
Discovery and engineering of redox enzymes · light production halogenation magnetosensing and...
24
18-10-2017 1 Discovery and engineering of redox enzymes Marco Fraaije Molecular Enzymology University of Groningen [email protected] www.gecco-biotech.com HORIZON 2020 SUMMER SCHOOL 2017 Biocatalysis as a Key Enabling Technology CarbaZymes / ROBOX / Metafluidics EU Horizon 2020 Innovation Action “ROBOX” Expanding the industrial use of Robust Oxidative Biocatalysts for the conversion and production of alcohols. “The research for this work has received funding from the European Union (EU) project ROBOX (grant agreement n°635734) under EU’s Horizon 2020 Programme Research and Innovation actions H2020-LEIT BIO-2014-1. Any statement made herein reflects only the author’s views. The European Union is not liable for any use that may be made of the information contained herein.” flavoenzymes: versatile biocats see Youtube for a movie on enzyme engineering: search for ‘enzyme’ and ‘robox’
Transcript of Discovery and engineering of redox enzymes · light production halogenation magnetosensing and...

18-10-2017
1
Discovery and engineering
of redox enzymes
Marco Fraaije
Molecular Enzymology
University of Groningen
www.gecco-biotech.com
HORIZON 2020 SUMMER SCHOOL 2017
Biocatalysis as a Key Enabling Technology
CarbaZymes / ROBOX / Metafluidics
EU Horizon 2020 Innovation Action “ROBOX”
Expanding the industrial use of Robust Oxidative Biocatalysts for the conversion and production of alcohols.
“The research for this work has received funding from the European Union (EU) project ROBOX (grant agreement n°635734) under EU’s Horizon 2020 Programme Research and Innovation actions
H2020-LEIT BIO-2014-1. Any statement made herein reflects only the author’s views. The European Union is not liable for any use that may be made of the information contained herein.”
flavoenzymes: versatile biocats
see Youtube for a movie on enzyme engineering:
search for ‘enzyme’ and ‘robox’

18-10-2017
2
focus on flavin-dependent enzymes
why?
flavins are the most versatile cofactors
oxidation
reduction light sensinge- transporthydrolysis
halogenationlight production
magnetosensing
and more...
focus on flavin-dependent enzymes
18-10-2017
3
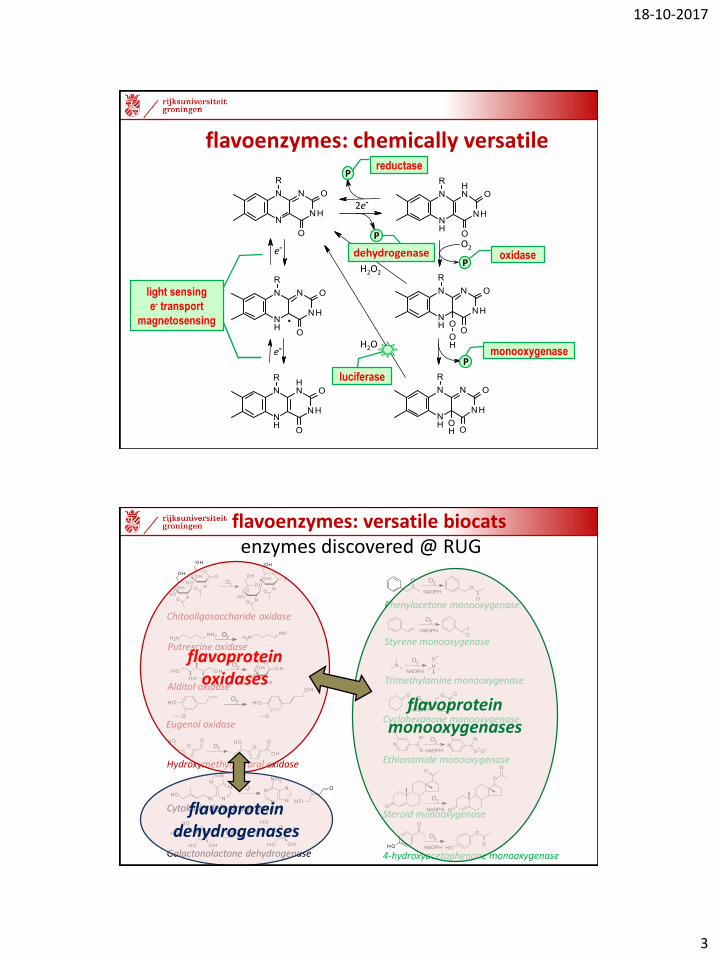
flavoenzymes: chemically versatile
.
e-
e-
2e-
H2O2
H2O
O2
P
Preductase
P
dehydrogenaseP
oxidase
luciferase
light sensing
e- transport
magnetosensing
PPmonooxygenase
Alditol oxidase
Putrescine oxidase
Chitooligosaccharide oxidase
Eugenol oxidase
Hydroxymethylfurfural oxidase
Cytokinin dehydrogenase
Galactonolactone dehydrogenase
Phenylacetone monooxygenase
Styrene monooxygenase
Trimethylamine monooxygenase
Cyclohexanone monooxygenase
Ethionamide monooxygenase
Steroid monooxygenase
4-hydroxyacetophenone monooxygenase
O2
O2
O2
O2
CytC
NADPH
O2
NADPH
O2
NADPH
O2
NADPH
O2
NADPH
O2
NADPH
O2
NADPH
O2
Q
O2
O2
enzymes discovered @ RUG
flavoproteinoxidases
flavoproteinmonooxygenases
flavoproteindehydrogenases
flavoenzymes: versatile biocats
18-10-2017
4
Alditol oxidase
Putrescine oxidase
Chitooligosaccharide oxidase
Eugenol oxidase
Hydroxymethylfurfural oxidase
Cytokinin dehydrogenase
Galactonolactone dehydrogenase
Phenylacetone monooxygenase
Styrene monooxygenase
Trimethylamine monooxygenase
Cyclohexanone monooxygenase
Ethionamide monooxygenase
Steroid monooxygenase
4-hydroxyacetophenone monooxygenase
O2
O2
O2
O2
CytC
NADPH
O2
NADPH
O2
NADPH
O2
NADPH
O2
NADPH
O2
NADPH
O2
NADPH
O2
Q
O2
O2
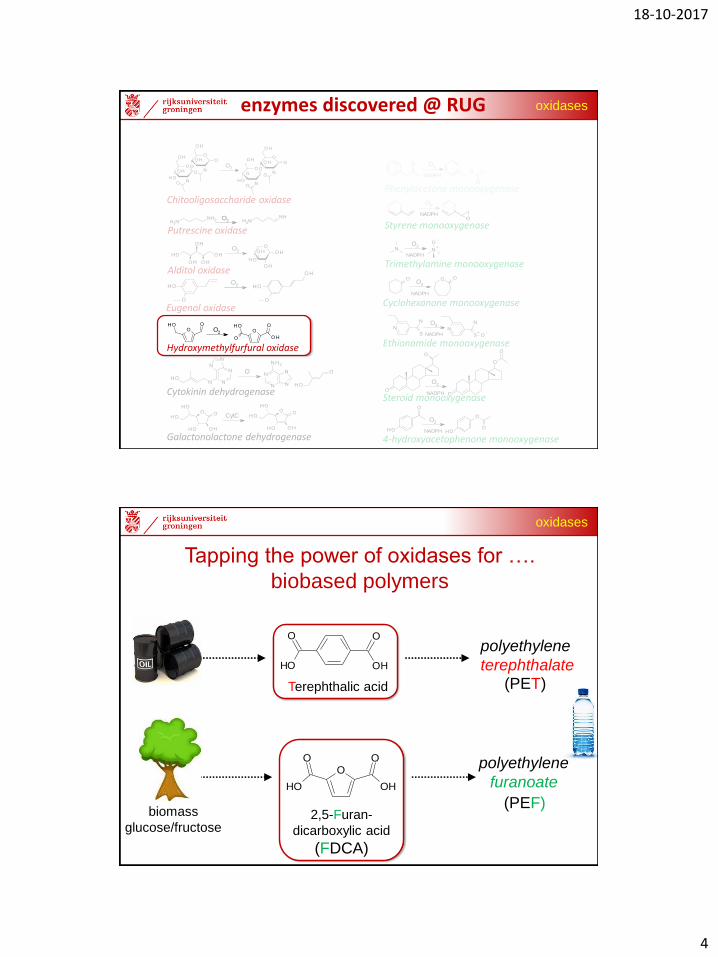
oxidasesenzymes discovered @ RUG
Tapping the power of oxidases for ….
biobased polymers
2,5-Furan-
dicarboxylic acid
(FDCA)
biomass
glucose/fructose
(PEF)
O
OH
OO
OH
polyethylene
furanoate
Terephthalic acid (PET)
OH
OO
OH
polyethylene
terephthalate
oxidases
18-10-2017
5
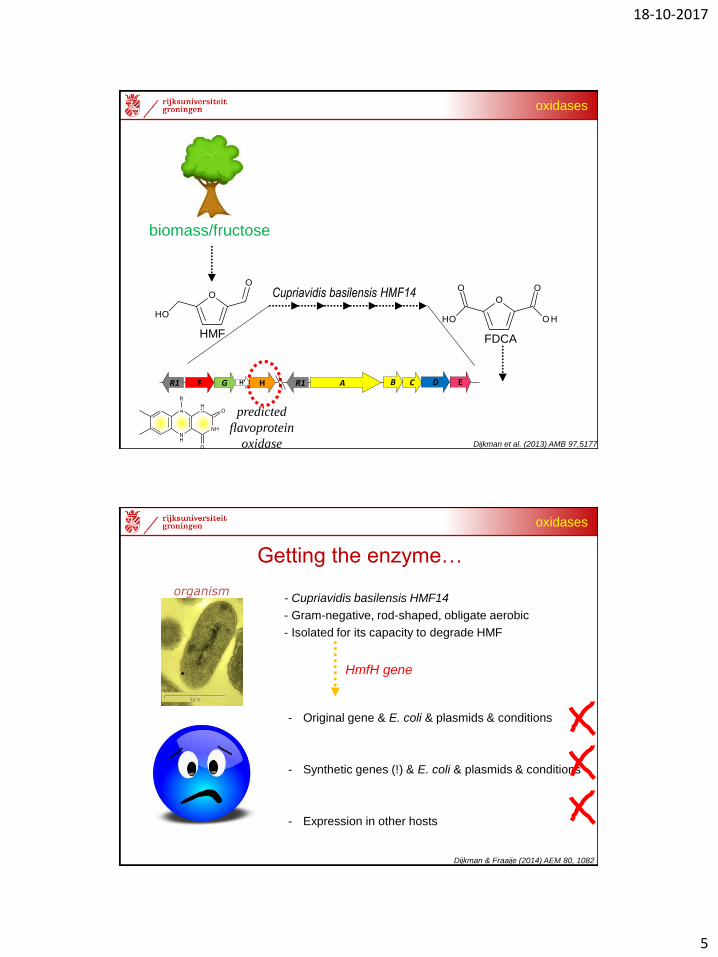
Cupriavidis basilensis HMF14
Dijkman et al. (2013) AMB 97,5177
HMF FDCA
biomass/fructose
predicted
flavoprotein
oxidase
oxidases
R1 F G H R1 A B C D EH′
O
NH
N
R
NH
O
NH
- Original gene & E. coli & plasmids & conditions
- Synthetic genes (!) & E. coli & plasmids & conditions
- Expression in other hosts
- Cupriavidis basilensis HMF14
- Gram-negative, rod-shaped, obligate aerobic
- Isolated for its capacity to degrade HMF
HmfH gene
organism
Getting the enzyme…
Dijkman & Fraaije (2014) AEM 80, 1082
oxidases

18-10-2017
6
- overexpression in Escherichia coli (100 mg/L culture!)
- soluble and stable monomeric protein
- FAD cofactor as prosthetic group
- Methylovorus sp. Strain MP688: methylotroph
- genome: 2.9 Mb (55 % GC), 2719 ORFs
organism
HmfH homolog (46 % protein seq. ident.)
Getting the enzyme!
Dijkman & Fraaije (2014) AEM 80, 1082
oxidases
- Methylovorus sp. Strain MP688: methylotroph
- genome: 2.9 Mb (55 % GC), 2719 ORFs
organism
HmfH homolog (gene ordered)
HMFOHMFOHMFO
HMF FDCA
Testing the enzyme!
Dijkman & Fraaije (2014) AEM 80, 1082
oxidases

18-10-2017
7
OH
SH
O
S
OH
OH
OOHOH
OH
O
OH
O
O
SH SH
HMFO also oxidizes thiols!
HMFO takes many compounds
Example substrates
Example reactions
Ewing, Dijkman et al. (2014) Angewandte 53, 13206
O
SH SH
O
SH SH
oxidases
HMF-OH HMF
HMF-acid
FFA FDCA
Crystal structure exploited to tune activity
Pathway of HMF oxidation by HMFO
OOHHO
OOHO
OOHO
HO
OOO
OOO
HO
OOO
HO OH
double mutant was designed
V367R-W466F HFMO
10000x active on FFA!
Crystal structure of HMFO
substrate
oxidases
Dijkman et al. (2015) ACS Catalysis 5, 1833

18-10-2017
8
Alditol oxidase
Putrescine oxidase
Chitooligosaccharide oxidase
Eugenol oxidase
Hydroxymethylfurfural oxidase
Cytokinin dehydrogenase
Galactonolactone dehydrogenase
O2
O2
O2
O2
CytC
Q
O2
O2
O2
Baron et al. PNAS 2009
oxidases
enzymes discovered @ RUG
Alditol oxidase
• hyperexpression in E. coli
(400 mg/L)
• soluble monomeric protein
• FAD cofactor incorporated
• active with alditols
• Streptomyces coelicolor A3(2): soil bacterium
• notable for production of pharmaceutically useful compounds
• genome: 8.7 Mb (72 % GC), 7825 ORFs
Heuts et al. J. Biol.Chem. 2007
Discovery of alditol oxidase (AldO)
xylitol oxidase homolog identified (76% seq. ident.)
predicted to contain histidyl-FAD
extract!
protein flavin
oxidases

18-10-2017
9
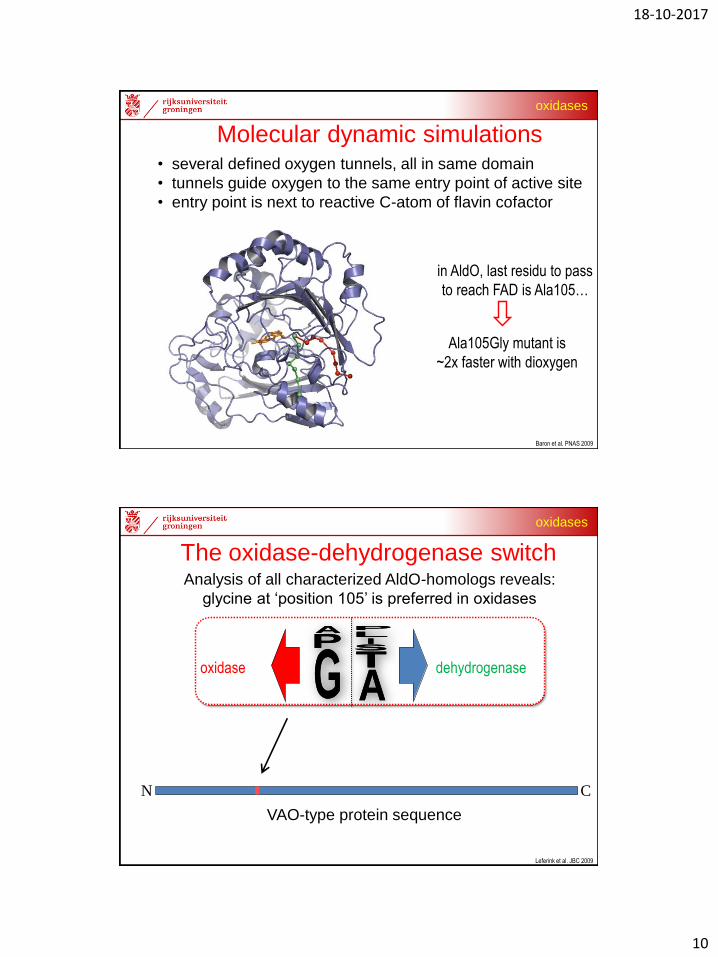
specific tunnel(s) or, random diffusion to active site?
What determines whether a flavoprotein
can use dioxygen as electron acceptor??
To be or not to be an oxidase
Alditol Oxidase: ideal model enzyme• expression system (high amounts, mutants)
• stable monomeric enzyme, facile purification
• full kinetic mechanism elucidated
• structures: high resolution & complexed structures
1. perform enhanced-statistics MD simulations
2. verify experimentally
Baron et al. PNAS 2009
oxidases
Baron et al.
PNAS 2009
• 1 monomer AldO (all atoms), waters, salt, dioxygens
• 50 ns unbiased calculations (~2 months)
• 500 pathways
Molecular dynamic simulations
oxidases
18-10-2017
10
Baron et al. PNAS 2009
Molecular dynamic simulations• several defined oxygen tunnels, all in same domain
• tunnels guide oxygen to the same entry point of active site
• entry point is next to reactive C-atom of flavin cofactor
in AldO, last residu to pass
to reach FAD is Ala105…
Ala105Gly mutant is
~2x faster with dioxygen
oxidases
Leferink et al. JBC 2009
IL
A
GP S
A
P
T
oxidase dehydrogenase
IL
A
GP S
A
P
T
oxidase dehydrogenase
oxidase dehydrogenase
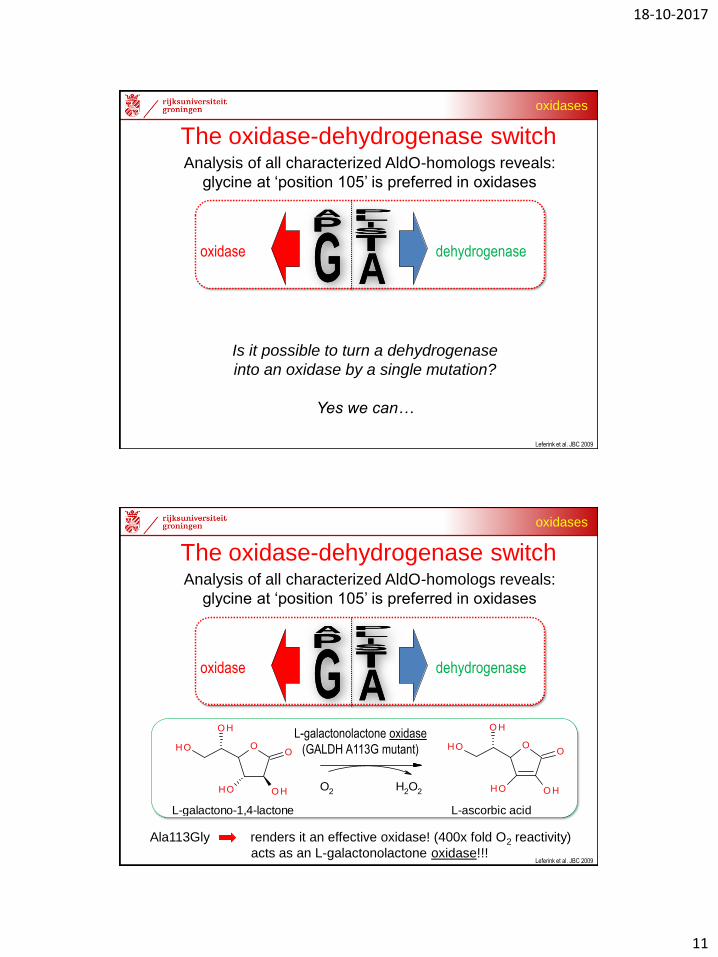
Analysis of all characterized AldO-homologs reveals:
glycine at ‘position 105’ is preferred in oxidases
The oxidase-dehydrogenase switch
N C
VAO-type protein sequence
oxidases
18-10-2017
11
Leferink et al. JBC 2009
IL
A
GP S
A
P
T
oxidase dehydrogenase
IL
A
GP S
A
P
T
oxidase dehydrogenase
oxidase dehydrogenase
Analysis of all characterized AldO-homologs reveals:
glycine at ‘position 105’ is preferred in oxidases
The oxidase-dehydrogenase switch
Is it possible to turn a dehydrogenase
into an oxidase by a single mutation?
Yes we can…
oxidases
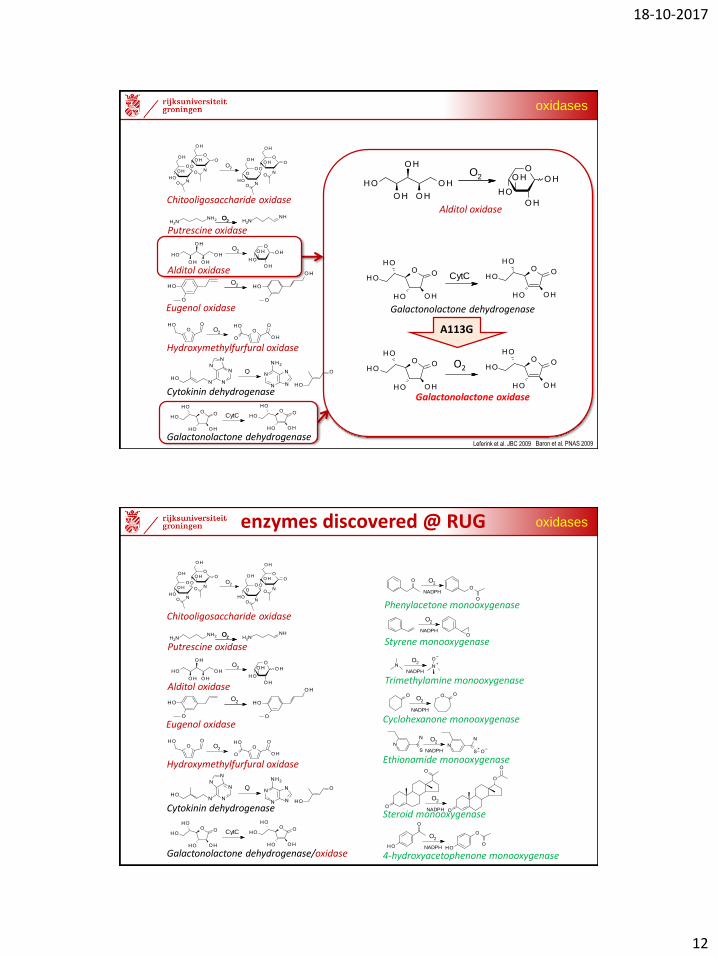
L-galactonolactone oxidase
(GALDH A113G mutant)
Ala113Gly renders it an effective oxidase! (400x fold O2 reactivity)
acts as an L-galactonolactone oxidase!!!
O2 H2O2
L-galactono-1,4-lactone L-ascorbic acid
IL
A
GP S
A
P
T
oxidase dehydrogenase
IL
A
GP S
A
P
T
oxidase dehydrogenase
Analysis of all characterized AldO-homologs reveals:
glycine at ‘position 105’ is preferred in oxidases
The oxidase-dehydrogenase switch
Leferink et al. JBC 2009
oxidases
oxidase dehydrogenase
18-10-2017
12
Alditol oxidase
Putrescine oxidase
Chitooligosaccharide oxidase
Eugenol oxidase
Hydroxymethylfurfural oxidase
Cytokinin dehydrogenase
Galactonolactone dehydrogenase
O2
O2
O2
O2
CytC
Q
O2
O2
Baron et al. PNAS 2009
Galactonolactone dehydrogenase
Galactonolactone oxidase
A113G
Leferink et al. JBC 2009
oxidases
O2
Alditol oxidase
CytC
CytCO2
Alditol oxidase
Putrescine oxidase
Chitooligosaccharide oxidase
Eugenol oxidase
Hydroxymethylfurfural oxidase
Cytokinin dehydrogenase
Galactonolactone dehydrogenase/oxidase
Phenylacetone monooxygenase
Styrene monooxygenase
Trimethylamine monooxygenase
Cyclohexanone monooxygenase
Ethionamide monooxygenase
Steroid monooxygenase
4-hydroxyacetophenone monooxygenase
O2
O2
O2
O2
CytC
NADPH
O2
NADPH
O2
NADPH
O2
NADPH
O2
NADPH
O2
NADPH
O2
NADPH
O2
Q
O2
O2
oxidasesenzymes discovered @ RUG

18-10-2017
13
Phenylacetone monooxygenase
Styrene monooxygenase
Trimethylamine monooxygenase
Cyclohexanone monooxygenase
Ethionamide monooxygenase
Steroid monooxygenase
4-hydroxyacetophenone monooxygenase
NADPH
O2
NADPH
O2
NADPH
O2
NADPH
O2
NADPH
O2
NADPH
O2
NADPH
O2
NADPHO2
spontaneous
indigo blue
trimethylaminemonooxygenase
monooxygenasesenzymes discovered @ RUG
Alditol oxidase
Putrescine oxidase
Chitooligosaccharide oxidase
Eugenol oxidase
Hydroxymethylfurfural oxidase
Cytokinin dehydrogenase
Galactonolactone dehydrogenase/oxidase
Phenylacetone monooxygenase
Styrene monooxygenase
Trimethylamine monooxygenase
Cyclohexanone monooxygenase
Ethionamide monooxygenase
Steroid monooxygenase
4-hydroxyacetophenone monooxygenase
O2
O2
O2
O2
CytC
NADPH
O2
NADPH
O2
NADPH
O2
NADPH
O2
NADPH
O2
NADPH
O2
NADPH
O2
Q
O2
O2
monooxygenasesenzymes discovered @ RUG

18-10-2017
14
- overexpression in Escherichia coli
- soluble monomeric protein (62 kDa)
- stable at high temperatures
- tolerant to solvents
- first BVMO crystal structure
- mainly active on aromatic substrates
- Thermobifida fusca: moderate thermophilic soil bacterium
- typical growth temperature: 55°C
- genome: 3.7 Mb (67 % GC), 4587 ORFs
organism
enzyme
motif-based BVMO gene identification
Malito et al., Proc. Natl. Acad. Sci. USA 2004; Fraaije et al., Appl.Microbiol.Biotechnol. 2005; de Gonzalo et al., Tetrahedron: Asymmetry 2005
Phenylacetone monooxygenase (PAMO)
PAMO
NADPH + O2
monooxygenases
- newly accepted substrates by PAMO M446G
- formation of indigo blue
M446 G446
Redesign of substrate binding pocket
Torres Pazmiño et al., Adv. Synth. Catal. 2007
spontaneous
monooxygenases
18-10-2017
15
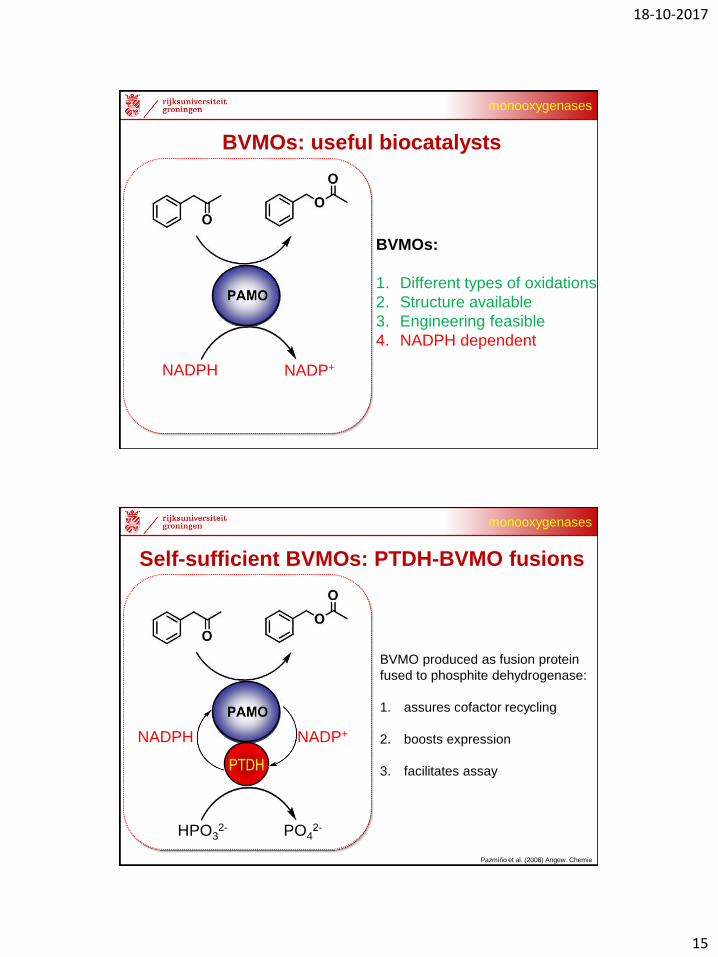
BVMOs: useful biocatalysts
BVMOs:
1. Different types of oxidations
2. Structure available
3. Engineering feasible
4. NADPH dependent
NADP+NADPH
monooxygenases
Pazmiño et al. (2008) Angew. Chemie
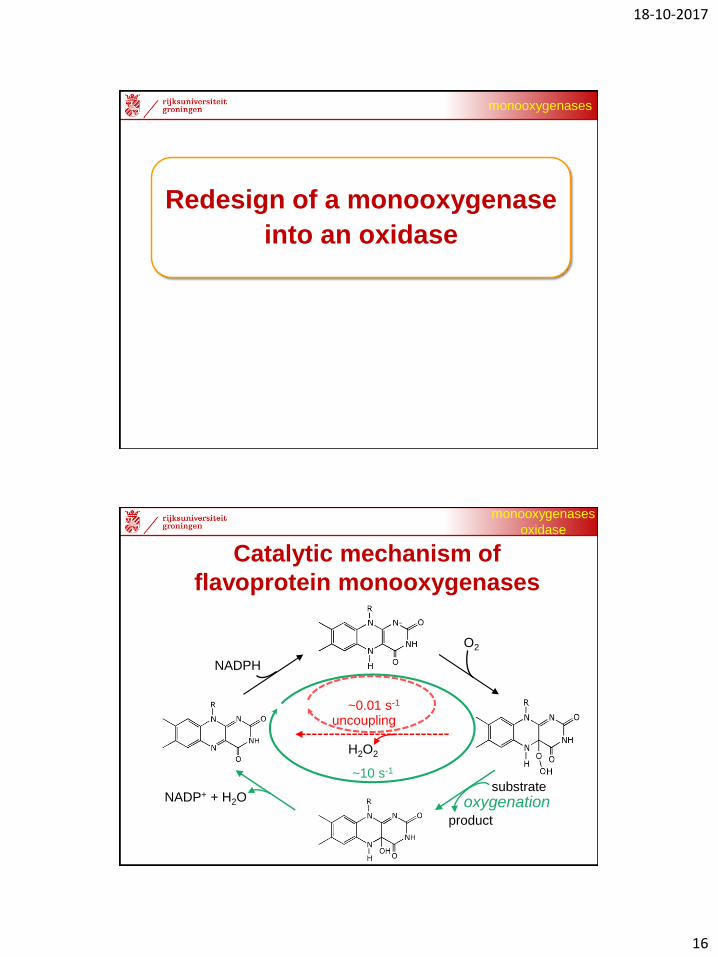
Self-sufficient BVMOs: PTDH-BVMO fusions
BVMO produced as fusion protein
fused to phosphite dehydrogenase:
1. assures cofactor recycling
2. boosts expression
3. facilitates assay
NADP+NADPH
HPO32- PO4
2-
PTDH
monooxygenases
18-10-2017
16
Redesign of a monooxygenase
into an oxidase
monooxygenases
O2
substrate
product
oxygenation
H2O2
uncoupling
~10 s-1
~0.01 s-1
NADPH
NADP+ + H2O
Catalytic mechanism of
flavoprotein monooxygenases
monooxygenases
oxidase
18-10-2017
17
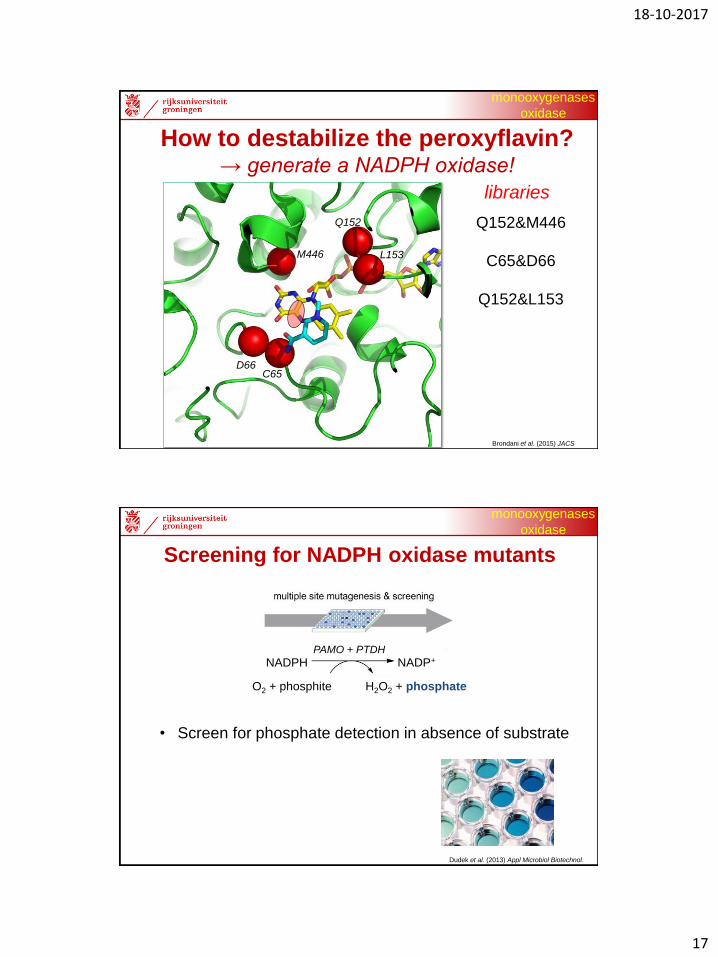
How to destabilize the peroxyflavin?→ generate a NADPH oxidase!
M446
D66C65
Q152
L153
Q152&M446
C65&D66
Q152&L153
libraries
Brondani et al. (2015) JACS
monooxygenases
oxidase
Screening for NADPH oxidase mutants
Dudek et al. (2013) Appl Microbiol Biotechnol.
• Screen for phosphate detection in absence of substrate
O2 + phosphite H2O2 + phosphate
PAMO + PTDH
NADPH NADP+
monooxygenases
oxidase
18-10-2017
18
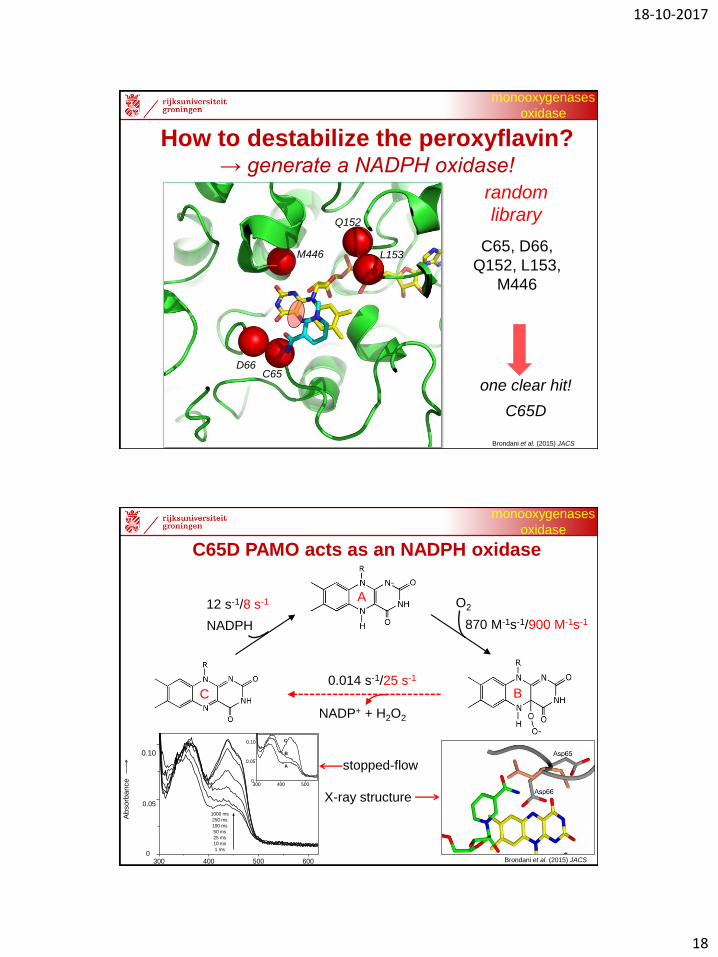
How to destabilize the peroxyflavin?→ generate a NADPH oxidase!
M446
D66C65
Q152
L153C65, D66,
Q152, L153,
M446
random
library
Brondani et al. (2015) JACS
one clear hit!
C65D
monooxygenases
oxidase
O2
NADPH
12 s-1/8 s-1
870 M-1s-1/900 M-1s-1
0.014 s-1/25 s-1
300 400 500
C
B
A
0
0.05
0.10
0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0,1707 0,1992 0,2337 0,2499 0,2585 0,2683 0,2759 0,2783 0,2673 0,2371 0,195 0,1499 0,1122 0,08504 0,06668 0,05791 0,05395 0,05377 0,05457 0,05709 0,05894 0,06186 0,0649 0,06758 0,07027 0,07334 0,07641 0,0797 0,08479 0,08656 0,08731 0,09024 0,09237 0,09462 0,09629 0,09798 0,09962 0,1002 0,1012 0,1014 0,1013 0,101 0,10070,09987 0,0989 0,09881 0,09543 0,09267 0,08955 0,0859 0,08219 0,07857 0,07508 0,0723 0,06965 0,06833 0,06763 0,06824 0,0674 0,06857 0,07022 0,0723 0,07331 0,07701 0,07913 0,08108 0,08305 0,08511 0,08703 0,08915 0,09166 0,09432 0,09693 0,09948 0,1016 0,1034 0,1037 0,1034 0,1022 0,1004 0,09826 0,09595 0,09504 0,09305 0,09167 0,09058 0,08955 0,08832 0,08681 0,08384 0,08061 0,07665 0,07111 0,065 0,0583 0,05092 0,04415 0,03839 0,03319 0,02885 0,02484 0,02157 0,01948 0,01726 0,0157 0,01436 0,01343 0,01286 0,01238 0,01165 0,01124 0,01112 0,01076 0,01052 0,0104 0,0103 0,009932 0,01034 0,009797 0,009924 0,009539 0,009067 0,009629 0,009535 0,009507 0,009338 0,009366 0,009258 0,009386 0,009071 0,008959 0,009001 0,008983 0,008922 0,008943 0,008815 0,008809 0,00879 0,008201 0,00871 0,008674 0,008581 0,008626 0,008482 0,008468 0,008356 0,008249 0,00821 0,007589 0,00817 0,008168 0,00812 0,00805 0,008052 0,008045 0,007974 0,007953 0,007926 0,007777 0,007741 0,007871 0,007815 0,007762 0,007188 0,007695 0,007656 0,007644 0,007566 0,007552 0,008283 0,007455 0,007907 0,007371 0,0073870,007201 0,007247 0,007198 0,007253 0,007043 0,007941 0,007006 0,006899 0,00816 0,006965 0,006962 0,007049 0,007396 0,007527 0,0069880,006753 0,005635 0,006573 0,006803 0,006619 0,006724 0,006727 0,006678 0,006706 0,006592 0,007664 0,006608 0,005515 0,006576 0,006541 0,006363 0,006179 0,005133 0,006413 0,007522 0,006422 0,006101 0,006509 0,006215 0,00631 0,00779 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0,1707 0,1992 0,2337 0,2499 0,2585 0,2683 0,2759 0,2783 0,2673 0,2371 0,195 0,1499 0,1122 0,08504 0,06668 0,05791 0,05395 0,05377 0,05457 0,05709 0,05894 0,06186 0,0649 0,06758 0,07027 0,07334 0,07641 0,0797 0,08479 0,08656 0,08731 0,09024 0,09237 0,09462 0,09629 0,09798 0,09962 0,1002 0,1012 0,1014 0,1013 0,101 0,10070,09987 0,0989 0,09881 0,09543 0,09267 0,08955 0,0859 0,08219 0,07857 0,07508 0,0723 0,06965 0,06833 0,06763 0,06824 0,0674 0,06857 0,07022 0,0723 0,07331 0,07701 0,07913 0,08108 0,08305 0,08511 0,08703 0,08915 0,09166 0,09432 0,09693 0,09948 0,1016 0,1034 0,1037 0,1034 0,1022 0,1004 0,09826 0,09595 0,09504 0,09305 0,09167 0,09058 0,08955 0,08832 0,08681 0,08384 0,08061 0,07665 0,07111 0,065 0,0583 0,05092 0,04415 0,03839 0,03319 0,02885 0,02484 0,02157 0,01948 0,01726 0,0157 0,01436 0,01343 0,01286 0,01238 0,01165 0,01124 0,01112 0,01076 0,01052 0,0104 0,0103 0,009932 0,01034 0,009797 0,009924 0,009539 0,009067 0,009629 0,009535 0,009507 0,009338 0,009366 0,009258 0,009386 0,009071 0,008959 0,009001 0,008983 0,008922 0,008943 0,008815 0,008809 0,00879 0,008201 0,00871 0,008674 0,008581 0,008626 0,008482 0,008468 0,008356 0,008249 0,00821 0,007589 0,00817 0,008168 0,00812 0,00805 0,008052 0,008045 0,007974 0,007953 0,007926 0,007777 0,007741 0,007871 0,007815 0,007762 0,007188 0,007695 0,007656 0,007644 0,007566 0,007552 0,008283 0,007455 0,007907 0,007371 0,0073870,007201 0,007247 0,007198 0,007253 0,007043 0,007941 0,007006 0,006899 0,00816 0,006965 0,006962 0,007049 0,007396 0,007527 0,0069880,006753 0,005635 0,006573 0,006803 0,006619 0,006724 0,006727 0,006678 0,006706 0,006592 0,007664 0,006608 0,005515 0,006576 0,006541 0,006363 0,006179 0,005133 0,006413 0,007522 0,006422 0,006101 0,006509 0,006215 0,00631 0,00779
300 400 500 600
1000 ms
250 ms
100 ms
50 ms
25 ms
10 ms
1 ms0
0.05
0.10
Ab
so
rba
nce
Wavelength (nm)
Brondani et al. (2015) JACS
C65D PAMO acts as an NADPH oxidase
A
BC
stopped-flow
X-ray structure
NADP+ + H2O2
monooxygenases
oxidase

18-10-2017
19
Brondani et al. (2015) JACS
C65D PAMO can be used for cofactor regeneration
Substrate Product ADH Temp. (0C)
Time (h)
Conversion (%)
T. brockii 40 24 100a
Evo-1.1.270 30 24 100b
Evo-1.1.270 30 10 50
(ee > 99%)b
Substrate Product ADH Temp. (0C)
Time (h)
Conversion (%)
T. brockii 40 24 100a
Evo-1.1.270 30 24 100b
Evo-1.1.270 30 10 50
(ee > 99%)b
monooxygenases
oxidase
Switching a vitamin-binding protein
into an enzyme
cofactorengineering
18-10-2017
20
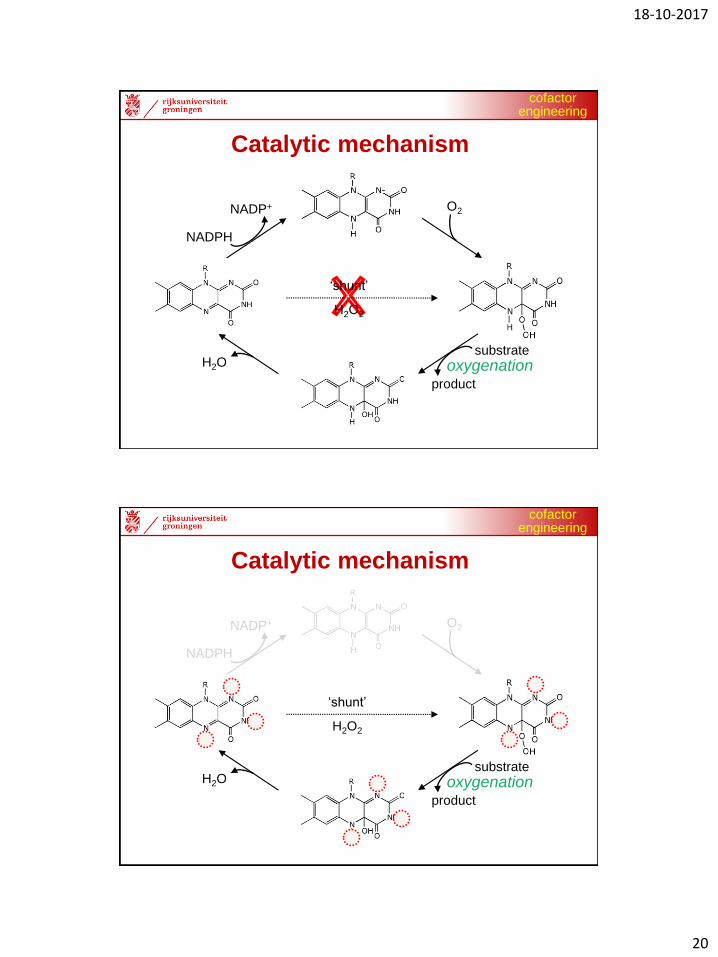
NADPH
NADP+ O2
substrate
product
oxygenationH2O
‘shunt’
H2O2
Catalytic mechanism
cofactorengineering
NADPH
NADP+ O2
substrate
product
oxygenationH2O
‘shunt’
H2O2
Catalytic mechanism
cofactorengineering

18-10-2017
21
• source: Gallus gallus
• protein: riboflavin-binding protein
• 29 kDa, monomeric
• 10 mg/egg!
• high affinity for flavins (nM range)
• apo preparation protocols
riboflavin-binding protein
Turning riboflavin-binding protein
into an enzyme
cofactorengineering
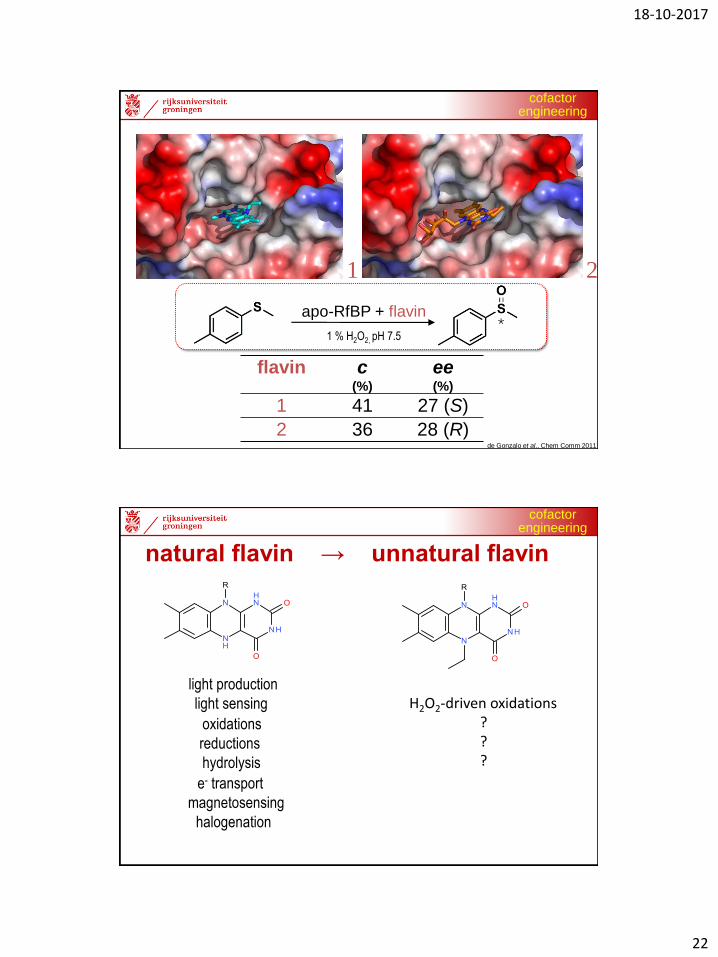
1 2
alkylated lumiflavin alkylated riboflavin
apo-RfBP + flavin
1 % H2O2, pH 7.5 *
de Gonzalo et al., Chem Comm 2011
flavin c(%)
ee(%)
1 41 27 (S)
2 36 28 (R)
Unnatural flavins
cofactorengineering
18-10-2017
22
1 2
*apo-RfBP + flavin
1 % H2O2, pH 7.5
flavin c(%)
ee(%)
1 41 27 (S)
2 36 28 (R)de Gonzalo et al., Chem Comm 2011
cofactorengineering
natural flavin → unnatural flavin
oxidations
reductions
light sensing
e- transport
hydrolysis
halogenation
light production
magnetosensing
H2O2-driven oxidations???
cofactorengineering

18-10-2017
23
EncM
enterocin
Teufel et al. (2013) Nature 503:552-556 Payne et al. (2015) Nature 522:497-501White et al. (2015) Nature 522:502-506
N
N
O
N H
NH
O
R
new natural flavins!
(de)carboxylationsFavorskii rearrangement
cofactorengineering
Conclusions
Flavin-containing enzymes: versatile biocatalysts
to be continued…
• Flavoenzymes are highly tuneable
Insight into enzyme mechanism can be extremely helpful
to introduce new activities:
dehydrogenase → oxidase (single mutation!)
monooxygenase → oxidase (single mutation!)
monooxygenase-dehydrogenase fusion
flavin cofactors can be engineered
flavoenzymes

18-10-2017
24
Marco FraaijeMolecular Enzymology
University of Groningen
The Netherlands
Andrea Mattevi
University of Pavia
Willem van Berkel
Wageningen University
Riccardo Baron
UC San Diego
Acknowledgements
funding collaborators materials
Fraaije group - September 2016www.dinosaurusmuseum.nl
www.gecco-biotech.com


















