
Direct conversion of human fibroblasts into functional ... · Direct conversion of human...
6
Direct conversion of human fibroblasts into functional osteoblasts by defined factors Kenta Yamamoto a,b , Tsunao Kishida a , Yoshiki Sato a,b , Keisuke Nishioka a , Akika Ejima a , Hiroyoshi Fujiwara c , Toshikazu Kubo c , Toshiro Yamamoto b , Narisato Kanamura b , and Osam Mazda a,1 Departments of a Immunology, b Dental Medicine, and c Orthopedics, Kyoto Prefectural University of Medicine, Kamigyo-ku, Kyoto 602-8566, Japan Edited by Darwin J. Prockop, Texas A&M Health Science Center, Temple, TX, and approved March 24, 2015 (received for review October 30, 2014) Osteoblasts produce calcified bone matrix and contribute to bone formation and remodeling. In this study, we established a proce- dure to directly convert human fibroblasts into osteoblasts by trans- ducing some defined factors and culturing in osteogenic medium. Osteoblast-specific transcription factors, Runt-related transcription factor 2 (Runx2), and Osterix, in combination with Octamer-binding transcription factor 3/4 (Oct4) and L-Myc (RXOL) transduction, con- verted ∼80% of the fibroblasts into osteocalcin-producing cells. The directly converted osteoblasts (dOBs) induced by RXOL displayed a similar gene expression profile as normal human osteoblasts and contributed to bone repair after transplantation into immunodefi- cient mice at artificial bone defect lesions. The dOBs expressed en- dogenous Runx2 and Osterix, and did not require continuous expression of the exogenous genes to maintain their phenotype. Another combination, Oct4 plus L-Myc (OL), also induced fibroblasts to produce bone matrix, but the OL-transduced cells did not express Osterix and exhibited a more distant gene expression profile to osteoblasts compared with RXOL-transduced cells. These findings strongly suggest successful direct reprogramming of fibroblasts into functional osteoblasts by RXOL, a technology that may provide bone regeneration therapy against bone disorders. osteoblasts | regenerative medicine | direct reprogramming O steoblasts play a central role in bone formation and remod- eling by producing type I collagen, osteopontin, osteocalcin, and bone sialoprotein (BSP), and calcifying these bone matrixes (1). They are also involved in hematopoiesis, phosphate metabo- lism, and glucose metabolism (2). Osteoblasts are derived from mesenchymal progenitor cells that are common precursors shared by chondrocytes, adipocytes, and myoblasts (3). The differentiation of osteoblasts is regulated by various transcription factors, in- cluding Runt-related transcription factor 2 [Runx2, also known as core-binding factor subunit α-1 (Cbfα-1)] (4, 5), Osterix (6, 7), Distal-less homeobox 5 (Dlx5) (8), and activation transcription factor 4 (ATF-4) (8). A functional decline in osteoblasts relative to osteoclasts results in imbalance between bone formation and re- sorption and may cause osteolytic pathological conditions, such as osteoporosis (9), alveolar bone resorption associated with perio- dontitis (10), and bone lysing associated with bone tumors, in- cluding multiple myeloma (11). It has been demonstrated that forced expression of combinations of some transcription factors, such as Octamer-binding transcrip- tion factor 3/4 (Oct4), Sox2, Klf-4, and c-Myc (reprogramming factors), induces immortality and pluripotency in mammalian so- matic cells (12, 13). The generation of induced pluripotent stem (iPS) cells clearly indicates that genome-wide epigenetic pro- gramming can be drastically changed in somatic cells by a small number of transcription factors that may have key regulatory roles in cell fate decisions (14, 15). Recent studies have reported that direct conversion, or direct reprogramming, of somatic cells into another differentiated line- age can be achieved without passing an intermediate pluripotent stage by introducing a set of transcription factors that are pivotal for development of the destination cells. These conversions in- clude direct conversion of murine fibroblasts into cardiomyocytes (16, 17), neurons (18–20), chondrocytes (21), and hepatocytes (22, 23), as well as of human fibroblasts into cardiomyocytes (24, 25), neurons (19, 26), and hematopoietic cells (27). Although the efficiencies of conversion are generally low at present, with only 0.005–30% of fibroblasts successfully converted into the desired cells (16–20, 22–24, 26), this technology may become quite useful for regenerative therapy against a variety of human diseases. To date, however, there have been no reports on the induction of human osteoblasts from another differentiated somatic cell lineage by means of direct reprogramming procedures. If this can be achieved, then a sufficient number of osteoblasts may be obtained from the somatic cells of a patient with osteolytic disease and then transplanted back into this patient. Moreover, such cell- based therapies might be applied to other clinical settings as well, such as repair of large bone defect caused by trauma or by surgical resection of osteosarcoma or metastatic bone tumors. In the present study, we attempted to directly convert human fibroblasts into osteoblasts by genetically introducing osteoblast- specific transcription factors and reprogramming factors. We also assessed the capability of the resulting directly converted osteoblasts (dOBs) to contribute to bone formation in vivo. Results Human Fibroblasts Were Induced to Produce Calcified Bone Matrix by Defined Factors. We focused on two transcription factors, Runx-2 (R) and Osterix (X), which are critical regulators of osteoblast Significance Osteoblasts produce bone matrix and contribute to bone remodeling. We have established a procedure to directly con- vert human fibroblasts into osteoblasts by transducing some defined factors and culturing in osteogenic medium. Osteo- blast-specific transcription factors, Runx2 and Osterix, in com- bination with Oct4 and L-Myc, drastically induced fibroblasts to produce calcified bone matrix and express osteoblast-specific markers. The directly converted osteoblasts (dOBs) showed similar gene expression profiles as normal osteoblasts and con- tributed to bone repair after transplantation into mice with bone defects. Furthermore, dOBs did not require continuous expres- sion of the exogenous genes to maintain their phenotype. These findings strongly suggest successful direct reprogramming of fibroblasts into osteoblasts, which may be applicable to bone regeneration therapy. Author contributions: K.Y., T. Kishida, and O.M. designed research; K.Y., T. Kishida, Y.S., and A.E. performed research; H.F., T. Kubo, T.Y., and N.K. contributed new reagents/ analytic tools; K.Y. and K.N. analyzed data; and K.Y. and O.M. wrote the paper. The authors declare no conflict of interest. This article is a PNAS Direct Submission. Data deposition: The microarray data reported in this paper have been deposited in the Gene Expression Omnibus (GEO) database, www.ncbi.nlm.nih.gov/geo (accession no. GSE52817). 1 To whom correspondence should be addressed. Email: [email protected]. This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10. 1073/pnas.1420713112/-/DCSupplemental. 6152–6157 | PNAS | May 12, 2015 | vol. 112 | no. 19 www.pnas.org/cgi/doi/10.1073/pnas.1420713112
Transcript of Direct conversion of human fibroblasts into functional ... · Direct conversion of human...

Direct conversion of human fibroblasts into functionalosteoblasts by defined factorsKenta Yamamotoa,b, Tsunao Kishidaa, Yoshiki Satoa,b, Keisuke Nishiokaa, Akika Ejimaa, Hiroyoshi Fujiwarac,Toshikazu Kuboc, Toshiro Yamamotob, Narisato Kanamurab, and Osam Mazdaa,1
Departments of aImmunology, bDental Medicine, and cOrthopedics, Kyoto Prefectural University of Medicine, Kamigyo-ku, Kyoto 602-8566, Japan
Edited by Darwin J. Prockop, Texas A&M Health Science Center, Temple, TX, and approved March 24, 2015 (received for review October 30, 2014)
Osteoblasts produce calcified bone matrix and contribute to boneformation and remodeling. In this study, we established a proce-dure to directly convert human fibroblasts into osteoblasts by trans-ducing some defined factors and culturing in osteogenic medium.Osteoblast-specific transcription factors, Runt-related transcriptionfactor 2 (Runx2), and Osterix, in combination with Octamer-bindingtranscription factor 3/4 (Oct4) and L-Myc (RXOL) transduction, con-verted ∼80% of the fibroblasts into osteocalcin-producing cells. Thedirectly converted osteoblasts (dOBs) induced by RXOL displayed asimilar gene expression profile as normal human osteoblasts andcontributed to bone repair after transplantation into immunodefi-cient mice at artificial bone defect lesions. The dOBs expressed en-dogenous Runx2 and Osterix, and did not require continuousexpression of the exogenous genes to maintain their phenotype.Another combination, Oct4 plus L-Myc (OL), also induced fibroblaststo produce bone matrix, but the OL-transduced cells did not expressOsterix and exhibited a more distant gene expression profile toosteoblasts compared with RXOL-transduced cells. These findingsstrongly suggest successful direct reprogramming of fibroblastsinto functional osteoblasts by RXOL, a technology that may providebone regeneration therapy against bone disorders.
osteoblasts | regenerative medicine | direct reprogramming
Osteoblasts play a central role in bone formation and remod-eling by producing type I collagen, osteopontin, osteocalcin,
and bone sialoprotein (BSP), and calcifying these bone matrixes(1). They are also involved in hematopoiesis, phosphate metabo-lism, and glucose metabolism (2). Osteoblasts are derived frommesenchymal progenitor cells that are common precursors sharedby chondrocytes, adipocytes, and myoblasts (3). The differentiationof osteoblasts is regulated by various transcription factors, in-cluding Runt-related transcription factor 2 [Runx2, also known ascore-binding factor subunit α-1 (Cbfα-1)] (4, 5), Osterix (6, 7),Distal-less homeobox 5 (Dlx5) (8), and activation transcriptionfactor 4 (ATF-4) (8). A functional decline in osteoblasts relative toosteoclasts results in imbalance between bone formation and re-sorption and may cause osteolytic pathological conditions, such asosteoporosis (9), alveolar bone resorption associated with perio-dontitis (10), and bone lysing associated with bone tumors, in-cluding multiple myeloma (11).It has been demonstrated that forced expression of combinations
of some transcription factors, such as Octamer-binding transcrip-tion factor 3/4 (Oct4), Sox2, Klf-4, and c-Myc (reprogrammingfactors), induces immortality and pluripotency in mammalian so-matic cells (12, 13). The generation of induced pluripotent stem(iPS) cells clearly indicates that genome-wide epigenetic pro-gramming can be drastically changed in somatic cells by a smallnumber of transcription factors that may have key regulatoryroles in cell fate decisions (14, 15).Recent studies have reported that direct conversion, or direct
reprogramming, of somatic cells into another differentiated line-age can be achieved without passing an intermediate pluripotentstage by introducing a set of transcription factors that are pivotalfor development of the destination cells. These conversions in-clude direct conversion of murine fibroblasts into cardiomyocytes
(16, 17), neurons (18–20), chondrocytes (21), and hepatocytes(22, 23), as well as of human fibroblasts into cardiomyocytes (24,25), neurons (19, 26), and hematopoietic cells (27). Although theefficiencies of conversion are generally low at present, with only0.005–30% of fibroblasts successfully converted into the desiredcells (16–20, 22–24, 26), this technology may become quite usefulfor regenerative therapy against a variety of human diseases.To date, however, there have been no reports on the induction
of human osteoblasts from another differentiated somatic celllineage by means of direct reprogramming procedures. If this canbe achieved, then a sufficient number of osteoblasts may beobtained from the somatic cells of a patient with osteolytic diseaseand then transplanted back into this patient. Moreover, such cell-based therapies might be applied to other clinical settings as well,such as repair of large bone defect caused by trauma or by surgicalresection of osteosarcoma or metastatic bone tumors.In the present study, we attempted to directly convert human
fibroblasts into osteoblasts by genetically introducing osteoblast-specific transcription factors and reprogramming factors. Wealso assessed the capability of the resulting directly convertedosteoblasts (dOBs) to contribute to bone formation in vivo.
ResultsHuman Fibroblasts Were Induced to Produce Calcified Bone Matrix byDefined Factors. We focused on two transcription factors, Runx-2(R) and Osterix (X), which are critical regulators of osteoblast
Significance
Osteoblasts produce bone matrix and contribute to boneremodeling. We have established a procedure to directly con-vert human fibroblasts into osteoblasts by transducing somedefined factors and culturing in osteogenic medium. Osteo-blast-specific transcription factors, Runx2 and Osterix, in com-bination with Oct4 and L-Myc, drastically induced fibroblasts toproduce calcified bone matrix and express osteoblast-specificmarkers. The directly converted osteoblasts (dOBs) showedsimilar gene expression profiles as normal osteoblasts and con-tributed to bone repair after transplantation into mice with bonedefects. Furthermore, dOBs did not require continuous expres-sion of the exogenous genes to maintain their phenotype. Thesefindings strongly suggest successful direct reprogramming offibroblasts into osteoblasts, which may be applicable to boneregeneration therapy.
Author contributions: K.Y., T. Kishida, and O.M. designed research; K.Y., T. Kishida, Y.S.,and A.E. performed research; H.F., T. Kubo, T.Y., and N.K. contributed new reagents/analytic tools; K.Y. and K.N. analyzed data; and K.Y. and O.M. wrote the paper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.
Data deposition: The microarray data reported in this paper have been deposited in theGene Expression Omnibus (GEO) database, www.ncbi.nlm.nih.gov/geo (accession no.GSE52817).1To whom correspondence should be addressed. Email: [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1420713112/-/DCSupplemental.
6152–6157 | PNAS | May 12, 2015 | vol. 112 | no. 19 www.pnas.org/cgi/doi/10.1073/pnas.1420713112
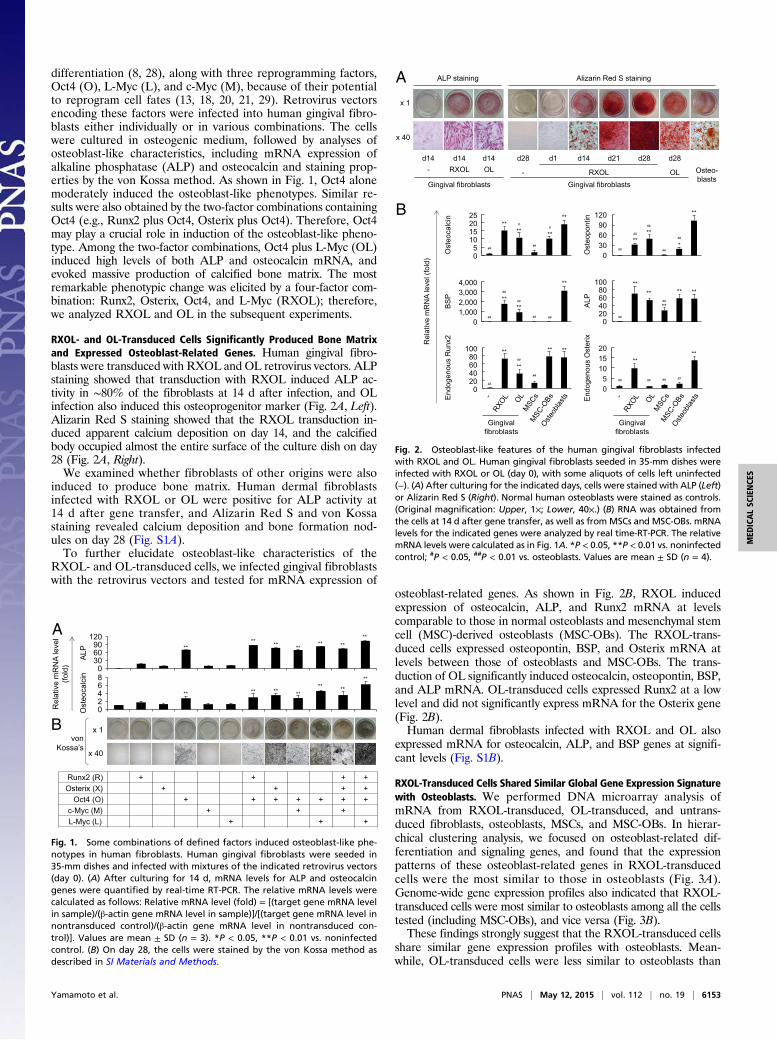
differentiation (8, 28), along with three reprogramming factors,Oct4 (O), L-Myc (L), and c-Myc (M), because of their potentialto reprogram cell fates (13, 18, 20, 21, 29). Retrovirus vectorsencoding these factors were infected into human gingival fibro-blasts either individually or in various combinations. The cellswere cultured in osteogenic medium, followed by analyses ofosteoblast-like characteristics, including mRNA expression ofalkaline phosphatase (ALP) and osteocalcin and staining prop-erties by the von Kossa method. As shown in Fig. 1, Oct4 alonemoderately induced the osteoblast-like phenotypes. Similar re-sults were also obtained by the two-factor combinations containingOct4 (e.g., Runx2 plus Oct4, Osterix plus Oct4). Therefore, Oct4may play a crucial role in induction of the osteoblast-like pheno-type. Among the two-factor combinations, Oct4 plus L-Myc (OL)induced high levels of both ALP and osteocalcin mRNA, andevoked massive production of calcified bone matrix. The mostremarkable phenotypic change was elicited by a four-factor com-bination: Runx2, Osterix, Oct4, and L-Myc (RXOL); therefore,we analyzed RXOL and OL in the subsequent experiments.
RXOL- and OL-Transduced Cells Significantly Produced Bone Matrixand Expressed Osteoblast-Related Genes. Human gingival fibro-blasts were transduced with RXOL and OL retrovirus vectors. ALPstaining showed that transduction with RXOL induced ALP ac-tivity in ∼80% of the fibroblasts at 14 d after infection, and OLinfection also induced this osteoprogenitor marker (Fig. 2A, Left).Alizarin Red S staining showed that the RXOL transduction in-duced apparent calcium deposition on day 14, and the calcifiedbody occupied almost the entire surface of the culture dish on day28 (Fig. 2A, Right).We examined whether fibroblasts of other origins were also
induced to produce bone matrix. Human dermal fibroblastsinfected with RXOL or OL were positive for ALP activity at14 d after gene transfer, and Alizarin Red S and von Kossastaining revealed calcium deposition and bone formation nod-ules on day 28 (Fig. S1A).To further elucidate osteoblast-like characteristics of the
RXOL- and OL-transduced cells, we infected gingival fibroblastswith the retrovirus vectors and tested for mRNA expression of
osteoblast-related genes. As shown in Fig. 2B, RXOL inducedexpression of osteocalcin, ALP, and Runx2 mRNA at levelscomparable to those in normal osteoblasts and mesenchymal stemcell (MSC)-derived osteoblasts (MSC-OBs). The RXOL-trans-duced cells expressed osteopontin, BSP, and Osterix mRNA atlevels between those of osteoblasts and MSC-OBs. The trans-duction of OL significantly induced osteocalcin, osteopontin, BSP,and ALP mRNA. OL-transduced cells expressed Runx2 at a lowlevel and did not significantly express mRNA for the Osterix gene(Fig. 2B).Human dermal fibroblasts infected with RXOL and OL also
expressed mRNA for osteocalcin, ALP, and BSP genes at signifi-cant levels (Fig. S1B).
RXOL-Transduced Cells Shared Similar Global Gene Expression Signaturewith Osteoblasts. We performed DNA microarray analysis ofmRNA from RXOL-transduced, OL-transduced, and untrans-duced fibroblasts, osteoblasts, MSCs, and MSC-OBs. In hierar-chical clustering analysis, we focused on osteoblast-related dif-ferentiation and signaling genes, and found that the expressionpatterns of these osteoblast-related genes in RXOL-transducedcells were the most similar to those in osteoblasts (Fig. 3A).Genome-wide gene expression profiles also indicated that RXOL-transduced cells were most similar to osteoblasts among all the cellstested (including MSC-OBs), and vice versa (Fig. 3B).These findings strongly suggest that the RXOL-transduced cells
share similar gene expression profiles with osteoblasts. Mean-while, OL-transduced cells were less similar to osteoblasts than
Runx2 (R) + + + + Osterix (X) + + + +
Oct4 (O) + + + + + + + c-Myc (M) + + +
L-Myc (L) + + +
x 1
x 40
von Kossa’s
** ** ** ** ** **
**
Ost
eoca
lcin
A
LP ** ** ** ** **
** **
Rel
ativ
e m
RN
A le
vel
(fold
)
A
B
120 90 60 30 0
6 4 2 0
8
Fig. 1. Some combinations of defined factors induced osteoblast-like phe-notypes in human fibroblasts. Human gingival fibroblasts were seeded in35-mm dishes and infected with mixtures of the indicated retrovirus vectors(day 0). (A) After culturing for 14 d, mRNA levels for ALP and osteocalcingenes were quantified by real-time RT-PCR. The relative mRNA levels werecalculated as follows: Relative mRNA level (fold) = [(target gene mRNA levelin sample)/(β-actin gene mRNA level in sample)]/[(target gene mRNA level innontransduced control)/(β-actin gene mRNA level in nontransduced con-trol)]. Values are mean ± SD (n = 3). *P < 0.05, **P < 0.01 vs. noninfectedcontrol. (B) On day 28, the cells were stained by the von Kossa method asdescribed in SI Materials and Methods.
End
ogen
ous
Run
x2
End
ogen
ous
Ost
erix
A
LP
BS
P
Ost
eopo
ntin
Ost
eoca
lcin
** ** **
** #
#
## ##
** **
**
##
##
##
##
##
** **
**
*
##
##
## ## ##
**
**
** ** **
##
##
**
**
** **
##
##
##
** **
## ## ## ##
Rel
ativ
e m
RN
A le
vel (
fold
)
A
RXOL Osteo- blasts
OL Gingival fibroblasts
-
d1 d14 d21 d28 d28
x 1
x 40
Alizarin Red S staining
d14 d14 d14
Gingival fibroblasts
ALP staining
d28 - RXOL OL
B 25 20 15 10 5 0
4,000 3,000 2,000 1,000
0
100 80 60 40 20 0
120 90 60 30 0
100 80 60 40
0
20 15 10 5 0
20
Fig. 2. Osteoblast-like features of the human gingival fibroblasts infectedwith RXOL and OL. Human gingival fibroblasts seeded in 35-mm dishes wereinfected with RXOL or OL (day 0), with some aliquots of cells left uninfected(−). (A) After culturing for the indicated days, cells were stained with ALP (Left)or Alizarin Red S (Right). Normal human osteoblasts were stained as controls.(Original magnification: Upper, 1×; Lower, 40×.) (B) RNA was obtained fromthe cells at 14 d after gene transfer, as well as fromMSCs and MSC-OBs. mRNAlevels for the indicated genes were analyzed by real time-RT-PCR. The relativemRNA levels were calculated as in Fig. 1A. *P < 0.05, **P < 0.01 vs. noninfectedcontrol; #P < 0.05, ##P < 0.01 vs. osteoblasts. Values are mean ± SD (n = 4).
Yamamoto et al. PNAS | May 12, 2015 | vol. 112 | no. 19 | 6153
MED
ICALSC
IENCE
S
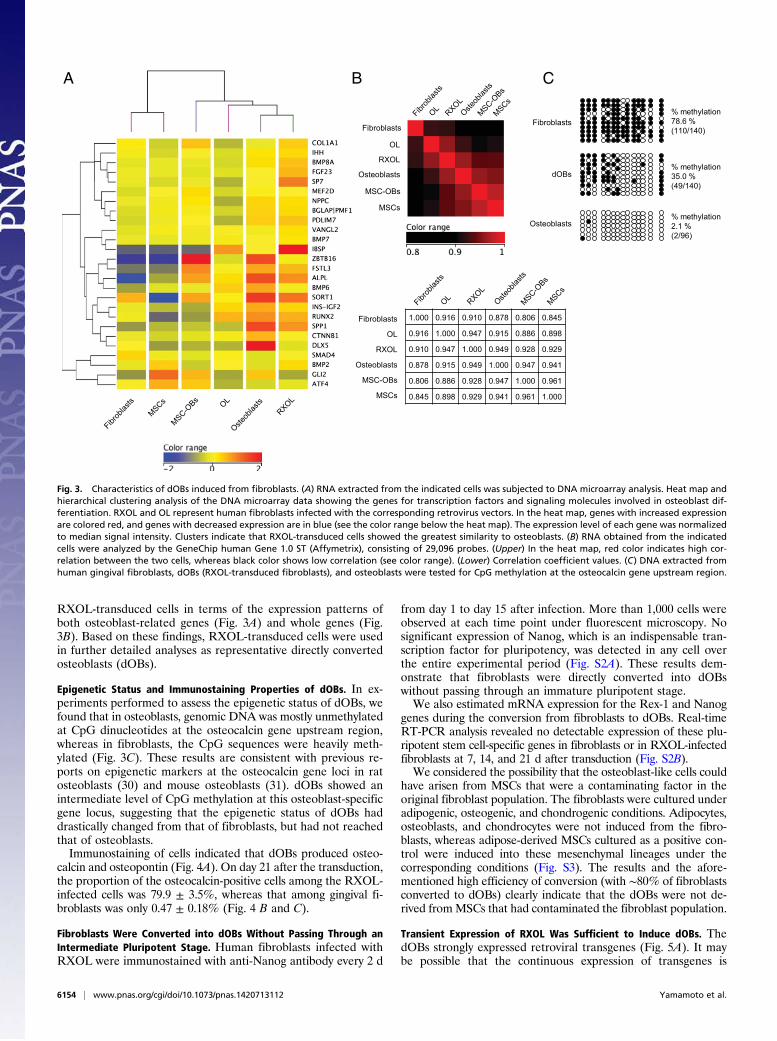
RXOL-transduced cells in terms of the expression patterns ofboth osteoblast-related genes (Fig. 3A) and whole genes (Fig.3B). Based on these findings, RXOL-transduced cells were usedin further detailed analyses as representative directly convertedosteoblasts (dOBs).
Epigenetic Status and Immunostaining Properties of dOBs. In ex-periments performed to assess the epigenetic status of dOBs, wefound that in osteoblasts, genomic DNA was mostly unmethylatedat CpG dinucleotides at the osteocalcin gene upstream region,whereas in fibroblasts, the CpG sequences were heavily meth-ylated (Fig. 3C). These results are consistent with previous re-ports on epigenetic markers at the osteocalcin gene loci in ratosteoblasts (30) and mouse osteoblasts (31). dOBs showed anintermediate level of CpG methylation at this osteoblast-specificgene locus, suggesting that the epigenetic status of dOBs haddrastically changed from that of fibroblasts, but had not reachedthat of osteoblasts.Immunostaining of cells indicated that dOBs produced osteo-
calcin and osteopontin (Fig. 4A). On day 21 after the transduction,the proportion of the osteocalcin-positive cells among the RXOL-infected cells was 79.9 ± 3.5%, whereas that among gingival fi-broblasts was only 0.47 ± 0.18% (Fig. 4 B and C).
Fibroblasts Were Converted into dOBs Without Passing Through anIntermediate Pluripotent Stage. Human fibroblasts infected withRXOL were immunostained with anti-Nanog antibody every 2 d
from day 1 to day 15 after infection. More than 1,000 cells wereobserved at each time point under fluorescent microscopy. Nosignificant expression of Nanog, which is an indispensable tran-scription factor for pluripotency, was detected in any cell overthe entire experimental period (Fig. S2A). These results dem-onstrate that fibroblasts were directly converted into dOBswithout passing through an immature pluripotent stage.We also estimated mRNA expression for the Rex-1 and Nanog
genes during the conversion from fibroblasts to dOBs. Real-timeRT-PCR analysis revealed no detectable expression of these plu-ripotent stem cell-specific genes in fibroblasts or in RXOL-infectedfibroblasts at 7, 14, and 21 d after transduction (Fig. S2B).We considered the possibility that the osteoblast-like cells could
have arisen from MSCs that were a contaminating factor in theoriginal fibroblast population. The fibroblasts were cultured underadipogenic, osteogenic, and chondrogenic conditions. Adipocytes,osteoblasts, and chondrocytes were not induced from the fibro-blasts, whereas adipose-derived MSCs cultured as a positive con-trol were induced into these mesenchymal lineages under thecorresponding conditions (Fig. S3). The results and the afore-mentioned high efficiency of conversion (with ∼80% of fibroblastsconverted to dOBs) clearly indicate that the dOBs were not de-rived fromMSCs that had contaminated the fibroblast population.
Transient Expression of RXOL Was Sufficient to Induce dOBs. ThedOBs strongly expressed retroviral transgenes (Fig. 5A). It maybe possible that the continuous expression of transgenes is
Fibroblasts
dOBs
Osteoblasts
% methylation 78.6 % (110/140)
% methylation 35.0 % (49/140)
% methylation 2.1 % (2/96)
C B A
1.000 0.916 0.910 0.878 0.806 0.845
0.916 1.000 0.947 0.915 0.886 0.898
0.910 0.947 1.000 0.949 0.928 0.929
0.878 0.915 0.949 1.000 0.947 0.941
0.806 0.886 0.928 0.947 1.000 0.961
0.845 0.898 0.929 0.941 0.961 1.000
Osteoblasts
Fibroblasts
OL
MSC-OBs
MSCs
RXOL
Fibroblasts
OL
Osteoblasts
MSC-OBs
MSCs
RXOL
Fig. 3. Characteristics of dOBs induced from fibroblasts. (A) RNA extracted from the indicated cells was subjected to DNA microarray analysis. Heat map andhierarchical clustering analysis of the DNA microarray data showing the genes for transcription factors and signaling molecules involved in osteoblast dif-ferentiation. RXOL and OL represent human fibroblasts infected with the corresponding retrovirus vectors. In the heat map, genes with increased expressionare colored red, and genes with decreased expression are in blue (see the color range below the heat map). The expression level of each gene was normalizedto median signal intensity. Clusters indicate that RXOL-transduced cells showed the greatest similarity to osteoblasts. (B) RNA obtained from the indicatedcells were analyzed by the GeneChip human Gene 1.0 ST (Affymetrix), consisting of 29,096 probes. (Upper) In the heat map, red color indicates high cor-relation between the two cells, whereas black color shows low correlation (see color range). (Lower) Correlation coefficient values. (C) DNA extracted fromhuman gingival fibroblasts, dOBs (RXOL-transduced fibroblasts), and osteoblasts were tested for CpG methylation at the osteocalcin gene upstream region.
6154 | www.pnas.org/cgi/doi/10.1073/pnas.1420713112 Yamamoto et al.
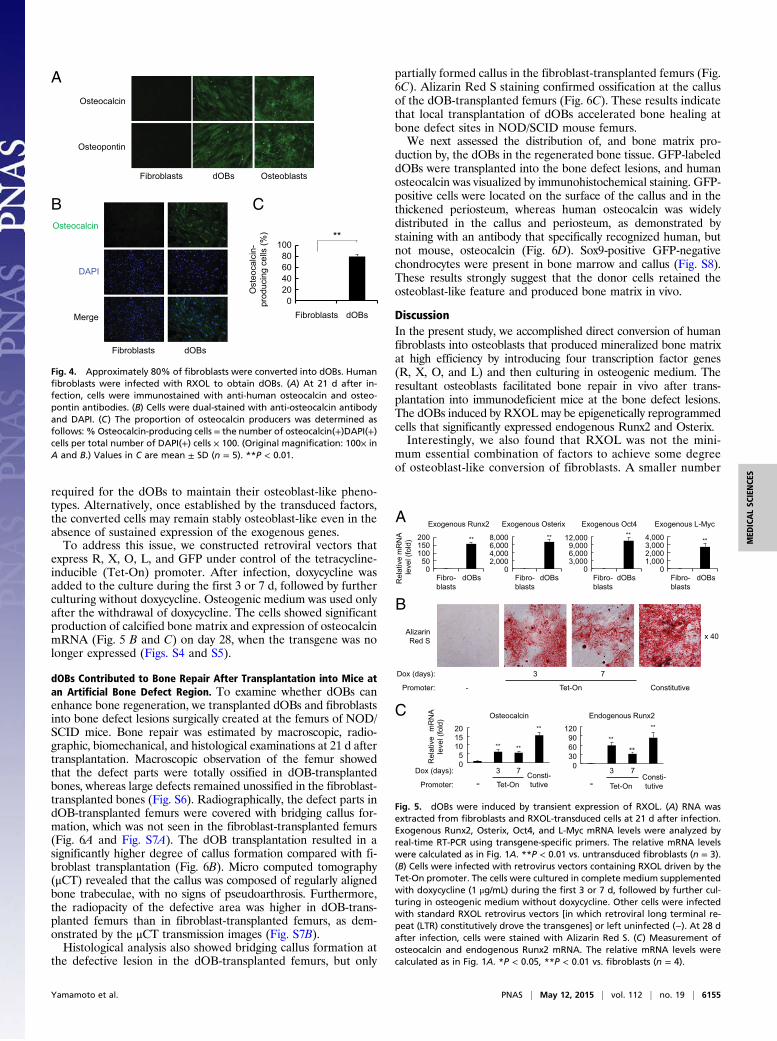
required for the dOBs to maintain their osteoblast-like pheno-types. Alternatively, once established by the transduced factors,the converted cells may remain stably osteoblast-like even in theabsence of sustained expression of the exogenous genes.To address this issue, we constructed retroviral vectors that
express R, X, O, L, and GFP under control of the tetracycline-inducible (Tet-On) promoter. After infection, doxycycline wasadded to the culture during the first 3 or 7 d, followed by furtherculturing without doxycycline. Osteogenic medium was used onlyafter the withdrawal of doxycycline. The cells showed significantproduction of calcified bone matrix and expression of osteocalcinmRNA (Fig. 5 B and C) on day 28, when the transgene was nolonger expressed (Figs. S4 and S5).
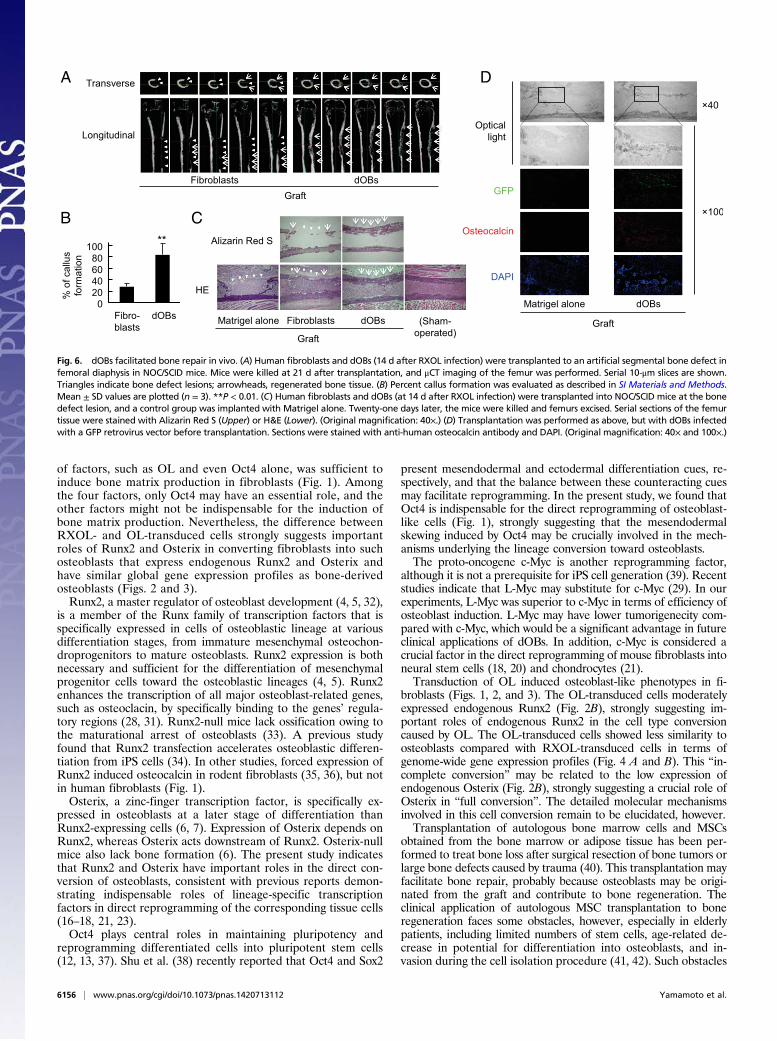
dOBs Contributed to Bone Repair After Transplantation into Mice atan Artificial Bone Defect Region. To examine whether dOBs canenhance bone regeneration, we transplanted dOBs and fibroblastsinto bone defect lesions surgically created at the femurs of NOD/SCID mice. Bone repair was estimated by macroscopic, radio-graphic, biomechanical, and histological examinations at 21 d aftertransplantation. Macroscopic observation of the femur showedthat the defect parts were totally ossified in dOB-transplantedbones, whereas large defects remained unossified in the fibroblast-transplanted bones (Fig. S6). Radiographically, the defect parts indOB-transplanted femurs were covered with bridging callus for-mation, which was not seen in the fibroblast-transplanted femurs(Fig. 6A and Fig. S7A). The dOB transplantation resulted in asignificantly higher degree of callus formation compared with fi-broblast transplantation (Fig. 6B). Micro computed tomography(μCT) revealed that the callus was composed of regularly alignedbone trabeculae, with no signs of pseudoarthrosis. Furthermore,the radiopacity of the defective area was higher in dOB-trans-planted femurs than in fibroblast-transplanted femurs, as dem-onstrated by the μCT transmission images (Fig. S7B).Histological analysis also showed bridging callus formation at
the defective lesion in the dOB-transplanted femurs, but only
partially formed callus in the fibroblast-transplanted femurs (Fig.6C). Alizarin Red S staining confirmed ossification at the callusof the dOB-transplanted femurs (Fig. 6C). These results indicatethat local transplantation of dOBs accelerated bone healing atbone defect sites in NOD/SCID mouse femurs.We next assessed the distribution of, and bone matrix pro-
duction by, the dOBs in the regenerated bone tissue. GFP-labeleddOBs were transplanted into the bone defect lesions, and humanosteocalcin was visualized by immunohistochemical staining. GFP-positive cells were located on the surface of the callus and in thethickened periosteum, whereas human osteocalcin was widelydistributed in the callus and periosteum, as demonstrated bystaining with an antibody that specifically recognized human, butnot mouse, osteocalcin (Fig. 6D). Sox9-positive GFP-negativechondrocytes were present in bone marrow and callus (Fig. S8).These results strongly suggest that the donor cells retained theosteoblast-like feature and produced bone matrix in vivo.
DiscussionIn the present study, we accomplished direct conversion of humanfibroblasts into osteoblasts that produced mineralized bone matrixat high efficiency by introducing four transcription factor genes(R, X, O, and L) and then culturing in osteogenic medium. Theresultant osteoblasts facilitated bone repair in vivo after trans-plantation into immunodeficient mice at the bone defect lesions.The dOBs induced by RXOLmay be epigenetically reprogrammedcells that significantly expressed endogenous Runx2 and Osterix.Interestingly, we also found that RXOL was not the mini-
mum essential combination of factors to achieve some degreeof osteoblast-like conversion of fibroblasts. A smaller number
Fibroblasts dOBs
Osteocalcin
DAPI
Merge
B
A
Osteoblasts dOBs
Osteocalcin
Osteopontin
Fibroblasts
C
100
0
40
Fibroblasts dOBs
Ost
eoca
lcin
-pr
oduc
ing
cells
(%)
60
20
80
**
Fig. 4. Approximately 80% of fibroblasts were converted into dOBs. Humanfibroblasts were infected with RXOL to obtain dOBs. (A) At 21 d after in-fection, cells were immunostained with anti-human osteocalcin and osteo-pontin antibodies. (B) Cells were dual-stained with anti-osteocalcin antibodyand DAPI. (C) The proportion of osteocalcin producers was determined asfollows: % Osteocalcin-producing cells = the number of osteocalcin(+)DAPI(+)cells per total number of DAPI(+) cells × 100. (Original magnification: 100× inA and B.) Values in C are mean ± SD (n = 5). **P < 0.01.
A
B
Rel
ativ
e m
RN
A
leve
l (fo
ld)
Exogenous Runx2 Exogenous Osterix
** **
Exogenous Oct4 Exogenous L-Myc **
**
Fibro- blasts
dOBs dOBs dOBs dOBs Fibro- blasts
Fibro- blasts
Fibro- blasts
Alizarin Red S x 40
-
7 3
Constitutive Tet-On
Dox (days):
Promoter:
C
4,000 3,000 2,000 1,000
0
200 150 100
50 0
8,000 6,000 4,000 2,000
0
12,000 9,000 6,000 3,000
0
- 7 3
Consti- tutive Tet-On
Endogenous Runx2 **
**
**
120 90 60 30 0
- 7 3 Consti-
tutive Tet-On
Dox (days):
Promoter:
Osteocalcin
** **
**
Rel
ativ
em
RN
A le
vel (
fold
)
20 15 10
5 0
Fig. 5. dOBs were induced by transient expression of RXOL. (A) RNA wasextracted from fibroblasts and RXOL-transduced cells at 21 d after infection.Exogenous Runx2, Osterix, Oct4, and L-Myc mRNA levels were analyzed byreal-time RT-PCR using transgene-specific primers. The relative mRNA levelswere calculated as in Fig. 1A. **P < 0.01 vs. untransduced fibroblasts (n = 3).(B) Cells were infected with retrovirus vectors containing RXOL driven by theTet-On promoter. The cells were cultured in complete medium supplementedwith doxycycline (1 μg/mL) during the first 3 or 7 d, followed by further cul-turing in osteogenic medium without doxycycline. Other cells were infectedwith standard RXOL retrovirus vectors [in which retroviral long terminal re-peat (LTR) constitutively drove the transgenes] or left uninfected (−). At 28 dafter infection, cells were stained with Alizarin Red S. (C) Measurement ofosteocalcin and endogenous Runx2 mRNA. The relative mRNA levels werecalculated as in Fig. 1A. *P < 0.05, **P < 0.01 vs. fibroblasts (n = 4).
Yamamoto et al. PNAS | May 12, 2015 | vol. 112 | no. 19 | 6155
MED
ICALSC
IENCE
S
of factors, such as OL and even Oct4 alone, was sufficient toinduce bone matrix production in fibroblasts (Fig. 1). Amongthe four factors, only Oct4 may have an essential role, and theother factors might not be indispensable for the induction ofbone matrix production. Nevertheless, the difference betweenRXOL- and OL-transduced cells strongly suggests importantroles of Runx2 and Osterix in converting fibroblasts into suchosteoblasts that express endogenous Runx2 and Osterix andhave similar global gene expression profiles as bone-derivedosteoblasts (Figs. 2 and 3).Runx2, a master regulator of osteoblast development (4, 5, 32),
is a member of the Runx family of transcription factors that isspecifically expressed in cells of osteoblastic lineage at variousdifferentiation stages, from immature mesenchymal osteochon-droprogenitors to mature osteoblasts. Runx2 expression is bothnecessary and sufficient for the differentiation of mesenchymalprogenitor cells toward the osteoblastic lineages (4, 5). Runx2enhances the transcription of all major osteoblast-related genes,such as osteoclacin, by specifically binding to the genes’ regula-tory regions (28, 31). Runx2-null mice lack ossification owing tothe maturational arrest of osteoblasts (33). A previous studyfound that Runx2 transfection accelerates osteoblastic differen-tiation from iPS cells (34). In other studies, forced expression ofRunx2 induced osteocalcin in rodent fibroblasts (35, 36), but notin human fibroblasts (Fig. 1).Osterix, a zinc-finger transcription factor, is specifically ex-
pressed in osteoblasts at a later stage of differentiation thanRunx2-expressing cells (6, 7). Expression of Osterix depends onRunx2, whereas Osterix acts downstream of Runx2. Osterix-nullmice also lack bone formation (6). The present study indicatesthat Runx2 and Osterix have important roles in the direct con-version of osteoblasts, consistent with previous reports demon-strating indispensable roles of lineage-specific transcriptionfactors in direct reprogramming of the corresponding tissue cells(16–18, 21, 23).Oct4 plays central roles in maintaining pluripotency and
reprogramming differentiated cells into pluripotent stem cells(12, 13, 37). Shu et al. (38) recently reported that Oct4 and Sox2
present mesendodermal and ectodermal differentiation cues, re-spectively, and that the balance between these counteracting cuesmay facilitate reprogramming. In the present study, we found thatOct4 is indispensable for the direct reprogramming of osteoblast-like cells (Fig. 1), strongly suggesting that the mesendodermalskewing induced by Oct4 may be crucially involved in the mech-anisms underlying the lineage conversion toward osteoblasts.The proto-oncogene c-Myc is another reprogramming factor,
although it is not a prerequisite for iPS cell generation (39). Recentstudies indicate that L-Myc may substitute for c-Myc (29). In ourexperiments, L-Myc was superior to c-Myc in terms of efficiency ofosteoblast induction. L-Myc may have lower tumorigenecity com-pared with c-Myc, which would be a significant advantage in futureclinical applications of dOBs. In addition, c-Myc is considered acrucial factor in the direct reprogramming of mouse fibroblasts intoneural stem cells (18, 20) and chondrocytes (21).Transduction of OL induced osteoblast-like phenotypes in fi-
broblasts (Figs. 1, 2, and 3). The OL-transduced cells moderatelyexpressed endogenous Runx2 (Fig. 2B), strongly suggesting im-portant roles of endogenous Runx2 in the cell type conversioncaused by OL. The OL-transduced cells showed less similarity toosteoblasts compared with RXOL-transduced cells in terms ofgenome-wide gene expression profiles (Fig. 4 A and B). This “in-complete conversion” may be related to the low expression ofendogenous Osterix (Fig. 2B), strongly suggesting a crucial role ofOsterix in “full conversion”. The detailed molecular mechanismsinvolved in this cell conversion remain to be elucidated, however.Transplantation of autologous bone marrow cells and MSCs
obtained from the bone marrow or adipose tissue has been per-formed to treat bone loss after surgical resection of bone tumors orlarge bone defects caused by trauma (40). This transplantation mayfacilitate bone repair, probably because osteoblasts may be origi-nated from the graft and contribute to bone regeneration. Theclinical application of autologous MSC transplantation to boneregeneration faces some obstacles, however, especially in elderlypatients, including limited numbers of stem cells, age-related de-crease in potential for differentiation into osteoblasts, and in-vasion during the cell isolation procedure (41, 42). Such obstacles
A
Graft
Longitudinal
sBOdstsalborbiF
Transverse
HE
C
Fibroblasts dOBs (Sham-operated) Graft
Alizarin Red S
Matrigel alone
B
D
Matrigel alone dOBs
Graft
% o
f cal
lus
form
atio
n
Fibro-blasts
dOBs
** 100 80 60 40
0 20
Fig. 6. dOBs facilitated bone repair in vivo. (A) Human fibroblasts and dOBs (14 d after RXOL infection) were transplanted to an artificial segmental bone defect infemoral diaphysis in NOC/SCID mice. Mice were killed at 21 d after transplantation, and μCT imaging of the femur was performed. Serial 10-μm slices are shown.Triangles indicate bone defect lesions; arrowheads, regenerated bone tissue. (B) Percent callus formation was evaluated as described in SI Materials and Methods.Mean ± SD values are plotted (n = 3). **P < 0.01. (C) Human fibroblasts and dOBs (at 14 d after RXOL infection) were transplanted into NOC/SCID mice at the bonedefect lesion, and a control group was implanted with Matrigel alone. Twenty-one days later, the mice were killed and femurs excised. Serial sections of the femurtissue were stained with Alizarin Red S (Upper) or H&E (Lower). (Original magnification: 40×.) (D) Transplantation was performed as above, but with dOBs infectedwith a GFP retrovirus vector before transplantation. Sections were stained with anti-human osteocalcin antibody and DAPI. (Original magnification: 40× and 100×.)
6156 | www.pnas.org/cgi/doi/10.1073/pnas.1420713112 Yamamoto et al.

may be avoided by applying the direct reprogramming technol-ogy, in which a large number of functional osteoblasts can beobtained from fibroblasts isolated from patients or allogenic do-nors by a less-invasive procedure (43, 44). Fibroblasts have a highproliferative capacity, which does not decline with increasingdonor age (43, 45). Thus, direct reprogramming may provideanother option for such patients in whom MSC transplantationmight not be an appropriate therapy.Osteoblasts induced from a patient’s own somatic cells or al-
logenic cells also may be applicable to cell-based therapy againstbone resorptive conditions associated with osteoporosis, bonedisruption caused by rheumatoid arthritis, and alveolar bone re-sorption due to periodontitis. This therapy may help increase thepatient’s quality of life and capacity to perform activities ofdaily living.
Materials and MethodsFor direct reprogramming, human fibroblasts were retrovirally transducedwithtranscriptional factors and cultured in DMEM supplemented with 50 μg/mLascorbic acid, 10 mM β-glycerol phosphate, 100 nM dexamethasone, and 10%FBS (osteogenic medium). NHost cells (normal human osteoblasts derived fromfemur of a 39-y-old male) were purchased from Lonza. Detailed informationon cells, retroviral vectors, infection, induction into osteoblasts, cell staining,real-time RT-PCR, immunostaining, DNA microarray analysis, bisulfite se-quencing, surgical procedure, cell transplantation, radiographic and histolog-ical assessment, and data analysis is provided in SI Materials and Methods. Thesequences of real-time RT-PCR primers are provided in Table S1.
ACKNOWLEDGMENTS. We thank Dr. Wilfred Germeraad for a critical readingof the manuscript and Hiroaki Ichioka and Ken-ichi Honjo for technical support.This work was supported by grants from the Japan Science and TechnologyAgency (PRESTO and A-Step); the Japanese Ministry of Education, Culture,Sports, Science and Technology; and the Suzuken Memorial Foundation.
1. Neve A, Corrado A, Cantatore FP (2011) Osteoblast physiology in normal and path-ological conditions. Cell Tissue Res 343(2):289–302.
2. Bethel M, Srour EF, Kacena MA (2011) Hematopoietic cell regulation of osteoblastproliferation and differentiation. Curr Osteoporos Rep 9(2):96–102.
3. Pittenger MF, et al. (1999) Multilineage potential of adult human mesenchymal stemcells. Science 284(5411):143–147.
4. Dalle Carbonare L, Innamorati G, Valenti MT (2012) Transcription factor Runx2 and itsapplication to bone tissue engineering. Stem Cell Rev 8(3):891–897.
5. Komori T (2010) Regulation of bone development and extracellular matrix proteingenes by RUNX2. Cell Tissue Res 339(1):189–195.
6. Nakashima K, et al. (2002) The novel zinc finger-containing transcription factor os-terix is required for osteoblast differentiation and bone formation. Cell 108(1):17–29.
7. Sinha KM, Zhou X (2013) Genetic and molecular control of osterix in skeletal for-mation. J Cell Biochem 114(5):975–984.
8. Papachroni KK, Karatzas DN, Papavassiliou KA, Basdra EK, Papavassiliou AG (2009)Mechanotransduction in osteoblast regulation and bone disease. Trends Mol Med15(5):208–216.
9. Rachner TD, Khosla S, Hofbauer LC (2011) Osteoporosis: Now and the future. Lancet377(9773):1276–1287.
10. Bartold PM, Cantley MD, Haynes DR (2010) Mechanisms and control of pathologicbone loss in periodontitis. Periodontol 2000 53:55–69.
11. Buckle CH, et al. (2012) Soluble rank ligand produced by myeloma cells causes gen-eralised bone loss in multiple myeloma. PLoS ONE 7(8):e41127.
12. Takahashi K, Yamanaka S (2006) Induction of pluripotent stem cells from mouseembryonic and adult fibroblast cultures by defined factors. Cell 126(4):663–676.
13. Takahashi K, et al. (2007) Induction of pluripotent stem cells from adult human fi-broblasts by defined factors. Cell 131(5):861–872.
14. Meissner A (2010) Epigenetic modifications in pluripotent and differentiated cells.Nat Biotechnol 28(10):1079–1088.
15. Papp B, Plath K (2011) Reprogramming to pluripotency: Stepwise resetting of theepigenetic landscape. Cell Res 21(3):486–501.
16. Ieda M, et al. (2010) Direct reprogramming of fibroblasts into functional car-diomyocytes by defined factors. Cell 142(3):375–386.
17. Inagawa K, Ieda M (2013) Direct reprogramming of mouse fibroblasts into cardiacmyocytes. J Cardiovasc Transl Res 6(1):37–45.
18. Han DW, et al. (2012) Direct reprogramming of fibroblasts into neural stem cells bydefined factors. Cell Stem Cell 10(4):465–472.
19. Kim J, Ambasudhan R, Ding S (2012) Direct lineage reprogramming to neural cells.Curr Opin Neurobiol 22(5):778–784.
20. Thier M, et al. (2012) Direct conversion of fibroblasts into stably expandable neuralstem cells. Cell Stem Cell 10(4):473–479.
21. Hiramatsu K, et al. (2011) Generation of hyaline cartilaginous tissue frommouse adultdermal fibroblast culture by defined factors. J Clin Invest 121(2):640–657.
22. Huang P, et al. (2011) Induction of functional hepatocyte-like cells from mouse fi-broblasts by defined factors. Nature 475(7356):386–389.
23. Sekiya S, Suzuki A (2011) Direct conversion of mouse fibroblasts to hepatocyte-likecells by defined factors. Nature 475(7356):390–393.
24. Nam YJ, et al. (2013) Reprogramming of human fibroblasts toward a cardiac fate.Proc Natl Acad Sci USA 110(14):5588–5593.
25. Wada R, et al. (2013) Induction of human cardiomyocyte-like cells from fibroblasts bydefined factors. Proc Natl Acad Sci USA 110(31):12667–12672.
26. Pang ZP, et al. (2011) Induction of human neuronal cells by defined transcriptionfactors. Nature 476(7359):220–223.
27. Szabo E, et al. (2010) Direct conversion of human fibroblasts to multilineage blood
progenitors. Nature 468(7323):521–526.28. Ducy P, Zhang R, Geoffroy V, Ridall AL, Karsenty G (1997) Osf2/Cbfa1: A transcrip-
tional activator of osteoblast differentiation. Cell 89(5):747–754.29. Nakagawa M, Takizawa N, Narita M, Ichisaka T, Yamanaka S (2010) Promotion of
direct reprogramming by transformation-deficient Myc. Proc Natl Acad Sci USA
107(32):14152–14157.30. Villagra A, et al. (2002) Reduced CpG methylation is associated with transcriptional
activation of the bone-specific rat osteocalcin gene in osteoblasts. J Cell Biochem85(1):112–122.
31. Xiao G, et al. (2005) Cooperative interactions between activating transcription factor
4 and Runx2/Cbfa1 stimulate osteoblast-specific osteocalcin gene expression. J BiolChem 280(35):30689–30696.
32. Komori T (2006) Regulation of osteoblast differentiation by transcription factors.J Cell Biochem 99(5):1233–1239.
33. Komori T, et al. (1997) Targeted disruption of Cbfa1 results in a complete lack of bone
formation owing to maturational arrest of osteoblasts. Cell 89(5):755–764.34. Tashiro K, et al. (2009) Efficient adipocyte and osteoblast differentiation from mouse
induced pluripotent stem cells by adenoviral transduction. Stem Cells 27(8):1802–1811.
35. Phillips JE, Guldberg RE, García AJ (2007) Dermal fibroblasts genetically modified to
express Runx2/Cbfa1 as a mineralizing cell source for bone tissue engineering. TissueEng 13(8):2029–2040.
36. Takahashi T (2011) Overexpression of Runx2 and MKP-1 stimulates trans-differentiation of 3T3-L1 preadipocytes into bone-forming osteoblasts in vitro. CalcifTissue Int 88(4):336–347.
37. Yamanaka S (2007) Strategies and new developments in the generation of patient-specific pluripotent stem cells. Cell Stem Cell 1(1):39–49.
38. Shu J, et al. (2013) Induction of pluripotency in mouse somatic cells with lineagespecifiers. Cell 153(5):963–975.
39. Nakagawa M, et al. (2008) Generation of induced pluripotent stem cells without Mycfrom mouse and human fibroblasts. Nat Biotechnol 26(1):101–106.
40. Liu Y, Wu J, Zhu Y, Han J (2014) Therapeutic application of mesenchymal stem cells in
bone and joint diseases. Clin Exp Med 14(1):13–24.41. Kim M, et al. (2012) Age-related alterations in mesenchymal stem cells related to shift
in differentiation from osteogenic to adipogenic potential: Implication to age-asso-ciated bone diseases and defects. Mech Ageing Dev 133(5):215–225.
42. Zhou S, et al. (2008) Age-related intrinsic changes in human bone marrow-derived
mesenchymal stem cells and their differentiation to osteoblasts. Aging Cell 7(3):335–343.
43. Huschtscha LI, et al. (2012) Enhanced isolation of fibroblasts from human skin ex-plants. Biotechniques 53(4):239–244.
44. Shay JW, Wright WE (2000) Hayflick, his limit, and cellular ageing. Nat Rev Mol Cell
Biol 1(1):72–76.45. Cristofalo VJ, Allen RG, Pignolo RJ, Martin BG, Beck JC (1998) Relationship between
donor age and the replicative lifespan of human cells in culture: A reevaluation. ProcNatl Acad Sci USA 95(18):10614–10619.
Yamamoto et al. PNAS | May 12, 2015 | vol. 112 | no. 19 | 6157
MED
ICALSC
IENCE
S














![Neuronal Transcription Factors Induce Conversion of Human ... · induces mouse fibroblasts to become functional neurons [12]. Other transcription factors, such as Ngn2 or Dlx1, are](https://static.fdocuments.in/doc/165x107/60418e3cb320ed3248628b47/neuronal-transcription-factors-induce-conversion-of-human-induces-mouse-fibroblasts.jpg)


