
Differential Effect of Hypolipidemic Drugs on Lipoprotein...
20
Differential Effect of Hypolipidemic Drugs on Lipoprotein-Associated Phospholipase A 2 Vasilios G. Saougos, Afroditi P. Tambaki, Mihalis Kalogirou, Michael Kostapanos, Irene F. Gazi, Robert L. Wolfert, Moses Elisaf, Alexandros D. Tselepis Objective—Lipoprotein-associated phospholipase A 2 (Lp-PLA 2 ) is a predictor for incident atherosclerotic disease. We investigated the effect of 3 hypolipidemic drugs that exert their action through different mechanisms on plasma and lipoprotein-associated Lp-PLA 2 activity and mass. Methods and Results—In 50 patients with Type IIA dyslipidemia were administered rosuvastatin (10 mg daily), whereas in 50 Type IIA dyslipidemic patients exhibiting intolerance to previous statin therapy were administered ezetimibe as monotherapy (10 mg daily). Fifty patients with Type IV dyslipidemia were given micronised fenofibrate (200 mg daily). Low- and high-density lipoprotein (LDL and HDL, respectively) subclass analysis was performed electrophoretically, whereas lipoprotein subfractions were isolated by ultracentrifugation. Ezetimibe reduced plasma Lp-PLA 2 activity and mass attributable to the reduction in plasma levels of all LDL subfractions. Rosuvastatin reduced enzyme activity and mass because of the decrease in plasma levels of all LDL subfractions and especially the Lp-PLA 2 on dense LDL subfraction (LDL-5). Fenofibrate preferentially reduced the Lp-PLA 2 activity and mass associated with the VLDLIDL and LDL-5 subfractions. Among studied drugs only fenofibrate increased HDL-associated Lp-PLA 2 (HDL-Lp-PLA 2 ) activity and mass attributable to a preferential increase in Lp-PLA 2 associated with the HDL-3c subfraction. Conclusion—Ezetimibe, rosuvastatin, and fenofibrate reduce Lp-PLA 2 activity and mass associated with the atherogenic apoB-lipoproteins. Furthermore, fenofibrate improves the enzyme specific activity on apoB-lipoproteins and induces the HDL-Lp-PLA 2 . The clinical implications of these effects remain to be established. (Arterioscler Thromb Vasc Biol. 2007;27:2236-2243.) Key Words: hyperlipidemia lipoproteins PAF-acetylhydrolase Lp-PLA 2 ezetimibe fenofibrate rosuvastatin P latelet-activating factor (PAF) acetylhydrolase exhibits a Ca 2 -independent phospholipase A 2 activity and degrades PAF and oxidized phospholipids by catalyzing the hydrolysis of the ester bond at the sn-2 position. 1 PAF-acetylhydrolase in plasma is complexed to lipoproteins 2 ; thus it is also referred as lipoprotein-associated phospholipase A 2 (Lp-PLA 2 ). 3 Lp- PLA 2 is associated mainly with apolipoprotein B (apoB)- containing lipoproteins and primarily with low-density li- poprotein (LDL), whereas a small proportion of circulating enzyme activity is also associated with high-density lipopro- tein (HDL). 1,2 he majority of the LDL-associated Lp-PLA 2 activity is bound to the atherogenic small-dense LDL (sdLDL) particles, 2,4,5 and we recently showed that the enzyme activity is a marker of sdLDL particles in plasma. 6 Lp-PLA 2 is principally produced by hematopoietic cells including monocytes-macrophages. 7,8 Lp-PLA 2 has been identified in atherosclerotic plaques 9 ; however, its role in atherosclerosis is still under investigation. In this regard, it is suggested that this enzyme might have an antiinflammatory role because it degrades and inactivates proinflammatory PAF and oxidized phospholipids 10,11 ; other studies showed that Lp-PLA 2 may have a proinflammatory and proathero- genic role 12 because it generates lysophosphatidylcholine (lysoPC) 3,13 and bioactive oxidized fatty residues. 3 Data from large White population studies demonstrated an independent association between plasma Lp-PLA 2 with cardio- vascular disease (CVD) risk. In this regard a recent metaanalysis showed that Lp-PLA 2 is significantly associated with CVD, and the risk estimate appears to be relatively unaffected by adjust- ment for conventional CVD risk factors. 14 In contrast to total plasma enzyme, which mainly represents the LDL-associated Lp-PLA 2 , several lines of evidence suggest that HDL-associated Lp-PLA 2 activity, although at low levels, contributes to the antiatherogenic effects of this lipoprotein. 1 However, the clinical value of HDL-associated Lp-PLA 2 as a potent inhibitor of the atherosclerotic process remains to be established. Among the various agents used to treat patients with CVD, only drugs that affect lipid metabolism can significantly Original received September 19, 2006; final version accepted July 4, 2007. From the Department of Internal Medicine, Medical School (V.G.S., M.Ka., M.Ko., I.F.G., M.E.), and the Laboratory of Biochemistry, Department of Chemistry (A.P.T., A.D.T.), University of Ioannina, Greece; and dia Dexus Inc (R.L.W.), South San Francisco, Calif. Correspondence to Prof Alexandros D. Tselepis, Laboratory of Biochemistry, Department of Chemistry, University of Ioannina, 45110 Ioannina, Greece. E-mail [email protected] © 2007 American Heart Association, Inc. Arterioscler Thromb Vasc Biol. is available at http://atvb.ahajournals.org DOI: 10.1161/ATVBAHA.107.147280 2236 by guest on July 13, 2018 http://atvb.ahajournals.org/ Downloaded from by guest on July 13, 2018 http://atvb.ahajournals.org/ Downloaded from by guest on July 13, 2018 http://atvb.ahajournals.org/ Downloaded from by guest on July 13, 2018 http://atvb.ahajournals.org/ Downloaded from by guest on July 13, 2018 http://atvb.ahajournals.org/ Downloaded from by guest on July 13, 2018 http://atvb.ahajournals.org/ Downloaded from by guest on July 13, 2018 http://atvb.ahajournals.org/ Downloaded from by guest on July 13, 2018 http://atvb.ahajournals.org/ Downloaded from by guest on July 13, 2018 http://atvb.ahajournals.org/ Downloaded from by guest on July 13, 2018 http://atvb.ahajournals.org/ Downloaded from by guest on July 13, 2018 http://atvb.ahajournals.org/ Downloaded from by guest on July 13, 2018 http://atvb.ahajournals.org/ Downloaded from by guest on July 13, 2018 http://atvb.ahajournals.org/ Downloaded from
Transcript of Differential Effect of Hypolipidemic Drugs on Lipoprotein...

Differential Effect of Hypolipidemic Drugs onLipoprotein-Associated Phospholipase A2
Vasilios G. Saougos, Afroditi P. Tambaki, Mihalis Kalogirou, Michael Kostapanos, Irene F. Gazi,Robert L. Wolfert, Moses Elisaf, Alexandros D. Tselepis
Objective—Lipoprotein-associated phospholipase A2 (Lp-PLA2) is a predictor for incident atherosclerotic disease. Weinvestigated the effect of 3 hypolipidemic drugs that exert their action through different mechanisms on plasma andlipoprotein-associated Lp-PLA2 activity and mass.
Methods and Results—In 50 patients with Type IIA dyslipidemia were administered rosuvastatin (10 mg daily), whereasin 50 Type IIA dyslipidemic patients exhibiting intolerance to previous statin therapy were administered ezetimibe asmonotherapy (10 mg daily). Fifty patients with Type IV dyslipidemia were given micronised fenofibrate (200 mg daily).Low- and high-density lipoprotein (LDL and HDL, respectively) subclass analysis was performed electrophoretically,whereas lipoprotein subfractions were isolated by ultracentrifugation. Ezetimibe reduced plasma Lp-PLA2 activity andmass attributable to the reduction in plasma levels of all LDL subfractions. Rosuvastatin reduced enzyme activity andmass because of the decrease in plasma levels of all LDL subfractions and especially the Lp-PLA2 on dense LDLsubfraction (LDL-5). Fenofibrate preferentially reduced the Lp-PLA2 activity and mass associated with the VLDL�IDLand LDL-5 subfractions. Among studied drugs only fenofibrate increased HDL-associated Lp-PLA2 (HDL-Lp-PLA2)activity and mass attributable to a preferential increase in Lp-PLA2 associated with the HDL-3c subfraction.
Conclusion—Ezetimibe, rosuvastatin, and fenofibrate reduce Lp-PLA2 activity and mass associated with the atherogenicapoB-lipoproteins. Furthermore, fenofibrate improves the enzyme specific activity on apoB-lipoproteins and induces theHDL-Lp-PLA2. The clinical implications of these effects remain to be established. (Arterioscler Thromb Vasc Biol.2007;27:2236-2243.)
Key Words: hyperlipidemia � lipoproteins � PAF-acetylhydrolase � Lp-PLA2 � ezetimibe� fenofibrate � rosuvastatin
Platelet-activating factor (PAF) acetylhydrolase exhibits aCa2�-independent phospholipase A2 activity and degrades
PAF and oxidized phospholipids by catalyzing the hydrolysisof the ester bond at the sn-2 position.1 PAF-acetylhydrolase inplasma is complexed to lipoproteins2; thus it is also referredas lipoprotein-associated phospholipase A2 (Lp-PLA2).3 Lp-PLA2 is associated mainly with apolipoprotein B (apoB)-containing lipoproteins and primarily with low-density li-poprotein (LDL), whereas a small proportion of circulatingenzyme activity is also associated with high-density lipopro-tein (HDL).1,2 �he majority of the LDL-associated Lp-PLA2
activity is bound to the atherogenic small-dense LDL(sdLDL) particles,2,4,5 and we recently showed that theenzyme activity is a marker of sdLDL particles in plasma.6
Lp-PLA2 is principally produced by hematopoietic cellsincluding monocytes-macrophages.7,8 Lp-PLA2 has beenidentified in atherosclerotic plaques9; however, its role inatherosclerosis is still under investigation. In this regard, it issuggested that this enzyme might have an antiinflammatory
role because it degrades and inactivates proinflammatoryPAF and oxidized phospholipids10,11; other studies showedthat Lp-PLA2 may have a proinflammatory and proathero-genic role12 because it generates lysophosphatidylcholine(lysoPC)3,13 and bioactive oxidized fatty residues.3
Data from large White population studies demonstrated anindependent association between plasma Lp-PLA2 with cardio-vascular disease (CVD) risk. In this regard a recent metaanalysisshowed that Lp-PLA2 is significantly associated with CVD, andthe risk estimate appears to be relatively unaffected by adjust-ment for conventional CVD risk factors.14 In contrast to totalplasma enzyme, which mainly represents the LDL-associatedLp-PLA2, several lines of evidence suggest that HDL-associatedLp-PLA2 activity, although at low levels, contributes to theantiatherogenic effects of this lipoprotein.1 However, the clinicalvalue of HDL-associated Lp-PLA2 as a potent inhibitor of theatherosclerotic process remains to be established.
Among the various agents used to treat patients with CVD,only drugs that affect lipid metabolism can significantly
Original received September 19, 2006; final version accepted July 4, 2007.From the Department of Internal Medicine, Medical School (V.G.S., M.Ka., M.Ko., I.F.G., M.E.), and the Laboratory of Biochemistry, Department
of Chemistry (A.P.T., A.D.T.), University of Ioannina, Greece; and dia Dexus Inc (R.L.W.), South San Francisco, Calif.Correspondence to Prof Alexandros D. Tselepis, Laboratory of Biochemistry, Department of Chemistry, University of Ioannina, 45110 Ioannina,
Greece. E-mail [email protected]© 2007 American Heart Association, Inc.
Arterioscler Thromb Vasc Biol. is available at http://atvb.ahajournals.org DOI: 10.1161/ATVBAHA.107.147280
2236
by guest on July 13, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on July 13, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on July 13, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on July 13, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on July 13, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on July 13, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on July 13, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on July 13, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on July 13, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on July 13, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on July 13, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on July 13, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on July 13, 2018http://atvb.ahajournals.org/
Dow
nloaded from

influence plasma Lp-PLA2.15,16 Thus several statins (atorva-statin, lovastatin, simvastatin, and fluvastatin) reduce theenzyme activity in plasma in parallel to a reduction inLDL-cholesterol levels.17–21 In contrast, pravastatin increasedplasma Lp-PLA2 activity,22 whereas other investigators sug-gested that pravastatin reduced plasma Lp-PLA2 mass.23
Fibrates reduce plasma Lp-PLA2 activity but significantlyincrease the HDL-associated Lp-PLA2 activity.16,24 Thesedata suggest that there are significant differences on the effectof various hypolipidemic drugs on Lp-PLA2 activity or massin total plasma and in lipoprotein subspecies. To providemore insights into the effect of hypolipidemic drugs onplasma Lp-PLA2, we investigated the effect of 3 agents thatexert their action through different mechanisms (rosuvastatin,ezetimibe, and fenofibrate) on Lp-PLA2 activity and mass intotal plasma and in lipoprotein subspecies in hyperlipidemicpatients.
Materials and Methods
PatientsOne hundred and fifty hyperlipidemic patients participated in thepresent study. According to their lipid levels, patients were dividedinto the following groups: (1) primary hypercholesterolemia (typeIIA dyslipidemia group, n�50, who were administered rosuvastatin),(2) type IIA dyslipidemia (n�50, who were administered ezetimibe),and (3) primary hypertriglyceridemia (type IV dyslipidemia, n�50,who were given micronized fenofibrate). Details on patient selectioncriteria, patient characteristics, and treatment are provided in theData Supplement, available online at http://atvb.ahajournals.org.
Biochemical ParametersLipoprotein subclass analysis was performed electrophoretically byuse of high-resolution 3% polyacrylamide gel tubes and the Li-poprint LDL System (Quantimetrix). Subfractionation of plasmalipoproteins by was performed by isopycnic density gradient ultra-centrifugation. Lp-PLA2 activity was measured by the trichloroaceticacid (TCA) precipitation procedure with the use of [3H]-PAF (100�mol/L final concentration) as a substrate. Lp-PLA2 mass wasdetermined by use of a dual monoclonal antibody immunoassaystandardized to recombinant Lp-PLA2 (PLAC test; diaDexus, Inc).Data are presented as mean�SD, except for Lp(a), which wasexpressed as the median and range. Statistical analysis was per-formed using SPSS 13.0 softpack. Details on the methodology usedand statistical analysis are provided in the Data Supplement, avail-able online at http://atvb.ahajournals.org.
ResultsEffect of Hypolipidemic Therapy on SerumLipid ProfileAs shown in supplemental Table I (available online athttp://atvb.ahajournals.org), ezetimibe significantly decreasedserum total cholesterol, LDL-cholesterol, non–HDL-choles-terol, and apoB levels. Furthermore, ezetimibe induced aslight but significant reduction in serum HDL-cholesterollevels whereas it did not affect apoA-I levels. Rosuvastatinsignificantly decreased serum triglycerides, total cholesterol,LDL-cholesterol, non–HDL-cholesterol, and apoB levels.The reduction in these serum lipid parameters was moreprofound compared with the ezetimibe group. Fenofibratesignificantly decreased serum levels of triglycerides, totalcholesterol, LDL-cholesterol, non–HDL-cholesterol, andapoB. Furthermore, fenofibrate induced a significant increasein the serum levels of HDL-cholesterol and apoA-I, a phe-nomenon not observed in the other 2 groups. Finally, noalterations in the baseline Lp(a) levels were induced by anylipid-lowering therapy (supplemental Table I).
Effect of Hypolipidemic Therapy onLipoprotein SubclassesType IV dyslipidemic patients had higher plasma levels ofVLDL-cholesterol and lower levels of IDL-cholesterol andbuoyant LDL-cholesterol at baseline compared with type IIAdyslipidemic patients of either the ezetimibe or the rosuvas-tatin group. Type IV dyslipidemic patients had significantlyhigher baseline levels of sdLDL-cholesterol and a higherproportion of sdLDL whereas the mean LDL size was lowercompared with type IIA patients. Treatment with eitherezetimibe or rosuvastatin significantly reduced the mass of allapoB-containing lipoprotein subclasses, with the exception ofVLDL-cholesterol, which was not reduced by ezetimibe.However, neither drug affected sdLDL proportion and meanLDL size. Fenofibrate significantly reduced VLDL-cholester-ol levels but it did not affect IDL-cholesterol or buoyantLDL-cholesterol levels. Finally, fenofibrate reduced sdLDL-cholesterol levels (and therefore the proportion of sdLDL)and increased mean LDL size (Table 1).
Type IV dyslipidemic patients had lower levels of HDL-2and HDL-3 subclasses at baseline compared with type IIA
Table 1. Effect of Hypolipidemic Drugs on ApoB- and apoA-I–Containing Lipoprotein Subclasses
Parameter
Ezetimibe (n�50) Rosuvastatin (n�50) Fenofibrate (n�50)
Baseline Posttreatment Baseline Posttreatment Baseline Posttreatment
VLDL-cholesterol, mmol/L 1.3�0.2 1.2�0.2 1.3�0.2 1.0�0.2† 1.6�0.3§ 1.2�0.3†
IDL-cholesterol, mmol/L 1.6�0.3 1.4�0.3* 1.8�0.4 1.1�0.3† 1.2�0.3§ 1.3�0.4
Buoyant LDL-cholesterol, mmol/L 3.5�0.8 2.9�0.6* 3.9�0.8 2.5�0.7‡ 2.6�1.0§ 2.6�1.0
sdLDL-cholesterol, mmol/L 0.20�0.08 0.13�0.08* 0.23�0.10 0.16�0.05* 0.52�0.30� 0.23�0.03†
sdLDL proportion, % 5�2 4�2 4�2 4�2 12�4� 6�2†
Mean LDL size, Å 269�4 269�3 266�3 267�4 258�4§ 270�3†
HDL-2 cholesterol, mmol/L 0.57�0.23 0.65�0.23 0.54�0.18 0.52�0.16 0.28�0.16§ 0.34�0.16*
HDL-3 cholesterol, mmol/L 1.2�0.3 1.1�0.2* 1.2�0.2 1.2�0.2 1.0�0.2§ 1.1�0.2†
Data represent the mean�SD values. *P�0.05, †P�0.01, and ‡P�0.001 compared with baseline values. §P�0.01 and �P�0.001compared with the baseline values of either ezetimibe or rosuvastatin group.
Saougos et al Hypolipidemic Drugs and Lp-PLA2 2237
by guest on July 13, 2018http://atvb.ahajournals.org/
Dow
nloaded from

patients of either ezetimibe or rosuvastatin group. Ezetimibesignificantly reduced the concentration of small HDL-3subclass without affecting the concentrations of the largeHDL-2 subclass. In contrast, rosuvastatin did not affect themass of either HDL subclass. Finally, fenofibrate signifi-cantly increased the mass of both HDL subclasses (Table 1).
Plasma Lp-PLA2 Activity and MassEzetimibe significantly decreased total plasma and non-HDL–Lp-PLA2 activity and mass (Table 2); however, it didnot affect the enzyme specific activity or the Lp-PLA2
activity to apoB ratio (in nmol/mg/min, 0.58�0.13 beforeversus 0.60�0.15 posttreatment) and the Lp-PLA2 mass toapoB ratio (in ng/mg, 3.75�0.51 before versus 3.83�0.98posttreatment). Rosuvastatin significantly reduced totalplasma and non-HDL Lp-PLA2 activity and mass; thesereductions were more pronounced compared with thoseinduced by ezetimibe (Table 2). Like ezetimibe, rosuvastatindid not alter the enzyme specific activity or the ratios ofLp-PLA2 activity or mass to apoB. Fenofibrate significantlyreduced total plasma and non-HDL Lp-PLA2 activity. Impor-tantly, fenofibrate induced a significant increase in the totalplasma and non-HDL enzyme specific activity (Table 2).Baseline and posttreatment corrrelations between non-HDL–Lp-PLA2 activity and mass are shown in Figure 1.
Ezetimibe induced a slight but significant reduction inHDL–Lp-PLA2 activity and mass but it did not affect theenzyme specific activity or the HDL-Lp-PLA2 activity toapoA-I ratio (in nmol/mg/min, 0.021�0.005 before versus0.021�0.006 posttreatment) and the HDL-Lp-PLA2 mass toapoA-I ratio (in ng/mg, 0.52�0.17 before versus 0.49�0.18posttreatment). By contrast rosuvastatin did not affect eitherof the above parameters.
Importantly, fenofibrate increased HDL-Lp-PLA2 activ-ity and mass but it did not affect the enzyme specificactivity (Table 2). Furthermore, fenofibrate significantlyincreased the ratios of HDL-Lp-PLA2 activity to apoA-I (innmol/mg/min, 0.019�0.008 before versus 0.030�0.009posttreatment, P�0.03) or HDL-Lp-PLA2 mass to apoA-I(in ng/mg, 0.48�0.16 before versus 0.69�0.21 posttreat-ment P�0.03).
Lp-PLA2 Activity and Mass onLipoprotein SubfractionsWe further investigated the effect of hypolipidemic drugs onthe Lp-PLA2 activity and mass associated with the apoB- and
Table 2. Effect of Hypolipidemic Drugs on Plasma and Lipoprotein-Associated Lp-PLA2 Activity, Mass, andSpecific Activity
Parameter
Ezetimibe (n�50) Rosuvastatin (n�50) Fenofibrate (n�50)
Baseline Posttreatment Baseline Posttreatment Baseline Posttreatment
Plasma Lp-PLA2 activity, nmol/mL/min 59.7�10.5 50.7�8.9* 66.0�15.4 44.6�11.5‡ 59�13.8 48.2�12.4‡
Plasma Lp-PLA2 mass, ng/mL 397�72 325�68* 449�59 320�74‡ 456�73 310.4�77.2‡
Plasma Lp-PLA2 specific activity, nmol/ng/min 0.15�0.04 0.16�0.06 0.15�0.03 0.14�0.04 0.13�0.05 0.17�0.06†
Non-HDL-Lp-PLA2 activity, nmol/mL/min 56.7�10.3 48.1�8.8* 63.5�21.4 42.2�16.7‡ 55.7�14.0 44.9�12.8‡
Non-HDL-Lp-PLA2 mass, ng/mL 324�71 264�54* 386�77 260�52.0‡ 411�65 234�73.2‡
Non-HDL-Lp-PLA2 specific activity, nmol/ng/min 0.17�0.05 0.18�0.08 0.16�0.04 0.16�0.05 0.13�0.06 0.17�0.08*
HDL-Lp-PLA2 activity, nmol/mL/min 2.95�0.82 2.57�0.64* 2.41�0.72 2.40�0.61 2.10�0.80 3.23�0.74†
HDL-Lp-PLA2 mass, ng/mL 72.7�21.5 61.1�19.8* 63.0�24.3 57.2�19.8 49.5�16.5 76.4�30.6†
HDL-Lp-PLA2 specific activity, nmol/ng/min 0.04�0.01 0.04�0.02 0.04�0.01 0.04�0.01 0.04�0.02 0.04�0.01
Data represent the mean�SD. *P�0.05, †P�0.01, and ‡P�0.001 compared with baseline values.
Figure 1. Correlation between non-HDL–Lp-PLA2 mass andactivity at baseline and after treatment with ezetimibe,rosuvastatin, or fenofibrate.
2238 Arterioscler Thromb Vasc Biol. October 2007
by guest on July 13, 2018http://atvb.ahajournals.org/
Dow
nloaded from

apoA-I-containing lipoprotein subspecies, isolated by densitygradient ultracentrifugation. Among the apoB-containing li-poproteins, Lp-PLA2 activity and mass were preferentiallyassociated with the dense LDL-5 subfraction in all patientgroups at baseline (Figure 2A and 2B), a finding that is inaccordance to our previous results.6,17,24 Ezetimibe signifi-cantly reduced the enzyme activity and mass (expressed permL of plasma) associated with all apoB-containing lipopro-tein subfractions (Figure 2A and 2B), whereas it did not affectthe enzyme activity or mass when it was expressed per mg ofprotein (data not shown).
Rosuvastatin reduced the enzyme activity and mass(expressed per mL of plasma) associated with all apoB-containing lipoprotein subfractions (Figure 2A and 2B).Remarkably, it significantly reduced the enzyme activity
and mass (expressed per mg of protein) in the dense LDL-5subfraction (Lp-PLA2 activity, nmol/mg/min: 38�13 atbaseline versus 20�9 posttreatment, P�0.03; Lp-PLA2
mass, ng/mg: 95�29 at baseline versus 50�18 posttreat-ment, P�0.03). Neither ezetimibe nor rosuvastatin af-fected the enzyme specific activity of each apoB lipopro-tein subfraction (Figure 2C). It should be noted that theLp-PLA2 specific activity at baseline in apoB-containinglipoprotein subfractions ranges from 0.3 (LDL-5) to 2.3nmol/ng/min (LDL-3) (Figure 2) whereas the non-HDLenzyme specific activity in the same patient groups islower and ranges from 0.12 to 0.22 nmol/ng/min (Table 2).However, when the enzyme specific activity in LDL-5 andLDL-3 was determined in the presence of human serumalbumin or total plasma proteins (prepared as described in
Figure 2. Bar graphs illustrating the effect of ezetimibe androsuvastatin on the Lp-PLA2 activity (A), Lp-PLA2 mass (B), andLp-PLA2 specific activity (C) associated with apoB-containinglipoprotein subspecies in patients with Type IIA dyslipidemia.Lipoprotein subspecies were fractionated by isopycnic densitygradient ultracentrifugation of plasma. Enzymatic activity wasdetermined by the trichloroacetic acid (TCA) precipitation proce-dure and enzyme mass by use of a dual monoclonal antibodyimmunoassay. Lp-PLA2 specific activity was expressed as aratio of the enzyme activity to the enzyme mass. Values repre-sent the mean�SD. *P�0.01 compared with respective baselinevalues.
0
1
2
3
4
5
6
7
8
9
Lp-P
LA2 a
ctiv
ity
(nm
ol/m
L pl
asm
a/m
in)
A
*
*
0
5
10
15
20
25
30
Lp-P
LA2 m
ass
(ng/
mL
plas
ma)
B
*
*
0
0,2
0,4
0,6
0,8
1
1,2
1,4
Spec
ific
activ
ity (n
mol
/ng/
min
)
BaselineFenofibrate
BaselineFenofibrate
BaselineFenofibrate
C
**
Figure 3. Bar graphs illustrating the effect of fenofibrate ther-apy on the Lp-PLA2 activity (A), Lp-PLA2 mass (B), and Lp-PLA2 specific activity (C) associated withVLDL�IDL and dense LDL-5 subspecies in patients with TypeIV dyslipidemia. Lipoprotein subspecies were fractionated byisopycnic density gradient ultracentrifugation of plasma. Enzy-matic activity was determined by the trichloroacetic acid(TCA) precipitation procedure and enzyme mass by use of adual monoclonal antibody immunoassay. Lp-PLA2 specificactivity was expressed as a ratio of the enzyme activity to theenzyme mass. Values represent the mean�SD. *P�0.001 and**P�0.01 compared with respective baseline values.
Saougos et al Hypolipidemic Drugs and Lp-PLA2 2239
by guest on July 13, 2018http://atvb.ahajournals.org/
Dow
nloaded from
the methods section), it was reduced in a dose-dependentmanner. Thus at an albumin or total plasma proteinconcentration of 6 g/dL a significant reduction of 50�8%in both subfractions for both treatments was observed,suggesting that plasma proteins, primarily albumin, signif-icantly affect the enzyme activity a finding, which is inaccordance with previously published results.25 Thus theLp-PLA2 specific activity in LDL-5 in the presence of 6g/dL albumin is 0.15�0.02 nmol/ng/min being similar tothat of the non-HDL–Lp-PLA2, a finding consistent withthe preferential association of the non-HDL enzyme withLDL-5.
Fenofibrate significantly reduced the Lp-PLA2 activity andmass associated with the VLDL�IDL subfraction (Figure 3Aand 3B), whereas it did not affect the enzyme-specific activityof this subfraction (Figure 3C). Fenofibrate did not affect theactivity, mass, or specific activity of the enzyme associatedwith large and intermediate LDL particles (LDL-1 to LDL-4);however, it significantly reduced the enzyme activity andmass associated with LDL-5 (Figure 3A and 3B). Further-more, fenofibrate induced a significant increase in the spe-cific activity of Lp-PLA2 associated with this subfraction(Figure 3C).
Finally, it should be noted that no detectable amounts ofLp(a) were found in any lipoprotein subfraction at baseline orafter treatment with any hypolipidemic drug. Thus it isunlikely that the alterations in the Lp-PLA2 associated withLDL subfractions induced by hypolipidemic therapy areinfluenced by changes in the Lp(a) levels and in the Lp(a)-associated Lp-PLA2.
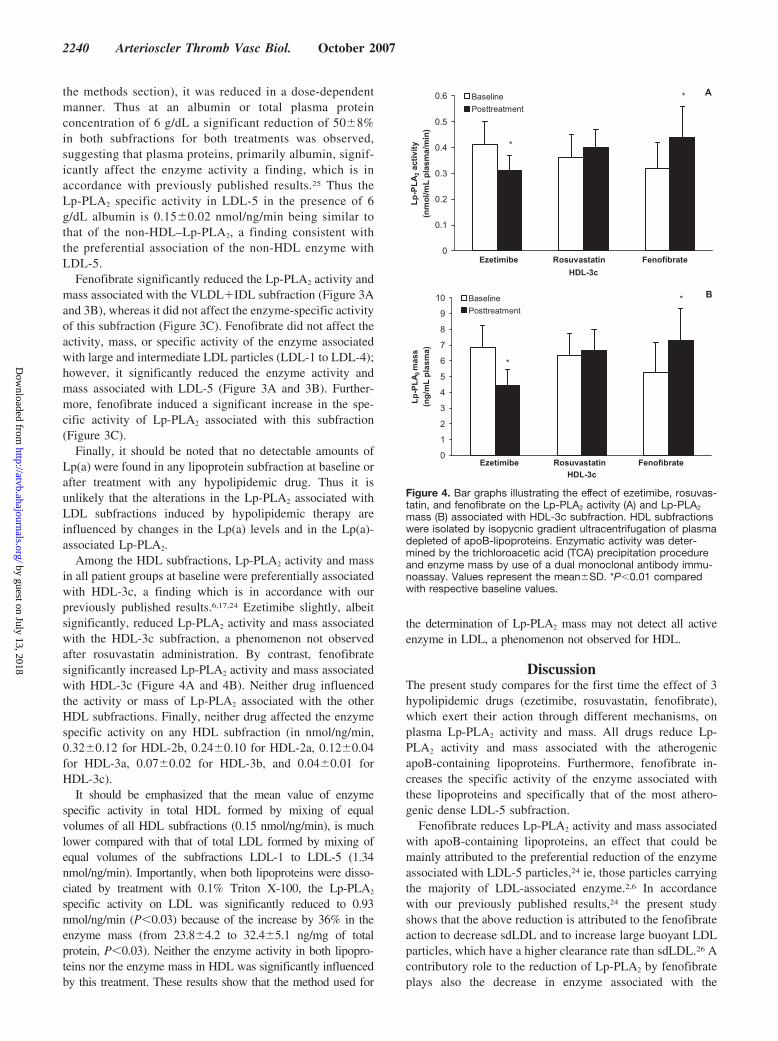
Among the HDL subfractions, Lp-PLA2 activity and massin all patient groups at baseline were preferentially associatedwith HDL-3c, a finding which is in accordance with ourpreviously published results.6,17,24 Ezetimibe slightly, albeitsignificantly, reduced Lp-PLA2 activity and mass associatedwith the HDL-3c subfraction, a phenomenon not observedafter rosuvastatin administration. By contrast, fenofibratesignificantly increased Lp-PLA2 activity and mass associatedwith HDL-3c (Figure 4A and 4B). Neither drug influencedthe activity or mass of Lp-PLA2 associated with the otherHDL subfractions. Finally, neither drug affected the enzymespecific activity on any HDL subfraction (in nmol/ng/min,0.32�0.12 for HDL-2b, 0.24�0.10 for HDL-2a, 0.12�0.04for HDL-3a, 0.07�0.02 for HDL-3b, and 0.04�0.01 forHDL-3c).
It should be emphasized that the mean value of enzymespecific activity in total HDL formed by mixing of equalvolumes of all HDL subfractions (0.15 nmol/ng/min), is muchlower compared with that of total LDL formed by mixing ofequal volumes of the subfractions LDL-1 to LDL-5 (1.34nmol/ng/min). Importantly, when both lipoproteins were disso-ciated by treatment with 0.1% Triton X-100, the Lp-PLA2
specific activity on LDL was significantly reduced to 0.93nmol/ng/min (P�0.03) because of the increase by 36% in theenzyme mass (from 23.8�4.2 to 32.4�5.1 ng/mg of totalprotein, P�0.03). Neither the enzyme activity in both lipopro-teins nor the enzyme mass in HDL was significantly influencedby this treatment. These results show that the method used for
the determination of Lp-PLA2 mass may not detect all activeenzyme in LDL, a phenomenon not observed for HDL.
DiscussionThe present study compares for the first time the effect of 3hypolipidemic drugs (ezetimibe, rosuvastatin, fenofibrate),which exert their action through different mechanisms, onplasma Lp-PLA2 activity and mass. All drugs reduce Lp-PLA2 activity and mass associated with the atherogenicapoB-containing lipoproteins. Furthermore, fenofibrate in-creases the specific activity of the enzyme associated withthese lipoproteins and specifically that of the most athero-genic dense LDL-5 subfraction.
Fenofibrate reduces Lp-PLA2 activity and mass associatedwith apoB-containing lipoproteins, an effect that could bemainly attributed to the preferential reduction of the enzymeassociated with LDL-5 particles,24 ie, those particles carryingthe majority of LDL-associated enzyme.2,6 In accordancewith our previously published results,24 the present studyshows that the above reduction is attributed to the fenofibrateaction to decrease sdLDL and to increase large buoyant LDLparticles, which have a higher clearance rate than sdLDL.26 Acontributory role to the reduction of Lp-PLA2 by fenofibrateplays also the decrease in enzyme associated with the
0
0.1
0.2
0.3
0.4
0.5
0.6
Ezetimibe Rosuvastatin FenofibrateHDL-3c
Lp-P
LA2 a
ctiv
ity
(nm
ol/m
L pl
asm
a/m
in)
BaselinePosttreatment
A
*
*
0
1
2
3
4
5
6
7
8
9
10
Ezetimibe Rosuvastatin FenofibrateHDL-3c
Lp-P
LA2m
ass
(ng/
mL
plas
ma)
BaselinePosttreatment
B
*
*
Figure 4. Bar graphs illustrating the effect of ezetimibe, rosuvas-tatin, and fenofibrate on the Lp-PLA2 activity (A) and Lp-PLA2
mass (B) associated with HDL-3c subfraction. HDL subfractionswere isolated by isopycnic gradient ultracentrifugation of plasmadepleted of apoB-lipoproteins. Enzymatic activity was deter-mined by the trichloroacetic acid (TCA) precipitation procedureand enzyme mass by use of a dual monoclonal antibody immu-noassay. Values represent the mean�SD. *P�0.01 comparedwith respective baseline values.
2240 Arterioscler Thromb Vasc Biol. October 2007
by guest on July 13, 2018http://atvb.ahajournals.org/
Dow
nloaded from

triglyceride-rich VLDL�IDL subfraction (attributable to thedrug-induced reduction in the plasma concentration of thissubfraction).
An important observation of the present study is that thenon-HDL–Lp-PLA2 specific activity is significantly in-creased by fenofibrate. According to our previous results, thespecific activity of Lp-PLA2 associated with large buoyantLDL is higher than that of either LDL-5 or VLDL�IDLsubfraction.6 In this regard, the results of the present studyshow that the method used for the determination of Lp-PLA2
mass may not detect all active enzyme in LDL, a phenome-non not observed for HDL, suggesting that structural differ-ences among lipoprotein species may significantly influencethe determination of enzyme mass, a hypothesis that needsfurther investigation. Based on the above observations, wemay suggest that the increase in non-HDL–Lp-PLA2 specificactivity by fenofibrate is attributed to the drug-inducedpreferential reduction in the enzyme associated with LDL-5and VLDL�IDL subfractions. A contributory role in theabove phenomenon may also play the fenofibrate-inducedincrease in the specific activity of Lp-PLA2 associated withLDL-5.
In accordance with our previously published results,24
fenofibrate treatment increases the HDL–Lp-PLA2 activity. Italso increases the HDL–Lp-PLA2 mass, thus it does not affectthe enzyme specific activity. This effect is attributable to thedrug-induced increase in plasma levels of both HDL-2 andHDL-3 subspecies as well as to the preferential enrichment ofthe HDL-3c in Lp-PLA2. We had previously suggested thatthe latter phenomenon is attributed to enzyme transfer fromtriglyceride-rich apoB-containing lipoproteins to HDL duringtheir enhanced lipolysis by lipoprotein lipase induced byfenofibrate.24,27 Although the role of the HDL-Lp-PLA2 inhumans has not been established yet, data from in vitroexperiments as well as in vivo studies in animal modelssuggest that this enzyme may significantly contribute to theantiatherogenic effects of HDL (reviewed in1). Consequently,the increase of HDL-Lp-PLA2 induced by fenofibrate mayrepresent an important antiatherogenic effect of this drug, ahypothesis that needs further investigation.
The present study further demonstrates that the adminis-tration of rosuvastatin in type IIA dyslipidemic patientssignificantly reduces Lp-PLA2 activity and mass associatedwith apoB-containing lipoproteins. This reduction is thehighest observed among all statins used in previous stud-ies,16–23 and it could be primarily attributed to the drug-induced reduction in the plasma concentration of all LDLsubfractions and to the preferential reduction of Lp-PLA2
associated with LDL-5. It has been suggested that simvastatinreduces LDL-associated Lp-PLA2 not only through thereceptor-mediated removal of LDL but also through areceptor-independent clearance of the lipid and enzymecontents of LDL.20 This mechanism may explain our resultson the preferential reduction in Lp-PLA2 associated withLDL-5 (expressed per mg of protein) induced by rosuvastatin.
Ezetimibe is a drug that acts by inhibiting the absorption ofcholesterol at the brush border of the intestinal wall.28,29 Thepresent study shows for the first time that it reduces theplasma levels of Lp-PLA2 mass and activity (although to a
lesser extent compared with rosuvastatin and fenofibrate), byreducing the plasma concentration of all apoB-containinglipoprotein subfractions. Furthermore, ezetimibe induces aslight but significant decrease in the HDL-Lp-PLA2 activityand mass. This follows the lowering effect of ezetimibe onplasma HDL-cholesterol levels a finding, which is not con-sistent in all studies30 and may reflect the relatively highpretreatment levels of HDL-cholesterol in our populationresulting in a regression to the mean effect. Becauseezetimibe does not influence the ratio of enzyme activity ormass to apoA-I levels, we suggest that the reduction inHDL-3 plasma concentration may account for the ezetimibe-induced decrease in the HDL-Lp-PLA2. This is furthersupported by the finding that ezetimibe decreases the enzymeassociated only with HDL-3c subfraction. It should be notedthat all patients treated with ezetimibe were statin-intolerant,therefore the above results may not be representative of othergroups given ezetimibe.
In addition to LDL and HDL, another carrier of Lp-PLA2
in plasma is Lp(a). Interestingly, we31 and others32 havedemonstrated that Lp(a) is enriched in Lp-PLA2 comparedwith LDL. However, Lp(a) can influence the distribution ofLp-PLA2 between LDL and HDL in plasma only when itsplasma levels exceed 8 mg/dL.31 Thus, it is unlikely that theLp(a)-associated Lp-PLA2 could influence the enzymechanges induced by hypolipidemic drugs, because the presentresults showed that no alterations in Lp(a) levels wereinduced by any lipid-lowering therapy and no patient exhib-ited a baseline or posttreatment Lp(a) levels above 8 mg/dL.Finally, no detectable amounts of Lp(a) were found in anylipoprotein subfraction at baseline or after treatment with anyhypolipidemic drug.
Clinical studies have shown an independent associationbetween plasma levels of Lp-PLA2 mass or activity andCVD.14 The present study further shows that the determina-tion of Lp-PLA2 mass, activity, and specific activity onindividual lipoprotein subfractions may be pathophysiologi-cally and clinically important. In this regard, we havepreviously shown that the preferential enzyme distribution onsdLDL particles compared with large buoyant apoB-containing has as a consequence an increased production oflysoPC, the main metabolite of Lp-PLA2, during oxidation ofthis subfraction.13 Several studies have supported the impor-tant role of lysoPC in atherogenesis,33 and more recently itwas shown that local coronary production of lysoPC isassociated with endothelial dysfunction and early atheroscle-rosis.34 Furthermore, the preferential association of Lp-PLA2
with HDL-3 subfraction as compared with other apoA-I–containing lipoproteins may contribute to the antiinflamma-tory and antioxidant effects of these particles.1 Finally theLp(a)-associated Lp-PLA2 may play an important role in themetabolism of oxidized phospholipids in humans, in view ofemerging data showing that oxidized phospholipids in plasmaare preferentially sequestered on Lp(a).35 Taking into accountour previous results showing that the type of dyslipidemiaand the underlying metabolic defect significantly influencethe enzyme distribution among lipoprotein subspecies,5,17,24
we suggest that the determination of enzyme parameters onspecific lipoprotein subspecies may provide useful informa-
Saougos et al Hypolipidemic Drugs and Lp-PLA2 2241
by guest on July 13, 2018http://atvb.ahajournals.org/
Dow
nloaded from

tion on both pathophysiological and clinical basis, in additionto the valuable information provided from the measurementof Lp-PLA2 mass and activity in total plasma.
A limitation of the present study could be the selection ofpatients who were allocated to different therapeutic agentsaccording to the NCEP ATPIII goals that resulted in popu-lations with different types of dyslipidemias. Thus it must beacknowledged that the different lipid abnormalities observedbetween Type IIA and Type IV patient groups may havecontributed to the differential effect on the lipoprotein-associated Lp-PLA2 levels of fenofibrate (administered inType IV patients) compared with rosuvastatin or ezetimibe(given to Type IIA individuals).
In conclusion the present study demonstrates for the firsttime that fenofibrate, rosuvastatin, and ezetimibe (actingthrough different mechanisms) reduce Lp-PLA2 activity andmass associated with the atherogenic apoB-containing li-poproteins, the rosuvastatin exhibiting the most potent effect.Additionally, fenofibrate increases the specific activity of thenon-HDL–Lp-PLA2 as well as the HDL–Lp-PLA2 mass andactivity. The clinical implications of these effects remain tobe established.
AcknowledgmentsWe are grateful to Dr Konstantinos G. Lagos and Dr Eleni S. Nakoufor skilled technical assistance.
Sources of FundingThis work was supported by research grants from the Greek Ministryof Research and Technology (PENED program 2003 ED 643).
DisclosuresNone.
References1. Tselepis AD, Chapman MJ. Inflammation, bioactive lipids and athero-
sclerosis: potential roles of a lipoprotein-associated phospholipase A2,platelet-activating factor acetylhydrolase. Atheroscler Suppl. 2002;3:57–68.
2. Tselepis AD, Dentan C, Karabina SA, Chapman MJ, Ninio E. PAF-degrading acetylhydrolase is preferentially associated with dense LDLand VLDL-1 in human plasma: catalytic characteristics and relation to themonocyte-derived enzyme. Arterioscler Thromb Vasc Biol. 1995;15:1764–1773.
3. Macphee CH, Moores KE, Boyd HF, Dhanak D, Ife RJ, Leach CA, LeakeDS, Milliner KJ, Patterson RA, Sucklling KE, Tew DG, Hickey DM.Lipoprotein-associated phospholipase A2, platelet-activating factoracetylhydrolase, generates two bioactive products during the oxidation oflow-density lipoprotein: use of a novel inhibitor. Biochem J. 1999;338:479–487.
4. Karabina SA, Liapikos TA, Grekas G, Goudevenos J, Tselepis AD.Distribution of PAF-acetylhydrolase activity in human plasma lowdensity lipoprotein subfractions. Biochim Biophys Acta. 1994;1213:34–38.
5. Tsimihodimos V, Karabina SA, Tambaki AP, Bairaktari E, Miltiadous G,Goudevenos JA, Kariolou MA, Chapman MJ, Tselepis AD, Elisaf M.Altered distribution of platelet activating factor- acetylhydrolase activitybetween LDL and HDL as a function of the severity of hypercholester-olemia. J Lipid Res. 2002;43:256–263.
6. Gazi I, Lourida ES, Filippatos T, Tsimihodimos V, Elisaf M, TselepisAD. Lipoprotein-associated phospholipase A2 activity is a marker ofsmall, dense LDL particles in human plasma. Clin Chem. 2005;51:2264–2273.
7. Asano K, Okamoto S, Fukunaga K, Shiomi T, Mori T, Iwata M, Ikeda Y,Yamaguchi K. Cellular source (s) of platelet-activating-factor acetylhy-drolase activity in plasma. Biochem Biophys Res Commun. 1999;261:511–514.
8. Stafforini DM, Elstad MR, McIntyre TM, Zimmerman GA, Prescott SM.Human macrophages secret platelet-activating factor acetylhydrolase.J Biol Chem. 1990;265:9682–9687.
9. Hakkinen T, Luoma JS, Hiltunen MO, Macphee CH, Milliner KJ, Patel L,Rice SQ, Tew DG, Karkola K, Yla-Herttuala S. Lipoprotein-associatedphospholipase A(2), platelet-activating factor acetylhydrolase, isexpressed by macrophages in human and rabbit atherosclerotic lesions.Arterioscler Thromb Vasc Biol. 1999;19:2909–2917.
10. Tjoelker LW, Wilder C, Eberhardt C, Stafforini DM, Dietsch G, SchimpfB, Hooper S, Le Trong H, Cousens LS, Zimmerman GA, Yamadat Y,McIntyre TM, Prescott SM, Gray PW. Anti-inflammatory properties of aplatelet-activating factor acetylhydrolase. Nature. 1995;374:549–553.
11. Lee C, Sigari F, Segrado T, Horkko S, Hama S, Subbaiah PV, Miwa M,Navab M, Witztum JL, Reaven PD. All apoB-containing lipoproteinsinduce monocyte chemotaxis and adhesion when minimally modified.Modulation of lipoprotein bioactivity by platelet-activating factor acetyl-hydrolase. Arterioscler Thromb Vasc Biol. 1999;19:1437–1446.
12. Carpenter KL, Dennis IF, Challis IR, Osborn DP, Macphee CH, LeakeDS, Arends MJ, Mitschinson MJ. Inhibition of lipoprotein-associatedphospholipase A2 diminishes the death-inducing effects of oxidized LDLon human monocyte-macrophages. FEBS Lett. 2001;505:357–363.
13. Karabina SA, Elisaf M, Bairaktari E, Tzallas C, Siamopoulos KC,Tselepis AD. Increased activity of platelet-activating factor acetylhy-drolase in low-density lipoprotein subfractions induces enhanced lyso-phosphatidylcholine production during oxidation in patients with het-erozygous familial hypercholesterolemia. Eur J Clin Invest. 1997;27:595–602.
14. Garza CAMV, McConnell JP, Somers VK, Kullo IJ, Lopez-Jimenez F.Association between lipoprotein-associated phospholipase A2 and car-diovascular disease: a systematic review. Mayo Clin Proc. 2007;82:159–165.
15. Elisaf M, Tselepis AD. Effect of hypolipidemic drugs on lipoprotein-associated platelet activating factor acetylhydrolase. Implication for ath-erosclerosis. Biochem Pharmacol. 2003;66:2069–2073.
16. Tambaki AP, Rizos E, Tsimihodimos V, Tselepis AD, Elisaf M. Effect ofantihypertensive and hypolipidemic drugs on plasma and HDL-associatedPAF-acetylhydrolase activity. J Cardiovasc Pharmacol Ther. 2004;9:91–95.
17. Tsimihodimos V, Karabina SA, Tambaki AP, Bairaktari E, GoudevenosJA, Chapman MJ, Elisaf M, Tselepis AD. Atorvastatin preferentiallyreduces LDL-associated platelet-activating factor acetylhydrolase activityin dyslipidemias of type IIA and type IIB. Arterioscler Thromb Vasc Biol.2002;22:306–311.
18. Schaefer EJ, McNamara JR, Asztalos BF, Tayler T, Daly JA, Gleason JL,Seman LJ, Ferrari A, Rubenstein JJ. Effects of atorvastatin versus otherstatins on fasting and postprandial C-reactive protein and lipoprotein-associated phospholipase A2 in patients with coronary heart diseaseversus control subjects. Am J Cardiol. 2005;95:1025–1032.
19. Kudolo GB, Bressler P, DeFronzo RA. Plasma PAF acetylhydrolase innon-insulin dependent diabetes mellitus and obesity: effect of hyperinsu-linemia and lovastatin treatment. J Lipid Mediat Cell Signal. 1997;17:97–113.
20. Zhang B, Fan P, Shimoji E, Itabe H, Miura SI, Uehara Y, Matsunaga A,Keijiro Saku K. Modulating effects of cholesterol feeding and simvastatintreatment on platelet-activating factor acetylhydrolase activity and lyso-phosphatidylcholine concentration. Atherosclerosis. 2006;186:291–301.
21. Winkler K, Abletshauser C, Friedrich I, Hoffmann MM, Wieland H, MarzW. Fluvastatin slow release lowers platelet activating factor acetylhydrolase activity: a placebo controlled trial in patients with Type 2diabetes. J Clin Endocrinol Metab. 2004;89:1153–1159.
22. O’Donoghue M, Morrow DA, Sabatine MS, Murphy SA, McCabe CH,Cannon CP, Braunwald E. Lipoprotein-associated phospholipase A2 andits association with cardiovascular outcomes in patients with acutecoronary syndromes in the PROVE IT-TIMI 22 (PRavastatin OratorVastatin Evaluation and Infection Therapy-Thrombolysis In Myo-cardial Infarction) trial. Circulation. 2006;113:1745–1752.
23. Albert MA, Glynn RJ, Wolfert RL, Ridker PM. The effect of statin therapyon lipoprotein associated phospholipase A2 levels. Atherosclerosis. 2005;182:193–198.
24. Tsimihodimos V, Kakafika A, Tambaki AP, Bairaktari E, Chapman MJ,Elisaf M, Tselepis AD. Fenofibrate induces HDL-associated PAF-AH butattenuates enzyme activity associated with apoB-containing lipoproteins.J Lipid Res. 2003;44:927–934.
25. Min JHWC, Aoki J, Arai H, Inoue K, Paul L, Gelb MH. Platelet-activating factor acetylhydrolases: broad substrate specificity and
2242 Arterioscler Thromb Vasc Biol. October 2007
by guest on July 13, 2018http://atvb.ahajournals.org/
Dow
nloaded from

lipoprotein binding does not modulate the catalytic properties of theplasma enzyme. Biochemistry. 2001;40:4539–4549.
26. Lund-Katz S, Laplaud PM, Phillips MC, Chapman MJ. Apolipoprotein B100confirmation and particle change in human LDL-subspecies: implication forLDL-receptor interaction. Biochemistry. 1998;37:128567–112874.
27. Fruchart JC. Peroxisome proliferator-activated receptor-a activation andhigh-density lipoprotein metabolism. Am J Cardiol. 2001;88:24N–29N.
28. van Heek M, Farley C, Compton DS, Hoos L, Alton KB, Sybertz EJ,Davis HRJ. Comparison of the activity and disposition of the novelcholesterol absorption inhibitor, SCH58235, and its glucuronide,SCH60663. Br J Pharmacol. 2000;129:1748–1754.
29. Knopp RH, Gitter H, Truitt T, Bays H, Manion CV, Lipka LJ, LeBeautAP, Suresh R, Yang B, Veltri EP. Effects of ezetimibe, a new cholesterolabsorption inhibitor, on plasma lipids in patients with primary hypercho-lesterolemia. Eur Heart J. 2003;24:729–741.
30. Gonzalez-Ortiz MM-AE, Kam-Ramos AM, Hernandez-Salazar E,Ramos-Zavala MG. Effect of ezetimibe on insulin sensitivity and lipidprofile in obese and dyslipidaemic patients. Cardiovasc Drugs Ther.2006;20:143–146.
31. Karabina SA, Elisaf MS, Goudevenos J, Siamopoulos KC, Sideris D,Tselepis AD. PAF-acetylhydrolase activity of Lp (a) before and duringCu(2�)-induced oxidative modification in vitro. Atherosclerosis 1996;125:121–134.
32. Blencowe C, Hermetter A, Kostner GM, Deigner HP. Enhanced asso-ciation of platelet-activating factor acetylhydrolase with lipoprotein (a) incomparison with low density lipoprotein. J Biol Chem. 1995;270:31151–31157.
33. Kougias P, Chai H, Lin PH, Lumsden AB, Yao Q, Chen C. Lysophos-phatidylcholine and secretory phospholipas A2 in vascular disease:mediators of endothelial dysfunction and atherosclerosis. Med Sci Monit.2006;12:RA5–RA16.
34. Lavi S, Mc Connell JP, Rihal CS, Prasad A, Mathew V, Lerman LO,Lerman A. Local production of lipoprotein-associated phospholipase A2and lysophosphatidylocholine in the coronary circulation: associationwith early coronary atherosclerosis and endothelial dysfunction inhumans. Circulation. 2007;115:2715–2721.
35. Tsimikas S, Brilakis ES, Miller ER, McConnell JP, Lennon RJ, KornmanKS, Witztum JL, Berger PB. Oxidized phospholipids, Lp(a) lipoprotein,and coronary artery disease. N Engl J Med. 2005;353:46–57.
Saougos et al Hypolipidemic Drugs and Lp-PLA2 2243
by guest on July 13, 2018http://atvb.ahajournals.org/
Dow
nloaded from

Gazi, Robert L. Wolfert, Moses Elisaf and Alexandros D. TselepisVasilios G. Saougos, Afroditi P. Tambaki, Mihalis Kalogirou, Michael Kostapanos, Irene F.
2Differential Effect of Hypolipidemic Drugs on Lipoprotein-Associated Phospholipase A
Print ISSN: 1079-5642. Online ISSN: 1524-4636 Copyright © 2007 American Heart Association, Inc. All rights reserved.
Greenville Avenue, Dallas, TX 75231is published by the American Heart Association, 7272Arteriosclerosis, Thrombosis, and Vascular Biology
doi: 10.1161/ATVBAHA.107.1472802007;27:2236-2243; originally published online July 26, 2007;Arterioscler Thromb Vasc Biol.
http://atvb.ahajournals.org/content/27/10/2236World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://atvb.ahajournals.org/content/suppl/2007/09/20/ATVBAHA.107.147280.DC1Data Supplement (unedited) at:
http://atvb.ahajournals.org//subscriptions/
at: is onlineArteriosclerosis, Thrombosis, and Vascular Biology Information about subscribing to Subscriptions:
http://www.lww.com/reprints
Information about reprints can be found online at: Reprints:
document. Question and AnswerPermissions and Rightspage under Services. Further information about this process is available in the
which permission is being requested is located, click Request Permissions in the middle column of the WebCopyright Clearance Center, not the Editorial Office. Once the online version of the published article for
can be obtained via RightsLink, a service of theArteriosclerosis, Thrombosis, and Vascular Biologyin Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions:
by guest on July 13, 2018http://atvb.ahajournals.org/
Dow
nloaded from

Methods
Patients
One hundred and fifty hyperlipidaemic patients attending the Outpatient Lipid
Clinic of the University Hospital of Ioannina participated in the present study.
Exclusion criteria were: prior atherosclerotic disease (myocardial infarction, unstable
angina, ischemic stroke, peripheral arterial disease, percutaneous transluminal
coronary angioplasty and coronary artery bypass graft), diagnosed diabetes mellitus
(fasting glucose >6.9 mmol/L), liver disease (serum aminotranferase activity greater
than 3-fold, e.g. >120 IU/L, normal range 5-40 IU/L), renal disease (serum creatinine
levels greater than 132 mmol/L, normal range 53-106 mmol/L) and hypothyroidism
(TSH greater than 5 μIU/ml, normal range 0.5-4.8 μIU/L). Moreover, patients
receiving drugs that could affect lipid metabolism as well as renal or hepatic function
were also excluded from the present study. Finally, all patients with serum levels of
lipoprotein (a) [Lp(a)] over the cut-off point of 8 mg/dL were excluded from the study
since according to our previously published results, higher concentrations of Lp(a)
significantly affect the distribution of Lp-PLA2 among plasma lipoprotein
subfractions separated by gradient ultracentrifugation1 After the initial screening,
patients were advised to follow the National Cholesterol Education Program (NCEP)
step 1 diet for three months2. At the end of this period, a complete laboratory baseline
analysis was performed. According to their lipid levels, patients were divided into the
following groups: 1. Primary hypercholesterolaemia [Type IIA dyslipidaemia, i.e.
serum levels of low-density lipoprotein cholesterol (LDL-cholesterol) >160 mg/dL
(4.1 mmol/L) and serum triglyceride levels <200 mg/dL (2.3mmol/L), n= 50, who
were administered rosuvastatin (10 mg daily)]. 2. Type IIA dyslipidaemia [n= 50,
intolerant in previous statin therapy [expressed as elevated creatine kinase (CK) levels

>5 ULN or liver enzymes >3 ULN], who were administered ezetimibe (10 mg daily)].
3. Primary hypertriglyceridaemia [type IV dyslipidaemia, i.e. serum levels of LDL-
cholesterol <160 mg/dL (4.1 mmol/L) and serum triglyceride levels ≥200 mg/dL (2.3
mmol/L), n= 50, who were given micronised fenofibrate (200 mg daily)] in order to
achieve the NCEP Adult Treatment Panel III goals for the LDL-cholesterol levels2
(Table I). After 2 months of active treatment, a second blood analysis was performed.
The Ethics Committee of the University Hospital of Ioannina gave its approval for the
conduction of the study and every participant gave written consent.
Biochemical parameters
All lipid and lipoprotein determinations were carried out after an overnight fast.
Serum levels of total cholesterol, HDL-cholesterol and triglycerides were determined
using an Olympus AU 600 analyzer (Olympus Diagnostica, Hamburg, Germany).
LDL-cholesterol was calculated using the Friedewald formula. Apolipoproteins (apo)
B and A-I were measured with a Behring Nephelometer BN100 (Liederbach,
Germany). The protein content of the lipoprotein subfractions was measured by the
bicinchoninic acid method (Pierce) 3. Lipoprotein [a] (Lp[a]) levels in serum and in
lipoprotein subfractions were measured by an enzyme immunoassay method (Macra
Lp[a], Terumo Medical Corporation Diagnostic Division, Elkton, MD, USA) as we
have previously described 1.
Electrophoretic analysis of apoB and apoA-I lipoprotein subclasses
Lipoprotein subclass analysis, was performed electrophoretically by use of high-
resolution 3% polyacrylamide gel tubes and the Lipoprint LDL System
(Quantimetrix, Redondo Beach, CA), following the instructions provided by the
2

manufacturer4. Analysis of the apoB-containing lipoproteins was performed as we
recently described5. After electrophoresis, very low-density lipoprotein (VLDL)
remained in the origin [retention factor (Rf) = 0.0], whereas HDL migrated at the front
(Rf = 1.0). In between, several bands can be detected: MID bands C, B, and A, which
correspond mainly to intermediate-density lipoprotein (IDL), as well as up to 7 LDL
bands. The LDL1 and LDL2 bands correspond to large, buoyant LDL particles,
whereas bands LDL3 to LDL7 correspond to sdLDL particles. We determined the
cholesterol mass of each apoB-lipoprotein subfraction, the mean LDL particle size (in
Å), and the proportion (%) of the cholesterol mass of sdLDL particles over the total
LDL cholesterol mass.
Analysis of the apoA-I-containing lipoproteins was performed using the same
electrophoretic procedure with some modifications. Briefly, 25 µL of sample was
mixed with 300 µL of Lipoprint Loading Gel and placed upon the upper part of the
high resolution 3% polyacrylamide gel. After 30 minutes of photopolymerization in
room temperature, electrophoresis was performed for 50 minutes with 3 mA for each
gel tube. After electrophoresis, VLDL and LDL remained in the origin (Rf = 0.0)
whereas albumin migrated at the front (Rf = 1.0). In between, up to 9 bands of HDL
can be detected whose Rf’s are 0.05, 0.10, 0.15, 0.20, 0.25, 0.29, 0.38, 0.48 and 0.53.
Bands 1 to 3 (Rf = 0.05 to 0.15) correspond to large HDL-2 subfraction, whereas
bands 4 to 9 (Rf = 0.20 to 0.53) comprised the small HDL-3 subfraction. The
cholesterol mass of each HDL subfraction was determined as previously described5.
Subfractionation of plasma lipoproteins by ultracentrifugation
Lipoproteins were fractionated by isopycnic density gradient ultracentrifugation as
previously described6. To study the effect of hypolipidemic drugs on Lp-PLA2
3

activity and mass associated with apoB-containing lipoprotein subspecies, total
plasma was subjected to isopycnic density gradient ultracentrifugation as previously
described6. According to previously published data, a proportion of LDL-bound Lp-
PLA2 dissociates and is redistributed to apoA-I-containing lipoprotein subspecies,
during ultracentrifugation of total plasma7. Thus, to avoid any contamination of the
HDL-associated Lp-PLA2 with the LDL-associated enzyme during
ultracentrifugation, we studied the effect of hypolipidemic drugs on Lp-PLA2
associated with the HDL subspecies, using plasma depleted of apoB-lipoproteins, ie
the HDL-containing supernatant, after treatment of plasma with magnesium chloride–
dextran sulfate (to precipitate all apoB-containing lipoproteins)3. When plasma was
used, the following apoB-containing subfractions were prepared: VLDL - IDL (d =
1.019 g/mL), LDL-1 (d = 1.019–1.023 g/mL), LDL-2 (d = 1.023–1.029 g/mL), LDL-
3 (d = 1.029 –1.039 g/mL), LDL-4 (d = 1.039–1.050 g/mL), and LDL-5 (d = 1.050–
1.063 g/mL). When the HDL-containing supernatant was used, the following apoA-I-
containing subfractions were prepared: HDL-2b (d = 1.063–1.091 g/mL), HDL-2a (d
= 1.091–1.100 g/mL), HDL-3a (d = 1.100–1.133 g/mL), HDL-3b (d = 1.133–1.156
g/mL), and HDL-3c (d = 1.156–1.179 g/mL). All subfractions were dialyzed
extensively at 4°C in 10 mmol/L phosphate-buffered saline, pH 7.4 (PBS) containing
2 mmol/L EDTA, filter-sterilized, and maintained at 4°C under nitrogen until
analysis. Under these storage conditions, no oxidation has been reported to occur3.
Measurement of Lp-PLA2 activity
Lp-PLA2 activity in total plasma, in apoB-depleted plasma, after the
sedimentation of all apo B-containing lipoproteins with dextran sulfate-magnesium
chloride (HDL-Lp-PLA2 activity), as well as in lipoprotein subfractions, was
4

determined by the trichloroacetic acid precipitation procedure using [3H]-PAF (100
μmol/L final concentration) as a substrate6,8. Lp-PLA2 activity was expressed as nmol
PAF degraded per min per mL of plasma or mg of LDL subfraction protein. The non-
HDL Lp-PLA2 activity was calculated by subtracting the HDL-Lp-PLA2 activity from
the total plasma enzyme activity. In separate experiments, Lp-PLA2 activity was
determined in LDL subfractions LDL-3 and LDL-5 of type IIA dyslipidaemic patients
at baseline (using 8 μg of protein) in the presence of either human serum albumin (at
concentrations ranging from 4 to 8 g/dL) or the total plasma proteins (at
concentrations ranging from 4 to 8 g/dL) prepared by mixing of equal volumes from
the gradient fractions 25-30 (d = 1.190-1.210 g/mL) isolated by ultracentrifugation of
the HDL-containing plasma supernatant (as described above). According to our
previously published results these subfractions do not contain any Lp-PLA2 activity6.
In other experiments equal volumes of the five LDL subfractions (LDL-1 to LDL-5)
or the 5 HDL subfractions HDL-2b to HDL-3c from type IIA dyslipidaemic patients
at baseline, were mixed to form the total LDL or total HDL. Subsequently, each
lipoprotein was treated with 0.1% Triton X-100 to dissociate the particle. Lp-PLA2
activity was determined before and after Triton treatment using 8 μg of protein as the
source of the enzyme. The final Triton concentration in the enzyme assay was 0.01%.
It should be noted that according to our previously published results the above Triton
concentration does not affect the enzyme activity9.
To study the recovery of Lp-PLA2 activity after ultracentrifugation in the presence
and absence of either albumin or Triton X-100, we mixed equal volumes from the
gradient fractions 1-24 containing the total enzyme activity. Lp-PLA2 activity was
determined after treatment of the reconstituted gradient fractions with either albumin
6 g/dL or 0.1% Triton X-100. Enzyme activities were determined as above. In the
5

absence of any agent, the enzyme recovery (compared to enzyme activity in total
plasma) was 150±15%. In the presence of albumin or Triton X-100 the enzyme
recovery was 90±5% or 146±12% respectively.
Measurement of Lp-PLA2 mass
Lp-PLA2 mass in total plasma, in apoB-depleted plasma (HDL Lp-PLA2 mass),
as well as in lipoprotein subfractions, was determined by a dual monoclonal antibody
immunoassay standardized to recombinant Lp-PLA2 (PLAC test, diaDexus, Inc.),
following the manufacturer instructions, as we recently described5. The non-HDL-
Lp-PLA2 mass was calculated by subtracting the HDL-Lp-PLA2 mass from the total
plasma enzyme mass5. In separate experiments, Lp-PLA2 mass was determined in
LDL subfractions LDL-3 and LDL-5 in the presence of either human serum albumin
(at concentrations ranging from 4 to 8 g/dL) or the total plasma proteins (at
concentrations ranging from 4 to 8 g/dL) prepared as described above. In other
experiments Lp-PLA2 mass was determined in the total LDL or total HDL prepared
as described above, before or after treatment with 0.1% Triton X-100. It should be
noted that the presence of the Triton did not affect the standard calibrator curves of
enzyme mass assay. Lp-PLA2 specific activity was expressed as a ratio of the
enzyme activity to the enzyme mass (nmol/ng/min)5. To study the recovery of Lp-
PLA2 mass after ultracentrifugation in the presence and absence of either albumin or
Triton X-100, we mixed equal volumes from the gradient fractions 1-24 containing
the total enzyme mass. Lp-PLA2 mass was determined after treatment of the
reconstituted gradient fractions with either albumin 6 g/dL or 0.1% Triton X-100. In
the absence of any agent the enzyme recovery (compared to enzyme mass in total
6

plasma) was 94±4%. In the presence of albumin or Triton X-100 the enzyme
recovery was 91±6% or 119±16%, respectively.
Statistical analysis
Data are presented as mean ± SD, except for Lp(a) which was expressed as the
median and range. Statistical analysis was performed using paired Student’s t-test for
comparisons between baseline and posttreatment values, while one-way analysis of
variance (ANOVA) followed by LSD test was used for comparisons between
individual groups. Because of the skewed distribution of Lp(a), the nonparametric
Wilcoxon matched-pairs test was used for comparisons between baseline and post
treatment values. Yates’s corrected chi-square test was used for differences in
proportions. A p value of less than 0.05 was considered to be significant. All analyses
were carried out using SPSS 13.0 softpack.
7

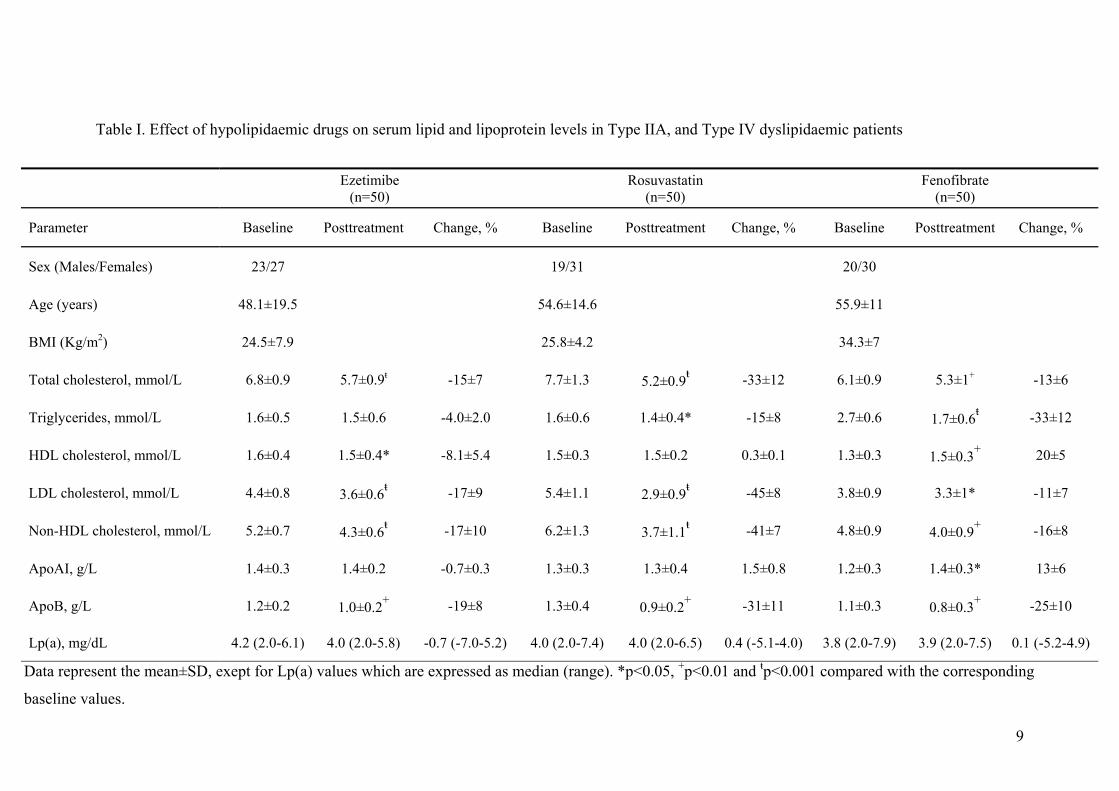
Table I. Effect of hypolipidaemic drugs on serum lipid and lipoprotein levels in Type IIA, and Type IV dyslipidaemic patients
Εzetimibe (n=50)
Rosuvastatin (n=50)
Fenofibrate (n=50)
Parameter Baseline Posttreatment Change, % Baseline Posttreatment Change, % Baseline Posttreatment Change, %
Sex (Males/Females) 23/27 19/31 20/30
Age (years) 48.1±19.5 54.6±14.6 55.9±11
BMI (Kg/m2) 24.5±7.9 25.8±4.2 34.3±7
Total cholesterol, mmol/L 6.8±0.9 5.7±0.9ŧ -15±7 7.7±1.3 5.2±0.9ŧ -33±12 6.1±0.9 5.3±1+ -13±6
Triglycerides, mmol/L 1.6±0.5 1.5±0.6 -4.0±2.0 1.6±0.6 1.4±0.4* -15±8 2.7±0.6 1.7±0.6ŧ -33±12
HDL cholesterol, mmol/L 1.6±0.4 1.5±0.4* -8.1±5.4 1.5±0.3 1.5±0.2 0.3±0.1 1.3±0.3 1.5±0.3+ 20±5
LDL cholesterol, mmol/L 4.4±0.8 3.6±0.6ŧ -17±9 5.4±1.1 2.9±0.9ŧ -45±8 3.8±0.9 3.3±1* -11±7
Non-HDL cholesterol, mmol/L 5.2±0.7 4.3±0.6ŧ -17±10 6.2±1.3 3.7±1.1ŧ -41±7 4.8±0.9 4.0±0.9+ -16±8
ApoAI, g/L 1.4±0.3 1.4±0.2 -0.7±0.3 1.3±0.3 1.3±0.4 1.5±0.8 1.2±0.3 1.4±0.3* 13±6
ApoB, g/L 1.2±0.2 1.0±0.2+ -19±8 1.3±0.4 0.9±0.2+ -31±11 1.1±0.3 0.8±0.3+ -25±10
Lp(a), mg/dL 4.2 (2.0-6.1) 4.0 (2.0-5.8) -0.7 (-7.0-5.2) 4.0 (2.0-7.4) 4.0 (2.0-6.5) 0.4 (-5.1-4.0) 3.8 (2.0-7.9) 3.9 (2.0-7.5)
Data represent the mean±SD, exept for Lp(a) values which are expressed as median (range). *p<0.05, +p<0.01 and ŧp<0.001 compared with the corresponding
baseline values.
0.1 (-5.2-4.9)
9

References
1. Karabina SA, Elisaf MC, Goudevenos J, Siamopoulos KC, Sideris D, Tselepis AD.
PAF-acetylhydrolase activity of Lp(a) before and during Cu(2+)-induced oxidative
modification in vitro. Atherosclerosis 1996; 125: 121-134.
2. Expert Panel on Detection, Evaluation and Treatment of High Blood Cholesterol in
Adults. Executive summary of the third report of the National Cholesterol Education
Program (NCEP) Expert Panel on Detection, Evaluation, and Treatment of High Blood
Cholesterol in Adults (Adult Treatment Panel III). JAMA. 2001;285:2486-2497.
3. Tsimihodimos V, Kakafika A, Tambaki AP, Bairaktari E, Chapman MJ, Elisaf M,
Tselepis AD. Fenofibrate induces HDL-associated PAF-AH but attenuates enzyme
activity associated with apoB-containing lipoproteins. J Lipid Res. 2003;44:927-934.
4. Hoefner DM, Hodel SD, O'Brien JF, Branum EL, Sun D, Meissner I, McConnell JP.
Development of a rapid, quantitative method for LDL subfractionation with use of the
Quantimetrix Lipoprint LDL System. Clin Chem. 2001;47:266-274.
5. Gazi I, Lourida ES, Filippatos T, Tsimihodimos V, Elisaf M, Tselepis AD. Lipoprotein-
associated phospholipase A2 activity is a marker of small, dense LDL particles in
human plasma. Clin Chem. 2005;51:2264-2273.
6. Tselepis AD, Dentan C, Karabina SA, Chapman MJ, Ninio E. PAF-degrading
acetylhydrolase is preferentially associated with dense LDL and VLDL-1 in human
plasma: catalytic characteristics and relation to the monocyte-derived enzyme.
Arterioscler Thromb Vasc Biol. 1995;15:1764-1773.
7. McCall, M. R., M. LaBelle, T. M. Forte, R. M. Krauss, Y. Takanami, and D. L. Tribble.
Dissociable and nondissociable forms of platelet-activating factor acetylhydrolase in
human plasma LDL: implications for LDL oxidative susceptibility. Biochim. Biophys.
Acta. 1999;1437: 23–36.
10

8. Tsimihodimos V, Karabina SA, Tambaki AP, Bairaktari E, Miltiadous G, Goudevenos
JA, Kariolou MA, Chapman MJ, Tselepis AD, Elisaf M. Altered distribution of platelet
activating factor- acetylhydrolase activity between LDL and HDL as a function of the
severity of hypercholesterolemia. J Lipid Res. 2002;43:256-263.
9. Tselepis AD, Karabina S-A, Stengel D, Piedagnel R, Chapman MJ, Ninio E. N-linked
glycosylation of macrophage-derived PAF-AH is a major determinant of enzyme
association with plasma HDL. J Lipid Res. 2001;42:1645-1654.
11


















