, the microsomes were fully activated by theaddition of alamethicin (5% protein concentrations). Before the glucuronida-tion reactions were started, all the samples were incubated at 37°C for 5 min,and then the reaction was initiated by the addition of UDPGA to a finalconcentration of 4 mM. The incubations were performed for different times, asspecified below, and they were terminated by the addition of 100 �l ofmethanol and then transferred to an ice bath and kept there for 20 min. Thesamples were then centrifuged at 20,000g for 10 min to remove protein.Control incubations were performed either without microsomes, withoutUDPGA, or without magnolol. The assays were analyzed by both ultra-fastliquid chromatography (UFLC)-diode array detector (DAD) and UFLC-MS.
To ensure the formation of glucuronide, the hydrolysis of product in theincubation mixture with �-glucuronidase was also performed. After incubationfor glucuronidation for 0.5 h in 200 �l of reaction mixture (without D-saccharicacid 1,4-lactone), 200 �l of 0.15 M acetate buffer (pH 5.0) with or without�-glucuronidase (1800 Fishman units) was added to the reaction mixture, andthen the incubation was performed at 37°C for another 0.5 h. The reaction was
FIG. 1. Glucuronidation pathway of magnolol.
530 ZHU ET AL.
at ASPE
T Journals on June 30, 2018
dmd.aspetjournals.org
Dow
nloaded from

quenched by the addition of 200 �l of methanol and centrifuged at 20,000g for10 min; the supernatant was analyzed by UFLC-DAD.
Analysis of magnolol glucuronidation samples. Magnolol and its glucuro-nide were analyzed by a UFLC spectrometry system (Shimadzu, Kyoto,Japan), equipped with two LC-20AD pumps, a DGU-20A3 vacuum degasser,a SIL-20ACHT autosampler, a CTO-20AC column oven, an SPD-M 20ADAD, a CBM-20A communications bus module, a mass detector (2010EV)with an electrospray ionization (ESI) interface, and a computer equipped withUFLC-MS solution software (version 3.41; Shimadzu). A Shim-pack XR-ODS(75.0 � 2.0 mm i.d., 2.2 �m; Shimadzu) analytical column with an ODS guardcolumn (5 � 2.0 mm i.d., 2.2 �m; Shimadzu) was used to separate magnololand its glucuronide. Column temperature was kept at 40°C. The mobile phasewas acetonitrile (A) and 0.2% formic acid (B) at a flow rate of 0.3 ml/min, withthe following gradient: 0 to 9 min, 90% B to 30% B; 9 to 12.5 min, 5% B; and12.5 to 16 min, balance to 90% B. The glucuronidation sample was stable over72 h at 4°C. Magnolol glucuronidation was quantified by the standard curve ofthe glucuronide at the detector wavelength of 290 nm, which was linear from0.1 to 20 �M (correlation coefficient was �0.999). The quantitative methoddisplayed good sensitivity, with the limit of detection of magnolol glucuronideless than 0.5 ng. The method also displayed good reproducibility, with theintraday and interday variances both less than 3%.
Mass detection was performed on a Shimadzu LCMS-2010EV instrumentwith an ESI interface both in positive and negative ion mode (ESI�) from m/z100 to 800. The detector voltage was set at �1.55 kV and �1.55 kV forpositive and negative ion detection, respectively. The curved desolvation linetemperature and the block heater temperature were both set at 250°C, whereasthe curved desolvation line voltage was set at 40 V. Other MS detectionconditions were as follows: interface voltage, �4.5 kV and �4.0 kV forpositive and negative ion detection, respectively; nebulizing gas (N2) flow, 1.5l/min; and drying gas (N2) pressure, 0.06 MPa. Data processing was performedusing the LC-MS solution software (version 3.41; Shimadzu).
Probe glucuronidation assays and analysis. Ethinylestradiol (100 �M) wasincubated with UGT1A1, HIM, or HLM for 30 min, with the final proteinconcentrations of 0.25 mg/ml. Ethinylestradiol glucuronidation samples wereanalyzed on the UFLC system (see above). A Shim-pack XR-ODS (50.0mm � 2.0 mm i.d., 2.2 �m; Shimadzu) analytical column with an ODS guardcolumn (5 mm � 2.0 mm i.d., 2.2 �m; Shimadzu) was used and kept at 40°C.The mobile phase consisted of acetonitrile (A) and 0.2% formic acid (B) at aflow rate of 0.3 ml/min, with the following gradient: 0 to 9 min, 90% B to 30%B; 9 to 12.5 min, 5% B; and 12.5 to 16 min, balance to 90% B. Theethinylestradiol glucuronide was detected by UV absorbance at 250 nm.
Propofol (200 �M) was incubated with UGT1A9 or HLM for 20 min, withthe final protein concentration of 0.5 mg/ml. Propofol glucuronide was ana-lyzed by LC-MS as described previously (Liang et al., 2011).
AZT (5 mM) was incubated with either UGT2B7 (commercial isoform),HLM, or HIM for 20 min, with the final protein concentrations of 0.5 mg/ml.AZT glucuronidation samples were analyzed on an HPLC system (Shimadzu),equipped with a SCL-10A system controller, two LC-10AT pumps, a SIL-10Aautosampler, and a SPD-10AVP UV detector. A C-18 column (4.6 � 250 mm,5 mm, Kromasil) was used to separate AZT and its glucuronide. The mobilephase was acetonitrile (A) and 0.2% formic acid (B) at a flow rate of 1.0ml/min, with an isocratic gradient: 0 to 25 min 90% B. The AZT glucuronidewas detected by UV absorbance at 260 nm.
Dopamine (10 mM) was incubated with either UGT1A10 (commercialisoform) or HIM for 40 min, with the final protein concentrations of 0.5 mg/ml.Glucuronidation samples were analyzed on a Shimadzu LCMS-2010EV in-strument with an ESI interface (see above). A self-made polar-copolymerizedstationary phase named C18HCE (150 � 4.6 mm, 5 mm) was used. Milliporewater containing 0.2% formic acid was used as the mobile phase. The ESIprobe voltage was set at �1.5 kV for detection. Mass spectrometry data werecollected at m/z 281 in the select ion monitoring mode with negative ESI todetect the dopamine glucuronide.
Biosynthesis of metabolites and NMR analysis. The metabolites were bio-synthesized using mixed liver microsomes from rat and human (90% rat livermicrosomes and 10% HLM) and purified for structure elucidation and quan-titative analysis. In brief, 200 �M magnolol was incubated with mixed livermicrosomes (0.5 mg of protein/ml), 50 mM Tris-HCl (pH 7.4), 5 mM MgCl2,Brij 58 (0.1 mg/mg protein), and 2 mM UDPGA in 200 ml of reaction mixtures
for 4 h at 37°C. The reaction was terminated by adding 100 ml of methanol,and the vessel was then put in an ice bath for 20 min. Proteins were removedby centrifugation at 20,000g for 30 min at 4°C, and the combined supernatantswere loaded on a SPE cartridge (C18 and anion exchange resin, 1000 mg;Dalian Sipore, Dalian, China). The SPE cartridge was preconditioned bysequential washing with 6 ml of methanol and 6 ml of Millipore water. Aftersample loading, the SPE cartridge was sequentially eluted with 6 ml ofMillipore water, 12 ml of methanol, and 12 ml of methanol containing 5%formic acid. The entire process was monitored by UFLC, and the metaboliteswere assembled in methanol containing 5% formic acid. After vacuum evap-oration, 3.2 mg of metabolites were obtained, and the purity was greater than95% by UFLC-DAD analysis.
The structure of the metabolite was determined by NMR spectra including1H NMR and 13C NMR. All experiments were performed on an INOVA-400NMR spectrometer (Varian, Inc., Palo Alto, CA). The purified metaboliteswere stored at �20°C before dissolving in dimethyl sulfoxide-d6 (Euriso-Top,Saint-Aubin, France) for NMR analysis. Chemical shifts were given on the �scale and referenced to tetramethylsilane at 0 ppm for 1H NMR (400 MHz) and13C NMR (100 MHz).
UGT isoform screening. The commercial recombinant UGT isoforms 1A1,1A3, 1A4, 1A6, 1A7, 1A8, 1A9, 1A10, 2B4, 2B7, 2B15, and 2B17 wereassayed for magnolol glucuronidation activity, in the presence of 12, 128, or256 �M substrate. The protein concentrations were 0.1 mg of protein/ml, andthe incubations were performed at 37°C for 60 min.
Kinetic analysis. To obtain the kinetic parameters in HLM and HIM,magnolol (0–100 �M) was incubated with pooled HLM (0.01 mg/ml) andHIM (0.04 mg/ml) at 37°C for 20 min. Preliminary experiments were per-formed to be sure that the formation of the glucuronides was in the linear rangeof both time (0–60 min) and protein concentrations (0.01–0.1 mg/ml). Kineticanalysis was also performed for UGT1A1, -1A3, -1A7, -1A9, and -2B7, whichexhibited significant magnolol glucuronidation activity. Magnolol (0–250�M) was incubated with commercial UGT1A1 (0.1 mg of protein/ml),UGT1A3 (0.1 mg of protein/ml), UGT1A7 (0.025 mg of protein/ml), UGT1A9(0.04 mg of protein/ml), and UGT2B7 (0.0125 mg of protein/ml) for 20 min,respectively. The kinetic models that were used to analyze the results wereeither substrate inhibition (eq. 1) or biphasic kinetics (eq. 2):
v �Vmax�S�
Ks � �S� � �S�2/Ksi(1)
where v is the rate of the reaction, [S] is the substrate concentration, Vmax is themaximum velocity estimate, Ks is the substrate affinity constant, and Ksi is thesubstrate inhibition constant.
v �Vmax1�S�
Ks1 � �S��
Vmax2�S�
Ks2 � �S�(2)
where Vmax1 and Vmax2 represent the estimated maximum velocity for the twometabolic phases and Ks1 and Ks2 represent the affinity constants for the twometabolic phases.
All incubations were performed in three independent experiments in dupli-cate. Kinetic constants were obtained using Origin 7.5 (OriginLab Corp.,Northampton, MA) and are reported as value S.E. of the parameter estimate.
Chemical inhibition studies. To better understand the roles of UGT enzymesin hepatic and intestinal magnolol glucuronidation, chemical inhibition studieswith diclofenac, phenylbutazone, and niflumic acid were performed.
Chemical inhibition for recombinant UGT enzymes. In the DICP laboratory,magnolol (6 �M) was incubated with individual isoforms of UGT1A1, -1A3,-1A7, -1A9, and -2B7 in the presence or absence of the inhibitors: diclofenac(10 and 100 �M), phenylbutazone (100 and 500 �M), and niflumic acid (10and 100 �M). Commercial UGT1A8 and UGT1A10 were not included herebecause of very low magnolol glucuronidation activity.
Chemical inhibition for HLM. To test the inhibitory effects of the inhibitorson magnolol glucuronidation in HLM, incubations of magnolol (6 �M) withHLM in the presence or absence of diclofenac (10 and 100 �M), phenylbu-tazone (100 and 500 �M), and niflumic acid (10 and 100 �M) were performed,respectively.
Given the possible dependence of contributions of UGTs on substrateconcentration, inhibition of diclofenac (0–200 �M) of magnolol glucuronida-
531MAGNOLOL GLUCURONIDATION IN HLM AND HIM
at ASPE
T Journals on June 30, 2018
dmd.aspetjournals.org
Dow
nloaded from

tion by HLM and UGT2B7 was investigated at three different magnololconcentrations (6, 12, and 24 �M) to get a clearer insight into the roles ofUGT2B7 in liver. The IC50 (inhibitor concentration that inhibits 50% ofcontrol activity) values for inhibition of diclofenac on magnolol glucuronida-tion by HLM and UGT2B7 were determined by nonlinear curve fitting withOrigin 7.5 as described previously and then were compared with each other(Zhang et al., 2009).
Chemical inhibition for HIM. To test the inhibitory effects of these inhib-itors (above) on magnolol glucuronidation in HIM, incubations of magnolol (6�M) with HIM in the presence or absence of diclofenac (10 and 100 �M),phenylbutazone (100 and 500 �M), and niflumic acid (10 and 100 �M) wereperformed, respectively.
RAF approach application. To achieve an accurate understanding of theroles of UGT enzymes in hepatic and intestinal glucuronidation, magnololglucuronidation activity was scaled from recombinant systems to HLM andHIM through the RAF approach. The RAF value was calculated by dividingthe velocity of the probe reaction in HLM or HIM by that in the recombinantenzyme (eq. 3) (Crespi and Miller, 1999):
RAFi �Vprobe reaction in HLM or HIM
Vprobe reaction in recombinant UGT(3)
The magnolol glucuronidation rate of the individual isoforms in HLM orHIM (Vi) is estimated by multiplying the reaction rate in the recombinantenzyme (vi) by the corresponding RAF value (RAFi) (eq. 4):
Vi � vi RAFi (4)
The relative contribution of the individual isoform (Contributioni) is calcu-lated by dividing Vi by the magnolol glucuronidation rate in HLM or HIM(Vtotal) (eq. 5):
Contributioni �Vi
Vtotal 100% (5)
Calculations of RAF values. Glucuronidation rates of ethinylestradiol,propofol, and AZT were reported to be probe reactions for catalytic activity ofhuman UGT1A1, UGT1A9, and UGT2B7, respectively, in human liver (Courtet al., 2003; Soars et al., 2003). In addition, ethinylestradiol, dopamine, andAZT can be used as probe substrates for UGT1A1, UGT1A10, and UGT2B7,respectively, in human small intestine (Court et al., 2003; Soars et al., 2003;Itaaho et al., 2009). The glucuronidation rates of propofol and AZT werequantified by using a standard curve made with authentic compounds. Becauseno ethinylestradiol and dopamine glucuronide was commercially available forthe DCIP laboratory, areas under the glucuronide peak were used to calculatethe RAF values in these cases.
Contributions of individual isoforms. Given possible dependence of contri-butions of UGT enzymes on concentrations of the parent compound, magnololglucuronidation rates at different substrate concentrations (6, 12, and 24 �M)by HLM, HIM, or recombinant UGT isoforms were measured simultaneouslywith probe reactions. After multiplying magnolol glucuronidation rates inrecombinant UGT (vi) by the corresponding RAF values (eq. 4), rates of theindividual UGT isoforms in HLM or HIM (Vi), respectively, were obtained.After Vi was divided by magnolol glucuronidation rates in HLM or HIM (Vtotal)(eq. 5), relative contributions of individual isoforms (Contributioni) to hepaticor intestinal glucuronidation, respectively, were obtained.
Methods and Assays: Helsinki Laboratory. Magnolol glucuronidationassays. In the magnolol glucuronidation assays in the Helsinki laboratory, theprotein concentrations were either 0.02 mg/ml UGT1A10* or HIM* or 0.05mg/ml UGT2B7*. The 100-�l reaction mixtures contained 50 mM sodiumphosphate buffer (pH 7.4), 10 mM MgCl2, alamethicin (5% of the proteinconcentration) when HIM* was assayed, and 5 mM UDPGA. All the reactionmixtures for the HIM* reactions (before UDPGA addition but in the presenceof alamethicin) were preincubated for 30 min on ice. Before reaction initiationby the addition of UDPGA, all the reaction mixtures were incubated at 37°Cfor 5 min, followed by UDPGA addition and subsequent incubation at 37°C for30 min. The assays were terminated by the addition of 60 �l of methanol-4 Mperchloric acid (5:1) mix and cooling on ice for 10 min, followed by centrif-
ugation at 13,000 rpm for 10 min in a bench top centrifuge. The resultingsupernatants were analyzed to detect the glucuronides.
Magnolol glucuronidation samples were analyzed by both UV absorbance(290 nm) and fluorescence (excitation was at 225 nm and emission at 353 nm)using a Shimadzu LC-10 model HPLC system equipped with a ChromolithSpeedROD RP-18e (50 � 4.6 mm) column at 40°C. The mobile phase wasacetonitrile (A) and 0.1% formic acid (B) at a flow rate of 1 ml/min, with thefollowing gradient: 1 to 8 min, 70% B to 5% B; 8 to 8.5 min, 5% B; 8.5 to 8.6min, 5% B to 70% B; and 8.6 to 10 min, 70% B. Because of the lack ofauthentic magnolol glucuronide standard in the Helsinki laboratory, the stan-dard curve for quantification of the magnolol glucuronide in the differentsamples was based on magnolol UV absorbance at 290 nm. The DCIPlaboratory found that the excitation of the glucuronide at 290 nm is 1.2 timeshigher than the corresponding value for magnolol, and the rates were adjustedaccordingly.
Magnolol glucuronidation kinetics was performed for UGT1A10* in the 2to 200 �M substrate concentration range. To get the kinetic constants, sub-strate inhibition (eq. 1) and the Michaelis-Menten model (eq. 6) were used:
v �Vmax �S�
Km � �S�(6)
where v is the rate of the reaction, [S] is the substrate concentration, Vmax is themaximum velocity estimate, and Km is the apparent affinity constant.
The chemical inhibition experiments in the Helsinki laboratory were per-formed for UGT1A10*. The concentrations of chemical inhibitors and mag-nolol were the same as those in the DICP laboratory, whereas other incubationconditions were the same as those described above for the magnolol glucu-ronidation assays in the Helsinki laboratory (above).
Probe glucuronidation assays. The AZT glucuronidation rates of UGT2B7*and HIM* and the dopamine glucuronidation rates of UGT1A10* and HIM*were measured. The incubation mixture contained 0.5 mg of protein/ml en-zyme. The AZT concentration in the respective analyses was 5 mM, and thedopamine concentration in the respective experiments was 10 mM. AZTglucuronidation samples were analyzed by ultraperformance liquid chroma-tography-MS. The system consisted of a Waters Acquity ultraperformanceliquid chromatograph equipped with an Acquity BEH C18 column (1.7 �m,2.1 � 100 mm) and an Acquity PDA detector and a Waters Xevo Q-Tof massspectrometer. Mass spectrometry was performed in negative ion mode fromm/z 100 to 1000, as described previously (Zhang et al., 2011). Dopamineglucuronidation was analyzed using a Waters Acquity ultraperformance liquidchromatograph and an Agilent 6410 triple-quadrupole mass spectrometer, asdescribed previously (Uutela et al., 2009).
Results
Identification of Magnolol Glucuronidation. Incubation of mag-nolol with either HLM or HIM in the presence of UDPGA yielded asingle glucuronide peak. This glucuronide was absent in controlsamples without microsomes, UDPGA, or magnolol. Mass spectrom-etry in the negative ion mode showed an m/z value of 441.1 for thedeprotonated metabolite, a value that corresponds well to magnolol(266 � 1) with m/z 176 of the glucuronosyl substitution (Fig. 2).Treatment with �-glucuronidase further indicated that the metabolitewas a �-glucuronide (data not shown).
The magnolol glucuronide was biosynthesized and purified forstructure elucidation, as well as for use as a standard in differentanalyses. The glucuronide structure determination was performed byNMR, using both 1H NMR and 13C NMR analyses (Table 1). Incomparison with magnolol, the 13C NMR spectrum of the glucuroniderevealed an upfield shift at C-2 to � 152.7 (� �0.4), which isprobably due to a glycosidation shift of a phenolic compound. In thesugar moiety, the G1 protons and carbons exhibited characteristicchemical shifts near 5 and 100 ppm, whereas the G6 carbon(�COOH) showed chemical shifts of approximately 170 ppm. Takentogether, the evidence clearly demonstrated that the location of the
532 ZHU ET AL.
at ASPE
T Journals on June 30, 2018
dmd.aspetjournals.org
Dow
nloaded from

glucuronic acid substitution was at the C-2 phenolic group of mag-nolol (Fig. 1).
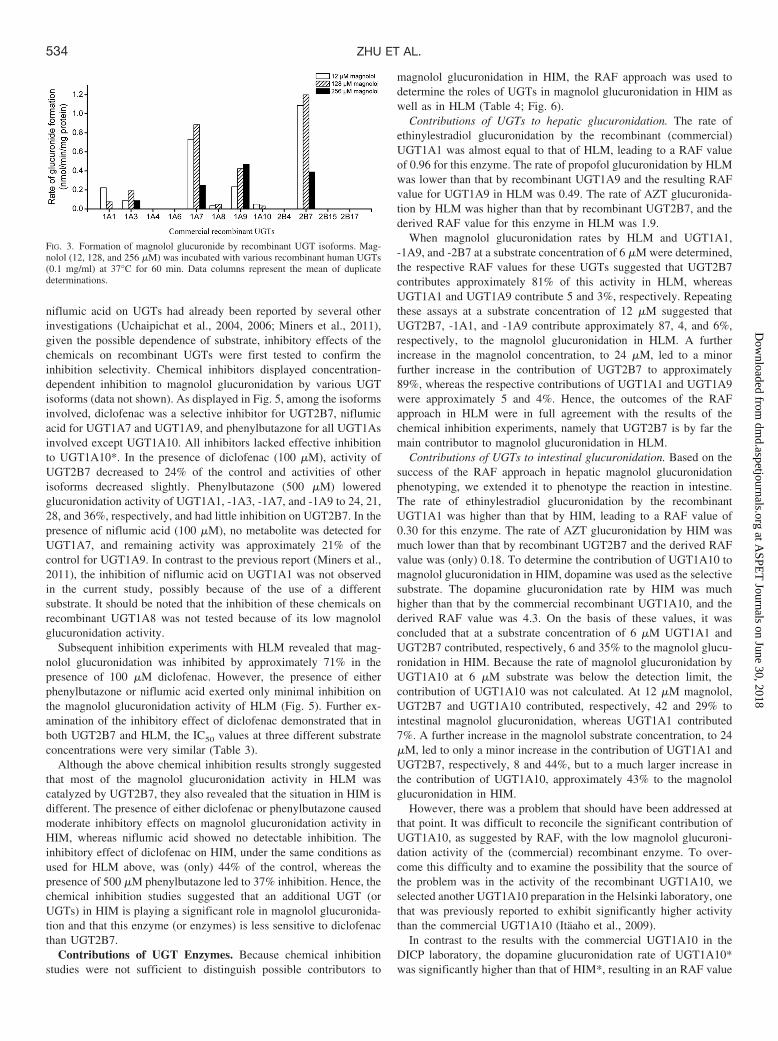
UGT Isoform(s) Screening. A panel of 12 commercial recombi-nant human UGT isoforms was screened for their magnolol glucu-ronidation activity. The results demonstrated that UGT1A1, -1A3,-1A7, -1A8, -1A9, -1A10, and -2B7 could catalyze this reaction,whereas UGT1A4, -1A6, -2B4, -2B15, and -2B17 did not exhibit anyactivity toward this compound. Of these, UGT2B7 showed the highestmagnolol glucuronidation activity, whereas UGT1A8 and UGT1A10exhibited very low magnolol glucuronidation activity (Fig. 3). Thescreening assays were performed using three different substrate con-centrations (12, 128 and 256 �M) to detect all UGTs with magnololglucuronidation activity, without missing the isoform whose activityis highly sensitive to substrate inhibition. The screening results (Fig.3) indeed gave strong indications for substrate inhibition in many ofthe UGTs, a property that was then studied in more detail.
Kinetic Analyses. Kinetic analyses were performed in HLM, HIM,and commercial recombinant UGT1A1, -1A3, -1A7, -1A9, and -2B7.
The results were best simulated by the substrate inhibition kineticmodel for all the tested enzymes and microsomes, with the exceptionof UGT1A9 (Fig. 4). In the case of recombinant UGT1A9, biphasickinetics was observed rather than substrate inhibition (Fig. 4). In thecase of UGT1A10, self-expressed enzyme (termed UGT1A10*) ratherthan commercial enzyme was used to perform the kinetic analysis.Results indicated that substrate inhibition and the Michaelis-Mentenmodel could both describe it well (Table 2). For recombinantUGT1A8, the magnolol glucuronidation activity of this enzyme wastoo low to get reliable kinetic constants (data not shown). Kineticparameters are listed in Table 2.
Chemical Inhibition Studies. Further information on the contri-bution of individual UGTs to magnolol glucuronidation in HLM orHIM may be obtained from UGT inhibition experiments with iso-form-specific inhibitors. Therefore, we tried to inhibit magnololglucuronidation by the main UGTs, using different inhibitors.
Chemical Inhibition for the Commercial Recombinant UGTEnzymes. Although inhibition of diclofenac, phenylbutazone, and
FIG. 2. Representative LC profiles of magnololand its glucuronide. The glucuronide and mag-nolol were eluted at 5.6 and 7.7 min, respec-tively. The m/z of the glucuronide in negativeion mode is displayed as the inset. mAU, mil-liabsorbance units.
TABLE 1
Proton and carbon NMR chemical shift assignments for magnolol and its metabolite
PositionMagnolol Metabolite
�1H �13C �1H �13C
1, 1� 128.4, 125.62, 2� 152.7, 152.75, 5� 131.5, 132.43, 3� 6.79–6.93 (6H, m) 116.1 6.81–7.09 (6H, m) 114.9, 116.44, 4� 128.4 128.4, 128.56, 6� 130.1 130.5, 132.47, 7� 3.26–3.34 (4H, m) 39.8 3.26–3.33 (4H, m) 39.5, 39.78, 8� 5.90–5.94 (2H, m) 138.8 5.89–5.98 (2H, m) 138.4, 138.89, 9� 4.99–5.08 (4H, m) 115.7 4.98–5.06 (4H, m) 115.7, 116.0G1 5.10 (1H, d, J � 8.0) 100.6G2 3.28 (1H, m) 70.2G3 3.30 (1H, m) 75.9G4 3.30 (1H, m) 71.7G5 4.01 (1H, d, J � 8.0) 75.9G6 170.6
533MAGNOLOL GLUCURONIDATION IN HLM AND HIM
at ASPE
T Journals on June 30, 2018
dmd.aspetjournals.org
Dow
nloaded from
niflumic acid on UGTs had already been reported by several otherinvestigations (Uchaipichat et al., 2004, 2006; Miners et al., 2011),given the possible dependence of substrate, inhibitory effects of thechemicals on recombinant UGTs were first tested to confirm theinhibition selectivity. Chemical inhibitors displayed concentration-dependent inhibition to magnolol glucuronidation by various UGTisoforms (data not shown). As displayed in Fig. 5, among the isoformsinvolved, diclofenac was a selective inhibitor for UGT2B7, niflumicacid for UGT1A7 and UGT1A9, and phenylbutazone for all UGT1Asinvolved except UGT1A10. All inhibitors lacked effective inhibitionto UGT1A10*. In the presence of diclofenac (100 �M), activity ofUGT2B7 decreased to 24% of the control and activities of otherisoforms decreased slightly. Phenylbutazone (500 �M) loweredglucuronidation activity of UGT1A1, -1A3, -1A7, and -1A9 to 24, 21,28, and 36%, respectively, and had little inhibition on UGT2B7. In thepresence of niflumic acid (100 �M), no metabolite was detected forUGT1A7, and remaining activity was approximately 21% of thecontrol for UGT1A9. In contrast to the previous report (Miners et al.,2011), the inhibition of niflumic acid on UGT1A1 was not observedin the current study, possibly because of the use of a differentsubstrate. It should be noted that the inhibition of these chemicals onrecombinant UGT1A8 was not tested because of its low magnololglucuronidation activity.
Subsequent inhibition experiments with HLM revealed that mag-nolol glucuronidation was inhibited by approximately 71% in thepresence of 100 �M diclofenac. However, the presence of eitherphenylbutazone or niflumic acid exerted only minimal inhibition onthe magnolol glucuronidation activity of HLM (Fig. 5). Further ex-amination of the inhibitory effect of diclofenac demonstrated that inboth UGT2B7 and HLM, the IC50 values at three different substrateconcentrations were very similar (Table 3).
Although the above chemical inhibition results strongly suggestedthat most of the magnolol glucuronidation activity in HLM wascatalyzed by UGT2B7, they also revealed that the situation in HIM isdifferent. The presence of either diclofenac or phenylbutazone causedmoderate inhibitory effects on magnolol glucuronidation activity inHIM, whereas niflumic acid showed no detectable inhibition. Theinhibitory effect of diclofenac on HIM, under the same conditions asused for HLM above, was (only) 44% of the control, whereas thepresence of 500 �M phenylbutazone led to 37% inhibition. Hence, thechemical inhibition studies suggested that an additional UGT (orUGTs) in HIM is playing a significant role in magnolol glucuronida-tion and that this enzyme (or enzymes) is less sensitive to diclofenacthan UGT2B7.
Contributions of UGT Enzymes. Because chemical inhibitionstudies were not sufficient to distinguish possible contributors to
magnolol glucuronidation in HIM, the RAF approach was used todetermine the roles of UGTs in magnolol glucuronidation in HIM aswell as in HLM (Table 4; Fig. 6).
Contributions of UGTs to hepatic glucuronidation. The rate ofethinylestradiol glucuronidation by the recombinant (commercial)UGT1A1 was almost equal to that of HLM, leading to a RAF valueof 0.96 for this enzyme. The rate of propofol glucuronidation by HLMwas lower than that by recombinant UGT1A9 and the resulting RAFvalue for UGT1A9 in HLM was 0.49. The rate of AZT glucuronida-tion by HLM was higher than that by recombinant UGT2B7, and thederived RAF value for this enzyme in HLM was 1.9.
When magnolol glucuronidation rates by HLM and UGT1A1,-1A9, and -2B7 at a substrate concentration of 6 �M were determined,the respective RAF values for these UGTs suggested that UGT2B7contributes approximately 81% of this activity in HLM, whereasUGT1A1 and UGT1A9 contribute 5 and 3%, respectively. Repeatingthese assays at a substrate concentration of 12 �M suggested thatUGT2B7, -1A1, and -1A9 contribute approximately 87, 4, and 6%,respectively, to the magnolol glucuronidation in HLM. A furtherincrease in the magnolol concentration, to 24 �M, led to a minorfurther increase in the contribution of UGT2B7 to approximately89%, whereas the respective contributions of UGT1A1 and UGT1A9were approximately 5 and 4%. Hence, the outcomes of the RAFapproach in HLM were in full agreement with the results of thechemical inhibition experiments, namely that UGT2B7 is by far themain contributor to magnolol glucuronidation in HLM.
Contributions of UGTs to intestinal glucuronidation. Based on thesuccess of the RAF approach in hepatic magnolol glucuronidationphenotyping, we extended it to phenotype the reaction in intestine.The rate of ethinylestradiol glucuronidation by the recombinantUGT1A1 was higher than that by HIM, leading to a RAF value of0.30 for this enzyme. The rate of AZT glucuronidation by HIM wasmuch lower than that by recombinant UGT2B7 and the derived RAFvalue was (only) 0.18. To determine the contribution of UGT1A10 tomagnolol glucuronidation in HIM, dopamine was used as the selectivesubstrate. The dopamine glucuronidation rate by HIM was muchhigher than that by the commercial recombinant UGT1A10, and thederived RAF value was 4.3. On the basis of these values, it wasconcluded that at a substrate concentration of 6 �M UGT1A1 andUGT2B7 contributed, respectively, 6 and 35% to the magnolol glucu-ronidation in HIM. Because the rate of magnolol glucuronidation byUGT1A10 at 6 �M substrate was below the detection limit, thecontribution of UGT1A10 was not calculated. At 12 �M magnolol,UGT2B7 and UGT1A10 contributed, respectively, 42 and 29% tointestinal magnolol glucuronidation, whereas UGT1A1 contributed7%. A further increase in the magnolol substrate concentration, to 24�M, led to only a minor increase in the contribution of UGT1A1 andUGT2B7, respectively, 8 and 44%, but to a much larger increase inthe contribution of UGT1A10, approximately 43% to the magnololglucuronidation in HIM.
However, there was a problem that should have been addressed atthat point. It was difficult to reconcile the significant contribution ofUGT1A10, as suggested by RAF, with the low magnolol glucuroni-dation activity of the (commercial) recombinant enzyme. To over-come this difficulty and to examine the possibility that the source ofthe problem was in the activity of the recombinant UGT1A10, weselected another UGT1A10 preparation in the Helsinki laboratory, onethat was previously reported to exhibit significantly higher activitythan the commercial UGT1A10 (Itaaho et al., 2009).
In contrast to the results with the commercial UGT1A10 in theDICP laboratory, the dopamine glucuronidation rate of UGT1A10*was significantly higher than that of HIM*, resulting in an RAF value
FIG. 3. Formation of magnolol glucuronide by recombinant UGT isoforms. Mag-nolol (12, 128, and 256 �M) was incubated with various recombinant human UGTs(0.1 mg/ml) at 37°C for 60 min. Data columns represent the mean of duplicatedeterminations.
534 ZHU ET AL.
at ASPE
T Journals on June 30, 2018
dmd.aspetjournals.org
Dow
nloaded from

of 0.28. Also different from the results in the DICP laboratory, in theHelsinki laboratory, the rate of AZT glucuronidation in HIM* washigher than that in UGT2B7*, and the RAF value for this enzyme in
HIM was found to be 1.3. On the basis of these RAF values, theHelsinki laboratory results indicated that at 5 �M magnolol,UGT1A10 contributed approximately 33% to magnolol glucuronida-
TABLE 2
Kinetic parameters for magnolol glucuronidation by HLM, HIM, and recombinant UGT isoforms
Data represent means S.E. of computer calculation.
Enzyme Sources Vmax Ks Ksi Goodness of Fit (R2)
nmol � min�1 � mg protein�1 �M �M
UGT1A1a 0.27 0.02 6.74 0.78 16.5 1.9 0.992UGT1A3a 2.14 0.19 138 14 11.5 1.2 0.999UGT1A7a 2.06 0.30 9.60 2.39 36.6 8.5 0.972UGT2B7a 7.70 1.89 19.4 6.0 6.45 1.94 0.991UGT1A9b 0.19 0.01 0.28 0.08 0.999
0.33 0.01 33.4 4.1UGT1A10*a 4.02 0.19 10.8 1.2 1314 589 0.995UGT1A10*c 3.62 0.08 8.55 0.85 0.989HLMa 34.4 14.8 27.6 14.4 6.78 3.47 0.979HIMa 2.50 0.43 4.70 1.44 19.7 5.6 0.957
a Data were fit into the substrate inhibition model (eq. 1).b Data were fit into the biphasic kinetic model (eq. 2).c Data was fit into the Michaelis-Menten model (eq. 6).
FIG. 4. Kinetics of magnolol glucuronidation by HLM, HIM, and the commercial recombinant UGT1A1, -1A3, -1A7, -1A9, and -2B7 (DICP laboratory) and UGT1A10*(Helsinki laboratory). Eadie-Hofstee plots (V/S � V) are displayed as insets. Data points represent the mean of triplicate independent determinations, and error bars representthe calculated S.D.
535MAGNOLOL GLUCURONIDATION IN HLM AND HIM
at ASPE
T Journals on June 30, 2018
dmd.aspetjournals.org
Dow
nloaded from

tion in HIM, whereas UGT2B7 contributed 27%. At a 20 �M mag-nolol concentration, the contributions of UGT1A10 and UGT2B7were 26 and 37%, respectively.
Hence, regardless of the use of different enzyme preparations andmicrosomes samples, the results in both the DICP and Helsinkilaboratories showed that UGT1A10, alongside UGT2B7, is a signif-icant contributor to magnolol glucuronidation in HIM (Fig. 6).
Discussion
This study is focused on the characterization of the glucuronidationof magnolol in human liver and intestine. Both HLM and HIM caneffectively catalyze magnolol glucuronidation, forming the 2-O-monoglucuronide. Magnolol glucuronidation exhibits two distinctfeatures. First, multiple UGT isoforms, including several hepaticUGTs (UGT1A1, -1A3, -1A9, and -2B7) and a few extrahepaticenzymes (UGT1A7, -1A8, and -1A10), are all involved in magnololglucuronidation. Second, nearly all UGTs, with the possible exceptionof UGT1A10, exhibit atypical kinetics toward magnolol glucuronida-tion (Table 2). Among them, UGT1A9 is an interesting exception thatexhibits biphasic kinetics, a rare interaction model with the substrate.
Because multiple UGTs are involved in magnolol glucuronidation,and each enzyme shows atypical kinetics, it is quite challenging tocorrectly determine the relative contributions of individual isoforms inliver and intestine. Because only a limited number of UGT selectiveinhibitors are currently available, magnolol glucuronidation in therecombinant UGT is essential to estimate the relative contributions ofindividual isoforms. However, the rate of magnolol glucuronidationby a recombinant UGT rarely reflects the true glucuronidation rate ofthe individual UGTs in HLM and HIM. This finding is fully exem-plified in this study. The rate of a marker reaction in the recombinantenzyme differed a lot from that in tissue microsomes, making the RAF
value rarely approximately 1. Thus, the glucuronidation rate by therecombinant enzyme often gives misleading information of the con-tribution of this isoform in native tissues, such as magnolol glucu-ronidation by commercial UGT1A10. The RAF approach offered anew possibility to closely estimate the roles of individual UGTs inhuman liver and intestine.
The RAF approach is seldom used in UGT phenotyping. A possiblereason for this may be that determination of a RAF value requiresenough specific substrates for the different UGTs, with selectivity ofat least 80% (Crespi and Miller, 1999). Nevertheless, whereas thereare still only a few fully specific substrates for individual UGTs, it iseasier to find suitable substrates when the expression pattern of theUGTs in the tissue tested is taken into account. In the current study,magnolol glucuronidation phenotyping in HLM and HIM was sepa-rately conducted, under full consideration of expression of UGTs inliver and intestine. The following substrates were used: ethinylestra-diol for UGT1A1 in HLM and HIM (Soars et al., 2003), propofol forUGT1A9 in HLM (Soars et al., 2004), dopamine for UGT1A10 inHIM (Itaaho et al., 2009), and AZT for UGT2B7 in HLM and HIM(Court et al., 2003).
According to the tissue-specific UGT expression pattern (Ohno andNakajin, 2009), UGT1A7, -1A8, and -1A10 can be excluded fromhepatic magnolol glucuronidation. Among the other UGTs, the RAFapproach indicated that UGT2B7 was almost solely responsible forthe hepatic magnolol glucuronidation, with only limited contributionfrom UGT1A1, -1A3, and -1A9. The contribution of UGT2B7 to theliver reaction was in full agreement with the chemical inhibitionresults. Diclofenac showed potent inhibition to both HLM and recom-binant UGT2B7 and even similar inhibition IC50 values (Table 3). Inparticular, at a 24 �M substrate concentration, the reaction rate beganto decrease in assays with recombinant UGT2B7. It was assumed thatthe contribution of UGT2B7 would decrease; however, diclofenacalso gave inhibition values similar to those of UGT2B7 and HLM,strongly suggesting that UGT2B7 plays the predominant role in theliver reaction through the whole concentration range. In addition, verylittle inhibition was exerted by the non-UGT2B7 inhibitors on HLM(phenylbutazone and niflumic acid). The consistencies of these ex-periments fully validated use of the RAF approach, leading to itsfurther application in intestinal reaction phenotyping.
Compared with the hepatic magnolol glucuronidation, the intestinalreaction was more complicated. According to the tissue-specificUGTs expression pattern (Ohno and Nakajin, 2009), it can be as-sumed that all magnolol glucuronidation UGTs contribute to intestinalmagnolol glucuronidation. Both diclofenac and phenylbutazoneshowed moderate inhibition to magnolol glucuronidation in HIM,indicating that the reaction in HIM was mediated by multiple isoformswith comparable contributions. Of note, niflumic acid, a potent inhib-itor of UGT1A7 and UGT1A9, exhibited no inhibition to intestinalmagnolol glucuronidation. Thus, UGT1A7 and UGT1A9 contributedlittle to the intestinal reaction, despite the recombinant enzymes,particularly UGT1A7, exhibiting considerable magnolol glucuronida-
TABLE 3
IC50 values for inhibitory effects of diclofenac on magnolol glucuronidation byHLM and UGT2B7
Data are means S.E. of computer calculation.
MagnololIC50
HLM UGT2B7
�M
6 �M 23.2 3.0 24.6 2.812 �M 38.6 6.2 31.1 4.724 �M 46.3 5.2 39.6 7.1
TABLE 4
RAF values for major isoforms
IsoformsRAF Values
HLM (DICP) HIM (DICP) HIM (Helsinki)a
UGT1A1 0.96 0.30UGT1A9 0.49UGT1A10 4.3 0.28UGT2B7 1.9 0.18 1.3
a In the Helsinki laboratory, the recombinant enzymes are UGT1A10* and UGT2B7*,whereas in the DICP laboratory the recombinant enzymes are commercial isoforms.
FIG. 5. Inhibitory effects of phenylbutazone (500 �M), diclofenac (100 �M), andniflumic acid (100 �M) toward glucuronidation of magnolol (6 �M) by HLM, HIM,and recombinant UGT1A1, -1A3, -1A7, -1A9, -1A10*, and -2B7. Data columnsrepresent the means of duplicate determinations.
536 ZHU ET AL.
at ASPE
T Journals on June 30, 2018
dmd.aspetjournals.org
Dow
nloaded from

tion activities. This result may be evidence of low expression ofUGT1A7 and UGT1A9 in human intestine (Strassburg et al., 1998,2000).
Chemical inhibition failed to give further detailed information onthe contribution of individual UGTs to intestinal magnolol glucuroni-dation. The RAF approach indicated that both UGT2B7 andUGT1A10 contributed significantly, whereas UGT1A1 contributedinsignificantly (less than 10%). Of interest, the significant contribu-tion of UGT1A10 is contrary to the assays with commercial recom-binant enzyme that exhibited the low magnolol glucuronidation ac-tivity. To further investigate whether UGT1A10 acts as a significantcontributor, a more active enzyme (UGT1A10*) was used to performmagnolol glucuronidation assays. Assays with UGT1A10* are inagreement with the RAF results for the commercial recombinantenzyme.
It should be noted here that the RAF values are dramaticallydifferent between the two laboratories because different sources ofUGT1A10, UGT2B7, and HIM were used. Take UGT1A10, forexample. Because the commercial recombinant UGT1A10 showedmuch lower activity than the self-constructed one (UGT1A10*), theRAF value for UGT1A10 was much higher when the commercialenzyme was used. The differences in RAF values may be caused bythe different levels of UGT1A10 expression and the percentage ofactive UGT1A10 as well as other factors in the enzyme sources used.These differences in sample activity between laboratories did notundermine the usefulness of the RAF methodology. On the contrary,it may even make this approach more important. The results of this
study demonstrated how successful the methodology was for deci-phering the contributions of individual UGTs from assays with onepoor recombinant enzyme without being aware of the activity prob-lem. Although recombinant (commercial) UGT1A10 showed pooractivity, the RAF approach did not underestimate the true role of thisenzyme in intestinal magnolol glucuronidation.
This study provided a very good example of the powerful value ofapplication of the RAF approach in detecting individual UGTs thatcontribute to specific reactions in human tissues. However, one itemof note is that the respective roles of UGT1A3 and UGT1A8 inintestine glucuronidation are still not clear in the current study be-cause of the lack of a selective inhibitor and substrate. On the otherhand, according to this study, the total contributions of UGT1A1,-1A10, and -2B7 to intestinal magnolol glucuronidation are more than80%. Thus, the roles of UGT1A3 and UGT1A8 are insignificant in theintestinal reaction. One more item of note is that because not all UGTsare currently commercial available, several other UGTs (includingUGT1A5, -2B10, -2B11, -2A1, and -2A2) can also potentially cata-lyze the magnolol glucuronidation. However, they contribute little tothe reaction in human liver and intestine, either based on this study oron their low expression level in these two tissues (Ohno and Nakajin,2009; Sneitz et al., 2009).
This study not only presented the power of RAF in glucuronidationphenotyping but also issued an alarm for the application of magnolol.From this study, it can be concluded that UGT1A10 and UGT2B7play important roles in magnolol glucuronidation. Magnolol glucu-ronidation occurs at a relatively low rate in those with impaired
FIG. 6. Relative contributions of UGT isoforms to HLM (A, DICP laboratory) and HIM (B, DICP laboratory; C, Helsinki laboratory). because of the low activity of thecommercial recombinant UGT1A10, the contribution of this isoform to intestinal magnolol glucuronidation (6 �M) is not separately displayed and is included in thecontribution of others (B).
537MAGNOLOL GLUCURONIDATION IN HLM AND HIM
at ASPE
T Journals on June 30, 2018
dmd.aspetjournals.org
Dow
nloaded from

glucuronidation activities of these two enzymes. Previous studiesindicated that glucuronidation activity of UGT2B7 was low in chil-dren (Strassburg et al., 2002; Zaya et al., 2006). Of note, children maytake a high dose of magnolol (1.64 mg/kg per day). It can be antici-pated that magnolol may reach a relatively high concentration inplasma. Based on this study, magnolol showed high affinity forseveral UGTs with low Ks or Ksi values, in particular for UGT1A9,with one Ks value less than 1 �M (Table 2), suggesting that magnololmay possibly act as an potent UGT inhibitor. The risk of magnololinducing undesired drug-drug interactions through inhibition of UGTenzymes should be evaluated, particularly in children with an over-dose of this compound.
In summary, multiple UGT isoforms are involved in magnololglucuronidation, and most of them exhibit atypical kinetics. The RAFapproach is proved to be a powerful tool in glucuronidation pheno-typing on this condition, compensating for a lack of selective UGTinhibitors. The results clearly indicate UGT2B7 is a predominantisoform in magnolol glucuronidation in human liver, whereasUGT1A10 and UGT2B7 are significant contributors in intestine.
Acknowledgments
We thank Dr. Xiaochi Ma from Dalian Medical University for help in NMRanalysis and Joann Mosorin of the Helsinki laboratory for recombinant UGTpreparations.
Authorship Contributions
Participated in research design: Zhu, Ge, H. Zhang, Liu, Finel, and Yang.Conducted experiments: Zhu, H. Zhang, Liang, and Fang.Contributed new reagents or analytic tools: Zhu and Ge.Performed data analysis: Zhu, H. Zhang, Y. Zhang, and Dong.Wrote or contributed to the writing of the manuscript: Zhu, Ge, H. Zhang,
He, Finel, and Yang.
References
Court MH, Krishnaswamy S, Hao Q, Duan SX, Patten CJ, Von Moltke LL, and Greenblatt DJ(2003) Evaluation of 3�-azido-3�-deoxythymidine, morphine, and codeine as probe substratesfor UDP-glucuronosyltransferase 2B7 (UGT2B7) in human liver microsomes: specificity andinfluence of the UGT2B7*2 polymorphism. Drug Metab Dispos 31:1125–1133.
Crespi CL and Miller VP (1999) The use of heterologously expressed drug metabolizingenzymes—state of the art and prospects for the future. Pharmacol Ther 84:121–131.
Fakhrudin N, Ladurner A, Atanasov AG, Heiss EH, Baumgartner L, Markt P, Schuster D,Ellmerer EP, Wolber G, Rollinger JM, et al. (2010) Computer-aided discovery, validation, andmechanistic characterization of novel neolignan activators of peroxisome proliferator-activated receptor �. Mol Pharmacol 77:559–566.
Greenberg M, Urnezis P, and Tian M (2007) Compressed mints and chewing gum containingmagnolia bark extract are effective against bacteria responsible for oral malodor. J Agric FoodChem 55:9465–9469.
Hattori M, Endo Y, Takebe S, Kobashi K, Fukasaku N, and Namba T (1986) Metabolism ofmagnolol from Magnoliae cortex. II. Absorption, metabolism and excretion of [ring-14C]magnolol in rats. Chem Pharm Bull 34:158–167.
Homma M, Oka K, Kobayashi H, Niitsuma T, Yamamoto S, Itoh H, and Takahashi N (1993)Liquid chromatographic determination of magnolol in urine collected from volunteers after asingle dose of saiboku-to, an oriental herbal medicine for bronchial asthma. J Pharm Phar-macol 45:839–841.
Itaaho K, Court MH, Uutela P, Kostiainen R, Radominska-Pandya A, and Finel M (2009)Dopamine is a low-affinity and high-specificity substrate for the human UDP-glucuronosyltransferase 1A10. Drug Metab Dispos 37:768–775.
Kurkela M, Patana AS, Mackenzie PI, Court MH, Tate CG, Hirvonen J, Goldman A, and FinelM (2007) Interactions with other human UDP-glucuronosyltransferases attenuate the conse-quences of the Y485D mutation on the activity and substrate affinity of UGT1A6. Pharma-cogenet Genomics 17:115–126.
Kuuranne T, Kurkela M, Thevis M, Schanzer W, Finel M, and Kostiainen R (2003) Glucuroni-dation of anabolic androgenic steroids by recombinant human UDP-glucuronosyltransferases.Drug Metab Dispos 31:1117–1124.
Lee YJ, Lee YM, Lee CK, Jung JK, Han SB, and Hong JT (2011) Therapeutic applications ofcompounds in the Magnolia family. Pharmacol Ther 130:157–176.
Li W, Liu Y, He YQ, Zhang JW, Gao Y, Ge GB, Liu HX, Huo H, Liu HT, Wang LM, et al.(2008) Characterization of triptolide hydroxylation by cytochrome P450 in human and rat livermicrosomes. Xenobiotica 38:1551–1565.
Liang SC, Ge GB, Liu HX, Shang HT, Wei H, Fang ZZ, Zhu LL, Mao YX, and Yang L (2011)Determination of propofol UDP-glucuronosyltransferase (UGT) activities in hepatic micro-somes from different species by UFLC-ESI-MS. J Pharm Biomed Anal 54:236–241.
Mackenzie PI, Bock KW, Burchell B, Guillemette C, Ikushiro S, Iyanagi T, Miners JO, OwensIS, and Nebert DW (2005) Nomenclature update for the mammalian UDP glycosyltransferase(UGT) gene superfamily. Pharmacogenet Genomics 15:677–685.
Miners JO, Bowalgaha K, Elliot DJ, Baranczewski P, and Knights KM (2011) Characterizationof niflumic acid as a selective inhibitor of human liver microsomal UDP-glucuronosyltrans-ferase 1A9: application to the reaction phenotyping of acetaminophen glucuronidation. DrugMetab Dispos 39:644–652.
Miners JO, Mackenzie PI, and Knights KM (2010) The prediction of drug-glucuronidationparameters in humans: UDP-glucuronosyltransferase enzyme-selective substrate and inhibitorprobes for reaction phenotyping and in vitro-in vivo extrapolation of drug clearance anddrug-drug interaction potential. Drug Metab Rev 42:196–208.
Ohno S and Nakajin S (2009) Determination of mRNA expression of human UDP-glucuronosyltransferases and application for localization in various human tissues by real-timereverse transcriptase-polymerase chain reaction. Drug Metab Dispos 37:32–40.
Rouguieg K, Picard N, Sauvage FL, Gaulier JM, and Marquet P (2010) Contribution of thedifferent UDP-glucuronosyltransferase (UGT) isoforms to buprenorphine and norbuprenor-phine metabolism and relationship with the main UGT polymorphisms in a bank of humanliver microsomes. Drug Metab Dispos 38:40–45.
Sneitz N, Court MH, Zhang XL, Laajanen K, Yee KK, Dalton P, Ding XX, and Finel M (2009)Human UDP-glucuronosyltransferase UGT2A2: cDNA construction, expression, and func-tional characterization in comparison with UGT2A1 and UGT2A3. Pharmacogenet Genomics19:923–934.
Soars MG, Petullo DM, Eckstein JA, Kasper SC, and Wrighton SA (2004) An assessment ofUDP-glucuronosyltransferase induction using primary human hepatocytes. Drug Metab Dis-pos 32:140–148.
Soars MG, Ring BJ, and Wrighton SA (2003) The effect of incubation conditions on the enzymekinetics of UDP-glucuronosyltransferases. Drug Metab Dispos 31:762–767.
Strassburg CP, Kneip S, Topp J, Obermayer-Straub P, Barut A, Tukey RH, and Manns MP(2000) Polymorphic gene regulation and interindividual variation of UDP-glucuronosyltrans-ferase activity in human small intestine. J Biol Chem 275:36164–36171.
Strassburg CP, Manns MP, and Tukey RH (1998) Expression of the UDP-glucuronosyltrans-ferase 1A locus in human colon. Identification and characterization of the novel extrahepaticUGT1A8. J Biol Chem 273:8719–8726.
Strassburg CP, Strassburg A, Kneip S, Barut A, Tukey RH, Rodeck B, and Manns MP (2002)Developmental aspects of human hepatic drug glucuronidation in young children and adults.Gut 50:259–265.
Toide K, Terauchi Y, Fujii T, Yamazaki H, and Kamataki T (2004) Uridine diphosphatesugar-selective conjugation of an aldose reductase inhibitor (AS-3201) by UDP-glucuronosyltransferase 2B subfamily in human liver microsomes. Biochem Pharmacol 67:1269–1278.
Tsai TH, Chou CJ, and Chen CF (1995) Glucuronidation of magnolol assessed using HPLC/fluorescence. Planta Medica 61:491–492.
Uchaipichat V, Mackenzie PI, Elliot DJ, and Miners JO (2006) Selectivity of substrate (triflu-operazine) and inhibitor (amitriptyline, androsterone, canrenoic acid, hecogenin, phenylbuta-zone, quinidine, quinine, and sulfinpyrazone) “probes” for human UDP-glucuronosyltrans-ferases. Drug Metab Dispos 34:449–456.
Uchaipichat V, Mackenzie PI, Guo XH, Gardner-Stephen D, Galetin A, Houston JB, and MinersJO (2004) Human UDP-glucuronosyltransferases: isoform selectivity and kinetics of 4-meth-ylumbelliferone and 1-naphthol glucuronidation, effects of organic solvents, and inhibition bydiclofenac and probenecid. Drug Metab Dispos 32:413–423.
Uutela P, Karhu L, Piepponen P, Kaenmaki M, Ketola RA, and Kostiainen R (2009) Discoveryof dopamine glucuronide in rat and mouse brain microdialysis samples using liquid chroma-tography tandem mass spectrometry. Anal Chem 81:427–434.
Wells PG, Mackenzie PI, Chowdhury JR, Guillemette C, Gregory PA, Ishii Y, Hansen AJ,Kessler FK, Kim PM, Chowdhury NR, et al. (2004) Glucuronidation and the UDP-glucuronosyltransferases in health and disease. Drug Metab Dispos 32:281–290.
Zaya MJ, Hines RN, and Stevens JC (2006) Epirubicin glucuronidation and UGT2B7 develop-mental expression. Drug Metab Dispos 34:2097–2101.
Zhang H, Tolonen A, Rousu T, Hirvonen J, and Finel M (2011) Effects of cell differentiation andassay conditions on the UDP-glucuronosyltransferase activity in Caco-2 cells. Drug MetabDispos 39:456–464.
Zhang YY, Liu Y, Zhang JW, Ge GB, Wang LM, Sun J, and Yang L (2009) Characterization ofhuman cytochrome P450 isoforms involved in the metabolism of 7-epi-paclitaxel. Xenobiotica39:283–292.
Address correspondence to: Dr. Ling Yang, Laboratory of PharmaceuticalResource Discovery, Dalian Institute of Chemical Physics, Chinese Academy ofSciences, 457 Zhongshan Rd., Dalian, 116023 China. E-mail: [email protected]
538 ZHU ET AL.
at ASPE
T Journals on June 30, 2018
dmd.aspetjournals.org
Dow
nloaded from


















