
DIHYDROOROTATE DEHYDROGENASE OF Toxoplasma gondii
83
i DIHYDROOROTATE DEHYDROGENASE OF Toxoplasma gondii: KINETIC CHARACTERIZATION AND INTRACELLULAR LOCALIZATION by MIRYAM ANDREA HORTUA TRIANA DISSERTATION Submitted to the Graduate School of Universidad de los Andes, Bogota, Colombia in partial fulfillment of the requirements for the degree of DOCTOR OF PHILOSOPHY 2010 MAJOR: Biological Sciences
Transcript of DIHYDROOROTATE DEHYDROGENASE OF Toxoplasma gondii

i
DIHYDROOROTATE DEHYDROGENASE OF Toxoplasma gondii:
KINETIC CHARACTERIZATION AND INTRACELLULAR LOCALIZATION
by
MIRYAM ANDREA HORTUA TRIANA
DISSERTATION
Submitted to the Graduate School
of Universidad de los Andes,
Bogota, Colombia
in partial fulfillment of the requirements
for the degree of
DOCTOR OF PHILOSOPHY
2010
MAJOR: Biological Sciences

ii
UNIVERSIDAD DE LOS ANDES
FACULTAD DE CIENCIAS BIOLOGICAS
Name of Student: Miryam Andrea Hortua Triana
Department: Biology
Title of thesis: Dihydroorotate dehydrogenase of Toxoplasma gondii: Kinetic
charactherization and intracellular localization
Director of Thesis: Barbara H. Zimmermann, Ph.D.
Approved by thesis committee:
________________________________________________
Barbara H. Zimmermann, Ph. D.
________________________________________________
David R. Evans, Ph. D.
________________________________________________
Hedeel Guy Evans, Ph. D.
________________________________________________
Monika Löffler, Ph. D.
________________________________________________
Chad Leidy, Ph. D.
Date: Friday, June 25 de 2010

iii
ACKNOWLEDGMENTS
Words are not enough to express my gratitude to my adviser, Dr. Barbara H.
Zimmermann, thanks for your guidance and for the opportunity to learn from you in the lab. She
has mentored me through my work, and in doing so, has led me to develop a profound love for the
sciences. You have my respect and admiration.
I would like also to express my gratitude to my committee members, Drs. David Evans and
Hedeel Guy Evans, I had such a great experience in your lab, it increased my knowledge and passion
for science. To Dr. Monika Loffler, for opening my mind towards biochemical research and teaching
me to take care of all the details to improve the quality of my work. My work time in Wayne State
University and Phillips-Universität Marburg give me a clear vision of working science. Dr. Chad
Leidy, thanks for your suggestions, critiques and support.
Parts of my work could not have been completed without the collaboration of the
extraordinary members of the lab. Manuel Garavito, Heidy Narvaez, Gabriel Lozano, Debora
Primrose, Consuelo Rocha, Enrique Carmargo, Andres Tibabuzo part of the lab family Bioquimica y
Biologia Molecular de Pasitos (BBMP), thanks a lot for you help.
Also I would like to thank all the people that supported me through all this process like Damian
Kotsis, Song Chen, Elizabeth Mazco, Bettina Kowalski, Ute Beck, Elka Zameitat.
Thanks to my father Manuel Hortua, my mother Miryam Triana, my brother Muller
Hortua and Francisco Becerra for their love.
I especially want to thank God because without his help none of this would have been
possible.

iv
TABLE OF CONTENTS
ACKNOWLEDGMENTS ......................................................................................................................... iii
LIST OF FIGURES .................................................................................................................................. vii
LIST OF TABLES ................................................................................................................................... viii
CHAPTER 1 ................................................................................................................................................ 1
INTRODUCTION ....................................................................................................................................... 1
1.1 The Phylum Apicomplexa ............................................................................................. 2
1.2 Toxoplasma gondii .............................................................................................................. 2
1.2.1 Morphology ................................................................................................................... 3
1.2.2 Life cycle ....................................................................................................................... 4
1.2.3 Stage conversion ............................................................................................................ 5
1.2.4 Clonal lineages ............................................................................................................... 6
1.2.5 Epidemiology................................................................................................................. 7
1.2.6 Treatments ...................................................................................................................... 9
1.3 The Apicoplast .................................................................................................................... 9
1.4 Mitochondria .................................................................................................................... 10
1.5 Pyrimidine salvage ........................................................................................................... 11
1.6 Pyrimidine biosynthesis ................................................................................................... 12
1.7 Dihydroorotase dehydrogenase ....................................................................................... 15
CHAPTER 2 .............................................................................................................................................. 18
MATERIAL AND METHODS ............................................................................................................... 18
2.1 Parasite Culture................................................................................................................ 18

v
2.1.1 Tachyzoites grown in mice .......................................................................................... 18
2.1.2 Tachyzoites grown in tissue culture ............................................................................. 18
2.2 Expression and purification of truncated recombinant TgDHOD ............................... 19
2.3 Protein concentration determination .............................................................................. 21
2.4 Protein electrophoresis ..................................................................................................... 21
2.5 Western blotting ................................................................................................................ 23
2.6 Protein A-sepharose immunoprecipitation ..................................................................... 25
2.7 N-terminal sequencing ...................................................................................................... 26
2.8 Enzyme assays ................................................................................................................... 26
2.9 Immunolocalization .......................................................................................................... 28
2.10 Bioinformatics analyses .................................................................................................. 30
CHAPTER 3 .............................................................................................................................................. 31
RESULTS .................................................................................................................................................. 31
3.1 Protein purification ........................................................................................................... 32
3.2 His-tag Western blot visualized with anti-His tag antibodies. ..................................... 34
3.3 Immunoblots of T. gondii tachyzoite extracts ................................................................. 35
3.4 Cross-reaction of TgDHOD-VSSM with antibodies directed against HsDHOD. ....... 36
3.5 N-terminal sequencing ..................................................................................................... 36
3.6 Kinetic characterization ................................................................................................... 38
3.6.1 Kinetic constant ............................................................................................................ 38
3.6.2 Electron acceptors ......................................................................................................... 39

vi
3.6.3 Inhibitors ....................................................................................................................... 40
3.7 Preliminary immunolocalization of dihydroorotate dehydrogenase in T. gondii
tachyzoites ................................................................................................................................ 42
3.8 Predicted model and molecular docking ......................................................................... 47
CHAPTER 4 .............................................................................................................................................. 51
DISCUSSION ............................................................................................................................................ 51
SUMMARY ............................................................................................................................................... 61
REFERENCES .......................................................................................................................................... 62

vii
LIST OF FIGURES
Figure 1. Tachyzoite organelles ...................................................................................................... 4
Figure 2. Life cycle of Toxoplasma gondii ..................................................................................... 5
Figure 3. World seroprevalence of Toxoplasma gondii. ................................................................. 8
Figure 4. T.gondii DHOD amino acid sequences ......................................................................... 32
Figure 5. TgDHOD-VSSM recombinant protein purified protein by His-tag technology ........... 33
Figure 6. Western blot and Gel of purified TgDHOD-VSSM. .................................................... 34
Figure 7. Immunoblot of T. gondii tachyzoite extracts ................................................................. 35
Figure 8. TgDHOD protein cross-react with anti-HsDHOD antibodies ...................................... 36
Figure 9. Purification of a truncated fragment of TgDHOD from tachyzoite extracts ................. 37
Figure 10. Inhibition plots............................................................................................................. 41
Figure 11. Immunolocalization of TgDHOD in tachyzoites ........................................................ 43
Figure 12. Partial localization of TgDHOD to mitochondria ....................................................... 44
Figure 13. Intracellular Golgi-tagged tachyzoites ........................................................................ 45
Figure 14. Immunolocalization of TgDHOD in wild type RH strain ........................................... 46
Figure 15. Immunolocalization of TgDHOD................................................................................ 46
Figure 16. Immunolocalization of TgDHOD with purified anti-DHOD ..................................... 47
Figure 17. TgDHOD structural model .......................................................................................... 48
Figure 18. TgDHOD model for alpha helix A .............................................................................. 49
Figure 19. TgDHOD molecular docking with Redoxal A ............................................................ 50

viii
LIST OF TABLES
Table 1. Kinetic constants of purified recombinant TgDHOD-VSSM........................................ 38
Table 2. Use of electron acceptors by TgDHO-VSSM................................................................. 39
Table 3. Kinetic constant of electron acceptors ............................................................................ 40
Table 4. TgDHOD-VSSM activity in presence of putative inhibitors.......................................... 41
Table 5. IC50 values for DHODs .................................................................................................. 42

1
CHAPTER 1
INTRODUCTION
Toxoplasma and Plasmodium belong to the Phylum Apicomplexa and are parasites of
relevance to the human health. Plasmodium caused the death of eight hundred and eighty
thousand people in 2006 (WHO report 2008). Toxoplasma gondii generates severe clinical
symptoms in the affected population, immunodeficient patients and growing fetuses. Current
prophylactic treatments with sulfadiazine and pyrimethamine are effective, but cannot be used
for pregnant woman (Montoya & Remington, 2008), and cause severe side effects in some HIV
patients (Haverkos, 1987; Luft B et al., 1992; Montoya et al., 2004). Thus, there is a need to
identify new targets for design of less toxic and more effective drugs.
Previous work performed by Fox & Bzik (2001) showed that mutant parasites of T.
gondii lacking the first enzyme of the pyrimidine synthesis lose their virulence and replication
capacities. This points significance of pyrimidine synthesis in the parasite’s viability, and the
importance of identifying targets and designing new drugs to block this pathway.
T.gondii has become a model organism in the study of intracellular pathogens because it is
easy to handle in the laboratory with regard to genetic manipulation, cell markers and stability of
the transfectants. The research breakthrough obtained in T. gondii has been extrapolated to
Plasmodium and other organisms of this Phylum of importance in human and veterinary health.

2
1.1 The Phylum Apicomplexa
The Phylum Apicomplexa includes a group of parasites which have an apical complex of
secretory organelles that take part in the invasion of the host cell (Dlugonska, 2008 & Roos et al.
2000) and in the sexual division by endodiogeny and endopolygeny (Radke et al., 2001).
This Phylum includes causative agents of important parasitic diseases such as Plasmodium,
Eimeria, Cryptosporidium, Neospora, Theileria and Toxoplasma. One of the most important
characteristics of the Apicoplexans is the presence of an apicoplast (Wilson et al., 1993; Palmer,
1992), an organelle homologous to the chloroplast in plants. It is believed that this organelle was
acquired by the endosymbiosis of an alga (Archibald & Keeling, 2002; McFadeen & Dooren,
2004). Since this organelle is not present in mammals, it is a good target for specific new drugs.
Some studies have shown that mutant tachyzoites that lack apicoplast can invade the host cell but
can only replicate once, leading to the death of the parasite (He et al., 2001).
1.2 Toxoplasma gondii
T. gondii is the causative agent of toxoplasmosis. It is an obligate intracellular parasite with
worldwide distribution, and it is capable of using any warm-blooded animal as an intermediate
host (Dubey, 1986), including marine animals such as dolphins (Inskeep et al., 1990; Dubey et
al., 2008). T. gondii was first described by Nicolle and Manceaux in 1908. They isolated it from
the rodent Ctnodactilus gondii, from which the name was derived. Around the same time,
Splendore had reported cases in Brasil.

3
1.2.1 Morphology
In 1973, Frenkel was able to distinguish between three different stages of the parasite.
Tachyzoites replicate quickly and they are clinically related to the acute infection stage of the
disease. Bradyzoites replicate slowly and are the latent stage of the parasite found in tissue
cysts, and are thus associated with the chronic phase of the infection. Oocysts are only formed
in the intestine of the Felinidae family, where sexual reproduction takes place (Frenkel et al.,
1970).
The three stages are very similar structurally. The main differences are size, shape, inclusion
bodies, and organization of the organelles (Dubey, 1996; Dzierszinski et al., 2004). The parasite
has most of the organelles found in other eukaryotic cell. Golgi complex, ribosomes,
endoplasmic reticulum, one large mitochondria and one central nucleus are observed in the
ultrastructural image (se Souza et al., 1978). In addition, in tachyzoites and bradyzoites,
characteristic structures of Apicoplexa, known as rhoptrias and micronemas are found in the
apical area called the conoid. The tachyzoite has a crescent shape, with a size of ~6 x 3μm (Fig.
1), while the bradyzoite has a size of ~4 x 2μm. T. gondii has two other organelles, relatively
recently discovered, the apicoplast responsible for the lipid synthesis (Kohler et al., 1997), and a
plant-like vacuole apparently involved in the endocytic pathway (Miranda, et al., 2010).
Bradydizoites are less susceptible to destruction by proteolytic enzyme than are tachyzoites
(Jacobs et al., 1960). Drugs used in treatment against Toxoplasmosis do not affect the
bradyzoites in tissue cysts (Botero & Restrepo, 2003).

4
Figure 1. Tachyzoite organelles
Schema of tachyzoites from T.gondii (left), and thachyzoites immunofluorescence (right),
nucleus (blue) and apicoplast (yellow) stained (Modified from Black & Boothroyd, 2000).
1.2.2 Life cycle
The life cycle of T. gondii is divided in two: sexual and asexual replication. The first takes
place in the definitive host, members of the felinidae family. When a cat is infected, the parasite
is differentiated into microgametocyte and microgametocyte in the intestinal cells, the fusion of
both produces the oocyst formation.
The asexual component of the life cycle consists in the division of the parasite by
endodogeny or endopolygeny. When the intermediate host ingests infected meat, the parasite
can replicate rapidly in the form of a tachyzoites and then invade other tissues. The immune

5
response of the host can induce stage conversion to bradyzoite. The parasite can camouflage
itself within the cell and while still replicating slowly in the form of the bradyzoite.
Figure 2. Life cycle of Toxoplasma gondii
(Taken from Ferguson, 2010)
1.2.3 Stage conversion
Stage conversion has been studied in vivo as well as in vitro. In vivo the tachyzoite
changes to bradyzoite induced by NO (Bohne et al, 1994), which is produced by IFN-γ. It
affects the metabolism energetic of T. gondii (Bohne et al, 1994).

6
Stage conversion from the tachyzoite to the bradyzoite can be induced in vitro by stress
conditions such as high pH or high temperature, as well as exposure to inhibitors of
mitochondrial respiration, or by reduction of pyrimidine pools under conditions of low levels of
CO2. It appears from expression data that the slowly-replicating stage also sustains the
production of pyrimidines through biosynthesis and salvage pathways. Nevertheless, at the
moment the parasite interconverts to its rapid replicating stage it is likely that it will need to
increase the activity of pyrimidine biosynthesis.
Inhibitors that act on respiration and on the membrane potential in mitochondria, such as the
atovaquone, induce a change from tachyzoite to bradyzoite. A problem with these inhibitors is
that they do not eliminate the parasite, which continues to be viable even under conditions when
there is a decrease in the mitochondrial activity. There appear to be interactions, such as
exchange of metabolites, between the mitochondrion and the apicoplast; however these are not
yet well understood. The relative importance of the organelles in different stages is not clear,
although the bradyzoite appears to depend less on functional mitochondria. It is believed there is
interchange of metabolites between the different membranes. How these factors affect the
interconversion from tachyzoite to bradizoite are not well known.
1.2.4 Clonal lineages
Three clonal strains of T.gondii have been described.(Type I, II and III). They differ in
their virulence and epidemiologic occurrence. Strains Type I and III have been observed in cases
of congenital toxoplasmosis while Type II has been observed in patients with AIDS (Montoya &
Liesenfeld, 2004). Furthermore, some atypical strains difficult to classify have been reported.
Apparently these strains are highly virulent (Grigg & Suzuki, 2003). The increase in the number

7
of the atypical strains has generated the search for specific genetic characteristics to group them
in clonal lineages.
1.2.5 Epidemiology
Toxoplasmosis is a disease that can be asymptomatic in immunocompetent patients, as the
immune response can control the infection inducing the change from tachyzoite to bradyzoite.
This latent form can be of great importance in immunosuppressed patients. It is this group of
patients that has shown the greatest number of cases of reactivation of the disease. In Colombia,
47% of prevalence of toxoplasmosis has been recorded, and 63% of the patients with HIV were
serum positive for toxoplasma (Gomez-Marin et al., 1999). Seroprevalence varies depending on
the region and increases with the age of the patients (Juliao et al., 1988). In a hospital in Neiva,
of 83 HIV positive patients presenting cerebral problems, 37% were diagnosed with toxoplasmic
encephalitis (Avila, et al., 2007). In France 24.5% were reported (Bossi et al., 1998) and in USA
(Jones et al., 2002) 27% before Highly Active AntiRetroviral Treatment (HAART).

8
Figure 3. World seroprevalence of Toxoplasma gondii.
60% prevalence (dark red), 40-20% prevalence (light red), 10-20% blue and less 10% prevalence
(yellow). No data reported (white). Take form Pappas et al., 2009.
Toxoplasmosis is also very important when the infection takes place in pregnant women,
as the infection can be transmitted to the fetus. The first case of congenital toxoplasmosis was
described by Wolf in 1939. The gravity of the infection depends on the time of gestation. The
signs can be serious, causing blindness, brain calcifications, mental retardation or even death.
Studies performed in Colombia show that between 53 and 60% of pregnant women have
antibodies to toxoplasma and between 0.6 and 3% were infected during pregnancy (Gomez-
Marin, 2007). A prevalence of congenital toxoplasmosis of 0.62% (Gomez-Marin, et al., 2008)
was reported in the Quindío area. The former data show that there is high exposure to and
circulation of the parasite in Colombia has also seen in other parts of the world. For this reason,

9
it is important to perform genetic studies of the parasite in order to classify the types of parasites
associated with geographical distribution and populations affected.
1.2.6 Treatments
Although there have been advances in diagnosis that can differentiate between the clonal
types, there is only a single treatment for Toxoplasma. Prophylactic treatment is generally
limited to the use of pyremethamine, sulphadiazine and/or folic acid, but these drugs are only
directed against the tachyzoites as they have little effect on the bradyzoites. Another problem is
that these drugs are not well-assimilated by the patients with AIDS. This is an important reason
why the search for new targets is a priority in order to design more efficient drugs. New research
depends on an in-depth understanding of different metabolic pathways that may be used to block
the replication of the parasite. The synthesis of pyrimidines is one of the most promising
metabolic pathways to be used as a target. This pathway produces nucleotides and nucleosides
essential for the synthesis of DNA. It begins with the synthesis of the pyrimidine ring and
followed by the addition of the sugar.
1.3 The Apicoplast
The apicoplast is a circular organelle surrounded by four membranes (Köhler et al., 1997). It
contains a DNA conserved among the Apicoplexa, and which is approximately 35kb in T. gondii
(Gardner, 2005). The apicoplast also contains repeated coding sequences for rRNA as is also
observed in chloroplasts (Gillham, 1994). Most proteins found in the apicoplast are coded for by
nuclear genes. Some of the functions of this organelle are not yet known, although studies have
shown that it is responsible for the synthesis of lipids (fatty acids and isoprenoids) (Zuther et al.,

10
1999). Some studies on the redox state of the apicoplast have been performed (Seeber et al.,
2005). In P. falciparum the movement of electrons of NADPH to ferredoxin has been described
(Vollmer et al., 2001). It has been speculated that T.gondii presents a similar electron transport
system which would allow several biosynthetic processes to take place in this organelle. The
apicoplast and the mitochondrion have been studied as targets for the development of new drugs,
especially those that inhibit of DNA replication, transcription of the RNA, translation of proteins,
and the synthesis of fatty acids and isoprenoids (Waller et al., 2005).
The identification of proteins localized in the apicoplast by the isolation of this organelle
using differential centrifugation has not been. This is because the membrane of the apicoplast is
very sensitive and can be found in all the cellular fractions. It has also been observed that the
florescent markers used to tag proteins for immunolocalization can affect the cellular targeting of
the tagged protein (Carruthers, 2008, personal communication; Pino et al., 2007).
1.4 Mitochondria
The mitochondrion although this is one of the largest organelles within the cell, little is
known about their true function and metabolism in T. gondii although some studies have been
performed (Köhler et al., 2006). The parasite in contrast to higher eukaryotes, has one (or few)
mitochondria in the cytosol whose morphology may vary. Mitochondria as well as Gram-
negative bacteria have two membranes (inner and outer). In eukaryotes usually the
outermembrane and intermembranal space are essentially involved in the communication
between organelles, transport and preliminary processing of metabolites (Köhler et al., 2005).

11
This membrane is permeable to ions and molecules that cross the channels generated by
porins. Unlike the external membrane, the inner membrane is impermeable to most of these ions
and molecules. In the inner membrane takes place most of the reactions because it contains the
electron respiratory chain (complex I - IV), ADP / ATP translocase, ATP synthase and other
membrane transporters. Finally, the inner membrane contains the mitochondrial matrix pyruvate
dehydrogenase complex as well as the enzymes involved in metabolic pathway of B-oxidation of
fatty acids, citric acid cycle and amino acid oxidation route (Nelson et al. 2003).
In T. gondii the ability of mitochondria in the tachyzoite to carry out the functions of electron
transport coupled to respiration (Coombs et al., 1997) , generates a membrane potential sufficient
to carry out certain processes, their ability in the bradyzoites (less dependent on breathing) has
been questioned (Tomavo et la., 1995; Coombs et al., 1997). T. gondii has been proposed that the
apicoplast may contain components of the respiratory chain which are normally attached to the
inner membrane of mitochondria.
1.5 Pyrimidine salvage
There are two ways in which the parasite can meet its pyrimidine requirements, by the
salvage pathways (Iltzsch, 1973), or through de novo synthesis, as observed in mammalian cells.
The salvage pathways recycle nucleosides and nucleobases. In T. gondii this pathway is
represented only by three enzymes: uridine phophoribosyltransferase (UPRT) and a low activity
of cytidine and deoxycytidine deaminase.
Uracil, cytidne and deoxycytidine can be used as sources for uridine monophosphate
(Iltzsch, 1993). Cytidine and deoxycytidine are transformed to uridine and deoxyuridine,

12
respectively, by deamination. The nucleosides are then transformed to the nucleobase uracil
through the action of the uridine phosphorylase (Iltzsch, 1993). Uracil is metabolized to uridine
5’monophosphate by the T.gondii UPRTase. T. gondii UPRT is the only enzyme that can
salvage a nucleobase, uracil, to the nucleotide level (Iltzsch, 1993). The UPRTase catalyzes the
transfer of a ribosyl phosphate group to uracil to form UMP and PPi (Schummer et al., 1998).
The UPRT from T.gondii has been isolated and cloned (Carter et al., 1997).
In bradyzoites pyrimidine biosynthesis and salvage pathways are not well characterized,
even thought the expression of some of the enzymes of both pathways have been reported in this
stage of the parasite.
The T.gondii pyrimidine salvage pathway is different from that of its human host. The
human host salvages machinery can phosphorylate pyrimidine ribosides and deoxyribosides to
nucleosides 5’ monophosphates. T. gondii cannot do so because it lacks the pyrimidine
nucleoside kinase (El kouni et al., 1996). El Kouni group found differences in the binding host
and parasite uridine phosphorylases.
1.6 Pyrimidine biosynthesis
The synthesis of pyrimidine pathway produces UMP, which is a starting point for the
synthesis of other nucleotides such as adenine, thymine and uracil, is composed of six steps.
This pathway uses as precursors bicarbonate, glutamine, ATP, aspartate y phophoribosyl
pyrophosphate (PRPP) to produce uridine monophosphate (UMP). Studies in mammalian cells
by Patterson (1978), show that mutant cells with dysfunction in the first 3 enzymes of the

13
synthesis grew normally if uridine was supplied, thus showing that mammalian cells can be
sustained with the salvage pathways.
In contrast, studies by Fox & Bzik, show that mutant tachyzoites lacking the first enzyme
of the pathway, carbamoyl phosphate synthetase (CPSII), causes lose of virulence and replicating
capacity. This suggests that pyrimidine biosynthesis is essential for the growth, replication and
virulence of T. gondii. Additionally, Pfefferkorn (1978) reported evidence showing that the
contribution of pyrimidines by the salvage pathways in tachyzoites was not significant, since
mutant tachyzoites with a deficiency in UPRT were indistinguishable from wild-type parasites
under most conditions. A better understanding of the function and localization of pyrimidines in
the parasite could supply information that could lead to the design of inhibitors in different
functional stages for tachyzoites and/or bradyzoites.
Pyrimidine biosynthesis is conserved in mammals, plants, bacteria and T. gondii,
however various differences have been described. For example, mammalian cells have two
carbomoyl phosphate synthetases (CPSI and CPSII). CPS I provides carbamoyl phosphate for
arginine biosynthesis, while CPSII provides this intermediate for pyrimidine (Jones, 1980).
Additionally, mammalian CPS II is part of a multifunctional protein named CAD that contains
also the enzymes aspatate transcarbomilase (ATC) and dihidroorotase (DHO), the next enzymes
of the pathway. CAD is a polypeptide of approximately 243 kDa that includes also the
subdomain of glutaminase required for the CPSII activity. This multifunctional complex
localized in the cytoplasm is a point of regulation of the pathway, activated by PRPP and
inhibited by UTP (Evans & Guy, 2004).

14
T. gondii has a CPSII responsible for catalysis in pyrimidine biosynthesis but it is
auxotrophic for arginine biosynthesis and does not contain a CPSI (Fox et al., 2004). The
sequence that codes for CPSII in T. gondii is characterized by large introns, and differs in its C-
terminus, the site of allosteric regulation by PRPP (Fox & Bzik, 2003). However, inhibition by
UTP has been observed (Asai, 1983). The first three enzymes (CPSII, ATC and DHO) are
encoded by different genes which constitute independent polypeptides. A CAD complex is not
present in T. gondii, each activity step of activity appears to be independent. ATC has a
molecular mass of 46.8 kDa and does not appear to be regulated by nucleotides (Mejias-Torres &
Zimmermann, 2002), and DHO has a molecular mass of 44.2 kDa (Robles-Lopez, et al., 2006)
and was found in a cytosolic localization.
The first three enzymes in bacteria and plants are similar. In this case CPS I supplies
both the pyrimidine and arginine pathways (Doremus, 1986). CPS of plants has a glutamine and
amidotransferase subunits, and a regulatory activity is in the amino domain. CPS, ATC and
DHO of plant are encoded in the nucleus and targeted to the chloroplast (Kafer et al., 2004).
DHO amino acid sequence has a predicted chloroplast or mitochondria signal peptide in the N-
teminus (Zrenner et al., 2006).
The fifth and sixth enzymes of the pathway, orotate phosphoribosyltransferase (OPRT,
EC 2.4.2.10) which catalyzes the formation of oritidine 5’-monophosphate (OMP) from orotate
and 5-phosphoribosyl-1-pyrophosphate (PRPP) and orotidine 5’-monophosphate decarboxylase
(ODC, EC 4.1.1.23) which catalyzes the subsequent decarboxylation of OMP to uridine 5’-
monophosphate (UMP) have not been identified. In mammals these two enzymes are present in a
single gene, encoding the multifunctional protein uridine monophosphate synthethase (UMPS)
(Livingstone & Jones, 1987), in most prokaryotes (Jensen et al., 1984), yeast (Umezu et al.,

15
1971) and apicomplexan parasite such as P. falciparum (Krungkrai et al., 2004A; Krungkrai et
al., 2004B) the enzymes are coded by two separate and independent genes. As well, there are
reports of different events of horizontal gene transfer and fusion of these two enzymes in other
eukaryotic groups (Nara et al., 2000; Makiuchi et al., 2007). Previous works demonstrated the
potential of the T. gondii OPRT as a druggable target to find inhibitors that bind preferentially to
the parasite enzyme or mammalian enzymes (Javaid et al., 1999; Niedzwicki et al., 1984). There
are reports of different inhibitors of OPRT and ODC, in another apicomplexan parasite, P.
falciparum, which have high anti-malaria activities (Seymour et al., 1994; Scottet al., 1986) or
bind preferentially to the parasites enzyme (Krungkrai et al., 2004A; Krungkrai et al., 2004B;
Krungkrai et al., 2005; Bello et al., 2008).
1.7 Dihydroorotase dehydrogenase
The fourth enzyme of the de novo pathway, dihydroorotate dehydrogenase (DHOD, E.C.
1.3.3.1), catalyzing dihydroorotate oxidation to orotate, appears to be a promising target. This
enzyme is the target of drugs used for the treatment rheumatoid arthritis and other autoimmune
diseases (Kovarik & Burtin, 2003; Loffler et al., 2005), and is being intensively studied as an
antimalaria therapeutic target (Baldwin et al., 2002; Baldwin et al., 2005; Heikkila et al., 2007;
Phillips et al., 2008; Gujjar et al., 2009; Davies et al., 2009).
DHODs are classified into two families. Family 1 DHODs are soluble enzymes found in
Gram-positive bacteria, archaea, and lower eukaryotes. These are further subdivided into family
1A, FMN-containing homodimeric enzymes that use fumarate as the electron acceptor (Rowland
et al., 1997), and family 1B heterotetrameric enzymes that use FMN, FAD and iron/sulfur
clusters as redox centers, and NAD+ as the electron acceptor (Rowland et al., 2000; Nielsen et

16
al., 1996). Family 2 DHODs are membrane-associated and found in gram-negative bacteria and
eukaryotes. They are flavoproteins, usually located at outer surface of the plasma or inner
mitochondrial membranes, where they transfer electrons via FMN to quinones and are thus
linked to the respiratory chain.
Multiple sequence alignments of the T. gondii DHOD (TgDHOD) with DHODs from
other organisms indicate that it is most similar to family 2 enzymes (Sierra Pagan &
Zimmermann, 2003). An important difference between family 1 and 2 enzymes is that the latter
contain extended N-termini that play roles in targeting and membrane association (Rawls et al.,
2000; Hansen et al., 2004). The N-terminal extension of TgDHOD is comprised of ~157
residues on the N-terminal side of a predicted transmembrane segment, and is the longest found
to date (Sierra Pagan and Zimmermann, 2003). A slightly shorter N-terminal extension (~143
residues) is found in the DHOD from P. falciparum (PfDHOD), while a relatively short
extension (~ 13 residues) in the human enzyme. DHOD crystal structures of family 2 enzymes
are available for Escherichia coli (Nørager et al., 2002; Fagan et al., 2009), human (Liu et al.,
2000; Baumgartner et al., 2006), rat (Hansen et al., 2004) and P. falciparum (Hurt et al., 2006a;
Deng et al., 2009). The structures of the eukaryotic enzymes are of recombinant proteins
truncated at the N-termini, thereby eliminating targeting signals and the transmembrane segment
thought to serve as a membrane anchor. The structural studies reveal a large domain, highly
conserved among family 2 enzymes, consisting of an α/β barrel structure containing the active
site with a conserved catalytic serine (S175 in the E. coli enzyme, S215 in HsDHOD). The
serine is in close contact with the substrate, dihydroorotate, and has been proposed to remove a
proton from C5 (Liu et al., 2000; Nørager et al., 2002; Hansen et al., 2004). A second, smaller,
domain near the N-terminus is composed of two α-helices forming the entrance of a tunnel to the

17
dihydroorotate oxidation site. This domain is the binding site for leflunomide, brequinar, and
atovaquone and their derivatives and triazolopyrimidine derivatives, and is the predicted binding
site for ubiquinone (Hurt et al., 2006b). Differences in longitudes and orientations of the first
alpha-helix in the different DHODs (Nørager et al., 2002) suggest differences in ubiquinone- and
inhibitor-binding that could account for the observed differences in sensitivity to inhibitors (Hurt
et al., 2006b; Deng et al., 2009). Studies on the PfDHOD show that potent human DHOD
inhibitors have no significant effect on the parasite enzyme, and PfDHOD inhibitors are not
cytotoxic to kidney tissue (Patel et al., 2009). For these reasons, DHOD is likely to be a good
target for new anti-toxoplasma drugs.

18
CHAPTER 2
MATERIAL AND METHODS
2.1 Parasite Culture
2.1.1 Tachyzoites grown in mice
The T. gondii virulent strains RH and 5TN were maintained by serial passage in the
peritoneal cavity of CFW or Swiss mice (Derouin et al., 1987). The strain 5TN was the gift of
Claudia Herrera (Centro de Investigaciones en Microbiología y Parasitología Tropical,
Universidad de los Andes, Bogotá, Colombia) and mice were provided by the Universidad de los
Andes animal facility.
Peritoneal exudates containing tachyzoites were harvested three to five days after
intraperitoneal injection of seven week-old mice with 7.4 x 106 tachyzoites in PBS 1X medium
(1.8 mM potassium phosphate, 137 mM NaCl, 2.9 mM KCl, 10 mM sodium phosphate at pH
7.4). Tachyzoites were counted in a Neubauer chamber.
In our restriction fragment length polymorphism (RFLP) analyses of strain 5TN, the amplified
portions of the B1 and SAG1 genes (Howe et al.; 1997; Grigg and Boothroyd, 2001) gave the
same restriction patterns as a type I strain.
2.1.2 Tachyzoites grown in tissue culture
Human foreskin fibroblast (HFF) cells were grown at 37°C and 5% CO2 in 15 mL of D10
growth medium (Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% fetal
calf serum (FCS), 2 mM glutamine, 5000 IU/ml penicillin, and 5 mg/mL streptomycin) into T25

19
or T75 cell culture flasks. This cell line was maintained in a humidified incubator at 37oC and
5% CO2. The next day the medium was replaced with pre-warmed 15 mL of D10. When the
cells reached 95% confluency, they were subcultured. Infection with T.gondii freshly lysed from
host cells was done with just 50% cells confluency.
The amount of T.gondii lysed used for infection was determined by the flask size and the
desired time to cell lysis. After selection by viewing with a microscope, the flask with the best
parasite concentration was selected, and 1ml of lysed T.gondii was used to infect HFF culture in
T25 flask to achieve lysis overnight, while eight drops were used to achieve lysis in two days.
When frozen stock was required, a regular tissue culture of HFF cells was grown and
infected with T.gondii the day before desired freezing. The infected cells were washed 2 times
with 1X PBS, and then 0.05% trypsin was added per T25 flask. After incubated 2 min to permit
detachment cells, Trypsin was stopped with 1 mL DMEM + 50% FBS. Cells were collected in a
15 mL tube and kept on ice for 10 min, 0.5 mL DMEM + 20% dimethyl sulfoxide (DMSO) was
then added. 1mL volumes of this cells suspension were aliquoted into pre-cooled cryovials, wich
were then placed into a Styrofoam cooler and transferred to -70oC freezer. The next day
cryovials were transferred to a liquid nitrogen storage container.
2.2 Expression and purification of truncated recombinant TgDHOD
TgDHODpET19b-MIYS, TgDHODpET19b-FYEP, or TgDHODpET19b-VSSM (all are
recombinant truncated proteins, where the last four letters indicate the first four amino acids at
the truncated N-terminus) were transformed into BL21-CondoPlus(DE3)-RP cells (Stratagene)
for protein expression. In this E. coli strain, T7 RNA polymerase is under control of the lacUV5
promotor for easy protein expression, which is inducible with isopropylthio-β-galactoside

20
(IPTG). E.coli BL21 transformed with the plasmids were placed on LB plates with 100 µg/mL
ampicillin and incubated overnight at 37oC. The next day, colonies were transferred to 5 mL LB
media (Sambrook, Fritsch, Maniatis, 1989) in 15 mL tubes containing 100 µg/mL ampicillin.
The tubes were placed overnight in a shaker (225 rmp) at 37oC. The following day, the 5 mL
culture was inoculated into 500 mL of LB media containing 100 µg/mL ampicillin and was
incubated at 37°C.
When cultures reached OD600 = 0.5, 1 mM IPTG and 0.1 mM riboflavin 5’-phosphate
sodium salt (FMN) were added and the temperature was lowered to 25°C. FMN is a cofactor
required for DHOD catalysis; it is tightly bound to the enzyme (Baldwin et al., 2002).
Cells were harvested 20 hours after induction. The purification procedure was based on
that developed for recombinant Plasmodium falciparum DHODase (Baldwin et al., 2002) with
modifications. Cells were resuspended in 5 mL of buffer A (2 mM beta-mercaptoethanol, 2%
Triton X-100, 10% glycerol, 0.5 mM FMN, 50 mM Tris-HCl, pH 8.5) in the presence of 1 mM
phenylmethanesulfonyl flouride (PMSF) and 1 mM benzamidine. The solution was frozen at -
80°C, thawed, and submitted to sonication on ice for a total of 12 min (model W-220F sonicater,
Heat Systems Ultrasonics), followed by centrifugation at 10,000 x g and 4°C for 30 min to
remove cell debris. An ultracentrifugation step was found to be unnecessary.
Recombinant proteins were purified from the supernatant using Ni-NTA technology
(Qiagen). The nickel column was prepared by adding 1 mL resin into chromatography column.
The resin was equilibrated with 20 mL buffer B (buffer A containing 300 mM NaCl, and 20 mM
imidazole). Next, 5 mL of supernatant containing the protein was applied to the resin. The beads
were then washed 4 times with 5 mL buffer B containing 40 mM imidazole.

21
Fractions containing recombinant protein were eluted with buffer B. Prior to measuring
activity, recombinant protein-containing fractions were prepared by using PD-10 desalting
columns (GE Healthcare, 17-0851-01) equilibrated with 300 mM NaCl, 10% glycerol, 0.1%
Triton, 50 mM Tris-HCl, pH 8.5, at 4°C or by dialyzing against the same buffer at 4°C with three
buffer changes. SDS-PAGE was performed to check fractions containing recombinant protein.
2.3 Protein concentration determination
Protein concentration was measured by the bicinchoninic (BCA) protein assay kit (Pierce,
23227). This kit was selected because it is compatible with high Triton TX-100 (up to 5%)
concentrations. A standard curve was done with bovine serum albumin (BSA) using
concentrations between 0 to 225μg/mL, using the microplate in accordance with the
manufacturer’s instructions; 25 μL of each standard or sample was pipetted into a microplate
well. Then, 200 μL of working reagent (a mixture of 50 parts of BCA Reagent A with 1 part of
BCA Reagent B) was placed into each well and mixed on a plate shaker for 30 seconds. After
covering the plate was incubated for 30 min at 37oC. Finally, absorbance was measured at 595
nm in a microplate reader (BioRad, Model 680).
2.4 Protein electrophoresis
SDS-polyacrylamide gel electrophoresis (SDS-PAGE) was carried out on gels of 0.75mm
or 1mm thickness, using 7.5 – 15% gradient running gels with 5% stacking gels, with the buffer
system described by Laemmli (1970). The running gel was prepared with 1.25 mL running gel
buffer (1.5M Tris-HCl, pH 8.8), 0.05 mL 10% SDS and the appropriate amount of acrylamide

22
using a 30% stock solution (29.2% acrylamide w/v and 0.8% N,N-methylene-bis-acylamide
w/v). To polymerize the gel 0.2% 1,2 ethanediamine N1,N2,N3, N4-tetramethylethylenediamine
(TEMED, Promega) and 0.2% ammonium persulfate (APS, Promega) were added to the
solution, and it was immediately injected into 10 x 8 cm or 20 x 20 cm plates assembled on the
casting unit. A thin layer of water was then added to the surface of the running solution to
ensure a level surface. After the gel had polymerized the water was removed and the stacking
gel solution was added.
The stacking gel was prepared the same way as the running gel but with 0.64 mL
stacking gel buffer (1M Tris-HCl, pH 6.9). After the gel had polymerized; it was placed in 1X
running electrode buffer (0.195 M glycine, 0.025 M Tris-HCl and 0.003 M sodium dodecyl
sulfate). The electrophoresis was performed in BioRad Mini-PROTEAN or PROTEAN II xi
electrophoresis cells.
Protein samples were boiled for 10 min at 90oC in 5X loading buffer (0.275 M Tris-HCl,
17.5% glycerol, 0.008% bromphenol blue, 3 M 2-mercaptoethanol and 0.04 M
ethylenediaminetetraacetic acid (EDTA), pH 7.0). After, the samples were load into the gel and
the run gel started. The power supply was programmer at 80 V of voltage constant until the dye
front cross the stacking gel and then, voltage was sending to 165 V until the dye front reached
the bottom gel. The gel was stained if it was not used for Western blot. To stain the gel was
placed in a solution containing 0.1% Coomassie brilliant blue R-250 in 50% ethanol and 10%
acetic acid in distilled water. The gel was rinsed in distainer solution (30% ethanol and 10%
acetic acid in distilled water) several times.

23
2.5 Western blotting
Protein samples were electrotransferred from gels to 0.45 μm nitrocellulose membranes
(Biorad 162-0145) using a BioRad Mini Trans-blot System. The gel sandwich was prepared by
placing filter paper on the fiber pad and then placing the gel on the top of the filter paper. A pre-
wet piece of nitrocellulose was placed on the gel. Finally, another piece of filter paper was
placed on the top of the nitrocellulose membrane, which was covered with the fiber pad to
complete the sandwich. Care was taken during this process to ensure no air bubbles were present
in the sandwich. The gel cassette was closed and placed into the blot unit with the membrane
facing the anode. Electrotransfer was performed in 1X transfer buffer (20 mM Tris-HCl pH 8.3,
150 mM glycine and 20% ethanol) at 100 V for 1 hour.
When the transfer was complete, the membrane was removed from the sandwich and
incubated at room temperature with agitation for 45 min in 1X Tris Buffer Saline (TBS)/Milk
(10 mM Tris-HCl and 150 mM NaCl pH 7.4 and 5% non-fat dry milk Colanta). Afterwards, the
membranes were washed 3 times for 5 min with 1X TBS/Tween (1X TBS and 0.05% Tween 20),
followed by overnight incubation at room temperature with primary polyclonal antibodies
(Strategic Biosolutions, Newark, DE) directed against purified, truncated recombinant T. gondii
DHODase (TgDHOD-ALQD (Sierra Pagan and Zimmermann, 2002)) at a dilution of 1:7.500.
After 3 times more washes, the membrane was incubated at room temperature for 1 hour at a
1:1500 dilution of anti-rabbit IgG peroxidase conjugate produced (Sigma, A-8275). Finally, the
membranes were washed again 3 times for 5 min with 1X TBS/Tween.

24
Visualization was performed with 2.8 mM 4-chloro-1-naphthol, 0.01% H2O2, 16.7%
methanol in TBS. Antibodies produced in two different rabbits exhibited similar reactivities to
TgDHOD recombinant proteins.
Purified anti-TgDHOD antibodies in a dilution of 1:25.000 were used to against T. gondii
DHOD enzyme from lysed parasite. Extracelullar parasites from one 25 flask was collected in a
15 mL falcon tube and then washed three times with Hank’s-HEPES-EGTA (HHE) (Hank’s
Balanced Salt Solution (HBSS), 1M HEPES solution and 0.1M EGTA pH 7.5). Tachyzoites
pellet was resuspended in HHE and then concentration calculated by Multisizer™ 3 Coulter
Counter® (Beckman). 7.5x106 thachyzoites was denatured with 5x loading buffer and heat at
90oC for 10 min. SDS-PAGE and Western blotting was performed as was described before. The
secondary antibody used was horseradish peroxidase (HRP) conjugated goat anti-rabbit IgG
(Sigma) at a dilution of 1:5000. Chemiluminescence system was used to developing
(SuperSignal West Pico Chemiluminescent Substrate kit #34080. Pierce).
Another Western blot was performed with affinity-purified rabbit antibodies raised
against human DHOD. The membrane was exposed to a 1:500 dilution of anti-HsDHOD in 20
mM PBS, 3% BSA and 0.05% Tween-20. The secondary antibody used was horseradish
peroxidase (HRP) conjugated goat anti-rabbit IgG (Sigma) at a dilution of 1:20.000, followed by
visualization using an enhanced chemiluminescence (ECL) detection kit (# RPN2106 GE-
Healthcare).

25
2.6 Protein A-sepharose immunoprecipitation
Immunoprecipitation samples were prepared by resuspending 1.8 x 109
tachyzoites in a
lysis buffer containing 5 mM potassium phosphate, pH 7.0, 2X Halt TM Protease and
Phospahtase Inhibitor Cocktail (Pierce # 78443) supplemented with 0.06 mM chymostatin, 0.15
mM antipain, 0.10 mM leupeptin, 1 mM PMSF, 1 mM EDTA, 1 mM dithiothreitol, 2% Triton
X-100, and the solution was then subjected to 15 freeze/thaw cycles using with nitrogen and 8
sonication cycles (model W-220F sonicater, Heat Systems Ultrasonics). Lysates were
centrifuged at 4°C, 16.000 x g, 20 min.
The protein A sepharose (Sigma #P3391) was resuspended (250 mg per 2 mL) in
phosphate buffer pH 7.0 (38.2 mM K2HPO4 and 227.8 mM Na2PO4) for 15 min. The resin was
washed with 10 volumes 1X PBS to remove protein stabilizers (dextrans and lactose) and then
resuspended in 1 mL 1X PBS.
A volumen of 0.4 mL of lysate supernatant was incubated 5 hours with 3 μL polyclonal
antibodies directed against purified recombinant protein TgDHOD-ALQD (Sierra Pagan and
Zimmermann, 2002) frequently mixing on ice. Next, this solution containing the protein
bounded to the antibody was incubated at 4oC for 3 hours with 20 μL protein A sepharose
previously hydrated and washed. After 3 washes with 1X PBS (15 min at 1700 x g) TgDHOD
protein, antibody and resin complex were heat separated at 93oC for 15 min in the presence of
100 μL SDS-PAGE loading buffer.

26
2.7 N-terminal sequencing
SDS-PAGE electrophoresis was performed as described in section 2.4 using BioRad
PROTEAN II xi electrophoresis cells to get better bands separation. After, SDS-PAGE, the
protein was transferred to polyvinylidene difluoride (PVDF) membranes (pre-wet with 100%
methanol) using transfer buffer 20 mM Tris-HCl pH 8.3, 150 mM glycine and 20% methanol.
The membrane was stained for 20 min on shaker with a solution of 1% acetic acid, 20%
methanol and 0.2% Coomassie brilliant blue R-250 filtered that had been previously filtered.
Bands visualization was obtained after destaining 20 min with 30% methanol in freshly
deionized water.
After air-drying the membrane, the band at approximately 45 kDa was cut from the
membrane with a razor blade and sent to for N-terminal amino acid sequencing by Edman
degradation at the Yale University Keck Biotechnology Resource Laboratory.
2.8 Enzyme assays
Activities of purified recombinant proteins were measured by monitoring 2,6
dichlorophenol-indophenol (DCIP) reduction, a reaction coupled to dihydroorotate substrate via
quinone cosubstrate oxidation (Knecht & Löffler, 1998). DCIP reduction assays were performed
at 30°C in a reaction buffer containing 50 mM Tris-HCl pH 8.0, 150 mM KCl, 0.1% Triton X-
100, 1 mM dihydroorotate, 0.1 mM DCIP, 0.1 mM ubiquinone, with a concentration of 6.2 nM
of recombinant TgDHOD. (Zameitat et al., 2006). DCIP reduction was measured at a 600 nm (ε
= 18.800 M-1
cm-1
) (Zameitat et al., 2006). Since DCIP can accept electrons in absence of
ubiquinone, this background activity was measured (kcat = 28.1 ± 0.7 sec-1
) and was subtracted
from activities in the presence of the ubiquinones shown in Table 1. The activities of quinones at

27
0.1 mM, such as PQ0, Q10, and menadione (Ullrich et al., 2002; Zameitat et al., 2006), were
measured and compared to the activity in the presence of QD.
Ubiquinones were dissolved in absolute ethanol to make stock solution of 10 mM,
resulting in a final ethanol concentration of 1% in the assay. Q10 presented solubility problems;
to prevent precipitation the ethanol concentration in the reaction assay was raised to 10%, a
concentration that does not affect enzyme activity (Ullrich et al., 2002).
The kinetic constants of both substrates dihydroorotate and decylubiquinone (QD) were
determined by varying dihydroorotate concentration (5 µM – 1.0 mM) while keeping QD
constant at 100 µM, or by varying QD (0.1 µM – 100 µM) at a fixed dihydroorotate
concentration of 1mM. Saturation curves were also performed using 1 mM dihydroorotate, 0.1
mM DCIP, while varying concentrations (0.1 µM – 100 µM) of ubiquinone-0 (Q0), ubiquinone-6
(Q6), or ubiquinone-10 (Q10). The Michaelis-Menten equation v = Vmax x (S)/ (Km + (S)) was
used to calculate Km (SigmaPlot 8.0). The kcat was calculated from kcat = Vmax/(ET), where (ET) is
total enzyme concentration, based on one active site monomer.
An alternative assay measuring appearance of orotate at 30°C at 280 nm (ε = 7.500 M-
1cm
-1) (Zameitat et al., 2004) was performed using 1 mM fumarate as the electron acceptor
(Zameitat et al., 2004). The same assay was performed using 1 mM ferricyanide (K3Fe(CN)6) as
the electron acceptor at the isosbestic wavelength 450 nm (ε = 1.020 M-1
cm-1
) (Zameitat, et al.,
2007).
To determine the effect of inhibitors, DHOD activity was measured by the DCIP
reduction assay with QD as the ubiquinone in presence of the different inhibitors at 1mM
concentration. Fresh stock solutions of redoxal, 0.5mM toltrazuril, 0.1 mM atovoquone , A77-

28
1726 , 2-thenoyltrifluoroacetone (TTFA), brequinar, dichloroallyl lawson (DCL) and ectoine
were prepared. Stock solutions of the first three compounds were prepared in dimethyl sufoxide
(DMSO), while those of the remaining compounds were prepared in the assay buffer solution.
IC50 values were determined for A77-1726 and Redoxal. Fixed saturating concentrations
of the substrates dihydroorotate (1 mM) and QD (0.1 mM) were used, while inhibitor
concentrations were varied between 1 µM – 2 mM for A77-1726 and 0.5 µM - 1.25 mM for
Redoxal.
2.9 Immunolocalization
Three different T. gondii strains were used to make intracellular and extracellular slides, a
wild type strain, a strain with yellow fluorescent protein tagged acyl-carrier protein (YFP-ACP)
(Huynh and Carruthers, 2009) and a strain with red fluorescent protein tagged Golgi reassembly
stacking protein 55 (GRASP55) (Pelletier et al., 2002; Hartmann et al., 2006).
Slides with intracellular parasite were prepared growing human foresking fibroblast
(HFF) cells on eight-well glass Nunc Lab-Tek chamber slides (Thermo Scientific) 24 hours
before infection. HFF cells on slides were infected with fresh T. gondii culture (3 or 6 μl) in
invasion buffer (DMEM supplemented with 3% fetal bovine serum (FBS) and 10 mM HEPES)
and grown for 18 hours.
Extracellular parasites were prepared by passing twice through needles and filtering
through a 3 µm filter (Brydges and Carruthers, 2003). Extracellular parasites were attached on
eight-well glass Nunc Lab-Tek Chamber slides (Thermo Scientific), with BD Cell TakTM
adhesive (BD Biosciences) with approximately 100 µL of 1x107 parasites/mL in each well.

29
Both extra- and intracellular parasites were fixed with 4% formaldehyde for 20 min at room
temperature.
Slide preparation was carried out at room temperature unless otherwise noted, and
incubations were performed using gentle agitation. Parasites were permeabilized with 0.3%
Triton X-100 for 1 hr, washed three times with 1X phosphate-buffer saline (PBS) and blocked
with 10% BSA or 10% FBS for 30 minutes, or with 3% BSA for 1 hour, and washed three times
with 1% FBS, 1% NGS and 0.03% Triton X-100 in PBS pH 7.4. Slides were incubated with the
primary antibodies or pre-immune serum diluted 1:300 overnight at 4 °C. Slides were washed
and then incubated with secondary antibodies (Molecular Probes) conjugated with Alexa Fluor
594, diluted 1:1000, or conjugated with Alexa Fluor 488, diluted 1:3.000, for 1 hour, and washed
as before. Some slides were incubated with 4.0 µM 4’, 6-diamino-2-phenylindole dilactate
(DAPI) (Invitrogen) in 0.03% Triton X-100 PBS pH 7.4 after antibody incubations. Slides of
wild type parasites (RH strain) were stained with 100 nM Mitotracker Red in normal growth
medium (DMEM with 10% Cosmic Calf Serum). Slides were treated with the anti-fade agent
Mowiol (Calbiochem), and kept at 4°C in a humid chamber until observation. Slides were
examined with a Zeiss Apo Tome Microscope in Confocal mode using four channels at the
Microscopy Imaging Resources Laboratory in the Department of Pharmacology at Wayne State
University School of Medicine.
Anti-TgDHOD polyclonal antibodies were purified by affinity column using the
AminoLink® Immobilization kit (Pierce, #44890). The recombinant purified TgDHOD-VSSM
protein was immobilized in the column and used to couple the antibody. Purified anti-TgDHOD
antibodies were eluted at 150mM glycine pH 2.5 and neutralized with 1M Tris pH 9.0.

30
Inmunolocalization with purified anti-TgDHOD antibodies in a 1:400 dilution were
performed in extracellular and intracellular parasites as described before.
2.10 Bioinformatics analyses
Prediction of the DHOD 3D structure was performed by 3D-JIGSAW modeling program,
consulted on line (http://www.bmm.icnet.uk/servers/3djigsaw/). Human DHOD crystallographic
structure, reported in protein data bank (2PRM) was the template used to predict the DHOD T.
gondii 3D model. It was selected to be the best score in the structure aliment getting by the
program. Ramachandran parameters and energy conditions was considered to model evaluation.
Molecular graphics images were produced using the UCSF Chimera package from the
Resource for Biocomputing, Visualization, and Informatics at the University of California, San
Francisco (supported by NIH P41 RR-01081), (http://www.cgl.ucsf.edu/chimera/).
The 3D model DHOD model obtained of T. gondii was used as molecular receptor in
docker software (Doker 1.1.0, http://www.molegro.com) to test in silico redoxal and A77-1726,
inhibitors in vivo of TgDHOD. The cavity for the ligand was limited to 10Ao. The best 5
possible locations of the ligand were defined by most favorable energetic model.

31
CHAPTER 3
RESULTS
The dihydroorotase dihydrogenase of Toxoplasma gondii (TgDHOD) cDNA is composed
of 1045 nucleotides which are translated in 592 amino acids. The predicted molecular mass of
full-length protein was 65 kDa. In previous work Sierra-Pagan and Zimmermann (2002) showed
that a truncated form of TgDHOD recombinant protein (TgDHOD-MIYS) was able to
complement a DHOD-deficient E. coli (pyrD-) cell strain (Sierra-Pagan et al., 2003).
Nevertheless, complementation was not observed in the full length protein TgDHOD-MAPL or
in another N-terminally truncated recombinant protein TgDHOD-MGFS. These three forms
started with three different methionines present in the TgDHOD amino sequences. The last
four letters indicate the DHOD N-terminal sequences of the each protein (Fig 4).
10 20 30 40 50 60
MAPLTMHFQG RFALLRLPIS SGKPLCRETR VRRSGTRPVS ADNLSHARCV LPKCHSFCPA
70 80 90 100 110 120
GGMQESPEAR VTLSRGTSRN FGTFLTALGN DVHWKSAFPG ALLRTQIRKL SVSLHPRPGS
130 140 150 160 170 180
AESSRPSAGL PPKDVDPEEI ERIVRERTTR ERKANRRLVF LVLLLGTGVY CYSALQDVSS
190 200 210 220 230 240
MIYSFYEPVT SVLFRYFSSG PLDPETAHGY TMELAKRGWL PVDYDREESA LNVDINGLKF
250 260 270 280 290 300
LSPIGLAAGF DKHAEAPAAL LRMGFSFLEV GSITPKPQPG NPKPRLFRLY EDRSVINRFG
310 320 330 340 350 360
FNSNGADYAQ TQLEAFSEAR LRDPFTAQGV LGVSLGKNKT SEDAVADLRE GVKKLGRFAD
370 380 390 400 410 420
FLVVNLSSPN TPGLRSLQSA SHLAAIIDGV QEELDALDRQ AQAASQKQRN ERRRHGGNPE
430 440 450 460 470 480
ETKAFYANQT GRRPLFFVKI APDLSMEEKE SIAKVALEKN LDGFVVSNTT IQRPETLKSP
490 500 510 520 530 540
AKSETGGLSG RALKHLSTAC VSDMYKLTQG KLAIIATGGV ESGRDALDKI EAGASLVELY
550 560 570 580 590
SSMVYIGPQV ARRVKNELYH ALNEKGYKDV AAAVGRKHKH VPEKKLQAPK FD

32
Figure 4. T.gondii DHOD amino acid sequences
Full length protein (TgDHOD-MAPL) indicated with red letters. The N-terminal sequences of
the TgDHOD-ALQD and TgDHOD-MGFS insoluble and inactive recombinant proteins are
indicated with blue and pink letters. The N-terminal sequences of the TgDHOD-VSSM,
TgDHOD-MIYS and TgDHOD-FYEP soluble and active recombinant proteins are indicated
with orange, purple and green letters. A predicted transmembrane region is underlined.
An important transmembrane region in the N-terminus was predicted by the
transmembrane prediction server DAS (Distributed annotation system). The hydrophobic profile
showed a hydrophobic alpha-helix between 157 to 170 residues of the DHOD sequences (Fig 4).
The N-terminally truncated protein TgDHOD-ALQD with just 7 residues more than the
complementing form TgDHOD-MIYS was insoluble and inactive.
In the current work, new clones were constructed with different N-terminal truncations
using a different bacterial expression vector (pET19b, Novagen) to produce the active
recombinant proteins TgDHOD-pET19bVSSM, TgDHOD-pET19bMIYS, and TgDHOD-
pET19bFYEP. All of these recombinant proteins were present in supernatants of detergent
extracts.
3.1 Protein purification
TgDHOD-VSSM was selected for the characterization. The first 178 residues of the N-
terminus of the full-length protein were eliminated, and the resulting recombinant protein was
soluble and active. The predicted molecular mass of the enzyme was 48.3 kDa including the N-
terminal His-tag (MGHHHHHHHHHHSSGAIDDDDKHM).

33
The predicted TgDHOD-VSSM recombinant protein was purified from bacterial extracts
by his-tag technology, using the 10 histidines tagged at the N- terminus. The largest amount of
protein was eluted in the first fraction with 300 mM imidazole. A band of the expected
molecular mass band was observed at ~ 48 kDa (Fig. 5) in a 12% denaturing gel stained with
Coomassie blue.
Figure 5. TgDHOD-VSSM recombinant protein purified protein by His-tag technology
SDS-PAGE stained with Coomassie Brilliant Blue R. All fractions from the purification were
loaded onto the gel; flow thru (FT), four different washes (W1 to W4), and three elutions (E1 to
E3). TgDHOD-VSSM was eluted in the first two fractions (E1 – E2) with 300 mM imidazole.
An expected band of 48 kDa was observed.
The yield of purified recombinant TgDHOD-VSSM was approximately 2.8 mg/L of
bacterial culture. The specific activity measured for the purified TgDHOD-VSSM using QD as
electron acceptor was 83.8 U/mg, where one unit is defined as the amount of enzyme required to
catalyze the reduction of 1 µmol of substrate in 1 min.

34
3.2 His-tag Western blot visualized with anti-His tag antibodies.
Recombinant purified TgDHOD protein was transferred from a denaturing gel to
nitrocellulose membrane. The membrane was incubated 1 h with anti-His-tag primary antibody
at a dilution of 1:100 dilution and then 1 h with IgG peroxidase conjugate as secondary antibody
at a 1:1500 dilution. Visualization was performed with 4-chloro -1- naphthol. The antibodies
were available to react with the purified recombinant protein TgDHOD-VSSM detecting a band
of ~ 48 kDa (Fig. 6 Panel A) and the same size as the band observed in a gel of purified protein
(Fig.6 Panel B).
Figure 6. Western blot and Gel of purified TgDHOD-VSSM.
Panel A, Western blot of purified TgDHOD-VSSM recombinant protein reacted with the anti
His-tag antibodies; Panel B, Denaturing gel electrophoresis of purified TgDHOD –VSSM
recombinant protein (0.5μg) purified from bacterial extract.

35
3.3 Immunoblots of T. gondii tachyzoite extracts
A sample with ~7.5 X 106 extracellular tachyzoites of strain RH T.gondii was prepared
for a Western blot. The cell lysis was performed by heating the tachyzoites at 90oC in loading
buffer for 10 min.
Two bands reacting with purified anti-DHODase polyclonal purified antibodies were
observed in the lysed tachyzoite extract (Fig. 7). One band corresponded to the expected size of
full-length T. gondii DHOD, 65 kDa. The second band was more intense, and appeared to co-
migrate with a truncated recombinant T. gondii DHOD, TgDHOD-VSSM, of molecular mass
48.3 kDa. The presence of a truncated protein might reflect in vivo processing.
Figure 7. Immunoblot of T. gondii tachyzoite extracts
Immunoblot of T. gondii tachyzoite extracts incubated with purified antibodies directed against
purified recombinant TgDHOD-ALQD.

36
3.4 Cross-reaction of TgDHOD-VSSM with antibodies directed against HsDHOD.
A Western blot performed with purified polyclonal antibodies raised against human
DHOD (HsDHOD) showed a weak cross-reaction with purified TgDHOD-VSSM recombinant
protein (Fig. 8).
Figure 8. TgDHOD protein cross-react with anti-HsDHOD antibodies
A nitrocellulose membrane loaded with 0.4μg of TgDHOD and 0.4 ug of Rattus rattus DHOD
was treated with purified anti-HsDHOD human (1:500). TgDHOD-VSSM was found to weakly
cross-react with the antibodies.
3.5 N-terminal sequencing
Toxoplasma gondii DHOD was affinity-purified using protein A-sepharose. Supernatants
from lysed, detergent treated tachyzoites extracts (1.8 x 108 parasites) were incubated with
polyclonal anti-TgDHOD antibodies, and then incubated with protein A-sepharose. After
centrifuging, the sample was placed in loading buffer and boiled for 10 min at 90oC. To achieve

37
better separation between the heavy chain subunit of rabbit IgG (50 kDa) and the low molecular
mass TgDHOD band (~ 48 kDa), large format, gradient gel electrophoresis was performed, and
protein was transferred to a PVDF membrane.
On the membrane, an intense band with a size of ~ 48 kDa was observed, with a weaker
band corresponding to the expected size of rabbit IgG heavy chain migrating above it (Fig 9).
The lower band was excised for N-terminal amino acid sequencing by Edman degradation.
Figure 9. Purification of a truncated fragment of TgDHOD from tachyzoite extracts
A PVDF membrane containing polyclonal anti-TgDHOD antibodies (AB), recombinant purified
protein (TgDHOD-VSSM), and TgDHOD affinity purified by protein A-sepharose from
extracellular tachyzoites (Complex AB-DHOD) was stained with Coomassie. The antibodies
and TgDHOD complex were previously separated by electrophoresis using a gradient gel. The
red arrow labeled (b) indicates the band thought to be TgDHOD, while the blue arrow labeled (a)
shows the slightly higher molecular weight, less intense band correspondig to the IgG havy
chain.

38
The piece of PDVF membrane with the lower band has been already sent. We are waiting for
sequences results.
3.6 Kinetic characterization
3.6.1 Kinetic constant
The Km and Vmax values of the TgDHOD were measured by the oxidation of DHO to
orotate using as electron QD and DCIP. Kinetic constant of DHO and QD were determined
(Table 1) by varying DHO concentration while QD was kept constant, or vice versa as was
described in section 2.8.
The activity assay was performed in presence of 0.1% Triton X-100 to to solubilize and
stabilize the protein and the quinones. A reduction of 50% in activity was observed when the
activity assay was performed in buffer reaction without Triton
Since DCIP can accept electrons in absence of ubiquinone, this background activity was
measured and was subtracted from activities in the presence of the ubiquinones (Table 1).
Table 1. Kinetic constants of purified recombinant TgDHOD-VSSM
Varied co-substrate
Fixed substrate
kcat (s-1
)
Km (μM)
DCIP L-DHO / QD 28.1 ± 0.7 38.72 ± 3.06
L-DHO QD / DCIP 81.8 ± 1.1 60.3 ± 0.002
QD L-DHO / DCIP 89.2 ± 1.5 28.9 ± 1.6

39
3.6.2 Electron acceptors
TgDHOD activity was measured in presence of different alternatives electron acceptors.
Percent activities were calculated based on the activity measured with QD (taken as 100%),
which showed highest activity.
The formation of orotate was measured to determine TgDHOD activity using
ferrocyanide and fumarete as electron acceptors. Appropriate isobestic points were used to avoid
overlap between product and electron acceptor (ferrycianide at 420 nm and fumarete at 280 nm).
As expected, low activity was observed with fumarate, which is not usually an electron
acceptor for Type II DHODs. With the ubiquinone acceptors, 35 – 40 % higher activities were
observed (Table 2).
Table 2. Use of electron acceptors by TgDHO-VSSM
Electron acceptor
Enzyme activity ( % )
QD 100.0
Q0 44.1 ± 5.4
Q6 34.1 ± 2.1
Q10 20.6 ± 2.7
PQ0 35.2 ± 1.3
Menadione 22.9 ± 0.6
Ferricyanide 11.8 ± 0.6
Fumarate 0.3 ± 0.1

40
Kinetic constants for different quinones were determined for TgDHOD-VSSM using the
DCIP reduction assay (Table 3). The highest activity was observed when QD was used as the
electron acceptor. Kinetic constants were not determined for menadione, PQ0, and Q10.
Table 3. Kinetic constant of electron acceptors
Electron acceptor Kcat (s-1
) Km (μM)
QD 89.2 ± 1.5 28.9 ± 1.6
Q0 32.3 ± 1.7 99.2 ± 7.5
Q6 20.6 ± 1.5 51.4 ± 7.0
3.6.3 Inhibitors
Compounds known to inhibit to human or Plasmodioum DHODs were tested to evaluate
their effect on TgDHOD-VSSM (Table 4). High activity reduction was observed in presence of
Redoxal and A77-1726. Also, DCL also affected TgDHOD activity but to a lesser extent.
Controls for the reactions were performed with appropriate dissolvent (buffer or DMSO) used
for each inhibitor (Table 4). Percentage of inhibition was calculated in based on the control
reaction (taken as 100%).
Toltrazuril and atovoquone did not remain soluble at high concentrations. The inhibition
experiments were performed with 0.5mM of toltrazuril or 0.1mM atovoquone and in both cases
TgDHOD activity was affected only by 20%.

41
Table 4. TgDHOD-VSSM activity in presence of putative inhibitors
Compound Concentration
(mM)
Enzyme activity ( % ) (a)
Control(b)
Control+ DMSO
- 100.0
- 100.0
Redoxal 1 17.1 ± 3.7
Toltrazuril 0.5 82.9 ± 2.9
Atovoquone 0.1 80.6 ± 3.2
A77-1726 1 10.7 ± 3.0
TTFA 1 106.7 ± 6.4
Brequinar 1 100.2 ± 2.2
DCL 1 29.8 ± 2.9
(a)DCIP reduction assay with 1mM DHO, 0.1QD and 0.1mM DCIP in presence of the inhibitor.
(b)Activity without inhibitor was set as 100%.
The IC50 was determined for the best inhibitors, Redoxal and A77-1726. The calculated IC50
values were 91.2 µM ± 2.2 for A77-1726, and 253 µM ± 13.3 for Redoxal.
Figure 10. Inhibition plots

42
Inhibition plots of purified TgDHOD-VSSM recombinant protein activity in presence of A77-
1726 (left) and Redoxal (right).
IC50 values obtained for redoxal and A77-1726 were compared with the IC50 published in the
literature for Plasmodium and human DHOD. Difference in sensitivity to inhibitors was
observed for the three DHODs.
Table 5. IC50 values for DHODs
Compound
IC50
(μM)
TgDHOD PgDHOD HsDHOD
A77-1726 91.2 ± 2.2 190 ± 10 (a)
1 ± 0.1 (c)
0.26 ± 0.1 (a)
Redoxal 253.0 ± 13.3 71 ± 5 (b)
0.013 ± 0.0005 (b)
DCL - 220 ± 30 (b)
0.067 ± 0.0064 (d)
Brequinar - - 0.010 ± 0.0009 (d)
(a)
Heikkila et al., 2007 (b)
Baldwin et al., 2002 (c)
Knecht et al., 1998 (d)
Knecht et al., 2000
3.7 Preliminary immunolocalization of dihydroorotate dehydrogenase in T. gondii
tachyzoites
Initial immunolocalization experiments with anti-DHOD antibodies suggested a possible
co-localization of the DHOD with DAPI-stained apicoplast (data not showed). To further clarify
these observations we undertook experiments with parasites containing labeled apicoplast or
mitochondria. In intracellular or extracellular parasites treated with anti-DHOD antibodies,

43
fluorescence (red) was observed that appeared to co-localize with apicoplasts visualized by
yellow fluorescent protein-tagged acyl carrier protein (yellow, YFP-ACP). Apicoplast shape
observed in yellow was consistent with the red shape detected with the anti-TgDHOD antibodies
(Fig. 11).
Figure 11. Immunolocalization of TgDHOD in tachyzoites
Immunolocalization of TgDHOD in tachyzoites containing yellow fluorescent protein tagged acyl-carrier
protein (ACP-YFP). Panel A, In extracellular tachyzoites, fluorescence of the anti-DHOD antibodies (red)
showed apparent colocalization with the apicoplast (yellow). Panel B, In intracellular tachyzoites, a
similar apparent colocalization was observed. The white bar indicates 5 µm.

44
In some of the parasites additional fluorescence was observed that did not appear to co-
localize with ACP. Experiments with MitoTracker Red CMXRos showed possible
colocalization of mitochondria and anti-DHOD antibodies (green) in ~ 10% of tachyzoites (Fig.
12).
Figure 12. Partial localization of TgDHOD to mitochondria
Partial localization of TgDHOD to mitochondria. Tachyzoites were incubated with MitoTracker
(red) and anti-DHOD antibodies (green). Although green fluorescence is observed
predominantly in circular structures, partial mitochondrial localization was detected in some
tachyzoites (white arrows). The white bar indicates 5 µm.

45
Inmunoflorescense with Golgi-labeled intracellular tachyzoites were performed to check
whether the enzyme colocalized with this parasite organelle. GRASP-mRFP transfected
parasites and the polyclonal antibodies were used. No apparent colocalization was observed (Fig
13).
Figure 13. Intracellular Golgi-tagged tachyzoites
Intracellular Golgi-tagged tachyzoites labeled with GRASP-mRFP (red), were incubated with
anti-DHOD antibodies (green). Overlap was not observed (The Golgi-labeled parasite strain was
the gift of Dr. David S. Roos, University of Pennsylvania, Philadelphia, PA, USA)
Polyclonal antibodies anti-TgDHOD were purified by affinity column and then new
immunolocalization experiments were performed. As before, the unpurified antibodies once
again appeared to show apicoplast colocalization in the RH (Fig 14) and the apicoplast labeled
(ACP-YFP) (Fig 15) strains. In contrast, the purified antibodies did not produce a specific
localization pattern (Fig 16) even though they recognized the two bands in Western blots of
parasite extracts.
Anti-DHOD GRASP-mRFP Overlap
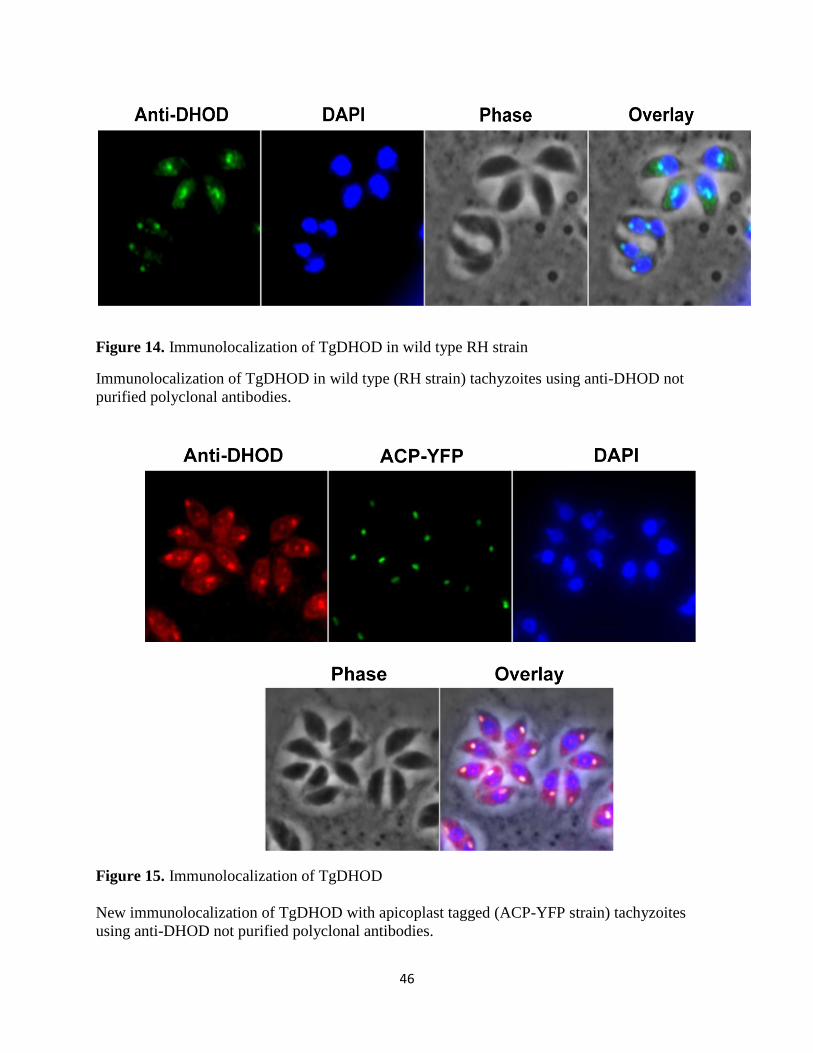
46
Figure 14. Immunolocalization of TgDHOD in wild type RH strain
Immunolocalization of TgDHOD in wild type (RH strain) tachyzoites using anti-DHOD not
purified polyclonal antibodies.
Figure 15. Immunolocalization of TgDHOD
New immunolocalization of TgDHOD with apicoplast tagged (ACP-YFP strain) tachyzoites
using anti-DHOD not purified polyclonal antibodies.
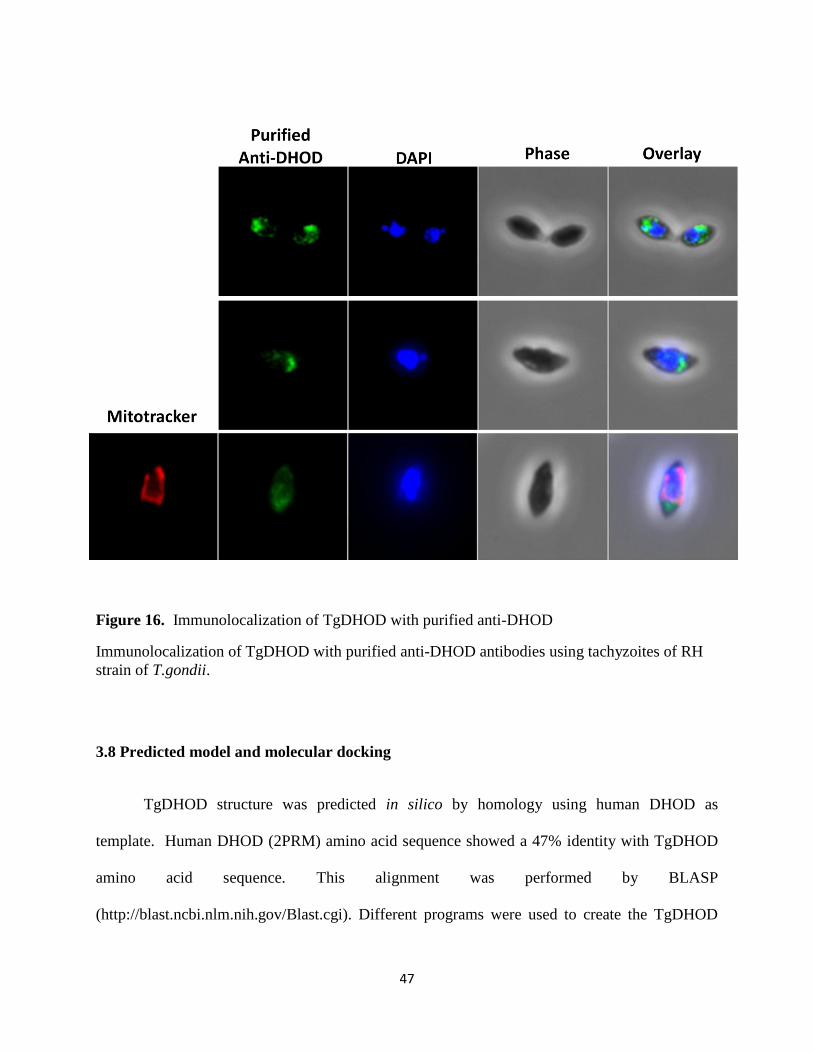
47
Figure 16. Immunolocalization of TgDHOD with purified anti-DHOD
Immunolocalization of TgDHOD with purified anti-DHOD antibodies using tachyzoites of RH
strain of T.gondii.
3.8 Predicted model and molecular docking
TgDHOD structure was predicted in silico by homology using human DHOD as
template. Human DHOD (2PRM) amino acid sequence showed a 47% identity with TgDHOD
amino acid sequence. This alignment was performed by BLASP
(http://blast.ncbi.nlm.nih.gov/Blast.cgi). Different programs were used to create the TgDHOD

48
model, but it was not possible to obtain the complete model however using 3D-JIGSAW
modeling program a model starting in the Y170 amino acid was achieved. This model presented
90% of the amino acid residues that fall within the acceptable region of the Ramachandran plot.
The non predicted area of the N-terminal secondary structure was determined by PSIPRED
(http://bioinf.cs.ucl.ac.uk/psipred/).
PSIPRED showed that N-terminal belongs to an alpha-helix apparently related with the
electron acceptor binding site (fig. 17).
Figure 17. TgDHOD structural model
A. TgDHOD predicted structure (Purple), amino acids involved in the FMN binding (Green) B.
Amino acids involved in the Orotate binding (light pink) C. Serine amino acids conserved in all
type II DHODs (turquoise).

49
Active site of the protein is conserved in all the DHODases type II. Substrate and cofactor
region is shown in the figure 18.
Figure 18. TgDHOD model for alpha helix A
TgDHOD model for alpha helix A. TgDHOD predicted structure alpha helix (Purple) and beta
sheets (Yellow), amino acids involved in the FMN binding (Green), amino acids involved in the
Orotate binding (light pink), Serine amino acids conserved in all type II DHODs (Blue) and
Alpha helix A (Dark pink).

50
Figure 19. TgDHOD molecular docking with Redoxal A
TgDHOD molecular docking with Redoxal A. Electrostatic surface model of TgDHOD showing the
expected cavity of the electron binding acceptor (green), Negative charge amino acids (blue) and positive
chare amino acids (red). B. TgDHOD showed as backbone ribbons. Redoxal inhibitor (stick) is located in
the predicted electron acceptor cavity. C. Surface of the cavity were the inhibitor might be located (green
mesh). D. Electrostatic interaction polar, non polar and neutral, between the inhibitor and the nearby
amino acids. E. Hydrogen bonding formation between the inhibitor (yellow stick) and the residues
localized 5Å. F. Hydrophobic interactions between the residues located at 5Å of the inhibitor overlap in
the surface of the protein.

51
CHAPTER 4
DISCUSSION
Pyrimidine biosynthesis has been proposed as a drug target for Apicomplexan parasites
(Gero and O’Sullivan, 1990; McFadden and Boothroyd, 2001; Fox and Bzik, 2002; Baldwin et
al., 2005; Hyde, 2007). Plasmodium depends completely on de novo pyrimidine biosynthesis for
pyrimidine nucleotides because it lacks salvage enzymes. Although T. gondii is able to salvage
uracil using uracil phosphoribosyltransferase (Pfefferkorn, 1978; Iltzsch, 1993), this enzyme is
nonessential (Donald & Roos, 1995). In contrast, T. gondii parasites lacking the first enzyme of
the pyrimidine biosynthetic pathway are avirulent in mice and are unable to replicate in cell
culture, suggesting that enzymes of the pathway might be good targets (Bzik & Fox, 2002).
DHOD, the fourth enzyme in the pathway is being intensively studied as a potential target in
Plasmodium (Gujjar et al., 2009, Davies et al., 2009, Patel et al., 2008). The production of an
active, purified, recombinant TgDHOD has allowed us to begin the kinetic characterization of
the enzyme, a fundamental step for defining the effects of potential inhibitors.
A summary of the kinetic parameters determined for the TgDHOD-VSSM recombinant
protein follows. TgDHOD exhibited the lowest Kms for QD and Q6 (~ 30 - 50 µM), and a higher
Km for Q0 (~ 100 µM) (Table 1). Baldwin and co-workers observe similar trends for
recombinant DHOD from Plasmodium falciparum (PfDHOD), which exhibits lower Kms for Q4,
Q6 and QD (~ 15 – 20 µM), and higher Kms for quinones lacking isoprenoid chains, such as Q0 or
menadione (~ 55 - 115 µM). The Kms for ubiquinones synthesized by P. falciparum, Q7, Q8 and
Q9 (de Macedo et al., 2002; Tonhosolo et al., 2005) have not been determined but are probably in

52
the former range. Q10 is the only ubiquinone present in humans and most higher animals (Ellis et
al., 1994). The ubiquinones present in T. gondii have not yet been identified. HsDHOD exhibits
a Km for QD of 14 µM, similar to the values measured for the parasite enzymes (Knecht et al.,
1996). The Km for dihydroorotate for TgDHOD with QD as the co-substrate was 60 µM, in the
same range as that reported for PfDHOD, 30 – 90 µM (Baldwin et al., 2005; Heikkila et al.,
2007). The Km for human DHOD for dihydroorotate, 9.5 µM, is lower (Ullrich et al., 2001).
The kcat measured for TgDHOD, 90 s-1
, was similar to that of HsDHOD, 107 s-1
(Ullrich
et al., 2001), and 6 to 11-fold higher (Table 1) than those measured for recombinant PfDHOD
proteins (Baldwin et al., 2005; Heikkila et al., 2007). The lower activity for the Plasmodium
enzyme can be accounted for, in part, by differences in assay conditions, including a lower
temperature. In addition, the position of the truncation of the recombinant Plasmodium enzyme
appears to affect activity (Heikkila et al., 2007). This was also found to be the case for the
activity of the T. gondii DHOD; a recombinant protein starting at A174 was inactive (Sierra
Pagan and Zimmermann, 2002), while those starting at V178, M181, and F185 were all active.
We tested the effect of compounds known to inhibit HsDHOD on our purified
recombinant enzyme. TgDHOD was unaffected by 1 mM brequinar, which inhibits HsDHOD
with IC50 = 0.010 µM (Knecht et al., 2000) (Table 3). The related compound, DCL (IC50 = 0.067
µM for HsDHOD (Knecht et al., 2000), IC50 = 220 µM for PfDHOD (Baldwin et al., 2002))
decreased the activity of TgDHOD to 30% at 1 mM (Table 3). Another related compound,
Redoxal (IC50 = 0.013 µM for HsDHOD (Baldwin et al., 2002), IC50 = 71 µM for PfDHOD
(Baldwin et al., 2002)), inhibited TgDHOD with an IC50 = 253 µM (Fig. 2). A77-1726, the
active metabolite of leflunomide, a drug used to treat rheumatoid arthritis, inhibits HsDHOD
with IC50 = 0.26 µM (Heikkila et al., 2007). The parasite enzymes exhibit roughly 3 orders of

53
magnitude less affinity for A77-1726, with IC50 = 190 µM for PfDHOD (Heikkila et al., 2007))
and IC50 = 91 µM for TgDHOD (Fig. 2). Other compounds were also tested. TTFA, shown to
bind to ubiquinone sites in the structure of mitochondrial Complex II (Sun et al., 2005), had no
effect on TgDHOD activity. Atovaquone, an inhibitor that binds to the ubiquinol oxidation
pocket of Complex III (IC50 = 0.04 µM for Saccharomyces cerevisiae complex III, IC50 = 0.4
µM for human complex III (Kessl et al., 2003), IC50 = 14.5 µM for HsDHOD), and a drug used
in the treatment of toxoplasmosis, malaria, and pneumocystis pneumonia, had a relatively small
effect on the activity of TgDHOD at a concentration 0.1 mM. Toltrazuril, an anticoccidial drug
(Haberkorn, 1996), and an inhibitor of rat DHOD, also had a small effect on TgDHOD activity at
0.5 mM (Table 3). It is evident from many studies (Baldwin et al., 2002; Baldwin et al., 2005;
Heikkila et al., 2007; Phillips et al., 2008; Gujjar et al., 2009; Davies et al., 2009) that there are
significant differences between the binding sites of PfDHOD and HsDHOD that can be exploited
for producing Plasmodium-specific inhibitors. Our limited experimental data on DHOD
inhibitors suggests that this will also be the case for TgDHOD. We built a homology model of
the TgDHOD amino acid sequence using the 3D-JIGSAW server (Bate et al., 2001) in the
automatic modeling mode, using as templates each of the available type 2 DHOD structures.
Regardless of the template used, the resulting TgDHOD model could only be generated after
residue Y170, leaving unmodeled the first α-helix of the small domain, the binding site of many
inhibitors and the predicted binding site of the ubiquinone cofactor. This failure to generate a
structural prediction for the complete TgDHOD small domain indicates a likely structural
dissimilarity between the TgDHOD and other type 2 DHODs, which could be exploited to
design T. gondii-specific inhibitors, and which may have implications for the binding of electron
acceptors in vivo.

54
To date, all known type 2 eukaryotic DHODs have been found to be associated with
mitochondria. In these preliminary localization studies we observed the TgDHOD in T. gondii
mitochondria, but we also observed a second, unexpected, apparent localization to the parasite’s
apicoplast. Organellar targeting signals generally are found in a protein’s N-terminus. The
bioinformatic analyses of the TgDHOD N-terminus showed that targeting predictions favor
mitochondrial localization, however, unlike the PfDHOD and HsDHOD, the predicted
probabilities for plastid localization were not zero. Different cleavage sites were predicted, but
there was no consensus among the different programs. Predictions for T. gondii proteins are
complicated by the paucity of T. gondii data incorporated into the analysis programs.
The observation of a truncated T. gondii DHODase antibody-reacting band on
immunoblots of tachyzoite extracts may reflect in vivo processing, although proteolysis during
extraction cannot be excluded. Denaturing gel electrophoresis of P. falciparum DHOD purified
from parasite extracts reveals a band with a molecular mass of 56 kDa (Krungkrai, 1995), while
the predicted molecular mass from the protein sequence is 65.6 kDa (XP_966023.1). The N-
terminus of the mammalian DHOD contains a typical bipartite signal comprised of cationic
sequences and a hydrophobic region that direct localization to the intermembrane space and
insertion into the inner mitochondrial membrane, however, no cleavage of mammalian DHOD
sequences are observed in vivo or in vitro (Rawls et al., 2000).
Although well-defined bipartite N-terminal sequences direct proteins to the lumen of the
apicoplast in T. gondii (Waller & McFadden, 2005), only a handful of membrane-associated
apicoplast proteins have been studied in T. gondii and P. falciparum, and their trafficking
appears to be governed by diverse types of signaling (Parsons et al., 2009; Lim et al., 2009).
Some, like the proteins of the lumen, appear to contain bipartite signal sequences and undergo N-

55
terminal processing to remove a transit peptide, presumably using import mechanisms similar to
those employed by proteins of the apicoplast lumen (Parsons et al., 2009). Other apicoplast
membrane proteins do not contain bipartite signal sequences. In these proteins, a lack of
processing is observed in some cases, while cleavage of C- and N-terminal sequences occurs in
other cases (Parsons et al., 2009).
Our preliminary results point to dual-targeting of TgDHOD. Several proteins from T.
gondii are targeted to both apicoplast and mitochondrion (Sheiner and Soldati-Favre, 2008).
Two mechanisms for dual targeting have been proposed. In the first, an ambiguous signal
peptide is thought to interact inefficiently with the signal recognition particle responsible for
routing proteins via the endoplasmic reticulum to the apicoplast, as appears to be the case for an
isozyme of superoxide dismutase (SOD2) that is present in both organelles (Pino et al., 2007,
Brydges & Carruthers, 2003). A second mechanism is the production of different proteins from
the same gene by initiating translation at different methionines. An example is the recently
discovered membrane-associated isozyme of pyruvate kinase (PK2) that is localized to both to
apicoplast and mitochondrion (Saito et al., 2008). Three methionines are present in the N-
terminus of TgDHOD at residues 1, 6, and 63. Bioinformatic analyses of a sequence starting at
M63 shows an increased predicted probability of plastid localization (0.560 compared to 0.385
for the full length sequence) and decreased predicted probability of mitochondrial localization
(0.165 compared to 0.593 for the full length sequence) (Table 4). As stated above, however,
localization predictions for T. gondii should be viewed with caution.
If T. gondii DHOD indeed localizes to the apicoplast as well as to the mitochondrion, as
suggested by our initial studies, then an appropriate electron acceptor(s) should be present in that
organelle. The most likely electron acceptor in vivo would be a quinone. In higher eukaryotic

56
cells, ubiquinone and its reduced form, ubiquinol, are not limited to the mitochondrial inner
membrane, and are found in membranes of the endoplasmic reticulum, the Golgi apparatus,
lysosomes, peroxisomes, and the plasma membrane and (Ernster and Dallner. 1995). Moreover,
ubiquinone is present in membranes of Entamoeba histolytica, Giardia lamblia, and
Trichomonas foetus, parasites that do not have mitochondria (Ellis et al., 1994). T. gondii is
probably capable of quinone synthesis, since enzymes of the DOXP/MEP pathway producing the
precursor isoprenoids are present in its genome (Clastre et al., 2007). In Plasmodium, isoprenoid
synthesis occurs in the apicoplast (Wiesner and Jomaa, 2007), and this is also thought to be the
case with T. gondii. An enzyme catalyzing one of the subsequent steps in synthesis of quinone,
farnesyl-diphosphate synthase, has been localized to T. gondii mitochondria (Ling et al., 2007).
A close physical association between the apicoplast and mitochondrion is observed in G1 and
during apicoplast elongation (Nishi et al., 2008). Taking the aforementioned points into
consideration, it seems reasonable that quinones would be present in the membranes of the
apicoplast of T. gondii. Several metabolic pathways requiring a source of reducing equivalents
are present in the apicoplast, and a redox system comprised of ferredoxin and ferredoxin
reductase is also present (Vollmer et al., 2001). We speculate that if TgDHOD is indeed
associated with this organelle, it would be likely to play a role in redox buffering. The matrix of
the apicoplast of P. falciparum provides a highly redox-buffered reducing environment as
demonstrated by using the thiol of the acyl carrier protein as a redox indicator (Gallagher and
Prigge, 2009). To further explore the possible unusual localization of the Tg DHOD, other
approaches are needed such as immunolocalization with monoclonal instead of polyclonal
antibodies, or visualization of T. gondii transfected with epitope-tagged TgDHOD.

57
In mammals and other organisms with family 2 DHODases, electrons from
dihydroorotate oxidation are transferred to ubiquinone, thus contributing to oxidative
phosphorylation. Questions remain about the importance of oxidative phosphorylation in
different stages of T. gondii. The mitochondrial electron transport chain in T. gondii is
composed of homologs of respiratory complexes II, III, and IV (Moreno et al., 1998), while
complex I is replaced by a rotenone-insensitive type-II NADH dehydrogenase (Lin et al., 2008).
Experiments with mitochondrial dyes, such as Mitotracker, CMXRos, and DiOC6(3),
demonstrate mitochondrial membrane potential in both extra- and intracellular parasites (Melo et
al., 2000, Lin et al., 2009). In extracellular tachyzoites, under conditions when the electron
transport chain is inhibited by KCN, or in absence of oxygen, tachyzoites retain 60% -70%
motility, suggesting that glycolysis is the major source of ATP (Pomel et al., 2008).
Interestingly, early work by Asai and co-workers (1983) shows that tachyzoite extracts harvested
from peritoneal cavities of mice still retain 20% of DHOD activity in the presence of 1 mM KCN
(enough to inhibit complex IV (Vercesi et al., 1998)), while 5 mM KCN decreases the activity to
6%.
In mammals, the TCA cycle generates reducing equivalents for electron transport using
acetyl-CoA produced from pyruvate by the mitochondrial enzyme pyruvate dehydrogenase. In
T. gondii, the enzymes of the TCA cycle are located in the mitochondrion, however, the pyruvate
dehydrogenase is located in the apicoplast (Fleige et al., 2007). Seeber and co-workers propose
that T. gondii enzymes predicted to metabolize propionyl-CoA derived from degradation of
branched chain amino acids could provide succinate and oxaloacetate for the TCA cycle (Seeber
et al., 2008). Disruption of the T. gondii TCA cycle by knocking out succinyl-CoA synthetase
does not appear to cause a decrease in the membrane potential detected by MitoTracker (Fleige

58
et al., 2008), suggesting that other sources of reducing equivalents are important for the
reduction of the electron transport chain. In T. gondii, as is the case in other organisms,
respiratory chain-linked dehydrogenases, for example, glycerol-3-phosphate dehydrogenase, and
dihydroorotate dehydrogenase are also a source of electrons. Differences in the abilities of these
two dehydrogenases to reduce the electron transport chain have been observed between
extracellular and intracellular tachyzoites. According to work by Vercesi et al. (1998) with
digitonin-permeabilized extracellular tachyzoites, oxygen consumption is stimulated by addition
of ADP when malate or succinate are added as substrates, however, no stimulation was observed
in the presence of dihydroorotate or glycerol-3-phosphate. In contrast, both dihydroorotate or
glycerol-3-phosphate (as well as malate or succinate) are able to generate a mitochondrial inner
membrane potential in digitonin-permeabilized intracellular tachyzoites as shown by the uptake
of the lipophilic, cationic, fluorescent probe, DiOC6(3) (Lin et al., 2009).
The amount of reducing power provided by the oxidation of dihydroorotate depends on
the metabolic flux through the pyrimidine biosynthetic pathway. The pathway is active in
extracellular parasites isolated from mouse peritoneal cavity (Asai et al., 1983, Robles Lopez et
al., 2006). Since extracellular parasites are not dividing, the requirement for pyrimidines would
presumably be lower than in intracellular parasites. In mammalian cells, the dependence of cell
growth and division on the activity of de novo pyrimidine biosynthesis is well documented, and
the enzymes are controlled at the transcriptional and post-transcriptional levels (Evans & Guy,
2004). The rate-limiting step of the mammalian pathway, is catalyzed by the carbamoyl-
phosphate synthetase (CPSII) activity of the multifunctional enzyme, CAD, that catalyzes the
first three steps in the pathway in mammals (Evans & Guy, 2004). In resting cells, the CPSII
activity is feedback inhibited by UTP and activated by phosphoribosyl 5’-pyrophosphate (PRPP)

59
(Evans & Guy, 2004). When cells start dividing, CPSII undergoes phosphorylation by MAP
kinase, resulting in the loss of feedback inhibition by UTP, and leading to an eight-fold increase
in flux through the pathway in exponentially growing cells compared to resting cells (Sigoillot et
al., 2002). In T. gondii, less is known about the regulation of the pyrimidine biosynthetic
pathway. Asai and co-workers (1983) found that the activity of TgCPSII partially purified from
parasite extracts was inhibited by UTP, but no activation was observed in the presence of PRPP.
Recent studies with TgCPSII minigenes show that mutations or deletions in the proposed
regulatory regions of the CPSII C-terminus decrease, and in some cases eliminate, the ability of
the minigene to complement CPSII knockout parasites (Fox et al., 2009).
Whether or not the pyrimidine biosynthetic pathway is operational in the slowly-dividing
bradyzoites has not yet been demonstrated. Several observations are relevant to this discussion.
Inhibitors of mitochondrial respiration induce differentiation from tachyzoites to bradyzoites in
cell cultures in the presence of uridine (Bohne et al., 1994; Tomavo and Boothroyd, 1995).
Nevertheless, bradyzoites probably have low levels of mitochondrial activity because
atovoquone, an inhibitor of respiratory complex III, is active in vitro against bradyzoites
(Ferguson et al., 1994). Furthermore, a survey of ToxoDB reveals the presence of expressed
sequence tags (ESTs) in bradyzoites for complexes II (TGME49_015590, TGME49_015280), III
(TGME49_046540, TGME49_088750), IV (TGME49_055060, TGME49_026590,
TGME49_110470), and cytochorome c (TGME49_019750, TGME49_029420). ESTs of the
pyrimidine biosynthetic enzymes catalyzing steps 1 (TGME49_015260), 3 (TGME49_093610),
4 (TgDHOD, TGME49_010790), and 6 (TGME49_59690), and ESTs of uracil phosphoribosyl
transferase, the principal pyrimidine salvage enzyme, have been found in bradyzoites. In
general, the number of ESTs of bradyzoite origin is much lower than the number from

60
tachyzoites. An exception to this trend is apparent for TgDHOD (7 ESTs from bradyzoites, 8
ESTs from tachyzoites), although whether this reflects the relative expression levels of the
enzyme in the two stages remains to be seen.
The availability of an active, recombinant TgDHOD will contribute to clarifying the role
of pyrimidine synthesis in different stages of T. gondii, and makes possible the characterization
of potential inhibitors, opening the possibility for future crystallization and structure
determination for the enzyme.

61
SUMMARY
The pyrimidine biosynthesis pathway in the protozoan pathogen Toxoplasma gondii is essential
for parasite growth during infection. To investigate the properties of dihydroorotate
dehydrogenase (TgDHOD), the fourth enzyme in the T. gondii pyrimidine pathway, we
expressed and purified recombinant TgDHOD. TgDHOD exhibited a specific activity of 83.8
U/mg, a kcat of 89.2 sec-1
± 1.5, a Km = 60.3 ± 0.002 µM for L-dihydroorotate, and a Km = 28.9
µM ± 1.8 for decylubiquinone (QD). Quinones lacking or having short isoprenoid side chains
yielded lower kcats and higher Kms than QD. As expected, fumarate was a poor electron acceptor
for this family 2 DHOD. The IC50s determined for Redoxal and A77-1726 were 253.3 μM ±
13.3 and 91.2 μM ± 2.2, respectively. The enzyme was not significantly affected by brequinar or
TTFA, known inhibitors of human DHOD, or by atovaquone. TgDHOD exhibits a 157-residue
N-terminal extension, consistent with potential organellar targeting, but bioinformatic analysis
failed to reveal a consensus subcellular destination. Preliminary immunolocalization studies of
TgDHOD performed in intra- and extracellular parasites treated with the polyclonal antibodies
raised against purified recombinant TgDHOD exhibited fluorescence that appeared to colocalize
with the apicoplast in tachyzoites expressing yellow fluorescent protein-tagged acyl-carrier
protein. Anti-DHOD fluorescence was observed in ~10% of mitochondria stained with
Mitotracker Red in extracellular parasites. These findings suggest that TgDHOD is associated
with both the apicoplast and mitochondrion, making it a member of a growing list of metabolic
enzymes that are dual targeted in T. gondii.

62
REFERENCES
Asai T, O'Sullivan WJ, Kobayashi M, Gero AM, Yokogawa M, Tatibana M. Enzymes of the de
novo pyrimidine biosynthetic pathway in Toxoplasma gondii. Mol Biochem Parasitol.
1983;7:89-100.
Baumgartner R, Walloschek M, Kralik M, Gotschlich A, Tasler S, Mies J, Leban J. Dual
binding mode of a novel series of DHODH inhibitors. J Med Chem. 2006;49:1239-47.
PMID 16480261
Baldwin J, Farajallah AM, Malmquist NA, Rathod PK, Phillips MA. Malarial dihydroorotate
dehydrogenase. Substrate and inhibitor specificity. J Biol Chem. 2002;277:41827-34.
PMID: 12189151
Baldwin J, Michnoff CH, Malmquist NA, White J, Roth MG, Rathod PK, Phillips MA. High-
throughput screening for potent and selective inhibitors of Plasmodium falciparum
dihydroorotate dehydrogenase. J Biol Chem. 2005;280:21847-53. PMID: 15795226
Bernacchia A, Biondi A, Genova ML, Lenaz G, Falasca A. The various sources of
mitochondrial oxygen radicals: a minireview. Toxicol Mech Methods. 2004;14:25-30.
PMID: 20021118
Bobrowski M, Liwo A, Hirao K. Theoretical study of the energetics of the reactions of triplet
dioxygen with hydroquinone, semiquinone, and their protonated forms: relation to the
mechanism of superoxide generation in the respiratory chain. J Phys Chem B.
2007;111:3543-9. PMID: 17388501

63
Bohne W, Heesemann J, Gross U. Reduced replication of Toxoplasma gondii is necessary for
induction of bradyzoite-specific antigens: a possible role for nitric oxide in triggering stage
conversion. Infect Immun. 1994;62:1761-7. PMID: 8168938
Brydges SD, Carruthers VB. Mutation of an unusual mitochondrial targeting sequence of
SODB2 produces multiple targeting fates in Toxoplasma gondii. J Cell Sci.
2003;116:4675-85.
Claros MG, Vincens P. Computational method to predict mitochondrially imported proteins and
their targeting sequences. Eur J Biochem. 1996;241:779-86. PMID: 8944766
Clastre M, Goubard A, Prel A, Mincheva Z, Viaud-Massuart MC, Bout D, Rideau M, Velge-
Roussel F, Laurent F. The methylerythritol phosphate pathway for isoprenoid biosynthesis
in coccidia: presence and sensitivity to fosmidomycin. Exp Parasitol. 2007;116:375-84.
PMID: 17399705
Contreras-Moreira,B., Bates,P.A. (2002) Domain Fishing: a first step in protein comparative
modelling. Bioinformatics 18: 1141-1142.
Cserzö M, Wallin E, Simon I, von Heijne G, Elofsson A. Prediction of transmembrane alpha-
helices in prokaryotic membrane proteins: the dense alignment surface method. Protein
Eng. 1997;10:673-6. PMID: 9278280
Davies M, Heikkilä T, McConkey GA, Fishwick CW, Parsons MR, Johnson AP. Structure-
based design, synthesis, and characterization of inhibitors of human and Plasmodium
falciparum dihydroorotate dehydrogenases. J Med Chem. 2009;52:2683-93. PMID:
19351152

64
de Macedo CS, Uhrig ML, Kimura EA, Katzin AM. Characterization of the isoprenoid chain of
coenzyme Q in Plasmodium falciparum. FEMS Microbiol Lett. 2002;207:13-20. PMID:
11886744
Deng X, Gujjar R, El Mazouni F, Kaminsky W, Malmquist NA, Goldsmith EJ, Rathod PK,
Phillips MA. Structural plasticity of malaria dihydroorotate dehydrogenase allows
selective binding of diverse chemical scaffolds. J Biol Chem. 2009;284:26999-7009.
PMID: 19640844
Derouin F, Mazeron MC, Garin YJ. Comparative study of tissue culture and mouse inoculation
methods for demonstration of Toxoplasma gondii. J Clin Microbiol. 1987;25:1597-600
Donald RG, Roos DS. Insertional mutagenesis and marker rescue in a protozoan parasite:
cloning of the uracil phosphoribosyltransferase locus from Toxoplasma gondii. Proc Natl
Acad Sci U S A. 1995;92:5749-53. PMID: 7777580
Ellis JE, Setchell KD, Kaneshiro ES. Detection of ubiquinone in parasitic and free-living
protozoa, including species devoid of mitochondria. Mol Biochem Parasitol. 1994;65:213-
24. PMID: 7969263
Emanuelsson O, Brunak S, von Heijne G, Nielsen H. Locating proteins in the cell using TargetP,
SignalP and related tools. Nat Protoc. 2007:2:953-71. PMID: 17446895
Emanuelsson O, Nielsen H, von Heijne G. ChloroP, a neural network-based method for
predicting chloroplast transit peptides and their cleavage sites. Protein Sci. 1999;8:978-84.
PMID: 10338008

65
Ernster L, Dallner G. Biochemical, physiological and medical aspects of ubiquinone function.
Biochim Biophys Acta. 1995;1271:195-204. PMID: 7599208
Evans DR, Guy HI. Mammalian pyrimidine biosynthesis: fresh insights into an ancient pathway.
J Biol Chem. 2004;279:33035-8. PMID: 15096496
Ferguson DJ, Huskinson-Mark J, Araujo FG, Remington JS. An ultrastructural study of the
effect of treatment with atovaquone in brains of mice chronically infected with the ME49
strain of Toxoplasma gondii. Int J Exp Pathol. 1994;75:111-6. PMID: 8199003
Fleige T, Fischer K, Ferguson DJ, Gross U, Bohne W. Carbohydrate metabolism in the
Toxoplasma gondii apicoplast: localization of three glycolytic isoenzymes, the single
pyruvate dehydrogenase complex, and a plastid phosphate translocator. Eukaryot Cell.
2007;6:984-96. PMID: 17449654
Fleige T, Pfaff N, Gross U, Bohne W. Localisation of gluconeogenesis and tricarboxylic acid
(TCA)-cycle enzymes and first functional analysis of the TCA cycle in Toxoplasma gondii.
Int J Parasitol. 2008;38:1121-32. PMID: 18336823
Fox BA, Bzik DJ. De novo pyrimidine biosynthesis is required for virulence of Toxoplasma
gondii. Nature. 2002;415:926-9.
Fox BA, Ristuccia JG, Bzik DJ. Genetic identification of essential indels and domains in
carbamoyl phosphate synthetase II of Toxoplasma gondii. Int J Parasitol. 2009;39:533-9.
PMID: 18992249

66
Gallagher JR, Prigge ST.Plasmodium falciparum acyl carrier protein crystal structures in
disulfide-linked and reduced states and their prevalence during blood stage growth.
Proteins. 2009 Aug 17. (Epub ahead of print) PMID: 19768685
Gero AM, O'Sullivan WJ. Blood Cells. Purines and pyrimidines in malarial parasites.
1990;16:467-84. PMID: 2257323
Grigg ME, Boothroyd JC. Rapid identification of virulent type I strains of the protozoan
pathogen Toxoplasma gondii by PCR-restriction fragment length polymorphism analysis at
the B1 gene. J Clin Microbiol 2001;39:398-400.
Gujjar R, Marwaha A, El Mazouni F, White J, White KL, Creason S, Shackleford DM, Baldwin
J, Charman WN, Buckner FS, Charman S, Rathod PK, Phillips MA. Identification of a
metabolically stable triazolopyrimidine-based dihydroorotate dehydrogenase inhibitor with
antimalarial activity in mice. J Med Chem. 2009;52:1864-72. PMID: 19296651
Haberkorn A. Chemotherapy of human and animal coccidioses: state and perspectives. Parasitol
Res. 1996;82:193-9. PMID: 8801548
Hansen M, Le Nours J, Johansson E, Antal T, Ullrich A, Löffler M, Larsen S. Inhibitor binding
in a class 2 dihydroorotate dehydrogenase causes variations in the membrane-associated N-
terminal domain. Protein Sci. 2004;13:1031-42. PMID: 15044733
Haverkos HW. Assessment of therapy for toxoplasma encephalitis. The TE Study Group. Am J
Med. 1987;82:907-14. PMID: 3578360

67
Heikkilä T, Ramsey C, Davies M, Galtier C, Stead AM, Johnson AP, Fishwick CW, Boa AN,
McConkey GA. Design and synthesis of potent inhibitors of the malaria parasite
dihydroorotate dehydrogenase. J Med Chem. 2007;50:186-91.PMID: 17228860
Howe DK, Honore S, Derouin F, Sibley LD. Determination of genotypes of Toxoplasma gondii
strains isolated from patients with toxoplasmosis. J Clin Microbiol. 1997;35:1411-4.
Hurt DE, Widom J, Clardy J. Structure of Plasmodium falciparum dihydroorotate
dehydrogenase with a bound inhibitor. Acta Crystallogr D Biol Crystallogr. 2006;62(Pt
3):312-23. PMID: 16510978
Hurt DE, Sutton AE, Clardy J. Brequinar derivatives and species-specific drug design for
dihydroorotate dehydrogenase. Bioorg Med Chem Lett. 2006; 15;16(6):1610-5. Epub 2006
Jan 10.PMID: 16406782
Huynh MH, Carruthers VB. Tagging of endogenous genes in a Toxoplasma gondii strain
lacking Ku80. Eukaryot Cell. 2009;8:530-9. PMID: 19218426
Hyde JE. Targeting purine and pyrimidine metabolism in human apicomplexan parasites. Curr
Drug Targets. 2007;8:31-47. PMID: 17266529
Iltzsch MH. Pyrimidine salvage pathways in Toxoplasma gondii. J Eukaryot Microbiol.
1993;40:24-8. PMID: 8457800
Jöckel J, Wendt B, Löffler M. Structural and functional comparison of agents interfering with
dihydroorotate, succinate and NADH oxidation of rat liver mitochondria. Biochem
Pharmacol. 1998;56:1053-60. PMID: 9776318

68
Kessl JJ, Lange BB, Merbitz-Zahradnik T, Zwicker K, Hill P, Meunier B, Pálsdóttir H, Hunte C,
Meshnick S, Trumpower BL. Molecular basis for atovaquone binding to the cytochrome
bc1 complex. J Biol Chem. 2003;278:31312-8. PMID: 12791689
Knecht W, Bergjohann U, Gonski S, Kirschbaum B, Löffler M. Functional expression of a
fragment of human dihydroorotate dehydrogenase by means of the baculovirus expression
vector system, and kinetic investigation of the purified recombinant enzyme. Eur J
Biochem. 1996;240:292-301. PMID: 8925840
Knecht W, Löffler M. Species-related inhibition of human and rat dihydroorotate dehydrogenase
by immunosuppressive isoxazol and cinchoninic acid derivatives. Biochem Pharmacol.
1998;56:1259-64. PMID: 9802339
Kovarik JM, Burtin P. Immunosuppressants in advanced clinical development for organ
transplantation and selected autoimmune diseases. Expert Opin Emerg Drugs. 2003;8:47-
62. PMID: 14610911
Kow RL, Whicher JR, McDonald CA, Palfey BA, Fagan RL. Disruption of the proton relay
network in the class 2 dihydroorotate dehydrogenase from Escherichia coli. Biochemistry.
2009;48:9801-9. PMID: 19694481
Laemmli UK. Cleavage of structural proteins during the assembly of the head of bacteriophage
T4. Nature. 1970;227:680-5.
Lim L, Kalnon M, McFadden GI. New proteins in the apicoplast membranes:time to rethink
apicoplast protein targeting. Trends Parasitol 2009; 25:197-200.

69
Lin SS, Gross U, Bohne W. Type II NADH dehydrogenase inhibitor 1-hydroxy-2-dodecyl-
4(1H)quinolone leads to collapse of mitochondrial inner-membrane potential and ATP
depletion in Toxoplasma gondii. Eukaryot Cell. 2009;8:877-87. PMID: 19286986
Lin SS, Kerscher S, Saleh A, Brandt U, Gross U, Bohne W. The Toxoplasma gondii type-II
NADH dehydrogenase TgNDH2-I is inhibited by 1-hydroxy-2-alkyl-4(1H)quinolones.
Biochim Biophys Acta. 2008;1777:1455-62. PMID: 18786503
Ling Y, Li ZH, Miranda K, Oldfield E, Moreno SN. The farnesyl-diphosphate/geranylgeranyl-
diphosphate synthase of Toxoplasma gondii is a bifunctional enzyme and a molecular
target of bisphosphonates. J Biol Chem. 2007;282:30804-16. PMID: 17724033
Liu S, Neidhardt EA, Grossman TH, Ocain T, Clardy J. Structures of human dihydroorotate
dehydrogenase in complex with antiproliferative agents. Structure. 2000;8:25-33 PMID:
10673429
Löffler M, Fairbanks LD, Zameitat E, Marinaki AM, Simmonds HA.Pyrimidine pathways in
health and disease. Trends Mol Med. 2005;11:430-7. PMID: 16098809
Luft BJ, Remington JS. Toxoplasmic encephalitis in AIDS. Clin Infect Dis. 1992;15:211-22.
PMID: 1520757. PMID 11859373
Melo EJ, Attias M, De Souza W. The single mitochondrion of tachyzoites of Toxoplasma
gondii. J Struct Biol. 2000;130:27-33. PMID: 10806088
McFadden DC, Camps M, Boothroyd JC. Resistance as a tool in the study of old and new drug
targets in Toxoplasma. Drug Resist Updat. 2001;4:79-84. PMID: 11512524

70
Montoya JG, Remington JS. Management of Toxoplasma gondii infection during pregnancy.
Clin Infect Dis. 2008;47:554-66. PMID: 18624630
Montoya JG, Liesenfeld O. Toxoplasmosis. Lancet. 2004;363:1965-76. PMID: 15194258
Nielsen FS, Rowland P, Larsen S, Jensen KF. Purification and characterization of
dihydroorotate dehydrogenase A from Lactococcus lactis, crystallization and preliminary
X-ray diffraction studies of the enzyme. Protein Sci. 1996;5:852-6. PMID: 8732756
Nishi M, Hu K, Murray JM, Roos DS. Organellar dynamics during the cell cycle of Toxoplasma
gondii. J Cell Sci. 2008;121:1559-68. PMID: 18411248
Nørager S, Jensen KF, Björnberg O, Larsen S. E. coli dihydroorotate dehydrogenase reveals
structural and functional distinctions between different classes of dihydroorotate
dehydrogenases. Structure. 2002;10:1211-23.PMID: 12220493
Patel V, Booker M, Kramer M, Ross L, Celatka CA, Kennedy LM, Dvorin JD, Duraisingh MT,
Sliz P, Wirth DF, Clardy J. Identification and characterization of small molecule inhibitors
of Plasmodium falciparum dihydroorotate dehydrogenase. J Biol Chem. 2008;283:35078-
85. PMID: 18842591
Parsons M, Karnataki A, Derocher AE. Evolving insights into protein trafficking to the multiple
compartments of the apicomplexan plastid. J Eukaryot Microbiol. 2009;56:214-20. PMID:
1952734
Pfefferkorn ER. Toxoplasma gondii: the enzymic defect of a mutant resistant to 5-
fluorodeoxyuridine. Exp Parasitol. 1978;44:26-35. PMID: 146608

71
Phillips MA, Gujjar R, Malmquist NA, White J, El Mazouni F, Baldwin J, Rathod PK.
Triazolopyrimidine-Based Dihydroorotate Dehydrogenase Inhibitors with Potent and
Selective Activity against the Malaria Parasite Plasmodium falciparum. J Med Chem.
2008;51:3649-53. PMID: 18522386
Pino P, Foth BJ, Kwok LY, Sheiner L, Schepers R, Soldati T, Soldati-Favre D. Dual targeting of
antioxidant and metabolic enzymes to the mitochondrion and the apicoplast of Toxoplasma
gondii. PLoS Pathog. 2007;3:e115. PMID: 17784785
Pomel S, Luk FC, Beckers CJ. Host cell egress and invasion induce marked relocations of
glycolytic enzymes in Toxoplasma gondii tachyzoites. PLoS Pathog. 2008;4:e1000188.
PMID: 18949028
Pettersen EF, Goddard TD, Huang CC, Couch GS, Greenblatt DM, Meng EC, Ferrin TE. J
Comput Chem. UCSF Chimera--a visualization system for exploratory research and
analysis. 2004 Oct;25(13):1605-12.
Rawls J, Knecht W, Diekert K, Lill R, Löffler M. Requirements for the mitochondrial import
and localization of dihydroorotate dehydrogenase. Eur J Biochem. 2000;267:2079-87.
PMID: 10727948
Rowland P, Nielsen FS, Jensen KF, Larsen S. The crystal structure of the flavin containing
enzyme dihydroorotate dehydrogenase A from Lactococcus lactis. Structure. 1997;5:239-
52. PMID: 9032071

72
Robles Lopez SM, Hortua Triana MA, Zimmermann BH. Cloning and preliminary
characterization of the dihydroorotase from Toxoplasma gondii. Mol Biochem Parasitol.
2006;148:93-8. PMID: 16621066
Rowland P, Nørager S, Jensen KF, Larsen S. Structure of dihydroorotate dehydrogenase B:
electron transfer between two flavin groups bridged by an iron-sulphur cluster. Structure.
2000;8:1227-38. PMID: 11188687
Saito T, Nishi M, Lim MI, Wu B, Maeda T, Hashimoto H, Takeuchi T, Roos DS, Asai T. A
Novel GDP-dependent Pyruvate Kinase Isozyme from Toxoplasma gondii Localizes to
Both the Apicoplast and the Mitochondrion. J Biol Chem. 2008;283:14041-52. PMID:
18326043
Sambrook J, Fritsch EF, Maniatis T. Molecular Cloning: A Laboratory Manuel, 2nd
ed. Cold
Spring Harbor Laboratory Press; 1989.
Schwede T, Kopp J, Guex N, Peitsch MC. SWISS-MODEL: An automated protein homology-
modeling server. Nucleic Acids Res. 2003;31:3381-5. PMID: 12824332
Seeber F, Limenitakis J, Soldati-Favre D. Apicomplexan mitochondrial metabolism: a story of
gains, losses and retentions. Trends Parasitol. 2008;24:468-78 PMID: 18775675
Sheiner L, Soldati-Favre D. Protein trafficking inside Toxoplasma gondii. Traffic. 2008;9:636-
46. PMID: 18331382
Sigoillot FD, Evans DR, Guy HI. Growth-dependent regulation of mammalian pyrimidine
biosynthesis by the protein kinase A and MAPK signaling cascades. J Biol Chem.
2002;277:15745-51. PMID: 11872754

73
Sierra Pagan ML, Zimmermann BH. Cloning and expression of the dihydroorotate
dehydrogenase from Toxoplasma gondii. Biochim Biophys Acta 2003;1637:178-181.
Small I, Peeters N, Legeai F, Lurin C. Predotar: A tool for rapidly screening proteomes for N-
terminal targeting sequences. Proteomics. 2004;4:1581-90. PMID: 15174128
Sun F, Huo X, Zhai Y, Wang A, Xu J, Su D, Bartlam M, Rao Z. Crystal structure of
mitochondrial respiratory membrane protein complex II. Cell. 2005;121:1043-57. PMID:
15989954
Tomavo S, Boothroyd JC. Interconnection between organellar functions, development and drug
resistance in the protozoan parasite, Toxoplasma gondii. Int J Parasitol. 1995;25:1293-9.
PMID: 8635881
Tonhosolo R, D'Alexandri FL, Genta FA, Wunderlich G, Gozzo FC, Eberlin MN, Peres VJ,
Kimura EA, Katzin AM. Identification, molecular cloning and functional characterization
of an octaprenyl pyrophosphate synthase in intra-erythrocytic stages of Plasmodium
falciparum. Biochem J. 2005;392:117-26. PMID: 15984931
Ullrich A, Knecht W, Piskur J, Löffler M. Plant dihydroortate dehydrogenase differs
significantly in substrate specificity and inhibition from the animal enzymes. FEBS Lett.
2002;529:346-50. PMID: 12372626
Ullrich A, Knecht W, Fries M, Löffler M. Recombinant expression of N-terminal truncated
mutants of the membrane bound mouse, rat and human flavoenzyme dihydroorotate
dehydrogenase. A versatile tool to rate inhibitor effects? Eur J Biochem. 2001;268:1861-8.
PMID: 11248707

74
Vercesi AE, Rodrigues CO, Uyemura SA, Zhong L, Moreno SN. Respiration and oxidative
phosphorylation in the apicomplexan parasite Toxoplasma gondii. J Biol Chem.
1998;273:31040-7. PMID: 9813002
Vollmer M, Thomsen N, Wiek S, Seeber F. Apicomplexan parasites possess distinct nuclear-
encoded, but apicoplast-localized, plant-type ferredoxin-NADP+ reductase and ferredoxin.
J Biol Chem. 2001;276:5483-90. PMID: 11056177
Waller RF, McFadden GI. The apicoplast: a review of the derived plastid of apicomplexan
parasites. Curr Issues Mol Biol. 2005;7:57-79. PMID: 15580780
Weiss LM, Dubey JP. Toxoplasmosis: A history of clinical observations. Int J Parasitol.
2009;39:895-901.
Zameitat E, Gojković Z, Knecht W, Piskur J, Löffler M. Biochemical characterization of
recombinant dihydroorotate dehydrogenase from the opportunistic pathogenic yeast
Candida Albicans. FEBS J. 2006;273:3183-91. PMID: 16774642
Zameitat E, Knecht W, Piskur J, Löffler M. Two different dihydroorotate dehydrogenases from
yeast Saccharomyces kluyyeri. FEBS Lett. 2004;568:129-34. PMID: 15196933
Zameitat E, Freymark G, Dietz CD, Löffler M, Bölker M. Functional expression of human
dihydroorotate dehydrogenase (DHODH) in pyr4 mutants of ustilago maydis allows target
validation of DHODH inhibitors in vivo. Appl Environ Microbiol. 2007;73:3371-9. PMID:
17369345

75
Zimmermann BH, Evans DR. Cloning, overexpression, and characterization of the functional
dihydroorotase domain of the mammalian multifunctional protein CAD. Biochemistry.
1993;32:1519-27.


![Primerdesign Ltd TM Toxoplasma gondii - Home : genesig · Toxoplasma gondii is a species of parasitic protozoa in the genus Toxoplasma.[1] The definitivehostofT.gondiiisthecat,buttheparasitecanbecarriedbythevastmajorityof](https://static.fdocuments.in/doc/165x107/5cc21bb288c993ed078d60da/primerdesign-ltd-tm-toxoplasma-gondii-home-toxoplasma-gondii-is-a-species.jpg)







![Research Article Toxoplasma gondii Pave the Road for Dementia?downloads.hindawi.com/journals/jpr/2020/8859857.pdf · including Toxoplasma gondii (T. gondii) [6], Herpes simplex virus-1](https://static.fdocuments.in/doc/165x107/5f9376ed91220772b35c9b7d/research-article-toxoplasma-gondii-pave-the-road-for-dementia-including-toxoplasma.jpg)

![Toxoplasma gondii - PCRmax · Toxoplasma gondii is a species of parasitic protozoa in the genus Toxoplasma.[1] The definitive host of T. gondii is the cat, but the parasite can be](https://static.fdocuments.in/doc/165x107/5cc21bb288c993ed078d60e8/toxoplasma-gondii-toxoplasma-gondii-is-a-species-of-parasitic-protozoa-in.jpg)





