
2011-01-05 Dissertation Final Version Dissertation Fi… · 2011-01-05 Dissertation Final Version
Upload
alina-ivanCategory
view
62download
1
RUNNING HEAD: SPECIFIC INTERFERENCE WITHIN THE RECONSOLIDATION WINDOW
Are declarative memories sensitive to specific post-retrieval interference?
620036721, University of Exeter

2 SPECIFIC INTERFERENCE WITHIN THE RECONSOLIDATION WINDOW
Acknowledgements
With thanks to my supervisor, Dr. Nicolas Dumay, for his encouragement and support during the
completion of this project.

3 SPECIFIC INTERFERENCE WITHIN THE RECONSOLIDATION WINDOW
Abstract
According to the reconsolidation hypothesis, the retrieval of a consolidated memory may
briefly return it to a plastic state, making it vulnerable to retroactive interference. However,
Dumay and Tremblett (in prep.) found no reduction in memory strength for declarative
memories that were reactivated before 12 hours of unspecific interference from daily
encoding. Since the reconsolidation hypothesis cannot accommodate for such findings, we
sought to examine whether retrieved memories are altered by specific interference (i.e.,
novel, yet similar learning). We reasoned that finding reconsolidation effects a day after
reactivated memories have been exposed to specific interference would challenge the
assumption that consolidation and reconsolidation operate under similar boundary conditions.
Participants learned two lists of novel words (e.g., frenzylk) on day one. On day two, they
reactivated one list, then learned corruptors (e.g., frenzyl) for all words memorized on the
previous day. We tested reconsolidation-related effects on day three, expecting weaker
memories for the non-overlapping phoneme (e.g., ‘k’ in frenzylk) in reactivated compared to
non-reactivated words. Notwithstanding our predictions, cued recall data showed an overall
memory boost for reactivated compared to non-reactivated words, and no difference in the
percentage of omissions on the last letter. Similarly, phoneme monitoring data revealed
slightly stronger memories for reactivated compared to non-reactivated words. Since past
research provides no clear guide regarding experimental parameters that unequivocally
induce reconsolidation effects, our results allow several interpretations. We discuss our
findings in relation to available research on boundary conditions that constrain
reconsolidation.

4 SPECIFIC INTERFERENCE WITHIN THE RECONSOLIDATION WINDOW
Newly formed memory traces are vulnerable to interference from subsequent encoding
(Müller & Pilzecker, 1900). Yet, during a period of reduced interference, such as sleep, they
undergo a stabilisation process (Wixted, 2004). This process is referred to as consolidation, and
it has traditionally been thought to keep memories unchanged (McGaugh, 1966, 2000). However,
in the 1960s, Misanin, Miller, and Lewis (1968) posited that reactivation renders memories labile
again, contradicting the canonic consolidation view. Consistent with this notion, a seminal study
by Nader, Schafe, and LeDoux (2000) showed that a protein synthesis inhibitor can block a
consolidated memory upon reactivation. There is now relatively wide consensus about the
reconsolidation hypothesis, which states that reactivation renders memories in a state of
temporary plasticity that is followed by another period of stabilisation (Dudai & Eisenberg,
2004). Despite sharing many features with consolidation, the reconsolidation process is not
considered to be merely another cycle of consolidation (Dudai, 2012; Lee, Everitt, & Thomas,
2004; Tronson & Taylor, 2007). While consolidation strengthens a newly formed memory to
make it available for later retrieval, reconsolidation is thought to facilitate memory updating
(e.g., Lee, 2010; Merlo, Milton, & Everitt, 2015; Tronel & Alberini, 2007; Alberini, 2011; Hardt,
Einarsson, & Nader, 2010).
Neuroscientific research has provided accumulating evidence in favour of the
reconsolidation hypothesis. Amnesia-inducing drugs have been shown to block consolidated
memories upon retrieval in various species (Alberini & LeDoux, 2013). Using fear conditioning
paradigms, several studies confirmed that reconsolidation also occurs in humans. Soeter and
Kindt (2010) used propranolol to manipulate the reconsolidation of learned fear responses. A day
after learning a fear association, the experimental group received propranolol before memory
reactivation, while a control group received propranolol without reactivating the target memory.

5 SPECIFIC INTERFERENCE WITHIN THE RECONSOLIDATION WINDOW
A session of extinction on day three showed reduced fear expression in the experimental group,
but not in the control group. Importantly, similar results were obtained one month after the
original learning took place, suggesting that the reconsolidation manipulation blocked the return
of fear. The possibility to reduce the strength of fear memories has generated substantial interest
in reconsolidation. This interest partly arouse from its potential applications to reduce the
physiological reactions in disorders caused by maladaptive fear memories (Agren, 2014).
Notably, reconsolidation-based treatments have already been reported to induce attenuation of
trauma symptoms in PTSD patients (Brunet et al., 2008; Brunet et al., 2011; Poundja, Sanche,
Tremblay, & Brunet, 2012).
Further evidence for reconsolidation comes from the observation that new encoding
following reactivation can impair the original memory (Walker, Brakefield, Hobson, &
Stickgold, 2003; Boccia, Blake, Acosta, & Baratti, 2005). Walker and colleagues (2003) were
among the first to study reconsolidation in humans using a behavioural interference paradigm.
On day one, subjects were trained on a finger-tapping sequence. The following day, the
reactivation group rehearsed the sequence and immediately learned a second sequence, while the
non-reactivation (i.e., control) group learned the second sequence without rehearsing the initial
sequence. A test on the third day revealed impaired recall of the initial sequence in the
reactivation group, but intact knowledge in the non-reactivation group. These findings suggest
that interference from learning similar information upon memory reactivation can lead to
impaired recall. Similar results were obtained in a series of studies looking at the reconsolidation
of procedural memories (e.g., Censor, Dimyan, & Cohen, 2010, 2014).
Interference paradigms offer a means to study memory reconsolidation non-invasively.
They are particularly useful for targeting declarative and procedural memories, since no safe

6 SPECIFIC INTERFERENCE WITHIN THE RECONSOLIDATION WINDOW
pharmacological blocker is currently known to interfere with their reconsolidation in humans
(Schiller & Phelps, 2011). These paradigms have been traditionally used in consolidation
research (Müller & Pilzecker, 1900; Robertson, 2012), where unspecific interference has been
documented to interrupt the consolidation process in a time-dependent manner (Wixted, 2004).
The current view in the consolidation literature is that newly encoded memories and to-be-
consolidated memories compete for plasticity-related mechanisms, which prevents initial
memories from being consolidated (Mednick et al., 2011). Surprisingly, and in contrast with
consolidation research, a study in our laboratories demonstrated that unspecific interference does
not affect declarative memories rendered plastic by reactivation (Dumay & Tremblett, in prep.).
The day following encoding of nonsense words, half of the subjects reactivated one list in the
morning, subjecting it to 12 hours of unspecific interference from daily mental activity. The rest
of participants reactivated the same list before sleep, exposing the memory trace to minimal
interference. On day three, explicit and implicit memory measures revealed that the expected
reduction in memory strength for reactivated memories did not take place. This suggests that
reactivated memories are robust to unspecific interference. Two concerns arise.
One concern is that reconsolidation might not occur in declarative memories. However,
there are several reports of reconsolidation in declarative memories using behavioural
interference paradigms (e.g., Chan & LaPaglia, 2013; Jacques & Schacter, 2013; Forcato et al.,
2007; Hupbach, Hardt, Gomez, & Nadel, 2008). Indeed, some studies are susceptible to
alternative explanations due to the nature of the paradigms employed. Using a typical three-day
reconsolidation paradigm, Forcato and colleagues (2007) had subjects reactivate overnight-
consolidated lists through a pair associate (e.g., OEN-SRO) coupled with context cues (e.g., light
colour). On the third day, they found that learning a second list up to six hours after reactivation

7 SPECIFIC INTERFERENCE WITHIN THE RECONSOLIDATION WINDOW
lead to less retrieval-induced forgetting (i.e., a phenomenon observed only when a retrieved
memory is intact). However, Deliens and colleagues (2013) found that learning a pair-associate
that is similar to an already-learnt pair-associate activated the latter and interfered with its
encoding. It may be that results from the Forcato and associates (2007) study might have arisen
from interference at encoding, rather than reconsolidation. Furthermore, an alternative model
(Sederberg, Gershman, Polyn, & Norman, 2011) was proposed to explain reports of
reconsolidation in episodic memories by Hupbach and colleagues (2008, see Agren, 2014).
Despite the existence of some questionable reconsolidation demonstrations, a recent study has
provided evidence for reconsolidation in declarative memories using electroconvulsive therapy,
which produces seizure activity (Kroes et al., 2014). These researchers found that
electroconvulsive therapy applied after memory reactivation blocked the reconsolidation of
emotional declarative emotional memories in reactivated, but not in non-reactivated memories in
patients with depression. This study provides support for the notion that emotional declarative
memories can, in fact, be altered during the reconsolidation window.
Rather than concluding that declarative memories do not undergo reconsolidation based
on negative findings, another question that arises from the work of Dumay and Tremblett (in
prep.) would be whether a memory that is destabilised by reactivation is sensitive to particular
types of retroactive interference. This is the focus of the current investigation. Most studies that
observed reconsolidation effects using interference paradigms employed an interfering agent that
overlapped to a certain degree with the reactivated memory. In the Walker and associates (2003)
study, for instance, the new motor-tapping sequence (e.g., 2-3-1-4-2) was performed using the
same fingers as the original sequence (e.g., 4-1-3-2-4). In another study that reported
reconsolidation effects, participants were trained participants on a list of nonsense syllables that

8 SPECIFIC INTERFERENCE WITHIN THE RECONSOLIDATION WINDOW
had the same length as the corrupting learning (e.g., ITE-OBN vs. OEN-SRO), both being in
rioplantese Spanish (Forcato, Argibay, Pedreira, & Maldonado, 2009). Furthermore, research
using conditioning paradigms on rats has used post-reactivation learning shares features with the
original ‘conditioned stimulus – unconditioned response’ contingency (e.g., Boccia et al., 2005;
Boccia, Blake, Krawczyk, & Baratti, 2010).
A number of researchers advocate for the need to have a certain degree of similarity
between target memory and corrupting information to detect reconsolidation (e.g., Besnard,
Caboche, & Laroche, 2012; Hupbach, 2011). Besnard and colleagues (2012) proposed a model
to explain how similarity between the previously memorised experiences and potential
interference at memory recall may engage competing consolidation and reconsolidation
mechanisms. He argued that a high level of similarity results in updating the previous memory,
while a low level of similarity reduces the likelihood of engaging in reconsolidation by triggering
cellular mechanisms responsible for consolidation. According to this model, if consolidation is
initiated, then the assumed interfering information is stored in an independent memory trace,
separate from the original memory. This theory could explain findings that learning new
information upon retrieval engages consolidation in some instances (Tronel, Milekic, & Alberini,
2005) and reconsolidation in others (Lee, 2010).
The notion that reconsolidation has ‘boundary conditions’ has been proposed to name
parameters that constrain the observation of the reconsolidation process. Due to these constraints,
reconsolidation research has obtained a mixed bag of results. For example, old memories were
found not to be sensitive to reconsolidation manipulations (Suzuki et al., 2004; Milekic &
Alberini, 2002), neither were strong memories (e.g., Lee, Ciano, Thomas, & Everitt, 2005;
Debiec, LeDoux, & Nader, 2002), memories that do not encounter a mismatch between original

9 SPECIFIC INTERFERENCE WITHIN THE RECONSOLIDATION WINDOW
memory and expected outcome at reactivation (e.g., Sevenster, Beckers, & Kindt, 2013; Morris
et al., 2006; Pedreira, Pérez-Cuesta, & Maldonado, 2004), and memories reactivated indirectly
(Debiec, Doyère, Nader, & LeDoux, 2006). However, there are opposing findings for some
reports of boundary conditions. Older and stronger memories reconsolidate under different sets
of experimental parameters, such as larger doses of amnesic agents or longer reactivation
sessions (Debiec & LeDoux, 2004; Bustos, Maldonado, & Molina, 2009). The literature on
boundary conditions that inhibit reconsolidation has inconsistencies and lacks comprehensive
accounts. It thus needing further attention.
Of special relevance for the current investigation is a recent study which sought to
compare the effects of specific and unspecific post-retrieval interference (Chan & LaPaglia,
2013; Experiments 4 & 5). Subjects learned about a false terrorist attack from a video. The
following day, half of the subjects reactivated the memory trace, while the rest performed a
distractor task. After five minutes, they found impaired memory in the misinformation condition,
but not in the distractor condition. The effect was not observed when the same parameters were
used to examine non-specific interference (i.e., misinformation items did not target the
reactivated memory, being related to a story about drug trafficking). The authors asserted that the
interference agent needs to be specific to the original memory to observe reconsolidation.
Although Chan and LaPaglia’s (2013) interpretation is of interest, their results do not
necessarily demonstrate that the original memory has been modified through reconsolidation.
Testing memory immediately after the relearning phase is at odds with the reconsolidation
hypothesis, which states that the results of the reactivation-corruption manipulation should be
visible only after reconsolidation has been completed (Nader et al., 2000). In numerous studies,
reconsolidation appears to take at least two to four hours (Stickgold & Walker, 2007). There is

10 SPECIFIC INTERFERENCE WITHIN THE RECONSOLIDATION WINDOW
evidence that behaviours remained unchanged immediately after exposure to interference
(Walker et al., 2003), and also two (Suzuki et al., 2004) or four hours (Duvarci & Nader, 2004;
Debiec & LeDoux, 2004) after blocking reconsolidation. Thus, memory performance measured
five minutes before the end of interference phase might not reflect the effects of a completed
reconsolidation process in the Chan and LaPaglia (2013) study. Rather, it might be that new
learning was established as a separate memory trace and competed for reactivation with the
original memory (cf. Deliens et al., 2013).
Current study
To our knowledge, reports of reconsolidation published to date employed specific
interference. Based on this observation and on the model proposed by Besnard and associates
(2012), we assess whether declarative memories are sensitive to specific interference upon
retrieval. Finding reconsolidation effects when interference is specific, rather that unspecific
(other factors held constant) would demonstrate that having similarity between target learning
and corrupting information is a boundary condition for reconsolidation. We therefore adopt the
same learning and reactivation procedures from Dumay & Tremblett (in prep.). We employed a
three-day paradigm that aims to alter previously consolidated lexical memories following their
reactivation (Fig. 1). The use of a three-day paradigm, instead of a two-day paradigm as in Chan
and LaPaglia (2013) avoids potential problems with estimating the end of the reconsolidation
phase.

11 SPECIFIC INTERFERENCE WITHIN THE RECONSOLIDATION WINDOW
On day one, subjects formed memories of novel words (e.g., frenzylk). Half of these
memories were reactivated on day two, before learning corruptors (i.e., novel , but similar
words) for all memories encoded the previous day (e.g., frenzyl). On day three, memory for
reactivated and non-reactivated words was assessed using three tasks. In the phoneme
monitoring task, subjects detected phonemes in spoken words. The task measures how existing
phonological knowledge about the word stimulus contributes to phoneme recognition. Stronger
memories facilitate speed and accuracy (cf. Frauenfelder, Segui, & Dijkstra, 1990). We predict
that detection of the non-overlapping phoneme (“k” in frenzilk) would be slower and more liable
to errors in the case of reactivated compared to non-reactivated words. We then tested explicit
recall with a cued recall task. Recalling word endings is expected to be poorer for reactivated
compared to non-reactivated words (Fig. 2). Moreover, we posit that there will be more errors on
the last phoneme for reactivated compared to non-reactivated words.

12 SPECIFIC INTERFERENCE WITHIN THE RECONSOLIDATION WINDOW
A pause detection (i.e., lexical interference) task was lastly employed. In this task,
participants detected short silences in target words. This was a necessary measure as it shows
whether the information presented in the assumed reconsolidation window replaces the original
memory, or it has been stored in a separate memory trace. The rationale is that upon hearing a
word, the same input elicits parallel processing of multiple phonologically-similar words until
one passes the uniqueness point (McClelland & Elman, 1986). If one has memory for both
frenzylk and frenzyl, the two memories will compete for activation when presented with the
spoken word frenzylk. The speed at which the pause is detected depends on the degree of the
lexical activity existent at the point where the pause is inserted (Mattys & Clark, 2002). Hence, if
one has separate memories for target words and corruptors (rather than one updated memory),
the activation of the phonological neighbour would produce interference, thus slowing down or
even impeding pause detection. Accordingly, our expectation is that there will be slower reaction
times and more errors for pauses embedded into non-reactivated compared to reactivated words
(Fig. 3).

13 SPECIFIC INTERFERENCE WITHIN THE RECONSOLIDATION WINDOW
If as predicted, the data would constitute the first unambiguous evidence that specific
interference facilitates reconsolidation. As a more general implication, these predicted results
would add to the accumulating body of evidence that reconsolidation is not merely another cycle
of consolidation (e.g., Dudai, 2012).
Method
Subjects
Thirty subjects aged 19-25 (M=21.37; SD=1.92) were tested (15 females, 15 males).
They were native UK-English speakers (i.e., had UK-English as their first learnt and most skilled
language) and had no known hearing, language, sleep, or memory deficits. They consented to
keep their usual sleeping and drinking patterns throughout the experiment, and not to exert
mental effort between sessions. Subjects were students at the University of Exeter, recruited

14 SPECIFIC INTERFERENCE WITHIN THE RECONSOLIDATION WINDOW
through convenience sampling. Sweets or course credits were given for their participation. This
study was approved by University of Exeter’s Ethics Committee (Appendix A).
Design
A within-subjects design was used to compare memory strength for reactivated versus
non-reactivated words. Forty-eight words were equally split into three separate lists (A, B, and
C). Each participant learned two of these lists (e.g., A and B) on day one. On day two, they
reactivated one list (e.g., A or B). Lastly, on day three, we tested memory for both lists and for
baseline words. The baseline was set to ensure that performance changed depending on learning
and memory strength, and not due to task proficiency or to other extraneous factors. All three
lists (i.e., A, B, and C) had each possible role (i.e., reactivated, non-reactivated, and baseline) in
turn among six groups of participants.
Baseline and target words were further divided into two lists for the pause detection task
(i.e., pause-present and pause-absent words). In the sound detection task each word appeared
twice, first time requiring a ‘yes’ response and the second time requiring a ‘no’ response.
No effect was expected on negative responses in phoneme monitoring, since they engage
checking mechanisms that conflict with the memory facilitation effect (cf. Frauenfelder &
Peeters, 1990). In terms of pause detection, we recorded responses on pause-absent trials
although they were not required in Mattys and Clark’s (2002) original pause detection task. The
rationale is that inhibition or failure to detect pause absence result from the allocation of general
resources to lexical competition (cf. Gaskell & Dumay, 2003). An advantage is that processing
can proceed uninterrupted, unlike in the case of pause-present trials where the pause may slightly

15 SPECIFIC INTERFERENCE WITHIN THE RECONSOLIDATION WINDOW
disrupt processing when resources are not taken up completely by competition (Gaskell &
Dumay, 2003).
Materials
Stimuli were adapted from Dumay and Gaskell (2012). There were 48 target novel
words, all derived from existing bi-syllabic English words by adding a two-consonant suffix to a
stem (e.g., “frenzy” + “lk”). Corruptors differed from target words by lacking the final consonant
(e.g., “frenzy” + “l”). They were recorded separately, being compatible with target words.
Additionally, compatible recordings of the stem words were used for cued recall and pause
detection.
All stimuli were presented over Beyer Dynamic DT 770 Pro/80 headphones. Subjects
used a Trust 1200 gamepad to start and respond to the phoneme monitoring task and to the pause
detection task. For cued recall, responses were recorded by a Sennheiser pc151 microphone. A
soundproof computer booth was allocated per subject. For practical reasons, the booth was
chosen randomly for each session. However, all computer booths were identical and excluded
subjects’ belongings.
Procedure
Procedure is schematically represented in Fig. 1. Stimuli were presented through DMDX,
which also collected latencies and voice recordings.
Day 1 (after 6 pm). After signing a consent form that detailed experimental procedure
and inclusion criteria (Appendix B), subjects performed a sound detection task (Fig. 4). A prompt
instructed them to decide quickly and accurately if spoken words presented over the headphones
contained the sound presented on the screen. It also required subjects to learn the words at their
best because they will be asked to perform a memory test. A 1000-ms break followed each

16 SPECIFIC INTERFERENCE WITHIN THE RECONSOLIDATION WINDOW
stimulus presentation. Six target sounds (/L/, /K/, /M/, /S/, /T/, /N/) were used for six set
repetitions (i.e., each set was repeated 36 times as in Dumay & Gaskell, 2012). Break lengths
between blocks lasted until participants decided to press a key. The task encouraged attention to
each trial and took approximately 40 minutes.
Day 2 (after 6 pm). Subjects retrieved half of the words by responding to the same sound
detection task. Each set was reactivated six times, once with each target sound. The retrieval
session took approximately 10 minutes. The same sound detection task ran subjects through
corruptors for each of the 32 target words for 36 times with the same instructions.
Day 3 (before 12 pm). Subjects performed a series of memory tasks. In the phoneme
monitoring task (Fig. 5), a phoneme appeared quickly on the screen (200 ms). Subjects were
required to respond promptly and accurately if the spoken word that followed contained this
phoneme or not. The phoneme appeared at a random position.

17 SPECIFIC INTERFERENCE WITHIN THE RECONSOLIDATION WINDOW
A cued recall task followed. Upon hearing stems (e.g., frenzy), subjects articulated what
they recalled from the respective word learnt on day one.
Finally, a pause detection task was performed. The task started with 167 practice trials.
Quick and accurate ‘yes’ or ‘no’ decisions were required depending whether the spoken word
contained a short silence. A 200-ms artificial pause was inserted after the stem (e.g., frenzy_lk).
Subjects had 3 s to respond from stimulus onset, and a 1-s break before the next trial. Response
latencies were measured starting from pause onset.
After completing the experiment, subjects were asked to report unusual sleeping times or
alcohol consumption over the duration of the experiment. Oral debriefing followed.
Results
Responses were missing for three subjects in phoneme monitoring, three in cued recall,
and one in pause detection. All analyses on proportional data (e.g., error rates) were conducted
after performing arcsine transformations to achieve normality.

18 SPECIFIC INTERFERENCE WITHIN THE RECONSOLIDATION WINDOW
Phoneme monitoring
Performance on phoneme monitoring was analysed through a repeated measures
ANOVA. We examined how response latencies and error rates vary as a function of word
condition (baseline, non-reactivated, or reactivated). Responses given earlier than 200 ms or
2000 ms after stimulus presentation were not recorded. The analysis included only trials that
contained the target phoneme.
Response latencies. Analysis of response latencies included only correct trials. The word
learning manipulation was successful (F2,56=16.340, p<.001), as revealed by the significantly
longer response latencies to baseline words compared to non-reactivated words and reactivated
words (see Fig. 6a for mean response latencies). The difference was significant both in the case
of non-reactivated words (F1,28=17.173, p<.001) and reactivated words (F1,28=27.890, p<.001).
However, latencies to reactivated words did not differ significantly from those to non-reactivated
words (F1,28=1.515, p=.227). Surprisingly, it appears that reactivation slightly boosted memory
strength (see Fig. 6a).
19 SPECIFIC INTERFERENCE WITHIN THE RECONSOLIDATION WINDOW
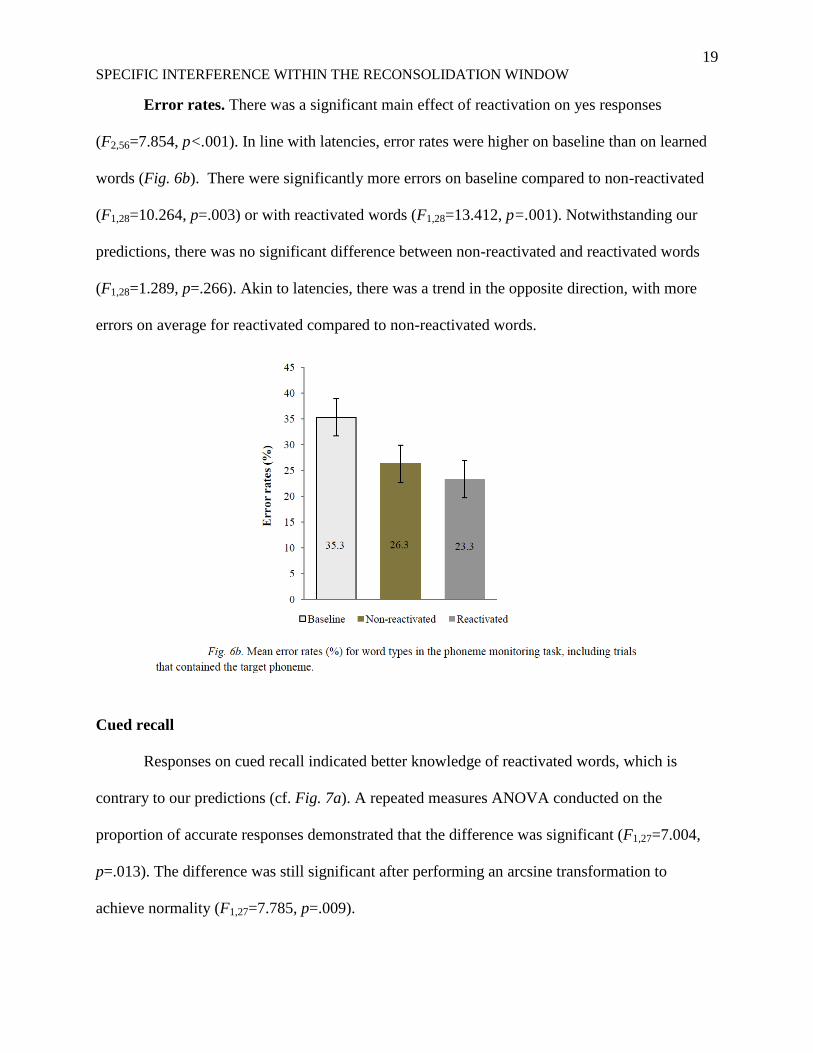
Error rates. There was a significant main effect of reactivation on yes responses
(F2,56=7.854, p<.001). In line with latencies, error rates were higher on baseline than on learned
words (Fig. 6b). There were significantly more errors on baseline compared to non-reactivated
(F1,28=10.264, p=.003) or with reactivated words (F1,28=13.412, p=.001). Notwithstanding our
predictions, there was no significant difference between non-reactivated and reactivated words
(F1,28=1.289, p=.266). Akin to latencies, there was a trend in the opposite direction, with more
errors on average for reactivated compared to non-reactivated words.
Cued recall
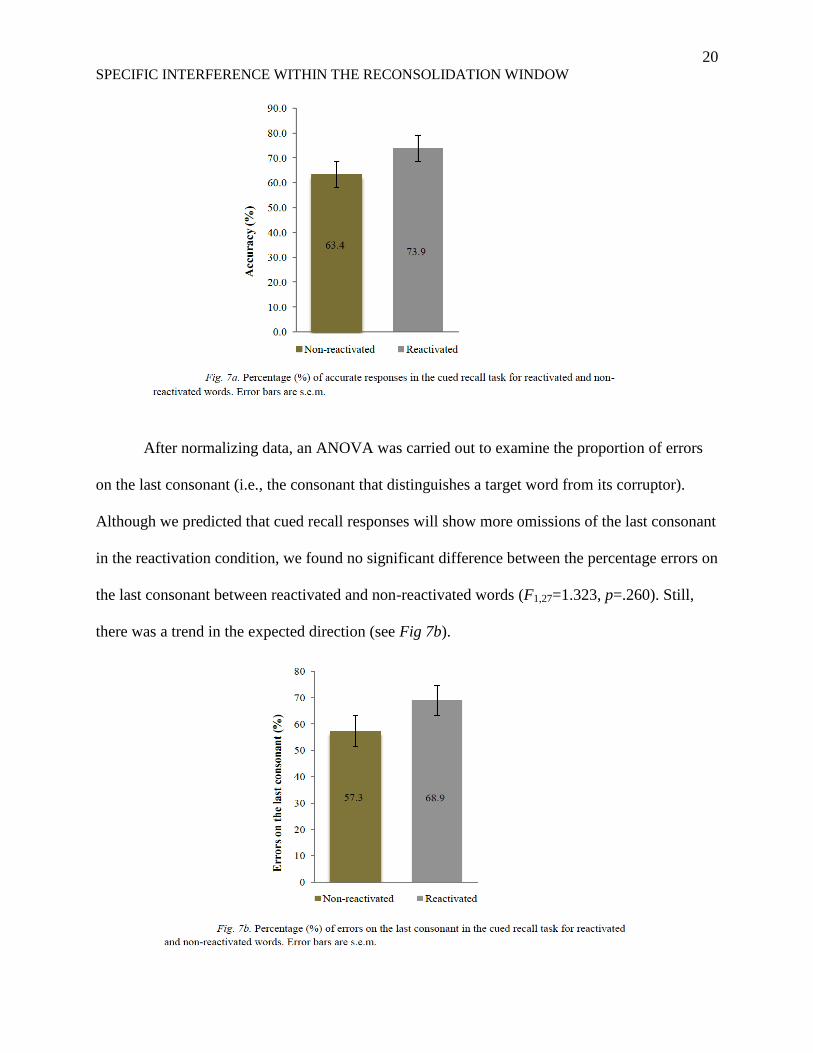
Responses on cued recall indicated better knowledge of reactivated words, which is
contrary to our predictions (cf. Fig. 7a). A repeated measures ANOVA conducted on the
proportion of accurate responses demonstrated that the difference was significant (F1,27=7.004,
p=.013). The difference was still significant after performing an arcsine transformation to
achieve normality (F1,27=7.785, p=.009).
20 SPECIFIC INTERFERENCE WITHIN THE RECONSOLIDATION WINDOW
After normalizing data, an ANOVA was carried out to examine the proportion of errors
on the last consonant (i.e., the consonant that distinguishes a target word from its corruptor).
Although we predicted that cued recall responses will show more omissions of the last consonant
in the reactivation condition, we found no significant difference between the percentage errors on
the last consonant between reactivated and non-reactivated words (F1,27=1.323, p=.260). Still,
there was a trend in the expected direction (see Fig 7b).

21 SPECIFIC INTERFERENCE WITHIN THE RECONSOLIDATION WINDOW
Pause detection
A repeated measures ANOVA was used to analyse performance on pause-present and
pause-absent trials. We assessed how lexical interference varies as a function of word condition
(i.e., reactivated, non-reactivated, baseline).
Response latencies. Only correct trials were included in the analysis. We recorded
responses between 200 ms and 2000 ms after stimulus presentation. There was a significant main
effect of word category on pause-present trials (Fig 8a; F2,58=3.512, p=.036). At odds with
predictions, subjects detected pauses in the reactivated words significantly more slowly than
those in the non-reactivated words (Fig. 8a, F1,29=4.395, p=.045). Yet, only performance on non-
reactivated words significantly differed from performance on baseline words (F1,29=6.640,
p=.015), with significantly faster responses on non-reactivated words. There was no significant
difference between reactivated words and baseline words (F1,29=.046, p<.832). Although there
was no significant main effect observed in the pause-absent trials, the trend was in line with
pause-present responses (Fig. 8a; F2,58=.492, p=.614).

22 SPECIFIC INTERFERENCE WITHIN THE RECONSOLIDATION WINDOW
Error rates. There was a significant effect of word category (F2,58=8.484, p<.001).
Analogous to latency scores, there were significantly more errors on reactivated words compared
to non-reactivated words (Fig 8b; F1,29=16.842; p<.001). In contrast with latencies, percentage
errors for non-reactivated words did not differ from those for baseline words (F1,29=.290,
p=.594). Only reactivated words differed from baseline words (F1,29=15.140, p<.001). Again,
there was no significant main effect of word category on pause-absent trials (F2,58=2.090,
p=.132), although responses aligned on average with performance on pause-resent trials.
Discussion
Knowledge of boundary conditions is crucial for theorizing reconsolidation, yet a
comprehensive account of these is missing. This study aimed to assess whether specific
interference can alter memories following reactivation. Interference specificity has been a
common denominator of studies that have obtained evidence for reconsolidation to date (e.g.,
Walker et al., 2003; Boccia et al., 2005; Boccia et al., 2010). To demonstrate unequivocally that

23 SPECIFIC INTERFERENCE WITHIN THE RECONSOLIDATION WINDOW
specificity of interference is a boundary condition for reconsolidation, one should be able to find
evidence of reconsolidation when using specific, but not unspecific interference – other factors
being held constant. A previous study has shown that unspecific interference does not influence
the reconsolidation process (Dumay & Tremblett, in prep.). Thus, we adapted the learning and
reactivation procedure used by Dumay and Tremblett (in prep.) in attempting to make a case that
specific interference is needed to observe reconsolidation. On the third experimental day, we
compared memories for reactivated words (which were expected to be in a plastic state prior to
potential interference) with memory for non-reactivated words (which were expected to be
robust to interference).
We expected that learning corruptors after memory reactivation would update memory
for reactivated words. Notwithstanding this prediction, we found that errors on the non-
overlapping consonant in cued-recall did not significantly differ between reactivation and non-
reactivation word conditions. We also anticipated that cued recall and phoneme monitoring
performance would reveal stronger memories for non-reactivated compared to reactivated words.
In sharp contrast to our expectations, cued recall responses indicated better memory strength for
reactivated than non-reactivated words. Phoneme monitoring revealed a similar pattern of
performance in the form of a non-significant trend: There were longer latencies and more errors
on non-reactivated words compared to reactivated words.
Our last prediction was that non-reactivated words and their respective corruptors would
be stored in separate memory traces, while reactivated words would blend with corruptors in one
updated memory. Thus, we anticipated that reactivated words would receive less interference in
a lexical competition task (i.e., the pause detection task) compared to non-reactivated words.
However, we found that reactivated words received more interference than non-reactivated

24 SPECIFIC INTERFERENCE WITHIN THE RECONSOLIDATION WINDOW
words (expressed as longer latencies and more errors in pause detection). This suggests that,
contrary to our prediction, reactivated words and corruptors were stored in separate memories.
In sum, it appears that reactivated memories were updated in strength instead of memory
content. We propose that this finding does not necessarily rebut the reconsolidation hypothesis.
To reconcile the discrepancy between our results and results obtained in previous reconsolidation
reports (Walker et al., 2003), we must question the ability of our experimental parameters to
initiate reconsolidation. At the moment, there is no reconsolidation protocol found to reliably
destabilize reactivated memories (Merlo et al., 2015). Drawing on available findings on
boundary conditions of reconsolidation, we derive at least four interpretations of our data.
First, we speculate that the reactivation protocol could have prevented reconsolidation
from proceeding. A number of studies have demonstrated that successful engagement in
reconsolidation processes depends on having a mismatch between what is expected to occur
upon reactivation and what actually occurs (Pedreira et al., 2004; Forcato et al., 2009; Sevenster
et al., 2013; Jacques & Schacter, 2013). This boundary condition has been regarded as prediction
error. The underlying principle is that encountering prediction error signals that the initial
learning is no longer adequate and new learning is required instead (Sevenster et al., 2013). In
the present study, the reactivation session consisted of the same experience that took place
during the learning phase, which means that there was no prediction error during reactivation.
This may have caused the memory to strengthen itself because information of the same quality
was acquired during the plastic period of the reactivated memory. In support of this argument, a
recent study found that lack of prediction error at reactivation resulted in memory strengthening
(Jaques & Schacter, 2013). In this study, subjects went on a museum tour carrying a camera that
was taking photos automatically. On the following day, they viewed photos that were

25 SPECIFIC INTERFERENCE WITHIN THE RECONSOLIDATION WINDOW
presumably taken by their camera. Half of the photos matched the encoding experience, while
the other half were modified versions of the original photos, being altered in angle, height, or
timing. A recognition test on the third day revealed that memories were stronger when
reactivation matched, rather than mismatched with encoding. Importantly, they found that the
mismatching retrieval session was also associated with increased false recognition of photos
illustrating stops that participants did not experience during the museum tour. The latter indicates
that having prediction error when retrieving an episodic memory can result in memory updating.
Based on these findings, it is tempting to speculate that the lack of prediction error might have
brought to the strengthening effect observed in reactivated words.
A second interpretation derives from limitations in how the prediction error hypothesis is
currently formulated. Most studies that used prediction error at reactivation proceeded on the
assumption that prediction error should occur throughout the reactivation session in order to
detect reconsolidation. However, Walker and associates (2003) provided evidence for
reconsolidation despite reactivating memories through a short reiteration of the learning phase.
Nonetheless, it can be argued that prediction error was still generated in this study. The study
used a target sequence (4-1-3-2-4) that overlapped with the corrupting sequence (2-3-1-4-2) in
content and sequence length, but differed in aspects such as finger repetition. On a similar note,
our study has attempted to induce prediction error by replacing the presentation of the
consolidated word frenzylk during sound detection with the novel word frenzyl. The
inconsistency between our findings and those by Walker and colleagues (2003) might arise from
using different degrees of similarity between target memories and corrupting learning. According
to the prediction error hypothesis, a large degree of similarity may prevent a mismatch from

26 SPECIFIC INTERFERENCE WITHIN THE RECONSOLIDATION WINDOW
occurring simply because only a small amount of information would be added to the original
memory.
As such, it is conceivable that our procedure might have failed to induce prediction error
by using corruptors that highly overlapped with target information. Future studies that aim to
investigate the role of interference specificity may benefit from using corruptors that overlap to
various degrees with initial learning. If further research does not validate the prediction that less
similarity allows reconsolidation to proceed, it may be that other characteristics of reactivation
deter the successful engagement in reconsolidation. Ideally, future research should find a method
of measuring the optimum degree of similarity without depending on the occurrence of
reconsolidation itself (which is constrained by other boundary conditions).
Thirdly, reactivation length may have influenced our results. Several associative memory
studies have found that repeated cue presentations upon memory reactivation lead to extinction
instead of the expected reconsolidation effects (Inda, Muravieva, & Alberini, 2011; Lee, Milton,
& Everitt, 2006). A shorter reactivation session, in contrast, resulted in reconsolidation.
Extinction represents a decrease in a conditioned response (Quirk & Mueller, 2008). Unlike
reconsolidation, extinction is underpinned by a mechanism that enables the formation of a
separate memory which, in turn, inhibits the previously learnt association. It has recently been
demonstrated that these two processes are mutually exclusive (Merlo et al., 2014). This study
showed that repeated cue presentations lead to the termination of the reconsolidation process.
Building on these findings, if the reactivation session was too long in our study, then the
corruptors appeared outside the reconsolidation window. Following this line of thought,
corruptors were actually learnt when memories were no longer plastic. As a result, they did not
produce the expected shift in the initial memory.

27 SPECIFIC INTERFERENCE WITHIN THE RECONSOLIDATION WINDOW
The increase in memory strength concurrent with reactivation might have then resulted
from rehearsing the initial memory during the reconsolidation window. Several studies have
obtained strengthening effects using different reactivation durations and parameters of memory
strength and age (Inda et al., 2011; Rodriguez-Ortiz, Balderas, Garcia-DeLaTorre, & Bermudez-
Rattoni, 2012; Lee, 2008; Karpicke & Roediger, 2008; Roediger & Butler, 2011, Coccoz,
Maldonado, & Delorenzi, 2011). It still remains an open question how the reactivation length
varies as a function of memory strength and memory age. At this stage, we cannot draw
definitive conclusions about the adequacy of our reactivation length.
Lastly, we should consider the possibility that the reactivation session has been too short
to induce memory destabilisation. This interpretation is supported by findings by Bustos and
colleagues (2009). In this study, one-minute reactivation of a strong associative fear memory did
not induce reconsolidation, while a three-to-five-minutes reactivation session successfully led to
reconsolidation. This suggests that insufficient reactivation leaves reactivated memories in an
insensitive state. It is tempting to argue that the consolidation of corruptors as separate memory
traces arouse from the fact that memories were not destabilised from the very beginning.
However, our results show a memory boost concurrent with reactivation. This position would
predict that memory strengthening occurs irrespective of the fact that memories are in a plastic or
in an insensitive state. If memory strengthening cannot occur outside the reconsolidation
window, then the memory has been successfully destabilised in this instance. While such cases
have yet not been reported, available evidence only shows a correlation between reconsolidation
and memory strengthening, and not a causal effect (Alberini, 2011).

28 SPECIFIC INTERFERENCE WITHIN THE RECONSOLIDATION WINDOW
Conclusion and Future Directions
In conclusion, we show that the manipulation of reconsolidation through exposure to
similar learning after reactivation can boost the initial memory. Although we expected that
specific interference would affect memory reconsolidation, our results did not support this
prediction. This study highlights a number of gaps in the literature that reduce the feasibility of
disrupting reconsolidation. Past research has obtained mixed results using different reactivation
paradigms. Determining the characteristics of reactivation that induce reconsolidation would be a
fruitful avenue for research. In the absence of a framework that could guide the choice of
reactivation duration for declarative memory reconsolidation, it is impossible to make firm
assertions about the ability of the retrieval session to induce reconsolidation based on associative
memory research per se. Also, we cannot fully disambiguate between an explanation that
pertains to lack of prediction error at reactivation and one that pertains to reactivation length.
While research has been clear about the necessity to induce prediction error, how to induce
prediction error remains an open question. Outside the issues with the retrieval session, it
remains to be established whether a high degree of similarity between corruptors and target
memory can conflict with prediction error. It is difficult to draw conclusions about the right
amount of similarity based on reconsolidation itself. If anything, determining boundary
conditions for reconsolidation would be a fruitful avenue for research.
The present study is the first to investigate the need to use specific interference to observe
reconsolidation using a three-day procedure. The use of a three-day procedure, instead of a two-
day procedure as in Chan and LaPaglia (2013) avoids potential difficulties with estimating the
timeframe of the reconsolidation window. An understanding of the limits and operating
conditions of reconsolidation is crucial for understanding mechanisms responsible for plasticity

29 SPECIFIC INTERFERENCE WITHIN THE RECONSOLIDATION WINDOW
in human memories. In addition to its theoretical value, the possibility to alter memories could
also have therapeutic value. Retrieval-induced plasticity could be manipulated to alter
consolidated pathogenic memories for PTSD (Brunet et al., 2011). Since situations that offer
opportunities for memory updating can alternatively strengthen the original memory and lead to
new learning, it is essential to be able to select optimal reactivation and interference specificity
parameters in order to attain the expected memory outcome.

30 SPECIFIC INTERFERENCE WITHIN THE RECONSOLIDATION WINDOW
References
Agren, T. (2014). Human reconsolidation: A reactivation and update. Brain Research
Bulletin, 105, 70-82.
Alberini, C. M. (2011). The role of reconsolidation and the dynamic process of long-term
memory formation and storage. Frontiers in Behavioral Neuroscience, 5.
Alberini, C. M., & LeDoux, J. E. (2013). Memory reconsolidation. Current Biology, 23(17),
746-750.
Besnard, A., Caboche, J., & Laroche, S. (2012). Reconsolidation of memory: A decade of
debate. Progress in Neurobiology, 99(1), 61-80.
Boccia, M. M., Blake, M. G., Acosta, G. B., & Baratti, C. M. (2005). Memory consolidation
and reconsolidation of an inhibitory avoidance task in mice: Effects of a new different
learning task. Neuroscience, 135(1), 19-29.
Boccia, M. M., Blake, M. G., Krawczyk, M. C., & Baratti, C. M. (2010). Hippocampal
alpha7 nicotinic receptors modulate memory reconsolidation of an inhibitory
avoidance task in mice. Neuroscience, 171(2), 531-543.
Brunet, A., Orr, S. P., Tremblay, J., Robertson, K., Nader, K., & Pitman, R. K. (2008). Effect
of post-retrieval propranolol on psychophysiologic responding during subsequent
script-driven traumatic imagery in post-traumatic stress disorder. Journal of
Psychiatric Research, 42(6), 503-506.
Brunet, A., Poundja, J., Tremblay, J., Bui, É., Thomas, É., Orr, S. P., ... & Pitman, R. K.
(2011). Trauma reactivation under the influence of propranolol decreases
posttraumatic stress symptoms and disorder: Three open-label trials. Journal of
Clinical Psychopharmacology, 31(4), 547-550.

31 SPECIFIC INTERFERENCE WITHIN THE RECONSOLIDATION WINDOW
Bustos, S. G., Maldonado, H., & Molina, V. A. (2009). Disruptive effect of midazolam on
fear memory reconsolidation: Decisive influence of reactivation time span and
memory age. Neuropsychopharmacology, 34(2), 446-457.
Censor, N., Dimyan, M. A., & Cohen, L. G. (2010). Modification of existing human motor
memories is enabled by primary cortical processing during memory
reactivation. Current Biology, 20(17), 1545-1549.
Censor, N., Dimyan, M.A., & Cohen, L. G. (2014). Cortico-subcortical neuronal circuitry
associated with reconsolidation of human procedural memories. Cortex, 58, 281-288.
Chan, J. C., & LaPaglia, J. A. (2013). Impairing existing declarative memory in humans by
disrupting reconsolidation. Proceedings of the National Academy of
Sciences, 110(23), 9309-9313.
Coccoz, V., Maldonado, H., & Delorenzi, A. (2011). The enhancement of reconsolidation
with a naturalistic mild stressor improves the expression of a declarative memory in
humans. Neuroscience, 185, 61-72.
Debiec, J., Doyère, V., Nader, K., & LeDoux, J. E. (2006). Directly reactivated, but not
indirectly reactivated, memories undergo reconsolidation in the
amygdala. Proceedings of the National Academy of Sciences of the United States of
America, 103(9), 3428-3433.
Debiec, J., & LeDoux, J. E. (2004). Disruption of reconsolidation but not consolidation of
auditory fear conditioning by noradrenergic blockade in the
amygdala. Neuroscience, 129(2), 267-272.
Debiec, J., LeDoux, J. E., & Nader, K. (2002). Cellular and systems reconsolidation in the
hippocampus. Neuron, 36(3), 527-538.

32 SPECIFIC INTERFERENCE WITHIN THE RECONSOLIDATION WINDOW
Deliens, G., Schmitz, R., Caudron, I., Mary, A., Leproult, R., & Peigneux, P. (2013). Does
recall after sleep-dependent memory consolidation reinstate sensitivity to retroactive
interference. PloS one, 8(7), e68727.
Dudai, Y. (2012). The restless engram: Consolidations never end. Annual Review of
Neuroscience, 35, 227-247.
Dudai, Y., & Eisenberg, M. (2004). Rites of passage of the engram: reconsolidation and the
lingering consolidation hypothesis. Neuron, 44(1), 93-100.
Dumay, N. & Tremblett, N. (in prep.). Do declarative memories reconsolidate whenever
recalled? Unpublished thesis.
Dumay, N., & Gaskell, M. G. (2012). Overnight lexical consolidation revealed by speech
segmentation. Cognition, 123(1), 119-132.
Duvarci, S., & Nader, K. (2004). Characterization of fear memory reconsolidation. The
Journal of Neuroscience, 24(42), 9269-9275.
Forcato, C., Argibay, P. F., Pedreira, M. E., & Maldonado, H. (2009). Human
reconsolidation does not always occur when a memory is retrieved: the relevance of
the reminder structure. Neurobiology of Learning and Memory, 91(1), 50-57.
Forcato, C., Burgos, V. L., Argibay, P. F., Molina, V. A., Pedreira, M. E., & Maldonado, H.
(2007). Reconsolidation of declarative memory in humans. Learning &
Memory, 14(4), 295-303.
Frauenfelder, U. & Peeters. G. (1990). Lexical segmentation in TRACE: An exercise in
simulation. In G.E. Altmann (Ed.), Cognitive models of speech processing.
Cambridge, MA: MIT Press.

33 SPECIFIC INTERFERENCE WITHIN THE RECONSOLIDATION WINDOW
Frauenfelder, U., Segui, J., & Dijkstra, T. (1990). Lexical effects in phonemic processing:
Facilitatory or inhibitory? Journal of Experimental Psychology: Human Perception
and Performance, 16(1), 77.
Gaskell, M. G., & Dumay, N. (2003). Lexical competition and the acquisition of novel
words. Cognition, 89(2), 105-132.
Hardt, O., Einarsson, E. Ö., & Nader, K. (2010). A bridge over troubled water:
reconsolidation as a link between cognitive and neuroscientific memory research
traditions. Annual Review of Psychology, 61, 141-167.
Hupbach, A. (2011). The specific outcomes of reactivation-induced memory changes depend
on the degree of competition between old and new information. Frontiers in
Behavioral Neuroscience, 5, 33.
Hupbach, A., Hardt, O., Gomez, R., & Nadel, L. (2008). The dynamics of memory: Context-
dependent updating. Learning & Memory, 15(8), 574-579.
Inda, M. C., Muravieva, E. V., & Alberini, C. M. (2011). Memory retrieval and the passage
of time: from reconsolidation and strengthening to extinction. The Journal of
Neuroscience, 31(5), 1635-1643.
Jacques, P. L. S., & Schacter, D. L. (2013). Modifying memory selectively enhancing and
updating personal memories for a museum tour by reactivating them. Psychological
Science, 24(4), 537-543.
Karpicke, J. D., & Roediger, H. L. (2008). The critical importance of retrieval for learning.
Science, 319(5865), 966-968.
Kroes, M. C., Tendolkar, I., van Wingen, G. A., van Waarde, J. A., Strange, B. A., &
Fernández, G. (2014). An electroconvulsive therapy procedure impairs

34 SPECIFIC INTERFERENCE WITHIN THE RECONSOLIDATION WINDOW
reconsolidation of episodic memories in humans. Nature Neuroscience, 17(2), 204-
206.
Lee, J. L. (2008). Memory reconsolidation mediates the strengthening of memories by
additional learning. Nature Neuroscience, 11(11), 1264-1266.
Lee, J. L. (2010). Memory reconsolidation mediates the updating of hippocampal memory
content. Frontiers in Behavioral Neuroscience, 4.
Lee, J. L., Ciano, P., Thomas, K. L., & Everitt, B. J. (2005). Disrupting reconsolidation of
drug memories reduces cocaine-seeking behavior. Neuron, 47(6), 795-801.
Lee, J. L., Everitt, B. J., & Thomas, K. L. (2004). Independent cellular processes for
hippocampal memory consolidation and reconsolidation. Science, 304(5672), 839-
843.
Lee, J. L., Milton, A. L., & Everitt, B. J. (2006). Reconsolidation and extinction of
conditioned fear: inhibition and potentiation. The Journal of Neuroscience, 26(39),
10051-10056.
Mattys, S. L., & Clark, J. H. (2002). Lexical activity in speech processing: Evidence from
pause detection. Journal of Memory and Language, 47(3), 343-359.
McClelland, J. L., & Elman, J. L. (1986). The TRACE model of speech
perception. Cognitive Psychology, 18(1), 1-86.
McGaugh, J. L. (1966). Time-dependent process in memory storage. Science, 15 (1351).
McGaugh, J. L. (2000). Memory – a century of consolidation. Science, 287(5451), 248-251.
Mednick, S. C., Cai, D. J., Shuman, T., Anagnostaras, S., & Wixted, J. T. (2011). An
opportunistic theory of cellular and systems consolidation. Trends in
Neurosciences, 34(10), 504-514.

35 SPECIFIC INTERFERENCE WITHIN THE RECONSOLIDATION WINDOW
Merlo, E., Milton, A. L., & Everitt, B. J. (2015). Enhancing cognition by affecting memory
reconsolidation. Current Opinion in Behavioral Sciences, 4, 41-47.
Merlo, E., Milton, A. L., Goozée, Z. Y., Theobald, D. E., & Everitt, B. J. (2014).
Reconsolidation and extinction are dissociable and mutually exclusive processes:
Behavioral and molecular evidence. The Journal of Neuroscience, 34(7), 2422-2431.
Milekic, M. H., & Alberini, C. M. (2002). Temporally graded requirement for protein
synthesis following memory reactivation. Neuron, 36(3), 521-525.
Misanin, J. R., Miller, R. R., & Lewis, D. J. (1968). Retrograde amnesia produced by
electroconvulsive shock after reactivation of a consolidated memory trace. Science,
160(3827), 554-555.
Morris, R. G., Inglis, J., Ainge, J. A., Olverman, H. J., Tulloch, J., Dudai, Y., & Kelly, P. A.
(2006). Memory reconsolidation: Sensitivity of spatial memory to inhibition of
protein synthesis in dorsal hippocampus during encoding and
retrieval. Neuron, 50(3), 479-489.
Müller, G. E., & Pilzecker, A. (1900). Experimental contributions to the theory of memory. Z
Psychol Z Angew Psychol, 1, 1-288.
Nader, K., Schafe, G. E., & LeDoux, J. E. (2000). Fear memories require protein synthesis in
the amygdala for reconsolidation after retrieval. Nature, 406(6797), 722-726.
Pedreira, M. E., Pérez-Cuesta, L. M., & Maldonado, H. (2004). Mismatch between what is
expected and what actually occurs triggers memory reconsolidation or
extinction. Learning & Memory, 11(5), 579-585.

36 SPECIFIC INTERFERENCE WITHIN THE RECONSOLIDATION WINDOW
Poundja, J., Sanche, S., Tremblay, J., & Brunet, A. (2012). Trauma reactivation under the
influence of propranolol: An examination of clinical predictors. European Journal of
Psychotraumatology, 3.
Quirk, G. J., & Mueller, D. (2008). Neural mechanisms of extinction learning and retrieval.
Neuropsychopharmacology, 33(1), 56-72.
Robertson, E. M. (2012). New insights in human memory interference and
consolidation. Current Biology, 22(2), R66-R71.
Rodriguez-Ortiz, C. J., Balderas, I., Garcia-DeLaTorre, P., & Bermudez-Rattoni, F. (2012).
Taste aversion memory reconsolidation is independent of its retrieval. Neurobiology
of Learning and Memory, 98(3), 215-219.
Roediger, H. L., & Butler, A. C. (2011). The critical role of retrieval practice in long-term
retention. Trends in Cognitive Sciences, 15(1), 20-27.
Schiller, D., & Phelps, E. A. (2011). Does reconsolidation occur in humans? Frontiers in
Behavioral Neuroscience, 5, 24.
Sederberg, P. B., Gershman, S. J., Polyn, S. M., & Norman, K. A. (2011). Human memory
reconsolidation can be explained using the temporal context model. Psychonomic
Bulletin & Review, 18(3), 455-468.
Sevenster, D., Beckers, T., & Kindt, M. (2013). Prediction error governs pharmacologically
induced amnesia for learned fear. Science, 339(6121), 830-833.
Soeter, M., & Kindt, M. (2010). Dissociating response systems: Erasing fear from memory.
Neurobiology of Learning and Memory, 94(1), 30-41.
Stickgold, R., & Walker, M. P. (2007). Sleep-dependent memory consolidation and
reconsolidation. Sleep Medicine, 8(4), 331-343.

37 SPECIFIC INTERFERENCE WITHIN THE RECONSOLIDATION WINDOW
Suzuki, A., Josselyn, S. A., Frankland, P. W., Masushige, S., Silva, A. J., & Kida, S. (2004).
Memory reconsolidation and extinction have distinct temporal and biochemical
signatures. The Journal of Neuroscience, 24(20), 4787-4795.
Tronel, S., & Alberini, C. M. (2007). Persistent disruption of a traumatic memory by
postretrieval inactivation of glucocorticoid receptors in the amygdala. Biological
Psychiatry, 62(1), 33-39.
Tronel, S., Milekic, M. H., & Alberini, C. M. (2005). Linking new information to a
reactivated memory requires consolidation and not reconsolidation
mechanisms. PLoS biology, 3(9), 1630.
Tronson, N. C., & Taylor, J. R. (2007). Molecular mechanisms of memory
reconsolidation. Nature Reviews Neuroscience, 8(4), 262-275.
Walker, M. P., Brakefield, T., Hobson, J. A., & Stickgold, R. (2003). Dissociable stages of
human memory consolidation and reconsolidation. Nature, 425(6958), 616-620.
Wixted, J. T. (2004). The psychology and neuroscience of forgetting. Annual Review of
Psychology, 55, 235-269.

38 SPECIFIC INTERFERENCE WITHIN THE RECONSOLIDATION WINDOW
Appendix A
Ethics approval

39 SPECIFIC INTERFERENCE WITHIN THE RECONSOLIDATION WINDOW
Appendix B
Consent form


















