
DirectTransferof -SynucleinfromNeurontoAstroglia ... · DirectTransferof...
12
Direct Transfer of -Synuclein from Neuron to Astroglia Causes Inflammatory Responses in Synucleinopathies * □ S Received for publication, October 31, 2009, and in revised form, December 23, 2009 Published, JBC Papers in Press, January 13, 2010, DOI 10.1074/jbc.M109.081125 He-Jin Lee ‡ , Ji-Eun Suk § , Christina Patrick ¶ , Eun-Jin Bae § , Ji-Hoon Cho , Sangchul Rho , Daehee Hwang , Eliezer Masliah ¶ , and Seung-Jae Lee §1 From the § Department of Biomedical Science and Technology, IBST, and ‡ Department of Anatomy, School of Medicine, Konkuk University, Seoul 143-701, Korea, the ¶ Department of Neurosciences and Pathology, School of Medicine, University of California San Diego, La Jolla, California 92093, and the School of Interdisciplinary Bioscience and Bioengineering and Department of Chemical Engineering, Pohang University of Science and Technology, Pohang, Kyoungbuk 790-784, Korea Abnormal neuronal aggregation of -synuclein is implicated in the development of many neurological disorders, including Par- kinson disease and dementia with Lewy bodies. Glial cells also show extensive -synuclein pathology and may contribute to disease progression. However, the mechanism that produces the glial -synuclein pathology and the interaction between neurons and glia in the disease-inflicted microenvironment remain unknown. Here, we show that -synuclein proteins released from neuronal cells are taken up by astrocytes through endocytosis and form inclusion bodies. The glial accumulation of -synuclein through the transmission of the neuronal protein was also demonstrated in a transgenic mouse model expressing human -synuclein. Fur- thermore, astrocytes that were exposed to neuronal -synuclein underwent changes in the gene expression profile reflecting an inflammatory response. Induction of pro-inflammatory cytokines and chemokines correlated with the extent of glial accumulation of -synuclein. Together, these results suggest that astroglial -synuclein pathology is produced by direct transmission of neu- ronal -synuclein aggregates, causing inflammatory responses. This transmission step is thus an important mediator of pathogenic glial responses and could qualify as a new therapeutic target. Parkinson disease (PD) 2 is a complex neurodegenerative dis- ease related to aging process. Clinically, it is characterized by a variety of motor and non-motor symptoms, and pathologically by selective neurodegeneration and the occurrence of Lewy bodies in central and peripheral nervous tissues (1). Evidence from genetic, pathologic, and biochemical studies suggests that abnormal deposition of the neuronal protein -synuclein produces the pathogenesis of PD (2). Aggregated forms of -synuclein are the major component of Lewy bodies (3), which are also found in other neurological diseases, such as dementia with Lewy bodies (DLB) and Alzheimer disease (AD). Further- more, recent studies showed extensive deposition of filamen- tous -synuclein aggregates in astroglia and oligodendrocytes in PD, DLB, and multiple system atrophy (4 – 8), even though these cells express very low levels of the protein (9 –11). The mechanism of inclusion body formation in neurons and glia and the role in disease onset and progression remain largely unknown. -Synuclein is a cytosolic protein with no known signal sequence. Previously, we showed that small amounts of the pro- tein, both overexpressed and endogenous, are present in the lumen of vesicles and secreted from neuronal cells by exocyto- sis (12). Both monomeric and the aggregated forms were found in vesicles and exocytosed. Monomeric and oligomeric forms of -synuclein have been detected in human cerebrospinal fluid and blood plasma (13), and the levels of the oligomeric forms were elevated in PD patients compared with control cases (14). Together, these results suggest the pathogenic actions of -synuclein extend to the extracellular space and neighboring cells. Consistent with this hypothesis, it was shown that extra- cellular exposure of cultured microglia to aggregated or nitrated -synuclein produced some signs of activated micro- glia (15, 16). More recently, it was shown that -synuclein aggregates released from neuronal cells can be transferred to neighboring neurons forming Lewy-like inclusions (17), pro- viding mechanistic basis for the development of Lewy pathol- ogy in mesencephalic transplants in PD patients (18, 19). Despite these exciting new findings, the pathophysiological roles of the secreted -synuclein remain largely unclear. Here, we report that -synuclein proteins released from neu- ronal cells are readily endocytosed by astrocytes. When the internalized -synuclein accumulates in astrocytes, the cells produce glial inclusions and inflammatory responses. There- fore, when neurons are under protein conformational stresses forming -synuclein aggregates, secreted forms of -synuclein * This work was supported, in whole or in part, by National Institutes of Health Grants AG5131 and AG18440. This work was also supported by the Disease Network Research Program (Grant 20090084180) from the National Research Foundation of Korea funded by the Ministry of Edu- cation, Science and Technology, Republic of Korea and by the Korea Science and Engineering Foundation funded by the Korea government (Grant 20090083737). The microarray data reported in this paper have been deposited to the Gene Expression Omnibus (GEO) data base under accession number GSE11574. □ S The on-line version of this article (available at http://www.jbc.org) contains supplemental Figs. S1–S8 and Tables 1– 4. 1 To whom correspondence should be addressed: Dept. of Biomedical Sci- ence and Technology, Konkuk University, 1 Hwayang-dong, Gwangjin-gu, Seoul 143-701, Korea. Tel.: 82-2-450-4166; Fax: 82-2-447-5683; E-mail: [email protected]. 2 The abbreviations used are: PD, Parkinson disease; DLB, dementia with Lewy bodies; AD, Alzheimer disease; APP, amyloid precursor protein; GFP, green fluorescent protein; DEG, differentially expressed gene; IL, interleukin; adeno, adenoviral vector; tg, transgenic; PDGF, platelet-derived growth factor; RT, reverse transcription; TNF, tumor necrosis factor; GOBP, GO Bio- logical Process; JAK, Janis kinase; STAT, signal transducers and activators of transcription; -syn, -synuclein; GFAP, glial fibrillary acidic protein; KEGG, Kyoto Encyclopedia of Genes and Genomes. THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 285, NO. 12, pp. 9262–9272, March 19, 2010 © 2010 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A. 9262 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 285 • NUMBER 12 • MARCH 19, 2010 by guest on June 5, 2018 http://www.jbc.org/ Downloaded from
Transcript of DirectTransferof -SynucleinfromNeurontoAstroglia ... · DirectTransferof...

Direct Transfer of �-Synuclein from Neuron to AstrogliaCauses Inflammatory Responses in Synucleinopathies*□S
Received for publication, October 31, 2009, and in revised form, December 23, 2009 Published, JBC Papers in Press, January 13, 2010, DOI 10.1074/jbc.M109.081125
He-Jin Lee‡, Ji-Eun Suk§, Christina Patrick¶, Eun-Jin Bae§, Ji-Hoon Cho�, Sangchul Rho�, Daehee Hwang�,Eliezer Masliah¶, and Seung-Jae Lee§1
From the §Department of Biomedical Science and Technology, IBST, and ‡Department of Anatomy, School of Medicine, KonkukUniversity, Seoul 143-701, Korea, the ¶Department of Neurosciences and Pathology, School of Medicine, University of CaliforniaSan Diego, La Jolla, California 92093, and the �School of Interdisciplinary Bioscience and Bioengineering and Department ofChemical Engineering, Pohang University of Science and Technology, Pohang, Kyoungbuk 790-784, Korea
Abnormal neuronal aggregationof�-synuclein is implicated inthe development of many neurological disorders, including Par-kinsondiseaseanddementiawithLewybodies.Glialcellsalsoshowextensive �-synuclein pathology and may contribute to diseaseprogression. However, the mechanism that produces the glial�-synuclein pathology and the interaction between neurons andglia in the disease-inflicted microenvironment remain unknown.Here, we show that �-synuclein proteins released from neuronalcells are taken up by astrocytes through endocytosis and forminclusion bodies. The glial accumulation of �-synuclein throughthe transmission of the neuronal proteinwas also demonstrated ina transgenic mouse model expressing human �-synuclein. Fur-thermore, astrocytes that were exposed to neuronal �-synucleinunderwent changes in the gene expression profile reflecting aninflammatory response. Induction of pro-inflammatory cytokinesand chemokines correlated with the extent of glial accumulationof �-synuclein. Together, these results suggest that astroglial�-synuclein pathology is produced by direct transmission of neu-ronal �-synuclein aggregates, causing inflammatory responses.This transmissionstep is thusanimportantmediatorofpathogenicglial responses and could qualify as a new therapeutic target.
Parkinson disease (PD)2 is a complex neurodegenerative dis-ease related to aging process. Clinically, it is characterized by a
variety of motor and non-motor symptoms, and pathologicallyby selective neurodegeneration and the occurrence of Lewybodies in central and peripheral nervous tissues (1). Evidencefrom genetic, pathologic, and biochemical studies suggests thatabnormal deposition of the neuronal protein �-synucleinproduces the pathogenesis of PD (2). Aggregated forms of�-synuclein are themajor component of Lewy bodies (3), whichare also found in other neurological diseases, such as dementiawith Lewy bodies (DLB) and Alzheimer disease (AD). Further-more, recent studies showed extensive deposition of filamen-tous �-synuclein aggregates in astroglia and oligodendrocytesin PD, DLB, and multiple system atrophy (4–8), even thoughthese cells express very low levels of the protein (9–11). Themechanism of inclusion body formation in neurons and gliaand the role in disease onset and progression remain largelyunknown.
�-Synuclein is a cytosolic protein with no known signalsequence. Previously, we showed that small amounts of the pro-tein, both overexpressed and endogenous, are present in thelumen of vesicles and secreted from neuronal cells by exocyto-sis (12). Bothmonomeric and the aggregated forms were foundin vesicles and exocytosed.Monomeric and oligomeric forms of�-synuclein have been detected in human cerebrospinal fluidand blood plasma (13), and the levels of the oligomeric formswere elevated in PD patients compared with control cases (14).Together, these results suggest the pathogenic actions of�-synuclein extend to the extracellular space and neighboringcells. Consistent with this hypothesis, it was shown that extra-cellular exposure of cultured microglia to aggregated ornitrated �-synuclein produced some signs of activated micro-glia (15, 16). More recently, it was shown that �-synucleinaggregates released from neuronal cells can be transferred toneighboring neurons forming Lewy-like inclusions (17), pro-viding mechanistic basis for the development of Lewy pathol-ogy in mesencephalic transplants in PD patients (18, 19).Despite these exciting new findings, the pathophysiologicalroles of the secreted �-synuclein remain largely unclear.Here, we report that�-synuclein proteins released fromneu-
ronal cells are readily endocytosed by astrocytes. When theinternalized �-synuclein accumulates in astrocytes, the cellsproduce glial inclusions and inflammatory responses. There-fore, when neurons are under protein conformational stressesforming �-synuclein aggregates, secreted forms of �-synuclein
* This work was supported, in whole or in part, by National Institutes ofHealth Grants AG5131 and AG18440. This work was also supported bythe Disease Network Research Program (Grant 20090084180) from theNational Research Foundation of Korea funded by the Ministry of Edu-cation, Science and Technology, Republic of Korea and by the KoreaScience and Engineering Foundation funded by the Korea government(Grant 20090083737).
The microarray data reported in this paper have been deposited to the GeneExpression Omnibus (GEO) data base under accession number GSE11574.
□S The on-line version of this article (available at http://www.jbc.org) containssupplemental Figs. S1–S8 and Tables 1– 4.
1 To whom correspondence should be addressed: Dept. of Biomedical Sci-ence and Technology, Konkuk University, 1 Hwayang-dong, Gwangjin-gu,Seoul 143-701, Korea. Tel.: 82-2-450-4166; Fax: 82-2-447-5683; E-mail:[email protected].
2 The abbreviations used are: PD, Parkinson disease; DLB, dementia with Lewybodies; AD, Alzheimer disease; APP, amyloid precursor protein; GFP, greenfluorescent protein; DEG, differentially expressed gene; IL, interleukin;adeno, adenoviral vector; tg, transgenic; PDGF, platelet-derived growthfactor; RT, reverse transcription; TNF, tumor necrosis factor; GOBP, GO Bio-logical Process; JAK, Janis kinase; STAT, signal transducers and activators oftranscription; �-syn, �-synuclein; GFAP, glial fibrillary acidic protein; KEGG,Kyoto Encyclopedia of Genes and Genomes.
THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 285, NO. 12, pp. 9262–9272, March 19, 2010© 2010 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A.
9262 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 285 • NUMBER 12 • MARCH 19, 2010
by guest on June 5, 2018http://w
ww
.jbc.org/D
ownloaded from

potentially act as a messenger between neurons and astroglia,inducing neuroinflammatory responses.
EXPERIMENTAL PROCEDURES
Materials—All-trans-retinoic acid, proteinase K, TritonX-100, thioflavin S, and protease inhibitor mixture were pur-chased from Sigma. Bafilomycin A1 was purchased fromCalbiochem (San Diego, CA). The Dynamin K44A/pCB1vector was a kind gift from Dr. Marc Caron. Recombinanthuman wild-type �-synuclein was purchased from ATGen(Sungnam, Korea). The following primary antibodies wereused: �-synuclein polyclonal antibody (Cell Signaling Tech-nology, Beverly, MA), �-synuclein monoclonal antibody(BD Biosciences, San Diego, CA), Myc polyclonal antibody(Abcam, Cambridge, MA), anti-dynamin antibody (BD Bio-sciences), anti-GFPN terminus antibody (Cell SignalingTechnol-ogy), anti-GFAP antibody (Abcam), anti-S100 antibody(Abcam),anti-GLUT1 antibody (Abcam), anti-Hsp70/Hsc70 antibody(Stressgen,Victoria,BritishColumbia,Canada), anti-20Santibody(Calbiochem), anti-ubiquitin antibody (Chemicon, Temecula,CA), anti-IL-1� antibody (Abcam), anti-IL-1� antibody (Abcam),anti-IL-6 antibody (Abcam), and anti-�-actin antibody. A mono-clonal antibody for LAMP2 (H4B4) developedbyDrs.August andHildreth and �-tubulin antibody (E7) were obtained from theDevelopmental Studies Hybridoma Bank developed under theauspices of the NICHD, National Institutes of Health and main-tained by The University of Iowa, Dept. of Biological Sciences,Iowa City, IA.Cell Culture and �-Synuclein Expression—Maintenance of
the human neuroblastoma cell line SH-SY5Y and differentia-tion of SH-SY5Y cells were previously described (20). Primaryastrocytes were obtained from 1-day-old Sprague-Dawley ratsas previously described (21) and maintained in Dulbecco’smodified Eagle’s medium/F-12 medium (Invitrogen) with 10%fetal bovine serum (HyClone Laboratories) and penicillin/streptomycin (Invitrogen). Briefly, dissected cortices withoutmeninges were broken into smaller pieces by pipetting in Dul-becco’s modified Eagle’s medium/F-12 medium, and trypsinwas added. After DNaseI digestion, mediumwas added to inac-tivate trypsin, and cells were centrifuged at 500 � g for 2 min.The cell pellet was resuspended with fresh medium before fil-tration through a 120-�mNylon mesh filter. Cells were grownin poly-D-lysine-coated flasks for 3–4 days before splitting intonew flasks. On day 7 after preparation, cells grown in T75 flaskswere shaken vigorously at least 15 times to detach microgliacells from astrocyte layer and removed. Cells were cultured foranother 1�2 weeks, split onto new dishes when confluent,before they were used for experiments. Contaminant microgliacells in these cultures were 2.3 � 0.09%.For �-synuclein expression in differentiated SH-SY5Y cells,
a recombinant adenoviral vector containing human �-synuclein cDNA (adeno/�-syn), �-galactosidase (adeno/lacZ,supplied from BD Biosciences), and C-terminal-tagged human�-synuclein (adeno/�-syn-MycHis) were used as previouslydescribed (20, 22). For co-culture of cells, primary astrocyteswere added to SH-SY5Y cells infected with adeno/�-syn,adeno/lacZ, or adeno/�-syn-MycHis on day 3 of infection andco-cultured for 3 days or as indicated. For expression of
dynamin K44A and GFP, primary astrocytes were transfectedwith dynamin K44A/pCB1 or pEGFP (Clontech) using Lipo-fectamine 2000 before culturing with SH-SY5Y cells.Inhibition of Internalized �-Synuclein Degradation—For
inhibition of internalized�-synuclein degradation, bafilomycinA1 was added to a final concentration of 50 nM on day 2 ofinfection for 15 h.Preparation of Cell Extract and Conditioned Medium—For
extraction of cellular proteins, cells were washed twice in ice-cold PBS and harvested in PBS containing 1% triton X-100 andprotease inhibitor mixture. After incubation on ice for 10 min,the cell extract was centrifuged at 16,000� g for 10min and thetriton-soluble (supernatant) and -insoluble (pellet) fractionswere obtained.For collection of the conditioned medium, differentiated
SH-SY5Y cells infected with adeno/�-syn were used. On day 2of infection, themediumwas replacedwith serum-free Dulbec-co’s modified Eagle’s medium after washing twice with Dulbec-co’s modified Eagle’s medium. After overnight incubation at37 °C, the conditioned medium was collected and centrifugedat 1,000 � g for 10 min, and the supernatant was centrifugedfurther at 10,000 � g to remove debris and dead cells. Therecovered supernatant was concentrated 50- to 100-fold usingAmicon 10K MWCO filters (Millipore, Billerica, MA). Theconcentrations of non-tagged and Myc-tagged �-synuclein inthe conditioned medium measured by enzyme-linked immu-nosorbent assay were 4.23 � 1.2 �g/ml and 83.1 � 27.3 ng/ml,respectively. The concentrations of �-synuclein in themedia ofrat primary neuron cultures were measured by quantitativeWestern blotting with recombinant human �-synuclein stan-dards and were approximately �3 ng/ml.Depletion of �-Synuclein from Conditioned Medium—The
conditioned medium from polyhistidine-tagged �-synuclein-overexpressing SH-SY5Ycellswas prepared as described above.The conditioned medium was incubated with Talon beads (BDBiosciences) to deplete �-synuclein, and the beads were re-moved by centrifugation.Western Blotting—Western blotting was performed using a
monoclonal anti �-synuclein antibody (Syn-1, BD Biosciences)as previously described (22). Images were obtained and quanti-fied using a luminescent image analyzer LAS-3000 (Fujifilm,Japan) and Multi Gauge (v3.0) software.
�-Synuclein Transgenic Mice and Treatments—For thisstudy, a total of 12 heterozygous transgenic (tg) mice (Line D)expressing human �-synuclein under the regulatory control ofthe platelet-derived growth factor-� (PDGF�) promoter (23) (6months old) and 12 non-transgenic age-matched littermateswere used. These animals were selected because they displayabnormal accumulation of detergent-insoluble �-synucleinand develop �-synuclein-immunoreactive inclusion-like struc-tures in the brain. Although some nuclear staining has beenobserved in this model, distinct cytoplasmic inclusion-likestructures have been consistently identified by confocal andelectron microscopy (23, 24). Furthermore, these animals alsodisplay neurodegenerative and motor deficits that mimic cer-tain aspects of PD and DLB. Comparisons for the patterns of�-synuclein distribution were performed in the brains of ourmodel mimicking AD-like pathology by expressing the human
Neuron-to-Astroglia Transmission of �-Synuclein
MARCH 19, 2010 • VOLUME 285 • NUMBER 12 JOURNAL OF BIOLOGICAL CHEMISTRY 9263
by guest on June 5, 2018http://w
ww
.jbc.org/D
ownloaded from

mutated amyloid precursor protein (APP tg, line 41) (n � 6, 6months old) under the thy1 promoter (25). In addition, studiesof the re-distribution of �-synuclein were performed in thePDGF�-syn-GFP tg mouse model (line 78) (n � 6, 6 monthsold) (26), because in thesemice the fused GFP protein expectedto reduce the neuronal release of �-synuclein compared thePGDF�-syn tg lines (line D). Animal protocols were approvedby the University of California San Diego Institutional AnimalUse and Care Committee.Cases and Neuropathological Evaluation—For this study
we included a total of 18 cases (for detail see supplemen-tal Table S4); of these, 4 were non-demented controls, 8 wereDLB cases, and the other 6 were AD cases. Autopsy materialwas obtained from patients studied neurologically and psycho-metrically at the Alzheimer Disease Research Center/Univer-sity of California, San Diego.For each case, paraffin sections from 10% buffered formalin-
fixed neocortical, limbic system, and subcortical materialstained with hematoxylin and eosin and thioflavin-S were usedfor routine neuropathological analysis (27) that included Braakstage (28). The diagnosis of DLB was based on the clinical pres-entation of dementia and the pathological findings of Lewybodies in the locus coeruleus, substantia nigra, or nucleus basa-lis of Meynert, as well as in cortical and subcortical regions.Lewy bodies were detected using hematoxylin and eosin anti-ubiquitin and anti-�-synuclein antibodies as recommended bythe Consortium on DLB criteria for a pathologic diagnosis ofDLB (29). In addition to the presence of Lewy bodies, the greatmajority of these cases display sufficient plaques and tangles tobe classified as Braak stages III–IV. Specifically, DLB cases hadabundant plaques in the neocortex and limbic system but fewertangles compared with AD cases.Immunohistochemistry and Laser Scanning Confocal Micro-
scopy—The procedure for immunofluorescence cell staininghas been described elsewhere (22). Nuclei were stained withTOPRO-3 dye (Invitrogen), and the coverslips were mountedon slides in Antifade reagent (Invitrogen). Cells were observedwith an Olympus FV1000 confocal laser scanning microscope.For three-dimensional analysis of images, serial images throughz axis were obtained and analyzed using FV10-ASW software(Olympus).For immunohistochemistry of tissue sections, free-floating
40-�m-thick vibratome sections were washed with Tris-buff-ered saline (pH 7.4), pre-treated in 3% H2O2, and blocked with10% serum (Vector), 3% bovine serum albumin (Sigma), and0.2% gelatin in Tris-buffered saline-Triton X-100. Sectionswere incubated at 4 °C overnight with the rabbit polyclonalantibody against human �-synuclein (Chemicon), and thenincubated with secondary antibody (1:75, Vector Laboratories,Burlingame, CA), followed by avidin D-horseradish peroxidase(ABC Elite, Vector) and reacted with diaminobenzidine (0.2mg/ml) in 50 mM Tris (pH 7.4) with 0.001% H2O2. Controlexperiments consisted of incubation with pre-immune rabbitserum.Double-immunofluorescence analyses of tissue sectionswere performed utilizing the Tyramide Signal AmplificationTM-Direct (Red) system (PerkinElmer Life Sciences). Specificity ofthis system was tested by deleting each primary antibody. Forthis purpose, sections were double labeled with the mono-
clonal antibodies against �-synuclein (1:20,000, Cell Signaling)detected with Tyramide Red and either S100 detectedwith fluorescein isothiocyanate-conjugated secondary antibod-ies (1:75, Vector). All sections were processed simultaneouslyunder the same conditions, and experiments were performedtwice for reproducibility. Sections were imaged with a Zeiss63� (numerical aperture, 1.4) objective on an Axiovert 35microscope (Zeiss, Germany) with an attachedMRC1024 laserscanning confocal microscope system (Bio-Rad).In Situ Hybridization—In situ hybridization was carried out
using human �-synuclein sense and antisense Riboprobeslabeledwith 35S, essentially as described previously (30). Briefly,paraffin sections were treated with Proteinase K, incubated inthe prehybridization solution, and hybridized with Riboprobesat 60 °C. Sections were exposed to film and developed 3 days to2weeks later. These sectionswere then dipped inKodakNTB-2emulsion and developed 10 days later.RNA Preparation and RT-PCR—Total RNA was prepared
from primary astrocytes using the RNeasy mini kit (Qiagen).The RNA was used for microarray and RT-PCR analysis. ForRT-PCR, 2 �g of total RNAwas used to synthesize cDNA usingiScript cDNA synthesis kit (Bio-Rad) following the manufac-turer’s protocol. Two microliters of synthesized cDNA wasused for PCR using ImmomixRed (Bioline) with primers andreaction temperatures as indicated in supplemental Table 3.Cytokine Assay—Rat 8-plex cytokine kit was obtained from
Panomics (Fremont, CA). Primary astrocytes grown in a12-well plate were treated with LacZ or �-synuclein condi-tioned medium for 24 h. The medium from astrocytes werecollected and centrifuged at 10,000 � g for 20 min, and thesupernatant was obtained. Quantification of TNF� andCXCL1was performed according to the manufacturer’s instructions.The result was read using a Bio-Plex 200 system (Bio-Rad).Microarray Analysis—The integrity of the total RNA ex-
tracted from primary astrocytes was assessed using a Bioana-lyzer 2100 (Agilent, Santa Clara, CA), and the RNA integritynumbers were 10 for all samples. 500 ng of high quality totalRNA was amplified with a TotalPrep amplification kit(Ambion, Austin, TX), and 750 ng of amplified cRNA washybridized to the RatRef-12 expression BeadChip for 16 h. Thehybridized array was scanned with a BeadArray Reader (Illu-mina). The microarray data are deposited in a GEO data baseunder accession number GSE11574.Statistical Analysis of the Microarray Data—Using the Illu-
mina RatRef-12 Expression BeadChip, we obtained expressionlevels for nearly 22,000 genes in astrocytes exposed for 6 and24 h to conditioned media from cultures of �-synuclein andlacZ-expressing cells. There were three array replicates donefor each condition (astrocytes treated for 6 and 24 h with lacZand �-synuclein containing conditioned media). The probeintensitieswere normalized at log2 scale using quantile normal-ization in BeadArray 1.6, an R/Bioconductor package. Probeannotation was done using lumi 1.4, an R/Bioconductor pack-age providing an integrated solution for Illumina microarraydata analysis. Before identifying differentially expressed genes,we first applied a Gaussian mixture modeling technique to thenormalized data to determine present/absent calls as follows. Inthe mixture modeling, 1) two Gaussian probability density
Neuron-to-Astroglia Transmission of �-Synuclein
9264 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 285 • NUMBER 12 • MARCH 19, 2010
by guest on June 5, 2018http://w
ww
.jbc.org/D
ownloaded from

functions, one for absent and the other for present probes, werefitted to the distribution of the observed probe intensity and 2)among the 22,523 probes, 11,039 probes, whose maximumintensities in all 12 samples were higher than the thresholdintensity, where two fitted Gaussian probability density func-tions meet, were determined to be present.Then, we identified differentially expressed genes (DEGs)
between astrocytes treated with �-synuclein and lacZ contain-ing conditioned media independently for two time points (6and 24 h) by an integrative statistical test combining the p val-ues from three statistical hypothesis testing methods: Studenttwo-tailed t test, Wilcoxon rank-sum test, and median differ-ence test. In each test, an empirical null probability densityfunction was estimated for the corresponding statistic (T, ranksum, and median difference) computed from all possible per-mutations of the samples. A p value for the observed statistic ofeach probe was then computed using the empirical probabilitydensity function. For each probe, p values resulting from threestatistical tests were combined as an overall p value using metaanalysis (Stouffer’s method) as described in Hwang et al. (31).DEGs were selected as the genes with overall p values of �0.01.Also, false positives from potential bias toward p values fromparticular hypothesis testing in meta-analysis were reduced byremoving genes that showed -fold differences � 1.5 betweensamples treated with �-synuclein and lacZ containing condi-tioned media. We performed a functional enrichment analysisto associate the DEGs with GO Biological Processes (GOBPs),using BiNGO, a Cytoscape Plugin, and KEGG pathways, usingthe Database for Annotation, Visualization and Integrated Dis-covery (see Ref. 32). Several GOBPs and KEGG pathways sta-tistically enriched (p � 0.05) by the DEGs were identified (seesupplemental Tables 1 and 2). Finally, network analysis wasdone using Cytoscape (33) with information of the DEGs asso-ciated with GOBPs and KEGG pathways related to inflamma-tory responses and known protein-protein interactions fromBIND andHPRD (both available on-line). Human andmouseprotein-protein interactions were translated into those in ratvia ortholog ID mapping using ortholog information in theMouse Genome Informatics (MGI) data base (availableon-line).Statistical Analyses of Quantified Data—Values in the fig-
ures are expressed asmeans� S.E. Differenceswere consideredsignificant if p values were �0.05. For determination of statis-tical significance, values were compared by one-way analysis ofvariance with Tukey’s post test or Student’s t test using Graph-Pad InStat version 1.01 GraphPad software. The correlationcoefficients (Pearson r) and the p values were determined bylinear regression analysis.
FIGURE 1. Uptake of neuronal cell-derived �-synuclein from conditionedmedium by astrocytes. A, Western analysis of �-synuclein uptake in primaryastrocytes. �-Synuclein-containing conditioned medium was obtained fromSH-SY5Y culture and added to the primary astrocytes for the indicated times.Tx-s, Triton X-100-soluble; Tx-in, Triton X-100-insoluble. B, immunofluores-cence staining of primary astrocytes (GLUT1-positive) treated with the condi-tioned media used in C. Top panels: LacZ medium. Middle panels: Myc-His-tagged �-synuclein (�-synMH) medium. Bottom panels: depleted medium. Aperinuclear inclusion body produced by the internalized �-synuclein is indi-cated by an arrow in the middle panel. Scale bars: 20 �m. Experiments wererepeated three times. C, depletion of �-synMH from the conditioned medium
of differentiated SH-SY5Y cells expressing �-synMH. D, uptake of neuronalcell-derived �-synuclein by astrocytes. Conditioned media from LacZ- or�-synMH-expressing cells, or the latter with �-synMH depleted were incu-bated with rat astrocytes for the indicated times. Note that both monomericand aggregated forms (represented by the smear between the 37- and 180-kDa markers) of �-synuclein were taken up from the �-synMH medium.
Neuron-to-Astroglia Transmission of �-Synuclein
MARCH 19, 2010 • VOLUME 285 • NUMBER 12 JOURNAL OF BIOLOGICAL CHEMISTRY 9265
by guest on June 5, 2018http://w
ww
.jbc.org/D
ownloaded from

RESULTS
Transmission of�-Synuclein toAstrocytes and Inclusion BodyFormation—Recent studies show that cytoplasmic �-synucleindeposition occurs in both neurons and astrocytes in PD (4, 6).Considering the fact that astrocytes express no or low levels of�-synuclein on their own and are able to take up recombinant�-synuclein aggregates from the culturemedia (34), we hypoth-esized that neuronal �-synuclein aggregates are transmitted toadjacent astrocytes and form pathological inclusion bodies. Totest this hypothesis, the conditioned medium containingsecreted �-synuclein from differentiated SH-SY5Y cells wasadded to primary astrocytes. The secreted �-synuclein wasreadily internalized by the astrocytes, as judged by Westernblotting (Fig. 1A) and immunofluorescence microscopy (Fig.1B). The presence of �-synuclein inside astrocytes was verifiedby proteinase K resistance (supplemental Fig. S1). Internalizedproteins were found in both the Triton-soluble and -insolublefractions, and a portion of these appeared as SDS-stablehigh molecular weight complexes (Fig. 1A and supple-mental Fig. S2), suggesting internalized �-synuclein maintainsits aberrant conformation after internalization. Some astro-cytes accumulated the internalized �-synuclein in large jux-
tanuclear inclusion bodies (Fig. 1B).Approximately 30% of cells took up�-synuclein, and �4.6% of thesecells showed large inclusion bodies.As a control, when �-synuclein wasdepleted from the conditionedmedium (Fig. 1C), this protein wasnot detected in astrocytes (Fig. 1, Band D). These results exclude thepossibility of transmission of �-synuclein vector to the astrocytesand demonstrate that neuronal cell-derived �-synuclein can be taken upby astrocytes and accumulate ininclusion bodies.Next, transmission of �-synuclein
from neuronal cells to astrocyteswas directly demonstrated in a co-culture system consisting of pri-mary rat astrocytes and differenti-ated SH-SY5Y cells expressinghuman �-synuclein (Fig. 2 andsupplemental Fig. S3). Antibodiesspecific to the human sequence of�-synuclein enabled detection ofneuronal �-synuclein only, andthe rat astrocytes were identifiedby themarkers GFAP, GLUT1, andS100. In this co-culture, human�-synuclein was observed in the ratastrocytes, consistent with transmis-sion of �-synuclein from the neuro-blastoma (supplemental Fig. S3).Z-stack imaging of these cells con-firmed the presence of internalized�-synuclein in the astroglial cyto-
plasm (Fig. 2,A andB). Co-culture of rat astrocyteswith SH-SY5Ycells expressingLacZgenedidnot show�-synucleinaccumulationin astrocytes (supplemental Fig. S3D). Transmission of endoge-nous neuronal �-synuclein from the primary cortical neurons toastrocytes was also demonstrated (Fig. 2C).
The transferwas time-dependent andwidespread; by day 5 ofco-culture, �80% of astrocytes received neuronal cell-derived�-synuclein (Fig. 2, D and E, and supplemental Fig. S3E).Changes in the distribution of internalized �-synuclein werealso observed over time. Initially, the internalized �-synucleinwas distributed in vesicular patterns, which evolved to largerpunctates and finally showed large perinuclear inclusions.Among the �-synuclein-positive astrocytes, up to 10% of cellsdeveloped round inclusion bodies (Fig. 2E). Expression of�-synuclein in SH-SY5Y neuroblastoma cells was maintainedduring this 5-day period (supplemental Fig. S4).The �-synuclein-positive perinuclear inclusions shared
some characteristics with Lewy bodies and PD-associatedastroglial inclusions (Fig. 3). More than 90% of the astrocyteperinuclear inclusions were stained with thioflavin S, an indi-cator of fibrillar aggregates (Fig. 3,A and E). A large proportionof these inclusions contained some of the proteins found in
FIGURE 2. Cell-to-cell transmission of �-synuclein to primary astrocytes in co-culture. A and B, primaryastrocytes were co-cultured with differentiated SH-SY5Y cells overexpressing �-synuclein. Serial imagesthrough z axis were obtained and analyzed to show that transmitted �-synuclein (green) reside inside ofastrocytes (red: GFAP). Areas in the white boxes are enlarged (left panels, xy images; right panels, z-stackedimages seen through the y axis). C, transfer of neuronal endogenous �-synuclein to astrocytes. Rat primarycortical neurons were co-cultured with astrocytes. Note that the �-synuclein polyclonal antibody used in thisexperiment also recognizes rat sequences. Top panels: primary astrocytes cultured with neurons. Bottom pan-els: primary astrocytes only. White perforated lines indicate outlines of astrocyte periphery. D, primary astrocyteswere cultured with differentiated SH-SY5Y cells overexpressing �-synuclein and fixed at the indicated times.The control indicates a co-culture with SH-SY5Y cells without �-synuclein overexpression. Arrows: perinuclearinclusions. E, quantitative analyses of �-synuclein transfer (left graph) and glial inclusion formation (rightgraph). Differentiated SH-SY5Y cells overexpressing human �-synuclein were co-cultured with rat primaryastrocytes. The transfer was measured by fluorescence intensities of �-synuclein staining in astrocytes, and theinclusion formation was measured by the percentage of astrocytes with inclusions among the cells with trans-ferred �-synuclein. Experiments were repeated four times, and more than hundred cells were counted perslide. All scale bars, 20 �m (*, p � 0.05; **, p � 0.01; ***, p � 0.001).
Neuron-to-Astroglia Transmission of �-Synuclein
9266 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 285 • NUMBER 12 • MARCH 19, 2010
by guest on June 5, 2018http://w
ww
.jbc.org/D
ownloaded from

Lewy bodies, such as ubiquitin and the 20 S proteasome �-sub-unit, and to a lesser degree, Hsp/Hsc70 (Fig. 3, B–E). Theseresults suggest that neuron-to-astrocyte transmission of aber-rant �-synuclein may be a mechanism of astroglial �-synucleindeposition in PD and other Lewy body diseases.To determine if the transfer involved endocytosis, astrocytes
were transfected with the dominant negative dynamin-1 K44Amutant, which inhibits endocytic vesicle formation (35), andco-cultured with SH-SY5Y cells expressing �-synuclein (Fig. 4,A–C). The �-synuclein fluorescence intensity in astrocytes wasreduced �4-fold in dynamin-1 K44A-expressing cells, com-pared with non-transfected cells, consistent with endocy-tosis-mediated �-synuclein transmission. As a control, GFPtransfection had no effect on �-synuclein transmission(supplemental Fig. S5). Further analysis of internalized�-synuclein showed that most of the �-synuclein co-localizedwith the lysosome marker LAMP-2 (Fig. 4D). This indicatesthat internalized �-synuclein moves through the endosomalpathway and is normally destined to be degraded in thelysosome.Neuron-to-Glia �-Synuclein Transmission and Astroglial
Inclusion Body Formation in Vivo—To verify the neuron-to-astrocyte transmission in a physiological context, we exam-ined the brain sections of tg mice overexpressing human�-synuclein under neuronal promoter (PDGF�). In non-tgcontrols and PDGF-APP tg mice (see “Experimental Proce-dures”), �-synuclein immunolabeling was localized to the pre-synaptic terminals in the neuropil (Fig. 5A). Although abundantglial cells are present around the plaques in the APP tg model,�-synuclein accumulated in the dystrophic neurites but not inastroglial cells. In the PDGF�-syn tg mice, abundant�-synuclein accumulation was observed not only in neuronsbut also in glial cells (Fig. 5A). This glial accumulation of�-synuclein was more abundant in the deeper layers of thecortex and along the myelinated axons. The accumulation of�-synuclein in astroglial cells was considerably suppressed inPDGF�-syn-green fluorescent protein (GFP) tg mice (Fig. 5, Aand B). In this line, most of the �-synuclein labeling wasrestricted to axons and neuronal cell bodies. This is consis-
tent with our in vitro studies showing that tagging �-synuclein with a variant of GFP reduces the secretion of�-synuclein.3 Previously, we have shown that, unlike
3 A. Jang, H.-J. Lee, J.-E. Suk, J.-W. Jung, K.-P. Kim, and S.-J. Lee, submitted forpublication.
FIGURE 3. Characterization of astroglial �-synuclein inclusions. A–D, inclu-sion bodies in co-cultured primary astrocytes were stained with thioflavin S (A) orwith antibodies for ubiquitin (B), Hsp70/Hsc70 (C), and 20 S proteasome �-sub-unit (D). Inclusion bodies that are positive for the markers are indicated witharrowheads, and the ones negative are marked with arrows. Scale bars, 20 �m.E, quantitative analysis of inclusion bodies for each marker. Experiments wererepeated three times, and more than one hundred cells were counted per slide.
FIGURE 4. Endocytosis-dependent transfer of �-synuclein to primaryastrocytes. A–C, effects of dynamin-1 K44A mutant. Primary astrocytes weretransfected with a dominant negative K44A mutant of dynamin-1 and co-cultured with �-synuclein-expressing SH-SY5Y cells. The images in A show anexample of astrocyte expressing dynamin K44A with little �-synuclein trans-fer. The images in B show an astrocyte on the same coverslip without dynaminK44A expression, but with transferred �-synuclein (arrow). The graph in Cshows quantitative analysis of the transferred �-synuclein fluorescence inK44A-expressing (�) or untransfected (�) astrocytes. Error bars represent S.E.(****, p � 0.0001). D, co-staining of the transferred �-synuclein with the lateendosomal/lysosomal marker Lamp-2 in astrocytes in co-culture. Arrows indi-cate the co-staining of �-synuclein and Lamp-2. The bottom images are theenlarged views of the blue rectangle area in the upper images. Scale bars, 20�m. Experiments were repeated four times, and more than one hundred cellswere counted per slide.
Neuron-to-Astroglia Transmission of �-Synuclein
MARCH 19, 2010 • VOLUME 285 • NUMBER 12 JOURNAL OF BIOLOGICAL CHEMISTRY 9267
by guest on June 5, 2018http://w
ww
.jbc.org/D
ownloaded from

�-synuclein, little �-synuclein was secreted from cells (12).Consistently, in �-synuclein tg mice, both the endogenous�-synuclein and �-synuclein were exclusively localized tothe neurons and no glial immunoreactivity was detected(Fig. 5A).Similar to the findings in the mouse models, �-synuclein
immunoreactivity in the temporal cortex from human controland AD cases was present in the presynaptic terminals in theneuropil and in dystrophic neurites around the plaques, respec-tively (supplemental Fig. S6). In contrast, in DLB cases�-synuclein immunoreactivity was present in the Lewy bodiesand astroglial cells in the deeper layers of the neocortex and thewhite-gray matter junction. Double labeling and confocalmicroscopy studies confirmed that both in DLB cases as well as
in PDGF�-syn tg mice, glial cellsexpressing S100 and GFAP dis-played �-synuclein accumulation(Fig. 5, B and D). In contrast, no�-synuclein was detected in astro-cytes in the PDGF�-syn-GFP tgmice. To rule out the possibility oftransgene expression in astrocytesin PDGF�-syn tg mice, the cell typedistribution of human �-synucleinmRNAs were examined by in situhybridization. Consistent with theuse of neuron-specific promoter,human �-synuclein mRNAs werefoundmostly in neurons, not in glialcells (Fig. 5, C and D). Together,these results verify that astroglialinclusion bodies are formed by theaccumulation of �-synuclein pro-teins transmitted from neurons.Gene Expression Changes in
Astrocytes—To determine how as-trocytes respond to extracellular�-synuclein, gene expression pro-files were established and analyzedfor nearly 22,000 rat genes usingthe Illumina RatRef-12 ExpressionBeadChip. Total RNA was isolatedfrom astrocytes exposed for 6 or24 h to conditioned medium fromcultures of �-synuclein or lacZ-ex-pressing cells. About 1,116 DEGswere identified (supplementalFig. S7). The expression of abouthalf of these genes was up-regulated(red), whereas the other half showeddown-regulation (Fig. 6A). Manyinflammatory cytokines, such asIL-1�, -1�, and -6, and CC, CXC,CX3C-type chemokines, showedincreased expression. These cyto-kines typically are involved in theinflammatory responses in the cen-tral nervous system. Other DEGs
are involved in antigen processing and presentation, toll-likereceptor signaling, apoptosis, mitogen-activated protein kinasesignaling, the cell cycle, complement cascade, and leukocytetransendothelial migration, and many of these are importantfor brain immune responses (see supplemental Tables 1 and 2and supplemental Fig. S8).To confirm the inflammatory response, we constructed a
hypothetical protein network (Fig. 6B), using the inflamma-tion-related DEGs from the current study and informationabout protein-protein interactions from BIND andHPRD. Thenetwork node proteins were clustered into several networkmodules that represent the enrichedGOBP terms and enrichedKEGG pathways. The astroglial protein network was perturbedby exposure to neuronal cell-derived �-synuclein and showed
FIGURE 5. In vivo evidence for neuron-to-glia transmission of �-synuclein in tg models. A, immunocyto-chemical distribution of �-synuclein in the neocortical deeper layers in non-tg control, PDGF-APP tg, PDGF�-syn tg, PDGF�-syn-GFP, and �-syn tg. In the APP tg mice �-synuclein immunoreactivity is localized in dystro-phic neurites around the plaque (arrow), whereas in the PDGF�-syn tg, �-synuclein accumulates both inneuronal (arrows) and glial cells (*). No glial accumulation of �-synuclein was detected in PDGF�-syn-GFP and�-syn tg. Experiments were performed in triplicate, and the representative images are shown. B, double label-ing and confocal microscopy analysis, showing that, in DLB cases as well as in PDGF�-syn tg mice, glial cellsexpressing S100 displayed human �-synuclein accumulation (arrows). In contrast, in the PDGF�-syn-GFP tgmice no GFP signal (associated with �-synuclein) was detected in S100-positive cells. At least three indepen-dent experiments were performed, and the representative images are shown. C, in situ hybridization for thedetection of human �-synuclein mRNA. Human �-synuclein is expressed mostly in neurons (top and middlepanels). D, quantification of data represented in B (top panel) and C (bottom panel). For counting of glial cellswith accumulated �-synuclein (top panel), PDGF�-syn tg mice (n � 6), PDGF�-syn-GFP mice (n � 4), DLB cases(n � 8), and control cases (n � 6) were analyzed. The graph on the bottom panel presents % cells containinghuman �-synuclein mRNA. Six mice of each group were analyzed. For each mouse 2 sections were evaluated,and from each section 5 fields, on average, 75–100 cells were counted. White bars, NeuN-positive neurons;black bars, S100-positive astrocytes. Scale bars, 20 �m (**, p � 0.01; ***, p � 0.001).
Neuron-to-Astroglia Transmission of �-Synuclein
9268 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 285 • NUMBER 12 • MARCH 19, 2010
by guest on June 5, 2018http://w
ww
.jbc.org/D
ownloaded from

up-regulation of several cellular processes associated withinflammation and innate immunity. The modules involved inthis network include Toll-like receptor signaling, cytokines andchemokines, antigen processing and presentation, cell migra-tion, and the JAK-STAT pathway. Based on this network, inastrocytes exposed to neuron-derived �-synuclein aggregates,Toll-like receptor signaling appears to be activated, as evi-denced by the increased expression of TLR2, CD14, NFKB1/2,and RELA. Activated NF-�B complexes potentially induce the
transcription of several cytokines and chemokines (IL1B, IL6,CCL3/4/5, and CXCL4/10/11/12) (36), which in turn activaterepresentative components of the inflammation and innateimmunity-related modules. These include: (i) antigen proc-essing and presentation (PSME1/2, TAP1/2, TAPBP, CD74,CTSB, and MHC1/II), (ii) cell migration, (ICAM1, VCAM1,CXCL12, NCF1C, CYBA, and MMP9), and (iii) the JAK-STAT pathway activated by the up-regulated interleukins(37–40).
FIGURE 6. Gene expression changes in astrocytes exposed to neuronal cell-derived �-synuclein. A, a heatmap showing expression patterns of the identifiedDEGs at both 6 and 24 h. The colors represent the relative increase (red) and decrease (green) in expression levels of the corresponding genes in primary astrocytestreated with �-synuclein-containing conditioned medium from SH-SY5Y cultures, compared with those treated with the conditioned medium from LacZ-expressingcells. B, a hypothetical protein network constructed with some of the identified DEGs enriched by GOBPs or KEGG pathways. The network node colors represent theincrease (red) or the decrease (green) in expression levels of the corresponding genes. The center and border colors of the network node proteins represent the geneexpression changes at 6 and 24 h, respectively. The genes selected as DEGs are indicated by bold-italic fonts. The solid and dotted lines represent the direct and indirectprotein-protein interactions, respectively. Transcriptional regulations are represented by blue double-dashed lines. The shaded regions represent biological functionalmodules. C, RT-PCR analysis of the DEGs. Total RNAs were obtained from primary astrocytes treated with the conditioned media from LacZ (cont) or �-synuclein(�S)-expressing SH-SY5Y cultures. Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) and �-actin were used as control. Numbers on the right are the sizes of PCRproducts in base pairs. D, correlation between transmitted �-synuclein and cytokine induction in astrocytes. After immunofluorescence labeling, fluorescenceintensities per �m2 of cytokines and transmitted �-synuclein in each individual astrocyte were plotted, and linear regression was used (open circle, IL-1�; blue triangle,IL-1�; and red circle, IL-6).
Neuron-to-Astroglia Transmission of �-Synuclein
MARCH 19, 2010 • VOLUME 285 • NUMBER 12 JOURNAL OF BIOLOGICAL CHEMISTRY 9269
by guest on June 5, 2018http://w
ww
.jbc.org/D
ownloaded from
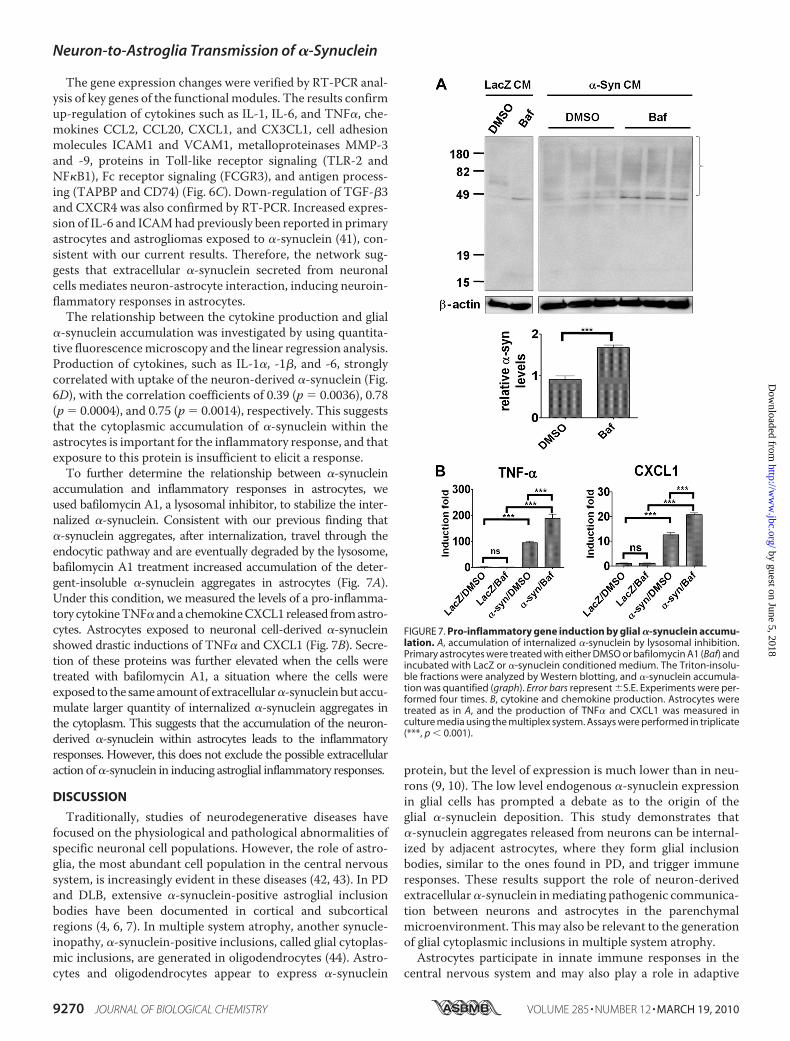
The gene expression changes were verified by RT-PCR anal-ysis of key genes of the functional modules. The results confirmup-regulation of cytokines such as IL-1, IL-6, and TNF�, che-mokines CCL2, CCL20, CXCL1, and CX3CL1, cell adhesionmolecules ICAM1 and VCAM1, metalloproteinases MMP-3and -9, proteins in Toll-like receptor signaling (TLR-2 andNF�B1), Fc receptor signaling (FCGR3), and antigen process-ing (TAPBP and CD74) (Fig. 6C). Down-regulation of TGF-�3and CXCR4 was also confirmed by RT-PCR. Increased expres-sion of IL-6 and ICAMhad previously been reported in primaryastrocytes and astrogliomas exposed to �-synuclein (41), con-sistent with our current results. Therefore, the network sug-gests that extracellular �-synuclein secreted from neuronalcells mediates neuron-astrocyte interaction, inducing neuroin-flammatory responses in astrocytes.The relationship between the cytokine production and glial
�-synuclein accumulation was investigated by using quantita-tive fluorescencemicroscopy and the linear regression analysis.Production of cytokines, such as IL-1�, -1�, and -6, stronglycorrelated with uptake of the neuron-derived �-synuclein (Fig.6D), with the correlation coefficients of 0.39 (p � 0.0036), 0.78(p � 0.0004), and 0.75 (p � 0.0014), respectively. This suggeststhat the cytoplasmic accumulation of �-synuclein within theastrocytes is important for the inflammatory response, and thatexposure to this protein is insufficient to elicit a response.To further determine the relationship between �-synuclein
accumulation and inflammatory responses in astrocytes, weused bafilomycin A1, a lysosomal inhibitor, to stabilize the inter-nalized �-synuclein. Consistent with our previous finding that�-synuclein aggregates, after internalization, travel through theendocytic pathway and are eventually degraded by the lysosome,bafilomycin A1 treatment increased accumulation of the deter-gent-insoluble �-synuclein aggregates in astrocytes (Fig. 7A).Under this condition, we measured the levels of a pro-inflamma-tory cytokineTNF� andachemokineCXCL1released fromastro-cytes. Astrocytes exposed to neuronal cell-derived �-synucleinshowed drastic inductions of TNF� and CXCL1 (Fig. 7B). Secre-tion of these proteins was further elevated when the cells weretreated with bafilomycin A1, a situation where the cells wereexposed to the sameamountof extracellular�-synucleinbut accu-mulate larger quantity of internalized �-synuclein aggregates inthe cytoplasm. This suggests that the accumulation of the neuron-derived �-synuclein within astrocytes leads to the inflammatoryresponses. However, this does not exclude the possible extracellularaction of�-synuclein in inducing astroglial inflammatory responses.
DISCUSSION
Traditionally, studies of neurodegenerative diseases havefocused on the physiological and pathological abnormalities ofspecific neuronal cell populations. However, the role of astro-glia, the most abundant cell population in the central nervoussystem, is increasingly evident in these diseases (42, 43). In PDand DLB, extensive �-synuclein-positive astroglial inclusionbodies have been documented in cortical and subcorticalregions (4, 6, 7). In multiple system atrophy, another synucle-inopathy, �-synuclein-positive inclusions, called glial cytoplas-mic inclusions, are generated in oligodendrocytes (44). Astro-cytes and oligodendrocytes appear to express �-synuclein
protein, but the level of expression is much lower than in neu-rons (9, 10). The low level endogenous �-synuclein expressionin glial cells has prompted a debate as to the origin of theglial �-synuclein deposition. This study demonstrates that�-synuclein aggregates released from neurons can be internal-ized by adjacent astrocytes, where they form glial inclusionbodies, similar to the ones found in PD, and trigger immuneresponses. These results support the role of neuron-derivedextracellular�-synuclein inmediating pathogenic communica-tion between neurons and astrocytes in the parenchymalmicroenvironment. This may also be relevant to the generationof glial cytoplasmic inclusions in multiple system atrophy.Astrocytes participate in innate immune responses in the
central nervous system and may also play a role in adaptive
FIGURE 7. Pro-inflammatory gene induction by glial �-synuclein accumu-lation. A, accumulation of internalized �-synuclein by lysosomal inhibition.Primary astrocytes were treated with either DMSO or bafilomycin A1 (Baf) andincubated with LacZ or �-synuclein conditioned medium. The Triton-insolu-ble fractions were analyzed by Western blotting, and �-synuclein accumula-tion was quantified (graph). Error bars represent �S.E. Experiments were per-formed four times. B, cytokine and chemokine production. Astrocytes weretreated as in A, and the production of TNF� and CXCL1 was measured inculture media using the multiplex system. Assays were performed in triplicate(***, p � 0.001).
Neuron-to-Astroglia Transmission of �-Synuclein
9270 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 285 • NUMBER 12 • MARCH 19, 2010
by guest on June 5, 2018http://w
ww
.jbc.org/D
ownloaded from

immunity (45). Although evidence is emerging to support a rolefor astrocytic neuroinflammatory responses in neurodegenera-tive disease, the endogenous triggers of the disease-associatedimmune responses remain elusive. Our study has identified anovel pathway that links defects in a specific neuronal protein,�-synuclein, to pathological and neuroinflammatory astrocyteresponses, including induction of cytokines and chemokines aswell as cell adhesionmolecules involved in cellmigration.How-ever, this study does not exclude the potential contribution ofother proteins co-secreted with �-synuclein, although thesecreted proteins from the control cells expressing LacZ genedo not show such effects. The glial pathology and associatedglial responses are also typical features of “tauopathies,” includ-ing AD, progressive supranuclear palsy, corticobasal degenera-tion, and Pick disease (46–50). Therefore, the work presentedhere potentially identifies a fundamental principle of a widespectrum of neurological diseases.Chronic inflammation is a critical component in progression of
manyneurodegenerative diseases, includingPD.Microglia are themajor resident immune cells of the central nervous systemandareresponsible toa largeextent for elevated inflammatory reactions indisease-inflictedbrains (51, 52). Recent studies show that extracel-lular treatmentwith aggregated recombinant�-synuclein causes avariety of inflammatory responses in rodent and humanmicroglia(15, 53–55).However, it remains tobe addressedwhetherneuron-derived �-synuclein can cause similar microglial responses. Inaddition to thedirect effect of�-synucleinonmicroglia, astrocytesmaymediatemicroglial activation. In a transgenicmousemodel ofamyotrophic lateral sclerosis, astrocyte-specific deletion of themutant superoxide dismutase 1 gene did not affect disease onset,but it significantly delayed microglial activation and late diseaseprogression (56). This implies that astrocytes control the timing ofmicroglial activation, leading to accelerated neuronal damage anddisease progression. These results warrant future studies of thecommunication between astrocytes and microglia in inflamma-tory reactions triggered by neuron-derived �-synuclein.In our previous study, we suggested that endocytosis of extra-
cellular�-synucleinaggregates ispartof its clearancemechanisms,based on the fact that the internalized proteins traffic along theendosomal pathway and eventually are degraded in the lyso-some (35). The current study shows that astrocytes take upneuro-nal cell-derived �-synuclein by endocytosis and target the proteinto the lysosome. This suggests that neuron-to-glia transmission of�-synuclein is anormal processwhereby astroglia clear potentiallyharmful protein aggregates from the parenchymal microenviron-ment. Recent studies have shown astrocyte-mediated clearance ofthe A� protein, which implies that astrocytes serve as potentialelimination sites for toxic extracellular protein aggregates (57, 58).However, in case of sustained exposure to protein conformationalstresses, neuronal release of �-synuclein aggregates could persistto the extent that it exceeds the capacity of glial cell lysosomaldegradation, causing theaggregates toaccumulate in theglial cyto-plasm. Indeed, we showed that lysosomal inhibition led to theincreased accumulation of �-synuclein aggregates in astrocytes.Importantly, the glial inflammatory responses correlate with theextent of �-synuclein aggregate deposition, further emphasizingthe significance of aggregate clearance in astrocytes.
Release of neuronal �-synuclein is promoted under stress con-ditions and can occur without apparent membrane damage; thisoccurs via vesicle-mediated exocytosis under the conditionwheremembrane leakage is undetectable.3 However, high level expres-sionof�-synucleindidcauseneuronal cell deatheventually, and insuch a situation, the neuron-to-astroglia transmission would bemore robust. Therefore, although transmissionof�-synuclein canoccur without apparent cell death, degenerating neurons acutelyprovide large amounts of �-synuclein and therebymay acceleratethe transmission. Because neurons are progressively degeneratingin disease-inflicted brain, we speculate that �-synuclein proteinsreleased fromdegeneratingneuronsmightplaya significant role inpropagation of �-synuclein pathology.In conclusion, �-synuclein proteins released from neurons
may be the source of astroglial �-synuclein pathology and animportant mediator of astroglial inflammatory responses.�-Synuclein is a typical cytosolic protein, and therefore, discus-sions on its role in the pathogenesis of PD and other synucle-inopathies have largely been limited within the neurons thathighly express this protein. However, we propose an entirelynovel non-cell autonomous mechanism of action for�-synuclein in disease onset and progression. The proposedmodel can explain some of the unexpected, yet significantexperimental findings such as the protective effects of�-synuclein immunization in a transgenic mouse model (24).Our study provides the critical knowledge base for futuremechanistic studies and development of immunization thera-pies. Recently, neuron-to-neuron transmission of �-synucleinaggregates has been demonstrated in tissue cultures and intransgenic mice with neuronal progenitor cell grafts (17).Transferred �-synuclein induces Lewy-like inclusion bodiesand apoptotic changes in the recipient neurons. Therefore,neuron-derived �-synuclein, perhaps when in excess, is trans-ferred and accumulates in neurons and glia, inducing patholog-ical inclusion formation and degenerative changes. These datacan explain the host-to-graft propagation of Lewy-like pathol-ogy in long term mesencephalic transplants in PD (18, 19) andthe stereotypic topographical progression of Lewy pathology inPD suggested by Braak and colleagues (59). This non-cellautonomous transmission behavior of disease-linked aggrega-tion-prone proteins has also been reported with polyglutamineproteins and Tau (60–62), and the templated transmission ofthe conformational properties has been demonstrated withrecombinant Tau and �-synuclein proteins (63, 64). Therefore,increasing our knowledge of cell-to-cell transmission of�-synuclein between brain cells can provide new insights intothe mechanism of disease progression and identify moleculartargets for diagnosis and therapeutic intervention in PD andother synucleinopathies.
Acknowledgments—We thank Junghwa Seo, Lei Cho, and JongminKim for technical assistance and Hyunjung Lim for consultation onimage handling.
REFERENCES1. Dauer, W., and Przedborski, S. (2003) Neuron 39, 889–9092. Cookson, M. R. (2005) Annu. Rev. Biochem. 74, 29–523. Spillantini, M. G., Crowther, R. A., Jakes, R., Hasegawa, M., and Goedert,
Neuron-to-Astroglia Transmission of �-Synuclein
MARCH 19, 2010 • VOLUME 285 • NUMBER 12 JOURNAL OF BIOLOGICAL CHEMISTRY 9271
by guest on June 5, 2018http://w
ww
.jbc.org/D
ownloaded from

M. (1998) Proc. Natl. Acad. Sci. U.S.A. 95, 6469–64734. Braak, H., Sastre, M., and Del Tredici, K. (2007) Acta Neuropathol. 114,
231–2415. Terada, S., Ishizu, H., Yokota, O., Tsuchiya, K., Nakashima, H., Ishihara,
T., Fujita, D., Ueda, K., Ikeda, K., and Kuroda, S. (2003)Acta Neuropathol.105, 163–169
6. Wakabayashi, K., Hayashi, S., Yoshimoto,M., Kudo, H., and Takahashi, H.(2000) Acta Neuropathol. 99, 14–20
7. Croisier, E., and Graeber, M. B. (2006) Acta Neuropathol. 112, 517–5308. Tu, P. H., Galvin, J. E., Baba,M., Giasson, B., Tomita, T., Leight, S., Nakajo,
S., Iwatsubo, T., Trojanowski, J. Q., and Lee, V.M. (1998)Ann. Neurol. 44,415–422
9. Miller, D. W., Johnson, J. M., Solano, S. M., Hollingsworth, Z. R., Stan-daert, D. G., and Young, A. B. (2005) J. Neural Transm. 112, 1613–1624
10. Mori, F., Tanji, K., Yoshimoto, M., Takahashi, H., and Wakabayashi, K.(2002) Exp. Neurol. 176, 98–104
11. Solano, S. M., Miller, D. W., Augood, S. J., Young, A. B., and Penney, J. B.,Jr. (2000) Ann. Neurol. 47, 201–210
12. Lee, H. J., Patel, S., and Lee, S. J. (2005) J. Neurosci. 25, 6016–602413. El-Agnaf, O. M., Salem, S. A., Paleologou, K. E., Cooper, L. J., Fullwood,
N. J., Gibson, M. J., Curran, M. D., Court, J. A., Mann, D. M., Ikeda, S.,Cookson, M. R., Hardy, J., and Allsop, D. (2003) FASEB J. 17, 1945–1947
14. El-Agnaf, O. M., Salem, S. A., Paleologou, K. E., Curran, M. D., Gibson,M. J., Court, J. A., Schlossmacher,M.G., andAllsop,D. (2006)FASEB J.20,419–425
15. Zhang,W.,Wang, T., Pei, Z.,Miller, D. S.,Wu, X., Block,M. L.,Wilson, B.,Zhang, W., Zhou, Y., Hong, J. S., and Zhang, J. (2005) FASEB J. 19,533–542
16. Reynolds, A.D., Glanzer, J. G., Kadiu, I., Ricardo-Dukelow,M., Chaudhuri,A., Ciborowski, P., Cerny, R., Gelman, B., Thomas,M. P.,Mosley, R. L., andGendelman, H. E. (2008) J. Neurochem. 104, 1504–1525
17. Desplats, P., Lee, H. J., Bae, E. J., Patrick, C., Rockenstein, E., Crews, L.,Spencer, B., Masliah, E., and Lee, S. J. (2009) Proc. Natl. Acad. Sci. U.S.A.106, 13010–13015
18. Kordower, J. H., Chu, Y., Hauser, R. A., Freeman, T. B., andOlanow, C.W.(2008) Nat. Med. 14, 504–506
19. Li, J. Y., Englund, E., Holton, J. L., Soulet, D., Hagell, P., Lees, A. J., Lashley,T., Quinn, N. P., Rehncrona, S., Bjorklund, A., Widner, H., Revesz, T.,Lindvall, O., and Brundin, P. (2008) Nat. Med. 14, 501–503
20. Lee, H. J., Khoshaghideh, F., Patel, S., and Lee, S. J. (2004) J. Neurosci. 24,1888–1896
21. Shin, C. Y., Choi, J.W., Ryu, J. R., Ryu, J. H., Kim,W., Kim,H., andKo, K.H.(2001) Brain Res. 902, 198–204
22. Lee, H. J., Shin, S. Y., Choi, C., Lee, Y. H., and Lee, S. J. (2002) J. Biol. Chem.277, 5411–5417
23. Masliah, E., Rockenstein, E., Veinbergs, I., Mallory, M., Hashimoto, M.,Takeda, A., Sagara, Y., Sisk, A., and Mucke, L. (2000) Science 287,1265–1269
24. Masliah, E., Rockenstein, E., Adame, A., Alford,M., Crews, L., Hashimoto,M., Seubert, P., Lee,M., Goldstein, J., Chilcote, T., Games, D., and Schenk,D. (2005) Neuron 46, 857–868
25. Rockenstein, E., Mallory, M., Mante, M., Alford, M., Windisch, M.,Moessler, H., and Masliah, E. (2002) J. Neural Transm. Suppl., 327–336
26. Rockenstein, E., Schwach, G., Ingolic, E., Adame, A., Crews, L.,Mante,M.,Pfragner, R., Schreiner, E., Windisch, M., and Masliah, E. (2005) J. Neuro-sci. Res. 80, 247–259
27. Hansen, L. A., Daniel, S. E., Wilcock, G. K., and Love, S. (1998) J. Neurol.Neurosurg. Psychiatry 64, 653–656
28. Braak, H., and Braak, E. (1991) Acta Neuropathol. 82, 239–25929. McKeith, I. G., Galasko, D., Kosaka, K., Perry, E. K., Dickson, D.W., Hansen,
L.A., Salmon,D. P., Lowe, J.,Mirra, S. S., Byrne, E. J., Lennox,G.,Quinn,N. P.,Edwardson, J. A., Ince, P. G., Bergeron, C., Burns, A., Miller, B. L., Lovestone,S., Collerton, D., Jansen, E. N., Ballard, C., de Vos, R. A., Wilcock, G. K.,Jellinger, K. A., and Perry, R. H. (1996)Neurology 47, 1113–1124
30. Rockenstein, E., Mallory, M., Hashimoto, M., Song, D., Shults, C. W.,Lang, I., and Masliah, E. (2002) J. Neurosci. Res. 68, 568–578
31. Hwang, D., Rust, A. G., Ramsey, S., Smith, J. J., Leslie, D.M.,Weston, A. D.,de Atauri, P., Aitchison, J. D., Hood, L., Siegel, A. F., and Bolouri, H. (2005)
Proc. Natl. Acad. Sci. U.S.A. 102, 17296–1730132. Dennis, G., Jr., Sherman, B. T., Hosack, D. A., Yang, J., Gao, W., Lane,
H. C., and Lempicki, R. A. (2003) Genome Biol. 4, P333. Shannon, P., Markiel, A., Ozier, O., Baliga, N. S., Wang, J. T., Ramage, D.,
Amin, N., Schwikowski, B., and Ideker, T. (2003) Genome Res. 13,2498–2504
34. Lee, H. J., Suk, J. E., Bae, E. J., and Lee, S. J. (2008) Biochem. Biophys. Res.Commun. 372, 423–428
35. Lee, H. J., Suk, J. E., Bae, E. J., Lee, J. H., Paik, S. R., and Lee, S. J. (2008) Int.J. Biochem. Cell Biol. 40, 1835–1849
36. Pahl, H. L. (1999) Oncogene 18, 6853–686637. Wright, K. L., White, L. C., Kelly, A., Beck, S., Trowsdale, J., and Ting, J. P.
(1995) J. Exp. Med. 181, 1459–147138. Bien, S., Ritter, C. A., Gratz, M., Sperker, B., Sonnemann, J., Beck, J. F., and
Kroemer, H. K. (2004)Mol. Pharmacol. 65, 1092–110239. Herrmann, F., Trowsdale, J., Huber, C., and Seliger, B. (2003) Immunoge-
netics 55, 379–38840. Ossendorp, F., Fu, N., Camps,M., Granucci, F., Gobin, S. J., van den Elsen,
P. J., Schuurhuis, D., Adema, G. J., Lipford, G. B., Chiba, T., Sijts, A.,Kloetzel, P. M., Ricciardi-Castagnoli, P., and Melief, C. J. (2005) J. Immu-nol. 174, 7815–7822
41. Klegeris, A., Giasson, B. I., Zhang, H., Maguire, J., Pelech, S., andMcGeer,P. L. (2006) FASEB J. 20, 2000–2008
42. Lobsiger, C. S., and Cleveland, D.W. (2007)Nat. Neurosci. 10, 1355–136043. Maragakis, N. J., and Rothstein, J. D. (2006) Nat. Clin. Pract. Neurol. 2,
679–68944. Wakabayashi, K., Hayashi, S., Kakita, A., Yamada, M., Toyoshima, Y., Yo-
shimoto, M., and Takahashi, H. (1998) Acta Neuropathol. 96, 445–45245. Farina, C., Aloisi, F., and Meinl, E. (2007) Trends Immunol. 28, 138–14546. Arai, T., Ikeda, K., Akiyama, H., Shikamoto, Y., Tsuchiya, K., Yagishita, S.,
Beach, T., Rogers, J., Schwab, C., and McGeer, P. L. (2001) Acta Neuro-pathol. 101, 167–173
47. Arima, K., Nakamura, M., Sunohara, N., Ogawa, M., Anno, M., Izumiyama,Y., Hirai, S., and Ikeda, K. (1997)Acta Neuropathol. 93, 558–566
48. Ferrer, I., Blanco, R., Carmona, M., and Puig, B. (2001) J. Neural Transm.108, 1397–1415
49. Komori, T. (1999) Brain Pathol. 9, 663–67950. Schofield, E., Kersaitis, C., Shepherd, C. E., Kril, J. J., and Halliday, G. M.
(2003) Brain 126, 827–84051. Block, M. L., Zecca, L., and Hong, J. S. (2007)Nat. Rev. Neurosci. 8, 57–6952. McGeer, P. L., and McGeer, E. G. (2008)Mov. Disord 23, 474–48353. Klegeris, A., Pelech, S., Giasson, B. I.,Maguire, J., Zhang,H.,McGeer, E.G.,
and McGeer, P. L. (2008) Neurobiol. Aging 29, 739–75254. Su, X., Maguire-Zeiss, K. A., Giuliano, R., Prifti, L., Venkatesh, K., and
Federoff, H. J. (2008) Neurobiol. Aging 29, 1690–170155. Thomas,M. P., Chartrand, K., Reynolds, A., Vitvitsky, V., Banerjee, R., and
Gendelman, H. E. (2007) J. Neurochem. 100, 503–51956. Yamanaka, K., Chun, S. J., Boillee, S., Fujimori-Tonou, N., Yamashita, H.,
Gutmann, D. H., Takahashi, R., Misawa, H., and Cleveland, D. W. (2008)Nat. Neurosci. 11, 251–253
57. Koistinaho,M., Lin, S.,Wu, X., Esterman,M., Koger, D., Hanson, J., Higgs,R., Liu, F., Malkani, S., Bales, K. R., and Paul, S. M. (2004) Nat. Med. 10,719–726
58. Wyss-Coray, T., Loike, J. D., Brionne, T. C., Lu, E., Anankov, R., Yan, F.,Silverstein, S. C., and Husemann, J. (2003) Nat. Med. 9, 453–457
59. Braak, H., Ghebremedhin, E., Rub, U., Bratzke, H., and Del Tredici, K.(2004) Cell Tissue Res. 318, 121–134
60. Clavaguera, F., Bolmont, T., Crowther, R. A., Abramowski, D., Frank, S.,Probst, A., Fraser, G., Stalder, A. K., Beibel,M., Staufenbiel,M., Jucker,M.,Goedert, M., and Tolnay, M. (2009) Nat. Cell Biol. 11, 909–913
61. Frost, B., Jacks, R. L., and Diamond, M. I. (2009) J. Biol. Chem. 284,12845–12852
62. Ren, P. H., Lauckner, J. E., Kachirskaia, I., Heuser, J. E., Melki, R., andKopito, R. R. (2009) Nat. Cell Biol. 11, 219–225
63. Frost, B., Ollesch, J., Wille, H., and Diamond, M. I. (2009) J. Biol. Chem.284, 3546–3551
64. Yonetani,M.,Nonaka, T.,Masuda,M., Inukai, Y.,Oikawa, T.,Hisanaga, S.,and Hasegawa, M. (2009) J. Biol. Chem. 284, 7940–7950
Neuron-to-Astroglia Transmission of �-Synuclein
9272 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 285 • NUMBER 12 • MARCH 19, 2010
by guest on June 5, 2018http://w
ww
.jbc.org/D
ownloaded from

Daehee Hwang, Eliezer Masliah and Seung-Jae LeeHe-Jin Lee, Ji-Eun Suk, Christina Patrick, Eun-Jin Bae, Ji-Hoon Cho, Sangchul Rho,
Responses in Synucleinopathies-Synuclein from Neuron to Astroglia Causes InflammatoryαDirect Transfer of
doi: 10.1074/jbc.M109.081125 originally published online January 13, 20102010, 285:9262-9272.J. Biol. Chem.
10.1074/jbc.M109.081125Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
Supplemental material:
http://www.jbc.org/content/suppl/2010/01/13/M109.081125.DC1
http://www.jbc.org/content/285/12/9262.full.html#ref-list-1
This article cites 63 references, 20 of which can be accessed free at
by guest on June 5, 2018http://w
ww
.jbc.org/D
ownloaded from


















