
Diffuse Noxious Inhibitory Controls (DNIC) in Animals...
10
Diffuse Noxious Inhibitory Controls (DNIC) in Animals andin Man Key words Acupuncturc, DNIC, Electro acupuncture, Netve fibres, Neuro-anatomy, Pain pathways, Stimulation inducedanalgesia. TENS. Introduction The transmission of nociceptive signalscan be modrrlated by powerful controls at as earlya stage as the fifst spinal relay. These controls include both segmental mechanisms and systems which involve supraspinal structures, and some of them can be triggered by somaesthetic stimuli(see relerences in Le Bars er a/.,1984; 1986). This last point is true for segmental mechanisms which can be triggered by stimulation of the corre- sponding dermatome : the responses of dorsal horn neurones to nociceptive stimuli can be inhibited by innocuous stimulation of large diarneter cutaneous fibres. lt is generally thought that these phenomena are triggeredby the activationof Aop fibres alone; however,numerous studieshave demonstrated that the activation of A8-fibres produces the most powerful seSmental inhibitions (Chung et a1.,1984; Lee et a/.,1985; Sj6lund,19B5; Woolf et a/., 1980). Such effects, which are essentially restricted to dermatomes, are deriveddirectlyfrom the properties of the receptive fields of dorsalhorn neurones. They could explain the hypo-al8esia which can be elicited by highfrequency, low intensity stimulation of peripheral netves ("Tftnscutaneous Electrical Stimulation", TENS.) and by some forms ol acupuncture or electro-acupuncture. lt should be noted however, that the time constantsof these clinical effects and of the electrophysiological phenomena are very different : patientscan gain pain relief which lasts for hoors after such strmulation \^hereds lhe :nhibilion ol neuroncs in animals or of nociceptive reflexes in man can end as soon asthe stimulation stops. However, another category of somaesthetic stimulus can also induce hypo-algesic ef{ects. Although it seems paradoxical at first, painful stimuli can diminish, or even mask, pain elicited by stimulationof a remote(extra-segmental) part of the body (see references in Le Bars et a/.,1984; Le Bars and Villanueva, 19BB). This phenomenon has been known of since ancient times and has even been usedduring surgical procedures on both man and domesticated animals. In the latter category, two examples arethe useof the twitch in horses and of nasalforcepsin cattle for performing caudectomies or castrations, both of which are potentially painful operations. The nature of the controls which underlie these observations is differentfrom that of the inhibitory phenomena described above which aretriggered by light stimuli and are essentially segmental. Accordingly, we have developed the working hypothesis that some of the neurones which are involved in the transmission of nociceptive signals can be inhibited by nociceptive stimulation of peripheral territories outsidetheir own excitatory receptive fields. That such an hypothesis is correct at as early a stage as the spinal cord was revealed by the finding that some dorsal horn neurones are stronsly inhibited when a nociceptive stimulus is applied to any part of the body,distinct from their excitatory receptive fields. Forconvenience, this phenomenon was iermed "Dif{use Noxious lnhibitory Controls" (DNIC). L Diffuse Noxious Inhibitory Controls in animals In the rat and the cat, the activity of certarn dorsal horn neurones can be strongly inhibited by noxious inputs. Sucheffects do not appear to be somatotopi- cally organised but apply to the whole body and affect all convergent neurones, whether in the dorsal hornof various segments of the spinal cord (Calvino et a/.,I 984;Le Bars et a/.,I 979aj Morton et a1., 1987) or in the trigeminal nucleus caudalis (Dickenson et a/., 1980a). By contrast, DNIC do not affect the other neuronal types which are found in thesestructures, i.e. noxious-specific, non-noxiousspecific, cold .responsive and proprioceptive neurones (Dickenson et a/.,I980; Le Bars ef a/.,1979b). The principal feature of DNIC is that they can be tri8gered by conditioning stimuliapplied to any part of the body distant from the excitatory receptive field of the neurone understudy, provided that the stimuliare clearlynoxious. Indeed, DNIC can be triggered by any heterotopic nociceptive stimulus whatever its type - mechanical, thermal, chemical, or electrical - whereasnon noxious stimuli are completelyineffective. With strong stimuli, the inhibitory effects are powerful and are followed by long-lasting after-effects which can persist for several minutes. 47 on 17 September 2018 by guest. Protected by copyright. http://aim.bmj.com/ Acupunct Med: first published as 10.1136/aim.9.2.47 on 1 January 1991. Downloaded from
Transcript of Diffuse Noxious Inhibitory Controls (DNIC) in Animals...

Diffuse Noxious Inhibitory Controls(DNIC) in Animals and in Man
Key wordsAcupuncturc, DNIC, Electro acupuncture, Netvefibres, Neuro-anatomy, Pain pathways, Stimulationinduced analgesia. TENS.
IntroductionThe transmission of nociceptive signals can bemodrrlated by powerful controls at as early a stageas the f i fst spinal relay. These controls include bothsegmental mechanisms and systems which involvesupraspinal structures, and some of them can betriggered by somaesthetic st imuli (see relerences inLe Bars er a/.,1984; 1 986).This last point is true for segmental mechanisms
which can be tr iggered by stimulation of the corre-sponding dermatome : the responses of dorsal hornneurones to nociceptive st imuli can be inhibited byinnocuous stimulation of large diarneter cutaneousfibres. l t is general ly thought that these phenomenaare triggered by the activation of Aop fibres alone;however, numerous studies have demonstrated thatthe activation of A8-fibres produces the mostpowerful seSmental inhibit ions (Chung et a1.,1984;Lee et a/.,1985; Sj6lund,19B5; Woolf et a/., 1980).Such effects, which are essentially restricted todermatomes, are derived directly from the propertiesof the receptive fields of dorsal horn neurones. Theycould explain the hypo-al8esia which can beelicited by high frequency, low intensity st imulationof peripheral netves ("Tftnscutaneous ElectricalStimulation", TENS.) and by some forms olacupuncture or electro-acupuncture. l t should benoted however, that the time constants of theseclinical effects and of the electrophysiologicalphenomena are very different : patients can gainpain rel ief which lasts for hoors after suchstrmulation \^hereds lhe :nhibi l ion ol neuroncs inanimals or of nociceptive reflexes in man can end assoon as the stimulation stops.
However, another category of somaestheticstimulus can also induce hypo-algesic ef{ects.Although it seems paradoxical at f irst, painful st imulican diminish, or even mask, pain el icited bystimulation of a remote (extra-segmental) part of thebody (see references in Le Bars et a/.,1984; Le Barsand Vil lanueva, 19BB). This phenomenon has beenknown of since ancient t imes and has even beenused during surgical procedures on both man anddomesticated animals. In the latter category, two
examples are the use of the twitch in horses and ofnasal forceps in cattle for performing caudectomiesor castrations, both of which are potential ly painfuloperations.The nature of the controls which underl ie these
observations is different from that of the inhibitoryphenomena described above which are tr iggered byl ight st imuli and are essential ly segmental.Accordingly, we have developed the workinghypothesis that some of the neurones which areinvolved in the transmission of nociceptive signalscan be inhibited by nociceptive st imulation ofperipheral terr i tories outside their own excitatoryreceptive f ields.That such an hypothesis is correct at as early a
stage as the spinal cord was revealed by the f indingthat some dorsal horn neurones are stronslyinhibited when a nociceptive st imulus is applied toany part of the body, dist inct from their excitatoryreceptive f ields. For convenience, this phenomenonwas iermed "Dif{use Noxious lnhibitory Controls"(DNIC) .
L Diffuse Noxious Inhibitory Controls in animalsIn the rat and the cat, the activity of certarn dorsalhorn neurones can be strongly inhibited by noxiousinputs. Such effects do not appear to be somatotopi-cal ly organised but apply to the whole body andaffect al l convergent neurones, whether in the dorsalhorn of various segments of the spinal cord (Calvinoet a/., I 984; Le Bars et a/., I 979aj Morton et a1., 1987)or in the tr igeminal nucleus caudalis (Dickenson eta/., 1980a). By contrast, DNIC do not affect the otherneuronal types which are found in these structures,i .e. noxious-specif ic, non-noxiousspecif ic, cold
.responsive and proprioceptive neurones (Dickensonet a/., I980; Le Bars ef a/.,1979b).The principal feature of DNIC is that they can be
tri8gered by condit ioning stimuliapplied to any partof the body distant from the excitatory receptivefield of the neurone under study, provided that thestimuli are clearly noxious. Indeed, DNIC can betriggered by any heterotopic nociceptive st imuluswhatever i ts type - mechanical, thermal, chemical,or electr ical - whereas non noxious stimuli arecompletely ineffective. With strong stimuli, theinhibitory effects are powerful and are followed bylong-lasting after-effects which can persist for severalm inutes.
47
on 17 Septem
ber 2018 by guest. Protected by copyright.
http://aim.bm
j.com/
Acupunct M
ed: first published as 10.1136/aim.9.2.47 on 1 January 1991. D
ownloaded from

When the generaJ characterist ics of DNIC areanalysed, one str ikinB feature is their capacity toaffect al l kinds of activity in convergent neurones,no matter whether i t is evoked by r-roxious or nonnoxious, natural or electr ical periphefal st imuli orby the direct microelectrophoretic application ofexcitatory amino acids (Vil lanueva et a/.,1984a;i 984b). Transcutaneous electr ical st inrul i applied tothe receptive f ield of converBent neurones activatelarBe (A) and thin (C) f ibr€s, and in studies of DNIC,suprathreshold currents have been employedsysternatical ly in lhis way, to evoke reproducible "C-f ibre responses" from convergent neurones. Alinoxious condit ioning stimuli tested to date, havemarkedly inhibited these responses (FEUre tr).
MeaD pet .entate inhib i tbns (ot . l inate) o i C f ibrc evakedrcspanses af triBertinal lupper cLtrc) ,itd lunbat tnwer .utve)canvcqent neurcnes, indu.cd by innercion of the tail in watelbaths at vatious tempetaturci labr.issn). Nore rbir no sigrtli.anrellects wetc ol)setred when 36"C ot 4(fC was applie.l ta th€tail but tlit lErc \yas a hi\hly sieniti(aDt corclauon t)er\|ecnnoxious tutnpeetutes (11 52'C) and the exrlnt of the inhibitionslFran Le Bars et a|.,1986).An examplc af the effect af DNIC upon rcspanss of a
canveryenl unit b A' and C-Ubrc activatian illustrated by a dot.lisplay analysis of the neurcndl rcspans€s with tine tunningupwatds. The appli.ation ai themal (innesion of the lail in52'C waIel, nechaDical (pih.h af the tail and nuzzle) andviscenl (8!E bn(lykinin i.p.) naxiaus nnnuli rcsultetl in nrontinhibitians af resparses to both A' and C fibtcs, indo.e.l bt)tanscutaneous ele.tti.:al stimulattan af tlle dbtal part ol thehindlimb. Note the lon9 dtrration af the aftercffects, especiallyfat the C-tihrc reqonses (frcn. l-e 8a6 et al., 1983).
DNIC are not observed in anaesthetised ordecerebrate animals in which the spinal cord hasbeen sectioned (Cadden et a/., l9B3; Le Bars eta/.,1979b; Morton et a|.,1987). l t is thereloreobvious that the mechanisms underlying DNIC arenot confined to the spinal cord and that supraspinalstructures must be involved. In this respect, i t isimportant to note that such a system rnust becompletely differenL froni segmental inhibitorysystems which work both in intact and in spinalanimals. Furlherrnore, seBmental inhibit ions can betriggered by the activation of jow thresholdafferents. DNIC are also very different from thepropriospinal inhibitory processes which can betrig8ered by noxious inputs (Cadden et a/.,1983;Fitzgerald, 1 9B2; Cerhart el a/.,1981).The peripheral and central mechanisms involved inDNIC are considered below
A. Peripheral mechanismsThe relationship between the intensity of a noxiousstimulus and lhe strength of the resultant DNIC wasinvestigated by studying the eflects of varioustemperatures applied to the lai l , on the CJibreresponses of lumbar and Lrigeminal convergentneurones to Lranscutaneous electr ical st imulation oftheir receptive f ields on lhe hindpaw or face (LeBafs et a/.,1981a; Vil lanueva and Le Bars,l 985). Asshown in F/gure 2, the threshold for producingDNIC was between 40'and .+4'C, and above thistemperature ( in the 44-52'C range), a hiShlysignjf icanl correlation existed between Lne conolt ioning lemperature and the extent of the inhibit ion.These data reinforce the hypothesis that DNIC aretr iggered specif ical ly by the activation of peripheralnociceplors whose signals are carried by 46- and Cfibres (Dubner and Beitel,1976; Lamore anoCampbell, l9lB; Torebj6rk er a/.,1984; Vil lanuevaand Le Bars, 1985).
In order to furlher investigale the types ofperipheral f ibres involved in DNtC, we tookadvantage of the facls :( i) that tr igeminal and spinal dorsar norn
neurones respond with relaLively steadydischarges 1.)) the eJectrophoretic applical ionol excitatory amino-acids, and
48
on 17 Septem
ber 2018 by guest. Protected by copyright.
http://aim.bm
j.com/
Acupunct M
ed: first published as 10.1136/aim.9.2.47 on 1 January 1991. D
ownloaded from

( i i) that DNIC act on convergent neurones by afinal postsynaptic inhibitory rnechanisminvolving hyperpolarisation of the neuronalmembrane (Vil lanueva et al.,19B1a; 1984b).
It was found that when tr igeminal convergentneurones were directly excited by the elec-trophoretic application of di homocysteate (DLH),the percutaneous application of single square-wave,electr ical st imuli (10rriA; 2ms) to the tai l alwaysinduced a biphasic depression of the resultantactivity (Bouhassira et al.,19a7). Both the early andlate components of this inhibit ion occuned withshorter latencies when lhe base rather than the t ip ofthe tai l was stimulated (F€ure 3). When the lwostimulation sites were I00 mm apart, i t was possibleto use these differences in latency, to estimate theconduction velocit ies of the peripheral f ibrestr iggerinS the inhibi l ions. For the onset of the earl ierand later con ponents of the inhibit ion, the meandifferences between the latencies from the two sitesof st imulation were | 3.6 and 147.7 ms respectively,which conesponds to peripheraJ conductionvelocit ies of 7.3 and o-7 m/s. Such conductionvelocit ies fal l into the Ab and C-fibre tanges,resPectively.AlthouSh peripherai unmyelinated and thin
myeJinated f ibres can respond to st imuli below thepain threshold (Adriansen ef a/.,1983; Cybels etal.,1979; forcbtotl< et a1.,1984; Van Hees andCybels,l972), the relationship between activity insuch f ibres and nociceptive reactions or pain, is aclassical one (Adriansen et a/.,1983; Dubner andBeitel,1976; Cybels et a/., 1979; Lamotte andCampbell, l97B; Torebjork et a/.,1984; Van Heesand Cybels,1972). However, the thresholds fortr iggering the A6 and C-fibre components of DNICwere found to be in the 0.25-0.5mA and 1 2mAranges respectively, which might suggest a contribution by non-nociceptive afferents-The importance of A8Jibre activation in the
production of analgesia or anlinociceptive effects bysomatic,^lectr ical st imulation has been suggested bysevefal authors (Chung et ai.,1984) Kawakita andFunakosh i , l 9B2 ; Lee e t a / . , 198 -5 ; S j c i l und ,19B5 ;Woolf et a/., l9B0). In this respect, our resultsshowed that, by comparison with the C f ibrecomponent, the A6jibre component of inhibit ionwas easier to el iciL and had a more corrsrarrrmagnitude and duration : i t was found with lowerintensit ies of percutaneous electr ical sl imulatron andrapidly reached its maximal effect when the currentwas increased. Applying slronger intensit ies ot
B(a)
l
I"l
Ex.rnpie of heterctapic a.tivatia. ol Aii- ind CJibrcs t.iiSctin1 inhibitians in a LtiFninal converyent neurcnc lfan Bouhassira ."t
A. s.lremart rep,cscrr.rtion ai the expetinental dcsi\n. Neurones with rc.eptive fields lacatcd ipsilatetally on the muzzle ||ercrecatde.l b the iSeninal nucleus c,iodalis. Th€ cantinuaus electrcphoteLlc appli.atian af an cx.iabty amino ackl, tll homocystei.
aci.l IDLH), i!1du.ed a stcady discharye hotn he neutune undet nudy. Ihe tepetilive applicatiDn af indivi.lual pct.utancaus,
eledn)l stim]li ol Jdequatc intensities ta the base la) or the tip (b) af the tail in<lDced biphasic depre$ions of a.tivity which
aiEcted conretgent neurctrcs er./ust'e/f.B- tndlvidual -"xanple oi thc biphastc inhibitott ptocest5 tti\Bercrl by rcpetilive, single, percutaneoLts, electrical stinuli (2ms
dLtntian, lAn)A, o.66Hz, 2A0ms del.y) applie.l Lo the basc (a) ot the tip (b) al tl)e tail on the dis.harge evaked in a tti9€ninal.onverye]! neunn€ by thc continuars cle.trcphareti. applicatian af Dt H (t 7nA). PeristitnLlus histogtans (bin width: lns lcli: 5nstight) \|ere conrtu.tudian l0o ials. The ea iet.a]ryonent oithe inhibnion kshow] in dehilin the l-.ft partafthe fi\ure, while
the whole biphaslc lnhibilion is slnwn on the tieht. Nole that bath .ompanenx appearc.l eatliet when the base G) as oppase.) tathe tip (b)oi the teil||as stinulatcd.
on 17 Septem
ber 2018 by guest. Protected by copyright.
http://aim.bm
j.com/
Acupunct M
ed: first published as 10.1136/aim.9.2.47 on 1 January 1991. D
ownloaded from
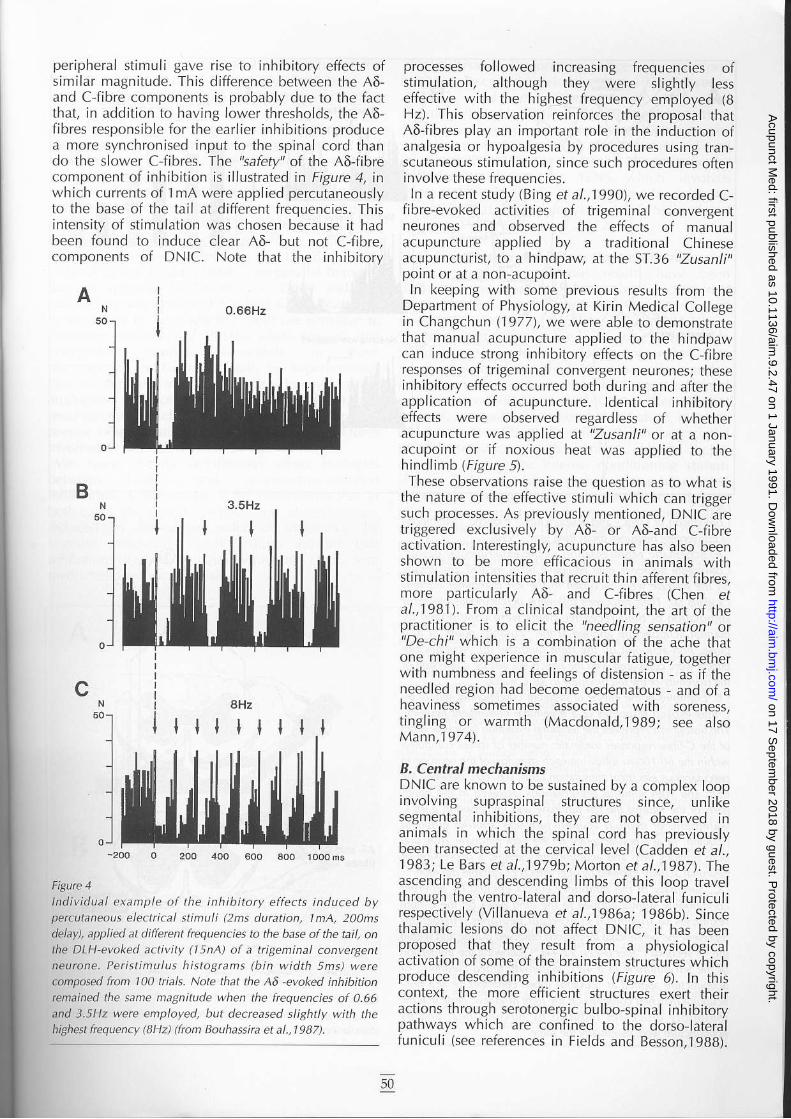
peripheral st imuli gave rise to inhibitory effects ofsimjlar magnitude- This difference b€tween the A6-and C f ibre cornponents is probably due to the {actthat, in addit ion to havinB lower thresholds, the ,46,f ibres responsible for the earl ier inhibit jons producea more synchronised input to the spinal cord thando the slower C-fibres. The '!atety" of the A6-librecomponent of inhjbit ion is i l lustrated in Figure I, inwhich currcnts of I mA were applied percutaneoLrslyto the base of the tai l at dif lefent frequencies. Thisintensity of st imuJalion was chosen because t l hadbeen found lo induce clear 46 but not CJibre,components of DNIC. Note that the inhibitory
A iO.66Hz
processes fol lowed increasinS frequencies ofst imulation, although they were sl ightly lesseffective with the highest frequency employed (BHz). This observation reinforces lhe proposal lhat46 f ibres play an important role in the induction ofJnJl8.\ i" o. h\pod/EF-'.r br prn, edure. r. irB ran-scutaneous stimulation, since such procedures ol leninvolve these frequencies.ln a recent study (Bing et a|.,1990), we recorded C-
fibre-evoked actjvi l ies of tr igeminal convergentneurones and observed the effects or rnanuaracupuncture applied by a lradit ional Chineseacupuncturist, to a hindpaw, aI the SL36 "Zusanli"point or at a non acupoint.
In keeping with some previous results f iom theDepartment of Physiology, at Kir in Medical ColJegein Changchun (1977), we were able to demonstratethat manual acupuncture applied to the hindpawcan induce strong inhibitofy effects on the C f ibreresponses ol tr igeminal convergent neurones; theseinhibitory effects occurred both during and after theapplication ol acupuncture. ldentical inhibitoryeffects were observed regardless of whetheracupuncture was applied at "Zusanli" ot at a non-acupoint or i l noxious heat was applied to thehindlimb (F/gure.t.These obsefvations raise the question as to what is
the nature of the effective st imuJi which can tr iggersuch proccsses. As previously mentioned, DNIC aretr iSgercd excJusively by 46- or 46 and CJibreactivation. lnterestingly, acupuncture has atso beenshown to be more eff icacious in animals withstimulation intensit ies that recruit thin afferent f ibres,more part icularly A5 and CJibfes (Chen eta/.,1981). From a cl inical standpoint, the art of thepractilioner is to elicit the "needling sensation" ol'De chl" which is a combination of the ache thatone mjght experience in rnLrscular fat igue, togetherwith nunrbness and feelings of distension - as i f theneedled region had become oedematous - and of aheaviness sometimes associated \\/ i th soreness,t inBJing or warmth (MacdonaId,t 989; see alsoMann , I974 ) .
R. Central mechanismsDNIC are known to be sustained by a complex loopinvolving supraspinal structures since, unlikeseSmental inhibit ions, they are not observ€d inanir.aJs in which the spinal cord has previouslybeen transected at the cervical level (Cadden et a/.,I983; Le Bafs et a/.,1979b; Morton et a/.,1987). TheascendinB and descending l imbs of this loop travelthrouBh the ventro lateral and dorso-lateral funicLrl irespectively (VilJanueva et al.,19B6a; 1986b). Sincethalarnic lesions do not affect DNIC, i t has beenproposed that they result from a physiologicalactivation of some of the brainstem structures whichprodLrce descending inhibit ions (FEUre 6). In thiscontext, the more efficient struclLrres exert theiractions through serotonerSic bulbo-spinal inhibitorypathways which are confined to the dorso lateralfunicul i (see references in Fields and Besson,19B8).
B
c
lnd i r idual exatnpie of the lnhib i to ty ef ie . ts induced bypercutaneaus eleclrical stinDli (2n! duntion, 1n1A, 20 rsdelay), applied at.liffercnt iequencies to the base ofthe tail, onthe DLH evaked a.tivlty (lsnA) of a ttiBeminal canvergenrneurone. Pet is t inu lus h is lo9 rns (b in width 5ns) wereconpased frcn 10A tiak. Note that the A5 -eyoked inhibi onrcnained the sane magnitude when the frcqucncies af 0.66and 3.5H2 were enpbyed, but.Jecteascd siightiy with thehighest hequency (\Hz) (ftani Boohassna et a|.,1987).
50
on 17 Septem
ber 2018 by guest. Protected by copyright.
http://aim.bm
j.com/
Acupunct M
ed: first published as 10.1136/aim.9.2.47 on 1 January 1991. D
ownloaded from
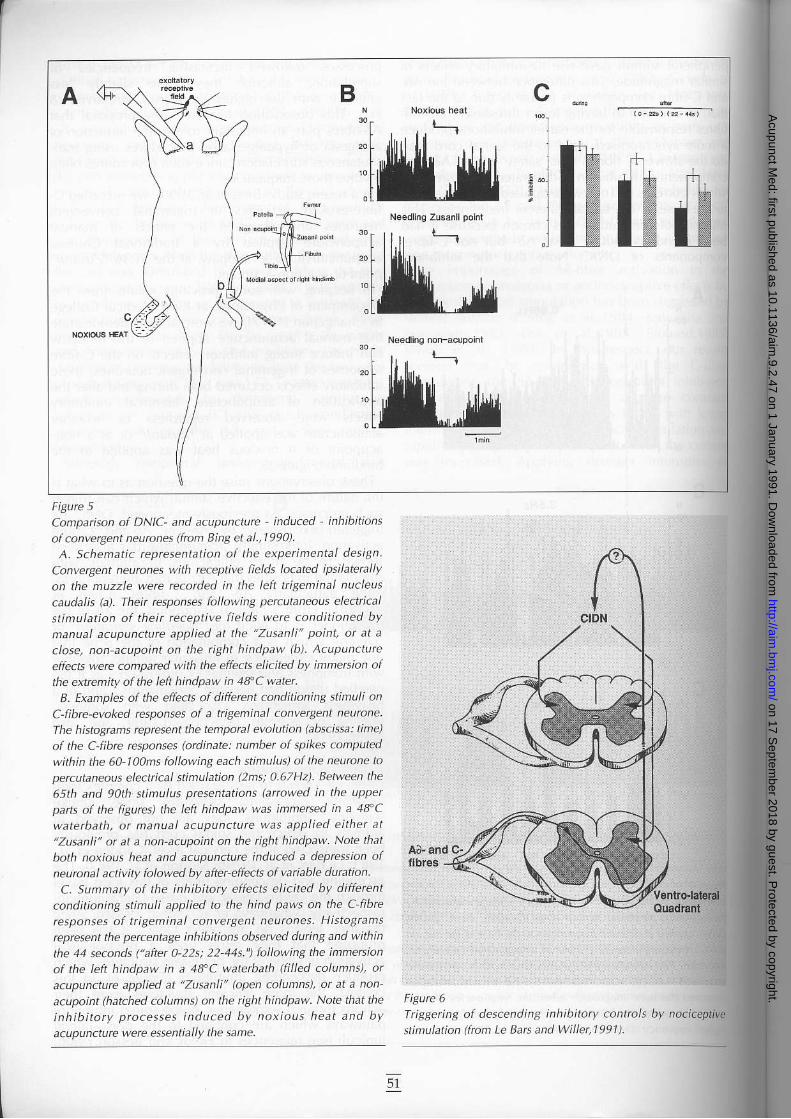
Compatisan af DNIC- and acupuncturc - induced inhibitions
of converyent neurones (fron ainS et a|.,1990).A. Schenatic reprcsentation of the experinental deslgn.
Convergent neurcnes with receptive fields located ipsilaterally
on the muzzle werc recorded in the left trigeminal nucleus
caudalis b). Then rcsponses following percutlneous etecticat
stimulatlon of their receptive fields werc conclitioned by
nanual acupuncturc applied at the "Zusanli" point, at at a
close, non acupoint on the right hindpaw (b). Acupuncture
etects were conpared with the effects elicite.J bl, inmersion ol
the extemity of the lelt hindpaw in 4n C wateLB. Exanples of the effects of ditrerent conditionin7 stimuli on
C-fibre-evoked responses of a ttigeminal converqent neurone.
The histoarans rcprcsent the temparal evolution (abscissa: tne)
of the C fibre responses (odinate: numbet of spikes canputed
within lhe 60 t oans fallowins each stinulus) af ke neunne t '
percutaneous electrical stimulatian (2ns; O.67Hz). Betuveen the
65th and 9Ath stinulus prcsentations larrowed in the upperpafts of the figurcs) the left hindpaw was inme5ed in a 4trC
wateftath, ot nanual acupuncturc v/as applied eithet atlzusanli'l ot at a non-acupointon the ight hinclpaw. Note that
both noxious heat and acupuncturc induced a depression ofneuronal activity folowed by aftercfiects of variable dutation
C. Sumnaty of the inhibitory effecE elicited by differcnl
conditioning stinuli applied to the hind paws on the CJibrc
rcspanses of triBeminal converSent neurones. Histograms
rcpresent the percentage inhihnions observed dutin7and within
the 41 seconds ("aker O-22s; 22 14s.") followin9 the immeBion
of the lell hindpaw in a 18'C waterbath (fi|led colunns), or
acupuncture applied at "Zusanli" (open columni, or at a non'
acupoint (hatched colunns) on the ti\ht hin.lpaw. Note that the
inhib i to ty processes induced by noxious heat and by
acupuncture were essentially the same.
t t t : , t : , : , , 1 ' . . t : , . . l ,
. ,11 l1 , ' ,1 . , , ,1 : . . t1 ;1 ; ,1 . , . ,
l
\ -alTr".
.],:::.].::.]i:ii.].::t.:.:::i]:,,.].::,::i.:]::::i:::::iiil !Geeh"B o l dp-""d 'np tnh bror \ \ontrc l . bstimulation (fron Le BaE and Willer199l).
l , ' '
i \ : i : i i
:it:{i:\r:
51
on 17 Septem
ber 2018 by guest. Protected by copyright.
http://aim.bm
j.com/
Acupunct M
ed: first published as 10.1136/aim.9.2.47 on 1 January 1991. D
ownloaded from

I t may be noted that electr ical st irnulation of theNucleus Raph6 MaSnus produces inhibit ions ofspinai convergent neurones which are as potent asDNIC.The hypothesis that these slructures part icipate in
tr iggering DNIC seems Lo be confirmed by thefindings that DNIC are profoundly reducedfol lowing : electrolyt ic lesions of Nucleus Raph6MaBnus, the administration of blockers of seroton-ergic receptors and section oi the dorso-laleralfuniculus; conversely, DNIC are polentiated by theadministration ol 5 hydroxytryptophane, a precursoroI serotonin (see references in Le Bars,19BB;Vil lanueva and Le Bars,l988). The paral lel betweenthe two types of inhibit ion i .e. DNIC and thatinduced by stimulation of Nucleus Raph6 MaSnus -is supported by the facts that both are sensit ive tonaloxone (Le Bars et a/.,1981b), which su8Bests thatendogenous opioids pa(icipaLe in thesephenomena, and both act f inal ly by hyperpolarisa-t ion of the neuronal membrane- Howevet theseanalogies do not prove the identi ly of themechanisms which are brought into play; theprecise determination of the supraspinal structuresinvolved in DNIC has sti lJ to be carried olrt-We have already emphasised some analogies
between DNIC and acupuncture-inducedinhibit ions. A further one is lhe observation that inboth cases, the inhibitory effects and after-effects arereduced by the sar.e order of magnitude bysystemic naloxore- These f indings indicate thatinhibit ions tr iggered by acupUncture and DNIC bothinvolve endogenous opioids.
The reversal of acupuncture analgesia by naloxonehas been described previously ( l iang,197B; Mayer eia|.,197/; and see references in Han andTerenius,l982; He,l9B7). Such an effect is padicuIarly obvious when low frequency, hiBh intensitystimuli are used (Huang et a1.,1986; Sj6lund andEriksson,l979; Wil ler et ai.,1982). Interestinlt ly, thehypoalgesia el icited by TENS which involvessegmental mechanisnrs - is not affected by nalorone(Hansson et a/.,1986; Lundeberg,l9B5; Sjolund andEriksson, 1979; WilJer et a/.,1 982)
Il. Diffuse Noxious lnhibitory Contrcls in manIn man, exactly analogous results have beenobtained in studies which have combinedpsychophysical measurements with the recordinB ofnociceptive reflexes (Wil ler et a/.,1984,19B9).Electr icaj st imulation of the sural ncrve al the anklesimultaneously indLrces a nociceptive rettex in aflexor muscle of the thigh (the Rrr reflex) and apainful sensation in the terf i tory of the nerve. Painfulheterotopic condit joninB stimuli, no matter whethertherrnal, mechanical or chemical in nature, increasethe thresholds of both the reflex and Lhe associatedpainful sensation, with strong€r effects beingobserved with more intense condit ioninS stimuJi(Figure a. fhese inhibitory phenomena are exertedselectively on nociceptive responses and do notaffect monosynaptic reflexes of proprioceptiveo ' iB rn . l he .p rp .u l t . d "mon . t r " t c l h r t ' 1 mr1 , Jpainful condit ioning stimuJus can depress both apre exist in€i pain and its associated nociceptivereflex at as early a stage as the f irst spinal relays ior
52
Rnr Reflex Erlclcnce lar DNIC ln n)an (iom Willet et rl.,t98t)A: [xperinental set-up use.] f.t clici )B a no.iceptive rcflex
lRt t t ) by n lnolat ian (s t ln t . ) o f the sLtn l netre 6.h. ) aDClte.orcths (rec.)frcn aver the l."ti Eiccps t'etnatis n)u!.]e (bi.).An exanple of the Rrt rc f lcx is shown i t the top af thediaBran (.alibratbns: harizantal 25tns; vertical IOOV). fhetef lex ercked by ju \ tathteshold (1.2 ! in tes th.eshalc l )stnnolation, is nudied befarc, dorinE, an<l nhet a 2 tninuteperiod of innlersion ot the ti\ht han.l in a thenlorc\ulate.l
B: Exanples a l the ef fccts af hetetatopl . thernatcondi t ian inE st i i lu la t ian an the Rrt te f lex. Each t t . .erepresents the aveLrye of lO sr..essive full ||ave rectiiiedresponses recoftie.t .lu.ing a oDe ninute pe.iod. Fot ea.hexpeinental sequen.e, tl1e tcmponlevalutbn is shawn frcn)back ta frcnt, the 2 ninute candi!laninE petiod 01cltcate.l byaraws)bei.g in the thld and foutth nrhutes.
Ihe nanlaintul tenpcatLrc (11"C, len did not nadiiy theteflex. By &nttast, thc painiul tempetaturcs (15,16,47"C)depresse.J the rcfk:x duting dnd .itet thc applicathn ot thecon.litbninB stinuli. Ibe exrer/ o/ rre{, dcpresiDrs tr.rrte tnperal D te depe ncl e nt.
Nate the lon\Clutatlan (araDn.l 1a nrnutc| of the inhtbnotrpost . . f fects fa l lowiDE the appl i .a t ian af the h i rhcsttenpeatu.e (17, ti1hr.
B
on 17 Septem
ber 2018 by guest. Protected by copyright.
http://aim.bm
j.com/
Acupunct M
ed: first published as 10.1136/aim.9.2.47 on 1 January 1991. D
ownloaded from

the transmission of nociceptive information.The fol lowing question arises: are the inhibitory
mechanisms purely spinal ot alternatively, do theyinvolve supraspinal structures? To answer this, wecompared the effects on the Rir ref lex in the riEhtleg, of nociceptive condit ioning stimuli applied tothe 4th and 5th f ingers of the Jeft hand in normalsubjects and tetrapleBic patients with Iesions oftraumatic origin at the C5, C6 or C7 level (RobyBrami e t a / . , 19B7) .In the normal subjects, as previously, the painful
condit ioning stimuli caused a strong depression ofboth the Rr reflex and the associated pain. Bycontrast, in the telrapJegic patients nociceptivestimulation of the same cutaneous terri tories, which,being in the CB and Tl dermatomes, were cl inicalJyunaffected by the spinal lesion, did not produce anydepression of the Rrrr ref lex recorded in thecontralateral leg. These results demonstrate that inman, as in animals, Lhe inhibitory e{fects tr iggeredby heterotopic nociceptive st imuli are most l ikelysustained by a loop which includes supraspinalstrucrures._lhese
results also lead to a subsidiary question : isi t possible to identify, or at least localise, thesesupraspinal strucLures? A tentative answel has beenobtained by observations made on patienLs withcerebral lesions causing contralateral hemianalgesia (De Bfoucker et a/.,1990). These werepatients with either a unilateral thalamic lesion(identif ied and delineated by a CT scan), or a lesionof the retro-ol ivary part of the medulla (Wallenberg'ssyndrome).In the forraer Broup, the Rrr reflex on the normal
side was strongly depressed, as in norrnal subjects,by nociceptive condit ioning stimuli applied to theaffected side which were not felt as painful. Bycontrast, in the patients with Wallenberg's syndromeno inhibit ions were observed when the nociceplivecondit ioning stimuli were applied to the affectedside, whereas i{ these stimuli were applied to thenormal side they tr ig8ered inhibitory effects andaftereffects very similar to those seen in normalsubjects.These results show that in humans, thalamic
structures and consequently spino-thalamicpathways are nol involved in DNIC whereasbrainstem probably fel icular structures seem toplay a key role in these phenomena.Finally, i t has been shown that naloxone can block
these inhibitory phenomena which are tr ig8ered bynociceptive st imulation (W;l ler er a/.,1990). Theremust therefore be at least one opioidergic l ink in Lhespino bulbo spinal loop which is implicated in theseinhibitory processes.Thus, the fol lowing fealures are shared by the Rll l
ref lex and associated painfuJ sensations in hunransand by dorsal horn convergent neuTones in the ratspinal cord:
l) The Rri ref lex and lhe responses of convergenlneurones to electr ical st imulation of their
cutaneous receptive f ields are similarlyinhibited by various heterotopic nociceptives t i rnu l i .The extent of the inhibit ions is directly relatedto the intensity of the condit ioning stirnulus.The inhibitions are followed by afler-effectswhich can last for several minutes.The i nh ib i on . a 'e med id led b1 a -p . robulbo-spinal loop, the ascendinS part ofwhich is composed of the spino reticular tractand synaptic relays in the brainstem. Inaddit ion, there is at least one opioderSic l inkin this loop both in the rat and in man.
) )
l )
4)
Such similari t ies al low one to conclude that theinhibitory processes observed in man and DNIC inthe rat share cor.mon mechanisms; the existence ofDNIC in humans is therefore more than l;kely.
lll. HypothesesThe data described above suggest that nociceptivestimuli, even though unquestionably perceived asbeing painflr l , activate ceftain inhibitory controlswhich originate in the brainstem.
In order to interpret the physiological meaning ofthese {indinBs one has to take into account aparadoxical property of converSent neurones. Theseunits do indeed respond to non nociceptive st imuli
f i r lrElH
' iNBA
Hypothet ica l in tetpretat ion af the p. lobal act iv i ty af a l l.anvergent neurones invalrc.l in nocicepLion at spinal andtri9ein)nal levels. At 'len", because ot the praPerties of theseneurones, such activiry would nat be neslisible and thos a basicsonaesthetic signal would be sent bwardt the brain lA). Anocicept ive facus wi l l act ivate sante canvetgent andnaciceptive specific neuranes lB), which in tutn {,i11 tnnsnit anex.itatoty signal towads suptaspinal centres (C). This willti19et DNIC (D), which will inhibit those conreryent neurarleswhi.h were nat dircctly affected by the ini al stinulus, and tllusthe back1rcund noise which constltubs Lhe basic sarnaesth-.ti.activity willbe rcduced ot abolished (E).
Marphine, eithet synenically at law.lases, intncerebrally, olin t td vent t ico lat ly b locks DNIC dnd thus restores thebackgrcund noise (f). At high synemn: doses ar lntdhe.ally,noryh)ne b locks the spinal . ransniss ion ot nocl .ept l telnformation and therchre futthet teduces lhe cantran lC).Electriel stinulation of same zones in the bninsLeni blo.k, tl)eactivities of the whole neuonal populatian and therclote elicltsstron' anal]sla (H). (Frcm Le Earc and Vlllanueva,l988).
53
on 17 Septem
ber 2018 by guest. Protected by copyright.
http://aim.bm
j.com/
Acupunct M
ed: first published as 10.1136/aim.9.2.47 on 1 January 1991. D
ownloaded from

(e.g. rubbing or hair movernents) and thus arerandomly bul permanently beinS activated by al i thesomaesthetic sl imuli arisinS from the environrnenl._lhe
resull ing basic somaesthetic activity wil l hetransmil led towafds hi: lhef centres and coLrJdconsli lLrte a "background noise", from which thebrain's centres could extract a signif icanllynociceptive message only with diff iculty (Figure 8A).D \ l ( , a L J d , n n \ t u t e ' l i l t e r b r n r i ' h . r p , , i ' . ,nociceptive signal would be extracted from thisbasic somaestheLic activity (Le Bars et a/., 1979b;1986 ; 1 9BB) .Indeed, when a noxjous focus appears in a region
of the body, both converSent and nociceptivcspecil ic neurones are activated (F/gure BB) and sendan excitatory signal lowards hi: lher centres (F/gureBO. The signal secondari ly activates DNIC (F/gureBD wh ich w i l l i nh ib i t a l l t hose sp ina l andtrigeminal conver8ent neutones whjch werc notdirectly concerned $,i lh the init ial st imulus (F/gurcBO. Such a rnechanism improves the "signal tonoise ratio" by increasing the contrast between theactivity of the segmental focus of excited ncuronesand the si lence of the remaining population. Thedestination of such a "pjcture", i ts recognit ion, andits processing by cerebral c-ontres remain unsolvedproblems. As an hypothesis, one can proposc thalthe brain is able to fecognise this picturc and thiswould inter Ihat DNIC constitute not only a f i l lerwhich al lorvs the extraction of the signal lor pain,bul also - and this is perhaps more imponanl - anampJif ier in lhe transmission system which increasesthe potential alarm function oi the nociceptivesignals. DurinS cl inical pain therefore, i t isconceivable that the global message sent byconverSent neurones becomes polymorphic, of evencomplex, and that a lar€lc variety ol pain syndromescould come about in this way.AccordinS to the above model, hypo- or hyper-
algesic effects could resuJt from manipulationsrn,hich affect excitatory and/or inhibitoryphenomena. An intcnsif ical ion ol the contrast effectshould faci l i tate the rcco€lni l ion oi nociceptivesiEnals b), hiSher centres ; consislenl wilh this, in amodel of chronic pain, thc arihri l ic rat, hyperal8esic phenomena occur together with anexacefbation of DNIC. ConverseJy, a reduction ofthe contrast should hinder the recoSnit ion of thesignals and thus produce an analSesic effect. lnorder to verify this hypothcsis, one can test whetheror not lhe reference analgesic drug, morphine, canproduce a recovery in thc somaeslhetic backgroundactivity which would normally be depressed byDNIC. In fact, DNIC have been found to beextremely sensit ive to the adr. inistration oT lowdoses of morphlrre (Le Bars el a/.,19B1 c).
In view of the polenlial ly impoftant roJe ofconVergent neurones in nociception, a second drrcctimp ication of the model is thaL there are tnteracttvephenomena betwcen nociceptive s;gnals iromromore , r r . d . oJ .hc bod r .nd he r r , p oe t ( cpn pd tn .with dist inct topographical or. lgins. Evidence for
such intefactions in anir.als have been repofted, butmore convincing observalions have been made inhumans with the common observation that "onep " i n , a n m t - k d n o l h c t ' . r o t , - 1 t J n o \ d l d r ' F
number of popular medical practices for rel ievingpain have becn based on this principle. Thesee.rlpir ical observations have been confirmed Lrndercondit ions of scicnti f ic objectivity and suchphenomena are often designated as "counter-itrilatian" or "caunter stimulatiou" (see feferences inLe Bars et a/.,1984; Le Bars and Vil lanveva,l9BB).DNIC pfobably represent, at least in part, thefunctjonal substrate for these observations; theexperiments in human described above confirm thishypolhesis.The question arises as to what m€chanisms
underl ie the hypoalgesic effects of transculaneouselectr ical nerve stimuial ion (TENS) ano etecrro-acupuncture, and whether they mighL sharecommon neuronal SuDstrales.As stated in the introduclion, i t seems possible to
disl inguish between two means of producinghypoalgesia by somatic st irnulation. TENS can beeifective when applied at high frequencies andintensjt ies below the pain threshold with ther-esLrlt inB pain rei ief being localised and oftenlimited to the stirnulated seSment (Andersson,1979).The eff iciency of this procedure can be explained byseSmental inhibitory pfocesses tr iggered by lowthreshold afferents, and lhe "gate control" theory hasbeen proposed as i ts neuronal basis (Melzack andWall,1965; Wall, l978). In some cases, acupunctureprobably activates such processes, especial ly whenthe needles are applied in the vicinity of the pain.
However, the mechanisms underlying TENS andacupunctLrre at sites with or without a segmentalreJationship to the soufce of pain are st i l l conlrover-sial and, in any case, cannol be explained with asinglc theory (Satfan and Coldstein,1973; Nathanand Rud8e,1974; Andersson and Holmgren,l975;jeans,l979). In this respect, i l has been shown thatstronger analgesic eflccts can be el icited wilh TENS,by using a cri t ical leveJ of st irnulation whichproduces an unpleasant, but not quite painfulsensation (Andersson, 1979; Melzack,1984). Thisreinlorces the idea that A6-f ibre activation isimportant in the production of analgesia by sornaticstimulation, especial iy i f one takes account of theiact that TENS is more effective and inducescxtraseBmental antinociceptive effects when 46libres are activated (ChunB et a/.,1984). We havealready mentioned that lhe pain rel ief produced byacupLrncture can have a widespread distr ibutionwhen the stimulation is strong enough to induce thefeeling of "De Chl", which is an unpleasantsensation that is probably related to activity in thinperipheral f ibres. FLrrthermore, i t has been shownlhat antinociceptive effects el icjted by acupunclureare stronger when the stimulation intensit ies aresuff icient to recruit A5 and C-fibres (Chen eta/.,1 981 ; Kawakita and Funakoshi, l 9B2).
on 17 Septem
ber 2018 by guest. Protected by copyright.
http://aim.bm
j.com/
Acupunct M
ed: first published as 10.1136/aim.9.2.47 on 1 January 1991. D
ownloaded from

3
We have proposed th.rt DNIC nlay well fonr thencural basis lor the pain-rel ieving effects ol thcseproceclures in which afferenl A5 or 46- and C f ibresplay an imporlant fo e. This suSScstion is slfonglysupported by lhe i i fdjng thal re8ardlcss of whelherlnh bit ions oi lr i€icminal converElcnt neurones arctr iggered by imrrrcfsion of a l) indpaw in 50"C watcr,or by manual st imu ation oi lhc ST.36 'Zusanli"
point on Lhe l ibial js irnterior muscle, lhey arc. L l L . , l , | . r ' l " r n r * r t , / , 8 r i , 1 . u ' b \ . \ . 1 . - i ,
AcknowledgementsIhis \/ork lvas suppofted by l ' lnslhut Nation.rl de aSant6 et de a Rccherche M-6dica e (INSERM), laDirection des Recherches el Etudes Te.hr] iqucs(DRET), and La Fondation pour la RecherchcM6d cale. The alr lhols are very Sratelul to Drs.Besson, Busse, Cadden, C.rlvino, Cesaro, Chaouch,ChitoLrr, De Broucker, Dickenson, Kraus,Pcschanski, Rivot, and Roby'l lrami ior their col l l l lbution to sornc aspects oI lhis wofl(, to Df. Caddcnior advice ir lhe preparation ol thc rnanuscripl, loMiss M.Ciryla lor thc typing, and lo Mr.E.Dehaussefor drawings and photogrnphy.
Daniel Le BarsLuis Villanueva
Didicr Bouhassiru/NS'RM U. 16l, 2 rLte cl'Al.sia
75011, Paris
J.C. willerC. H.U. Piti4-Salpdlriire
Laborilai re dc NeLt rolthysi ol ogi c91 Baulevad de I'116pi!.tl
75013 Pais
l . A d r a n s e n I , G y b e s l , H a n d $ , e r k e r l l O , V a n r r e e s l L l 9 t i J lRcsponse proper l e i o l th n myc inr te({ t \6) t bres In hunars(in Nves /.Neu'ophfsioi. .,19: '22Afdefsson SA 1r9791 Pain conlnr l by seisory st inru lat ion l f :Eonica J, L ebeskind jC and Albe- l - "$ard, €ds A. /v , r .es i ,P,)in Research and fltefri])v L Raven Prcss, New Yo'k: j69-
l t5Afdcr$or SA, Ho m8rcn t (1975) Of a.uP!ncturcanJlgesi . r . id th€ nrcchrr isnr of ta i i . Aner i .an launat o jCirh-.se,V.di.irc. l: lr I -l.lB n8 Z, V lanucv. i L , Le Bars D (1990) Acl f lnc lufe an. ld l f !se noxious inhib iLory contro ls : rao\on€ re! -pr b le. lepre$ion o i a.L iv l l ie5 o l l r i8ern in. l co rvefSenL neuron-"s.Neufoi.i.rr.e :lt: 8{l'l I 8Bolhrss i ra D, Lc Ears D, Vi l la i t - "va L 11987) lc terotof ici rcL i !a l ion o l A6 and C l ibres l r 88eE jnhlb i t on o i t r Eemi i r land sp nal corveBenl fe!rones if the tut. / Pr),'toi.lloDdl.t89: lA1 17C a d d c n S w , V l l r f u e l a L , C h t o u ' I � L e 8 a t r D ( l 9 8 l )Dcpression oi activitics oi dors.rl horn .ofveqent nc! onesbr- propr iospi .a l nc.hanisnrs l r iEg-" fed by nox o!s inputs :compar ison l r i th d f i !se nor io!s i ih ib tor ! contro ls l lN lC).
Cal ino B, V l .n leva L, Lc Bars D (198.1) Ihe heterotoPiccfi.'cts of v s.errl paii : bch.vilrlr3l an.l e ectroPhysio 08i ' i l Jpprc.ches in l rc i \ t . f ' . i r 2A:261 71Chen I , Taig i . Yan I { l98 l r Anr vs i5 o l a l lercnt f ibrcs ior theina ses of ac lpunct ! 'e .nalgesia. Kcxuc Totub.o. 26:
9 ChlnE lM, lee KH, Hor \ ' , [ndo K, \ / l i5 WD 1198,1)Frc loA i f l renc nB per iphtra ncrve n lmL . l o f produccdnhbi to i o l p f nratc spnotr r . f r i . 1r3.1 .e s . Paln. /91
277-. ) il0 De Bro! .ker T, Ccsaro P, Wi l l€ f lC, Le BrF D 1 l990) Di f i lse
Noxious l .h ib tory Contro s lDNlCl i . man in!o lenrenl orn 5pi fo r€t icu ar t rn. t l l .ah 1 I J :12) l )1
I I DePartner t o i l ih fs o lo8l . ( i r in rv le( c . r Col le8e,Ch.ngcl runi ( l97rr Th. inh ib t ion e i lec l an. l lhe modc ofact ion o i e - " . l roa. !puncture upon d is .h i r rges f rom thc painseis l ive .e s in spina t r Ben, ina nu. le !s . 5. i .5nr . l6 i 20:.+85 501
2 ) ick-" ison A l , Le Bars D ( i l l l l l Dt t lse rox iou! inh ibto.o i t ro ls iDNIC) in lo ! - " l f igenr lnothalan c Jn spinolharnr c ncurofes in Lhe r . l Exp. Bta in Res. 19:1 i1-81)
l l .Dic le.so. AH, Le Ba6 D, Besso. JM ( lg l l {Ja l Df f !scio\ors inhibtory cor t ro ls 1 l )Nrcr . EI fe. ts o i t rgcminJlf ! . - " !s ca lda is rcar ofes ) lhe r r l bra n A€in Res. 2r0:291 l0 i
1.1. t l rb fer R, Bci tc E ( l !76) l 'er iph-"ra rcuu .orenles ores. .pc bcravior i r rhe56 nronle. ! to iox o ls he. l lpPl edto t rc IJce. In : Bon.a lJ . A bc rcssard ) , eas. Adr?r . .5 t tPair Rpsert.h tDd Thctapr / Rrlen Pfer, Ne$, York: 5560
l5 Fe. ls l lL , Bcson lM i l !8€ l eds. "Pain nodr l . t ian.Ptogrc$ in Eait R."seatch,77, Elseuer, Annl{dnm
l 6 . F t z g c r a l d M ( 1 9 8 2 ) T h e . o f t r a l a t c a l r p r l l o l h - o d o r s ahonr oi thc spln.l cord in lhe deccrcbrate sr) na faL. Errl,rRas z l t i :275-87
l7.Ceh, .n KD, Yezi€Ek RP; Gicalcr J f CJ, Wi s WD (19El lihb i tory rccc l r t ive Iek ls ot pf imat€ spinotra rn ic l i r . l
c-"lls. / Ncu.opirrtol ?6: 1.Jil9 25l l l .Gybcls l , f . fdwerker lO, Van ccs I (1979) Conrpar ison
bellrce r the d scharSes oi hlman noc (eptile nerve libresind tlre sfbl,"cl! ral n8 of h s sc rsatioi. I.Ph'"siol.iLand).2t2: | 9 J-2116
l9.H.r lS, ler€nius L (1982) Neufochen ca bal rs ori.upf ncrurc .na 8csi.i. Arnu Rer.Phtnnaol.Toxn:.i 22:193 220
20.1|nssor i i ' , Lkblom A, Thonr$o r M, f je ier B ( l r l8 [ , )ln i l le ,ce o i i . oxone on rc l c f o f . ru le oro- ia. ia p! n byl . fsc! ra )eous e e. t r .a ncr !c ' n imt . l o . (TEN5l o i! ibraLion. / ' . rnr . 2 t : l2 l 9
I t rc .o! /s- . . l DNIC nedi . te . l i t lh ib i ta t r et te. t t an thc
a.ti,itic\ r,i tt ieenhn al .anrL\ Nent netonet baloe.nd snet ttlc
echnhint.Lian ai nabxane. Nole th-p patallel n.hrc at the tua. :LrvLs. t r l the reco|crr . to ! . . ] J0 4(rntn.tilled .n.|es: D-.edliry "Zuihlr" palntl
Optn.ttu'les: innosio, oi hildpas in naxkia hot \ratct.
'cr ,Euk 5 tbt .l-"talk oi thL' e\petinental sat up (tan1 Bing ct
55
on 17 Septem
ber 2018 by guest. Protected by copyright.
http://aim.bm
j.com/
Acupunct M
ed: first published as 10.1136/aim.9.2.47 on 1 January 1991. D
ownloaded from

21. He L (1987) l ivo lvement o i endogenols opio id pept ides inacupuncture analgesia. Pain. l1 :93 121
22.Huang Y WanB 0, Zheng J, L i D, Xie C (1986) AnalSesiceffects of several modes of electroacupunclure in monkeysand thei r .eve6al by naloxone. In: Zhang X and Chans Hleds. Reiear.h on Acupuncture, Moxibustion, anclAcupuncturc Anesthesia.Science Pres, Beijing: 60 9
23. Jeans ME ( l979) Rel ie f of chronic pain by br iet , in tense t ran-scutaneous elecrrical stimularion a doub e-blind study. 1n:Bonica ll, Liebeskind jC and Albe-Fessard D. A..tvances inPain Research and lhetapy .?. Raven Press, New York: 601
24. l ianS Z, Ye Q, Shen Y Zh! F, Tang S, L iang N, Z€nS X ( l978)EffecG of naloxone on experimental acupuncture :nalSesiaevnlunted by sen5ory decisian rheaty. Acta Zool.Sin. 24: 1-t 0
25.Kawaki ta K, Funakoshi M (1982) Suppresion ot the jawopeninS rei ex by conditioning A-deka fibfe stimulation ande ectroacupuncrure in the raL Exp.Neurol. 78:451-5.
26. Lamotte RH, Campbell JN (1978) Comparison of respons,"sof warm and nociceptive C fibre afferent in monkey withhuman judgements of thermal pain. J.Neurcphysial. 11:509 28
27.1e Bars D (1988) Serolonin and pain. In : Osborne N andH.rmon M, eds. Neuronal Serotorlr. Wiley, Chichesten I Zl-229
28. Le Ba15 D, Villanueva L (1988) Elecirophysiologicalevidence for rhe activation of descendinB inhibitory contro,by nocjceptive afferent prthways. PrcEress in BrainReseat.h : "Pain Ma.lulation". 77t 275 99
29. Le Bars D, Wi l ler lC ( l991) Physio loSy of pai . sensat ion. In :Pviars and F.Bonner eds. Pain in the periopeative perb.l.Blackwell, Oxford {in press)
l0.Le Bars D, Dickenson AH, Besson jM ( l9Z9a) Di f f !senoxio!s inhibitory contro s (DNIC) | l. Eftects on do$a homconve€ent ne!r ones in the tat . Pain.6:283-)04
l l .Le Ba6 D, Dickenson AH, Besson lM (1979b) Di f fusenoxious inhibitory controls (DNIC) : ll. Lack of effect on nonconver8ent neufones, supraspinal involvement andtheoret ica impl icat ions. Pain. 6:105 27
32. Le Bars D, Chi lo l r D, C ot AM ( l98la) The encodin8 ofthermal srimu i by dlff!se noxious inhibitory controls(DNIC). Aral, Rer. -?10:19.1 9
ll. Le BaE D, Ch tour D, Kraus E, Dickenson AH, Besson JM{1981b) Effect of ialoxone upon diffuse noxious inhibjtorycontrols (DNIC) in the rat. arcin Res. 201:387 102
34.1e Bars D, Chi tour D, Kraus E, Clot AM, Dickeison AH,Besson lM (1981c) The effect oi synemic morphine lponDi f iuse Noxio ls lnh ib i tory Contro ls (DNIC) in the rat :evidence for a IiftinS of .ertain descendinB inhibitorycontrols ofdo6al horn converSent neurones. Sfain Res. 2r5:257 74
35.1e Bi r rs D, Dickefson AH, Be$on lM (1983) Opiateanalgesia and des.ending control systems. Adv. PnlnRes.Ther. t )41 72
36.1e Bars D, Calv ino B, Vi l lanueva L, Cadden S (198a)PhysioloSlca approaches to counter'irritation phenomena.In:Tricklebank MD, Cuaon C eds. Stress-irduced a'alsesia.Wi ley, Chichener: 67 101
37 Le Bru D, Dickenson AH, Besson lM, V l lanueva L (1986)Aspecls of sensory processinS through conveGent neurons.ln: Yaksh TL etl. Spinal affercnt pfo.esslr8. Plenum.NewYork:467 504
l8.Lee HK, Ch!ns JM, Wi l l is WD (1985) Inhib i t ion o l pr imarespinothalamic tracl cells by TENS. /.NelrosurA. 2:276-87
19.Lundeber8 T i1985) Naloxone does not reveGe the painreducing effect of vibratory stim!lation. A.ta araesth. s.and.291212-6
40. Macdonald AJR (1989) Acupunct!rc analSesia and therapy.ln: Wall PD and Melzack R eds. Ie.{tbook o/ Pain. ChurchillL iv ingstone, EdinbuGh: 906J 9
4l .Mann F (1974) Acupuncture analgesia, repor t of 100exped ments. Af../.Araenh. .,/6: 3 6I -4
42. Me zack R (1984) Acuplnclure and related forms of folk
medicine. In: Wall PD and Melzack R eds. Iextbook ofPair.Churchi l l L i ! insstone, Edinbursh: 691 -700
4l .Melzack R, Wal l PD (1965) Pain mechanisms:a newrheoty. Science. | 50 : 97 1 -9
44 Morton CR, Maisch I, Zimmerniann M (1987) Difiusenoxious inhibitory controls of lumbar spinal neurons involvea supraspinal loop in the cat. Brain Res. 410:347-52
45 Nathan PW RudSe P (1974) Testjng the gate conlrol lheoryof pain in man. ./.Neuro/oA, Neurosur9ery ancl Psychiatry,37:1366 72
46.Roby Erami A, Busse B, Wi ler JC, Le Bars D (1987) Anelectrophysiological investigalion into the painielievinge{fects of heterotopic nociceptive stim!li : possibleinvolvemenl of a suplaspinal loop. Afain- 110: 69 80
47.Satran R, Coldste in MN (1971) Pain percepr ion : modi f ication and threshold of intolerance and conical potentials bycutaneous stimulalion. Sclence. 1 80. 1 201 -2
48. Sjolund BH (1985) Peripheral nerve supprerrion of Cjjbre'evoked flexion reflex in rats. Pan l:parameteB ofcontinuousslim!laiion. /.NeufosurA. 6l: 6l2 6
49 Sjd l !nd BH, Er ikson MBE (1979)The inf luenceol naloxoneon analSesia produced by peripheral conditioninEsiimulation. 8rrl, fies. ZJ: 295-301
s0.Torebjork HE, Lamotte RH, Robinson C {1984) Peripheraneural cotre ates of maSnitude ofcutaneo!s pain and hypeFalSesia : simultan€o!s recordin8s in humans of sensoryjudsements of pain and evoked rcsponses in nociceptorswith C fibrcs. /. Neuropf,ys io|. 5l: 325 9
sl. Van Hees J, Cybels JM (l972) Pain rclated to single afferentC{ibres trom human skin. Erain Res. 18t 397-4oO
52.Vi l lanueva L, Le Bars D (1985) The encoding of lhermastimuli applied ro ihe tail of rhe rat by loweing theexcitabilily of trigeminal convergent ne!rones. Ara- Rei.330:245 51
53. Villanueva L, Cadden 5W Le Bars D (1984a) Evidence thatdiffuse noxious inhibitory controls (DNIC) are mediated by afinal post synaptic inhibitory mechanism. Brain Res. 298:67,71
54 Vi l lanueva L, Cadden SW Le Ban D (198ab) Di f fusenoxio!s inhibitory controls (DNIC): evidence for posl-synapl ic nh b i t lon of t r igeminal n!c leu, caudal isconverSenr neurones. Efain Res. 321:165 A
55.Vi l lanueva L, Peschanski M, Calv ino B, Le Ba6 D (1985a)Ascending pathways in the spinalcord involv€d in triSgerinSof difiuse noxious inhibitory controk (DNIC) in the rat-J.Neurcphysial. s5. 34-5s
56. v i l lan leva L, Chi loLrr D, Le Bars D (1986b) Involvement o ithe do6olatera l funiculus in the descending spjnalproiecrions responsible for diffuse noxious jnhibitory, nnrro l - i , rp dr . / .v-ufoph) , io l .56. | |85 q5
57.Wal PD (1978)The gate contro l theory of pain mechanisms.q r a i n . 1 0 1 : 1 1 8
58.Wi l ler JC, Roby A, Boulu P, Boufeau F (1982) Comparat iveefiects of electroac!puncture and transcuianeo!s stimulationon rhe human bl ink ref lex. Pain.14:267-78
59.Wi ler jC, Roby A, Le Bars D (1984) Psychophysical andelectrophysiological approaches to the pain rclieving effectof heterotopic nociceptive slimuli- Brain.107:1095 102
60.Wi l ler Jc, Barranquero A, Kahn Mt Sal l iere D (1987) Painin sciatica depresses lower limb cuianeous reflexes to suralneNe sr im u lafjon. l. Neurol Neurosutg.Psychiatry. 50.1-5
6l.Wil er lC, De Broucker T, Le Bar, D (1989) The encoding ofnociceptive thermal stimuli by Diffuse Noxious InhibitoryControls (DNIC) in man. /.Neurcphysiol. 62:1028-38
62.Wi l lerJC, De BrcuckerT, Le Ba6 D (1990) Di f fuse NoxiousInhibitory Controk (DNIC) in man : involvement of anoplu op'Er l'k . ru,./.Phltndcol. 182: 147 5\
63.Woolf CJ, Mitchell D, Bafrett CD (1980) Antinocic€ptiveeffect ol peripheral segmental electrical stimulation in th€'ar . Pain.8:237 52
56
on 17 Septem
ber 2018 by guest. Protected by copyright.
http://aim.bm
j.com/
Acupunct M
ed: first published as 10.1136/aim.9.2.47 on 1 January 1991. D
ownloaded from


















