
DISCOVERY AND CHARACTERIZATION OF MICROBIAL ESTERASES FOR ... · DISCOVERY AND CHARACTERIZATION OF...
108
DISCOVERY AND CHARACTERIZATION OF MICROBIAL ESTERASES FOR FIBER MODIFICATION by Lijun Wang A thesis submitted in conformity with the requirements for the degree of Masters of Applied Science Department of Chemical Engineering and Applied Chemistry University of Toronto © Copyright by Lijun Wang 2009
Transcript of DISCOVERY AND CHARACTERIZATION OF MICROBIAL ESTERASES FOR ... · DISCOVERY AND CHARACTERIZATION OF...

DISCOVERY AND CHARACTERIZATION OF MICROBIAL ESTERASES FOR FIBER MODIFICATION
by
Lijun Wang
A thesis submitted in conformity with the requirements for the degree of Masters of Applied Science
Department of Chemical Engineering and Applied Chemistry University of Toronto
© Copyright by Lijun Wang 2009

ii
Discovery and Characterization of Microbial Esterases for Fiber
Modification
Lijun Wang
Masters of Applied Science
Department of Chemical Engineering and Applied Chemistry University of Toronto
2009
Abstract
Carboxyl esterases, particularly arylesterases, were predicted from 16 microbial genomes, and
then expressed in E. coli. Of the more than 175 cloned genes, 86 were expressed in soluble
form. These were screened for activity using a range of both commercial and natural substrates.
Forty-eight proteins were active on pNP-acetate at pH 8 whereas 38 proteins did not exhibit any
activity towards any substrates. Among the 48 active proteins, 20 proteins showed arylesterase
activity. To date, 8 bacterial esterases and 2 archaeal arylesterases were characterized in terms of
pH stability and optima, thermal inactivation, solvent stability, and kinetics. To our knowledge
there is only one other published report of arylesterases from archaea. The synthetic capability
of arylesterases can transform phenolic acids to value-added chemicals. Accordingly, this
project provides an arsenal of industrially significant activities that can extend the antioxidant
properties of lignin-derived molecules in a broader range of renewable products.

iii
Acknowledgments
I would like to thank my supervisor Prof. Emma Master for her guidance and encouragement. I
would like to thank the following people from SPiT lab (University of Toronto) that helped me
with gene selection, gene cloning, and protein purification and for providing guidance on
experimental techniques: Valentina Mavisakalyan, Alexander Yakunin, Greg Brown, and
Michael Proudfoot. I would also like to thank the following people that helped me with
experimental techniques, solving problems, allowed me access to their equipment, and for
providing guidance and encouragement throughout my project: Alex Tsai, Jacqueline McDonald,
Sonam Mahajan, Elizabeth Srokowski, Angelika Duffy, and everyone else in BioZone. Further,
I would like to thank Ping Hay Lam for his hard work as summer research student and his
contribution to this project. Finally, I would like to thank my family and friends for their
support and encouragement over the course of the project.

iv
Table of Contents
Abstract ........................................................................................................................................... ii
Acknowledgments.......................................................................................................................... iii
Table of Contents........................................................................................................................... iv
Abbreviations................................................................................................................................ vii
List of Tables ............................................................................................................................... viii
List of Figures ................................................................................................................................ ix
List of Appendices .......................................................................................................................... x
1 Introduction ................................................................................................................................ 1
1.1 Plant Biomass: Opportunities and Challenges for Renewable Materials ........................... 1
1.2 Thesis Objectives ................................................................................................................ 2
2 Literature Survey........................................................................................................................ 3
2.1 Lignocellulose Structure and Composition......................................................................... 3
2.2 Lignocellulose-active Enzymes .......................................................................................... 4
2.3 Carboxyl Esterases and Lipases.......................................................................................... 6
2.3.1 Distinguishing Lipases from Carboxyl Esterases ................................................... 6
2.3.2 Carboxyl Esterase and Lipase Structure ................................................................. 6
2.3.3 Proposed Mechanism of Esterases and Lipases...................................................... 8
2.3.4 Classification of Esterases and Lipases .................................................................. 8
2.3.5 Applications of Carboxyl Esterases and Lipases .................................................... 9
2.3.5.1 Applications in the Detergent Industry..................................................... 9
2.3.5.2 Applications in the Pharmaceutical Industry.......................................... 10
2.3.5.3 Applications in the Food Industry .......................................................... 10
2.3.5.4 Other Industrial Applications ................................................................. 10
2.3.6 Esterase and Lipase Catalyzed Transesterification............................................... 11

v
2.3.7 Potential of Carboxyl Esterases and Lipases for Lignocellulose Modification.... 12
3 Bacterial Genome Mining and Recombinant Expression of Putative Arylesterases ............... 15
3.1 Introduction....................................................................................................................... 15
3.2 Materials and Methods...................................................................................................... 16
3.2.1 Materials ............................................................................................................... 16
3.2.2 Sequence Selection of Putative Arylesterases ...................................................... 17
3.2.3 Gene Cloning ........................................................................................................ 17
3.2.4 Protein Purification ............................................................................................... 17
3.2.5 Primary Enzyme Assays Using p-Nitrophenyl (pNP) Substrates......................... 18
3.2.6 Primary Screen Using Natural Substrates............................................................. 19
3.2.7 General Properties of Arylesterases...................................................................... 19
3.2.7.1 Optimal pH and pH Stability .................................................................. 19
3.2.7.2 Thermal Inactivation .............................................................................. 19
3.2.8 Enzyme Kinetics ................................................................................................... 20
3.2.9 Construction of A Phylogenetic Tree ................................................................... 20
3.3 Results and Discussion ..................................................................................................... 20
3.3.1 Identification and High Throughput Production of Putative Arylesterases.......... 20
3.3.2 pH Optima of Purified Esterases........................................................................... 21
3.3.3 Thermal Inactivation of Purified Esterases........................................................... 25
3.3.4 Kinetic Analysis of Purified Esterases.................................................................. 25
3.3.5 Sequence Analyses for Improved Prediction of Arylesterases ............................. 25
3.4 Conclusions....................................................................................................................... 30
4 Solvent Stability of Bacterial Carboxyl Esterases ................................................................... 36
4.1 Introduction....................................................................................................................... 36
4.2 Materials and Methods...................................................................................................... 38
4.2.1 Materials ............................................................................................................... 38

vi
4.2.2 Effect of Detergents and Reducing-agents on Enzyme Stability.......................... 38
4.2.3 Effect of Organic Solvents on Enzyme Stability .................................................. 38
4.2.4 Effect of Ionic Liquids (ILs) on Enzyme Stability ............................................... 39
4.3 Results and Discussion ..................................................................................................... 40
4.3.1 Effects of Detergents, Chelators and Reducing-agents on Hydrolysis Activities 40
4.3.2 Effects of Organic Solvents on Hydrolysis Activities .......................................... 41
4.3.3 Effects of Ionic Liquids (ILs) on Hydrolysis Activities ....................................... 45
5 Isolation and Characterization of Two, Solvent-Tolerant, Thermophilic Archaeal Esterases. 49
5.1 Introduction: Extreme Environments: A Source of Unique Biocatalysts ......................... 49
5.2 Materials and Methods...................................................................................................... 50
5.3 Results and Discussion ..................................................................................................... 50
5.3.1 Esterase Activity with p-Nitrophenyl Substrates .................................................. 51
5.3.2 Effect of Additives and Organic Solvents on Esterase Stability........................... 57
5.3.3 Kinetic Properties.................................................................................................. 61
6 Conclusions and Recommendations ........................................................................................ 62
6.1 Summary ........................................................................................................................... 62
6.2 Recommendations............................................................................................................. 63
Bibliography ................................................................................................................................ 65
Appendices................................................................................................................................... 75

vii
Abbreviations
BLASTp
BTB
CAZymes
CBM
CE
DMSO
EDTA
GGL
GH
GlcA
GT
HEPES
HSL
IL
ITC
Lif
LiP
MeGlcA
MnP
NPS
PAF-AH
PCR
PL
pNP
SDS
SDS-PAGE
Basic Local Alignment Search Tool: protein-protein BLAST
Bromothymol blue
Carbohydrate-active enzymes
Carbohydrate Binding Module
Carbohydrate Esterase
Dimethyl Sulfoxide
Ethylenediaminetetraacetic Acid
Glycerol Glucose α-Lactose
Glycoside Hydrolase
α-D-glucosyluronic acid
Glycosyl Transferase
4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid
Hormone-Sensitive Lipases
Ionic Liquids
Isothermal Titration Calorimetry
Lipase-specific foldase
Lignin Peroxidase
4-O-methyl-α-D-glucosyluronic acid
Manganese Peroxidase
Ammonium Phosphate Sulfate
Platelet-Activating Factor Acetylhydrolase
Polymerase Chain Reaction
Polysaccharide Lyase
p-Nitrophenol
Sodium Dodecyl Sulfate
Sodium Dodecyl Sulfate Polyacrylamide Gel Electrophoresis

viii
List of Tables
Table 2.1.Differences between lipases and esterases...................................................................... 6
Table 2.2. Summary of Enzymes Involved in Polymer Synthesis and/or Modification............... 14
Table 3.1. Activities (µmol•min-1•mg protein-1) of enzymes at 0.5 mM concentration of each substrate. ....................................................................................................................................... 23
Table 3.2. General properties of 8 characterized proteins. ........................................................... 27
Table 3.3. Kinetic parameters of 8 characterized proteins............................................................ 28
Table 3.4. Kinetic parameters of the arylesterase EstA from Lactobacillus helveticus CNRZ32 and EstB from Lactobacillus casei LILA ..................................................................................... 28
Table 4.1. Effect of detergents, inhibitor, and disulfide reducing agent on enzyme stability. ..... 41
Table 4.2. Effect of various organic solvents on enzyme stability. .............................................. 43
Table 4.3. Effect of ionic liquids on enzyme stability. ................................................................. 48
Table 5.1. Specific activities of AF_est1 and AF_est2 on different substrates. ........................... 51
Table 5.2. Effects of various surfactants on AF_est1 and AF_est2 stability. ............................... 57
Table 5.3. Effects of various organic solvents on AF_est1 and AF_est2 stability. ...................... 58
Table 5.4. Effects of ionic liquids on AF_est1 and AF_est2 stability. ......................................... 60

ix
List of Figures
Fig. 2.1. Schematic representation of the α/β fold. ........................................................................ 7
Fig. 2.1. Crystal structure of a lipase from Candida rugosa (PDB: 1TRH) .................................. 7
Fig. 2.3. Schematic representation of esterification (a) and transesterification (b) reactions...... 11
Fig. 3.1. The distribution of enzyme activities that were detected using a variety of substrates……………………………………………………………………………..…………. 23
Fig. 3.2. Thermal inactivation of RP_est1 … .............................................................................. 29
Fig. 3.3. Kinetics of RP_est1.. ..................................................................................................... 29
Fig. 3.4. Amino acid sequence alignment of 8 characterized bacterial proteins. ........................ 31
Fig. 3.5. Amino acid sequence alignment of bacterial proteins that were active on phenyl acetate........................................................................................................................................................ 32
Fig. 3.6. Amino acid sequence alignment of proteins that were not active on any tested substrates....................................................................................................................................... 33
Fig. 3.7. A phylogenetic tree of 39 enzymes that showed activity on tested substrates. ............. 35
Fig. 4.1. Effect of DMSO on enzyme activity. ............................................................................ 46
Fig. 5.1. Substrate preference of AF_est1 and AF_est2 on pNP-esters....................................... 52
Fig. 5.2. Amino acid sequence alignment of AF_est1 and AF_est2 with other esterases from Archaeoglobus fulgidus. ............................................................................................................... 53
Fig. 5.3. Amino acid sequence alignment of AF_est1 and AF_est2 with an arylesterases from Sulfolobus solfataricus.................................................................................................................. 54
Fig. 5.4. pH profiles of AF_est1 (black) and AF_est2 (grey)...................................................... 55
Fig. 5.5. Effects of pH on AF_est1 (black) and AF_est2 (grey) stability.................................... 55
Fig. 5.6. Effect of thermal inactivation of AF_est1 and AF_est2................................................ 56

x
List of Appendices
Appendix A.1 Amino acid sequence alignment of proteins that were active on phenyl acetate. . 75
Appendix A.2 Amino acid sequence alignment of proteins that were not active on any substrates............................................................................................................... 78
Appendix A.3 Amino acid sequence alignment of AF_est1 and AF_est2 with 8 bacterial esterases that were characterized in Chapter 3. .................................................... 85
Appendix B.1 Summary of the 208 genes targeted for cloning, protein expression, purification, and biochemical characterization. .................................................... 87
Appendix B.2 The schematic representations of pNP-esters, phenyl acetate, and tributyrin. ...... 93
Appendix B.3 SDS-PAGE (15%) of the purified bacterial esterases that were characterized in this study. .............................................................................................................. 94
Appendix B.4 A standard curve of p-nitrophenol at pH 7. ........................................................... 95
Appendix B.5 Absorbance reading at 400 nm of the residual activity for PP_est3 after 5 h preincubation in organic solvents. Absorbance reading of blanks for organic solvents mixed in 50 mM posstassium phosphate buffer at pH 7......................... 96
Appendix B.6 SDS-PAGE (15%) of the purified archaeal esterases that were characterized in this study. .............................................................................................................. 98

1
1 Introduction
1.1 Plant Biomass: Opportunities and Challenges for Renewable Materials
It is now generally accepted that current consumption rates of easily extracted petroleum
resources are not sustainable given the finite nature of these reserves and detrimental
environmental impacts associated with its usage. This acknowledgement has led to a renewed
interest in developing petroleum-displacing sources of energy, chemicals and polymeric
materials.
The inherent polymeric structure and carbon-rich composition of lignocellulosic biomass means
it is an important resource for the production of renewable bioproducts [1-4]. In addition to
dedicated crops for energy and chemicals, lignocellulose can be obtained from a variety of
industrial wastes, including straw, hulls, stems, and stalks from agricultural crops, slash and
sawdust from forest industries, and municipal solid wastes (paper, cardboard, yard trash, wood
products) [5]. Although a large fraction of this biomass is currently disposed of by burning or
landfill, much of it could be converted into higher-value renewable products [6].
Thermochemical conversion and bioconversion are two strategies aimed at diversifying the range
of products generated from lignocellulose. In both cases, most research efforts have been aimed
at depolymerizing biomass to building blocks for biofuel production. However, it is anticipated
that the synthesis of high-value products from plant fiber is an important means to reducing
biofuel costs and stabilizing biofuel markets. The importance of high-value products from plant
biomass is perhaps particularly relevant to Northern climates where the biomass feedstocks are
slower to grow.
Cellulose, hemicellulose and lignin possess many active functional groups that are susceptible to
chemical reactions. Modifications on celluloses have been increasingly investigated over last
decades in order to confer new properties to fiber surfaces, such as water retention and surface
reactivity [2,3,7]. However, extensive chemical modification of cellulose can compromise the
integrity of cellulose microfibrils, such as in the case of super-adsorbing hydrogels [8].
Therefore, coating or lamination is used when retention of fiber strength is required [4]. These
coatings are used to improve the compatibility of cellulose fibrils with typical hydrophobic

2
matrix polymers [4]. While conventional coatings include synthetic resins [9], the common β-
(1,4)-linked backbone structure of cellulose and hemicellulose can be harnessed to create
biodegradable alternatives to synthetic resins. For example, xyloglucan endotransglycosylases
have been used to covalently link various functional groups to xyloglucan, which is then used to
coat cellulose fibrils [4, 10-12]. In this way, entirely “green” biocomposites can be synthesized
using both renewable compounds and catalysts. Similarly, lipases have been used to acetylate
cellulose, as well as catalyze regio-specific addition of aliphatic side-groups to xyloglucan
oligosaccharides [12]. Indeed, given their specificity and mild reaction requirements, enzymes
are ideal catalysts for plant fiber engineering since corresponding substrates are typically highly
functionalized, and resulting products are more likely to retain desired degree of polymerization
and crystallinity characteristics. Still, significant challenges to realizing the full potential of
lignocellulose fiber and its polymeric components include 1) extraction of intact biopolymers
from plant fibers; 2) specific, reproducible modification of biopolymer chemistry and properties,
and 3) the limited repertoire of industrially relevant enzymes to perform corresponding catalyses.
1.2 Thesis Objectives
The overall objective of my Master’s thesis is to increase the arsenal of industrially significant
enzymes for plant polymer engineering, taking advantage of microbial genome sequences and a
high-throughput protein production platform. More specifically, my objectives are to:
1. Discover novel carboxyl esterases (particularly arylesterases) from microorganisms with
publicly available genome sequences.
2. Recombinantly produce and characterize new esterases on the basis of substrate specificity,
optimal pH, pH stability, thermal inactivation, kinetics as well as solvent tolerance in a variety of
detergents, organic solvents, and ionic liquids.

3
2 Literature Survey
2.1 Lignocellulose Structure and Composition
Lignocellulose is the major structural component of plants and is comprised of the two most
abundant polymers on earth, cellulose and lignin [1]. In addition to cellulose and lignin,
lignocellulose contains different hemicelluloses, as well as varying amounts of pectin, protein
and waxes [2]. Cellulose is a linear polymer of β-(1,4)-D-glucose, where the repeating unit is
cellobiose. These glucan polymers adhere to each other through hydrogen bonding and
hydrophobic interactions to form crystalline microfibrils; microfibrils also associate to form
highly organized cellulose fibrils [3]. Hemicellulose represents approximately 20 to 35 % of the
lignocellulosic biomass, and has a branched structure consisting of a variety of sugars including
pentoses (xylose, arabinose), hexoses (mannose, glucose, galactose), and sugar acids [4]. This
class of molecules is also called crosslinking polysaccharides given their interaction with both
lignin and cellulose. Like cellulose, backbone sugars of hemicellulose are linked through β-
(1,4)-linked glycosidic bonds. Xylans are the most abundant hemicelluloses, which predominate
in secondary walls of hardwoods and the fibrous fraction of agricultural feedstocks. Xylan
consists of a homopolymeric backbone chain of β-(1,4)-D-xylopyranose units, and can be
branched by short α-D-glucosyluronic acid (GlcA), 4-O-methyl-α-D-glucosyluronic acid
(MeGlcA), α-L-arabinosyl, O-acetyl, feruloyl, or coumaroyl groups [3]. The frequency and
composition of these side groups depend on the source of xylan [5]. Mannan is the primary
hemicellulose in softwood fiber; the backbone structure of mannan consists of β-(1,4)-linked
mannose or a combination of glucose and mannose residues. The mannan backbone can be
substituted with side chains of α-1,6-linked galactose residues, resulting in four subfamilies:
linear mannan, glucomannan, galactomannan, and galactoglucomannan [6]. While xylan and
mannan are the major hemicelluloses in secondary cell walls of wood fiber, xyloglucan is the
primary hemicellulose in seeds and primary cell walls of plants [7]. Xyloglucan is composed of
a linear β-(1,4)-glucan backbone branched at C-6 with α-linked xylopyranosyl residues; most
xyloglucans contain XXXG or XXGG oligosaccharide repeats, where X and G refer to an
unbranched β-D-Glcp residue and an α-D-Xylp(1→6)-β-D-Glcp segment, respectively [7].

4
Lignin is a complex phenolic polymer that is composed of a combination of coniferyl alcohol,
sinapyl alcohol and/or para-coumaryl alcohol [2]. These monolignols are incorporated into
lignin in the form of p-hydroxyphenyl (H), guaiacyl (G), and syringyl (S), respectively [9].
Unlike cellulose and hemicellulose, the lignin structure is highly irregular. The relative
abundance of each monolignol depends on the plant species, cell type, as well as individual cell
wall layers; they are also influenced by developmental and environmental affects [9]. Generally,
hardwood lignins contain a mixture of G and S units and traces of H units, whereas softwood
lignins consist of mainly G units with low amounts of H units. In contrast, grass lignins show
comparable levels of G and S units and comparatively high amounts of H units [9].
Cellulose, hemicellulose and lignin are associated by a variety of covalent and non-covalent
linkages. The conventional model of lignocellulose describes hemicellulose as a crosslinking
polysaccharide that is hydrogen bound to cellulose and covalently linked to lignin [2,3]. While
hemicellulose is believed to impart flexibility to cellulose fibrils, lignin is believed to participate
in resistance to plant pathogens and facilitate water transport.
2.2 Lignocellulose-active Enzymes
Thermochemical and bioconversion are two approaches to processing biomass for the production
of sugars or the extraction of polymeric components from lignocellulose. Bioconversion
processes typically include an initial physical/chemical pretreatment of biomass followed by
biological conversion of resulting sugars and biopolymers to fuels, chemicals, and new materials.
In part because of lower temperatures and the specificity of enzyme-catalyzed reactions, key
advantages of bioconversion strategies are i) the potential to selectively extract lignocellulose
polymers and sugars, ii) low corrosive impact on process equipment; iii) minimal formation of
inhibitors to downstream fermentation processes, and iv) corresponding bioprocesses are often
renewable and have low energy requirements.
Carbohydrate-active enzymes (CAZymes) have been classified into over 100 glycoside
hydrolase (GH) families, 91 glycosyl transferase (GT) families, 16 carbohydrate esterase (CE)
families, 21 polysaccharide lyase (PL) families and 55 carbohydrate binding modules (CBM)
(http://www.cazy.org/). Renewed interest in applying these enzymes for the production of
fermentable sugars from biomass reflects widespread concern over greenhouse gases emitted

5
from transportation fuels, and efforts to increase societal and economic security through the
production of domestic biofuels [1]. Specific areas of research towards bioconversion of
lignocellulose to fermentable sugars includes reducing the cost of enzyme production,
discovering and engineering hydrolytic enzymes with increased specific activity on cellulosic
substrates, and development of enzyme suites that are tailored for the hydrolysis of specific
feedstocks [1]. Examples of CAZymes that are already used in industrial applications include
amylases, pectinases, and pectate lyases used for food processing, cellulases in laundry
detergents and by textile industries, and xylanases used for pulp bleaching [1,10]. Amylases and
cellulases are also used for biofuel production. Notably, these applications have harnessed the
depolymerization activity of CAZymes. Despite their industrial relevance, comparatively few
applications have been developed for CAZymes that have the potential to modify carbohydrates
through synthetic catalyses. An exception includes the development of a xyloglucan
endotransglycosylase to graft new chemistry to the surface of cellulose microfibrils [8,11].
Similarly, lipase enzymes have been used to acetylate cellulose, as well as catalyze the regio-
specific addition of aliphatic side-groups to xyloglucan oligosaccharides [11,12]. Further,
glycosyl transferases can synthesize novel oligosaccharides, some having significant clinical
relevance [13, 14]. However, these examples have yet to transfer to an industrial scale.
Lignin degradation is mainly catalyzed by extracellular, oxidative enzymes [15]. Lignin
peroxidases (LiPs), manganese peroxidases (MnPs), and laccases are the most studied enzymes
involved in ligninolysis. These enzymes require low molecular weight mediators to carry out
lignin degradation [16]. Other reductive enzymes such as cellobiose dehydrogenase, aryl alcohol
oxidases, and aryl alcohol dehydrogenases might also participate in ligninolysis [15]. Lignin that
is recovered during the conversion of biomass to fermentable sugars is typically burned to help
fuel the process. However, the development of higher-value products from lignin could improve
the economics of biofuel production from lignocellulose feedstocks. For instance, the biological
antioxidant properties of lignin could be further harnessed in functional foods, cosmetics, as well
as a broader range of lubricant oils [17]. To extend the utilization of lignin, it will be important
to adjust its water solubility characteristics. For instance, enzymatic esterification of lignin-
derived phenolic acids with aliphatic molecules can increase the miscibility of the product in
food with higher fat content. The synthetic potential of carboxyl esterases and lipases has been
demonstrated for this application [18].

6
2.3 Carboxyl Esterases and Lipases
2.3.1 Distinguishing Lipases from Carboxyl Esterases
Lipases and esterases are hydrolytic enzymes that cleave ester linkages through the addition of a
water molecule, producing a carboxylic acid and an alcohol. Esterases and lipases are lipolytic
enzymes but exhibit different substrate preferences (Table 2.1). While esterases catalyze the
hydrolysis of short to medium length aliphatic esters that are partially soluble in water, lipases
typically display maximal activity towards water insoluble long-chain esters. Interfacial
activation and deviant Michaelis-Menten kinetic curves are additional phenomena that are used
to distinguish lipases from esterases [19]. These phenomena have been correlated to the
presence of a flexible hydrophobic α-helix domain (lid or flap) in many lipases, which are
thought to expose the active site and direct substrate binding at the interface between water and
solvent phases [19] . Lipases also typically contain comparatively high numbers of non-polar
residues, Val, Leu and Ile [20]. It has been suggested that increased hydrophobic content may
improve lipase binding to hydrophobic substrate aggregates [20]. Finally, lipases have been
distinguished from esterases by having optimal activity at comparatively alkaline pH, and by
distinct surface electrostatic potential distributions at corresponding pH optima [21].
Table 2.1.Differences between lipases and esterases [19].
2.3.2 Carboxyl Esterase and Lipase Structure
Esterases and lipases acquire the α/β hydrolase fold, a structural conformation that is highly
conserved among other hydrolytic enzymes. The α/β hydrolase fold typically consists of 8 β-
strands connected by 6 α-helices (Fig. 2.1). An example of crystal structure for Candida rugosa
lipase is shown in Fig. 2.2. A conserved feature of α/β hydrolases is the nucleophilic elbow,

7
which displays a sharp γ turn with a nucleophilic serine residue positioned between a β-strand
and an α-helix [23]. Most of these enzymes also contain a catalytic triad consisting of a serine,
histidine and aspartate residue. Glutamate replaces aspartate in some lipases. In primary
sequences, the catalytic serine is usually part of the conserved pentapeptide Gly-X-Ser-X-Gly,
where X represents any amino acid. Exceptions include GDSL family esterases, where the
catalytic serine is presented in the conserved Gly-Asp-Ser-(Leu) motif found near the N-terminus
of the enzyme [24]. As mentioned above, the 3D structure of many lipases also reveals a lid
(flap) structure that covers the active site of the enzyme in the absence of substrate.
Fig. 2.1.Schematic representation of the α/β fold. Six α helices and 8 β strands are shown by cylinders and arrows, respectively. The catalytic triad is indicated by black dot. (A modified representation of Fig. 1 from [19])
Fig. 2.2. A crystal structure of a lipase from Candida rugosa (PDB: 1TRH). The structure was displayed by NewCartoon generated in Visual Molecular Dynamics program (Version 1.8.5).

8
2.3.3 Proposed Mechanism of Esterases and Lipases
Esterases and lipases share the same catalytic mechanism for ester hydrolysis [19]. The
mechanism involves four steps: 1) formation of a tetrahedral intermediate that is stabilized by the
catalytic His and Asp residues once the substrate is bound to the active serine (in lipases, this
step also involves movement of the lid structure); 2) the formation of an acyl-enzyme complex
after the release of the alcohol; 3) nucleophilic attack of the acyl-enzyme complex by water,
forming a second tetrahedral intermediate, and 4) the release of the acid product and free
enzyme. The reverse reaction (transesterification) is achieved by substituting water for an
alcohol, thereby forming new ester compounds.
2.3.4 Classification of Esterases and Lipases
Bacterial esterases and lipases were classified by Aprigny and Jaeger (1999) based mainly on a
comparison of primary amino acid sequences and some biological properties including how the
enzyme is secreted, requirements for lipase-specific foldases, and potential relationship to other
enzyme families [24]. A total of 53 sequences were compared and classified into 8 different
families. Family I (block L in ESTHER database, http://bioweb.ensam.inra.fr/ESTHER)
contains true lipases and is further divided into 6 subfamilies. Subfamilies I.1 and I.2 appear to
require a lipase-specific foldase (Lif) for proper folding and contain two conserved aspartic acid
residues involved in the Ca2+ binding and two cysteine residues that form a disulphide bridge
[24]. However, Subfamily I.1 and I.2 differ in size; subfamily I.1 enzymes range from 30 to 32
kDa, while subfamily I.2 are roughly 33 kDa and contain an insertion sequence that forms an
anti-parallel double β-strand at the protein surface. By contrast, subfamily I.3 are typically 50 to
60 kDa, and do not contain cysteine residues; subfamily I.4 are approximately 20 kDa, and
subfamily I.5 are approximately 45 kDa and usually display maximal activity between pH 7 to 9
and 30 to 50 °C [24,25]. The conserved pentapeptide sequence in Bacillus lipases from
subfamily I.4 and I.5 is characterized by Ala-X-Ser-X-Gly, whereby the first glycine residue is
replaced by an alanine residue. Finally, subfamily I.6 is usually secreted as a precursor protein
and cleaved extracellularly by a specific protease with processing beginning at the N-terminus.
Lipases and esterases with the conserved sequence Gly-Asp-Ser-(Leu) motif are classified in
family II. Family III (block L in ESTHER database) includes lipases that possess the conserved

9
catalytic triad and show 20 % identity with the human PAF-AH (platelet-activating factor
acetylhydrolase). By contrast, family IV (block H in ESTHER database) includes hormone-
sensitive lipases (HSL) that show significant sequence similarity to the mammalian HSL. The
remaining families also contain the conserved catalytic triad but are distinguished by sequence
similarity to other functional enzymes, including epoxide hydrolases, dehalogenases and
haloperoxidase for family V (block X in ESTHER database), eukaryotic lysophospholipases for
family VI (block X in ESTHER database), eukaryotic acetylcholine esterases and intestine/liver
carboxylesterases for family VII (block C in ESTHER database), and class C β-lactamases for
family VIII.
2.3.5 Applications of Carboxyl Esterases and Lipases
Esterases and lipases comprise a large group of hydrolytic enzymes, which function in both
hydrolysis and synthesis of ester bonds. They can be utilized in pharmaceutical synthesis, food
processing, detergent treatment, as well as kinetic resolution [26]. The interest in these enzymes
resides in the fact that they do not require any cofactors, are easy to acquire, show high regio-
and stereospecificity, and are usually stable and active in organic solvents [19].
2.3.5.1 Applications in the Detergent Industry
Owing to their ability to hydrolyze fats, esterases and lipases are the major enzyme additives in
industrial laundry and household detergents. For instance, the first commercial lipase,
LipolaseTM was introduced by Novo Nordisk in 1994, which was isolated from the fungus
Thermomyces lanuginosus and recombinantly expressed in Aspergillus oryzae [27]. In addition,
Genencor International introduced two bacterial lipases used as detergent additives in 1995:
LumafastTM from Pseudomonas mendocina and LipomaxTM from Pseudomonas alcaligenes [27].
Favorable enzymes properties for this application include low substrate specificity and high
tolerance to pH shifts, temperature and solvents. All these requirements can be resolved either
by screening naturally stable enzymes or enhancing enzyme properties through protein
engineering [28].

10
2.3.5.2 Applications in the Pharmaceutical Industry
Esterases and lipases catalyze a wide variety of chemo-, regio-, and stereo-specific
transformations, thereby providing great opportunities in organic syntheses. One example
includes synthesis of optically pure (S)- and/or (R)-ketoprofen [2-(3-benzoylphenyl) propionic
acid] by Trichosporon brassicae esterase. This product is used in the reduction of inflammation
and relief of pain caused by arthritis, sunburn, menstruation, and fever [29]. Another anti-
inflammatory drug, naproxen, is synthesized by carboxyl esterase NP from Bacillus subtilis [30].
(S)-naproxen generated in this process is 99 % enantiomerically pure, and the reaction yield is in
excess of 95 %. Other examples include stereospecific conversions in the production of
pharmaceutical intermediates such as taxol synthesis, throumboxane-A2-antagonist,
acetylcholine esterase inhibitors, anti-cholesterol drugs, anti-infective drugs, Ca2+ channel
blocker drugs, K channel blocking drugs, anti-arrhythmic agents and antiviral agents [31].
2.3.5.3 Applications in the Food Industry
The physical properties and nutritional value of triglycerides is greatly influenced by the
position, chain length and degree of unsaturation of the fatty acids [27]. Since fats and oils are
important constituents of foods, higher value fats can be synthesized using esterases or lipases in
a relatively inexpensive way. For instance, these enzymes can alter the position of fatty acid
chains in the glyceride substrate and replace one or more of fatty acids with more desirable ones.
A typical example in the food industry includes upgrading palm oil to achieve properties similar
to cocoa butter used in the production of chocolate [32]. Here, lipases from Rhizomucor miehei
have been used to substitute undesired palmitic acids with desired stearic acids. Other examples
include lipase-catalyzed removal of fat from meats, and extraction of polyunsaturated fatty acids
from plants and animals to be used as neutraceuticals.
2.3.5.4 Other Industrial Applications
In addition to the above-mentioned applications, esterases and lipases are employed in many
other industries, including perfume industries, pulp and paper processing, and degradation of
synthetic materials [31, 32]. A sub-group of esterases, in particular the arylesterases and feruloyl
esterases, are interesting candidates for the production of flavors and anti-oxidant additives from
lignin-derived phenolic acids [31]. For example, feruloyl esterases along with xylanases can

11
release ferulic acid from xylan, which can be converted to vanillin [19]. Feruloyl esterases
belong to carbohydrate esterase family 1, and have been isolated from many fiber-active
microorganisms, including Aspergillus niger, and Fusarium oxysporum [31]. In addition,
cholesterol esterases and polyurethenase have been used to degrade synthetic pollutants
including plastics, polyurethane, and polyesters. These enzymes were successfully isolated from
Pseudomonas chlororophis and Pseudomonas aeruginosa [33, 34].
2.3.6 Esterase and Lipase Catalyzed Transesterification
Esterases and lipases have demonstrated ability in organic chemical synthesis, catalyzing a wide
range of chemo-, regio-, and stereo-selective transformations. Corresponding (trans-)
esterification reactions are favored in the absence of water, whereby an ester bond is synthesized
between an alcohol and a carboxylic acid (or carboxylic ester) by synthesizing the acid or
exchanging the organic group (R3) of the ester with the organic group (R2) of the alcohol (Fig.
2.3). For instance, the commercial lipase, PS “Amano” SD, originated from Burkholderia
cepacia, catalyzes a variety of transesterification reactions and successfully resolves many
racemic mixtures [35].
Fig..2.3. Schematic representation of esterification (a) and transesterification (b) reactions. In an esterification, an ester is formed between a carboxylic acid and an alcohol by releasing a water molecule; in a transesterification, a new ester is form by exchanging the organic group (R3) of an ester with the organic group (R2) of the alcohol.
As mentioned, water content determines the reaction equilibrium (ie., synthesis vs. hydrolysis),
the distribution of products in the reaction vessel, as well as the thermo-inactivation of
biocatalysts. Researchers have demonstrated that enzymes show higher stability at elevated
temperatures in low water environments compared to aqueous solutions [36]. Organic solvents

12
are thus used as reaction media because they not only favor (trans-)esterification, but also
increase the solubility of organic substrates, thereby resulting in higher reaction rates.
Accordingly, in transesterification reactions, the only water present in the system is the water
bound to the enzyme itself; this water is essential to maintain the structural integrity of the
enzyme. A general agreement previously stated by Klibanov et al. is that enzymes can function
in predominantly nonaqueous environment or low water environment as long as the essential
water layer around them is not stripped off [36].
Enzyme activities can be correlated to the hydrophobicity of an organic solvent [36]. Log Poct is
defined as the logarithm of the partition coefficient of a solvent with respect to the octanol/water
two-phase system, and shows good correlation with enzyme activity [36]. Generally, organic
solvents with log P ≤ 2 are not good choices for reaction media because polar solvents may strip
off the essential water layer around the biocatalyst. Rather, organic solvents with log P ≥ 4 are
most suitable [36]. The effect of organic solvents with log P between 2 and 4 depends on the
nature of the biocatalyst. An exception includes the pancreatic lipase, which functions in all
tested organic solvents with log P ranging from -1.3 to 13.7. Nevertheless, this enzyme still
performs better in hydrophobic organic solvents compared to hydrophilic organic solvents.
Lanne et al. (1987) explain that the outer part of the pancreatic lipase is more hydrophobic than
other lipases, thereby binding its essential water more tightly [37].
2.3.7 Potential of Carboxyl Esterases and Lipases for Lignocellulose Modification
Canada has an abundance of underutilized biomass that can be harnessed for the production of
sustainable energy and materials. Our biomass resource provides an opportunity to reduce our
demand for petroleum feedstocks while satisfying the need to generate renewable and
environmental friendly products. Moreover, valorization of natural biopolymers using enzymes
represents a “green” process technology, ensuring that the product as well as its synthesis, are
environmentally sustainable.
To date, lipases, esterases, proteases, nitrilases and glycosidases have been developed for the
specific non-destructive functionalization of polymer surfaces (Table 2.2) [38]. Owing to their
broad specificity, high solvent-tolerance, and ability to catalyze transesterification reactions,
esterases and lipases display particular benefits for the production of value-added lignocellulosic

13
polymers. While enzymatic transesterification of sugars and short oligosaccharides has already
been demonstrated [39], similar examples of directly modifying long fibers rather than short
oligosaccharides are lacking. Similarly, there are few examples where enzymes have been used
to adjust the solubility characteristics of lignin-derived phenolics to harness their valuable
antioxidant properties in a broader range of products. An exception is the use of feruloyl
esterases to catalyze the esterification of phenolic acids with n-butanol and glycerol, for the
production of antioxidant nutritional supplements [20,40].
Given the current interest in conversion of lignocellulose-derived sugars to biofuel, arylesterases
that modify lignin and lignin-derived phenolic acids are particularly interesting targets for the
production of high-value co-products from biomass [41]. To advance enzyme-catalyzed
transesterification of lignin for the production of lignin derivatives with altered miscibility, it is
necessary to find arylesterases with new substrate specificities and high performance in different
industrial conditions. The following chapters will describe efforts to isolate and characterize
new arylesterases from different microorganisms. These data will be used to assess the potential
of each enzyme for synthetic modification of lignin.

14
Table 2.2. Summary of Enzymes Involved in Polymer Synthesis and/or Modification [38].

15
3 Bacterial Genome Mining and Recombinant Expression of Putative Arylesterases
3.1 Introduction
Lipases (EC 3.1.1.3) and esterases (EC 3.1.1.1) similarly hydrolyze carboxyl ester bonds and
generally exhibit broad substrate specificity. Under non-aqueous conditions, the reverse reaction
can be catalyzed, whereby esterification of hydroxyl groups on sugars, phenolics and alcohols
has been demonstrated [1-3]. Historically, lipases and esterases were distinguished by
preference for medium to long-chain triacylglycerols and short-chain fatty acyl esters,
respectively. Structural analyses of lipases also revealed the presence of a peptide loop that was
correlated to transformation of water-insoluble substrates and activation at lipid-water interfaces.
However, exceptions to these generalizations have been observed, leading some to suggest that
lipases should constitute any carboxyl ester hydrolase that has the capacity to hydrolyze long-
chain triacylglycerols.
The regioselectivity and stereospecificity of lipase and esterase catalyzed transesterification
reactions offer advantages over chemical syntheses, which often demonstrate poor selectivity and
formation of undesirable side products [4]. While several examples of enzyme-catalyzed
transesterification of sugars and fatty acids have been reported, there are few examples of
enzymatic esterification of phenolic compounds. Exceptions include the production of
antioxidant nutritional supplements through feruloyl esterase esterification of phenolic acids with
n-butanol and glycerol, respectively [5, 6]. Accordingly, there is considerable opportunity to
identify new arylesterases with ability to synthesize novel chemicals from lignin-derived
aromatic substrates. The development of value-added compounds from lignin is an important
means to reducing the cost biofuels from lignocellulose feedstocks, and creating new products
for forest and agricultural sectors [7]. At the same time, the multitude of genome sequences that
are publicly available enables the identification of new enzymes with targeted activities.
To date, there are 3056 isolated genomes, of which 923 are completed, 1143 are on draft
assembly, and 990 are in progress (http://www.ncbi.nlm.nih.gov/genomes/static/gpstat.html).
Most of these genomes were annotated based on the sequence similarity to proteins that are
biochemically characterized [8]. While this approach is quick and inexpensive, 40 to 60 % of

16
sequences fail to be assigned a function, and many open reading frames are incorrectly annotated
[9,10]. In addition to sequence similarity-based approaches, comparative genomics [11-15],
two-hybrid assays [16-18], microarrays [19-22], and three-dimensional protein structures [23-
25], are being applied to improve functional predictions of gene products. Arguably, the most
definitive approach to assign a molecular function to a predicted open reading frame is to isolate
and biochemically characterize the corresponding protein [26]. While heterologous production
of proteins and the development of appropriate activity screens is still challenging, high
throughput platforms for protein synthesis and characterization have successfully identified
erroneous annotations and enzyme activities initially missed by conventional sequence
comparisons [26].
Given the industrial significance of arylesterases and the difficulty of distinguishing esterases
from lipases based on sequence comparisons alone [27], high throughput production and
characterization of predicted esterases and lipases was conducted. Microbial genome sequences
were selected based on the environmental niche of the corresponding microorganism, and the
availability of genomic DNA from the American Type Culture Collection (ATCC). Bacterial
genomes were initially analyzed to maximize successful protein expression in E.coli [28, 29].
3.2 Materials and Methods
3.2.1 Materials
All bacterial genomes were purchased from the ATCC. The reagents for the autoinduction
medium were purchased from Bioshop (Canada). The commercial pNP-esters (pNP-acetate,
pNP-propanioate, pNP-butyrate, pNP-capraote, pNP-caprate, pNP-laurate, pNP-palmitate, pNP-
benzoate, and paraoxon) and natural substrates (phenyl acetate, olive oil, and tributyrin) were
purchased from Sigma (Canada). The chemicals used in preparing universal buffers (phosphoric
acid, acetic acid and boric acid) were also purchased from Sigma (Canada). All chemicals are
pure at molecular level and sterilized before use. The pNP-ester stock solutions (10 mM) used
for primary screens were prepared in DMSO.

17
3.2.2 Sequence Selection of Putative Arylesterases
A biochemically characterized and structurally resolved bacterial arylesterase (Swiss-Prot
accession: P22862) was identified by searching the BRENDA and Swiss-Prot databases. The
corresponding protein sequence served as a parent sequence for BLASTp search of the following
15 translated bacterial genomes: Agrobacterium tumefaciens C58, Clostridium acetobutylicum
824D-S, Clostridium thermocellum 27405D, Escherichia coli K12, Nitrosomonas europaea,
Pseudomonas aeuginosa, Pseudomonas putida KT2440, Pseudomonas syringae, Rhodococcus
sp. RHA1, Rhodopseudomonas palustris CGA009, Ralstonia solanacearum 11696D-S,
Streptomyces avermitilis, Streptomyces coelicolor, Sinorhizobium meliloti, and Thermotoga
maritima. In order to capture novel esterase activities, an E-value of 0.01 was used to first filter
enzyme targets. Corresponding sequences were then aligned to P22862 to confirm the presence
of conserved catalytic residues and sequence motifs [27, 30].
3.2.3 Gene Cloning
Forward and reverse primers were designed and synthesized for each gene target; all primers
were flanked by BseRI restriction sites (www.idtdna.com). The genes were amplified from
bacterial genomes purchased from the ATCC; PCR was performed using a proof reading DNA
polymerase (Pfx polymerase) and the following PCR cycles: denaturation at 95 °C for 15 sec,
annealing at 53 °C for 30 sec, and elongation at 68 °C for 1 min. PCR products were purified
using the QIAquick PCR purification kit (Qiagen), digested using BseRI, and then ligated to
p15Tv-L (GenBank accession EF456736) using T4 DNA ligase (Fermentas). Ligation products
were transformed into Escherichia coli DH5α by electroporation; plasmids were isolated from
transformants using the Qiaprep spin miniprep kits (Qiagen) and sequenced at TCAG DNA
sequencing facility (Toronto).
3.2.4 Protein Purification
Briefly, E. coli transformants were cultured at 37 °C in autoinduction media [31] containing 100
µg/mL of ampicillin. Autoinduction medium contained 1 mM MgSO4, 1x metal mix, 1x GGL,
and 1x NPS. The 1000x trace metal mixture contained 0.1 M FeCl3・6H2O, 1 M CaCl2, 1 M
ZnSO4・7H2O, 1 M MnCl2・4H2O, 0.2 M CoCl2・6H2O, 0.1 M CuCl2・2H2O, 0.2 M NiCl2・6H2O,

18
0.1 M Na2MoO4・5H2O, and 0.1 M Na2SeO3・5H2O. The stock of 20x NPS contained 0.5 M
(NH4)2SO4, 1 M KH2PO4, and 1 M Na2HPO4, while 50x GGL contained 0.5 % glycerol, 0.05 %
glucose, and 0.2 % α-lactose.
Protein expression was induced by reducing the temperature to 20 °C. The cells were harvested
after overnight incubation, suspended in binding buffer (300 mM NaCl, 50 mM HEPES, pH 7.5,
5 % Glycerol, 5 mM imidazole), and lysed by sonication. Cell extracts were collected by
centrifugation and incubated with Ni resin (Qiagen) for 2 h. The resin was then washed with 200
mL of washing buffer (0.5 M NaCl, 50 mM HEPES, pH 7.5, 5 % Glycerol, 30 mM imidazole)
and eluted with approximately 10 mL elution buffer (0.5 M NaCl, 50 mM HEPES, pH 7.5, 5 %
Glycerol, 250 mM imidazole). Protein concentrations were measured using the Bradford assay
and their purities were evaluated by 15 % SDS-PAGE.
3.2.5 Primary Enzyme Assays Using p-Nitrophenyl (pNP) Substrates
The standard assay was performed in 96-well plate format at 37 °C; final reaction volumes were
200 µL and contained 1 µg of purified protein. Reactions were prepared using an automated
liquid handler (TECANTM Freedom EVO 100 MCA) and product formation was measured using
a microplate spectrophotometer (TECANTM Infinite M200). Product absorbency was initially
measured at a specified wavelength after 30 min at 37 °C. Reactions showing positive activity
were then assayed every minute for the first 6 min, followed by every 5 min up to 30 min. The
amount of enzyme added was also adjusted to ensure that initial reaction rates were measured,
(i.e., 0.05 µg for RP_est1, 0.25 µg for RP_est2, 0.5 µg for RP_est3, 0.5 µg for PP_est1, 0.5 µg
for PP_est2, 0.5 µg for PP_est3, 0.5 µg for PA_est1 and 1 µg for SAV_est1). Enzyme dilutions
were prepared in 50 mM potassium phosphate buffer (pH 7). Reactions were prepared by mixing
1 volume of the 10 mM pNP-ester stock solution with 9 volumes of 100 mM sodium phosphate
buffer (pH 7), containing 2.5 % Triton X-100, followed by immediate vortexing [32]. The final
pNP-ester concentration in the reaction was 0.5 mM. Liberation of pNP was measured at 400
nm and one enzymatic unit is defined as the amount of enzyme required to produce 1 µmol of
pNP per min. pNP was thus used to generate a standard curve (0.002 µmol to 0.1 µmol). All
purified proteins were assayed at pH 6, 7 and 8 on pNP-acetate, pNP-caprate, pNP-palmitate, and
pNP-benzoate.

19
3.2.6 Primary Screen Using Natural Substrates
Phenyl acetate, olive oil and tributyrin were used as natural substrates. The production of acids
was detected by Bromothymol blue (BTB) as described by Mertinez-Martines et al. [33] with
minor modifications. In brief, 30 mM of each substrate was prepared in 5 mM sodium phosphate
buffer (pH 7.3) containing 0.01 % of BTB. Initial reactions contained 1 µg of purified protein.
Reactions were prepared in 96-well plates, and were incubated for 30 min at 37 °C, followed by
absorbance measurement at 616 nm. The specific activity of enzymes on phenyl acetate was
calculated by generating a standard curve using acetic acid (1.75 µmol to 17.5 µmol).
3.2.7 General Properties of Arylesterases
3.2.7.1 Optimal pH and pH Stability
The effect of pH was investigated using the Britton and Robinson's universal buffer system (50
mM phosphoric acid, acetic acid and boric acid, pH adjusted to 4 to 11 with NaOH) [34]. pNP-
acetate was used to evaluate optimal pH. To examine enzyme stability at different pH values
(pH 6, pH 7, pH 8, pH 9, pH 9.5), residual enzyme activity was measured after 24 h of
incubation at 37 °C. Residual enzyme activity was measured using standard reaction conditions
and pNP-acetate as the substrate. In brief, 10x concentrated enzymes (i.e. 0.5 µg/100 µL for
RP_est1, 2.5 µg/100 µL for RP_est2, 5 µg/100 µL for RP_est3, 5 µg/100 µL for PP_est1, 5
µg/100 µL for PP_est2, 5 µg/100 µL for PP_est3, 5 µg/100 µL for PA_est1 and 10 µg/100 µL for
SAV_est1) were preincubated in each pH buffer for 24 h at 37 °C. The residual activity was
measured by transferring 10 µL aliquots back to standard conditions, as described in section
3.2.5.
3.2.7.2 Thermal Inactivation
Residual enzyme activity was measured after incubation at 22 °C, 30 °C, 37 °C, 50 °C, 55 °C,
and 70 °C for up to 5 h. Enzyme samples were incubated in 50 mM potassium phosphate buffer
(pH 7). Residual enzyme activity was measured using standard reaction conditions and pNP-
acetate as the substrate. In brief, 10X concentrated enzymes (i.e. 0.5 µg/100 µL for RP_est1, 2.5
µg/100 µL for RP_est2, 5 µg/100 µL for RP_est3, 5 µg/100 µL for PP_est1, 5 µg/100 µL for
PP_est2, 5 µg/100 µL for PP_est3, 5 µg/100 µL for PA_est1 and 10 µg/100 µL for SAV_est1)

20
were preincubated at each temperature for up to 5 h. The residual activity was measured by
transferring 10 µL aliquots back to standard conditions, as described in section 3.2.5.
3.2.8 Enzyme Kinetics
Kinetic parameters were obtained at 37 °C and using the optimal pH and preferred substrate for
each enzyme. Substrate concentrations ranged from 0.05 mM to 1 mM with 50 µM increments
(i.e., 20 different substrate concentrations). The amount of each enzyme used per each reaction
was 0.05 µg for RP_est1, 0.1 µg for RP_est2, 0.5 µg for RP_est3, 0.5 µg for PP_est1, 0.5 µg for
PP_est2, 0.25 µg for PP_est3, 0.5 µg for PA_est1 and 0.5 µg for SAV_est1. Initial rates of each
reaction at different substrate concentration were measured every minute for total of 10 min and
rates of product formation (V) were determined using the pNP standard curve. The kinetic curve
was generated by plotting velocity against substrate concentration using the software GraphPad
Prism5 (GraphPad Software, Inc.). In this way, non-liner regression was used to determine the
kinetic parameters Vmax and KM using the Michaelis-Menten model.
3.2.9 Construction of a Phylogenetic Tree
Protein sequences were aligned using MUSCLE, and 500 bootstrapped data sets were generated
using SEQBOOT. Protein distance matrices were obtained using the PHYLIP 3.69 Neighbor-
Joining algorithm PRODIST (100 iterations), and trees were drawn using NEIGHBOR and a
consensus tree was generated using CONSENSE. The tree was viewed by PhyloWidget
(http://www.phylowidget.org/).
3.3 Results and Discussion
3.3.1 Identification and High Throughput Production of Putative Arylesterases
The objective of this study is to identify and isolate new arylesterases, given their potential to
modify phenolic acids and thereby synthesize high-value compounds from lignin. The ability of
certain esterases to modify hydroxyl groups present on sugars also means these enzymes could
be used to create new compounds from polysaccharides, including biomass-derived
hemicellulose and cellulose.

21
A total of 202 gene targets from 15 bacteria were selected based on overall sequence similarity to
a biochemically characterized arylesterase, as well as presence of conserved catalytic residues
and sequence motifs [35]. While lipases and esterases are difficult to distinguish at the sequence
level, differences in catalytic properties can be used to differentiate these enzymes. For instance,
lipases can be distinguished from esterases by their ability to hydrolyze insoluble substrates and
by interfacial activation [35]. Previous analyses also suggest that lipases can be distinguished
from esterases by optimal activity of the former at alkaline pH, and by correlating the
electrostatic potential of enzyme surfaces to pH optima [30]. In addition, preference for aliphatic
and aromatic substrates can be used to distinguish lipases, carboxyl esterases and arylesterases.
Therefore, to identify arylesterases encoded by the gene targets, each gene was cloned for
heterologous expression in E.coli. A total of 180 genes were successfully cloned and 84 of them
were expressed as soluble proteins in E.coli. All 84 soluble proteins were purified and then
screened using colorimetric assays; 46 proteins showed activity on pNP-acetate at pH 8 while 38
proteins did not exhibit any activity towards the test substrates (Fig. 3.1). Phenyl acetate is a
general substrate to identify arylesterase and 18 proteins showed activity towards this substrate.
The specific activity of these 18 proteins on aliphatic pNP-esters and phenyl acetate was
compared to evaluate whether they demonstrate arylesterase-like activity or lipase-like activity
(Table 3.1). All 18 bacterial enzymes showed significantly higher specific activity towards
phenyl acetate than the aliphatic pNP-esters, and none of these enzymes hydrolyzed olive oil to
detectable levels, which is a common screen for lipase activity. Accordingly, these 18 bacterial
enzymes can be classified as arylesterases [36-38]. Six arylesterases that were also active on
medium-chain aliphatic substrates were selected for further biochemical characterization. Two
enzymes with activity towards pNP-acetate only were also characterized for comparison. All of
the eight enzymes preferentially hydrolyzed short- and/or medium- chain substrates, which is
typical of esterases [35]. Notably, none of the eight esterases exhibited paraoxonase-like
activity. The general properties of these 8 enzymes are summarized in Table 3.2.
3.3.2 pH Optima of Purified Esterases
Given the structural similarity of phenyl acetate and pNP-acetate, and challenges associated with
changing the pH of BTB assays, pNP-acetate was used to measure the optimal pH and pH
stability of selected enzymes. Previous analyses that correlate electrostatic potential distribution

22
of enzyme surfaces to pH optima suggest that lipases can be distinguished from esterases by
optimal activity of the former at alkaline pH [39]. Notably, the pH optima of esterases isolated
in this study were either neutral or alkaline (Table 3.2). According to biochemical data for
arylesterases listed in the BRENDA enzyme database (http://www.brenda-enzymes.info), neutral
or basic pH optima were reported for both eukaryotic arylesterases [40, 41] and bacterial
arylesterases [36, 37]. This suggests that arylesterases may represent a unique subfamily of
esterases that share properties of both esterases and lipases. All esterases except RP_est3 and
SAV_est1 were stable in both acidic to alkaline conditions. Differences in pH stabilities likely
result from proton activation and deactivation of titratable amino acid residues [36]. By
comparing pH optima to predicted pI values of each enzyme, it appeared that the enzymes
isolated in this study are most active when a net negative charge is attained. This result is
consistent with the electrostatic study published by Petersen et al. [39], which explains that while
products are deprotonated once released from the enzyme active site, lipases and esterases can
still carry a negative potential and retain an active conformation. At a high pH, the active site
may also carry a negative potential, which repulses the product. Supporting analyses such as
predicting the electrostatic potential distribution of enzymes will be performed when
corresponding x-ray structures are solved.

23
Fig. 3.1. The distribution of enzyme activities that were detected using a variety of substrates. All assays were performed using standard conditions, and were incubated for 30 min at 37 °C. The number of enzymes that were active on each substrate is shown in parentheses.
Table 3.1. Activities (µmol•min-1•mg protein-1) of enzymes using 0.5 mM of substrate. Microorganisms ID Substrates
pNP acetate
pNP caprate
pNP palmitate
Phenyl acetate
b2593 0.46±0.01 - - - Escherichia coli K12 b2799 0.16±0.02 - - - NE0279 0.14±0.02 - - - Nitrosomonas
europaea NE2298 0.37±0.02 - - 3.17±0.27
PA1469 0.1±0.01 - - - PA2934 0.38±0.01 - - 3.25±0.86 PA2949 0.66±0.02 1.04±0.4 0.2±0.01 2.33±0.43
Pseudomonas aerigonosa
PA3132 0.25±0.01 - - - PP0364 0.48±0.02 - - - PP3645 0.8±0.01 1.02±0.16 4.96±0.44
Pseudomonas putida KT2440
PP5253 0.91±0.02 - - 5.83±0.49

24
PSPTO3914 0.23±0.00 - - - PSPPH3482 0.24±0.02 - - 3.27±0.18
Pseudomonas syringae PSPPH4522 0.49±0.02 - - 5.22±0.19
RHA1_ro00320 0.2±0.02 - - 3.13±0.29 RHA1_ro01401 0.19±0.00 - - 3.47±0.47 RHA1_ro03350 0.24±0.02 - - - RHA1_ro02607 0.41±0.02 - - 4.22±0.06 RHA1_ro04513 0.19±0.02 - - - RHA1_ro03520 0.06±0.02 - - - RHA1_ro01338 0.38±0.01 - - - RHA1_ro03603 0.23±0.01 - - - RHA1_ro08044 1.29±0.01 - - - RHA1_ro08081 1.61±0.01 - - 6.27±0.12
Rhodococcus sp. RHA1
RHA1_ro10146 1.46±0.02 - - - RPA0348 2.24±0.10 1.74±0.05 - 7.36±0.49 RPA1163 0.31±0.01 - - - RPA2105 0.1±0.02 0.6±0.01 - - RPA3430 0.63±0.01 0.68±0.01 - 3.53±0.37
Rhodopseudomonas palustris CGA009
RPA4646 1.2±0.02 0.24±0.00 - 9.06±0.24 RSc1135 0.23±0.02 - - - Ralstonia
solanacearum 11696D-S
RSc2250 0.05±0.01 - - -
SAV512 0.45±0.00 - - - SAV875 0.1±0.02 - - - SAV2722 0.13±0.02 - - - SAV3810 0.15±0.02 - - -
Streptomyces avermitilis
SAV5173 0.13±0.02 - - - SCO1989 0.1±0.02 - - - SCO3439 0.35±0.01 - - 3.55±0.24 SCO3566 0.33±0.01 - - 3.09±1.07 SCO3690 0.83±0.02 - - 4.8±0.20 SCO6712 0.03±0.01 - - -
Streptomyces coelicolor
SCO7440 0.3±0.02 - - 3.02±0.41 Sinorhizobium meliloti RA0563 0.14±0.01 - - -
TM0111 0.18±0.01 - - - Thermotoga maritima TM1350 0.68±0.03 - - -

25
All assays were performed using standard conditions, and were incubated for 30 min at 37°C. n=3; errors indicate standard derivation.
3.3.3 Thermal Inactivation of Purified Esterases
RP_est1, PP_est2 and PP_est3 exhibited the highest half-lives at 50 °C (Table 3.2). While
RP_est2 was stable at 50 °C after 5 h incubation, the half-life of the enzyme at 55 °C was 4 h.
By contrast, 10 min at 50 °C reduced RP_est3 and PA_est1 activity by 80 %, while PP_est1 was
entirely inactivated. Thermal inactivation is likely due to protein denaturation at high
temperature.
3.3.4 Kinetic Analysis of Purified Esterases
Activity data for lipases with soluble substrates typically display either hyperbolic or deviant
Michaelis-Menten kinetics [42, 43]. Since kinetic data collected in this study obeyed the
Michaelis-Menten equation, kinetic data further support classifying the corresponding enzymes
as esterases (Table 3.3). In comparison to EstA from Lactobacillus helveticus CNRZ32 and
EstB from Lactobacillus casei LILA [36] (Table 3.4), most esterases characterized in this study
showed comparatively low catalytic efficiency with pNP-esters. Two explanations might
account for this: 1) enzyme kinetics were not performed at the optimal temperature of each
enzyme owing to the instability of pNP-esters at high temperatures and alkaline pH, and 2) the
structure of the aromatic moiety of substrates is important for binding by the arylesterases
characterized in this study. The latter is supported by comparing the specific activity of each
enzyme using pNP-acetate and phenyl acetate, which differ only by a nitro functional group at
position 4 of the phenyl moiety (Table 3.1). Thus, future experiments will include enzyme
kinetics at optimal temperature with phenyl acetate as well as other phenolic esters such as
phenyl butyrate and phenyl palmitate.
3.3.5 Sequence Analyses for Improved Prediction of Arylesterases
The amino acid sequences for the eight characterized esterases were aligned by using the
MUSCLE algorithm (http://www.ebi.ac.uk/Tools/muscle/). All proteins except for RP_est1
exhibit the catalytic triad Ser-Asp-His with Ser in the conserved Gly-X-Ser-X-Gly pentapeptide
sequence (Fig. 3.2). By contrast, the primary sequence of RP_est1 contained two analogous
pentapeptide sequences, one upstream and one downstream of the conserved pentapeptide

26
sequence. At this stage, it is unclear whether one or both of the Ser residues participate in the
catalytic mechanism of RP_est1; the significance of this unusual sequence feature will be
explored in future studies using site-directed mutagenesis and 3D structure determination. Most
of these eight characterized enzyme targets also contain the HG sequence, which constitutes a
conserved oxyanion hole that contributes to charge stabilization of reaction intermediates [27].
Instead, RP_est1, PP_est1 and SAV_est1 contain TA, PG, and HY sequences, respectively. The
structure of the parent protein used in this study (Swiss-Prot number: P22862) predicts two Phe
residues that contribute to the substrate-binding site of this enzyme [44]. Neither Phe residue
was identified in predicted sequences of the eight enzymes characterized in this study. Rather,
random amino acid residues are presented at these two positions. However, a conserved Phe
residue was identified near the C-terminal end of the proteins. Additional experiments, including
site-directed mutagenesis, are required to elucidate the significance of a conserved Phe residue at
this position.
In an attempt to improve future predictions of arylesterases based on primary protein sequence
information, enzymes that were biohemically characterized, that were active on phenyl acetate,
and that were not active on any of the test substrates were aligned (Fig. 3.4, 3.5 and 3.6).
Unfortunately, no distinguishing features were revealed. A phylogenetic tree of 39 active
enzymes was also constructed, to determine whether enzymes active on phenyl acetate clustered
together (Fig. 3.7). However, clustering according to enzyme activity was not observed. Future
bioinformatics analyses will attempt to compare these proteins based on predicted secondary and
tertiary structures. In this way, correlations between substrate preference and the hydrophobicity
of residue in the active site cleft might be supported, which could strengthen Hidden Markov
Models for arylesterases. The difficulty to identify new motifs that distinguish arylesterases,
esterases and lipases underline the importance of heterologous protein production and
characterization efforts for accurate annotation of open reading frames in publicly available
sequence databases.

27
Table 3.2. General properties of 8 characterized proteins. Properties RP_est1 RP_est2 RP_est3 PP_est1 PP_est2 PP_est3 PA_est1 SAV_est1
ID RPA0348 RPA4646 RPA3430 PP0364 PP5253 PP3645 PA2949 SAV512
Molecular weight 32.4 kDa 26.2 kDa 27.2 kDa 26.3 kDa 29.9 kDa 31.1 kDa 34.8 kDa 28.5 kDa
Predicted pI 5.28 5.31 5.55 5.03 4.92 5.09 6.09 5.41
Substrate preference on pNP substrates
pNP acetate to pNP caproate
pNP acetate pNP caproate
pNP acetate
pNP acetate
pNP caproate
pNP butyrate
pNP acetate
Optimal pH 7 9 9 8 to 9 9 8 to 9 7 to 9 8
pH stability1 4 to 9 5 to 10 8 to 10 5 to 11 6 to 8 4 to 10 6 to 8 7 to 10
Half life at 50 °C 5 h Stable after 5 h2
<5 min 5-10 min 4 h 5 h <5 min 2 h
1. Residual activity was greater than 80 % with respect to the highest activity. 2. The half-life at 55 °C was 4 h. n=6, standard derivation are within 20 % of the averaged value. The half life was interpreted from the corresponding thermal inactivation curve. An example of thermal inactivation curve is shown in Fig. 3.2.

28
Table 3.3. Kinetic parameters of 8 characterized proteins.
The kinetics were performed at 37 °C. Kinetic parameters were calculated using GraphPad Prism5 (GraphPad Software, Inc.). All kinetic curves obeyed the Michaelis-Menten model and showed R2 of fitness above 0.96. n=3, errors indicate standard deviation. An example of a kinetic curve is shown in Fig. 3.3.
Table 3.4. Kinetic parameters of the arylesterase EstA from Lactobacillus helveticus CNRZ32 and EstB from Lactobacillus casei LILA EstA1 EstB1 Substrate Km
(mM) Specific activity at Vmax (µmol•min-1•mg protein-1)
Vmax/Km Km (mM)
Specific activity at Vmax (µmol•min-1•mg protein-1)
Vmax/Km
pNP-acetate 0.16 120 750 1.9 20 11 pNP-butyrate 0.17 150 880 0.38 17 45 pNP-caproate 0.065 3.4 51 0.029 16 550
Data from reference [36]. Kinetic assays were conducted at 30 °C for EstA and 35 °C for EstB. Reactions were performed at protein concentrations of 0.064 to 3.0 µg protein/ml. Kinetic constants were calculated from Hyperbola (Hyperbol.fit) program of Sigma Plot 3.0.
Proteins ID Assayed conditions Parameters pH pNP substrates Km (mM) Specific activity at Vmax
(µmol•min-1•mg protein-1) Vmax/Km kcat (s-1) kcat / Km(s-1•mM-1)
RP_est1 RPA0348 7 pNP butyrate 0.5±0.0 139.8±2.7 279.6 75.5 151.0 RP_est2 RPA4646 9 pNP acetate 1.4±0.2 120.6±9.9 83.2 52.7 37.6 RP_est3 RPA3430 9 pNP caproate 2.1±0.3 11.1±1.0 5.2 5.0 2.4 PP_est1 PP0364 8 pNP acetate 1.4±0.1 8.8±0.5 6.4 3.8 2.7 PP_est2 PP5253 9 pNP acetate 1.1±0.1 25.0±1.0 22.5 12.4 11.3 PP_est3 PP3645 8 pNP caproate 0.2±0.0 23.3±0.5 136.8 12.1 60.5 PA_est1 PA2949 8 pNP butyrate 5.4±1.7 38.6±9.6 7.2 22.4 4.1 SAV_est1 SAV512 8 pNP acetate 0.3±0.0 6.0 ±0.2 22.2 2.8 9.3

29
Fig. 3.2. Thermal inactivation of RP_est1. The residual activities of enzymes were measured after preincubation at corresponding temperatures for up to 5 h. Error bars indicate standard derivation.
Fig. 3.3. Kinetics of RP_est1. Rate of hydrolysis of pNP-butyrate in the range of 0 to 0.95 mM performed at pH 7 and 37 °C. Error bars indicate standard derivation.

30
3.4 Conclusions
In this study, a total of 180 genes were successfully cloned and 84 of them were expressed as
soluble proteins in E.coli. All 84 soluble proteins were purified and then screened using
colorimetric assays; 46 proteins showed activity on pNP-acetate at pH 8 while 38 proteins did
not exhibit any activity towards the test substrates. Amino acid sequence alignment revealed that
most of the proteins, whether they were active or not, contained the conserved catalytic triad,
suggesting that additional, unidentified biochemical features are required for esterases and lipase
activity. Meanwhile, 8 bacterial esterases have been characterized. All of the enzymes showed
optimal activity at neutral or alkaline pH and did not show paraoxonase activity. To our best
knowledge, all 6 arylesterases (RP_est1, RP_est2, RP_est3, PP_est2, PP_est3, and PA_est1)
represent the first arylesterases to be isolated and characterized from Rhodopseudomonas
palustris, Pseudomonas putida, and Pseudomonas aeuginosa, respectively. PP_est3
demonstrated the highest thermal stability and solvent stability, suggesting that it is the best
bacterial candidate for subsequent transesterification trials.

31
Fig. 3.4. Amino acid sequence alignment of 8 characterized bacterial proteins. The 8 bacterial protein sequences were aligned with the parent sequence (P22862). The putative catalytic triad residues and the putative residues contributed to oxyanion hole are highlighted. The identical and similar residues are marked by (*) and (:), respectively.

32
Fig. 3.5. Amino acid sequence alignment of bacterial proteins that were active on phenyl acetate. The putative catalytic triad residues and the putative residues contributed to oxyanion hole are highlighted. The identical and similar residues are marked by (*) and (:), respectively. The full-length sequence alignment is provided in Appendix A.1.

33
TM0820 27 KNA---GIRKVLFLYGGGSIKKNGVYDQVV...SVVDSAKAVAAGALYEGDIWDAF 122 TM0423 26 SRF---GERAFVVIDDFVD--KNVLGENFF...VEEETDVVV--GIGGGKTLDTAK 99 ATu2215 19 GEN---RVRHGFFTRQGGV--SEGLYQGLN...VHSPDVVVV--DDNYDGNRPQAD 99 PP0624 14 PAP--ASVRACVTTRQGGV--SLPPYETFN...VADADPTVV--AE------ADAS 91 PA3301 22 PSA--TAVGAVMLSHGMAE--HAGRYERLA...HPELPIFLL--GHSMGSYISMAY 121 PP4551 22 LPA-TPVKAVVLLAHGMAE--HAGRYQRLG...FPCTPLFLF--GHSMGSYIAQAY 122 PSPPH0357 30 GSP---EGLPVVFIHGGPG--SGCDAHSRC...LGIDKWVLF--GGSWGSTLALAY 119 PP3523 8 GKP-------------------MSKFDFQL...LGLDTVEHI--ETTLLGMITLNG 65 ATu2497 7 GRT-DAEAPTILLSSGLGG--SSAYWLPQI...LNLEKFHFM--GHALGGLIGLDI 97 RPA3731 24 GDV-DPDKPVALLLHGTGF--VGEIWDEIA...LGLSGIYGI--GHSAGATDLLLT 112 PSPPH1999 23 GPK---DGQPVIALHGWLD--NANSFARLA...LGWQRFALL--GHSLGAIISVLL 111 SAV0974 18 GSG-----PPVVFSHGWPL--NADAWDVQL...LDLRDAVVV--GHSTGGGEVARY 104 ATu5389 18 GPK---DAQPIVFHHGWPL--SSDDWDAQM...LDLKNAVHI--GHSTGGGEVARY 106 PA2717 18 GPR---DAQVIYFHHGWPL--SSDDWDAQM...LGVRGAIHV--GHSTGGGEVVHY 106 PA3053 60 APK-KANGRTILLMHGKNF--CAGTWERTI...LGVARASVI--GHSMGGMLATRY 151 PP2201 59 AQG-QANGHNVVLMHGKNF--CAATWETTI...LGVKQTIVL--GHSTGGMLATRY 150 SAV0923 13 GRP---DGPVVLLAHGFGC--DQNMWRLVV...LDLRDVAFV--GHSVSAMVGVLA 104 ATu2061 31 GPG-----PVLLMLHGFSD--TSRSFSIIE...MGVSRFFVL--GHSMGAMTAIEL 114 SAV2105 28 GTG-----PLVLLVHGFPE--SWYSWRHQL...LGERTAVIV--GHDWGSNIAATS 116 RPA2773 21 GEG-----PLVVLCHGWPE--LSYSWRHQI...LGETRAAII--GHDWGAPVAWHA 109 SAV141 23 GQG-----PLVLLLHGWPE--SWYSWRHQF...LGEEQAVVV--GHDWGAPVAWTT 111 RHA05146 24 GDG-----EPLVLIHGTAL--SHAIWRGFG...LDLPSAHVL--GYSLGGRVALAL 113 RHA00069 20 GEG-----ETLLLVHGMAG--SSATWRAVL...LDIERVTVI--GQSLGGGVAMQF 105 SAV1746 41 GEG-----PVLVLIHGIGD--SSATWAELI...LDIESATLV--GHSLGGGVAMQF 126 PP4540 17 GQG-----EPLVLLHGLGS--SCQDWELQV...LHTGPVHFV--GLSMGGMVGFQF 102 PSPPH2566 19 GQG-----HPVVLIHGVGL--NKEMWGGQI...LQLPQATVV--GFSMGGLVARAF 104 PA0231 17 GPA---GAPVLLLSNSLGT--DLGMWDTQI...LELPRVHFC--GLSMGGLIGQWL 104 PP1380 17 GPD---DAPVLVLSNSLGT--DLGMWDTQI...LDIQKAHFV--GLSMGGLIGQWL 104 PA3586 44 GRD---SDPALLLVMGLGG--QLIHWPDEV...LDIPQAHVL--GASMGGMIAQHI 154 PP2804 21 GHE---AAPAVLLIVGLGL--QLTYWPQAL...LHLEAAHVV--GMSMGGMIAQTL 125 SAV0519 14 GGC----GRPVVLLHGLAG--HAGEWDTLA...LALHRPVLV--GQSLGGHTAMLT 100 SC0267 17 GTG-----PTLVFLHYWGG--SARTWDLVV...AGITDYVLV--GHSMGGKVAQLV 101 SC5943 22 GDG-----VPLLLVHGGES--DRTQFATLR...LGLARAHLL--GTSFGGAVAQHA 107 PSPPH0033 17 GTG-----PVVLLGHSYLW--DKAMWSAQI...LNIERCSIV--GLSVGGMWGAIA 103 RPA1568 16 GKPFDASLPAAVFIHGAGF--DHSVWALQT...TGAQPAKLI--GHSMGSLIALEA 106 SAV7198 43 GKG-----PAVLLLHGFPH--TWELWTDVM...LGAGPAAVV--GIDAGTPPALLL 128 SAV1548 17 GPHDGDGGVPLVFIHGWTA--DRHRWDHQM...LEIDRFIPV--GHSMGGMIAQTL 105 SAV4410 11 GTQ-RDTSLPLVLVHGHPF--DRTMWAPQL...LKADRFVLA--GLSMGGQIAMEC 100 TM0820 280 IAHGAGLAIV...QFERFAKKIFGFE...SLKDAG---IPEEDIDKIVDNVMLLVE 365 TM0423 267 YLHGEKVAIG...EEVGLPTTLAEIG...AEKACDKNETIHNEPQPVTSKDVFFAL 350 ATu2215 218 TKAGVKAENL...----GLCTYPDEH...RFYSYR-RTTHRAEPDYGRQISAIAIL 362 PP0624 202 IRLAARGVTA...--------VYGGG...TVSDAR-FFSYRRTPQGGRFASLVWLD 244 PA3301 242 IIGGERDPVS...LGD-LADALRGAG...TYPEARHELFNESNRDAVTQDLIDWLE 301 PP4551 243 VIGGECDPVS...LTH-LADALRATG...VYPEARHEVLNETNRGEVIADILGWLE 302 PSPPH0357 259 IVHGRYDVIC...AWE-LHQNWPGSE...VIREAGHSASESGIADALVRAAAEIAR 313 PP3523 97 KFYGCL----...-------------...MVDGLG---------PAMRFDILPK-- 118 ATu2497 203 VVATKDDLLV...SLR-LAEGLPQSE...LLDFGA-HAVNITEPDLFNTRLLQFLL 256 RPA3731 246 IATAELSWPI...ATR-ASALIPGAS...AFDGVG-HCVAQEAPTQLLTALEEFAA 299 PSPPH1999 231 LMVASDGMLA...QRQELLSALPFDV...LAGGHHLHLNDEQGARSVAHCINRFFA 282 SAV0974 218 IAHGDADQIV...SAPKSAELVKDAT...VYSGAP-HGLTGAYEQEFNADLLNFIR 272 ATu5389 223 VLHGEDDQIV...AARKSIKLLKNGQ...TYPGFS-HGMLTVNADVLNQDLLAFIK 276 PA2717 21 VMHGDDDQIV...SGVLSAKLLRNGT...TYPGFP-HGMPTTQAEVINADLLAFIR 275 PA3053 263 LLIGEKDNTA...GKD-AARRIPQAT...EFPDLG-HTPQIQAPERFHQALLEGLQ 332 PP2201 262 LLIGDKDTTA...GPQ-VARLIPKGE...TFEGMG-HAPQIEEPVRFNRTLVEWLG 331 SAV0923 211 VLECAQDVIA...GAY-VHAAIPGSR...TLDATG-HCPQLSAPDATAQAITAFLG 264 ATu2061 215 CLAGSEDPLF...-RQQLFEAFPLAQ...TMPGHG-HNPHWENPQGVSTLVTEFFA 268 SAV2105 268 FIGGGLDAST...AIEAYPVTLPGLV...ILDGCG-HWLQQERPQDTNRLLTDWLA 324 RPA2773 255 FIAGSNDPVI...HLAAINRVLPNLK...IIDGAG-HWIQQEKPAEVNAALIEFLK 313 SAV141 265 YVVGDRDMVT...EIF-RGQGGPGNP...VLPGCG-HWTQQERPVEVNAALLDFLT 336 RHA05146 213 LFVGSNDA--...DTQRVAALIPGAS...ILPGFD-HSTAVAASPEVLAAVEPFLA 265 RHA00069 223 LIWGDSDGII...GYA-AHEAIPGSR...VLDGVG-HYPHLEDPAAVVEIIDDFVS 276 SAV1746 245 LLWGDRDSVV...AYG-AHEAMPGSR...IFEGAG-HFPFHTDPARFLALVEEFTG 298

34
PP4540 206 VIAADHDYT-...LKERYVALIPNAR...VVDDSR-HATPLDQPEVFNQTLLQFLA 258 PSPPH2566 210 IATGELDPGS...ARE-LAMRISGAE...ILPDQR-HMMPVESPRLVNQVLLDFFE 263 PA0231 209 IVAGSHDAVT...ARF-MQARIADAQ...LVEFAAAHLSNVEAGDAFSRRLVDFLL 261 PP1380 209 IVAGTQDVVT...GRF-MQAGIQGAE...YVDFPAAHLSNVEIGEAFSRRVLDFLL 261 PA3586 265 VIHGTADPLL...GVH-VAAHIRGSE...LIPGLA-HRFQEAFKEPLLAAVVPYLK 318 PP2804 238 VIHGDKDLMV...GFA-TASAIRGAN...LLPGMG-HDLPTCLSGTLLSLLTGHMR 291 SAV0519 203 TVLGQSGIIS...--ETMLRRLPVAT...SVPGAG-HDVHLERPAVLRHLIQEFLE 255 SC0267 202 VVAGEHDQVE...LRDNLVPYLARAD...VVPRTG-HLIPLEAPADLADAVTAFAT 256 SC5943 203 IVHGTQDRLA...GALLMERRIPNAE...PIEGGR-HGIAVEFADTVAHRVREFLG 256 PSPPH0033 215 VMCGDADIPR...TRE--MASLIGCP...LVPEAG-HIANLENPDFVSGALMTFLA 267 RPA1568 210 FILGERDMMT...GKT-LAAAISGSR...ILKGAG-HTMMVERPDEVLKALQQ--- 260 SAV7198 248 -AVGSHPVGA...QLRPFADDLTGH-...LVEDCG-HIIPLHRPDALLALLHPFLK 298 SAV1548 204 LVHGYYDIQL...MLR-MAKDYPDAV...IVDAG--HELPVEKPAELTSALDRFVT 256 SAV4410 203 VVVGADDTFT...AAA-MHAALPGAT...VIDGAA-HLPNLERPEEFNAALGEFLF 256 Fig. 3.6. Amino acid sequence alignment of proteins that were not active on any substrates. The putative catalytic triad residues and the putative residues contributed to oxyanion hole are highlighted. The identical and similar residues are marked by (*) and (:), respectively. The full-length sequence alignment is provided in Appendix A.2.

35
Fig. 3.7. A phylogenetic tree of 39 enzymes that showed activity on tested substrates. The 8 characteried proteins were highlighted. RP_est1: RPA0348; RP_est2: RPA4646; RP_est3: RPA3430; PP_est1: PP0364; PP_est2: PP5253; PP_est3: PP3645; PA_est1: PA2949; and SAV_est1: SAV512.

36
4 Solvent Stability of Bacterial Carboxyl Esterases
4.1 Introduction
Esterases are valuable catalysts for biotechnological applications since many are able to both
hydrolyze and synthesize ester linkages, depending on the reaction condition. Further, they
usually exhibit high regio- and stereo-specificity under mild reaction conditions, making them
ideal tools for green chemistry, particularly when applied for the production of chiral pure
compounds. While reactivity under mild conditions is attractive for energy and environmental
considerations, commercial applications typically demand enzymes that are stable at high
temperature, acidic or basic pH, or in the presence of organic solutions. For instance, non-
aqueous conditions are required to force lipases and esterases to function as synthetic catalysts,
rather than catalyze hydrolysis reactions. Further, many lipase and esterase substrates are
lipophilic, suggesting that reaction efficiencies can be improved in the presence of an organic
solvent. As a result, esterases that are stable in detergents, organic solvents, or ionic liquids are
particularly useful for most industrial applications [1]. Given the impact of temperature on
substrate solubility and reaction rates, thermal stable enzymes are also desirable [2,3]. Notably,
a high temperature process can improve reaction performance by reducing solvent viscosity and
minimizing microbial contamination [4].
While enzyme immobilization can stabilize enzyme activity [5, 6], enzymes with innate stability
are preferred for industrial applications. However, predicting temperature and solvent stability
from primary sequence and protein structural analyses is often inconclusive, particularly since
less than 1 % differences in protein sequences can lead to significant changes in the temperature
stability [7]. Accordingly, two strategies that are frequently carried out to optimize enzyme
performance include: 1) screening for natural solvent- and/or thermo- stable enzymes [8-10], and
2) random mutagenesis and selection of temperature stable or solvent tolerant enzymes
[4,11,12]. For example, solvent-stable lipases and proteases were previously isolated from
organic solvent-tolerant microorganisms grown on solvent-rich media [1,13]. It was
hypothesized that the enzymes secreted during growth on these carbon sources must be solvent-
stable [14]. Lipase LST-03 from Pseudomonas aeruginosa was isolated in this way [13], which
has a longer half-life in the presence of organic solvents like DMSO and cyclohexane than in the

37
absence of an organic solvent. Solvent tolerant enzymes have also been discovered by
measuring residual enzyme activity after pre-incubation in different solvents [15]. This approach
typically uses heterologously expressed and purified proteins, and incorporates high throughput
enzyme screens in the presence of different solvents. More recently, ionic liquids (ILs) have
become attractive solvents for enzyme-catalyzed reactions. ILs represent a specific class of
organic solvents, and are considered green solvents due to their low volatility, thermostability,
high polarity, and recyclability [16]. Many studies have demonstrated the potential of ILs to
promote enzyme-catalyzed synthetic reactions [16-19]. In particular, lipases and esterases have
been used with ILs, and exhibit comparatively high (enantio)selectivity and operational stability
compared to reactions performed in organic solvents [20-23].
While several publications demonstrate the synthetic potential of lipases and optimization of
these catalyses [24-26], less effort has focused on synthetic reactions catalyzed by esterases,
including arylesterases. Exceptions include transesterification of a phenolic acid by a feruloyl
esterase from Fusarium oxysporum, and esterification of glycerol with sinapic acid by a feruloyl
esterase from Aspergillus niger [27, 28]. Phenolic acids, including sinapic acids, are derived
from lignin, which is a by-product generated during the production of biofuels, as well as pulp
for paper. The production of value-added materials from lignin is anticipated to off-set the cost
of biofuels, and increase the long term viability of future biorefineries whose main product is
bioenergy [29]. Accordingly, to increase the arsenal of enzymes capable of synthesizing new
and industrially relevant chemicals from lignin, 15 publicly available bacterial genomes were
mined for putative arylesterase-encoding genes. Corresponding genes were recombinantly
expressed and characterized as described in Chapter 3. This chapter reports the stability of eight
esterases in 7 detergents, 14 organic solvents and 4 ILs. The stability of these esterases was
compared to that of two commercial lipases (PS “Amano” SD and AY “Amano” 30). This
analysis is the first step to identifying new enzymes and reaction conditions that synthesize new
products from lignin.

38
4.2 Materials and Methods
4.2.1 Materials
Ionic liquids, including [BMIM]BF4, [BMIM]PF6, [BMIM]CF3SO3 were purchased from
Solvent Innovation GmbH (Germany) and Cyphos 109 was kindly provided by Cytec Industries
Inc. (Canada). Tween 20, Tween 60, Tween 80, Triton X-100, SDS, EDTA, and 2-
mercaptoethanol were purchased from Sigma (Canada). Solvents, including methanol, ethanol,
isopropanol, 1-butanol, tert-amyl alcohol, acetone, acetonitrile, tetrahydrofuran, 1,6-dixaone,
DMSO, toluene, p-xylene, hexane, cyclohexane were also purchased from Sigma (Canada). PS
“Amano” SD and AY “Amano” 30 were kindly supplied by AMANO Enzyme Inc. The
recombinant enzymes used in this study were recombinantly expressed and purified as described
in Chapter 3.
4.2.2 Effect of Detergents and Reducing Agent on Enzyme Stability
The effect of several detergents on enzyme stability was analyzed by preincubating 0.5 µg to 1
µg of each enzyme in 50 mM potassium phosphate buffer (pH 7), containing 1 % (v/v) of Tween
20, Tween 60, Tween 80 or Triton X-100, or 1 mM of SDS, EDTA, or 2-mercaptoethanol.
Residual activity was measured after 5 h incubation at 37 °C. The enzyme amount added to each
reaction was adjusted to ensure that initial reaction rates were measured. The standard activity
assay was performed to measure residual activity, as described in Chapter 3 (Section 3.2.5). In
brief, 10X concentrated enzymes (i.e. 0.5 µg/100 µL for RP_est1, 2.5 µg/100 µL for RP_est2, 5
µg/100 µL for RP_est3, 5 µg/100 µL for PP_est1, 5 µg/100 µL for PP_est2, 5 µg/100 µL for
PP_est3, 5 µg/100 µL for PA_est1 and 10 µg/100 µL for SAV_est1) were preincubated in each
detergent for 5 h preincubaton at 37 °C. The residual activity was measured by transferring 10
µL aliquots back to the standard conditions. The effect of the small amount of detergent in the
10 µL aliquot on background absorbency and enzyme activity was evaluated and found to be
negligible.
4.2.3 Effect of Organic Solvents on Enzyme Stability
Residual enzyme activity was measured using the standard assay following 5 h of perincubation
at 37 °C in the presence of the following solvents: methanol, ethanol, isopropanol, 1-butanol,

39
tert-amyl alcohol, acetone, acetonitrile, tetrahydrofuran, 1,6-dixaone, DMSO, toluene, p-xylene,
hexane, cyclohexane. Organic solvents were prepared in 15 %, 30 % or 50 % (v/v) in 50 mM
potassium phosphate buffer (pH 7). The effect of DMSO on enzyme activity was studied in
more detail. Here, 0.5 µg to 1 µg of each enzyme was incubated in buffer containing 0 to 40 %
of DMSO with 5 % increments. Residual activity was measured after 24 h incubation at 37 °C.
Specific activities were measured in the presence of 5 % to 95 % of DMSO; pNP-acetate was
used as the substrate. In brief, 10X concentrated enzymes (i.e. 0.5 µg/100 µL for RP_est1, 2.5
µg/100 µL for RP_est2, 5 µg/100 µL for RP_est3, 5 µg/100 µL for PP_est1, 5 µg/100 µL for
PP_est2, 5 µg/100 µL for PP_est3, 5 µg/100 µL for PA_est1 and 10 µg/100 µL for SAV_est1)
were preincubated in each organic solvent mixture for 5 h preincubaton at 37 °C. The residual
activity was measured by transferring 10 µL aliquots back to the standard conditions, as
described in section 3.2.5. Slight changes in absorbance readings that could result from
partitioning of substrate between phases was subtracted (Appendix B.5).
4.2.4 Effect of Ionic Liquids (ILs) on Enzyme Stability
Three imidazolium-based ILs ([BMIM]BF4, [BMIM]PF6, [BMIM]CF3SO3), and one
phosphonium-based IL (Cyphos 109) were used to study the effect of ILs on enzyme stability.
The imidazolium ILs were selected since they are most commonly used in lipase-catalyzed
synthetic reactions [17-26]. Enzymes were incubated for 3 h at 37 °C in the presence of 40 %,
50 %, 60 %, 70 %, 85 %, or 100 % of each solvent mixed with DMSO. Residual enzyme
activity was measured using the standard activity assay. In brief, 10X concentrated enzymes (i.e.
0.5 µg/100 µL for RP_est1, 2.5 µg/100 µL for RP_est2, 5 µg/100 µL for RP_est3, 5 µg/100 µL
for PP_est1, 5 µg/100 µL for PP_est2, 5 µg/100 µL for PP_est3, 5 µg/100 µL for PA_est1 and 10
µg/100 µL for SAV_est1) were preincubated in each IL cosolvent for 3 h preincubaton at 37 °C.
The residual activity was measured by transferring 10 µL aliquots back to the standard
conditions, as described in section 3.2.5.

40
4.3 Results and Discussion
This chapter extends the work described in Chapter 3, focusing on enzyme stability in a variety
of detergents and organic solvents. This analysis was conducted to identify the best enzyme
candidates for future applications in synthetic reactions.
4.3.1 Effects of Detergents, Chelators and Reducing agents on Hydrolysis Activities
The effect of detergents (Tween 20, Tween 60, Tween 80, Triton X-100, and SDS), chelator
(EDTA), and disulfide reducing agent (2-mercaptoethanol) on enzyme activity is summarized in
Table 4.1. Esterases from Rhodopseudomonas palustris showed higher stability compared to
esterases from Pseudomonas putida, Pseudomonas aeruginosa and Streptomyces avermitilis.
Notably, preincubation of RP_est1 with Triton X-100 and SDS appeared to have an activation
effect. Similarly, while SDS inhibited most of esterases, RP_est1 was activated by over 75 %.
Increases in enzyme activity after incubation may be explained by partial unfolding of enzymes
or possible complexes formed between enzymes and micelles [30, 31]. Additional experiments
such as Isothermal Titration Calorimetry (ITC) are required to confirm this hypothesis. PA_est1
activity was reduced following incubation with detergents, but was not inhibited by EDTA or 2-
mercaptoethanol. Instead, the activity of PA_est1 was 20 to 30 % higher after incubation with
EDTA and 2-mercaptoethanol compared to the control. These observations suggest that the
enzyme is not a metalloprotein and probably does not contain cysteine disulfide bridges that are
required for enzyme activity [32, 33]. Indeed, the primary sequence of PA_est1 contains only
one cysteine residue that is unable to form a disulfide bond. To confirm if the enzymes are
metalloproteins, additional experiments should be performed as described by Hogbom et al. [34]
based on detection of chemiluminescence produced by luminol in the presence of certain metal
catalysts when mixed with a base and an oxidant, and color production of some metal complexes
of the chelator 4-(2-pyridylazo)resorcinol. PP_est2 and RP_est3 also contain only one cysteine
residue and SAV_est1 does not contain any cysteine residues. However, their stability was
slightly affected in the presence of 1 mM 2-mercaptoethanol. The presence of disulfide bridges
in esterases that contained more than one cysteine residue and were affected by EDTA (i.e.,
RP_est2, and PP_est1) will be evaluated using corresponding structural information, which is
currently being acquired. The activity of all esterases isolated from P. putida decreased after

41
preincubation with the four nonionic detergents, except for PP_est3, which was stable in the
presence of Triton X-100. The stability profiles of RP_est1 and RP_est2 were similar to the
commercial lipases PS-SD and AY-30, which were stable in the presence of detergents and
slightly inhibited by EDTA and 2-mercaptoethanol.
Table 4.1. Effect of detergents, inhibitor, and disulfide reducing agent on enzyme stability.
Residual activities were measured after 5 hr incubation at 37 °C. n= 6; errors indicate standard derivation. The values of Critical Micelle Concentration (CMC) for nonionic and ionic detergents were obtained from [39] and [40], respectively.
4.3.2 Effects of Organic Solvents on Hydrolysis Activities
All esterases were more stable in non-polar solvents (log P> 2.5) than in polar solvents (log P<
0.89) (Table 4.2). This is consistent with the general agreement that water miscible polar
solvents can strip off the water layer that forms around many proteins, which is essential for
activity [15]. In contrast, non-polar solvents may protect the microenvironment of enzymes,
thereby preventing loss of the water layer. Among the esterases that were screened in this study,
RP_est3 was the most stable in polar solvents. Even after preincubation with 50 % of all tested
Residual activity (%) Detergents CMC at 22°C
RP_ est1
RP_ est2
RP_ est3
PP_ est1
PP_ est2
PP_ est3
PA_ est1
SAV_ est1
None n/a 100 100 100 100 100 100 100 100 1 % v/v Tween 20
0.042 mM
131±5 96±14 64±8 31±8 17±1 82±24 24±4 78±9
1 % v/v Tween 60
0.022 mM
136±39 93±9 88±7 40±10 89±2 78±26 101±15 90±5
1 % v/v Tween 80
0.028 mM
135±9 100±4 91±8 64±9 80±3 50±3 32±8 70±1
1 % v/v Triton X-100
0.20 mM
174±8 93±16 90±9 27±8 58±3 110±10 25±7 67±16
1 mM SDS 5.29 mM
177±3 95±1 73±9 30±8 58±6 53±6 40±4 28±7
1 mM EDTA
n/a 93±1 85±9 80±21 40±8 66±15 98±9 130±16 88±4
1 mM 2-Mercaptoethanol
n/a 89±35 86±4 88±13 44±11 85±8 95±10 123±9 82±13

42
solvents, RP_est3 retained more than 50 % of its original activity. While solvent stability is
important during the equilibration step of enzymatic reactions performed in organic solvents,
enzyme activity in the presence of organic solvents must be evaluated independently. This is
because solvent stability is not well correlated to enzyme activity in the presence of the
corresponding solvent.
None of the esterases exhibited higher stability in organic solvents compared to the commercial
lipases PS-SD and AY-30. This could be attributed to the lid-like structure that many lipases
display close to the active site [35-37]. This lid structure, and surrounding non-polar residues,
could reduce the accessibility of polar solvents to active site residues.
Since lipase-catalyzed transesterification reactions are typically performed in up to 40 % DMSO,
the standard activity assay was performed in the presence of 0 to 95 % DMSO (Fig. 4.1). While
RP_est3 and PP_est3 were activated by preincubation with 15 to 30 % or 30 to 50 % of DMSO,
respectively, the specific activity of these enzymes decreased rapidly when measured in the
presence of more than 10 % DMSO. By contrast, in the presence of 40 % DMSO, PP_est2,
RP_est2, RP_est1 and SAV_est1 exhibited 70 %, 46 %, 66 %, 39 % of residual activity. Their
performance could be further improved by lyophilization and/or immobilization. DMSO is used
as a co-solvent to prevent hydrolysis reactions when using substrates that are soluble in aqueous
solutions. Accordingly, the relative stabilities and activities of arylesterases in non-polar
solvents including hexane, butanol and isopropanol might be more relevant to predicting their
ability to catalyze the esterification of lignin subunits with aliphatic molecules.

43
Table 4.2. Effect of various organic solvents on enzyme stability.
Residual activity (%) Organic solvents
LogP Conc. (v/v%) PS-SD AY-30 RP_
est1 RP_ est2
RP_ est3
PP_ est1
PP_ est2
PP_ est3
PA_ est1
SAV_ est1
None - - 100 100 100 100 100 100 100 100 100 100 15 105±10 85±5 112±25 83±6 138±12 42±10 78±3 100±14 77±11 103±17 30 105±8 69±4 118±18 94±6 171±7 32±7 54±2 111±11 28±7 103±4
DMSO -1.22
50 116±12 18±7 16±2 110±3 70±44 0±3 0±0 121±11 25±6 4±2 15 86±5 96±4 112±14 83±6 72±7 92±53 75±6 87±18 35±7 37±13 30 100±12 104±9 132±17 84±8 66±4 0±1 3±0 70±31 22±4 1±1
Methanol -0.76
50 105±12 75±14 139±26 3±0 68±10 0±2 0±1 103±9 20±3 1±1 15 102±12 57±9 86±2 85±7 98±8 0±3 0±0 97±8 35±8 6±2 30 111±26 35±5 12±2 7±2 91±11 1±2 0±0 78±6 33±4 1±1
1,6-Dixaone -0.42
50 125±26 4±5 0±2 5±1 114±5 5±4 0±0 1±1 38±7 5±1 15 87±4 29±46 109±8 85±2 72±4 0±2 47±2 86±29 21±2 123±8 30 70±12 98±22 111±7 3±0 84±13 0±2 0±1 110±38 20±2 2±2
Acetonitrile -0.34
50 95±13 12±9 0±1 3±1 84±12 0±21 0±5 0±3 23±5 0±12 15 94±5 66±11 94±11 98±5 70±5 0±3 12±7 115±14 18±6 53±24 30 110±16 38±8 79±17 6±2 72±10 0±3 0±1 95±14 19±3 0±1
Isopropanol -0.28
50 116±22 3±5 2±1 3±1 88±12 0±3 0±1 21±22 15±4 0±1 15 91±3 63±2 104±5 87±5 76±5 0±1 56±5 103±15 20±2 26±10 30 104±16 49±2 124±11 67±2 80±4 0±2 1±0 89±12 16±4 0±1
Ethanol -0.24
50 108±15 14±7 3±1 3±1 73±11 0±2 0±0 34±8 25±7 1±2 15 89±5 111±12 99±6 84±9 71±4 6±3 53±10 105±11 36±11 118±15 30 91±14 109±32 75±24 94±4 66±5 0±2 7±16 117±19 22±3 128±18
Acetone -0.23
50 96±30 72±10 172±12 3±0 72±9 0±2 0±0 86±8 19±4 4±3

44
15 89±3 105±12 118±4 81±3 79±9 0±1 0±1 108±21 24±3 119±16 30 87±3 115±8 119±8 3±0 84±12 0±2 0±1 89±7 24±4 6±1
Tetrahydro-furan
0.49
50 88±15 50±51 15±4 4±1 104±10 0±1 0±0 0±0 32±6 2±2 15 61±13 17±3 2±0 86±4 70±4 2±1 4±2 111±16 22±3 1±1 30 79±11 13±7 1±1 90±4 62±7 0±1 1±1 125±27 23±3 1±1
1-Butanol 0.8
50 114±27 20±6 2±1 88±28 68±12 2±2 1±0 94±20 24±4 1±1 15 96±8 34±11 86±14 94±4 80±4 0±3 0±0 125±11 26±3 0±2 30 86±19 4±5 4±1 99±4 69±7 0±3 0±0 133±23 25±4 0±1
Tert-amyl alcohol
0.89
50 106±8 0±6 3±0 87±33 72±16 0±6 0±0 164±26 24±4 0±3 15 95±51 119±4 101±7 83±6 101±9 38±5 15±12 44±57 105±3 111±16 30 100±55 131±13 122±7 83±8 119±6 94±18 99±5 81±62 130±7 127±18
Toluene 2.5
50 97±36 182±20 181±14 82±17 132±39 148±15 115±5 174±12 160±54 167±31 15 81±9 137±11 120±7 73±2 87±35 176±45 51±17 88±31 107±5 104±18 30 64±14 250±90 118±6 69±9 127±10 120±11 94±6 135±9 121±49 106±33
p-Xylene 3.1
50 65±28 323±70 169±14 79±14 116±40 320±183 117±7 194±34 178±54 135±31 15 73±8 154±10 112±5 72±8 125±8 11±15 42±47 63±56 126±17 116±16 30 85±7 190±30 140±6 77±8 131±18 176±74 78±35 56±41 142±21 147±15
Cyclohexane 3.2
50 77±25 20013 187±17 74±7 162±70 356±127 126±9 166±38 166±75 208±16 15 89±9 93±3 105±10 78±6 115±8 32±36 62±41 43±58 95±20 132±8 30 85±12 142±20 116±10 76±9 130±6 300±101 66±32 85±64 138±12 143±8
Hexane 3.9
50 97±17 195±14 165±12 79±14 147±55 285±70 120±14 202±22 143±74 187±19
Residual activities were measured after 5 h incubation at 37 °C. The column “conc. v/v %” indicates the percentage of corresponding organic solvents in 50 mM potassium phosphate buffer (pH 7). n= 6; errors indicate standard derivation.

45
4.3.3 Effects of Ionic Liquids (ILs) on Hydrolysis Activities
Ionic liquids offer new options for retaining enzyme activity during synthesis reactions that must
be performed in the absence of water. In this study, esterase stability was evaluated using three
typical imidazolium-based ILs and one phosphonium-based IL. A co-solvent system containing
DMSO and the selected IL was generated. None of the recombinant or commercial enzymes was
active in 40 % to 100 % of the phosphonium-based IL (Cyphos 109). Therefore, only the results
for imidazolium-based ILs are shown (Table 4.3). [BMIM]PF6 significantly destabilized all
esterases for P. putida and S. avermitilis. By contrast, RP_est2 and RP_est3 from R. palustris
retained 40 % and 60 % of activity after preincubation with 85 % of this IL. In general,
increasing the fraction of IL in DMSO/IL co-solvents to at least 85 % increased measured
residual activities (Table 4.3). An exception was SAV_est1, which retained 85 % activity in 100
% of DMSO. Overall, RP_est3 displayed higher stability than the commercial lipases PS-SD
and AY-30, suggesting that this enzyme is a particularly interesting target for application
development. Notably, RP_est3 appeared to be activated by preincubation with [BMIM]BF4 and
[BMIM]CF3SO3. A similar result was observed for AY-30 and PP_est3 with [BMIM]CF3SO3,
albeit to a lesser extent. The mechanism behind IL stabilization of enzymes remains unclear
[38]. However, two recent spectroscopic studies of protein thermostability in ILs suggest that a
stabilization effect may result from alteration in the protein hydration level and structural
compaction as well as other additional factors such as free volume contributions, ionic
interactions and confinement effects [38].
Owing to their limited commercial availability and the extensive use of mechanistically related
lipases as synthetic catalysts, the arylesterases characterized in this study constitute an important
set of new, industrially relevant catalysts. Their ability to catalyze hydrolysis reactions in the
presence of organic solvents, as well as esterification of lignin-derived compounds is currently
being evaluated.

46

47
Fig. 4.1. Effect of DMSO on enzyme activity. Enzyme activity was measured in the presence of 5 to 95 % DMSO at 37 °C using pNP-acetate as the substrate. n= 6; error bars indicate standard deviation.

48
Table 4.3. Effect of ionic liquids on enzyme stability. Residual activity (%) Ionic
liquids Conc. (v/v% in DMSO)
PS-SD
AY-30 RP_est1 RP_est2 RP_est3 PP_est1 PP_est2 PP_est3 PA_est1 SAV_est1
None - 100 100 100 100 100 100 100 100 100 100 100% DMSO
- 0±0 16±3 5±2 4±2 59±8 0±1 8±1 0±4 22±2 85±3
40 3±0 70±11 9±2 6±4 67±3 13±8 9±1 11±5 36±1 14±1 50 3±0 72±14 9±3 6±4 74±7 13±8 8±1 13±8 31±6 12±2 60 3±0 78±14 10±3 5±3 84±9 14±7 11±0 18±12 41±5 15±2 70 24±16 73±19 11±3 20±19 102±11 17±12 13±1 14±14 47±8 15±2 85 60±19 91±15 13±4 78±20 184±16 18±10 16±1 25±17 47±8 24±4
[BMIM]BF4
100 12±3 105±19 20±6 84±14 205±53 30±19 23±2 39±27 60±3 37±8 40 0±1 19±8 1±2 3±4 43±18 0±6 0±2 0±13 20±18 0±2 50 0±1 14±5 0±5 4±3 20±12 0±8 0±7 0±12 22±21 0±4 60 23±2 7±14 0±3 22±18 41±21 0±27 0±8 0±39 23±11 1±5 70 35±4 16±18 5±4 61±8 81±12 2±13 0±8 0±2 23±16 0±1 85 20±7 33±28 41±19 61±11 77±12 0±28 0±4 42±28 20±2 6±4
[BMIM]PF6
100 12±5 32±40 2±3 10±5 11±18 0±8 0±4 0±4 10±15 0±4 40 3±0 82±14 11±2 6±4 84±10 16±9 13±1 13±±53 34±9 13±1 50 4±0 93±8 11±2 6±6 94±17 20±11 15±1 18±10 50±7 14±1 60 6±1 100±8 13±2 9±2 108±22 25±14 18±2 56±22 54±7 16±1 70 44±9 116±7 21±3 35±12 119±13 27±16 20±2 84±44 61±11 19±3 85 40±3 135±14 51±9 44±19 176±19 42±21 37±6 120±44 81±13 28±4
[BMIM]CF3SO3
100 34±6 145±11 41±10 63±10 177±14 67±16 96±10 121±25 64±19 27±1 Residual activities were measured after 3 h incubation at 37 °C. The column “conc. v/v % in DMSO” indicates the percentage of corresponding ILs in DMSO during the incubation. n= 6; errors indicate standard derivation.

49
5 Isolation and Characterization of Two, Solvent-Tolerant, Thermophilic Archaeal Esterases
5.1 Introduction: Extreme Environments: A Source of Unique Biocatalysts
Archaea represent one of the three domains of living organisms. They are single celled
microogranisms, and display unique properties that the other domains do not share. For instance,
archaea are often found in extreme habitats such as high temperature or highly acidic or basic
environments. Organisms that withstand these environments, and indeed thrive in them, are
called extremophiles. For instance, Archaeoglobus fulgidus are hyperthermophiles and can be
found in hydrothermal vents and hot springs. Enzymes that are isolated from extremophiles
often exhibit superior performance in industrial applications [1]. Amongst extremophiles,
thermophilic archaea have attracted significant attention, especially for their production of
thermophilic enzymes [1]. A key example is Taq polymerase from Thermus aquaticus, an
enzyme ubiquitously used in PCR reactions, a core molecular biology technique [2]. There are
several advantages to performing biocatalyses at an elevated temperature, including enhanced
reaction rates, increased substrate solubility and decreased risks of contamination [1]. In
addition, thermophilic enzymes usually exhibit high resistance against many organic solvents
and detergents and are more resistant to proteolytic attack [1].
Despite the industrial relevance of temperature stable enzymes that can be retrieved from
hyperthermophilic archaea, most industrial enzymes that are currently used were isolated from
either bacteria or fungi. Archaeal enzymes that have received most attention to date include
amylases, cellulases, xylanases, and proteases; a handful of lipases and esterases have also been
reported [1,3]. The industrial interest in lipases and esterases resides in the fact that they
typically do not require cofactors, show high regio- and stereospecificity, are often
comparatively stable and active in organic solvents, and can perform both hydrolytic and
synthetic reactions [4].
More lipases than carboxyl esterases have been described to date, and at the time of writing, only
one example of an arylesterase from an archaeal species had been reported [5]. Additional
examples of arylesterases isolated from thermophilic archaea will enable the identification of

50
sequence motifs and structural features that determine substrate specificity and thermostability
within this class of enzymes. Understanding these biochemical determinants will greatly
facilitate subsequent optimization of substrate preference and stability of arylesterases, and
possibly other carboxyl esterases and lipases.
Herein, we report the cloning, recombinant expression, and characterization of two thermostable
esterases from A. fulgidus. The purified enzymes were most active on phenyl acetate and pNP-
acetate in comparison to other pNP-esters and retained over 80 % of activity after 5 h incubation
at up to 85 °C and in the presence of several detergents and organic solvents. These two
esterases exhibited significantly lower thermal inactivation than the bacterial esterases described
in Chapter 3. In contrast to the bacterial enzymes, they were also comparatively stable in both
polar and nonpolar solvents.
5.2 Materials and Methods
Genes for this study were isolated from the Archaeoglobus fulgidus genomic DNA purchased
from the ATCC (Cat. # 49558). Please refer to Chapter 3 for details regarding gene cloning,
recombinant expression in E. coli, protein purification, and enzyme characterization using pNP
substrates, as well as phenyl acetate. To evaluate enzyme activity at optimal pH, pNP-palmitate
was used as the substrate, since it is difficult to monitor pH-acetate at pH 10 and pH 11. Please
refer to Chapter 4 for details regarding enzyme stability measurements in the presence of
detergents, organic solvents and ionic liquids.
5.3 Results and Discussion
Six putative esterase genes from A. fulgidus were cloned and expressed in E. coli. Of these, two
were soluble, three were insoluble and one protein was not expressed. The two soluble proteins
(AF_est1 and AF_est2) were biochemically characterized to determine their substrate preference,
optimal pH, kinetic parameters, pH stability, thermal inactivation, as well as their stability in the
presence of various detergents, organic solvents, and ionic liquids.

51
5.3.1 Esterase Activity with p-Nitrophenyl Substrates
Lipases and esterases share similar conserved sequence motifs, and can be differentiated based
on substrate preference. Therefore, to investigate whether AF_est1 and AF_est2 were esterases
or lipases, the activities of these enzymes were screened using a series of short to long-chain
pNP-esters. Consistent with esterase activity, both enzymes showed preference for short-chain
pNP-esters, with the highest activity towards pNP-acetate (Fig. 5.1). To a lesser extent, AF_est1
and AF_est2 also hydrolyzed long-chain pNP-esters including pNP-palmitate, an activity that
was not observed by another esterase (Est-AF) isolated from A. fulgidus [6]. A comparison of
specific activities on all of the substrates tested indicated that both esterases preferred phenyl
acetate, a substrate that is commonly used to identify arylesterase (Table 5.1). Many
arylesterases are also paraoxonases, and are not inhibited by organophosphates, like paraoxon [7-
9]. However, AF_est1 and AF_est2 did not show any activity towards paraoxon, and so are not
paraoxonases.
Table 5.1. Activities (µmol•min-1•mg protein-1) of enzymes using 0.5 mM of substrate. Esterases Specific activities of the test substrates
pNP
acetate pNP
caproate pNP
palmitate pNP
benzoate Phenyl acetate
Tributyrin
Olive oil
AF_est1 0.74±0.01 0.36±0.01 0.08±0.02 n.a1 7.04±0.29 n.a n.a AF_est2 0.81±0.01 0.59±0.00 0.15±0.00 n.a 7.93±0.33 n.a n.a
1. n.a means not active.
All assays were performed using standard conditions, and were incubated for 30 min at 37°C. n=3; errors indicate standard derivation.
AF_est1 shared 20 % sequence identity with AF_est2 in a global alignment. The alignment of
AF_est1 and AF_est2 with three known archaeal esterases confirmed the presence of a
conserved catalytic triad sequence, consisting of Ser, Asp, and His, where the serine residue is
located in the conserved Gly-X-Ser-X-Gly pentapeptide sequence (Fig. 5.2). An X-ray structure
has been solved for the esterase from A. fulgidus (AFEST; O28558) [10]. AF_est1 and AF_est2
shared 8.6 % and 19.8 % sequence identity with AFEST, respectively. Based on the alignment
and positions of catalytic triad predicted from X-ray structure of AFEST, it showed that Ser95,
His210, Asp238 constitute the catalytic triad in AF_est1, and that Ser98, His214, and Asp243
constitute the catalytic triad in AF_est2. AF_est1 and AF_est2 sequences were also aligned to an

52
arylesterase isolated from Sulfolobus solfataricus (Fig. 5.3). Notably, the pentapeptide sequence
of S. solfataricus is slightly downstream compared to AF_est1 and AF_est2; AF_est1 and
AF_est2 shared 2.3 % and 13.2 % identity to this arylesterase, respectively. An alignment of
AF_est1 and AF_est2 to the eight bacterial esterases described in Chapter 3 showed that the
putative catalytic triad was perfectly aligned for all of them, except for RP_est1, which contained
two analogous pentapeptide sequences, one upstream and one downstream of the conserved
pentapeptide sequence. Features that distinguished the archaeal enzymes from the bacterial
enzymes were not identified by primary sequence comparisons (Appendix A.3). Future
bioinformatic analyses will attempt to identify residues that contribute to the thermal inactivation
of the archaeal enzymes.
Fig. 5.1. Substrate preference of AF_est1 and AF_est2 on pNP-esters. Activities of AF_est1 (black) and AF_est2 (grey) with various pNP-esters were determined photometrically at 400 nm at 37 °C. Enzymes were diluted in 50 mM potassium phosphate buffer (pH 7) and adjusted to ensure that initial reaction rates were measured in the kinetic cycle. n= 6; error bars indicate standard deviation.

53
Fig. 5.2. Amino acid sequence alignment of AF_est1 and AF_est2 with other esterases from Archaeoglobus fulgidus. Three esterases were listed by their Swiss-Prot accession numbers. The putative catalytic triad residues were highlighted. The identical and similar residues were marked by (*) and (:), respectively.

54
Fig. 5.3. Amino acid sequence alignment of AF_est1 and AF_est2 with an arylesterases from Sulfolobus solfataricus. The arylesterase was listed by their Swiss-Prot accession numbers) The putative catalytic triad residues were highlighted. The identical and similar residues were marked by (*) and (:), respectively.
AF_est1 had an optimal activity at pH 9.5 whereas AF_est2 showed highest activity between pH
9 and 10 (Fig. 5.4). Both enzymes also showed high stability in alkaline buffers, even at pH 11
(Fig. 5.5). These results indicate that AF_est1 and AF_est2 are more stable at alkaline pH than
the previously reported esterase from A. fulgidus, which lost activity above pH 8 [6]. AF_est1
and AF_est2 were also stable at high temperatures. Even after 5 h at 90 °C, AF_est1 and
AF_est2 retained 42 % and 52 % of activity, respectively (Fig. 5.6). Moreover, the activities of
AF_est1 and AF_est2 activities increased slightly after 2 h incubation at 75 to 84.5 °C. This
activation effect decreased for AF_est2 after 5 h incubation. In comparison, the thermal
stabilities of AF_est1 and AF_est2 were 4 to 5 times greater than Est-AF, an esterase previously
isolated from A. fulgidus [6]. However, both of them were not as stable as the arylesterase
isolated from S. solfataricus [5], which retained 52 % of its activity after 50 h of incubation at 90
°C.

55
Fig. 5.4. pH profiles of AF_est1 (black) and AF_est2 (grey). The activity was measured with 0.5 mM pNP-palmitate at 37 °C in the Britton and Robinson’s universal buffers with different pH. n= 6; error bars indicate standard deviation.
Fig. 5.5. Effects of pH on AF_est1 (black) and AF_est2 (grey) stability. The residual enzyme activity was measured with pNP-acetate at 37 °C after 24 h incubation in the Britton and

56
Robinson’s universal buffers with different pH at 37 °C. n= 6; error bars indicate standard deviation.
Fig. 5.6. Effect of thermal inactivation on AF_est1 and AF_est2. Enzymes were diluted in 50 mM potassium phosphate buffer (pH 7). The residual enzyme activity was measured with pNP-acetate at 37 °C after 2 h (black) and 5 h (grey) incubation in different temperatures. n=6; error bars indicate standard deviation.

57
5.3.2 Effect of Additives and Organic Solvents on Esterase Stability
Esterases, including arylesterases have significant industrial potential, since they have the
capacity to catalyze hydrolytic as well as synthetic reactions. To promote synthetic catalyses,
reactions are performed in the absence of water, and instead in organic solvents or more recently,
ionic liquids. Accordingly, enzymes that are stable in the presence of detergents, organic
solvents, and ionic liquids are particularly suitable for biotechnological applications. The
stability of AF_est1 and AF_est2 was investigated in the presence of detergents, a chelating
agent, a disulfide reducing agent, conventional organic solvents, and ionic liquids. Both
esterases retained over 85 % of activity after 5 h incubation in the presence of detergents, EDTA
and 2-mercaptoenthanol (Table 5.2). AF_est1 was slightly more stable than AF_est2, and
appeared to be activated in the presence of 1 mM SDS. Increases in enzyme activity after
incubation may be explained by partial unfolding of enzymes or possible complexes formed
between enzymes and micelles [5, 11].
Table 5.2. Effects of various surfactants on AF_est1 and AF_est2 stability. Residual activity of (%) Organic solvents CMC at 22°C
AF_est1 AF_est2 None n/a 100 100 1 % v/v Tween 20 0.042 mM 100±2 91±8 1 % v/v Tween 60 0.022 mM 113±20 102±16 1 % v/v Tween 80 0.028 mM 96±14 87±10 1 % v/v Triton X-100 0.20 mM 114±11 99±7 1 mM SDS 5.29 mM 131±1 101±8 1 mM EDTA n/a 114±1 104±20 1 mM 2-Mercaptoethanol n/a 105±16 93±11
Residual activities were measured after 5 h incubation at 37 °C. n= 6; errors indicate standard derivation. The values of Critical Micelle Concentration (CMC) for nonionic and ionic detergents were obtained from [20] and [21], respectively.
The destabilizing effects of the organic solvents were investigated using three solvent
concentrations (Table 5.3). With the exception of isopropanol, tetrahydrofuran, and acetonitrile,
AF_est1 and AF_est2 retained 70 % of activity after 5 h of incubation in 50 % of each solvent
(Table 5.3). This is consistent with the observation that thermophilic enzymes are typically
solvent tolerant, giving a great potential for these enzymes to catalyze reactions in organic media

58
[1,12]. Furthermore, both esterases were more stable in non-polar solvents with log P> 2.5 than
in polar solvent with log P< 0.89. A possible explanation for this observation was described in
Section 4.3.2, whereby polar solvents may strip off the essential water layer around the protein,
leading a destabilizing effect. By contrast, hydrophobic organic solvents appear to stabilize the
water layer at the protein surface, leading to reduced enzyme flexibility [13]. The activation
effect of several solvents is likely due to an improved packing of the polar side chains at the
protein surface [13].
While both the archaeal and bacterial esterases were reasonably stabile in non-polar solvents,
AF_est1 and AF_est2 exhibited higher stability in polar organic solvents compared to the
bacterial esterases. Similar to AFEST (O28558), the thermophilic carboxyl esterase from A.
fulgidus, AF_est1 and AF_est2 might contain additional intramolecular ion pairs, fewer loop
extensions, and a lower ratio of hydrophobic to charged amino acids at the protein surface,
compared to the bacterial enzymes [9]. Crystallization trials for AF_est1 and AF_est2 have
begun to test these predictions. Unfortunately, the solvent stability of AFEST and other archaeal
esterases has not been well characterized. Nevertheless, structural characteristics that lead to
solvent stability are often correlated to thermal stability.
Table 5.3. Effects of various organic solvents on AF_est1 and AF_est2 stability. Residual activity of (%) Organic solvents log P Conc. (v/v%)
AF_est1 AF_est2 None - - 100 100
15 91±7 92±9 30 93±3 85±3
DMSO -1.22
50 83±3 86±13 15 100±8 88±27 30 84±4 97±5
Methanol -0.76
50 81±26 90±7 15 90±11 106±6 30 96±9 94±8
1,6-Dixaone -0.42
50 79±18 64±5 15 77±11 75±10 30 78±6 85±2
Acetonitrile -0.34
50 54±8 46±8 15 62±11 69±7 Isopropanol -0.28 30 51±8 56±3

59
50 42±11 35±4 15 103±18 92±4 30 74±5 85±17
Ethanol -0.24
50 69±16 59±10 15 91±9 104±12 30 80±7 93±6
Acetone -0.23
50 68±12 63±7 15 95±13 113±13 30 75±5 72±4
Tetrahydrofuran 0.49
50 16±4 11±2 15 68±7 66±9 30 69±8 66±19
1-Butanol 0.8
50 72±4 54±36 15 74±3 74±3 30 71±11 79±16
Tert-amyl alcohol 0.89
50 91±1 62±41 15 109±5 121±27 30 124±6 115±26
Toluene 2.5
50 138±22 129±60 15 108±11 124±30 30 134±11 120±35
p-Xylene 3.1
50 144±17 130±58 15 128±10 134±28 30 160±29 129±30
Cyclohexane 3.2
50 113±70 135±71 15 103±22 116±19 30 169±44 134±31
Hexane 3.9
50 116±40 130±52 Residual activities were measured after 5 h incubation at 37 °C. The column “conc. v/v %” indicates the percentage of corresponding organic solvents in 50 mM potassium phosphate buffer, pH 7 during the incubation. n= 6; errors indicate standard derivation.
Ionic liquids (ILs) are intriguing alternatives to organic solvents given their low vapor pressure,
and potentially reduced effect on enzyme stability. Indeed, several studies have already
demonstrated the ability of lipases and esterases to catalyze synthetic reactions in the presence of
ionic liquids [14-17]. In this study, the impact of three imidazolium ILs ([BMIM]CF3SO3,
[BMIM]PF6, and [BMIM]BF4) and one phosphonium IL (Cyphos 109) on AF_est1 and AF_est2
stability was evaluated. The ILs were selected based on previous reports demonstrating their

60
ability to promote lipase-catalyzed synthetic reactions [14-17]. Little or no enzyme activity was
retained after incubation in the phosphonium-based ionic liquids, and so only results using the
imidazolium-based ILs are shown (Table 5.4). Both AF_est1 and AF_est2 exhibited higher
stability in the presence of [BMIM]CF3SO3 than the other two ILs. In most cases, the stability of
AF_est1 and AF_est2 increased with increasing concentrations of ionic liquids. Since previous
studies of lipase-catalyzed transesterification reactions in ILs used 100 % of the IL, or co-
solvents using up to 50 % of an organic solvent, the current results suggest that both AF_est1 and
AF_est2 are ideal candidates for application development. Accordingly, subsequent studies will
evaluate the ability of AF_est1 and AF_est2 to catalyze the transesterification of phenolic acids
or sugars with short-chain aliphatic compounds.
Table 5.4. Effects of ionic liquids on AF_est1 and AF_est2 stability. Residual activity (%) Ionic liquids Conc.
(v/v% in DMSO) AF_est1 AF_est2 None - 100 100 100% DMSO - 12±0 12±2
40 22±3 12±2 50 24±0 13±1 60 36±3 23±10 70 56±11 58±7 85 81±13 70±11
[BMIM]BF4
100 56±22 58±10 40 10±12 9±4 50 32±12 18±5 60 60±26 38±12 70 57±14 47±3 85 35±3 49±20
[BMIM]PF6
100 8±9 9±14 40 29±2 17±0 50 38±9 30±4 60 96±16 56±14 70 109±21 76±7 85 116±8 96±4
[BMIM]CF3SO3
100 138±28 109±44 Residual activities are measured after 3 h incubation at 37 °C. The column “conc. v/v % in DMSO” indicates the percentage of corresponding ILs in DMSO during the incubation. n= 6; errors indicate standard derivation .

61
5.3.3 Kinetic Properties
Enzyme kinetic parameters were measured to obtain more detailed characterization of AF_est1
and AF_est2. The enzymatic reactions were performed at 37 °C to facilitate the detection of
enzymatic hydrolysis of pNP-acetate at low substrate concentrations; autohydrolysis of pNP-
acetate is detected at higher temperatures [18]. The Km and kcat of AF_est1 and AF_est2 were
3.8 mM and 5.1 s-1, and 3.7 mM and 10.3 s-1, respectively. These kinetic parameters are
approximately 10 to 20 times lower than the thermophilic esterase from Bacillus acidocaldarius
and Sulfolobus shibatae, which were characterized using pNP-acetate at 37 °C (0.183 mM and
145 s-1, 0.013 mM, and 56.1 s-1, respectively) [18, 19]. The comparatively low activity of
AF_est1 and AF_est2 of pNP-acetate is not surprising given their preference for phenyl acetate.
The reaction conditions such as substrate preparation and reaction buffer may also be a factor.
The thermal and pH lability of phenyl acetate has complicated the collection of kinetic data using
this substrate. Nevertheless, efforts to obtain these data are still underway.
In conclusion, AF_est1 and AF_est2 were highly thermostable and were active at a wide range of
temperatures. This is especially important for industrial enzymes, which typically experience
high temperature conditions. Both AF_est1 and AF_est2 preferred short chain pNP-esters, and
were most active on phenyl acetate. In addition, they were stable in a variety of detergents and
organic solvents, a property required for organic syntheses. Amino acid sequence analysis
revealed that AF_est1 and AF_est2 contained the putative catalytic triad Ser, Asp and His, where
Ser was located in the conserved peptapeptide Gly-X-Ser-X-Gly. Future experiments will
investigate their potential to catalyze the transesterification of simple sugars or phenolic acids
with aliphatic compounds; crystallization trials are also underway.

62
6 Conclusions and Recommendations
6.1 Summary
Carboxyl esterases and lipases display broad but distinct substrate specificities, can perform
chemo-, regio-, and stereospecific hydrolytic and synthetic reactions, and are active under mild
reaction conditions. These properties make this class of enzymes important catalysts for
synthetic transformation of highly functionalized substrates, including biomass-derived
polymers. For example, arylesterases could be used to alter the solubility of lignin and lignin-
derived phenolic acids so their antioxidant properties can be harnessed in a broader range of
products.
In the present work, a total of 202 gene targets from 15 bacterial genomes were selected (Chapter
3). Exactly 180 genes were successfully cloned and 84 of them were expressed as soluble
proteins. Forty-six proteins were active on pNP-acetate at pH 8 whereas 38 proteins did not
exhibit any activity towards all substrates. Accordingly, the high throughput platform used for
protein synthesis successfully produced roughly 50 % of the novel enzyme targets, and more
than 50 % of proteins produced in soluble form were also active on predicted substrates. Among
the 46 active proteins, 18 proteins showed arylesterase activity. Six arylesterases that were also
active on medium-chain aliphatic substrates were selected for further biochemical
characterization. Two enzymes with activity towards pNP-acetate only were also characterized
for comparison. To our best knowledge, all 6 arylesterases (RP_est1, RP_est2, RP_est3, PP_est2,
PP_est3, and PA_est1) represent the first arylesterases to be isolated and characterized from
Rhodopseudomonas palustris, Pseudomonas putida, and Pseudomonas aeuginosa, respectively.
Further, a comparative sequence analysis of active and inactive proteins did not reveal sequence
features that would allow us to improve our prediction of arylesterases. Almost all of proteins,
either active or not active, contained the residues that comprise the catalytic triad and the HG
oxyanion sequence.
In addition to general properties (substrate specificity, optimal pH, pH stability, thermal
inactivation, kinetics), enzyme stability was assessed in a variety of detergents, organic solvents,

63
and ionic liquids (Chapter 4). In particular, RP_est1 and PP_est3 which showed the best kinetics
data were stable at 50 °C and were also stable in a range of solvents.
Two archreal esterases from Archaeoglobus fulgidus were also studied along with the bacterial
esterases (Chapter 5). These enzymes displayed high temperature stability and were stable in a
variety of detergents and organic solvents, satisfying a required property for organic syntheses.
In contrast to the bacterial enzymes described in Chapter 3, these two esterases exhibited
significantly lower thermal inactivation and were also comparatively stable in both polar and
nonpolar solvents, and ionic liquids. Notably, only one other arylesterase from an archaeal
species has been reported in the published literature.
The X-ray structural determination of all 10 characterized proteins (8 bacterial esterases and 2
archaeal esterases) is in progress. In the meanwhile, the potential for the most promising
enzymes, particularly PP_est3, AF_est1 and AF_est2, to catalyze the transesterification of simple
sugars and lignin-derived phenolic acids with hydrophobic compounds will be evaluated.
6.2 Recommendations
To expand on the discoveries presented in this study, the following research objectives are
recommended:
1. Obtain kinetic parameters for all enzymes using phenyl acetate at the optimal enzyme
temperature, as well as pH.
2. Evaluate structural changes of proteins at different temperatures by Circular Dichroism
and then correlate the results of thermal inactivation to thermostability.
3. Perform additional bioinformatic analyses to compare the active and non-active enzymes
isolated in this study and further attempt to identify sequence features that are correlated
to arylesterase activity. Moreover, compare the archaeal esterases with the bacterial
esterases to predict amino acid residues that contribute to the temperature and solvent
tolerance demonstrated by AF_est1 and AF_est2.
4. Analyze solved x-ray structures of the targeted enzymes to identify sequence and
structural features that contribute to substrate specificity.

64
5. Evaluate the unusual sequence feature of RP_est1 using site-directed mutagenesis and 3D
structure determination.
6. Perform additional experiments to explain the increase in RP_est1 activity after
incubation in 1 % SDS solution.
7. Follow the published procedures by Hogbom, et al. to confirm if 10 characterized
enzymes are metalloproteins.
8. Measure the hydrolytic activity of enzymes in the presence of organic solvents.
9. Follow published procedures to evaluate the potential of PP_est3, AF_est1 and AF_est2
to catalyze the transesterification of phenolic acids, as well as simple sugars. This might
require immobilizing the enzymes onto a solid support, such as celite. Here, methods
developed by a fourth year thesis student (Ping Hay Lam) can be followed.
10. Evaluate the enantiomeric selectivity of the new arylesterases.

65
Bibliography
Chapter 1 [1] J. George, M. S. Sreekala and S. Thomas, “A review on interface modification and characterization of natural fiber reinforced plastic composites,” Polymer Engineering & Science, vol. 41, pp. 1471-1485, 2001. [2] A. Kongdee, T. Bechtold and L. Teufel, “Modification of cellulose fiber with silk sericin,” Journal of Applied Polymer Science, vol. 96, pp. 1421-1428, 2005. [3] J. Borsa, T. Toth, E. Takacs and P. Hargittai, “Radiation modification of swollen and chemically modified cellulose,” Radiation Physics and Chemistry, vol. 67, pp. 509-512, 2003. [4] H. Brumer III, Q. Zhou, M. J. Baumann, T. T. Teeri and K. Carlsson, “Activation of crystalline cellulose surfaces through the chemoenzymatic modification of xyloglucan,” Journal of the American Chemical Society, vol. 126, pp. 5715-5721, 2004. [5] B. C. Saha and R. J. Bothast, “Enzymes in lignocellulosic biomass conversion,” Fuel and Energy Abstracts, vol. 39, pp. 36, 1998. [6] R. L. Howard, E. Abotsi, E. L. Jansen van Rensburg and S. Howard, “Lignocellulose biotechnology: issues of biconversion and enzyme production,” African Journal of Biotechnology, vol. 2, pp. 602-619, 2003. [7] J. Laine, T. Lindstrom, G. G. Nordmark and G. Risinger, “Studies on topochemical modification of cellulosic fibres – Part 2. The effect of carboxymethyl cellulose attachment on fibre swelling and paper strength,” Nordic Pulp & Paper Research Journal, col. 17, pp. 50-56, 2002. [8] F. Lenzi, A. Sannino, A. Borriello, F. Porro, D. Capitani and G. Mensitieri, “Probing the degree of crosslinking of a cellulose based superabsorbing hydrogel through traditional and NMR techniques,” Polymer, vol. 44, pp. 1577-1588, 2003. [9] J. F. Kennedy and M.Thorley, “Organic Coatings: Science and Technology: 2nd ed.; Z.W. Wicks, Jr., F.N. Jones, S.P. Pappas; Wiley, New York, 1999,” Carbohydrate Polymers, vol. 41, pp. 427, 2000. [10] T. T. Teeri, H. Brumer, G. Daniel and P. Gatenholm, “Biomimetic engineering of cellulose-based materials,” Trends in Biotechnology, vol. 25, pp. 299-306, 2007. [11] Q. Zhou, M. W. Rutland, T. T. Teeri and H. Brumer, “Xyloglucan in cellulose modification,” Cellulose, vol. 14, pp. 625-641, 2007. [12] M. T. Gustavsson, P. V. Persson, T. Iversen, M. Martinelle, K. Hult, T. T. Teeri and H. Brumer, “Modification of cellulose fiber surfaces by use of a lipase and a xyloglucan endotransglycosylase,” Biomacromolecules, vol. 6, pp. 196-203, 2005.
Chapter 2 [1] R. L. Howard, E. Abotsi, E. L. Jansen van Rensburg and S. Howard, “Lignocellulose biotechnology: issues of biconversion and enzyme production,” African Journal of Biotechnology, vol. 2, pp. 602-619, 2003. [2] S. Malherbe and T. E. Cloete, “Lignocellulose biodegradation: Fundamentals and applications," Reviews in Environmental Science and Biotechnology, vol. 1, pp. 105-114, 2002.

66
[3] N. Carpita and M. McCann, “The cell wall,” in Biochemistry and Molecular Biology of Plants B. Buchanan, W. Gruissem and R. Jones, Eds. Rockville, MD: American Society of Plant Physiologists, pp. 52-108, 2000 [4] B. C. Saha, “Hemicellulose bioconversion,” Journal of Industrial Microbiology and Biotechnology, vol. 30, pp. 279-291, 2003. [5] G. O. Aspinall, “Chemistry of cell wall polysaccharides,” Biochemistry of Plants : A Comprehensive Treatise, vol. 3, pp. 473-500, 1980. [6] L. R. S. Moreira and E. X. F. Filho, “An overview of mannan structure and mannan-degrading enzyme systems,” Applied Microbiology and Biotechnology, vol. 79, pp. 165-178, 2008. [7] Q. Zhou, M. W. Rutland, T. T. Teeri and H. Brumer, “Xyloglucan in cellulose modification,” Cellulose, vol. 14, pp. 625-641, 2007. [8] T. T. Teeri, H. Brumer, G. Daniel and P. Gatenholm, “Biomimetic engineering of cellulose-based materials,” Trends in Biotechnology, vol. 25, pp. 299-306, 2007. [9] W. Boerjan, J. Ralph and M. Baucher, “Lignin Biosynthesis,” Annual Review of Plant Biology, vol. 54, pp. 519-546, 2003. [10] M. K. Bhat, “Cellulases and related enzymes in biotechnology,” Biotechnology Advances, vol. 18, pp. 355-383, 2000. [11] M. T. Gustavsson, P. V. Persson, T. Iversen, M. Martinelle, K. Hult, T. T. Teeri and H. Brumer, “Modification of cellulose fiber surfaces by use of a lipase and a xyloglucan endotransglycosylase,” Biomacromolecules, vol. 6, pp. 196-203, 2005. [12] A. R. M. Yahya, W. A. Anderson and M. Moo-Young, “Ester synthesis in lipase-catalyzed reactions,” Enzyme and Microbial Technology, vol. 23, pp. 438-450, 1998. [13] D. H. G. Crout and VG. Vic, “Glycosidases and glycosyl transferases in glycoside and oligosaccharide synthesis,” Current Opinion in Chemical Biology, vol. 2, pp. 98-111, 1998. [14] A. Trincone and A. Glordano, “Glycosyl hydrolases and glycosyltransferases in the synthesis of oligosaccharides,” Current Organic Chemistry, vol. 10, pp. 1163-1193, 2006. [15] J. Perez, J. Munoz-Dorado, T. de la Rubia and J. Martinez, “Biodegradation and biological treatments of cellulose, hemicellulose and lignin: an overview,” International Microbiology, vol. 5, pp. 53-63, 2002. [16] M. Tuomela, M. Vikman, A. Hatakka and M. Itavaara, “Biodegradation of lignin in a compost environment: a review,” Bioresource Technology, vol. 72, pp. 169-183, 2000. [17] L. R. C. Barclay, F. Xi and J. Q. Norris, “Antioxidant properties of phenolic lignin model compounds,” Journal of Wood Chemistry and Technology, vol. 17, pp. 73-90, 1997. [18] E. Topakas, H. Stamatis, M. Mastihubova, P. Biely, D. Kekos, B. J. Macris and P. Christakopoulos, “Purification and characterization of a Fusarium oxysporum feruloyl esterase (FoFAE-I) catalyzing transesterification of phenolic acid esters,” Enzyme and Microbial Technology, vol. 33, pp. 729-737, 2003. [19] U. T. Bornscheuer, “Microbial carboxyl esterases: classification, properties and application in biocatalysis,” FEMS Microbiology Reviews, vol. 26, pp. 73-81, 2002. [20] P. Fojan, P. H. Jonson, M. T. N. Petersen and S. B. Petersen, “What distinguishes an esterase from a lipase: A novel structural approach,” Biochimie, vol. 82, pp. 1033-1041, 2000. [21] M. T. Neves Petersen, P. Fojan and S. B. Petersen, “How do lipases and esterases work: the electrostatic contribution,” Journal of Biotechnology, vol. 85, pp. 115-147, 2001. [22] J. Polaina and A. P. MacCabe, “Industrial enzymes: structure, function and applications / edited by Julio Polaina and Andrew P. MacCabe,” 2007.

67
[23] K. K. Kim, H. K. Song, D. H. Shin, K. Y. Hwang and S. W. Suh, “The crystal structure of a triacylglycerol lipase from Pseudomonas cepacia reveals a highly open conformation in the absence of a bound inhibitor,” Structure, vol. 5, pp. 173-185, 1997. [24] J. L. Arpigny and K. E. Jaeger, “Bacterial lipolytic enzymes: classification and properties,” The Biochemical Journal, vol. 343, pp. 177-183. 1999. [25] U. T. Bornscheuer, C. Bessler, R. Srinivas and S. Hari Krishna, “Optimizing lipases and related enzymes for efficient application,” Trends in Biotechnology, vol. 20, pp. 433-437, 2002. [26] S. B. Kim, W. Lee and Y. W. Ryu, “Cloning and characterization of thermostable esterase from Archaeoglobus fulgidus,” Journal of Microbiology, vol. 46, pp. 100-107, 2008. [27] Jaeger karl-erich and Reetz Manfred T., “Microbial lipases form versatile tools for biotechnology,” Trends in Biotechnology, vol. 16, pp. 396-403, 1998. [28] F. X. Malcata and North Atlantic Treaty Organization. Scientific Affairs Division., “Engineering of/with lipases / edited by F. Xavier Malcata,” 1996. [29] D. Shen, J. Xu, H. Wu and Y. Liu, “Significantly improved esterase activity of Trichosporon brassicae cells for ketoprofen resolution by 2-propanol treatment,” Journal of Molecular Catalysts B, vol. 18, pp. 219-224, 2002. [30] W. J. Quax and C. P. Broekhuizen, “Development of a new Bacillus carboxyl esterase for use in the resolution of chiral drugs,” Applied Microbiology and Biotechnology, vol. 41, pp. 425-431, 1994. [31] T. Panda and B. S. Gowrishankar, “Production and applications of esterases,” Applied Microbiology and Biotechnology, vol. 67, pp. 160-169, 2005. [32] R. Sharma, Y. Chisti and U. C. Banerjee, “Production, purification, characterization, and applications of lipases,” Biotechnology Advances, vol. 19, pp. 627-662, 2001. [33] G. T. Howard, B. Crother and J. Vicknair, “Cloning, nucleotide sequencing and characterization of a polyurethanase gene (pueB) from Pseudomonas chlororaphis,” International Biodeterioration & Biodegradation, vol. 47, pp. 141-149, 2001. [34] R. Jahangir, C. B. McCloskey, W. G. Mc Clung, R. S. Labow, J. L. Brash and J. P. Santerre, “The influence of protein adsorption and surface modifying macromolecules on the hydrolytic degradation of a poly(ether–urethane) by cholesterol esterase,” Biomaterials, vol. 24, pp. January, 2003. [35] E. Santaniello, P. Ferraboschi and P. Grisenti, “Lipase-catalyzed transesterification in organic solvents: Applications to the preparation of enantiomerically pure compounds,” Enzyme and Microbial Technology, vol. 15, pp. 367-382, 1993. [36] M. N. Gupta, “Enzyme function in organic solvents,” European Journal of Biochemistry, vol. 203, pp. 25-32, 1992. [37] C. Laane, S. Boeren, K. Vos and C. Veeger, “Rules for Optimization of Biocatalysis in Organic Solvents,” Biotechnology and Bioengineering, vol. 30, pp. 81-87, 1987. [38] G. M. Gubitz and A. C. Paulo, “New substrates for reliable enzymes: enzymatic modification of polymers,” Current Opinion in Biotechnology, vol. 14, pp. 577-582, 2003. [39] C. J. Hamilton, “Enzymes in preparative mono- and oligo-saccharide synthesis,” Natural Product Reports, vol. 21, pp. 365-385, 2004. [40] C. Vafiadi, E. Topakas, V. R. Nahmias, C. B. Faulds and P. Christakopoulos, “Feruloyl esterase-catalysed synthesis of glycerol sinapate using ionic liquids mixtures,” Journal of Biotechnology, vol. 139, pp. 124-129, 2009. [41] A. J. Ragauskas, C. K. Williams, B. H. Davison, G. Britovsek, J. Cairney, C. A. Eckert, W. J. Frederick Jr, J. P. Hallett, D. J. Leak, C. L. Liotta, J. R. Mielenz, R. Murphy, R. Templer and

68
T. Tschaplinski, “The path forward for biofuels and biomaterials,” Science (Wash. ), vol. 311, pp. 484-489, 2006.
Chapter 3 [1] C. Vafiadi, E. Topakas and P. Christakopoulos, "Regioselective esterase-catalyzed feruloylation of l-arabinobiose," Carbohydrate Research, vol. 341, pp. 1992-1997, 2006. [2] E. Topakas, C. Vafiadi and P. Christakopoulos, "Microbial production, characterization and applications of feruloyl esterases," Process Biochemistry, vol. 42, pp. 497-509, 2007. [3] F. J. Plou, M. A. Cruces, M. Ferrer, G. Fuentes, E. Pastor, M. Bernabé, M. Christensen, F. Comelles, J. L. Parra and A. Ballesteros, "Enzymatic acylation of di- and trisaccharides with fatty acids: choosing the appropriate enzyme, support and solvent," Journal of Biotechnology, vol. 96, pp. 55-66, 2002. [4] V. Dossat, D. Combes and A. Marty, "Lipase-catalysed transesterification of high oleic sunflower oil," Enzyme and Microbial Technology, vol. 30, pp. 90-94, 2002. [5] C. Vafiadi, E. Topakas, V. R. Nahmias, C. B. Faulds and P. Christakopoulos, "Feruloyl esterase-catalysed synthesis of glycerol sinapate using ionic liquids mixtures," Journal of Biotechnology, vol. 139, pp. 124-129, 2009. [6] Evangelos Topakas, Haralambos Stamatis, Maria Mastihubova, Peter Biely, Dimitris Kekos, Basil J. Macris and Paul Christakopoulos, "Purification and characterization of a Fusarium oxysporum feruloyl esterase (FoFAE-I) catalysing transesterification of phenolic acid esters," Enzyme and Microbial Technology, vol. 33, pp. 729-737, 2003. [7] A. J. Ragauskas, C. K. Williams, B. H. Davison, G. Britovsek, J. Cairney, C. A. Eckert, W. J. Frederick Jr, J. P. Hallett, D. J. Leak, C. L. Liotta, J. R. Mielenz, R. Murphy, R. Templer and T. Tschaplinski, "The path forward for biofuels and biomaterials," Science (Wash. ), vol. 311, pp. 484-489, 2006. [8] S. F. Altschul, T. L. Madden, A. A. Schaeffer, J. Zhang, Z. Zhang, W. Miller and D. J. Lipman, "Gapped BLAST and PSI-BLAST: A new generation of protein database search programs," Nucleic Acids Research, vol. 25, pp. 3389-3402, 1997. [9] M. L. Green and P. D. Karp, "A Bayesian method for identifying missing enzymes in predicted metabolic pathway databases," BMC Bioinformatics, vol. 5, pp. 76, 2004. [10] P. Liang, B. Labedan and M. Riley, "Physiological genomics of Escherichia coli protein families," Physiological Genomics, vol. 9, pp. 15-26, 2002. [11] T. Ryan Gregory, Rob DeSalle, "Comparative genomics in prokaryotes," in The Evolution of the Genome T. R. Gregory, Ed. Elsevier Academic Press, Amsterdam, Boston etc, 2005, pp. 585-675. [12] S. Brachat, F. S. Dietrich, S. Voegeli, Z. Zhang, L. Stuart, A. Lerch, K. Gates, T. Gaffney and P. Philippsen, "Reinvestigation of the Saccharomyces cerevisiae genome annotation by comparison to the genome of a related fungus: Ashbya gossypii," Genome Biology, vol. 4, pp. R45, 2003. [13] A. Filipski and S. Kumar, "Comparative genomics in eukaryotes," in The Evolution of the Genome T. R. Gregory, Ed. Elsevier Academic Press, Amsterdam, Boston etc, 2005, pp. 521-583. [14] M. Kellis, N. Patterson, M. Endrizzi, B. Birren and E. S. Lander, "Sequencing and comparison of yeast species to identify genes and regulatory elements," Nature, vol. 423, pp. 241-254, 2003.

69
[15] L. D. Stein, Z. R. Bao, D. Blasiar, T. Blumenthal, M. R. Brent, N. S. Chen, A. Chinwalla, L. Clarke, C. Clee, A. Coghlan, A. Coulson, P. D'Eustachio, D. H. A. Fitch, L. A. Fulton, R. E. Fulton, S. Griffiths-Jones, T. W. Harris, L. W. Hillier, R. Kamath, P. E. Kuwabara, E. R. Mardis, M. A. Marra, T. L. Miner, P. Minx, J. C. Mullikin, R. W. Plumb, J. Rogers, J. E. Schein, M. Sohrmann, J. Spieth, J. E. Stajich, C. C. Wei, D. Willey, R. K. Wilson, R. Durbin and R. H. Waterston, "The genome sequence of Caenorhabditis briggsae: A platform for comparative genomics," Public Library of Science Biology, vol. 1, pp. e45, 2003. [16] E. M. Phizicky and S. Fields, "Protein-protein interactions: methods for detection and analysis," Microbiology Review, vol. 59, pp. 94-123, 1995. [17] P. Uetz, L. Giot and G. Cagney, "A comprehensive analysis of protein-protein interactions in Saccharomyces cerevisiae," Nature, vol. 403, pp. 623-627, 2000. [18] D. Plewczynski and K. Ginalski, "The interactome: predicting the protein-protein interactions in cells," Cellular & Molecular Biology Letters, vol. 14, pp. 1-22, 2009. [19] P. F. Predki, "Functional protein microarrays: ripe for discovery," Current Opinion in Chemical Biology, vol. 8, pp. 8-13, 2004. [20] D. Stoll, M. Templin, J. Bachmann and T.O. Joos, "Protein microarrays: applications and future challenges," Current Opinion in Drug Discovery and Development, vol. 8, pp. 239-252, 2005. [21] H. Zhu and M. Snyder, "Protein arrays and microarrays," Current Opinion in Chemical Biology, vol. 5, pp. 40-45, 2001. [22] P. O. Brown and D. Botstein, "Exploring the new world of the genome with DNA microarrays," Nature Genetics, vol. 21, pp. 33-37, 1999. [23] M. Norin and M. Sundström, "Structural proteomics: developments in structure-to-function predictions," Trends in Biotechnology, vol. 20, pp. 79-84, 2002. [24] C. Zhang and S. H. Kim, "Overview of structural genomics: from structure to function," Current Opinion in Chemical Biology, vol. 7, pp. 28-32, 2003. [25] A. F. Yakunin, A. A. Yee, A. Savchenko, A. M. Edwards and C. H. Arrowsmith, "Structural proteomics: a tool for genome annotation," Current Opinion in Chemical Biology, vol. 8, pp. 42-48, 2004. [26] E. Kuznetsova, M. Proudfoot, S. A. Sanders, J. Reinking, A. Savchenko, C. H. Arrowsmith, A. M. Edwards and A. F. Yakunin, "Enzyme genomics: Application of general enzymatic screens to discover new enzymes," FEMS Microbiology Review, vol. 29, pp. 263-279, 2005. [27] H. Kwoun Kim, Y. Jung, W. Choi, H. S. Ryu, T. Oh and J. Lee, "Sequence-based approach to finding functional lipases from microbial genome databases," FEMS Microbiology Letter, vol. 235, pp. 349-355, 2004. [28] F. Baneyx, "Recombinant protein expression in Escherichia coli," Current Opinion in Biotechnology, vol. 10, pp. 411-421, 1999. [29] H. P. Sorensen and K. K. Mortensen, "Advanced genetic strategies for recombinant protein expression in Escherichia coli," Journal of Biotechnology, vol. 115, pp. 113-128, 2005. [30] P. Fojan, P. H. Jonson, M. T. N. Petersen and S. B. Petersen, "What distinguishes an esterase from a lipase: A novel structural approach," Biochimie, vol. 82, pp. 1033-1041, 2000. [31] F. W. Studier, "Protein production by auto-induction in high density shaking cultures," Protein Expression and Purification, vol. 41, pp. 207-234, 2005. [32] S. K. Ghatora, B. S. Chadha, H. S. Saini, M. K. Bhat and C. B. Faulds, "Diversity of plant cell wall esterases in thermophilic and thermotolerant fungi," Journal of Biotechnology, vol. 125, pp. 434-445, 2006.

70
[33] I. Martinez-Martinez, S. Montoro-Garcia, J. D. Lozada-Ramirez, A. Sanchez-Ferrer and F. Garcia-Carmona, "A colorimetric assay for the determination of acetyl xylan esterase or cephalosporin C acetyl esterase activities using 7-amino cephalosporanic acid, cephalosporin C, or acetylated xylan as substrate," Analytical Biochemistry, vol. 369, pp. 210-217, 2007. [34] A. Kosugi, K. Murashima and R. H. Doi, "Xylanase and acetyl xylan esterase activities of XynA, a key subunit of the Clostridium cellulovorans cellulosome for xylan degradation," Applied Environmental Microbiology, vol. 68, pp. 6399-6402, 2002. [35] U. T. Bornscheuer, "Microbial carboxyl esterases: classification, properties and application in biocatalysis," FEMS Microbiology Review, vol. 26, pp. 73-81, 2002. [36] K. M. Fenster, K. L. Parkin and J. L. Steele, "Nucleotide sequencing, purification, and biochemical properties of an arylesterase from Lactobacillus casei LILA," Journal of Dairy Science, vol. 86, pp. 2547-2557, 2003. [37] K. M. Fenster, K. L. Parkin and J. L. Steele, "Characterization of an arylesterase from Lactobacillus helveticus CNRZ32," Journal of Applied Microbiology, vol. 88, pp. 572-583, 2000. [38] J F Shaw, R C Chang, K H Chuang, Y T Yen, Y J Wang and and F G Wang, "Nucleotide sequence of a novel arylesterase gene from Vibro mimicus and characterization of the enzyme expressed in Escherichia coli." Biochemical Journal, vol. 298, pp. 675-680, 1994. [39] M. T. Neves Petersen, P. Fojan and S. B. Petersen, "How do lipases and esterases work: the electrostatic contribution," Journal of Biotechnology, vol. 85, pp. 115-147, 2001. [40] Bosmann HB, "Membrane marker enzymes. Characterization of an arylesterase of guinea pig cerebral cortex utilizing p-nitrophenyl acetate as substrate," Biochimica et Biophysica Acta, vol. 276, pp. 180-191, 1972. [41] Ecobichon DJ, "Characterization of the esterases of feline serum," Canadian Journal of Biochemistry, vol. 52, pp. 1073-1078, 1974. [42] H. Chahinian, L. Nini, E. Boitard, J. P. Dubes, L. C. Comeau and L. Sarda, "Distinction between esterases and lipases: a kinetic study with vinyl esters and TAG," Lipids, vol. 37, pp. 653-662, 2002. [43] K. Jaeger, S. Ransac, B. W. Dijkstra, C. Colson, M. Van Heuvel and O. Misset, "Bacterial lipases," FEMS Microbiology Review, vol. 15, pp. 29-63, 1994. [44] J. D. Cheeseman, A. Tocilj, S. Park, J. D. Schrag and R. J. Kazlauskas, "Structure of an aryl esterase from Pseudomonas fluorescens," Acta Crystallographica, vol. 60, pp. 1237-1243, 2004.
Chapter 4 [1] H. Ogino and H. Ishikawa, "Enzymes which are stable in the presence of organic solvents," Journal of Bioscience and Bioengineering, vol. 91, pp. 109-116, 2001. [2] B. Zentgraf and T. J. Ahern, "Practical importance of enzyme stability," Pure & Applied Chemistry, vol. 63, pp. 1527-1540, 1991. [3] K. M. Polizzi, A. S. Bommarius, J. M. Broering and J. F. Chaparro-Riggers, "Stability of biocatalysts," Current Opinion In Chemical Biology, vol. 11, pp. 220-225, 2007. [4] V. G. H. Eijsink, S. Gaseidnes, T. V. Borchert and B. van den Burg, "Directed evolution of enzyme stability," Biomolecular Engineering, vol. 22, pp. 21-30, 2005. [5] U. Hanefeld, L. Gardossi and E. Magner, "Understanding enzyme immobilisation," Chemical Society Reviews, vol. 38, pp. 453-468, 2009.

71
[6] C. Mateo, J. M. Palomo, G. Fernandez-Lorente, J. M. Guisan and R. Fernandez-Lafuente, "Improvement of enzyme activity, stability and selectivity via immobilization techniques," Enzyme and Microbial Technology, vol. 40, pp. 1451-1463, 2007. [7] G. N. Somero, "Proteins and temperature," Annual Review of Physiology, vol. 57, pp. 43-68, 1995. [8] E. Kuznetsova, M. Proudfoot, S. A. Sanders, J. Reinking, A. Savchenko, C. H. Arrowsmith, A. M. Edwards and A. F. Yakunin, "Enzyme genomics: Application of general enzymatic screens to discover new enzymes," FEMS Microbiology Review, vol. 29, pp. 263-279, 2005. [9] H. Kwoun Kim, Y. Jung, W. Choi, H. S. Ryu, T. Oh and J. Lee, "Sequence-based approach to finding functional lipases from microbial genome databases," FEMS Microbiology Letter, vol. 235, pp. 349-355, 2004. [10] M. Proudfoot, E. Kuznetsova, S. A. Sanders, C. F. Gonzalez, G. Brown, A. M. Edwards, C. H. Arrowsmith and A. F. Yakunin, "High throughput screening of purified proteins for enzymatic activity," Methods in Molecular Biology (Clifton, N. J. ), vol. 426, pp. 331-341, 2008. [11] V. G. H. Eijsink, A. Bjork, S. Gaseidnes, R. Sirevag, B. Synstad, B. v. d. Burg and G. Vriend, "Rational engineering of enzyme stability," Journal of Biotechnology, vol. 113, pp. 105-120, 2004. [12] U. T. Bornscheuer, "Immobilizing enzymes: How to create more suitable biocatalysts," Angewandte Chemie International Edition, vol. 42, pp. 3336-3337, 2003. [13] H. Ogino, K. Miyamoto, M. Yasuda, K. Ishimi and H. Ishikawa, "Growth of organic solvent-tolerant Pseudomonas aeruginosa LST-03 in the presence of various organic solvents and production of lipolytic enzyme in the presence of cyclohexane," Biochemical Engineering Journal, vol. 4, pp. 1-6, 1999. [14] H. Ogino, K. Miyamoto and H. Ishikawa, "Organic-solvent-tolerant bacterium which secretes organic-solvent-stable lipolytic enzyme," Applied and Environmental Microbiology, vol. 60, pp. 3884-3886, 1994. [15] M. N. Gupta, "Enzyme function in organic solvents," European Journal of Biochemistry, vol. 203, pp. 25-32, 1992. [16] K. W. Kim, B. Song, M. Y. Choi and M. J. Kim, "Biocatalysis in ionic liquids: Markedly enhanced enantioselectivity of lipase," Organic Letters, vol. 3, pp. 1507-1509, 2001. [17] R. Madeira Lau, F. van Rantwijk, K. R. Seddon and R. A. Sheldon, "Lipase-catalyzed reactions in ionic liquids," Organic Letters, vol. 2, pp. 4189-4191, 2000. [18] T. Itoh, E. Akasaki, K. Kudo and S. Shirakami, "Lipase-catalyzed enantioselective acylation in the ionic liquid solvent system: Reaction of enzyme anchored to the solvent," Chemistry Letters, vol. 30, pp. 262, 2001. [19] Nicole Kaftzik, Peter Wasserscheid and Udo Kragl, "Use of Ionic Liquids to Increase the Yield and Enzyme Stability in the β-Galactosidase Catalysed Synthesis of N-Acetyllactosamine," Organic Process Research & Development, vol. 6, pp. 553-557, 2002. [20] F. van Rantwijk, R. Madeira Lau and R. A. Sheldon, "Biocatalytic transformations in ionic liquids," Trends in Biotechnology, vol. 21, pp. 131-138, 2003. [21] A. P. de los Rios, F. J. Hernandez-Fernandez, F. Tomas-Alonso, D. Gomez and G. Villora, "Synthesis of esters in ionic liquids - The effect of vinyl esters and alcohols," Process Biochemistry, vol. 43, pp. 892-895, 2008. [22] U. Kragl, M. Eckstein and N. Kaftzik, "Enzyme catalysis in ionic liquids," Current Opinion In Biotechnology, vol. 13, pp. 565-571, 2002.

72
[23] S. Park and R. J. Kazlauskas, "Improved preparation and use of room-temperature ionic liquids in lipase-catalyzed enantio- and regioselective acylations," Journal of Organic Chemistry, vol. 66, pp. 8395-8401, 2001. [24] F. J. Plou, M. A. Cruces, M. Ferrer, G. Fuentes, E. Pastor, M. Bernabé, M. Christensen, F. Comelles, J. L. Parra and A. Ballesteros, "Enzymatic acylation of di- and trisaccharides with fatty acids: choosing the appropriate enzyme, support and solvent," Journal of Biotechnology, vol. 96, pp. 55-66, 2002. [25] J. L. Kaar, A. M. Jesionowski, J. A. Berberich, R. Moulton and A. J. Russell, "Impact of ionic liquid physical properties on lipase activity and stability," Journal of the American Chemical Society, vol. 125, pp. 4125-4131, 2003. [26] F. Ganske and U. T. Bornscheuer, "Lipase-catalyzed glucose fatty acid ester synthesis in ionic liquids," Journal of biotechnology, vol. 133, pp. 486-489, 2005. [27] E. Topakas, D. Kekos, B. J. Macris, P. Christakopoulos, H. Stamatis, M. Mastihubova and P. Biely, "Purification and characterization of a Fusarium oxysporum feruloyl esterase (FoFAE-I) catalysing transesterification of phenolic acid esters," Enzyme and Microbial Technology, vol. 33, pp. 729-737, 2003. [28] C. Vafiadi, E. Topakas, V. R. Nahmias, C. B. Faulds and P. Christakopoulos, "Feruloyl esterase-catalysed synthesis of glycerol sinapate using ionic liquids mixtures," Journal of Biotechnology, vol. 139, pp. 124-129, 2009. [29] Y. H. P. Zhang, "Reviving the carbohydrate economy via multi-product lignocellulose biorefineries," Journal of Industrial Microbiology and Biotechnology, vol. 35, pp. 367-375, 2008. [30] Y. J. Park, S. J. Yoon and H. B. Lee, "A novel thermostable arylesterase from the archaeon Sulfolobus solfataricus P1: Purification, Characterization, and Expression," Journal of Bacteriology, vol. 190, pp. 8086-8095, 2008. [31] C. M. L. Carvalho and J. M. S. Cabral, "Reverse micelles as reaction media for lipases," Biochimie, vol. 82, pp. 1063-1085, 2000. [32] M. B. Nthangeni, H. G. Patterton, A. van Tonder, W. P. Vergeer and D. Litthauer, "Over-expression and properties of a purified recombinant Bacillus licheniformis lipase: a comparative report on Bacillus lipases," Enzyme and Microbial Technology, vol. 28, pp. 705-712, 2001. [33] I. Karadzic, A. Masui, L. I. Zivkovic and N. Fujiwara, "Purification and characterization of an alkaline lipase from Pseudomonas aeruginosa isolated from putrid mineral cutting oil as component of metalworking fluid," Journal of Bioscience and Bioengineering, vol.102, pp. 82-89, 2006. [34] M. Hogbom, U. B. Ericsson, R. Lam, M. A. Bakali, E. Kuznetsova, P. Nordlund and D. B. Zamble, "A high throughput method for the detection of metalloproteins on a microgram scale," Molecular and Cellular Proteomics, vol. 4, pp. 827-834, 2005. [35] K. Nakamura, M. Kinoshita and A. Ohno, "Structure of solvent affects enantioselectivity of lipase-catalyzed transesterification," Tetrahedron, vol. 51, pp. 8799-8808, 1995. [36] M. Kinoshita and A. Ohno, "Factors influencing enantioselectivity of lipase-catalyzed hydrolysis," Tetrahedron, vol. 52, pp. 5397-5406, 1996. [37] M. J. Haas, D. J. Cichowicz, W. Jun and K. Scott, "The enzymatic hydrolysis of triglyceride-phospholipid mixtures in an organic solvent," Journal of the American Oil Chemists’ Society, vol. 72, pp. 519-525, 1995. [38] Z. Yang and W. Pan, "Ionic liquids: Green solvents for nonaqueous biocatalysis," Enzyme and Microbial Technology, vol. 37, pp. 19-28, 2005.

73
[39] A. Patist, S. S. Bhagwat, K. W. Penfield, P. Aikens and D. O. Shah, "On the measurement of critical micelle concentrations of pure and technical-grade nonionic surfactants," Journal of Surfactants and Detergents, vol. 3, pp. 53-58, 2000. [40] J. C. Jacquier and P. L. Desbène, "Determination of critical micelle concentration by capillary electrophoresis. Theoretical approach and validation," Journal of Chromatography A, vol. 718, pp. 167-175, 1995.
Chapter 5 [1] K. Egorova and G. Antranikian, "Industrial relevance of thermophilic Archaea," Current Opinion in Microbiology, vol. 8, pp. 648-655, 2005. [2] C. Vieille and G. J. Zeikus, "Hyperthermophilic enzymes: sources, uses, and molecular mechanisms for thermostability," Microbiology and Molecular Biology Reviews, vol. 65, pp. 1-43, 2001. [3] J. Eichler, "Biotechnological uses of archaeal extremozymes," Biotechnology Advances, Vol. 19, pp. 261-278, 2001. [4] U. T. Bornscheuer, "Microbial carboxyl esterases: classification, properties and application in biocatalysis," FEMS Microbiology Review, vol. 26, pp. 73-81, 2002. [5] Y. J. Park, S. J. Yoon and H. B. Lee, "A novel thermostable arylesterase from the archaeon Sulfolobus solfataricus P1: Purification, characterization, and expression," Journal of Bacteriology, vol. 190, pp. 8086-8095, 2008. [6] S. B. Kim, W. Lee and Y. W. Ryu, "Cloning and characterization of thermostable esterase from Archaeoglobus fulgidus," Journal of Microbiology, vol. 46, pp. 100-107, 2008. [7] S. Yair, B. Ofer, E. Arik, S. Shai, R. Yossi, D. Tzvika, and K. Amir, “Organophosphate degrading microorganisms and enzymes as biocatalysts in environmental and personal decontamination applications,” Critical Reviews in Biotechnology, vol. 28, pp. 265-275, 2008. [8] F. Marchegiani, M. Marra, F. Olivieri, M. Cardelli, R. W. James, M. Boemi and C. Franceschi, “Paraoxonase 1: Genetics and activities during aging,” Rejuvenation Research, vol. 11, pp. 113-127, 2008. [9] M. Mackness, H. M. Thompson, A. R. Hardy and C. H. Walker, “Distinction between ‘A’-esterases and arylesterases: Implication for esterase classification,” Biochemical Journal, vol. 245, pp. 293-296, 1987. [10] G. De Simone, V. Menchise, G. Manco, L. Mandrich, N. Sorrentino, D. Lang, M. Rossi and C. Pedone, "The crystal structure of a hyper-thermophilic carboxylesterase from the archaeon Archaeoglobus fulgidus " Journal of Molecular Biology, vol. 314, pp. 507-518, 2001. [11] C. M. L. Carvalho and J. M. S. Cabral, "Reverse micelles as reaction media for lipases," Biochimie, vol. 82, pp. 1063-1085, 2000. [12] G. A. Sellek and J. B. Chaudhuri, "Biocatalysis in organic media using enzymes from extremophiles," Enzyme and Microbial Technology, vol. 25, pp. 471-482, 1999. [13] P. Trodler and J. Pleiss, "Modeling structure and flexibility of Candida antarctica lipase B in organic solvents," BMC Structural Biology, vol. 8, pp. 9, 2008 [14] T. Itoh, E. Akasaki, K. Kudo and S. Shirakami, "Lipase-catalyzed enantioselective acylation in the ionic liquid solvent system: Reaction of enzyme anchored to the solvent," Chemistry Letters, vol. 30, pp. 262, 2001. [15] R. Madeira Lau, F. van Rantwijk, K. R. Seddon and R. A. Sheldon, "Lipase-catalyzed reactions in ionic liquids," Organic Letters, vol. 2, pp. 4189-4191, 2000.

74
[16] K. W. Kim, B. Song, M. Y. Choi and M. J. Kim, "Biocatalysis in ionic liquids: Markedly enhanced enantioselectivity of lipase," Organic Letters, vol. 3, pp. 1507-1509, 2001. [17] Y. Abe, K. Kude, S. Hayase, M. Kawatsura, K. Tsunashima and T. Itoh, "Design of phosphonium ionic liquids for lipase-catalyzed transesterification," Journal of Molecular Catalysis B: Enzymatic, vol. 51, pp. 81-85, 2008. [18] M. Giuseppe, D. G. Spartaco, D. R. Mario and R. Mosè, "Purification and characterization of a thermostable carboxylesterase from the thermoacidophilic eubacterium Bacillus acidocaldarius," European Journal of Biochemistry, vol. 221, pp. 965-972, 2005. [19] K. Ejima, J. Liu, Y. Oshima, K. Hirooka, S. Shimanuki, Y. Yokota, H. Hemmi, T. Nakayama and T. Nishino, "Molecular cloning and characterization of a thermostable carboxylesterase from an archaeon, Sulfolobus shibatae DSM5389: Non-linear kinetic behavior of a hormone-sensitive lipase family enzyme," Journal of Bioscience and Bioengineering, vol. 98, pp. 445-451, 2004. [20] A. Patist, S. S. Bhagwat, K. W. Penfield, P. Aikens and D. O. Shah, "On the measurement of critical micelle concentrations of pure and technical-grade nonionic surfactants," Journal of Surfactants and Detergents, vol. 3, pp. 53-58, 2000. [21] J. C. Jacquier and P. L. Desbène, "Determination of critical micelle concentration by capillary electrophoresis. Theoretical approach and validation," Journal of Chromatography A, vol. 718, pp. 167-175, 1995.

75
Appendices
Appendix A.1 Amino acid sequence alignment of proteins that were active on phenyl acetate.
RPA0348 ------------------------------------------------------------ RPA4646 ------------------------------------------------------------ PA2949 -----------------------------------------------MKRFLLGLVLLLA SCO3439 -----------------------------------------------------MGGLGAG SCO3566 ------------------------------------------------------MTGSST PA2934 -----------------------------------------------------MILDRLC SCO7440 -----------------------------------------------------------M RHA02607 ------------------------------------------------------------ RHA00320 ------------------------------------------------------------ SCO3690 ------------------------------------------------------------ NE2298 ------------------------------------------------------------ RHA01401 ------------------------------------------------------------ PP5253 ------------------------------------------------------------ PSPPH4522 MDKYNHAQAKARFDPKELIPLCPASSVPAGKYDEPMFQKITPEQYAHSSRLNYAKRQLPE PSPPH3482 ------------------------------------------------------------ RPA3430 ------------------------------------------------------------ PP3645 ------------------------------------------------------------ RHA08081 ------------------------------------------------------------ RPA0348 --------------MLAQGQRFEGLLCGDDSSRCEVVEMAKLTQFLISGACALLITATSA RPA4646 ----------------------------MSEAVMAGAEAFAFDGG---KTGVLLVHGFTG PA2949 VAAGVLYFVPATLLASVRTVERGLAGLSEHSVQVDNLEIAYLEGGSEKNPTLLLIHGFGA SCO3439 GALAGLGWAYADGGPLVADGKPPRAPAHEGAPLRDLPEVRSREGR--PEHTLTVATTRPV SCO3566 SPGQSPSPQHPNPTSVVRLGIPGGPEVTHRDVAANGARFHIAELG--DGPLVLLLHGFPQ PA2934 RGLLAGIALTFSLGGFAAEEFPVPNGFESAYREVDGVKLHYVKGG--QGPLVMLVHGFGQ SCO7440 TDDATAPSSSCDQNPVPGLPLHDLAGFTHRWVDAEGIRLHAVEGGRPAGPTVVLLAGFPQ RHA02607 --------------------------MREGSIRVGDRTITYLEAGDPGGPLVLHNHGGPS RHA00320 --------------MTTTKFDKAVGRYVHLTIDDVEYRVYYEESG--SGIPLLLQHTAGC SCO3690 ----------------------------MDVRDRNHVTV----TGRADGPVVMLAHGFGC NE2298 -----------------------------------MASIHIETTG--NGPDLVMLHGWAM RHA01401 ---------------------------MGTITTQDGTEIFYKDWG--TGQPIVFSHGWPL PP5253 ---------------------------MSTFVTRDGTSIYFKDWG--SGKPVLFSHGWPL PSPPH4522 LSAAAEGMHEPVHQSACPINPTSRGICMSTFNAKDGTEIYYKDWG--EGKPVLFSHGWPL PSPPH3482 ----------MSSSNLQTHSAQSFVQAPNKIATVAGIAFAYRESGLRGGVPLLLLNYWGA RPA3430 ----------------------------MPSFDHAGVNIAYLDEG--EGEPILLIHGFAS PP3645 -----------MACLPAAPDLQGAPSMIQPVAERTPAGTSYLDVG--QGQPVVLIHGVGL RHA08081 -----------------MTTTTDQSPEIAKTIDVNGIATNYHDVG--EGAPVVLIHGSGP RPA0348 IADPIAPAGKAIGQDGGGHT-IYQVDANGISIGYKLI---GQGAPMVMIM---GLGG--- RPA4646 SPQSMRYLGERLGE-----R-GYSI------LGPRLP---GHGVSPAA-----MAKT--- PA2949 DKD------NWLRFARPLTE-RYHV------VALDLP---GFGDSSKPQ----QASY--- SCO3439 VGGRRLHLDTYNGQLPGALL-RIRPGDRLRILLRNRMLPSGVPLNALP-----PLCA--- SCO3566 ------FWWTWRHQLVALADAGFRA------VAMDLR---GVGGSDRT-----PRGY--- PA2934 ------TWYEWHQLMPELAK-RFTV------IAPDLP---GLGQSEPP-----KTGY--- SCO7440 ------TWWAWRKVMPGLAA-RFRV------IAIDLP---GQGHSERP-----RGGY--- RHA02607 SRLEAELFDSH-----AKAN-GLRF------VCADRP---GIGGSDP------QPGR--- RHA0032 DGR---QYRHLL-EDEEITS-HFRV------ITWDLP---YHGRSLPPLSVRWWEQEYNL SCO3690 DQN---LWRL---VTPILER-DFTV------VLFDHV---GSGNSNLSA----WDPERYG NE2298 ------HSGVWDGVVESLSQ-RFRL------HQVDLP---GHGASRDC------------ RHA01401 SAD---DWDTQMLFFLQ--H-GYRV------IAHDRR---GHGRSTQT-----GDGH--- PP5253 DAD---MWDSQMEFLAS--R-GYRA------IAFDRR---GFGRSSQP-----WNGY--- PSPPH4522 DAD---MWEYQMEYLSS--R-GYRT------IAFDRR---GFGRSSQP-----WTGY--- PSPPH3482 VLD---DFDPR--IVDGLAT-RHHV------IAIDYR---GAGLSTG------TAPL--- RPA3430 NKNVNWVYPSWLSELKR--T-GRRV------IAIDNR---GHGESSKLYD---PNDY--- PP3645 NKE---MWGG---QFVGLAN-DYRV------IAYDML---GHGQSRVP-----AADT---

76
RHA0808 GVT---AWANWRTTIPHLAE-KFRV------IAPDIL---GFGYTERPD----GVEY--- RPA0348 TADNWPPQV---IEALSKNHQLILMDNRGMGHTTANDTPFSYPLFAADVIGL---LDA-- RPA4646 TANDWVNCAEDALLELSAKCDKTFVAGLSMGGTLSLYLAASHPDKVAGVIPINAAVQI-- PA2949 DVGTQAERVANFAAAIGVRR-LHLAGNS-MGGHIAALYAARHPEQVLSLALI--DNAG-- SCO3439 DKAADGTEHAHSAAGGPLHC-APEAGHQVVDGHTIIQQALATNLHTHGLQVSPSGSAD-- SCO3566 DPAGLALDITGVIRSLGEPD-AALVGHD-LGGYLAWTAAAMRPKLVRRLAVSSMPHPRRW PA2934 SGEQVAVYLHKLARQFSPDRPFDLVAHD-IGIWNTYPMVVKNQADIARLVYMEAPIPD-- SCO7440 DTHTVASRVQTALTALDVPK-YWLVGHD-VGAWVAFSLALKYEERLHGVALLDAGIPG-- RHA02607 TFEGWTDDLLLLADSFGAQR-FAVTGWSEGGPWALAAAAYLDPARLVNVVCIAGGNYGTF RHA00320 TQDFFMKFVVALAEALDCED-AVYMGSS-MGGNLAPDLALHYPGVFRAVIALEAALHS-- SCO3690 SLDGYVDDVLELCRELALGP-VTFVGHS-VSAMMGVLAAVREPEAFAGLVLL---APS-- NE2298 ALDSL-DQMTEVIADRLPGR-YSVCGWS-LGGQVAIRLALQAPERVQQLVLV---AST-- RHA01401 DMDHYADDLAELTAHLDLQD-AIHVGHSTGGGEVVHYLARHGESRVSKAALI--SAVP-- PP5253 DYDTFADDIAQLIEHLDLRD-VTLVGFSMGGGDVSRYIARHGSERVAGLVLL--GAVT-- PSPPH4522 DYDTFADDIAELIEHLDLHD-VTLVGFSMGGGDVTRYIAKYGSERVAKLALL--GSVT-- PSPPH3482 TIGEMARDTLELVRAMGYRQ-VDLIGFS-MGGFVAQDLALKAPDLVRKVILAGTGPAG-- RPA3430 TLEAMASDAVALMDHLGIAR-ADVMGYS-MGARIGANLARRQEQRVRSVVL---GGLG-- PP3645 PLEGYADQLAELLDHLQIAQ-ATVIGFS-MGGLVARAFALNYPQRLAALVVL-------- RHA08081 NSTTWTQHLVGLLDALGLDT-VSIVGNS-FGGSLALNIATKHPERVDRLVLM--GSVG-- . RPA0348 -----LGIKRSDVLGYSMGSTITQQLLLQYPD------------RFNKALIH-----ATS RPA4646 ----------------------------DSPD-----LAGLAYARGLPEFVP-------- PA2949 -----VMPARKSELFEDLERGENPLVVRQPED----------FQKLLDFVFVQQPPLPAP SCO3439 -----NVFVRLDP------LEDHQYAYDIPFD----------HPAGLHWYHP-------- SCO3566 -----RSAMLGDVRQSRAGSYVWGFQRPWVPE----RQLTADDGALVGRLLH-----DWS PA2934 -----ARIYRFPAFTAQGESLVWHFSFFAADDRLAETLIAGKERFFLEHFIK----SHSS SCO7440 -----ITLPDSIPTDPDRAWKTWHFAFHLVPE-LPETLLTGRERDYVDWFLK----VKTL RHA02607 GSNWAAKYLSSVDALGGRLELHFHPGFTLMYD-----VLGISATHFADRYAKAITQSACT RHA00320 -----PGFYLDYWFHPEISNDSKPAAMYALTS---------------------------- SCO3690 -----PCFVDDPDTGYRGG-----FSAADIEE-----LLDSLDANYLGWSGAMAPVIMGN NE2298 -----PCFVRRAD--WPWGMEDSTLTL-FMEN------LARDYTQTLNRFLT---LQVSG RHA01401 -----PLMVKTEA--NPGGLPKDVFD--DLQA-----QLAANRSKFYRDLPS-GPFYGFN PP5253 -----PVFGKRDD--NPDGVDLSVFE--GIRA-----GLRADRAQFIADFAT---PFYGL PSPPH4522 -----PFFLKTDD--NPEGVDKSVFD--GIKE-----GLLKDRAQFISDFAT---PFYGL PSPPH3482 -----------GHGIERIGALSWPLILKGLLR--------LRDPKVDMFFTS---TLNGR RPA3430 -----MGLLSNEG--RPGENVARALEADALDD---------------------------- PP3645 -----NSVFNRTPEQSAGVIARAAQAAELGPD-------ANVDAALDRWFSR-----EYK RHA08081 -----VPFEITDGLDAVWGFEPSLPAMRKLLD-----VFAYDRSLVNDELAE-------- RPA0348 TDGSNVAKALH-GRVPTDPIVARQVEATTHWKTPLDKL---------PSIDN----QVML RPA4646 GIGSDMVDTST-KELAYDQVPVTCVKQVMGLAATARALL--------PRIKC----PTLV PA2949 LKRYLGERAVA-ASAFNAQIFEQLRQRYIPLEPEL------------PKIEA----PTLL SCO3439 ---HHHGSTTH-QAWSGLAGPIVVEGDIDHVPEIAGMRE--------RTIVL----SMLR SCO3566 GPRLLDDDAVT-AYRRAMCIPSTAHCSVEPYRWLVRSLARPDGIQFYRRMKRPVRVPTLH PA2934 NTEVFSERLLD-LYARSYAKPHSLNASFEYYRALNESVRQNAE-LAKTRLQM----PTMT SCO7440 SPDTFDGAEID-HYAAAVAAEGGLSASLAYYRDAAESARRNHDALERGHLTV----PVLG RHA02607 ADREVLSDEKV-LDAFLRAGRECFRHGADGLVVDATMLYKAWP-FDMTKVTR----PVHF RHA00320 PTAPEVGRRET-TWVYSQGSPPVFKGDLHYYSVEHDLRETA------QNIDTSKT-AVYI SCO3690 PERPELGEELTNSFCRTDPDIARVFARVTFLSDNRADL---------APVRV----PTLV NE2298 SEDQARVLAWL-RKSILRGQPPTPATLQAGLKILQTSDLRA----ELNQVSQ----PVLL RHA01401 RPGVESSEAII-ANWWRQGMMGGAKAHYDGIVAFSQTDFTE----DLKKITV----PVLV PP5253 NHGQQVSQGVQ-TQTLNIALMASIKGTLDCVTAFSETDFRP----DMAKVDV----PTLV PSPPH4522 NKGQKVSEGVQ-AQTLNIALLASLKGTLDCVTAFSATDFRA----DMAKIDV----PTLV PSPPH3482 RAARNYLSRVK-ERTVDRDKPPTPRLLLRQLKAIKAWGKQPAQ--DLARLRV----PVLI RPA3430 ---VTDPVGRT-FRAFADQTRSDRKALAACMRGSRGLMSREDA----AQIAV----PVLI PP3645 AANPAQVAAI--RQVLASNDPQGYHTTYSLFA--TQDMYRAD---DLGSIQV----PTLI RHA08081 LRLAAATRPGV-QEAFSAMFPAPRQQGVDEMAVDETLI---------AGLTN----DTLI : RPA0348 VVGTADNVVGTESSKTIASAIPG-AWLVQ--FKGATHHLMYETPEGF-SAAALTFFETNE

77
RPA4646 INSRVDHVLSPANATVIANHVGS-SRIETLWLDNSYHVATIDNDKDLIAAEIDSFVTRNL PA2949 LWGDRDRVLDVSSIEVMRPLLKR-PSVVI--MENCGHVPMVERPEET-AQHYQAFLDGVR SCO3439 LDGNGEN-----------------PTAVV--LPTGGDDPFTTVPAVP-TRMVATLNGQLR SCO3566 LHGSLDPVMRTRSAAGSGQYVEAPYRWRL--FDGLGHFPHEEDPVAF-STELINWLKDPE PA2934 LAGGGHGGMGTFQLEQMKAYADD-VEGHV--LPGCGHWLPEECAAPM-NRLVIDFLSRGR SCO7440 VSGSHGSI--PDMAASIGPWAAN-ATGAV--IPQAGHFIPDEQPEAT-VKVLTAFIDYER RHA02607 WQGSADTLVPEIINKTVADKTPG-AVWHP--ISGGGHFIAVSHANDI-LALVANDLAPVQ RHA00320 LNGEYDFATGPEEAAELGAQIEG-AKVVP--MAGLGHFPMSEDAAQF-FRCIKPVLWEIR SCO3690 AQCSSDAIAPREVGAFVHAQIPA-SRLVT--LNATGHCPQLSAPQET-AAAIAAFAGGAD NE2298 IHGRNDVITPAGAADWMQQHLPR-ARLVL--FPHCGHAPFLSFPEQF-VSCFDAL----- RHA01401 MHSEDDQIVPYADAGPLSAKLLANGSLKT--YQDFPHGMPTTQAETI-NADLLAFLKDSY PP5253 IHGDDDQIVPFETTGKQAAELIRGAELKV--YAGAPHGFAVTHAQQL-NEDLLAFLQR-- PSPPH4522 IHGDGDQVVPFEASGKRAAELIKGAELKV--YPGAPHGFAVTHAEAL-NKDLLTFVQG-- PSPPH3482 VVGDSDVMVASELSRDMSQRLPQ-AQLVV--YQDAGHGCVFQYHADF-VSTALEFLA--- RPA3430 AVGTDDDVAG--SAHELGDIIPG-SQVLD--IPRRDHMRAVGD-RVY-KEGVVDFLARRP PP3645 ATGELDSGSTPAMTRQLAACIPG-ARSVV--LAEQRHMMPVEAPREV-NKMLLDFLTQAR RHA08081 VHGRDDQVIPLSNSLRLLELIDR-SQLHV--FGRCGHWVQIEHSARF-NSLIADFLSE-- RPA0348 TVTPKIEPNASVAPPPTQP RPA4646 ------------------- PA2949 NAQVAGR------------ SCO3439 PK----------------- SCO3566 PDR---------------- PA2934 ------------------- SCO7440 AE----------------- RHA02607 N------------------ RHA00320 DSAVPSKVLSATQ------ SCO3690 ------------------- NE2298 ------------------- RHA01401 EN----------------- PP5253 ------------------- PSPPH4522 ------------------- PSPPH3482 ------------------- RPA3430 ------------------- PP3645 TLTESAKGIVA-------- RHA08081 -------------------

78
Appendix A.2 Amino acid sequence alignment of proteins that were not active on any substrates.
TM0820 ----------------------------------MENFVFHNPTKIVFGRGTIPKIGEEI TM0423 ---------------------------------MITTTIFPGRYVQGAGAI--NILEEEL ATu2215 ---------------------------------------MQNATLPLPLQS--PLLAA-S PP0624 ----------------------------------------MSDLTQA------LLFPD-W PA3301 -------------------------------------MPAAAYWLPASDET--PLYTRHW PP4551 -------------------------------------MPHDAFWLPASEHC--SLYVHQW PSPPH0357 -----------------------------MQTLYPQIKPYARHDLAVEQPH--VLYVDES PP3523 -----------------------------------------------M-----EAFNQ-E ATu2497 -----------------------------------------------------MHFEV-H RPA3731 ----------------------------------MHLIHRHRLWIDNAGVR--IALVR-W PSPPH1999 -----------------------------------MTVAVEEVRLSLGHIE--LAAHL-Y SAV0974 ----------------------------------------MGTFTTDDGTE--IFYKD-W ATu5389 ----------------------------------------MGFVTTKDGTE--IFYKD-W PA2717 ----------------------------------------MGYVTTKDGVE--LFYKD-W PA3053 MSLPRLSLSLCLALACGPALASQAPVYGERLEGFDYAYPVHYLDFTSQGQPLSMAYLD-V PP2201 --MLRATALALACLASSSAIAAESPTYGPQLEGFSYPHPLKHFDFKSQGQALQMGYMDVP SAV0923 -----------------------------------------------MDIRRRNNVVV-T ATu2061 -----------------------------MTAPSLDNWEKRKRFVELPRKRQ-MAFIDTG SAV2105 ------------------------------MPQTPRTTALTHRLVSSPAGR--IHLVE-Q RPA2773 -------------------------------------MASTRSTITANGIE--LFLRE-Q SAV141 -----------------------------------MEGSLTHRFVDVNGVR--LHIAE-Q RHA05146 ----------------------------------MQTGRVQHATNPVDGVR--IAYKT-V RHA00069 --------------------------------------MSQLQHLALHGDA--VAYRL-S SAV1746 -----------------MVDVPPPRPRRALRLRPVGDGELRLHHRVVHGYR--RAYRM-A PP4540 -----------------------------------------MAYFEHEGCT--LHYEE-Y PSPPH2566 -----------------------------------------MIQLTAEHTPAGTSYLA-T PA0231 -----------------------------------------MPTVRLADGD--LNYSL-E PP1380 -----------------------------------------MAHLQLADGV--LNYQI-D PA3586 --------------MRAFIFLAALLCSLSSFAAARCDARVPTETVELGDVR--LAYQS-I PP2804 --------------------------------------MSTDRFCELAGERR-LCYRS-H SAV0519 --------------------------------------------MTRGGVR--LSCRD-W SC0267 --------------------------------------MSTTQHTLV------PVYDHRT SC5943 ------------------------------------MTKHRQETVRAGELD--VGFLD-S PSPPH0033 -----------------------------------------MPDLLIDGKT--LHYAD-Q RPA1568 ------------------------------------------MQIKVNGTD--TFVAT-G SAV7198 ---------------MCLSPLVTTPVSPPVSPTWRDRSMTVLRRVSVNGVE--LNVAL-A SAV1548 -----------------------------------------MPQLAVDGAG--MTYDD-E SAV4410 -----------------------------------------------MAPF--LAYED-K TM0820 KNA---GIRKVLFLYGGGSIKKNGVYDQVV--DSLKKHGIEWVEVSGVKPNPVL-----S TM0423 SRF---GERAFVVIDDFVD--KNVLGENFF-------SSFTKVRVNKQIFG--------- ATu2215 GEN---RVRHGFFTRQGGV--SEGLYQGLN--VGLGSHDEPEKVQENRRRV--------- PP0624 PAP--ASVRACVTTRQGGV--SLPPYETFNLGDHVG-DDPAAVAENRRRLS--------- PA3301 PSA--TAVGAVMLSHGMAE--HAGRYERLA--AALNAAGYHFYAIDQRGHG-RT-----A PP4551 LPA-TPVKAVVLLAHGMAE--HAGRYQRLG--QALSEAGFALFAADQRGHG-RT-----A PSPPH0357 GSP---EGLPVVFIHGGPG--SGCDAHSRC---YFDPNLYRIVTFDQRGCG-RS-----T PP3523 GKP-------------------MSKFDFQL-------------AYTIKPHN--------- ATu2497 GRT-DAEAPTILLSSGLGG--SSAYWLPQI--EALS-DHFRIVTYDHRGTG-RT-----G RPA3731 GDV-DPDKPVALLLHGTGF--VGEIWDEIA--NGLA-DRYTVYALDRRGHG-AS-----D PSPPH1999 GPK---DGQPVIALHGWLD--NANSFARLA--PRL--EGLQVVALDLAGHG-HS-----D SAV0974 GSG-----PPVVFSHGWPL--NADAWDVQL--RLVAENGYRAVAHDRRGHG-RS-----G ATu5389 GPK---DAQPIVFHHGWPL--SSDDWDAQM--LFFLSKGYRVVAHDRRGHG-RS-----A PA2717 GPR---DAQVIYFHHGWPL--SSDDWDAQM--LFFLAEGFRVVAHDRRGHG-RS-----S PA3053 APK-KANGRTILLMHGKNF--CAGTWERTI--DVLADAGYRVIAVDQVGFC-KS-----S

79
PP2201 AQG-QANGHNVVLMHGKNF--CAATWETTI--DALSKAGYRVIAPDQVGFC-TS-----S SAV0923 GRP---DGPVVLLAHGFGC--DQNMWRLVV--PALA-DDFRVVLFDYVGSG-RS-----D ATu2061 GPG-----PVLLMLHGFSD--TSRSFSIIE--PYF--QEYRLIAPDLPGHG-AS------ SAV2105 GTG-----PLVLLVHGFPE--SWYSWRHQL--PALAAAGYRAVAVDVRGYG-RS-----S RPA2773 GEG-----PLVVLCHGWPE--LSYSWRHQI--GALADAGYHVVAPDMRGFG-RS-----S SAV141 GQG-----PLVLLLHGWPE--SWYSWRHQF--GALAAAGYRVVAPDQRGYA-RS-----E RHA05146 GDG-----EPLVLIHGTAL--SHAIWRGFGYVAALR-DRYRLILVDLRGHG-CS-----D RHA00069 GEG-----ETLLLVHGMAG--SSATWRAVL--PQLA-RRYRVLAPDLPGHG-DS-----A SAV1746 GEG-----PVLVLIHGIGD--SSATWAELI--PDLA-RTHTVIAPDLLGHG-AS-----D PP4540 GQG-----EPLVLLHGLGS--SCQDWELQV--PLFS-RHYRVILMDIRGHG-RS-----D PSPPH2566 GQG-----HPVVLIHGVGL--NKEMWGGQI--VGLA-TNYQVIAYDMLGHG-AS-----P PA0231 GPA---GAPVLLLSNSLGT--DLGMWDTQI--PALT-AHFRVLRYDTRGHG-AS-----L PP1380 GPD---DAPVLVLSNSLGT--DLGMWDTQI--PLWS-QHFRVLRYDTRGHG-AS-----L PA3586 GRD---SDPALLLVMGLGG--QLIHWPDEV-VSALCEQGFRVIRYDNRDVGLSAWNVPVP PP2804 GHE---AAPAVLLIVGLGL--QLTYWPQAL-IDGLVERGFRVITLDNRDAG-RS-----F SAV0519 GGC----GRPVVLLHGLAG--HAGEWDTLA--GALS-PRYRVIAVDQRGHG-AS-----E SC0267 GTG-----PTLVFLHYWGG--SARTWDLVV--DRL--PGRAVVAVDFRGWG-RS-----R SC5943 GDG-----VPLLLVHGGES--DRTQFATLR--AGLG-AGIRAISYDQRDSG-IT-----V PSPPH0033 GTG-----PVVLLGHSYLW--DKAMWSAQI--DTLA-SRYRVIVPDLWGHGDSS-----G RPA1568 GKPFDASLPAAVFIHGAGF--DHSVWALQT--RWFAHHGFAVLAPDLPGHG-RS------ SAV7198 GKG-----PAVLLLHGFPH--TWELWTDVM--AGLS-ERYQVIAPDLRGFG-AS-----T SAV1548 GPHDGDGGVPLVFIHGWTA--DRHRWDHQM--AHFA-DKRRVVRLDLRGHG--------- SAV4410 GTQ-RDTSLPLVLVHGHPF--DRTMWAPQL--AAFA-PSRRVIAPDLRGYG-AS-----P TM0820 KV-------------HEAVEVAKKEKVEAVLGVGGG-SVVDSAKAVAAGALYEGDIWDAF TM0423 -----------------------GECSDEEIERLSGLVEEETDVVV--GIGGGKTLDTAK ATu2215 --------------------TQWFGLSADRLATVHQ-VHSPDVVVV--DDNYDGNRPQAD PP0624 --------------------EQFTIQPAWLKQVHGRVVADADPTVV--AE------ADAS PA3301 EADELGHFAD-----QGGWGKVVGDLASLNHHIRQQ-HPELPIFLL--GHSMGSYISMAY PP4551 ELGNLGLFAR-----HHGWNAVVNDLGCVAQHIGQQ-FPCTPLFLF--GHSMGSYIAQAY PSPPH0357 PH-------------ASLENNTTWKLVEDLERIREH-LGIDKWVLF--GGSWGSTLALAY PP3523 ---------------PA---YDETDAAQARLHLREK-LGLDTVEHI--ETTLLGMITLNG ATu2497 G--------------EVPTEGGISAMADDVLEIVSA-LNLEKFHFM--GHALGGLIGLDI RPA3731 K--------------PGR--YHFADFASDLAAAIET-LGLSGIYGI--GHSAGATDLLLT PSPPH1999 HR-------------PAGSSYALADYAFDVLQVAEQ-LGWQRFALL--GHSLGAIISVLL SAV0974 QP-------------WNG--NHMDTYADDLSQLIGA-LDLRDAVVV--GHSTGGGEVARY ATu5389 QV-------------ADG--HDMDHYAADAFAVVQA-LDLKNAVHI--GHSTGGGEVARY PA2717 QV-------------WDG--HDMDHYADDVAAVVER-LGVRGAIHV--GHSTGGGEVVHY PA3053 K--------------PAHYQYSFQQLAANTHALLER-LGVARASVI--GHSMGGMLATRY PP2201 K--------------PAHYQYSFQQLADNTHALLEQ-LGVKQTIVL--GHSTGGMLATRY SAV0923 LSAW-----------SEQRYSSLEGYALDVLEVCEE-LDLRDVAFV--GHSVSAMVGVLA ATu2061 ---------------SVGHGFHVADFAETIDRFLTL-MGVSRFFVL--GHSMGAMTAIEL SAV2105 RP-------------NAVHAYRMLDLVEDNVAVVHA-LGERTAVIV--GHDWGSNIAATS RPA2773 AP-------------QAVEAYSIFDLVGDMVALVAE-LGETRAAII--GHDWGAPVAWHA SAV141 QP-------------PDVASYTLLHLVGDVIGLIEE-LGEEQAVVV--GHDWGAPVAWTT RHA05146 KP-------------HDESAYAMDLVSGDVLAVLDH-LDLPSAHVL--GYSLGGRVALAL RHA00069 KP-------------RGD--YSLGAFAAWLRDLLNE-LDIERVTVI--GQSLGGGVAMQF SAV1746 KP-------------RAD--YSVAAYANGVRDLLAS-LDIESATLV--GHSLGGGVAMQF PP4540 KP-------------ADG--YQIATFSADLLALLEH-LHTGPVHFV--GLSMGGMVGFQF PSPPH2566 RP-------------DPD--TGLPGYAEQLRELLEH-LQLPQATVV--GFSMGGLVARAF PA0231 VT-------------PGP--YAIGQLGADVLALLDA-LELPRVHFC--GLSMGGLIGQWL PP1380 VT-------------EGP--YSIEQLGRDVLALLDG-LDIQKAHFV--GLSMGGLIGQWL PA3586 SSRLTYEVVRYRLGLPVSAPYTLTDMAGDALHLLDA-LDIPQAHVL--GASMGGMIAQHI PP2804 FTDVAPPTPLQQFLRKRTPGYDLGDMASDVIGLMDA-LHLEAAHVV--GMSMGGMIAQTL SAV0519 RF-------------PRE--VSRAAYVADVVAVLDQ-LALHRPVLV--GQSLGGHTAMLT SC0267 AL-------------PGP--YTLGRFADDTLAVLAD-AGITDYVLV--GHSMGGKVAQLV SC5943 NP-------------PVP--YTPEVLADDLVDLLDA-LGLARAHLL--GTSFGGAVAQHA

80
PSPPH0033 FP-------------EGT--RNLDDLARHALALLDH-LNIERCSIV--GLSVGGMWGAIA RPA1568 ---------------GGEALKTIAEMADWIAALLDA-TGAQPAKLI--GHSMGSLIALEA SAV7198 RA-------------KTG--YDAGSLADDAAALLET-LGAGPAAVV--GIDAGTPPALLL SAV1548 --E------------SGGSARTIDELAGDVIALLDH-LEIDRFIPV--GHSMGGMIAQTL SAV4410 VV-------------PGI--TPLSVFAEDIAALLDE-LKADRFVLA--GLSMGGQIAMEC TM0820 IGKY-----QIEKALPIFDVLTISATGTEMNGNAVITNEKTKEKYGVSSKA----LYPKV TM0423 AVAY-----KLKKPVVIVPTIASTDAPCSALSVIYTPNGEFKRYLFLPRNPDVVLVDTEI ATu2215 AMVT-ATP-GFVLGVLSADCGPILFADAQAGVVGAA----HAGWKGALFGV----LENTI PP0624 WTRQ---P-GIACTVMTADCLPALFCDRAGTRVAAA----HAGWRGLAGGV----LEATL PA3301 LLHH---S-CSLQGAILSGSNYQPQALYRIARL-------IARFERWRQGP----LGKSA PP4551 LLHH---S-GSLHGAILSGSNYQPAVLYRFARL-------IARLESWRQGP----LGKSA PSPPH0357 AQTH---P-DRVHALILRGIFLARQQEIDWFYQAGASRLFPDYWQDYVAPI----PLDER PP3523 TTLA---------------------------------------------DR----KREAE ATu2497 ALRQ---P-RLIDRLVLINAWSKADPHS------------GRCFDVRIELL----EKSGV RPA3731 AKLL---P-DRIARLFVMEPTVMDPSVVPGR-------------DARLSDQ----GEAAV PSPPH1999 ASSL---P-ERVTRLALIDGLLPLTGKAESA---------AERMGAAMQAQ----LELAN SAV0974 IGRH---GSGRVAKAVLVGAVPPIMLKTDANPEGLP----IEAFDAIRAGV----LSNRA ATu5389 VAKHGQKA-GRVAKAVLVSAVPPLMVKTTANPEGLP----IEVFDGFRSAL----AANRA PA2717 IARY---PDDPVPKAAIISAVPPLMVKTEGNPGGLP----KSVFDDLQAQL----AANRA PA3053 ALLY---P-RQVERLVLVNPIGLEDWKALGVP--------WRSVDDWYRRD----LQTSA PP2201 ALMY---P-QQVERLAMVNPIGLEDWKALGVP--------YRTVDQWYARE----LKLDA SAV0923 AQKA---P-ERFSRLVMVAPSPRYID--------------EDGYRGGFSAE----DIDEL ATu2061 AARR---S-SSVRGLALISGTLEPDFGTESK---------LARDILALRDP----IKPAG SAV2105 ALVR---P-DVFRAVGLLSVPYTPRGGPRPSEIFAGMGGDEEFYVSYFQEP----GRAEA RPA2773 AQFR---P-DLFAAVAGLSVPPPWRGKGPPLDQLRAAGI-TNFYWQYFQKL----GVAET SAV141 AMLR---P-DKVRAVAGLSIPPILPGGMVPPSITRTQYG-EGFYQVYFQQP----GVADA RHA05146 AVGA---P-ERLESLIVGGGSSRPQAGAFD----------RLFFPGCIDVL----ERDGM RHA00069 SYQH---P-ELCDRLVLIGSGGLGP---------------DVNWTLRLLAA----PGSEF SAV1746 AYQF---P-ERTERLILVSAGGV-----------------GREVNPVLRAV----SLPGA PP4540 AVDH---P-QWLRSLCIVNSAPEVKRRT------------RSDWAWWLKRW----SLARI PSPPH2566 ALEF---P-QLLAGLVILNSVFNRS---------------PEQRAGVIART----SQAAE PA0231 GIHA---G-ERLGRLVLCNTAAKIAS--------------DEVWNTRIDTV----LKGGE PP1380 GIHA---G-ERLHSLTLCNTAAKIAN--------------DEVWNTRIDTV----LKGGQ PA3586 ADMA---P-QRLLSLTLVMTSSGAEGLPAPS---------ESLLRLLARRE----AASRE PP2804 AARY---P-ERVSTLTSIFSTTGSLRVGQPA---------LKALFKLLRRP----PRNQH SAV0519 AAAH---P-HLAHALVLVEAGPGGPNPRVQAE--------IGGWLDAWPTP----FPSRE SC0267 AATR---P-AGLRAIVLVGSGPAKPAARV-----------TPEYREALSH-----AYDSA SC5943 ARRH---P-ERVASLVLVATTPSYAMG-------------SAAIDELLEMS----HEDRQ PSPPH0033 ALLA---P-ERITGLVLMDTYLGKESEA------------KKAYYFSLLDK----LEQAG RPA1568 AARH---P-AKVASLALIGTTSTMAVGP------------DLLKAAEANDP----AAIAM SAV7198 ALRR---P-GLVRRLVVMESLLGRLPGAESFLAG------GAPWWFGFHAV----PGLAE SAV1548 ALAH---P-ERIERLVLVNSIS------------------RMTYSRGRGLL----MAAST SAV4410 YRLF---P-ERIAGLVLADTFPTAETEAG-----------KRARGAMADRL----LREGM TM0820 SIIDPSVQFTLPKEQTVYGAVDAISH----------ILEYYFDGSSPEISNEIAEGTIRT TM0423 VAKAPARFLVAGMGDALATWFEAESC--------KQKYAPNMTGRLGSMTAYALARLCYE ATu2215 EAMVKLGASRERIAASLGPSISQDNY---------------------------------- PP0624 DRLALPP-------EEVLVW-LGPAI---------------------------------- PA3301 LIDFLSF-------GSFNKA-FKPNR-------------TAFDWLSRDP----------- PP4551 LIEWLSF-------GSFNNA-FKPNR-------------TAFDWLSRDP----------- PSPPH0357 DNILAAFHKRLTGPDQIAQMHAAKAW---------------STWEGRCATLRPN------ PP3523 KLVRDYIH---------------------------------------------------- ATu2497 DAFVKAQPLFLYPAAWMSEHQERLAR---------------------------------- RPA3731 SGARRRQAEFASAAAAFERYRSAPAF---------------APWSEYALRTYISHGFAHQ PSPPH1999 KKKPVYQDQDRAIQARMKGV-VAVSR---------------EAAELLAQR----------

81
SAV0974 QYYWDLS-------DQFYGF-NRPGA-------------EGTDGVR-------------- ATu5389 QFFRDVPA------GPFYGF-NRDGA-------------KVQEGVI-------------- PA2717 QFYQDIPA------GPFYGY-NRPGA-------------QPSEGIV-------------- PA3053 EGIRQYQQA-----TYYAGE-WRPEF---------------------------------- PP2201 EGVRAYERK-----TYYAGR-WKPEY---------------------------------- SAV0923 LTSLD---------SNYLGWSATMAP---------------------------------- ATu2061 GFLHDWYSC------------SRPVN---------------------------------- SAV2105 EIEPDVR-------GWLAGF-YAALSADTMPAPDAPDPHFVRRGGTLRERFPAGRLPAWL RPA2773 EFERDVASTMR---GMLCGGFADPGR-----------SLFVPEGRGFIGRSAASLPLPPW SAV141 EFAKDIP-------NSFRRF-LVGASGDN-PLGREPSPLVIPDGLGLLDIMPESPALPAW RHA05146 DAFLAQW-------NARRSWPIDAGT---------------------------------- RHA00069 VLPLVAPSAVRDAGNKVRGWLAAVGI---------------HSVRG-------------- SAV1746 HLMLSTL--------RLPGMRFQVGM-------FARLMRLLDTDLGQDA----------- PP4540 LSVETVGK------GLAERLFPKPQQ---------------------------------- PSPPH2566 HGPDANAG------EALSRWFSREYQ---------------------------------A PA0231 QAMRDLRD------ASVARW-FTAGF--------------------------------AE PP1380 QAMVDLRD------ASIARW-FTPGF--------------------------------AQ PA3586 QAVEQQADLL----AALGSPEVRDDR---------------------------------- PP2804 ESVRDYVDIMGLI-GSRLEW-NEPAL-------------------------------RDY SAV0519 SAVEFLG-------GGLVGEGWAAGL---------------------------------E SC0267 ESVAGAR-------DHVLTATELPAA---------------------------------- SC5943 QAAADYF--------------FTPEG---------------------------------- PSPPH0033 AFPEPLL-------DIVVPIFFRPGI-----------------------------DPQSP RPA1568 MTIWGLGPD-----AEIGGN-LAPGL---------------------------------- SAV7198 TVLEGNE-------ARYLDWFLDTGT--------------------------LGDGVRPA SAV1548 LVPFKLFVA-----ANIQRA-FAPGH---------------------------------- SAV4410 RGYAD---------EVLHRM-VAPYA---------------DADV--------------- TM0820 IMKMTERLIEKPDDYEARANLA----WSATIALNGTMAVGRRGGEWACHRIEHSLSALYD TM0423 TLLEYGVLAKRSVEEKSVTPALEKIVEANTLLSGLGFESGGLAAAHAIHNGLTVLENTHK ATu2215 --EVGPEYVARFTDIDPAYADY---------FAATERQGHALFDLKQFT----I-DRL-- PP0624 ------GPQAFEVGLEVRDAFTAVHPQAARAFVDGERPGKLLADIYELA----R------ PA3301 --QEVDRYVADPLCGFRCSNQL----WVDLLGGLADITPP--THLRQIDAD--L-PLL-- PP4551 --GEVDQYVNDPLCGFRCTNQL----WLDLLQGLAQISQP--GNLAQIDPN--L-PML-- PSPPH0357 --PQVVDRFAEPHRALSIARIE----CHYFMNKAFLEENQLIRDMPKIA----HLPAI-- PP3523 ------------------------------------------DALKELR----VLSTV-- ATu2497 ------DDAHGVAHFQGKTNVL----RRIAALRAFDID----ARLGEIG----N-PVL-- RPA3731 PDGRVRLLCSPDVETEILRPIF----EAMEQVYRGDERGNPFGWLRELG----C-PVR-- PSPPH1999 --GLMPVPGGYTWRSDSRLTLP-------SAIRFTDQQAM--AFVHGIR----C-PTQ-- SAV0974 --RAFWLWS----MQVGLKAAY----DCIEQFSEQDFT----EDLRRID----V-PTF-- ATu5389 --QNWWRQG----MMGGAKAHY----DGIKAFSETDQT----EDLTAIT----V-PTL-- PA2717 --RNWWRQG----MMGGAKAHY----DGIVAFSQTDFS----DDLKRID----I-PVL-- PA3053 --DRWVQMQAGMYRGKGRESVA----WNSALTYDMIFTQPVVYELDRLQ----M-PTL-- PP2201 --ERWVQMLVGLNKGPGHEAVA----WNSALIYDMIFTQPVYHEFKHLQ----M-PTL-- SAV0923 --VIMDNPDRPELGEELTASFCATDPDIARAFARTTFLSDSRQDLKSVA----V-PTL-- ATu2061 --EEFLFRMKRDAANMAATTWH----GVLKAFAETDLH----YSASKIN----V-PVL-- SAV2105 SEADLDVYAGEFERTGLSGALN----RYRAMDRDWEDLAP--FDGAPVR----Q-PSL-- RPA2773 LTEAELAFFIEQYKQSGFRGGL---NWYRNIDRNWELTSP--WQGAPIH----Q-PAA-- SAV141 LTEEDIQAYAEDFALHGERAFTGAFNWYRNIERNNELLAP--FRGRGID----V-PAL-- RHA05146 ------RAAFMANDAQALAAYM----RRSGVEPGVDD-----DVLRGIA----V-PTL-- RHA00069 --DEMWNAYSSLSDSDTRQAFL----RTLRAVVDHRGQAV--SALSRLYLNEGL-PTQ-- SAV1746 --PELLTLVDALPDVTSRSAFI----RTLRAVVDWRGQAV--TMLDRCYLTEGM-PTL-- PP4540 --ADLRQKMAQRWARNDKRAYL----KSFDAIVDWGVQ----ERIGQIH----C-PTL-- PSPPH2566 ANPAQIAAIRQNLASNDPQGYL----TTYKLFATQDMYRA--EDLGDIR----A-PTL-- PA0231 REPAQVERIVAMLAATSPQGYA----ANCAAVRDADFR----EQLGLVQ----A-PTL-- PP1380 AQAEQAQRICQMLAQTSPQGYA----GNCAAVRDADYR----EQLGRIQ----V-PAL-- PA3586 -QQLLLQAARSYDRAFNPEGVQ----RQLLAILAEPSRV---PLLNRLQ----V-PTL--

82
PP2804 AMQAWPRGGGEASNLGTARQIG-------AIINSGDRT----QELQRIR----C-PTL-- SAV0519 ERDGWWPCFDREIMVASLAEIAQRSFW---------------DEWARIT----C-PVL-- SC0267 ---LKARIVTDSRNVTDAARIE----WPLRGIAQDIT-----EHTRRID----V-PAL-- SC5943 -QAGQRARPAGTLTRRTPEQSV----RRHDAARRHEIR----DGLGGIT----A-PTL-- PSPPH0033 VYQAFRSALAGMNAEQLRQTVV---PLGRMIFGRDDWL----GLLEQLN----ADTTL-- RPA1568 ----WMHGGSVRVLEANRPGVIFNDLSACNDYKDAL------AAAAKVT----V-PTL-- SAV7198 VRDAFVRAYTGREALRCAFSYY----RALPVSATQIERA---FDTARLT----V-PTM-- SAV1548 -PREEVREYIRASSATPRDVVM----TYYAAMRSFDVL----DRVGEIR----M-PTL-- SAV4410 --QAHVHRMMTGTDPEGAAAAL----RGRAQRPDYR------DLLTRVS----V-PAL-- TM0820 IAHGAGLAIV--------FPAWMKYVYRKNPAQFERFAKKIFGFEGEGEELILKGIEAFK TM0423 YLHGEKVAIGVLASLFLTDKPRKMIEEVYSFCEEVGLPTTLAEIG-------------LD ATu2215 TKAGVKAENL--------------------------GLCTYPDEH--------------- PP0624 IRLAARGVTA------------------------------VYGGG-------------LC PA3301 IIGGERDPVS--------QGKR----------LGD-LADALRGAG-------------LR PP4551 VIGGECDPVS--------AGKR----------LTH-LADALRATGNRHVQ--------LR PSPPH0357 IVHGRYDVIC--------PLDN----------AWE-LHQNWPGSE-------------LQ PP3523 KFYGCL------------------------------------------------------ ATu2497 VVATKDDLLV--------PYTR----------SLR-LAEGLPQSE-------------LC RPA3731 IATAELSWPI--------YKEM----------ATR-ASALIPGAS-------------RI PSPPH1999 LMVASDGMLA----------------------QRQELLSALPFDV-------------ER SAV0974 IAHGDADQIV--------PLVA----------SAPKSAELVKDAT-------------LK ATu5389 VLHGEDDQIV--------PIAD----------AARKSIKLLKNGQ-------------IK PA2717 VMHGDDDQIV--------PYEN----------SGVLSAKLLRNGT-------------LK PA3053 LLIGEKDNTA--IGKDAAPAELKARLGNYAQLGKD-AARRIPQAT-------------LV PP2201 LLIGDKDTTA--IGSDIAPPEVKARLGNYAELGPQ-VARLIPKGE-------------LI SAV0923 VLECAQDVIA--------PREV----------GAY-VHAAIPGSR-------------LV ATu2061 CLAGSEDPLF--------DDSH-----------RQQLFEAFPLAQ-------------TV SAV2105 FIGGGLDAST-------QWLAD----------AIEAYPVTLPGLV-------------SS RPA2773 FIAGSNDPVI--------SDKMSGK-------HLAAINRVLPNLK-------------QK SAV141 YVVGDRDMVTSLRGPDGGPSLS----------EIF-RGQGGPGNP-------------LS RHA05146 LFVGSNDA----------ERIG----------DTQRVAALIPGAS-------------LT RHA00069 LIWGDSDGII--------PVAH----------GYA-AHEAIPGSR-------------LA SAV1746 LLWGDRDSVV--------PVRH----------AYG-AHEAMPGSR-------------LE PP4540 VIAADHDYT---------PIQ-----------LKERYVALIPNAR-------------LV PSPPH2566 IATGELDPGS--------TPEM----------ARE-LAMRISGAE-------------VA PA0231 IVAGSHDAVT--------TPDN----------ARF-MQARIADAQ--------------- PP1380 IVAGTQDVVT--------TPEH----------GRF-MQAGIQGAE--------------- PA3586 VIHGTADPLL--------PVMH----------GVH-VAAHIRGSE-------------LK PP2804 VIHGDKDLMV--------ATNG----------GFA-TASAIRGAN-------------LV SAV0519 TVLGQSGIIS--------PQES------------ETMLRRLPVAT-------------AA SC0267 VVAGEHDQVE--------PAGV----------LRDNLVPYLARAD-------------FV SC5943 IVHGTQDRLA--------PYE-----------GALLMERRIPNAE-------------LC PSPPH0033 VMCGDADIPR--------PPEE----------TRE--MASLIGCP-------------YV RPA1568 FILGERDMMT--------PTKN----------GKT-LAAAISGSR-------------TV SAV7198 -AVGSHPVGA--------ALER----------QLRPFADDLTGH---------------- SAV1548 LVHGYYDIQL--------PVSQ----------MLR-MAKDYPDAV-------------VR SAV4410 VVVGADDTFT--------PVAD----------AAA-MHAALPGAT-------------LR TM0820 NWLKKVGAPVSLKDAG---IPEEDIDKIVDNVMLLVEKNLKPKGASLGRIMVLEREDVRE TM0423 GVSDEDLMKVAEKACDKNETIHNEPQPVTSKDVFFALKAADRYGRMRKNLT--------- ATu2215 ----------RFYSYR-RTTHRAEPDYGRQISAIAILED--------------------- PP0624 ----------TVSDAR-FFSYRRTPQGGRFASLVWLDPR--------------------- PA3301 QVTLK-----TYPEARHELFNESNRDAVTQDLIDWLEQALRHRRDHSTKERT-------- PP4551 ----------VYPEARHEVLNETNRGEVIADILGWLEQALALGRPARSE----------- PSPPH0357 ----------VIREAGHSASESGIADALVRAAAEIARNLLDLPPEEA-------------

83
PP3523 ----------MVDGLG---------PAMRFDILPK------------------------- ATu2497 ----------LLDFGA-HAVNITEPDLFNTRLLQFLLPADQT------------------ RPA3731 ----------AFDGVG-HCVAQEAPTQLLTALEEFAAA---------------------- PSPPH1999 ----------LAGGHHLHLNDEQGARSVAHCINRFFAAS--------------------- SAV0974 ----------VYSGAP-HGLTGAYEQEFNADLLNFIRG---------------------- ATu5389 ----------TYPGFS-HGMLTVNADVLNQDLLAFIKG---------------------- PA2717 ----------TYPGFP-HGMPTTQAEVINADLLAFIRG---------------------- PA3053 ----------EFPDLG-HTPQIQAPERFHQALLEGLQTQP-------------------- PP2201 ----------TFEGMG-HAPQIEEPVRFNRTLVEWLGK---------------------- SAV0923 ----------TLDATG-HCPQLSAPDATAQAITAFLGATR-------------------- ATu2061 ----------TMPGHG-HNPHWENPQGVSTLVTEFFAEVMTDASPVRVETAADKR----- SAV2105 H---------ILDGCG-HWLQQERPQDTNRLLTDWLASLPG------------------- RPA2773 L---------IIDGAG-HWIQQEKPAEVNAALIEFLKENY-------------------- SAV141 AVAPQLQGPVVLPGCG-HWTQQERPVEVNAALLDFLTRIDGSAPR--------------- RHA05146 ----------ILPGFD-HSTAVAASPEVLAAVEPFLASV--------------------- RHA00069 ----------VLDGVG-HYPHLEDPAAVVEIIDDFVSTTPART----------------- SAV1746 ----------IFEGAG-HFPFHTDPARFLALVEEFTGTTRPAHWSREHWRELLRAGRPGS PP4540 ----------VVDDSR-HATPLDQPEVFNQTLLQFLAAASTSQGSLSPC----------- PSPPH2566 ----------ILPDQR-HMMPVESPRLVNQVLLDFFEKTGLDKLATAHNSIKGIVA---- PA0231 ----------LVEFAAAHLSNVEAGDAFSRRLVDFLLSA--------------------- PP1380 ----------YVDFPAAHLSNVEIGEAFSRRVLDFLLAH--------------------- PA3586 ----------LIPGLA-HRFQEAFKEPLLAAVVPYLKAHRQASVVQL------------- PP2804 ----------LLPGMG-HDLPTCLSGTLLSLLTGHMR----------------------- SAV0519 ----------SVPGAG-HDVHLERPAVLRHLIQEFLEENAAPRGPARRPGRGRDRGPGRL SC0267 ----------VVPRTG-HLIPLEAPADLADAVTAFATAV--------------------- SC5943 ----------PIEGGR-HGIAVEFADTVAHRVREFLGVEGHG------------------ PSPPH0033 ----------LVPEAG-HIANLENPDFVSGALMTFLARVNQQQG---------------- RPA1568 ----------ILKGAG-HTMMVERPDEVLKALQQ-------------------------- SAV7198 ----------LVEDCG-HIIPLHRPDALLALLHPFLKD---------------------- SAV1548 ----------IVDAG--HELPVEKPAELTSALDRFVTGQVGGVQLVG------------- SAV4410 ----------VIDGAA-HLPNLERPEEFNAALGEFLFSLSGV------------------ TM0820 ILKLAAK------------------- TM0423 -------------------------- ATu2215 -------------------------- PP0624 -------------------------- PA3301 -------------------------- PP4551 -------------------------- PSPPH0357 -------------------------- PP3523 -------------------------- ATu2497 -------------------------- RPA3731 -------------------------- PSPPH1999 -------------------------- SAV0974 -------------------------- ATu5389 -------------------------- PA2717 -------------------------- PA3053 -------------------------- PP2201 -------------------------- SAV0923 -------------------------- ATu2061 -------------------------- SAV2105 -------------------------- RPA2773 -------------------------- SAV141 -------------------------- RHA05146 -------------------------- RHA00069 -------------------------- SAV1746 AAGRPDTVRNRAVERDLRQASERSAT PP4540 --------------------------

84
PSPPH2566 -------------------------- PA0231 -------------------------- PP1380 -------------------------- PA3586 -------------------------- PP2804 -------------------------- SAV0519 RGGGRHLQPPPPA------------- SC0267 -------------------------- SC5943 -------------------------- PSPPH0033 -------------------------- RPA1568 -------------------------- SAV7198 -------------------------- SAV1548 -------------------------- SAV4410 --------------------------

85
Appendix A.3 Amino acid sequence alignment of AF_est1 and AF_est2 with 8 bacterial esterases that were characterized in Chapter 3.
AF_est2 -----------------------------------------MTLRTKDGLTLYTRRWDVE PP_est1 -----------------------------------------------------------M RP_est1 ------------------------MLAQGQRFEGLLCGDDSSRCEVVEMAKLTQFLISG- RP_est2 -----------------------------------------MSEAVMAGAEAFAF--DGG PA_est1 MKRFLLGLVLLLAVAAGVLYFVPATLLASVRTVERGLAGLSEHSVQVDNLEIA-YLEGGS AF_est1 -----------------------------------------MPYAD-NGVKIYYEVEDGG SAV_est1 ----------------------------------MTSQHVVGRFFDVQGGQVYAHVREGD PP_est2 ----------------------------------------MSTFVTRDGTSIY-FKDWGS RP_est3 -----------------------------------------MPSFDHAGVNIA-YLDEGE PP_est3 ------------------------MACLPAAPDLQGAPSMIQPVAERTPAGTS-YLDVGQ AF_est2 SPRAVICLVHGLGEHSG------RYEHVARFFNENG-----ISF------AAFDLRGHGR PP_est1 R--NRLVLLPGWGLGTA------ALEPLAASLRAQD-----ARLQVEL--MPLPELAHS- RP_est1 ---ACALLITATSAIAD------PIAPAGKAIGQDGGGHTIYQVDANGISIGYKLIGQGA RP_est2 K--TGVLLVHGFTGSPQ------SMRYLGERLGERG-----YSI------LGPRLPGHGV PA_est1 EKNPTLLLIHGFGADKD------NWLRFARPLT-ER-----YHV------VALDLPGFGD AF_est1 E--PAIVFVHGWTANMN------FWREQREYFK-GK-----HRM------LFIDNRGHGK SAV_est1 G--PALIFLHYWGGSRR------TWIPVLQRLD-PG-----QGF------VAYDQRGWGG PP_est2 G--KPVLFSHGWPLDAD------MWDSQMEFLASRG-----YRA------IAFDRRGFGR RP_est3 G--EPILLIHGFASNKNVNWVYPSWLSELKRTG--------RRV------IAIDNRGHGE PP_est3 G--QPVVLIHGVGLNKE------MWGGQFVGLA-ND-----YRV------IAYDMLGHGQ : . . AF_est2 SE--GKRGHAEYQQLMDDITLFLQSLDYDCPKI-LYGHSM-GGNLALNYILRYDPDIAGG PP_est1 ----------DVQAWIDHLDRKL------PNDVWLGGWSL-GGMLASALAHKRGDHCCGL RP_est1 PMVMIMGLGGTADNWPPQV---IEALSKNHQLILMDNRGM-GHTTANDTPFSYPLFAADV RP_est2 SP--AAMAKTTANDWVNCAEDALLELSAKCDKTFVAGLSM-GGTLSLYLAASHPDKVAGV PA_est1 SSK-PQQASYDVGTQAERVANFAAAIG--VRRLHLAGNSM-GGHIAALYAARHPEQVLSL AF_est1 SDKPFNRSFYEFDNFVSDLHAAVKDAS--FDRFVLVGHSF-GTMISMRYCVEHPGRVEAL SAV_est1 ST--SVPGPYDLEQLADDAQRVVDALG--YSRYVLVGHSM-GGKVAQILAARKPAGLRGV PP_est2 SS--QPWNGYDYDTFADDIAQLIEHLD--LRDVTLVGFSMGGGDVSRYIARHGSERVAGL RP_est3 SSKLYDPNDYTLEAMASDAVALMDHLG--IARADVMGYSM-GARIGANLARRQEQRVRSV PP_est3 SR--VPAADTPLEGYADQLAELLDHLQ--IAQATVIGFSM-GGLVARAFALNYPQRLAAL : . .: * . AF_est2 IISAPFLALPKELPKHLFFILKLLNVVAPSIQLSNGIDPNLISRDR--EVVEAY------ PP_est1 LTLASNPSFLARPD----WPHG------MAEDTFGTFLDGCRNHTQ--VTLKRFRTL--- RP_est1 IGLLDALGIKRSDVLG--YSMGSTITQQLLLQYPDRFNKALIHATS--TDGSNVAKA--- RP_est2 IPINAAVQIDSPDLAGLAYARG-------LPEFVPGIGSDMVDTST--KELA-------- PA_est1 ALIDNAGVMPARKSE-------------LFEDLERGENPLVVRQPE--DFQKLLDFVF-- AF_est1 VLIGGGARIQSLHRYG--YPIG-----RLFATLAYGISARIIANMAFGRKAGELRDW-GL SAV_est1 VLVAPAPP----------APIG------VTEQVQETVSHAYDNEEAVLQSIDLM------ PP_est2 VLLGAVTPVFGKRDD---NPDG------VDLSVFEGIRAGLRADRA--QFIADFATPFYG RP_est3 VLGGLGMGLLSNEGRP-----G--------ENVARALEADALDDVT--DPVGRTFRAFAD PP_est3 VVLNSVFNRTPEQSAG---------------VIARAAQAAELGPDA--NVDAALDRWFSR AF_est2 ---VSDPLVHDKISPRFILQSLE------------AGKWA--LENADRLRKPILLIHGTA PP_est1 ---CSDGALQPRTLLRQLGVGVPETDPLYLATGLEVLAKLDTREALQAYDGPQLHLFAGS RP_est1 ---LHGRVPTDPIVARQVEA---------------TTHWKTPLDKLPSIDNQVMLVVGTA RP_est2 ----YDQVPVTCVKQV-------------------MGLAATARALLPRIKCPTLVINSRV PA_est1 ---VQQPPLPAPLKRYLGERAVAASAFNAQIFEQLRQRYIPLEPELPKIEAPTLLLWGDR AF_est1 KEALENTPKHAALNTLW------------------TLTTVDLRDIAREIEKPTLIVVGKE SAV_est1 ---LTRGGLTPELRRQVVEDSLRGGDEARLEWPR-RGLVQDVSAGVSAIEVPVLVLAGSH

86
PP_est2 L--NHGQQVSQGVQTQTLNIALMASIKGTLDCVT-AFSETDFRPDMAKVDVPTLVIHGDD RP_est3 QTRSDRKALAACMRGS-------------------RGLMS--REDAAQIAVPVLIAVGTD PP_est3 EYKAANPAQVAAIRQVLASNDPQGYHTTYSLF---ATQDMYRADDLGSIQVPTLIATGEL : . AF_est2 DQITSYRASQEFAKRAGELCKFVS--YEGFYHEPHNEPEKERVLADMLKWIEEVI----- PP_est1 DALVP-AEAAKALSELLPDVEVGM--VEDSSHAFLLEYPQE-LAAGIKSFLHESGDD--- RP_est1 DNVVG-TESSKTIASAIPGAWLVQ--FKGATHHLMYETPEG-FSAAALTFFETNETVTPK RP_est2 DHVLS-PANATVIANHVGSSRIETLWLDNSYHVATIDNDKDLIAAEIDSFVTRNL----- PA_est1 DRVLD-VSSIEVMRPLLKRPSVVI--MENCGHVPMVERPEE-TAQHYQAFLDGVRNAQVA AF_est1 DALLP-VSKSEELSRLIKNSKMVI--VPDAGHCVMLEQPEI-VNRVLEEFIHTFSAMLIR SAV_est1 DKVDPPTVLADHLLPLIPTATLTV--LKDTGHLSPLEVPDQ-VAAHIGAFVAQL------ PP_est2 DQIVPFETTGKQAAELIRGAELKV--YAGAPHGFAVTHAQQ-LNEDLLAFLQR------- RP_est3 DDVAG-SA--HELGDIIPGSQVLD--IPRRDHMRAV-GDRV-YKEGVVDFLARRP----- PP_est3 DSGST-PAMTRQLAACIPGARSVV--LAEQRHMMPVEAPRE-VNKMLLDFLTQARTLTES * * :. AF_est2 -------------- PP_est1 -------------- RP_est1 IEPNASVAPPPTQP RP_est2 -------------- PA_est1 GR------------ AF_est1 A------------- SAV_est1 -------------- PP_est2 -------------- RP_est3 -------------- PP_est3 AKGIVA--------

87
Appendix B.1 The progress of 208 gene targets in gene cloning, protein expression and purification, and protein activity assay.
Full Name Protein ID Cloned Expressed Purified Active
activity AF_0865 1 1 1 1 AF_1706 1 insoluble AF_1753 1 1 1 1 AF_2336 1 insoluble AF_1537 1 0
Archaeoglobus fulgidus AF_0675 1 insoluble
Atu1572 1 0 Atu1814 1 0 Atu5389 1 1 1 0 Atu3664 1 0 Atu2061 1 1 1 0 Atu2497 1 1 1 0 Atu2679 1 0
Agrobacterium tumefaciens C58 Atu2215 1 1 1 0
CA_C2688 0 CA_P0097 0 CA_P0133 0 CA_C2936 0
Clostridium acetobutylicum 824D-S CA_C0816 0
Cthe_3133 0 Clostridium thermocellum 27405D Cthe_1390 0
b2593 1 1 1 1 Escherichia coli K12 b2799 1 1 1 1
NE2298 1 1 1 1 NE0456 0 NE2295 0 NE2346 0 NE1455 0
Nitrosomonas europaea NE0279 1 1 1 1
PA2717 1 1 1 0 PA4152 1 0 PA3226 1 0 PA0480 1 0 PA2949 1 1 1 1 PA0231 1 1 1 0
Pseudomonas aerugonosa
PA3053 1 1 1 0

88
PA3586 1 1 1 0 PA3301 1 1 1 0 PA3509 1 insoluble PA3132 1 1 1 1 PA0829 1 0 PA5057 1 insoluble PA3429 1 0 PA3994 1 insoluble PA3695 1 0 PA5080 1 insoluble PA2934 1 1 1 1 PA2856 1 0
PA1469 1 1 1 1
PP5253 1 1 1 1 PP4021 1 insoluble PP3645 1 1 1 1 PP4540 1 1 1 0 PP0553 1 insoluble PP1380 1 1 1 0 PP0364 1 1 1 1 PP2201 1 1 1 0 PP4624 1 0 PP4164 1 insoluble PP5028 1 insoluble PP2804 1 1 1 0 PP1979 1 0 PP4551 1 1 1 0 PP5117 1 0 PP2318 1 insoluble PP3523 1 1 1 0
Pseudomonas putida KT2440 PP0624 1 1 1 0
PSPPH_4522 1 1 1 1 PSPPH_3123 1 insoluble PSPPH_2566 1 1 1 0 PSPPH_1434 1 0 PSPPH_0033 1 1 1 0 PSPPH_3013 1 0 PSPPH_2235 1 insoluble PSPPH_1999 1 1 1 0
Pseudomonas syringae
PSPPH_3482 1 1 1 1

89
PSPPH_0357 1 1 1 0 PSPPH_3478 1 0 PSPPH_0378 1 insoluble PSPPH_2039 1 insoluble
PSPTO_3914
1 1 1 1 RHA1_ro01401 1 1 1 1 RHA1_ro02101 0 RHA1_ro01338 1 1 1 1 RHA1_ro02074 1 insoluble RHA1_ro08044 1 1 1 1 RHA1_ro03350 1 1 1 1 RHA1_ro04327 1 0 RHA1_ro03976 1 0 RHA1_ro10146 1 1 1 1 RHA1_ro08081 1 1 1 1 RHA1_ro05146 1 1 1 0 RHA1_ro10061 1 0 RHA1_ro10040 1 insoluble RHA1_ro03603 1 1 1 1 RHA1_ro02771 1 0 RHA1_ro01727 0 RHA1_ro06181 1 0 RHA1_ro03968 1 0 RHA1_ro00069 1 1 1 0 RHA1_ro08672 1 insoluble RHA1_ro06216 1 insoluble RHA1_ro06804 1 0 RHA1_ro01528 1 0 RHA1_ro06793 1 0 RHA1_ro08833 1 0 RHA1_ro04298 0 RHA1_ro04713 0 RHA1_ro01411 1 insoluble RHA1_ro03520 1 1 1 1 RHA1_ro09014 1 insoluble RHA1_ro01171 1 insoluble RHA1_ro00320 1 1 1 1 RHA1_ro04639 1 insoluble RHA1_ro00371 1 0 RHA1_ro04540 1 insoluble
Rhodococcus sp. RHA1 RHA1_ro02607 1 1 1 1
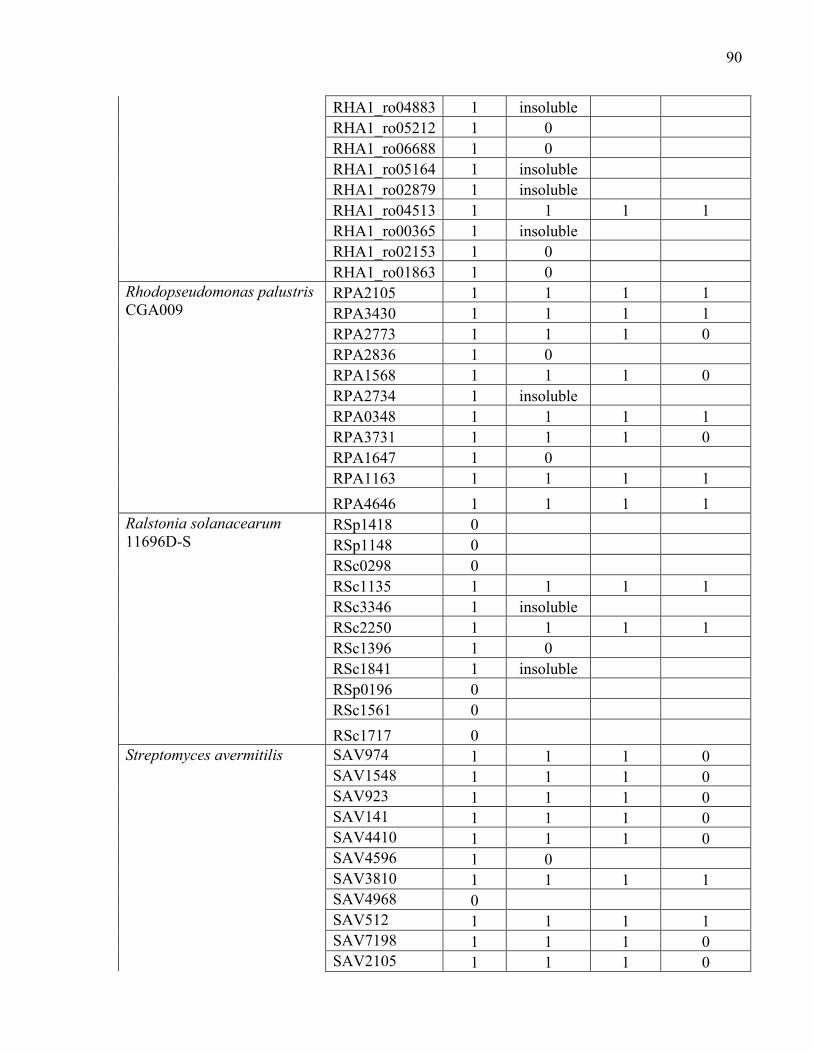
90
RHA1_ro04883 1 insoluble RHA1_ro05212 1 0 RHA1_ro06688 1 0 RHA1_ro05164 1 insoluble RHA1_ro02879 1 insoluble RHA1_ro04513 1 1 1 1 RHA1_ro00365 1 insoluble RHA1_ro02153 1 0
RHA1_ro01863 1 0 RPA2105 1 1 1 1 RPA3430 1 1 1 1 RPA2773 1 1 1 0 RPA2836 1 0 RPA1568 1 1 1 0 RPA2734 1 insoluble RPA0348 1 1 1 1 RPA3731 1 1 1 0 RPA1647 1 0 RPA1163 1 1 1 1
Rhodopseudomonas palustris CGA009 RPA4646 1 1 1 1
RSp1418 0 RSp1148 0 RSc0298 0 RSc1135 1 1 1 1 RSc3346 1 insoluble RSc2250 1 1 1 1 RSc1396 1 0 RSc1841 1 insoluble RSp0196 0 RSc1561 0
Ralstonia solanacearum 11696D-S RSc1717 0
SAV974 1 1 1 0 SAV1548 1 1 1 0 SAV923 1 1 1 0 SAV141 1 1 1 0 SAV4410 1 1 1 0 SAV4596 1 0 SAV3810 1 1 1 1 SAV4968 0 SAV512 1 1 1 1 SAV7198 1 1 1 0
Streptomyces avermitilis SAV2105 1 1 1 0

91
SAV519 1 1 1 0 SAV1256 1 0 SAV4779 1 0 SAV1746 1 1 1 0 SAV620 1 insoluble SAV3624 1 insoluble SAV5173 1 1 1 1 SAV1968 1 insoluble SAV5668 1 0 SAV298 1 insoluble SAV297 1 insoluble SAV875 1 1 1 1 SAV5711 1 insoluble SAV2164 1 0
SAV2722 1 1 1 1 SCO0465 1 insoluble SCO5943 1 1 1 0 SCO1097 1 insoluble SCO0939 1 insoluble SCO3566 1 1 1 1 SCO2886 1 insoluble SCO7245 1 insoluble SCO3690 1 1 1 1 SCO0267 1 1 1 0 SCO4746 1 0 SCO0427 1 insoluble SCO7205 1 insoluble SCO3233 1 0 SCO6277 1 insoluble SCO4479 1 insoluble SCO6697 1 insoluble SCO7440 1 1 1 1 SCO0878 1 insoluble SCO6612 1 insoluble SCO4392 1 0 SCO6966 1 0 SCO0526 1 insoluble SCO6339 1 0 SCO1989 1 1 1 1 SCO5986 1 0 SCO3171 1 insoluble
Streptomyces coelicolor SCO4395 1 insoluble

92
SCO3439 1 1 1 1 SCO6712 1 1 1 1 Sinorhizobium meliloti RA0563 1 1 1 1
TM_1350 1 1 1 1 TM_0423 1 1 1 0 TM_0820 1 1 1 0
Thermotoga maritima TM_0111 1 1 1 1 Total 208 186 86 86 48

93
Appendix B.2 The schematic representations of pNP-esters, phenyl acetate, and tributyrin.
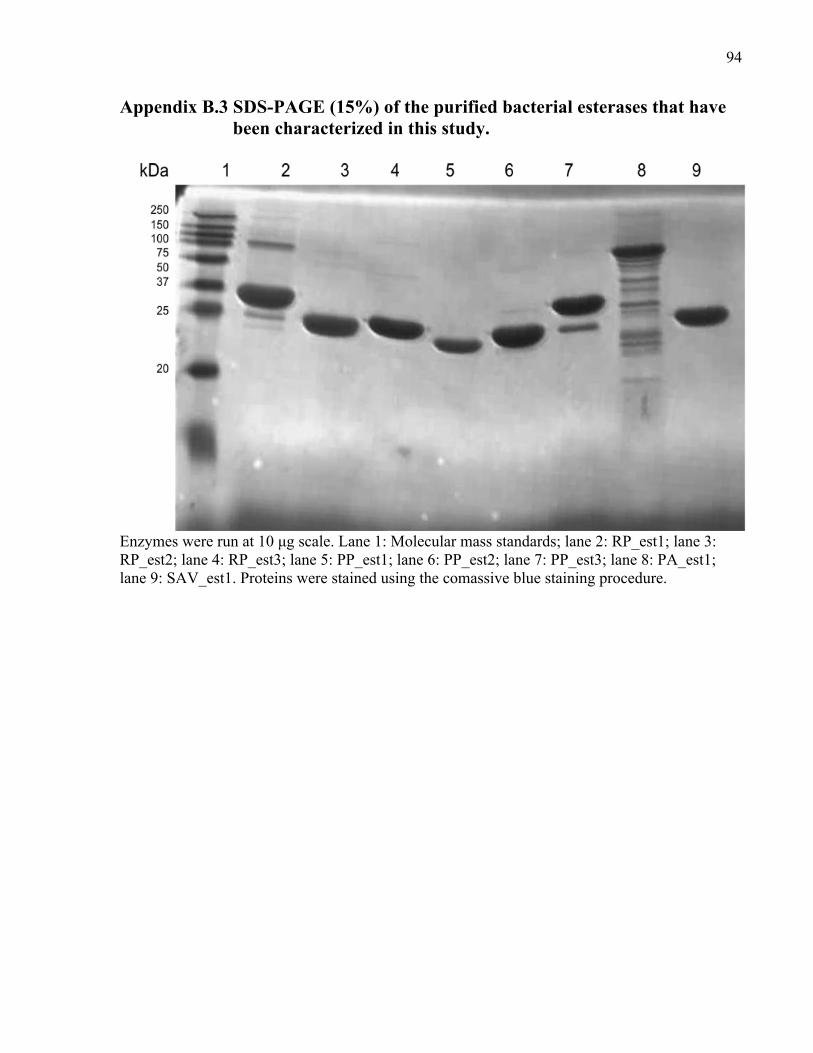
94
Appendix B.3 SDS-PAGE (15%) of the purified bacterial esterases that have been characterized in this study.
Enzymes were run at 10 µg scale. Lane 1: Molecular mass standards; lane 2: RP_est1; lane 3: RP_est2; lane 4: RP_est3; lane 5: PP_est1; lane 6: PP_est2; lane 7: PP_est3; lane 8: PA_est1; lane 9: SAV_est1. Proteins were stained using the comassive blue staining procedure.

95
Appendix B.4 A standard curve of p-nitrophenol at pH 7.

96
Appendix B.5 Absorbance reading at 400 nm of the residual activity for PP_est3 after 5 h preincubation in organic solvents.
1 2 3 4 5 6 7 8 9 10 11 12 A Buffer blank Buffer blank Control 0.1737 0.1769 0.6739 0.6693 0.6284 B Substrate blank Substrate blank Tert-amyl alcohol 0.2565 0.2523 0.745 0.7165 0.7242 0.7998 0.6134 0.6595 0.7316 0.8554 0.8678 C Std 10uM Std 10uM 1,6-Dixaone 0.2043 0.2032 0.6803 0.6681 0.6276 0.5529 0.55 0.5301 0.2705 0.2611 0.2565 D Std 25uM Std 25uM DMSO 0.2491 0.253 0.6828 0.5966 0.6485 0.6963 0.6987 0.6289 0.6984 0.7526 0.7084 E Std 50uM Std 50uM Toluene 0.3519 0.3472 0.7015 0.7666 0.7508 0.2298 0.2259 0.22 0.8923 0.9447 0.9714 F Std 75uM Std 75uM P-xylene 0.4585 0.4334 0.4335 0.4609 0.4562 0.7696 0.7875 0.8065 0.9885 1.0852 1.1407 G Std 100uM Std 100uM Hexane 0.5694 0.5393 0.2495 0.2339 0.2309 0.7768 0.797 0.7844 0.9711 1.0212 1.0343 H Std 500uM Std 500uM Cyclohexane 2.5616 2.3794 0.4979 0.4854 0.4786 0.5008 0.4886 0.4626 0.7621 1.0678 1.0508
15 % v/v organic solvent
30 % v/v organic solvent
50 % v/v organic solvent
Absorbance reading of blanks for organic solvents mixed in 50 mM potassium phosphate buffer at pH 7.
1 2 3 4 5 6 7 8 9 10 11 12 A 50 mM Potassium phosphate, pH 7 0.1533 0.1489 0.1501 B Tert-amyl alcohol

97
0.1471 0.307 0.2292 C 1,6-Dixaone 0.1387 0.1409 0.1464 D DMSO 0.1493 0.1414 0.1408 E Toluene 0.1283 0.1335 0.1369 F P-xylene 0.1287 0.1314 0.1379 G Hexane 0.1241 0.1209 0.12 H Cyclohexane 0.1337 0.1224 0.1222 15 % v/v organic solvent 30 % v/v organic solvent 50 % v/v organic solvent

98
Appendix B.6 SDS-PAGE (15%) of the purified archaeal esterases that have been characterized in this study.
Enzymes were run at 10 µg scale. Lane 1: Molecular mass standards; lane 2: AF_est1; lane 3: AF_est2. Proteins were stained using the comassive blue staining procedure.


















