
Dietary fish oil reduces the incidence of triggered arrhythmias in pig ventricular myocytes
9
Dietary fish oil reduces the incidence of triggered arrhythmias in pig ventricular myocytes Géza Berecki, PhD,* Hester M. Den Ruijter, MSc,* Arie O. Verkerk, PhD, Cees A. Schumacher, BSc, Antonius Baartscheer, PhD, Diane Bakker, BSc, Bastiaan J. Boukens, MSc, Antoni C.G. van Ginneken, PhD, Jan W.T. Fiolet, PhD, Tobias Opthof, PhD, Ruben Coronel, MD, PhD From the Experimental Cardiology Group (ECG), Center for Heart Failure Research, Academic Medical Center, University of Amsterdam, Amsterdam, The Netherlands. BACKGROUND Fish oil reduces the incidence of sudden cardiac death in postmyocardial infarction patients. Triggered activity is the principal mechanism of arrhythmogenesis under these conditions. OBJECTIVE The purpose of this study was to test whether dietary fish oil in pigs inhibits Ca 2 overload-induced triggered activity. METHODS Pigs were fed a diet of fish oil or sunflower oil for 8 weeks. Ventricular myocytes (3: fish oil, n 11; control: sun- flower oil, n 8) were isolated by enzymatic dissociation and used for patch clamp studies and intracellular Ca 2 recordings. Triggered activity was induced by rapid pacing in the presence of norepinephrine. RESULTS Dietary fish oil reduced the incidence of triggered ac- tion potentials and delayed afterdepolarizations compared to con- trol (9.1% in 3 and 84.6% in control, P .05), concomitant with a reduction in spontaneous Ca 2 release. Dietary fish oil prevented Ca 2 overload and reduced action potential prolongation in re- sponse to norepinephrine (APD 90 : 23.2 8.5 ms in 3 and 107.4 15.9 in control, P .05). 3 myocytes displayed de- creased sarcoplasmic reticulum Ca 2 content, reduced L-type Ca 2 current (I Ca,L ), and less recruitment of the Na /Ca 2 exchange current (I NCX ) in response to norepinephrine compared to control. In the absence of norepinephrine, the slow component of the delayed rectifier current (I Ks ) was larger in 3 myocytes. In the presence of norepinephrine, I Ks increased to the same level in 3 and control myocytes. CONCLUSION Dietary fish oil reduces the incidence of triggered activity and prevents Ca 2 overload and AP prolongation in re- sponse to norepinephrine. Fish oil may prevent arrhythmias in patients with heart failure. KEYWORDS Fish oil; 3-Polyunsaturated fatty acid; Triggered ac- tivity; Delayed afterdepolarization; Intracellular calcium; Norepi- nephrine; Arrhythmia; Electrophysiology (Heart Rhythm 2007;4:1452–1460) © 2007 Heart Rhythm Society. All rights reserved. Introduction Increased consumption of 3-polyunsaturated fatty acids (3-PUFAs) from fish reduces the incidence of sudden cardiac death in patients with recent myocardial infarc- tion, 1,2 suggesting that dietary fish oil is antiarrhythmic. Arrhythmias in the postmyocardial infarction setting are initiated by triggered activity 3,4 resulting from delayed af- terdepolarizations (DADs). DADs in human myocytes are due to Na /Ca 2 exchanger current (I NCX ) 5 following spon- taneous sarcoplasmic reticulum (SR) Ca 2 releases. 6,7 This release is facilitated by tachycardia and -adrenergic stim- ulation, because both elevate intracellular Ca 2 . 8 The anti- arrhythmic mechanism of fish oil supplementation in post- myocardial infarction patients is not completely understood. Dietary fish oil leads to incorporation of 3-PUFAs into the sarcolemma. 9 This leads to shortening of the ventric- ular action potential (AP) due to decreased L-type Ca 2 current (I Ca,L ) and I NCX and increased inward rectifier current (I K1 ) and I Ks , the slow component of the delayed rectifier current. 9 AP shortening leads to an increased diastolic interval, favoring removal of excess Ca 2 from the cytosol and reducing Ca 2 overload conditions. 8 In addition, an increase in I K1 and a reduction in I NCX protect against DADs. 7,10 Based on these factors, we hypothesize that dietary fish oil reduces the incidence of triggered activity under Ca 2 overload conditions. Therefore, we studied the occurrence of triggered APs, DADs, and spontaneous SR Ca 2 release after rapid pacing in the presence of norepinephrine in myocytes isolated from pigs fed a diet rich in fish oil (3) or sunflower oil (control). We also studied the changes in AP duration and cytosolic Ca 2 in response to norepineph- rine in 3 and control myocytes. Funded by the Wageningen Centre for Food Sciences, Wageningen University, Maastricht University, and TNO Nutrition and Food Research; The SEAFOODplus Program of the European Union (Grant 506359); and The Netherlands Heart Foundation (Grant 2003B079). * The first two authors contributed equally to this study. Address reprint requests and correspondence: Dr. Ruben Coronel, Experimental Cardiology Group (ECG), Center for Heart Failure Research, Academic Medical Center, University of Amsterdam, Room K2-112, Meibergdreef 9, 1105 AZ Amsterdam, The Netherlands. E-mail address: [email protected]. (Received June 5, 2007; accepted July 10, 2007.) 1547-5271/$ -see front matter © 2007 Heart Rhythm Society. All rights reserved. doi:10.1016/j.hrthm.2007.07.015
Transcript of Dietary fish oil reduces the incidence of triggered arrhythmias in pig ventricular myocytes

Di
GAA
FU
Bdp
Ofi
MwfluTn
RttaCs
II(ctAitdtru
UTT
EAMr
1
ietary fish oil reduces the incidence of triggered arrhythmiasn pig ventricular myocytes
éza Berecki, PhD,* Hester M. Den Ruijter, MSc,* Arie O. Verkerk, PhD, Cees A. Schumacher, BSc,ntonius Baartscheer, PhD, Diane Bakker, BSc, Bastiaan J. Boukens, MSc,ntoni C.G. van Ginneken, PhD, Jan W.T. Fiolet, PhD, Tobias Opthof, PhD, Ruben Coronel, MD, PhD
rom the Experimental Cardiology Group (ECG), Center for Heart Failure Research, Academic Medical Center,
niversity of Amsterdam, Amsterdam, The Netherlands.1cccIdpa
Casp
Ktn
(
ACKGROUND Fish oil reduces the incidence of sudden cardiaceath in postmyocardial infarction patients. Triggered activity is therincipal mechanism of arrhythmogenesis under these conditions.
BJECTIVE The purpose of this study was to test whether dietarysh oil in pigs inhibits Ca2� overload-induced triggered activity.
ETHODS Pigs were fed a diet of fish oil or sunflower oil for 8eeks. Ventricular myocytes (�3: fish oil, n � 11; control: sun-ower oil, n � 8) were isolated by enzymatic dissociation andsed for patch clamp studies and intracellular Ca2� recordings.riggered activity was induced by rapid pacing in the presence oforepinephrine.
ESULTS Dietary fish oil reduced the incidence of triggered ac-ion potentials and delayed afterdepolarizations compared to con-rol (9.1% in �3 and 84.6% in control, P �.05), concomitant withreduction in spontaneous Ca2� release. Dietary fish oil preventeda2� overload and reduced action potential prolongation in re-
am
tuccrdtap
[email protected]. (Received June 5, 2007; accepted July 10, 2007.)
547-5271/$ -see front matter © 2007 Heart Rhythm Society. All rights reserved
07.4 � 15.9 in control, P �.05). �3 myocytes displayed de-reased sarcoplasmic reticulum Ca2� content, reduced L-type Ca2�
urrent (ICa,L), and less recruitment of the Na�/Ca2� exchangeurrent (INCX) in response to norepinephrine compared to control.n the absence of norepinephrine, the slow component of theelayed rectifier current (IKs) was larger in �3 myocytes. In theresence of norepinephrine, IKs increased to the same level in �3nd control myocytes.
ONCLUSION Dietary fish oil reduces the incidence of triggeredctivity and prevents Ca2� overload and AP prolongation in re-ponse to norepinephrine. Fish oil may prevent arrhythmias inatients with heart failure.
EYWORDS Fish oil; �3-Polyunsaturated fatty acid; Triggered ac-ivity; Delayed afterdepolarization; Intracellular calcium; Norepi-ephrine; Arrhythmia; Electrophysiology
Heart Rhythm 2007;4:1452–1460) © 2007 Heart Rhythm Society.
ponse to norepinephrine (�APD90: 23.2 � 8.5 ms in �3 and All rights reserved.ntroductionncreased consumption of �3-polyunsaturated fatty acids�3-PUFAs) from fish reduces the incidence of suddenardiac death in patients with recent myocardial infarc-ion,1,2 suggesting that dietary fish oil is antiarrhythmic.rrhythmias in the postmyocardial infarction setting are
nitiated by triggered activity3,4 resulting from delayed af-erdepolarizations (DADs). DADs in human myocytes areue to Na�/Ca2� exchanger current (INCX)5 following spon-aneous sarcoplasmic reticulum (SR) Ca2� releases.6,7 Thiselease is facilitated by tachycardia and �-adrenergic stim-lation, because both elevate intracellular Ca2�.8 The anti-
Funded by the Wageningen Centre for Food Sciences, Wageningenniversity, Maastricht University, and TNO Nutrition and Food Research;he SEAFOODplus Program of the European Union (Grant 506359); andhe Netherlands Heart Foundation (Grant 2003B079).
*The first two authors contributed equally to this study.Address reprint requests and correspondence: Dr. Ruben Coronel,xperimental Cardiology Group (ECG), Center for Heart Failure Research,cademic Medical Center, University of Amsterdam, Room K2-112,eibergdreef 9, 1105 AZ Amsterdam, The Netherlands. E-mail address:
rrhythmic mechanism of fish oil supplementation in post-yocardial infarction patients is not completely understood.Dietary fish oil leads to incorporation of �3-PUFAs into
he sarcolemma.9 This leads to shortening of the ventric-lar action potential (AP) due to decreased L-type Ca2�
urrent (ICa,L) and INCX and increased inward rectifierurrent (IK1) and IKs, the slow component of the delayedectifier current.9 AP shortening leads to an increasediastolic interval, favoring removal of excess Ca2� fromhe cytosol and reducing Ca2� overload conditions.8 Inddition, an increase in IK1 and a reduction in INCX
rotect against DADs.7,10
Based on these factors, we hypothesize that dietary fishil reduces the incidence of triggered activity under Ca2�
verload conditions. Therefore, we studied the occurrencef triggered APs, DADs, and spontaneous SR Ca2� releasefter rapid pacing in the presence of norepinephrine inyocytes isolated from pigs fed a diet rich in fish oil (�3)
r sunflower oil (control). We also studied the changes inP duration and cytosolic Ca2� in response to norepineph-
ine in �3 and control myocytes.
. doi:10.1016/j.hrthm.2007.07.015

MCTH(([ntpstawp(bMccsmswfaC
EMtCtasldddtpdf
o9taaoecnwf
a
IIt(2m0(mDc
CImfiwnbas1Cnwro(rt
SDSt
Fpts
1453Berecki et al Fish-Oil Diet Reduces Incidence of Triggered Activity
ethodsell preparationhe investigation conforms to U.S. National Institutes ofealth guidelines (NIH Publication 85-23, 1996). Male pigs
age 7 weeks) received a diet rich in fish oil �3-PUFAseicosapentaenoic acid [EPA] and docosahexaenoic acidDHA]; n � 11) or sunflower oil (rich in �9-fatty acids;� 8) for 8 weeks. The difference between the two diets is
hat the fish oil diet contains 4% fish oil (�1.5 g EPA/DHAer 100 g feed), whereas the control diet contains 4%unflower oil (�3.7 g oleic acid). Analysis of the diet showshat all other components are equal, including the energynd total fat content.9 Subsequently, animals were sedatedith ketamine (500 mg IM, Nimatek, Animal Health), aza-erone (160 mg IM, Stresnil, Janssen-Cilag), and atropine0.5 mg IM, Centrafarm) and killed by injection of pento-arbital (20 mg/kg IV, Nembutal, Ceva Sante Animale).idmyocardial left ventricular myocytes were enzymati-
ally isolated as described previously.11 Small aliquots ofell suspension were placed in a recording chamber on thetage of an inverted microscope and allowed to adhere for 5inutes before superfusion was initiated. Quiescent rod-
haped cells with cross-striations and with smooth surfaceere selected for measurements. For each dataset, myocytes
rom 2 to 6 animals from each group were used. Separateliquots of myocytes were used for electrophysiology anda2� measurements.
lectrophysiologyembrane potentials and currents were recorded by ampho-
ericin-perforated patch clamp technique [Ca2�-activatedl� current (ICl(Ca)), IK1] or with the ruptured patch clamp
echnique [rapid delayed rectifier K� current (IKr), IKs, ICa,L,nd INCX] at 36° � 0.5°C with superfusion and pipetteolutions as described previously.9 Membrane currents wereow-pass filtered with a cutoff frequency of 1 or 3 kHz andigitized at 5 kHz. AP measurements were filtered andigitized at 5 and 10 kHz, respectively. Voltage control,ata acquisition, and analysis were accomplished using cus-om software. Potentials were corrected for liquid junctionotential. Cell membrane capacitance (Cm) was estimated asescribed previously.12 Series resistance was compensatedor by at least 80%.
APs were elicited at 0.5 to 3 Hz by 3-ms (1.5 � thresh-ld) current pulses through the patch pipette. AP duration at0% repolarization (APD90) and plateau level (defined ashe potential value recorded 60 ms after the upstroke) werenalyzed. AP parameters from six consecutive APs wereveraged. APs were measured in the absence and presencef norepinephrine (20 nM, Centrafarm, Etten-Leur, The Neth-rlands). In order to obtain steady-state conditions, AP re-ordings were started 1 minute after application of norepi-ephrine. Susceptibility to triggered activity and DADsere evoked by a 3-Hz (10-second) rapid pacing episode
ollowed by an 8-second pause (tracing period).Ion currents were measured in a paired experiment in the
bsence and presence norepinephrine in the same myocyte. s
Kr was measured as the residual current in the presence of
Ks blocker chromanol 293B (90 �M). IKs was measured ashe residual current in the presence of IKr blocker E-4031E-4031 5 �M; Eisai, Woodcliff Lake, NJ, USA; chromanol93B: Tocris Cookson, Ellisville, MO, USA).13 ICl(Ca) waseasured as the transient outward current sensitive to
.2 mM 4,4=diisothiocyanatostilbene-2,2=-disulfonic acidDIDS; Sigma-Aldrich, St. Louis, MO, USA). INCX waseasured as 10 mM Ni2�-sensitive current. The effects ofIDS and Ni2� are reversible.14,15 Current densities were
alculated by dividing current amplitudes by Cm.
ytosolic Ca2� measurementsntracellular Ca2� ([Ca2�]i) was measured in indo-1–loadedyocytes as described previously.16 Briefly, myocytes wereeld stimulated, and dual wavelength emission of indo-1as recorded [(405–440)/(505–540) nm, excitation at 340m]. Signals were digitized at 1 kHz and corrected forackground signals recorded from indo-1–free myocytes,fter which free [Ca2�]i was calculated. Each experimenttarted with a conditioning period of 2 minutes in which-Hz field stimulation was applied. This was followed bya2� transient recordings in the absence and presence oforepinephrine (20 nM) at 1-Hz stimulation. Recordingsere started 1 minute after the application of norepineph-
ine. Ca2� aftertransients were evoked by a 3-Hz (10-sec-nd) rapid pacing episode followed by an 8-second pausetracing period). SR Ca2� content was determined as theesponse to a rapid application of Tyrode’s solution con-aining caffeine (10 mM).
tatistical analysisata are given as mean � SEM (n � number of myocytes).tatistical significance was based on P �.05 using Student’s
-test, two-way repeated measures analysis of variance fol-
igure 1 Representative examples of the occurrence of triggered actionotentials (APs), delayed afterdepolarizations (DADs) (A) and Ca2� after-ransients (B) in a control and an �3 myocyte. The arrow indicates the lasttimulated AP or Ca2� transient preceding the tracing period. Stars indicate
pontaneous APs, DADs, and Ca2� aftertransients.
la
RMTDo2
FDFtsnastetac
o
aie
DCBisciCe�nrnaci[pnc�nc1
DiTmmeoR
Ta
T
C
b*
FBri
1454 Heart Rhythm, Vol 4, No 11, November 2007
owed by pairwise comparison using the Holm-Sidak test ifppropriate. Fisher’s exact test was used when appropriate.
esultsembrane phospholipidshe fish oil diet contained 1.5 g fish oil �3-PUFAs EPA/HA per 100g feed.9 �3-PUFAs in biopsies of the heart andf isolated myocytes from the pigs fed fish oil composed2% of the total lipid content vs �2% in control (P �.05).
ish oil reduces the incidence of triggered APs,ADs, and Ca2� aftertransientsigure 1 shows typical examples of the effect of fish oil on
he incidence of triggered APs, DADs, and Ca2� aftertran-ients following a rapid pacing protocol in the presence oforepinephrine. The arrows indicate the last stimulated APnd Ca2� transient of the rapid pacing protocol after whichtimulation was discontinued. In the control myocyte, ariggered AP (star) follows a DAD, whereas no triggeredvents occur in the �3 myocyte. The last stimulated Ca2�
ransient is followed by two Ca2� aftertransients (stars)fter the last stimulated Ca2� transient in the control myo-yte but not in the �3 myocyte.
The relative incidences of triggered APs and DADs andf Ca2� aftertransients (recorded from different myocytes)
able 1 Occurrence of triggered APs, DADs, and Ca2�
ftertransients in control and �3 myocytes
Control �3
riggered APs andDADs (%)
84.6(n � 13, N � 7)
9.1*(n � 11, N � 7)
a2� aftertransients (%) 53.8(n � 13, N � 3)
12.5*(n � 16, N � 4)
Values are expressed as percent of number of myocytes.AP � action potential; DAD � delayed afterdepolarization; n � num-
er of myocytes; N � number of pigs.P �.05.
igure 2 A: Typical example of Ca2� transients recorded from a control: Averaged diastolic and systolic [Ca2�]i in the absence and presence of no
ecorded from a control and an �3 myocyte in the absence (�NE) and prese
n control (n � 9) and �3 (n � 8) myocytes. *P �.05 for �NE vs �NE.re summarized In Table 1. They were significantly reducedn �3 myocytes compared to control myocytes (Fisher’sxact test, P �.05).
ietary fish oil reduces Ca2� overload and SRa2� content in response to norepinephrineecause Ca2� aftertransients and DADs reflect disturbances
n [Ca2�]i handling, our experiments imply that the re-ponse of [Ca2�]i to norepinephrine is impaired in �3 myo-ytes compared with control myocytes. Therefore, we stud-ed the effect of norepinephrine on Ca2� transients and SRa2� content in �3 and control myocytes. Figure 2A showsxamples of Ca2� transients recorded from a control and an3 myocyte before (�NE) and after (�NE) application oforepinephrine at 1 Hz. The effect of norepinephrine iseduced in the �3 myocyte. Figure 2B shows that no sig-ificant differences in diastolic and systolic [Ca2�]i in thebsence of norepinephrine between control and �3 myo-ytes in correspondence with our previous data.9 However,n the presence of norepinephrine, diastolic and systolicCa2�]i are significantly higher in the control myocyte com-ared to the �3 myocyte (Figure 2B). In the absence oforepinephrine, SR Ca2� content was not different betweenontrol and �3 myocytes (control: 150 nM � 34.8, n � 10;3: 104 nM � 36.7, n � 9; P � NS). In the presence oforepinephrine, SR Ca2� content was lower in �3 myocytesompared to control (control: 385 nM � 34.8, n � 10; �3:97 nM � 36.7 n � 9; P �.05).
ietary fish oil prevents AP prolongationn response to norepinephrinehe lower [Ca2�]i in the presence of norepinephrine in �3yocytes can be attributed to an increased efflux of Ca2�
ediated by a longer diastolic intervals caused by a short-ned AP. DAD-related triggered activity is also dependentn the duration of the cardiac AP (for review see Wit andosen8). Therefore, we hypothesized that �3 myocytes have
�3 myocyte in the absence (�NE) and presence (�NE) of norepinephrine.hrine. *P �.05 for control vs �3. C: Typical examples of action potentials
NE) of norepinephrine. D: Dependence of APD on stimulation frequency
and anrepinepnce (�
90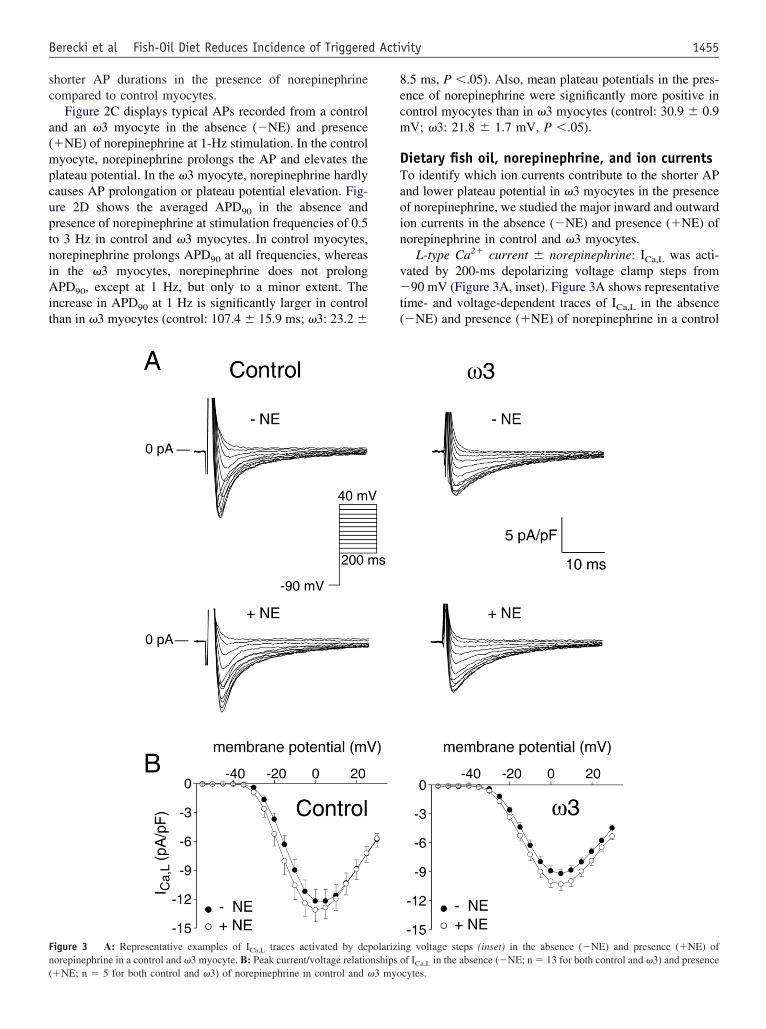
sc
a(mpcuptniAit
8ecm
DTaoin
v�t(
Fn(
1455Berecki et al Fish-Oil Diet Reduces Incidence of Triggered Activity
horter AP durations in the presence of norepinephrineompared to control myocytes.
Figure 2C displays typical APs recorded from a controlnd an �3 myocyte in the absence (�NE) and presence�NE) of norepinephrine at 1-Hz stimulation. In the controlyocyte, norepinephrine prolongs the AP and elevates the
lateau potential. In the �3 myocyte, norepinephrine hardlyauses AP prolongation or plateau potential elevation. Fig-re 2D shows the averaged APD90 in the absence andresence of norepinephrine at stimulation frequencies of 0.5o 3 Hz in control and �3 myocytes. In control myocytes,orepinephrine prolongs APD90 at all frequencies, whereasn the �3 myocytes, norepinephrine does not prolongPD90, except at 1 Hz, but only to a minor extent. The
ncrease in APD90 at 1 Hz is significantly larger in controlhan in �3 myocytes (control: 107.4 � 15.9 ms; �3: 23.2 �
igure 3 A: Representative examples of ICa,L traces activated by deporepinephrine in a control and �3 myocyte. B: Peak current/voltage relatio
�NE; n � 5 for both control and �3) of norepinephrine in control and �3 myo.5 ms, P �.05). Also, mean plateau potentials in the pres-nce of norepinephrine were significantly more positive inontrol myocytes than in �3 myocytes (control: 30.9 � 0.9V; �3: 21.8 � 1.7 mV, P �.05).
ietary fish oil, norepinephrine, and ion currentso identify which ion currents contribute to the shorter APnd lower plateau potential in �3 myocytes in the presencef norepinephrine, we studied the major inward and outwardon currents in the absence (�NE) and presence (�NE) oforepinephrine in control and �3 myocytes.
L-type Ca2� current � norepinephrine: ICa,L was acti-ated by 200-ms depolarizing voltage clamp steps from90 mV (Figure 3A, inset). Figure 3A shows representative
ime- and voltage-dependent traces of ICa,L in the absence�NE) and presence (�NE) of norepinephrine in a control
ng voltage steps (inset) in the absence (�NE) and presence (�NE) off I in the absence (�NE; n � 13 for both control and �3) and presence
olarizinships o
Ca,Lcytes.
av(idpaa�w7
r4ivp
cmdsct4Nm
osan
ha
FBc
1456 Heart Rhythm, Vol 4, No 11, November 2007
nd �3 myocyte. Figure 3B shows the average current/oltage (I/V) relationship of ICa,L in control (left) and �3right) in the absence and presence of norepinephrine. Bothn the absence and presence of norepinephrine, peak ICa,L
ensities at 0 mV are larger in control myocytes com-ared to �3 myocytes [control: �12 � 1.2 pA/pF (�NE)nd �13 � 1.2 pA/pF (�NE); �3: �9 � 0.5 pA/pF (�NE)nd �10 � 0.6 pA/pF (�NE), P �.05 for control vs3]. The relative norepinephrine-induced increase in ICa,L
as similar in the two groups (control: 12%.0 � 6.3%; �3:.5% � 7.0%, P � NS).
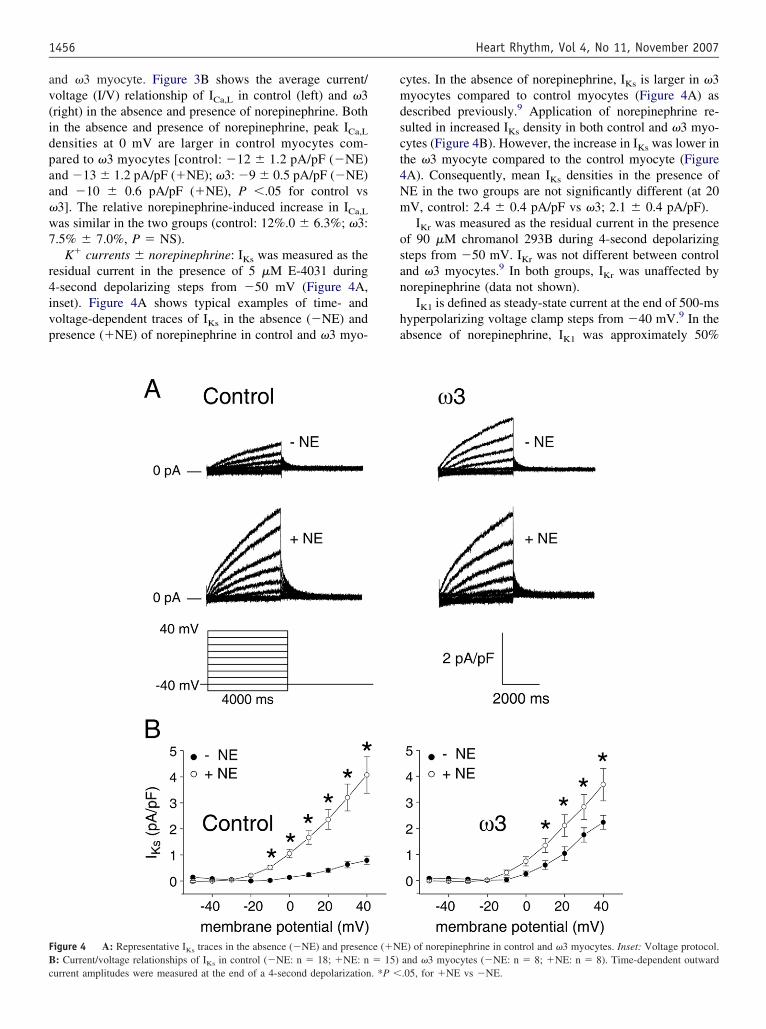
K� currents � norepinephrine: IKs was measured as theesidual current in the presence of 5 �M E-4031 during-second depolarizing steps from �50 mV (Figure 4A,nset). Figure 4A shows typical examples of time- andoltage-dependent traces of IKs in the absence (�NE) andresence (�NE) of norepinephrine in control and �3 myo-
igure 4 A: Representative IKs traces in the absence (�NE) and presenc: Current/voltage relationships of I in control (�NE: n � 18; �NE: n
Ksurrent amplitudes were measured at the end of a 4-second depolarization. *P �
ytes. In the absence of norepinephrine, IKs is larger in �3yocytes compared to control myocytes (Figure 4A) as
escribed previously.9 Application of norepinephrine re-ulted in increased IKs density in both control and �3 myo-ytes (Figure 4B). However, the increase in IKs was lower inhe �3 myocyte compared to the control myocyte (FigureA). Consequently, mean IKs densities in the presence ofE in the two groups are not significantly different (at 20V, control: 2.4 � 0.4 pA/pF vs �3; 2.1 � 0.4 pA/pF).IKr was measured as the residual current in the presence
f 90 �M chromanol 293B during 4-second depolarizingteps from �50 mV. IKr was not different between controlnd �3 myocytes.9 In both groups, IKr was unaffected byorepinephrine (data not shown).
IK1 is defined as steady-state current at the end of 500-msyperpolarizing voltage clamp steps from �40 mV.9 In thebsence of norepinephrine, IK1 was approximately 50%
E) of norepinephrine in control and �3 myocytes. Inset: Voltage protocol.and �3 myocytes (�NE: n � 8; �NE: n � 8). Time-dependent outward
e (�N� 15)
.05, for �NE vs �NE.

lI
stdp
mcsstra
T
I
AA
*
FB�
1457Berecki et al Fish-Oil Diet Reduces Incidence of Triggered Activity
arger due to the fish oil diet.9 Norepinephrine did not alter
K1 density (data not shown).INCX � norepinephrine: INCX was measured as the Ni2�-
ensitive current during a descending voltage ramp pro-ocol. Figure 5A shows representative time- and voltage-ependent traces of INCX in the absence (�NE) andresence (�NE) of norepinephrine in a control and an �3
able 2 INCX densities in the absence (�NE) and presence (�N
NCX
Control
�NE (n � 8, N � 3) �NE (n
t �40 mV (pA/pF) 1.3 � 0.1 2.7 �t �60 mV (pA/pF) �0.7 � 0.1 �2.0 �
Values are given as mean � SEM.I � Na�/Ca2� exchange current; n � number of myocytes; N � nu
igure 5 A: Representative INCX traces in the absence (�NE) and presen: Average current/voltage relationships of INCX in control (�NE: n � 8;NE.
NCX
P �.05 for �3 vs the corresponding control.
yocyte. Without norepinephrine, INCX is larger in theontrol myocyte compared to the �3 myocyte, as de-cribed previously.9 Application of norepinephrine re-ulted in significantly larger INCX in the control myocyteshan in the �3 myocyte. In the presence of norepineph-ine, INCX density values for both the reverse (outward)nd forward (inward) modes were significantly larger in
norepinephrine in control and �3 myocytes
�3
� 4) �NE (n � 9, N � 4) �NE (n � 8, N � 2)
0.7 � 0.1* 1.1 � 0.1*0.4 � 0.1* �0.5 � 0.1*
f pigs; NE � norepinephrine.
E) of norepinephrine in control and �3 myocytes. Inset: Voltage protocol.n � 8) and �3 myocytes (�NE: n � 9; �NE: n � 8). *P �.05, �NE vs
E) of
� 8, N
0.30.4
mber o
ce (�N�NE:
cF
isc5artFw(
DOtsF
pnfiIIncosno
pstoaF
FVI ps.
1458 Heart Rhythm, Vol 4, No 11, November 2007
ontrol myocytes compared to �3 myocytes (P �.05,igure 5B and Table 2).
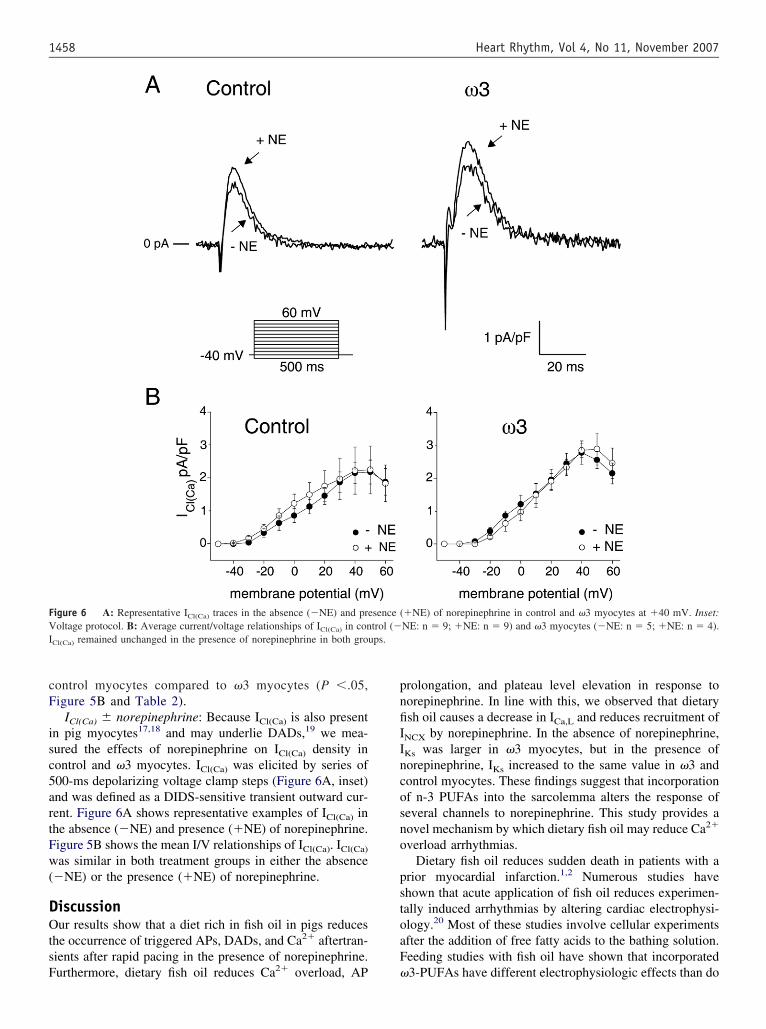
ICl(Ca) � norepinephrine: Because ICl(Ca) is also presentn pig myocytes17,18 and may underlie DADs,19 we mea-ured the effects of norepinephrine on ICl(Ca) density inontrol and �3 myocytes. ICl(Ca) was elicited by series of00-ms depolarizing voltage clamp steps (Figure 6A, inset)nd was defined as a DIDS-sensitive transient outward cur-ent. Figure 6A shows representative examples of ICl(Ca) inhe absence (�NE) and presence (�NE) of norepinephrine.igure 5B shows the mean I/V relationships of ICl(Ca). ICl(Ca)
as similar in both treatment groups in either the absence�NE) or the presence (�NE) of norepinephrine.
iscussionur results show that a diet rich in fish oil in pigs reduces
he occurrence of triggered APs, DADs, and Ca2� aftertran-ients after rapid pacing in the presence of norepinephrine.
igure 6 A: Representative ICl(Ca) traces in the absence (�NE) and preoltage protocol. B: Average current/voltage relationships of ICl(Ca) in con
Cl(Ca) remained unchanged in the presence of norepinephrine in both grou
urthermore, dietary fish oil reduces Ca2� overload, AP �
rolongation, and plateau level elevation in response toorepinephrine. In line with this, we observed that dietarysh oil causes a decrease in ICa,L and reduces recruitment of
NCX by norepinephrine. In the absence of norepinephrine,
Ks was larger in �3 myocytes, but in the presence oforepinephrine, IKs increased to the same value in �3 andontrol myocytes. These findings suggest that incorporationf n-3 PUFAs into the sarcolemma alters the response ofeveral channels to norepinephrine. This study provides aovel mechanism by which dietary fish oil may reduce Ca2�
verload arrhythmias.Dietary fish oil reduces sudden death in patients with a
rior myocardial infarction.1,2 Numerous studies havehown that acute application of fish oil reduces experimen-ally induced arrhythmias by altering cardiac electrophysi-logy.20 Most of these studies involve cellular experimentsfter the addition of free fatty acids to the bathing solution.eeding studies with fish oil have shown that incorporated
�NE) of norepinephrine in control and �3 myocytes at �40 mV. Inset:NE: n � 9; �NE: n � 9) and �3 myocytes (�NE: n � 5; �NE: n � 4).
sence (trol (�
3-PUFAs have different electrophysiologic effects than do

asom
TWiact[fioldr2nblicffiCcciSmcntArFic
AnUcPrcnciiarcrr
ItItI�dcIspccpA
ItrDb
nccrtpgm
STeptdstiotwaf
caeocpCep
1459Berecki et al Fish-Oil Diet Reduces Incidence of Triggered Activity
cutely administered �3-PUFAs.9,21,22 Our study was de-igned to investigate the effect of incorporated �3-PUFAsn norepinephrine-induced triggered activity in isolatedyocytes from pigs fed fish oil.
riggered activitye demonstrated that incorporated �3-PUFAs reduced the
ncidence of norepinephrine-induced triggered APs, DADs,nd Ca2� aftertransients. Elevation of diastolic Ca2� con-entration is directly related to the occurrence Ca2� after-ransients in heart failure.16,23 Indeed, diastolic and systolicCa2�]i in the presence of norepinephrine were lower in thesh oil group compared to control. A reduction in Ca2�
verload in the presence of norepinephrine may be due toess Ca2� influx, more Ca2� efflux, altered SR Ca2� han-ling, and/or differences in phosphorylation of ryanodineeceptors.16,24 Incorporated �3-PUFAs reduced ICa,L by0%9 with no difference in [Ca2�]i. In the presence oforepinephrine, ICa,L was increased to a similar extent inoth �3 and control myocytes (Figure 3), suggesting aimited role for ICa,L in the reduction of norepinephrine-nduced Ca2� overload. Ca2� efflux by INCX in �3 myo-ytes also was reduced compared to the control and there-ore could not contribute to increased Ca2� removal. Thisnding suggests that the explanation for the reduction ina2� overload in response to norepinephrine in �3 myo-ytes is mainly on the level of the SR. Indeed, SR Ca2�
ontent was lower in the �3 myocytes compared to controln the presence, but not in the absence, of norepinephrine.imilar to our finding in the absence of norepinephrine,embrane-incorporated �3-PUFAs did not affect SR Ca2�
ontent in rat myocytes.25 Our findings in the presence oforepinephrine imply that differences in phosphorylation ofhe ryanodine receptors and, for example, the SR Ca2�
TPase (SERCA) in �3 myocytes could account for theeduction in Ca2� overload and the lower SR Ca2� content.urther experiments are needed to establish the effect of
ncorporated �3-PUFAs on phosphorylation of receptors,hannels, and exchangers.
P changes in �3 and control myocytes duringorepinephrine exposurender control conditions, �3 myocytes have shorter APs
ompared to control myocytes due to incorporation of �3-UFAs into the sarcolemma.9 Application of norepineph-ine to control myocytes resulted in prolongation of theardiac AP and elevation of the action potential plateau, butot in �3 myocytes. The duration and the shape of theardiac AP is determined by a delicate balance betweennward and outward currents; therefore, small changes inon current densities can have a major impact on AP shapend duration. AP prolongation in response to norepineph-ine is another prominent feature of human ventricular myo-ytes.11 The lack of AP prolongation in �3 myocytes inesponse to norepinephrine can be explained by the reduced
ecruitment of individual ion currents. fion currents in �3 and control myocytes andheir response to norepinephrinencorporated �3-PUFAs caused a 60% reduction in INCX inhe absence of norepinephrine.9 Norepinephrine increased
NCX in both control and �3 myocytes, but much less in the3 myocytes (Figure 5). Because INCX is mostly inwarduring the repolarization phase of the cardiac AP, the de-reased INCX in �3 myocytes will lead to reduced inward
NCX during plateau and repolarization of the AP. The re-ulting smaller inward current could explain the reduced AProlongation in the �3 myocytes. In addition, the lowerytosolic [Ca2�]i in response to norepinephrine in �3 myo-ytes leads to even more reduced INCX and prevents AProlongation. Therefore, NCX is a key factor in the reducedP prolongation in �3 myocytes.Because INCX underlies generation of DADs, the reduced
NCX in �3 myocytes may also explain the reduction inriggered activity seen in our study. Any spontaneous Ca2�
elease in �3 myocytes results in a decreased incidence ofADs. The larger IK1 aids in stabilizing the resting mem-rane potential.9
Incorporated �3-PUFAs increased IKs by 70%.9 Norepi-ephrine enhanced IKs density in both control and �3 myo-ytes (Figure 4). The increase was much less in �3 myo-ytes, indicating that incorporated �3-PUFAs alter theesponse of the channel to norepinephrine. Consequently,he difference in IKs density between the two groups in theresence of norepinephrine was abolished (Figure 4), sug-esting a limited role in preventing AP prolongation in �3yocytes.
tudy implicationshe importance of the �-adrenergic system in the pathogen-sis and treatment of heart failure is well established, andharmacologic �-blockade confers survival benefit in pa-ients after myocardial infarction.26 Catecholamine-depen-ent arrhythmias, such as those associated with long QTyndrome and catecholaminergic polymorphic ventricularachycardia, are triggered by DADs that may degeneratento ventricular fibrillation.27–29 Our study supports fishil supplementation for catecholamine-induced ventricularachycardia (in postmyocardial infarction patients or thoseith long QT syndrome type 1) and provides a novel anti-
rrhythmic mechanism of fish oil in patients with heartailure and/or long QT syndrome.
A reduction in cytosolic calcium could further impairontractile function in patients with heart failure. However,study in monkeys showed that myocardial function is
nhanced rather than impaired after a prolonged diet of fishil.30 In addition, a cellular study on the effect of DHA onontractions and ICa,L showed that myocardial function wasreserved, although DHA did block ICa,L and Ca2�-induceda2� release.31 Therefore, we suppose that a detrimentalffect of fish oil on mechanical function in heart failureatients is limited, although further research on the safety of
sh oil supplementation in heart failure patients is needed.
CDascg�iTtDh
AWt
R
1
1
1
1
1
1
1
1
1
1
2
2
2
2
2
2
2
2
2
2
3
3
1460 Heart Rhythm, Vol 4, No 11, November 2007
onclusionietary fish oil reduces the incidence of triggered activity
nd prevents Ca2� overload and AP prolongation in re-ponse to norepinephrine. This corresponds with a de-reased ICa,L and reduced recruitment INCX in the fish oilroup. In the absence of norepinephrine, IKs was larger in3 myocytes, but in the presence of norepinephrine, IKs
ncreased to the same level in �3 and control myocytes.hese findings suggest that incorporated �3-PUFAs alter
he response of several ion channels to norepinephrine.ietary fish oil may prevent arrhythmias in patients witheart failure and/or the long QT syndrome.
cknowledgmentse thank Charly N.W. Belterman and Wim L. ter Smitte for
heir valuable help.
eferences1. Burr ML, Gilbert JF, Holliday RM, Elwood PC, Fehily AM, Rogers S, Sweet-
nam PM, Deadman NM. Effects of changes in fat, fish, and fibre intakes on deathand myocardial reinfarction: Diet And Reinfarction Trial (DART). Lancet 1989;334:757–761.
2. GISSI-Prevenzione Investigators. Dietary supplementation with n-3 polyunsat-urated fatty acids and vitamin E after myocardial infarction: results of theGISSI-Prevenzione trial. Lancet 1999;354:447–455.
3. Kimura S, Bassett AL, Kohya T, Kozlovskis PL, Myerburg RJ. Automaticity,triggered activity, and responses to adrenergic stimulation in cat subendocardialPurkinje fibers after healing of myocardial infarction. Circulation 1987;75:651–660.
4. Qin D, Zhang ZH, Caref EB, Boutjdir M, Jain P, El Sherif N. Cellular and ionicbasis of arrhythmias in postinfarction remodeled ventricular myocardium. CircRes 1996;79:461–473.
5. Verkerk AO, Veldkamp MW, Baartscheer A, Schumacher CA, Klopping C, vanGinneken ACG, Ravesloot JH. Ionic mechanism of delayed afterdepolarizationsin ventricular cells isolated from human end-stage failing hearts. Circulation2001;104:2728–2733.
6. Kass RS, Lederer WJ, Tsien RW, Weingart R. Role of calcium ions in transientinward currents and aftercontractions induced by strophanthidin in cardiacPurkinje fibres. J Physiol 1978;281:187–208.
7. Sipido KR, Bito V, Antoons G, Volders PG, Vos MA. Na/Ca exchange andcardiac ventricular arrhythmias. Ann N Y Acad Sci 2007;1099:348.
8. Wit AL, Rosen MR. Afterdepolarizations and triggered activity: distinction fromautomaticity as an arrhythmogenic mechanism. In: Fozzard HA, Haber E,Jennings RB, Katz AM, Morgan HE, The Heart and Cardiovascular System.Second edition. New York: Raven Press, 1992:2113–2163.
9. Verkerk AO, van Ginneken ACG, Berecki G, Den Ruijter HM, Schumacher CA,Veldkamp MW, Baartscheer A, Casini S, Opthof T, Hovenier R, Fiolet JWT,Zock PL, Coronel R. Incorporated sarcolemmal fish oil fatty acids shorten pigventricular action potentials. Cardiovasc Res 2006;70:509–520.
0. Nattel S, Maguy A, Le Bouter S, Yeh YH. Arrhythmogenic ion-channel remod-eling in the heart: Heart failure, myocardial infarction, and atrial fibrillation.Physiol Rev 2007;87:425–456.
1. Veldkamp MW, Verkerk AO, van Ginneken ACG, Baartscheer A, SchumacherC, de Jonge N, de Bakker JMT, Opthof T. Norepinephrine induces actionpotential prolongation and early afterdepolarizations in ventricular myocytes
isolated from human end-stage failing hearts. Eur Heart J 2001;22:955–963.2. Verkerk AO, Tan HL, Ravesloot JH. Ca2�-activated Cl� current reduces trans-mural electrical heterogeneity within the rabbit left ventricle. Acta PhysiolScand 2004;180:239–247.
3. Bosch RF, Gaspo R, Busch AE, Lang HJ, Li GR, Nattel S. Effects of thechromanol 293B, a selective blocker of the slow, component of the delayedrectifier K� current, on repolarization in human and guinea pig ventricularmyocytes. Cardiovasc Res 1998;38:441–450.
4. Hinde AK, Perchenet L, Hobai IA, Levi AJ, Hancox JC. Inhibition of Na/Caexchange by external Ni in guinea-pig ventricular myocytes at 37°C, dialysedinternally with cAMP-free and cAMP-containing solutions. Cell Calcium 1999;25:321–331.
5. Verkerk AO, Schumacher CA, van Ginneken AC, Veldkamp MW, RaveslootJH. Role of Ca2�-activated Cl� current in ventricular action potentials of sheepduring adrenoceptor stimulation. Exp Physiol 2001;86:151–159.
6. Baartscheer A, Schumacher CA, Belterman CNW, Coronel R, Fiolet JWT. SRcalcium handling and calcium after-transients in a rabbit model of heart failure.Cardiovasc Res 2003;58:99–108.
7. Li GR, Du XL, Siow YL, O K, Tse HF, Lau CP. Calcium-activated transientoutward chloride current and phase 1 repolarization of swine ventricular actionpotential. Cardiovasc Res 2003;58:89–98.
8. Li GR, Sun H, To J, Tse HF, Lau CP. Demonstration of calcium-activatedtransient outward chloride current and delayed rectifier potassium currents inswine atrial myocytes. J Moll Cell Cardiol 2004;36:495–504.
9. Verkerk AO, Veldkamp MW, Bouman LN, van Ginneken ACG. Calcium-activated Cl� current contributes to delayed afterdepolarizations in single Pur-kinje and ventricular myocytes. Circulation 2000;101:2639–2644.
0. Xiao YF, Sigg DC, Leaf A. The antiarrhythmic effect of n-3 polyunsaturatedfatty acids: modulation of cardiac ion channels as a potential mechanism. JMembr Biol 2005;206:141–154.
1. Den Ruijter HM, Berecki G, Opthof T, Verkerk AO, Zock PL, Coronel R. Pro-and antiarrhythmic properties of a diet rich in fish oil. Cardiovasc Res 2007;73:316–325.
2. Leifert WR, McMurchie EJ, Saint DA. Inhibition of cardiac sodium currents inadult rat myocytes by n-3 polyunsaturated fatty acids. J Physiol 1999;520:671–679.
3. Janse MJ. Electrophysiological changes in heart failure and their relationship toarrhythmogenesis. Cardiovasc Res 2004;61:208–217.
4. Zahradnikova A, Zahradnik I. Analysis of calcium-induced calcium release incardiac sarcoplasmic reticulum vesicles using models derived from single-channel data. Biochim Biophys Acta 1999;1418:268–284.
5. Leifert WR, Dorian CL, Jahangiri A, McMurchie EJ. Dietary fish oil preventsasynchronous contractility and alters Ca2� handling in adult rat cardiomyocytes.J Nutr Biochem 2001;12:365–376.
6. Rubart M, Zipes DP. Mechanisms of sudden cardiac death. J Clin Invest2005;115:2305–2315.
7. Marks AR, Priori S, Memmi M, Kontula K, Laitinen PJ. Involvement of thecardiac ryanodine receptor/calcium release channel in catecholaminergic poly-morphic ventricular tachycardia. J Cell Physiol 2002;190:1–6.
8. Paavola J, Viitasalo M, Laitinen-Forsblom PJ, Pasternack M, Swan H, TikkanenI, Toivonen L, Kontula K, Laine M. Mutant ryanodine receptors in cat-echolaminergic polymorphic ventricular tachycardia generate delayed afterde-polarizations due to increased propensity to Ca2� waves. Eur Heart J 2007;28:1135–1142.
9. Shimizu W, Antzelevitch C. Cellular basis for the ECG features of the LQT1form of the long-QT syndrome: effects of �-adrenergic agonists and antagonistsand sodium channel blockers on transmural dispersion of repolarization andTorsade de Pointes. Circulation 1998;98:2314–2322.
0. McLennan PL, Barnden LR, Bridle TM, Abeywardena MY, Charnock JS.Dietary fat modulation of left ventricular ejection fraction in the marmoset dueto enhanced filling. Cardiovasc Res 1992;26:871–877.
1. Ferrier GR, Redondo I, Zhu J, Murphy MG. Differential effects of docosahexae-2�
noic acid on contractions and L-type Ca current in adult cardiac myocytes.Cardiovasc Res 2002;54:601–610.


















