
Developmental cross talking in the ectomycorrhizal symbiosis: signals and communication genes
10
© New Phytologist (2001) 151 : 145 – 154 www.newphytologist.com 145 Review Blackwell Science Ltd Research review Developmental cross talking in the ectomycorrhizal symbiosis: signals and communication genes Francis Martin, Sébastien Duplessis, Frank Ditengou, Hubert Lagrange, Catherine Voiblet and Frédéric Lapeyrie Equipe de Microbiologie Forestière, Institut National de la Recherche Agronomique, Centre de Recherches de Nancy, 54280 Champenoux, France Author for correspondence Francis Martin Tel: + 33 383 39 40 89 Fax: + 33 383 39 40 69 Email: [email protected] Received: 20 November 2000 Accepted: 22 March 2001 Summary Development of ectomycorrhizas involves multiple genes that are implicated in a complex series of interdependent, sequential steps. Current research into ectomy- corrhiza development and functioning is aimed at understanding this plant–microbe interaction in a framework of the developmental and physiological processes that underlie colonization and morphogenesis. After a brief introduction to the ectomy- corrhizal symbiosis, the present article highlights recent work on the early signal exchange taking place between symbionts, and sketches the way functional geno- mics is altering our thinking about changes in gene expression during the early steps of the ectomycorrhiza development. © New Phytologist (2001) 151 : 145 –154 Key words: auxins, cytoskeleton, gene expression, hypaphorine, rhizosphere, root hairs, transduction pathways. Introduction One of the principal ecological niches of fungi is the inside of plant and animal tissues and cells. Although the majority of associations between fungi and eukaryotes is either commensal or, quite often, mutually beneficial, most studies of animal- and plant-associated fungi have dealt with those rare species that cause diseases. Our major interest focuses on ectomy- corrhizal fungi, which, unlike pathogenic parasites, have been associated with their host plant for an evolutionary length of time. The ectomycorrhizal symbiosis has evolved repeatedly over the last 130–180 Myr (LePage et al. , 1997) and has had major consequences for the diversification of both the myco- bionts and their hosts (Hibbett et al. , 2000). Ectomycorrhizal fungi mainly belong to the Basidiomycotina (agarics, bolets) (Fig. 1a), although many species are found within the Ascomycotina (truffles) and the switch between saprobic and mycorrhizal lifestyles probably happened convergently, and perhaps many times, during evolution of these fungal line- ages (Hibbett et al. , 2000). The first mycorrhizal associations must have been derived from earlier types of plant–fungus interactions, such as endophytic fungi in the bryophyte- like precursors of vascular plants (Selosse & Le Tacon, 1998). Ectomycorrhizal symbioses have a distinct host range allowing formation of ectomycorrhiza on a limited set of trees and shrubs (Smith & Read, 1997). Some of these are shown in Fig. 1(b). However, a given species of ectomycorrhizal fungus is usually able to establish a mutualistic symbiosis on a broad range of species, although highly specific interactions may occur (e.g. Rhizopogon vinicolor/Pseudotsuga mensiesii ). In temperate and boreal forests, up to 95% of the short roots form ectomycorrhizas. Ectomycorrhizas have a benefi- cial impact on plant growth in natural (Read, 1991) and agro- forestry ecosystems (Grove & Le Tacon, 1993). Central to the success of these symbioses is the exchange of nutrients between the symbionts (Smith & Read, 1997). The fungus
-
Upload
francis-martin -
Category
Documents
-
view
212 -
download
0
Transcript of Developmental cross talking in the ectomycorrhizal symbiosis: signals and communication genes

©
New Phytologist
(2001)
151
: 145–154
www.newphytologist.com
145
Review
Blackwell Science Ltd
Research review
Developmental cross talking in the ectomycorrhizal symbiosis: signals
and communication genes
Francis Martin, Sébastien Duplessis, Frank Ditengou, Hubert Lagrange, Catherine Voiblet and Frédéric Lapeyrie
Equipe de Microbiologie Forestière, Institut National de la Recherche Agronomique, Centre de
Recherches de Nancy, 54280 Champenoux, France
Author for correspondence
Francis Martin Tel:
+
33 383 39 40 89 Fax:
+
33 383 39 40 69 Email: [email protected]
Received:
20 November 2000
Accepted:
22 March 2001
Summary
Development of ectomycorrhizas involves multiple genes that are implicated in acomplex series of interdependent, sequential steps. Current research into ectomy-corrhiza development and functioning is aimed at understanding this plant–microbeinteraction in a framework of the developmental and physiological processes thatunderlie colonization and morphogenesis. After a brief introduction to the ectomy-corrhizal symbiosis, the present article highlights recent work on the early signalexchange taking place between symbionts, and sketches the way functional geno-mics is altering our thinking about changes in gene expression during the early stepsof the ectomycorrhiza development.
©
New Phytologist
(2001)
151
: 145–154
Key words:
auxins, cytoskeleton, gene expression, hypaphorine, rhizosphere, root hairs, transduction pathways.
Introduction
One of the principal ecological niches of fungi is the inside ofplant and animal tissues and cells. Although the majority ofassociations between fungi and eukaryotes is either commensalor, quite often, mutually beneficial, most studies of animal-and plant-associated fungi have dealt with those rare speciesthat cause diseases. Our major interest focuses on ectomy-corrhizal fungi, which, unlike pathogenic parasites, have beenassociated with their host plant for an evolutionary length oftime. The ectomycorrhizal symbiosis has evolved repeatedlyover the last 130–180 Myr (LePage
et al.
, 1997) and has hadmajor consequences for the diversification of both the myco-bionts and their hosts (Hibbett
et al.
, 2000). Ectomycorrhizalfungi mainly belong to the Basidiomycotina (agarics, bolets)(Fig. 1a), although many species are found within theAscomycotina (truffles) and the switch between saprobic andmycorrhizal lifestyles probably happened convergently, and
perhaps many times, during evolution of these fungal line-ages (Hibbett
et al.
, 2000). The first mycorrhizal associationsmust have been derived from earlier types of plant–fungusinteractions, such as endophytic fungi in the bryophyte-like precursors of vascular plants (Selosse & Le Tacon,1998). Ectomycorrhizal symbioses have a distinct host rangeallowing formation of ectomycorrhiza on a limited set of treesand shrubs (Smith & Read, 1997). Some of these are shownin Fig. 1(b). However, a given species of ectomycorrhizal fungusis usually able to establish a mutualistic symbiosis on a broadrange of species, although highly specific interactions mayoccur (e.g.
Rhizopogon vinicolor/Pseudotsuga mensiesii
).In temperate and boreal forests, up to 95% of the short
roots form ectomycorrhizas. Ectomycorrhizas have a benefi-cial impact on plant growth in natural (Read, 1991) and agro-forestry ecosystems (Grove & Le Tacon, 1993). Central tothe success of these symbioses is the exchange of nutrientsbetween the symbionts (Smith & Read, 1997). The fungus
NPH169.fm Page 145 Tuesday, May 29, 2001 5:12 PM

Research review
www.newphytologist.com
©
New Phytologist
(2001)
151
: 145–154
Review146
gains carbon from the plant while plant nutrient uptake ismediated via the fungus. In addition, the establishment ofthe symbiosis is required for the completion of the fungallife cycle (i.e. formation of fruiting bodies).
Differences in organogenetic programmes are reflected invariations in ectomycorrhizal root morphology, but overallthere are pronounced developmental similarities suggestingthat key developmental programmes are triggered in bothsymbionts. In all cases, the mycobiont must have the abilityto recognize and become associated with its host, escape thehost defense surveillance, and establish bi-directional nutri-ent transfers. Ectomycorrhiza development thus involvesmultiple genes that are implicated in a complex series ofinterdependent, sequential steps. Current research intoectomycorrhiza development and functioning is aimed atunderstanding this plant–microbe interaction in a frameworkof the developmental and physiological processes that underlycolonization and morphogenesis. After a brief introduction tothe ectomycorrhizal symbiosis, the present article aims tohighlight recent work on the early signal exchange takingplace between symbionts, and to sketch the way functionalgenomics is altering our thinking about changes in gene
expression during the early steps of the ectomycorrhizadevelopment.
The anatomy and development of ectomycorrhiza
Ectomycorrhizas are characterized structurally by the presenceof a dense mass of fungal hyphae forming a pseudoparen-chymatous tissue ensheathing the root (Fig. 1c). This is theHartig net of intercellular hyphæ, characterized by labyrinth-ine branching and an outward network of hyphae prospectingthe soil and gathering nutrients (Kottke & Oberwinkler,1987). The mantle of fungal tissue surrounding the hostlateral roots varies in composition from the characteristicpseudoparenchymatous tissue to a rather open-wefted arrange-ment of hyphae. Development of a mature mantle proceedsthrough a programmed series of events (Horan
et al.
, 1988;Martin
et al.
, 1997). Fungal hyphae originating from a soilpropagule or an older mycorrhiza penetrate into the root capcells and grow through them. Backwards from the tip theinvasion of root cap cells proceeds inwards until the hyphaereach the epidermal cells (Chilvers, 1968; Feugey
et al.
, 1999).The root cap tissue is progressively transformed by invasion
Fig. 1 The ectomycorrhizal symbiosis. (a) Boletus edulis, one of the numerous Basidiomycotina able to establish a mutualistic symbiosis with host trees (Photograph courtesy of F. Martin). (b) A seedling of Douglas fir (Pseudotsuga menziesii) colonized by the ectomycorrhizal basidiomycete Laccaria bicolor. The fungal mycelium has developed ectomycorrhizas on the root system and has produced a basidiocarp above ground (Photograph courtesy of P. Klett-Frey). (c) Short roots ensheathed by an ectomycorrhizal fungus. The yellow mantle covers the root tip and rhizomorphs (i.e. hyphal bundles) are extending in the medium (Photograph courtesy of J. Garbaye). (d) Transverse section of a Eucalyptus/Pisolithus ectomycorrhiza showing the external (EM) and internal (IM) mantles: the fungal hyphae have begun to penetrate between the epidermal cells of the root cortex (RC) to form the Hartig net (HN). Epidermal cells are radially enlarged. Extramatrical hyphae (EH) are exploring the medium (Photograph courtesy of G. Chevalier).
NPH169.fm Page 146 Tuesday, May 29, 2001 5:12 PM

Research review
©
New Phytologist
(2001)
151
: 145–154
www.newphytologist.com
Review 147
and dismemberment of the cells into the inner layers of themantle. Mantle formation therefore commences selectively inthe apical root region (Horan
et al.
1988). After attachmentonto epidermal cells, hyphae multiply to form a series oflayers. These aggregating hyphae, which could develop intoa biofilm of several hundred µm thick, then differentiateto form the mature mantle. The hyphae in these structuresare encased in an extracellular polysaccharide and proteinacousmatrix. Air and water channels that allow the flow of nutrientsinto the symbiosis innervate these structures (Ashford
et al.
,1989). There is a structural (Paris
et al.
, 1993) and physi-ological (Cairney & Burke, 1996) heterogeneity within theectomycorrhizal mantle, and between the mantle and theinward and outward fungal networks. These morphologicalanalyses of the infection process have shown a fairly complexinfluence of the root on the fungus, including a generalgrowth stimulus, a trophic response directing hyphal growthinwards towards the plant tissues and a morphogenetic effectleading to compact hyphal mantle development. In additionto putative morphogens, the supply of nutrients, the presenceof a physical support and the supply of O
2
likely play a role inmantle formation (Read & Armstrong, 1972; Martin
et al.
,1999). On the other hand, fungal hyphae stimulate lateralroot formation, dichotomy of the apical meristem in coniferspecies, and cytodifferentiation (radial elongation, root hairdecay) of root cells (Fig. 1d) (Horan
et al.
, 1988).As stressed above, morphogenesis of ectomycorrhiza
encompasses a series of complex and overlapping ontogenicprocesses in symbionts: switching off the fungal growthmode, initiation of lateral roots, aggregation of hyphæ, arrestof cell division in ensheathed roots, radial elongation of epi-dermal cells (Kottke & Oberwinkler, 1987; Horan
et al.
,1988; Feugey
et al.
, 1999). Morphological differentiation isaccompanied by the onset of novel metabolic organizationsin fungal and plant cells leading to the finished function-ing symbiotic organ (Martin & Botton, 1993; Cairney
et al.
,1989; Rygiewicz & Andersen, 1994; Hampp
et al.
, 1995).The entire purpose of the complex programme of multicellu-lar development in the ectomycorrhizal symbiosis is to extendthe function of the root system (Read, 1991; Smith & Read,1997). Extramatrical hyphae, the mantle and the intraradicu-lar hyphal network are active metabolic entities that provideessential nutrient resources (e.g. phosphate, nitrogen) to thehost plant. The contribution of essential nutrients is reciproc-ated by the provision of a stable carbohydrate-rich niche inthe roots for the fungal partner, making the relationship amutualistic symbiosis. This simple perspective does not dojustice to the challenge of defining this symbiosis, a complexphenotype that is dependent on the interactions between aspectrum of different host and microbial molecules. This mutu-ality of host and microbe, and its implicit coevolutionaryimplications, are key concepts. The distinction betweensaprobic and mycorrhizal behaviour is not a sharp cut-off(Hibbett
et al.
, 2000), but a continuum and must take into
account: ecological factors; variations in host surveillancemechanisms; and evolutionary factors that determine the cost(i.e. fitness) of the symbiosis (Fitter, 1991). The ecologicalperformance of ectomycorrhizal fungi is therefore a complexphenotype affected by many different traits and by environ-mental factors. Identification of the primary factors con-trolling the development of the symbiosis and its metabolicactivity (e.g. nutrient acquisition) will open the door tounderstanding the ecological fitness of the ectomycorrhizalsymbiosis.
Mutual signal exchange fires the developmental process
By contrast to some other plant–microbe interactions(Dénarié
et al.
, 1996; Moller & Chua, 1999), the nature ofthe signalling molecules and the molecular basis of signalperception and transduction in mycorrhiza are unknownor ill-defined. Identifying the processes that regulate theinformation flow between mycorrhizal fungi and host rootis an active research area. Host plants release into therhizosphere critical metabolites that are able to trigger basi-diospore germination (Fries
et al.
, 1987), growth of hyphaetowards the root (Horan & Chilvers, 1990) and the earlydevelopmental steps of mycorrhiza formation (Béguiristain& Lapeyrie, 1997; Salzer
et al.
, 1997a; Ditengou & Lapeyrie,2000). Molecules that control the interactions betweensymbionts can be classified as follows:
•
tropism of hyphae for host tissues (rhizospheric signals);
•
attachment and invasion of host tissues by hyphae(adhesins, hydrolases);
•
induction of organogenetic programmes in both fungal androot cells (hormones and secondary signals);
•
facilitating survival of the mycobiont despite plant defenseresponses;
•
coordinating strategies for exchanging carbon and othermetabolites (e.g. vitamins) for
in planta
colonization and forbalancing growth of the soil fungal web with its role ingathering minerals from the soil.
Rhizospheric signals
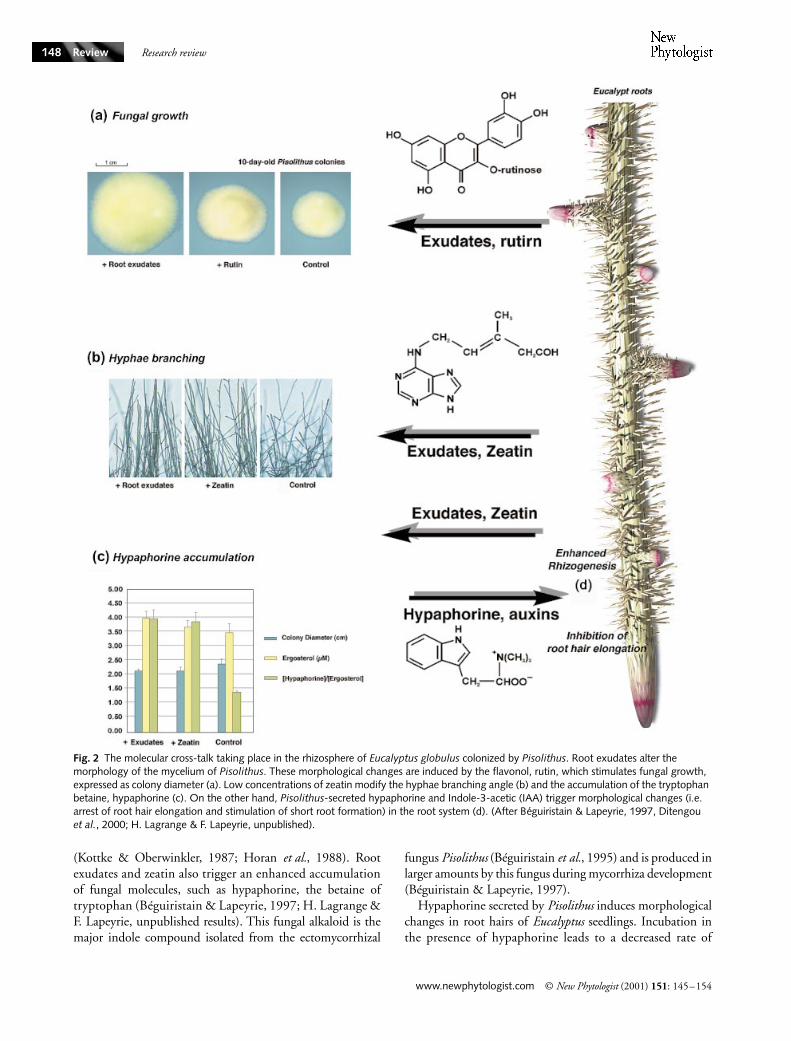
Based on current knowledge of the molecules released in otherplant–microbe interactions, the early plant host signalssecreted into the rhizosphere can include flavonoids, diterpenes,hormones and various nutrients. As shown in Fig. 2, host-released metabolites, such as the flavonol, rutin, and thecytokinin, zeatin, strikingly modified hyphal morphology(Lagrange
et al.
, 2001). When present in the growth med-ium at very low concentrations, rutin stimulated growth of
Pisolithus
(Fig. 2a), whereas the cytokinin, zeatin modified thehyphae branch angle (Fig. 2b). These rhizospheric moleculesare therefore able to induce morphological changes similar tothose observed during actual ectomycorrhizal development
NPH169.fm Page 147 Tuesday, May 29, 2001 5:12 PM
Research review
www.newphytologist.com
©
New Phytologist
(2001)
151
: 145–154
Review148
(Kottke & Oberwinkler, 1987; Horan
et al.
, 1988). Rootexudates and zeatin also trigger an enhanced accumulationof fungal molecules, such as hypaphorine, the betaine oftryptophan (Béguiristain & Lapeyrie, 1997; H. Lagrange &F. Lapeyrie, unpublished results). This fungal alkaloid is themajor indole compound isolated from the ectomycorrhizal
fungus
Pisolithus
(Béguiristain
et al.
, 1995) and is produced inlarger amounts by this fungus during mycorrhiza development(Béguiristain & Lapeyrie, 1997).
Hypaphorine secreted by
Pisolithus
induces morphologicalchanges in root hairs of
Eucalyptus
seedlings. Incubation inthe presence of hypaphorine leads to a decreased rate of
Fig. 2 The molecular cross-talk taking place in the rhizosphere of Eucalyptus globulus colonized by Pisolithus. Root exudates alter the morphology of the mycelium of Pisolithus. These morphological changes are induced by the flavonol, rutin, which stimulates fungal growth, expressed as colony diameter (a). Low concentrations of zeatin modify the hyphae branching angle (b) and the accumulation of the tryptophan betaine, hypaphorine (c). On the other hand, Pisolithus-secreted hypaphorine and Indole-3-acetic (IAA) trigger morphological changes (i.e. arrest of root hair elongation and stimulation of short root formation) in the root system (d). (After Béguiristain & Lapeyrie, 1997, Ditengou et al., 2000; H. Lagrange & F. Lapeyrie, unpublished).
NPH169.fm Page 148 Tuesday, May 29, 2001 5:12 PM

Research review
©
New Phytologist
(2001)
151
: 145–154
www.newphytologist.com
Review 149
elongation and a transitory swelling of the apex of the roothair (Ditengou
et al.
, 2000). Addition of indole-3-acetic acid(IAA) restores tip growth (Ditengou
et al.
, 2000) indicatingantagonistic effects of hypaphorine and auxin on the observedmorphological changes. Development of root hairs is dividedinto cell fate determination, hair initiation, and hair elonga-tion (Gilroy & Jones, 2000). The latter step is highly polar-ized, involving a narrow zone (< 10 µm) at the tip, where newmembrane and cell wall are built from fusion of secretoryvesicles (Geitmann & Emons, 2000). Growth in root hairs isassociated with an apex-high cytosolic free Ca
2
+
gradientgenerated by a local Ca
2
+
influx at the tip, which probablyhas a crucial role in the regulation of vesicle fusion (Geitmann& Emons, 2000). Whether the hypaphorine-induced cyto-skeleton changes are related to interactions with calciumchannels, cofilin/actin-depolymerizing proteins and/or auxinsignalling pathways (Ditengou
et al.
, 2000) is currently underinvestigation.
Root hairs are a key site for microbial interactions (Peterson& Farquhar, 1996) and it has been suggested that interactionbetween the ectomycorrhizal fungus and root hairs may playa role in the symbiosis development (Thomson
et al.
, 1989).However, this contention has been challenged (Horan
et al.
,1988). Mycorrhizal colonization is initiated in the root capregion and then propagates by an acropetal extension of rootand fungal tissues (Chilvers, 1968; Horan
et al.
, 1988). Onlynew epidermal cells, formed after fungal invasion of the apex,are involved in this infection process. The intercellular networkof hyphae (i.e. the Hartig net) develops only between epider-mal cells that undergo a radial elongation as a result of fungalcontact. In such circumstances, root hairs will not emerge ona colonized root surface, suggesting that their developmenthas been inhibited very early, either mechanically or chemic-ally (e.g. through the action of hypaphorine), by the fungus(Ditengou
et al.
, 2000).
Changes in hormonal balance
In addition to flavonoids, phytohormones, including auxins,cytokinins, abscisic acid and ethylene, are produced byectomycorrhizal fungi (Gogala, 1991). Many studies indicatethat changes in auxin balance are a prerequisite for mycorrhizaorganogenesis (Rupp
et al.
, 1989; Gay
et al.
, 1994; Karabaghli-Degron
et al.
, 1998; Kaska
et al.
, 1999). As can be seen inFig. 2, ectomycorrhizal fungi enhance proliferation of shortroots (Carnero Diaz
et al.
, 1996). In conifers, externally suppliedfungal exudates, extracts, or synthetic auxins partially mimicthe effect of mycelium in inducing root proliferation anddichotomous branching of lateral roots (Smith & Read, 1997).The presence of plant-derived tryptophan in the rhizospherecould be sufficient for ectomycorrhizal fungi to enhance thebiosynthesis of fungal IAA (Rupp
et al.
, 1989). However, recentstudies have shown that tryptophan synthase is dispensablefor auxin production by the nonmycorrhizal ascomycete
Aspergillus nidulans
and that auxin is not produced fromindole, an intermediate of the tryptophan synthase reaction(Eckert
et al.
, 2000). Pine inoculated with mutant strains of
Hebeloma cylindrosporum
that overproduce IAA generated anincreased number of ectomycorrhizal roots (Gay
et al.
, 1994)which presented a strikingly altered morphology: a multiseriateHartig net (Gea
et al.
, 1994). This confirms that somemorphogenetic steps controlling development of mycorrhizaare regulated by fungal IAA. Inhibitors of polar auxin trans-port, such as 2,3,5-triiodobenzoic acid (TIBA), restrict thestimulation of lateral root formation and the colonization ofthe tap-root cortex of conifer seedlings by ectomycorrhizalfungi (Karabaghli-Degron
et al.
, 1998).
Pisolithus
-secretedhypaphorine acts as a natural auxin antagonist (Ditengou& Lapeyrie, 2000). Whether tryptophan and/or other com-ponents of the plant exudates induces an IAA amplificationloop in the rhizospheric mycelium, synthesis of IAA must betightly controlled or compensated by other factors, since abovea certain concentration, exogenously supplied IAA inhibitsroot development.
This brief review has attempted to shed light on recentadvances in how the ectomycorrhizal symbionts respond torhizospheric signals, including hormones, and how thesevarious signals interact in order to execute the appropriatedevelopmental responses (Fig. 2). The multiplicity of identi-fied signals (auxins, alkaloids, cytokinins, phenylglycoside)has confirmed the complex network of cues controlling devel-opment of the symbiosis. These signals can act in a synergistic(rutin/zeatin) or antagonistic (IAA/hypaphorine) manner.Signal perception may culminate in the induction of down-stream target gene products whose expression probablyunderpins physiological and/or development responses (shortroot proliferation, alteration of hyphal branching). Althoughmany pieces of the puzzle remain to be elucidated, it seemsinescapable that the cross-talk between rhizospheric meta-bolites, the hormonal balance and signalling networks involv-ing Ca
2
+
play an important role in coordinating the executionof the appropriate responses and it will be fascinating tolearn more about the multiple facets of this communicationnetwork.
Walking in the promised land: how to escape host surveillance
It is well established that a variety of fungal products can elicitinducible plant defense responses in both host and nonhostplants, and that such responses can also be triggered by plantproducts released during cell-wall degradation (Boller, 1995).It seems a reasonable assumption that nonspecific oligo-saccharidic and proteinaceous elicitors are released duringroot colonization by ectomycorrhial fungi (Salzer
et al.
, 1996;1997c). Signal transduction pathways, related to the per-ception of microbial invasion of host plant tissues, includechanges in protein phosphorylation status, modifications in
NPH169.fm Page 149 Tuesday, May 29, 2001 5:12 PM

Research review
www.newphytologist.com
©
New Phytologist
(2001)
151
: 145–154
Review150
ion fluxes, increase in cytosolic Ca
2
+
concentration, depol-arization of the plasma membrane, production of reactiveoxygen species (ROS), and alterations in gene expression(Yang
et al.
, 1997). Post-translational modification of proteinsby phosphorylation is a general mechanism in the reception/transduction of signals originating from pathogens (Yang
et al.
,1997) and symbionts (Pingret
et al.
, 1998). Thus, it appearsto be of prime importance to identify the protein kinases,protein phosphatases and corresponding protein substratesthat are involved in the early steps of signal transductionin ectomycorrhiza.
Experimental evidence suggests that cell wall components(e.g. glucan and chitin fragments) of mycobionts bind to spe-cific sites on cells from host trees (e.g.
Picea abies
) (Salzer
et al.
,1997c). Treatments of cells of Norway spruce (
Picea abies
)with elicitors released from the ectomycorrhizal fungus
Hebeloma crustuliniforme
induce K
+
and Cl
–
effluxes into themedium followed by a Ca
2
+
influx into the cells within afew minutes (Schwacke & Hager, 1992; Salzer
et al.
, 1996).Phosphorylation of a 63-kDa and dephosphorylation of a65-kDa protein were identified using
in vivo
phospholabelingas little as 4 min after elicitor addition (Salzer
et al.
, 1997a;Hebe
et al.
, 1999). After a few minutes, extracellular alkalin-ization and a transient accumulation of ROS were observed(Schwacke & Hager, 1992; Salzer
et al.
, 1996; Salzer
et al.
,1997b). These initial signalling processes can also be triggeredby the G-protein activator, mastoparan, and the protein phos-phatase inhibitor, cantharidin and okadaic acid (Salzer
et al.
,1997a; Hebe
et al.
, 1999). Similar elicitor-induced reactionsare regarded as initial events of a hypersensitive response (HR)in plant–pathogen interactions (Yang
et al.
, 1997; Heath,2000), but the role of such rapid reactions in ectomycorrhizalinteractions remains to be elucidated.
Specific and nonspecific elicitors seem to trigger signallingcascades involving protein kinases, protein phosphatases, andelements of the mitogen-activated protein (MAP) kinasepathway (Heath, 2000). Some nonspecific, broad-spectrumdefenses are clearly mounted in hosts when the ectomycor-rhizal fungus penetrates directly into the root and digests itsway through the apoplastic space. For example, elicitors fromthe ectomycorrhizal fungi
Amanita muscaria
and
H. crustu-liniforme
induce the enzymes chitinase and peroxidase inspruce (
Picea abies
) cells (Sauter & Hager, 1989; Salzer &Hager, 1993), whereas cell-free extracts of
Pisolithus
elicitinduction of peroxidases and chitinases in
E. globulus
seed-lings (Albrecht
et al.
, 1994). Feugey
et al
. (1999) suggestedthat these induced defense responses may limit Hartig netformation. However, massive root colonization by the myco-biont does not induce the HR, a rapid death of cells at theinfection site that is associated with pathogen limitation aswell as with defense gene activation (Maleck & Dietrich,1999). This suggests that ectomycorrhizal fungi suppressdefense responses through yet unknown mechanisms. Rapidchanges in protein phosphorylation are inactivated by extra-
cellular spruce chitinases and
β
-1,3-glucanases (Salzer
et al.
,1997b) and cleavage of chitinous elicitors from
H. crustulini-forme by these host chitinases prevents induction of K+ andCl– release, extracellular alkalinization and synthesis of H2O2spruce cells (Salzer et al., 1997a). Elicitor-induced changes ofwall-bound and secreted peroxidase activities in suspension-cultured cells of spruce are attenuated by auxins (Mensen et al.,1998), suggesting interactions between elicitor and auxinsignalling cascades. However, it remains to be demonstratedthat these signalling networks occur in intact ectomycorrhiza.
Genes of transduction networks in symbionts
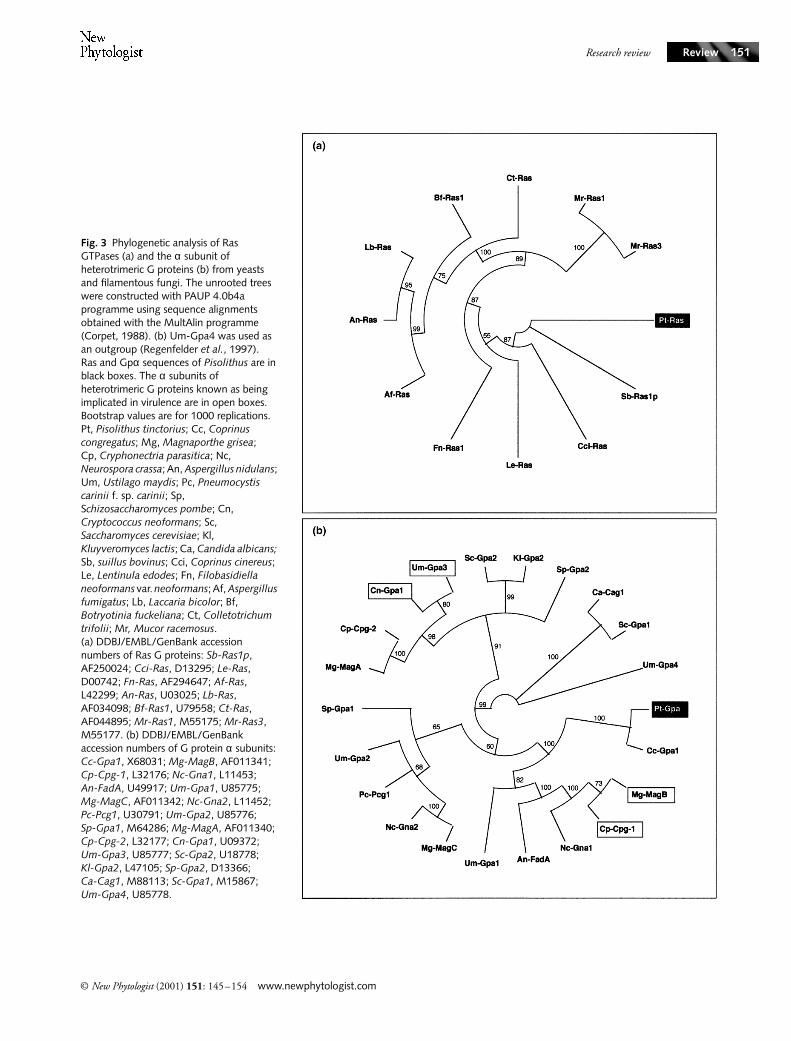
Formation of the symbiotic organ requires temporally andspatially controlled activity of genes and proteins participatingin the morphogenetic process. The proliferation of roots andfungal tissues, and the need to adapt to a rapidly changingenvironment (changes in pH, enhanced fluxes of nutrients,presence of ROS) require a variety of gene products. Sensingthis novel environment and coordination of fungal andplant developments might involved signalling networks.Fungal genes, such as PF6.2 and ras from Laccaria bicolor(Kim et al., 1999; G. Podila, pers. comm.) and ras fromPisolithus (Fig. 3a), are induced before any physical contact(S. Duplessis & H. Lagrange, unpublished), confirming thatdiffusible elicitors are involved in the early steps of theectomycorrhizal interaction.
Communication genes, such as heterotrimeric GTPases,ras, the Ca2+/calmodulin-dependent phosphoprotein phos-phatase (calcineurin), and serine/threonine kinases are expressedin 4-day-old Pisolithus/Eucalyptus ectomycorrhiza (Voibletet al., 2001); they represent about 13% of the cloned genes inthis symbiosis (Table 1). The sequence of the Pisolithus genethat codes for the α subunit of the heterotrimeric GTPase,Pt-Gpα, is very similar to that of the gene encoding thefungal Gpα involved in pathogenesis (Fig. 3b; Alspaugh et al.,1998; Kahmann et al., 1999), suggesting that it probablyplays a similar role in early signalling in the symbioticinteraction. The concentration of transcripts of Gpα wasincreased in 4 d-old E. globulus/Pisolithus ectomycorrhiza(Voiblet et al., 2001; S. Duplessis & F. Martin, unpub-lished). This up-regulation may partly result from changes inthe concentrations of C and N as suggested by the theenhanced expression of this gene in mycelia grown on C-richor N-depleted medium (S. Duplessis & F. Martin, unpub-lished). In addition to these changes in transcript concentra-tions, ectomycorrhizal development induces alteration inthe phosphorylation status of protein (Salzer et al., 1997c).The signal transduction pathways involved in linkingsymbiont contact with morphogenetic changes to the cellularmachinery remain to be elucidated. It appears that bothtranscriptional and post-translational regulations are involvedin the differentiation processes that are essential for symbiosisdevelopment.
NPH169.fm Page 150 Tuesday, May 29, 2001 5:12 PM
Research review
© New Phytologist (2001) 151: 145–154 www.newphytologist.com
Review 151
Fig. 3 Phylogenetic analysis of Ras GTPases (a) and the α subunit of heterotrimeric G proteins (b) from yeasts and filamentous fungi. The unrooted trees were constructed with PAUP 4.0b4a programme using sequence alignments obtained with the MultAlin programme (Corpet, 1988). (b) Um-Gpa4 was used as an outgroup (Regenfelder et al., 1997). Ras and Gpα sequences of Pisolithus are in black boxes. The α subunits of heterotrimeric G proteins known as being implicated in virulence are in open boxes. Bootstrap values are for 1000 replications. Pt, Pisolithus tinctorius; Cc, Coprinus congregatus; Mg, Magnaporthe grisea; Cp, Cryphonectria parasitica; Nc, Neurospora crassa; An, Aspergillus nidulans; Um, Ustilago maydis; Pc, Pneumocystis carinii f. sp. carinii; Sp, Schizosaccharomyces pombe; Cn, Cryptococcus neoformans; Sc, Saccharomyces cerevisiae; Kl, Kluyveromyces lactis; Ca, Candida albicans; Sb, suillus bovinus; Cci, Coprinus cinereus; Le, Lentinula edodes; Fn, Filobasidiella neoformans var. neoformans; Af, Aspergillus fumigatus; Lb, Laccaria bicolor; Bf, Botryotinia fuckeliana; Ct, Colletotrichum trifolii; Mr, Mucor racemosus. (a) DDBJ/EMBL/GenBank accession numbers of Ras G proteins: Sb-Ras1p, AF250024; Cci-Ras, D13295; Le-Ras, D00742; Fn-Ras, AF294647; Af-Ras, L42299; An-Ras, U03025; Lb-Ras, AF034098; Bf-Ras1, U79558; Ct-Ras, AF044895; Mr-Ras1, M55175; Mr-Ras3, M55177. (b) DDBJ/EMBL/GenBank accession numbers of G protein α subunits: Cc-Gpa1, X68031; Mg-MagB, AF011341; Cp-Cpg-1, L32176; Nc-Gna1, L11453; An-FadA, U49917; Um-Gpa1, U85775; Mg-MagC, AF011342; Nc-Gna2, L11452; Pc-Pcg1, U30791; Um-Gpa2, U85776; Sp-Gpa1, M64286; Mg-MagA, AF011340; Cp-Cpg-2, L32177; Cn-Gpa1, U09372; Um-Gpa3, U85777; Sc-Gpa2, U18778; Kl-Gpa2, L47105; Sp-Gpa2, D13366; Ca-Cag1, M88113; Sc-Gpa1, M15867; Um-Gpa4, U85778.
NPH169.fm Page 151 Tuesday, May 29, 2001 5:12 PM

Research review
www.newphytologist.com © New Phytologist (2001) 151: 145–154
Review152
Analysing transcription during early symbiosis development
The interaction between ectomycorrhizal fungi and roots isaccompanied by a series of molecular changes in both themycobiont and the host plant. These events result in alteredexpression of a number of proteins (Hilbert et al., 1991) andgenes (Voiblet et al., 2001). There is now an ever-growingnumber of technologies available for conducting large-scaleexpression studies, including sequencing-based methods, suchas the serial analysis of gene expression, and solid-supportapproaches, such as oligonucleotide and cDNA arrays(Strauss & Falkow, 1997; Bouchez & Höfte, 1998). Over thepast 3 yr, expression profiles have been reported for processesranging from fundamental cellular events such as cell cycle(Spellman et al., 1998) and oncogenesis (Alizadeh et al.,2000) to physiological challenges such a change in a nutrientsource (Wang et al., 2000) or insect-induced wounding ofleaves (Reymond et al., 2000). Transcriptional changes areobserved in as many as 10–20% of all genes during a givenprocess and only a fraction of these differences have beencharacterized previously. The simultaneous measurement ofseveral hundreds of transcripts allows more precise different-iation between related patterns of change, making it likelythat new topographical features of genetic regulation will beidentified. Elucidation of the nature of genes differentiallyexpressed during the development of ectomycorrhiza couldhelp in understanding the molecular basis of the early eventsin plant–ectomycorrhizal fungus interaction.
By using differential screening of about 500 arrayedcDNAs, we found 65 SR genes differentially expressed duringectomycorrhizal mantle formation (Voiblet et al., 2001). Thenumber of SR genes displaying similarity to genes involvedin cell wall and membrane synthesis, stress defence response,
protein degradation (in plant cells) and protein synthesis (inhyphae) suggests a highly dynamic environment in whichsymbionts are sending and receiving signals, are exposed tohigh levels of stress and are remodeling tissues. A strikingresult of these studies is the fact that all genes investigated arecommon to the nonsymbiotic and symbiotic stages. At thedevelopmental stage studied, symbiosis development does notinduce the expression of ectomycorrhiza-specific genes but,rather, a marked change in gene expression in the partners,suggesting that genes for vegetative development may havebeen recruited to function during mycorrhiza formation. Asimilar pattern has been shown for nodulation in alfalfa(Györgyey et al., 2000; Jiménez-Zurdo et al., 2000).
Conclusions
The nature of the signals released by the ectomycorrhizalsymbionts, how these signals are transduced within thepartners, and how these processes trigger the expression ofsymbiosis-regulated genes that assist in partner recognitionand the formation of symbiotic tissues are only beginning tobe understood. We can expect that new components of thetransduction pathways will soon be identified, facilitatingan understanding of the cross-talking between signallingnetworks. About 100 SR-genes have been identified invarious ectomycorrhiza associations, the products of whichmay play a role in recognition and attachment of the myco-biont onto root surfaces, formation of the symbiotic inter-face, signalling networks, protein turnover, organogenesis,and novel symbiotic metabolism. However, many questionsconcerning the differentiation of plant and fungal sym-biotic structures remain unanswered. Master regulatory genes(e.g. homeogenes) that may control morphogenesis in thesymbiosis have not yet been isolated and the biochemical
Clone ID Organism Ratio Best database match (species)
ud283 Fungus 4.0 PWP2/Transducin (Saccharomyces cerevisiae)9B9 Plant 3.4 Calmodulin (Daucus carota)8A7 Plant 2.7 ADP-ribosylation factor (Oryza sativa)8D10 Fungus 2.5 SHP1 protein phosphatase (Schizosaccharomyces pombe)11A6 Fungus 2.1 GTP-binding protein, α subunit (Coprinus congregatus)St92 Fungus 1.9 Putative histidine kinase (Arabidopsis thaliana)1D4 Fungus 1.8 GTP-binding protein YPT1 (Neurospora crassa)St114 Fungus 1.8 Serine/Threonine protein kinase (S. cerevisiae)5E9 Fungus 1.7 Rab11D/ras-related protein (Lotus japonicus)8D4 Fungus 1.5 Lectin receptor-like protein (A. thaliana)6C8 Fungus 1.5 PtCPC2, Gβ/RACK-like protein (Pisolithus tinctorius)EgPtdB24 Fungus 1.3 GTP-binding protein GTB1 (Mus musculus)7B5 Fungus 1.2 Ras1p (Suillus bovinus)St18 Fungus 1.2 SNF1 (carbon catabolite derepressing) (S. cerevisiae)8B8 Plant 1.0 ADP-ribosylation factor (Vigna unguiculata)8A1 Fungus 0.9 Serine/Threonine protein kinase (A. thaliana)11C5 Fungus 0.9 Serine/Threonine protein kinase (Caenorhabditis elegans)11D1 Fungus 0.9 Calcineurine Β subunit (N. crassa)
Table 1 Differential expression of genes of signalling pathways in ectomycorrhiza vs free-living partners. The columns represent: (1) the expressed sequence tag (EST) clone ID (2) the genomic origin (fungus or plant) (3) the ratio for the normalized hybridization values of transcripts expressed in the symbiotic tissues and in the free-living partners, and (4) the best database match (and corresponding species) (after Voiblet et al., 2001)
NPH169.fm Page 152 Tuesday, May 29, 2001 5:12 PM

Research review
© New Phytologist (2001) 151: 145–154 www.newphytologist.com
Review 153
activities associated with several SR genes are still unknown.Elucidation of the underlying mechanisms may lead to waysto intervene in this process. There are two routes for furtherwork. Firstly, ectomycorrhiza-regulated genes need to bestudied in order to determine their function in the develop-ment of the symbiosis by using reverse genetics. Secondly, thetranscriptional regulation of SR-genes needs to be analysed.What transcription factors regulate SR-gene expression, andhow is elicitor/signal-dependent activation of these tran-scription factors achieved? The answers to these questions willprovide further highlights into the signalling networks andearly gene regulation processes involved in ectomycorrhizadevelopment. Eventually, such studies will lead to a betterunderstanding of plant–microbe interactions and evolutionof plant–fungus associations.
Acknowledgements
This paper was presented at the 52nd Harden-New PhytologistConference ‘Signalling in plants’ held October 18–22, 2000,at the Wye College in Wye, Kent, UK. F.M. would like to thankJ. Gallon and C. Smith for their invitation to contribute tothis meeting and the New Phytologist Trust for its financialsupport. S.D., H.L. and C.V. were supported by DoctoralScholarships from the Ministère de l’Education Nationale, dela Recherche et de la Technologie. FAD was supported by afellowship from the Government of Gabon. We also appreci-ated partial support from the Groupement de Recherches etd’Etude des Génomes, the INRA Collaborative Research Pro-grammes in Microbiology, the European Commission INCO-DC Programme (contract number: ERBIC18CT-98319) andCOST-EUROSILVA-WG3–2000.
References
Albrecht C, Asselin A, Piché Y, Lapeyrie F. 1994. Chitinase activities are induced in Eucalyptus globulus roots by ectomycorrhizal or pathogenic fungi, during early colonization. Physiologia Plantarum 91: 104–110.
Alspaugh JA, Perfect JR, Heitman J. 1998. Signal transduction pathways regulating differentiation and pathogenicity of Cryptococcus neoformans. Fungal Genetics and Biology 25: 1–14.
Alizadeh AA, Eisen MB, Davis RE, Ma C, Lossos IS, Rosenwald A, Boldrick JG, Sabet H, Tran T, Yu X, Powell JI, Yang L, Marti GE, Moore T, Hudson J, Lu L, Lewis DB, Tibshirani R, Sherlock G, Chan WC, Greiner TC, Weisenburger DD, Armitage JO, Warnke R, Levy R, Wilson W, Grever MR, Byrd JC, Botstein D, Brown PO, Staudt LM. 2000. Distinct types of diffuse large B-cell lymphoma identified by gene expression profiling. Nature 403: 503–511.
Ashford AE, Allaway WG, Peterson CA, Cairney JWG. 1989. Nutrient transfer and the fungus–root interface. Australian Journal of Plant Physiology 16: 85–97.
Béguiristain T, Lapeyrie F. 1997. Host plant stimulates hypaphorine accumulation in Pisolithus tinctorius hyphae during ectomycorrhizal infection while excreted fungal hypaphorine controls root hair development. New Phytologist 136: 525–532.
Béguiristain T, Côté R, Rubini P, Jay-Allemand C, Lapeyrie F. 1995. Hypaphorine accumulation in hyphae of the ectomycorrhizal fungus Pisolithus tinctorius. Phytochemistry 40: 1089–1091.
Boller T. 1995. Chemoperception of microbial signals in plant cells. Annual Review of Plant Physiology and Plant Molecular Biology 46: 189–214.
Bouchez D, Höfte H. 1998. Functional genomics in plants. Plant Physiology 118: 725–732.
Cairney JWG, Burke RM. 1996. Physiological heterogeneity within fungal mycelia: an important concept for a functional understanding of the ectomycorrhizal symbiosis. New Phytologist 134: 685–695.
Cairney JWG, Ashford AE, Allaway WG. 1989. Distribution of photosynthetically fixed carbon within root systems of Eucalyptus pilularis plants ectomycorrhizal with Pisolithus tinctorius. New Phytologist 112: 495–500.
Carnero Diaz E, Martin F, Tagu D. 1996. Eucalypt α-tubulin: cDNA cloning and increased level of transcripts in ectomycorrhizal root system. Plant Molecular Biology 31: 905–910.
Chilvers GA. 1968. Low-power electron microscopy of the root cap region of Eucalypt mycorrhizas. New Phytologist 67: 663–665.
Corpet F. 1988. Multiple sequence alignment with hierarchical clustering. Nucleic Acid Research 16: 10881–10890.
Dénarié J, Debellé F, Promé JC. 1996. Rhizobium lipo-chitooligosaccharide nodulation factors: signaling molecules mediating recognition and morphogenesis. Annual Review of Biochemistry 65: 503–535.
Ditengou FA, Lapeyrie F. 2000. Hypaphorine from the ectomycorrhizal fungus Pisolithus tinctorius counteracts activities of indole-3-acetic acid and ethylene but not synthetic auxins in eucalypt seedlings. Molecular Plant–Microbe Interactions 13: 151–158.
Ditengou FA, Béguiristain T, Lapeyrie F. 2000. Root hair elongation is inhibited by hypaphorine, the indole alkaloid from the ectomycorrhizal fungus Pisolithus tinctorius, and restored by IAA. Planta. (In press.)
Eckert SE, Kubler E, Hoffmann B, Braus GH. 2000. The tryptophan synthase-encoding trpB gene of Aspergillus nidulans is regulated by the cross-pathway control system. Molecular and General Genetics 263: 867–876.
Feugey L, Strullu DG, Poupard P, Simoneau P. 1999. Induced defence responses limit Hartig net formation in ectomycorhizal birch roots. New Phytologist 144: 541–547.
Fitter AH. 1991. Costs and benefits of mycorrhizas – Implications for functioning under natural conditions. Experientia 47: 350–354.
Fries N, Serck-Hanssen K, Häll Dimberg L, Theander O. 1987. Abietic acid, an activator of basidiospore germination in ectomycorrhizal species of the genus Suillus (Boletaceae). Experimental Mycology 11: 360–363.
Gay G, Normand L, Marmeisse R, Sotta B, Debaud JC. 1994. Auxin overproducer mutants of Hebeloma cylindrosporum Romagnési have increased mycorrhizal activity. New Phytologist 128: 645–657.
Gea L, Normand L, Vian B, Gay G. 1994. Structural aspects of ectomycorrhiza of Pinus pinaster (Ait.) Sol. formed by an IAA-overproducer mutant of Hebeloma cylindrosporum Romagnési. New Phytologist 128: 659–670.
Geitmann A, Emons AMC. 2000. The cytoskeleton in plant and fungal cell tip growth. Journal of Microscopy 198: 218–245.
Gilroy S, Jones DL. 2000. Through form to function: root hair development and nutrient uptake. Trends in Plant Science 5: 56–60.
Gogala N. 1991. Regulation of mycorrhizal infection by hormonal factors produced by hosts and fungi. Experimentia 47: 331–339.
Grove TS, Le Tacon F. 1993. Mycorrhiza in plantation forestry. Advances in Plant Pathology 9: 191–227.
Györgyey J, Vaubert D, Jiménez-Zurdo JI, Charon C, Troussard L, Kondorosi A, Kondorosi E. 2000. Analysis of Medicago truncatula nodule expressed sequence tags. Molecular Plant – Microbe Interactions 13: 62–71.
Hampp R, Schaeffer C, Wallenda T, Stülten C, Johann P, Einig W. 1995. Changes in carbon partitioning or allocation due to ectomycorrhiza formation: biochemical evidence. Canadian Journal of Botany 73: S548–S556.
Heath M. 2000. Nonhost resistance and nonspecific plant defenses. Current Opinion in Plant Biology 3: 315–319.
NPH169.fm Page 153 Tuesday, May 29, 2001 5:12 PM

Research review
www.newphytologist.com © New Phytologist (2001) 151: 145–154
Review154
Hebe G, Hager A, Salzer P. 1999. Initial signalling processes induced by elicitors of ectomycorrhiza-forming fungi in spruce cells can also be triggered by G-protein-activating mastoparan and protein phosphatase-inhibiting cantharidin. Planta 207: 418–425.
Hibbett DS, Gilbert LB, Donoghue MJ. 2000. Evolutionary instability of ectomycorrhizal symbioses in basidiomycetes. Nature 407: 506–510.
Hilbert JL, Costa G, Martin F. 1991. Ectomycorrhizin synthesis and polypeptide changes during the early stage of eucalypt mycorrhiza development. Plant Physiology 97: 977–984.
Horan DP, Chilvers GA. 1990. Chemotropism; the key to ectomycorrhizal formation? New Phytologist 116: 297–301.
Horan DP, Chilvers GA, Lapeyrie FF. 1988. Time sequence of the infection process in eucalypt ectomycorrhizas. New Phytologist 109: 451–458.
Jimenez-Zurdo JI, Frugier F, Crespi MD, Kondorosi A. 2000. Expression profiles of 22 novel molecular markers for organogenetic pathways acting in alfalfa nodule development. Molecular Plant – Microbe Interactions 13: 96–106.
Kahmann R, Basse C, Feldbrügge M. 1999. Fungal-plant signalling in the Ustilago maydis-maize pathosystem. Current Opinion in Microbiology 2: 647–650.
Karabaghli-Degron C, Sotta B, Bonnet M, Gay G, Le Tacon F. 1998. The auxin transport inhibitor 2,3,5-triiodobenzoic acid (TIBA) inhibits the stimulation of in vitro lateral root formation and the colonization of the tap-root cortex of Norway spruce (Picea abies) seedlings by the ectomycorrhizal fungus Laccaria Bicolor. New Phytologist 140: 723–733.
Kaska DD, Myllylä R, Cooper JB. 1999. Auxin transport inhibitors act through ethylene to regulate dichotomous branching of lateral root meristems in pine. New Phytologist 142: 49–58.
Kim SJ, Hiremath ST, Podila GK. 1999. Cloning and identification of symbiosis-regulated genes from the ectomycorrhizal Laccaria bicolor. Mycological Research 103: 168–172.
Kottke I, Oberwinkler F. 1987. The cellular structure of the Hartig net: coenocytic and transfer cell-like organization. Nordic Journal of Botany 7: 85–95.
Lagrange H, Jay-Allemand C, Lapeyrie F. 2001. Rutin, the phenolglycoside from Eucalyptus root exudates, stimulates Pisolithus hyphal growth at picomolar concentrations. New Phytologist 150: 349–355.
LePage BA, Currah RS, Stockey RA, Rothwell GW. 1997. Fossil ectomycorrhizae from the middle eocene. American Journal of Botany 84: 410–412.
Maleck K, Dietrich RA. 1999. Defense on multiple fronts: how do plants cope with diverse enemies? Trends in Plant Science 4: 215–219.
Martin F, Botton B. 1993. Nitrogen metabolism of ectomycorrhizal fungi and ectomycorrhiza. Advances in Plant Pathology 9: 83–102.
Martin F, Lapeyrie F, Tagu D. 1997. Altered gene expression during ectomycorrhizal development. In: Tudzynski C, ed. The mycota, V, Part A, plant relationships. Berlin, Heidelberg, Germany: Springer-Verlag, 223–242.
Martin F, Laurent P, De Carvalho D, Voiblet C, Balestrini R, Bonfante P, Tagu D. 1999. Cell wall proteins of the ectomycorrhizal basidiomycete Pisolithus tinctorius: identification, function, and expression in symbiosis. Fungal Genetics and Biology 27: 161–174.
Mensen R, Hager A, Salzer P. 1998. Elicitor-induced changes of wall-bound and secreted peroxidase activities in suspension-cultured spruce (Picea abies) cells are attenuated by auxins. Physiologia Plantarum 102: 539–546.
Moller SG, Chua NH. 1999. Interactions and intersections of plant signaling pathays. Journal of Molecular Biology 293: 219–234.
Paris F, Dexheimer J, Lapeyrie F. 1993. Cytochemical evidence of a fungal cell wall alteration during infection of Eucalyptus roots by the ectomycorrhizal fungus Cenococcum geophilum. Archives of Microbiology 159: 526–529.
Peterson RL, Farquhar ML. 1996. Root hairs: specialized tubular cells extending root surfaces. Botanical Review 62: 1–40.
Pingret JL, Journet EP, Barker DG. 1998. Rhizobium nod factor signaling: evidence for a G protein-mediated transduction mechanism. Plant Cell 10: 659–671.
Read DJ. 1991. Mycorrhizas in ecosystems. Experientia 47: 376–390.Read DJ, Armstrong W. 1972. A relationship between oxygen transport and
the formation of ectotrophic mycorrhizal sheath in conifer seedlings. New Phytologist 71: 49–53.
Regenfelder E, Spellig T, Hartmann A, Lauenstein S, Bolker M, Kahmann R. 1997. G proteins in Ustilago maydis: transmission of multiple signals? EMBO Journal 16: 1934–1942.
Reymond P, Weber H, Damond M, Farmer EE. 2000. Differential gene expression in response to mechanical wounding and insect feeding in Arabidopsis. Plant Cell 12: 707–719.
Rupp LA, Mudge KW, Negm FB. 1989. Involvement of ethylene in ectomycorrhiza formation and dichotomous branching of roots of mugo pine seedlings. Canadian Journal of Botany 67: 477–482.
Rygiewicz PT, Andersen CP. 1994. Mycorrhizae alter quality and quantity of carbon allocated below ground. Nature 369: 58–60.
Salzer P, Hager A. 1993. Effect of auxins and ectomycorrhizal elicitors on wall bound proteins and enzymes of spruce (Picea abies (L.) Karst.) cells. Trees 8: 49–55.
Salzer P, Hebe G, Reith A, Zitterell-Haid B, Stransky H, Gaschler K, Hager A. 1996. Rapid reactions of spruce cells to elicitors released from the ectomycorrhizal fungus Hebeloma crustuliniforme, and inactivation of these elicitors by extracellular spruce cell enzymes. Planta 198: 118–126.
Salzer P, Hebe G, Hager A. 1997a. Cleavage of chitinous elicitors from the ectomycorrhizal fungus Hebeloma crustuliniforme by host chitinases prevents induction of K+ and Cl− release, extracellular alkalinization and H2O2 synthesis of Picea abies cells. Planta 203: 470–479.
Salzer P, Hübner B, Sirrenberg A, Hager A. 1997b. Differential effect of purified spruce chitinases and β-1,3-glucanases on the activity of elicitors from ectomycorrhizal fungi. Plant Physiology 114: 957–968.
Salzer P, Münzenberger B, Schwacke R, Kottke I, Hager A. 1997c. Signalling in ectomycorrhizal fungus-root interactions. In: Rennenberg H, Eschrich W, Ziegler H, eds. Trees – contributions to modern tree physiology. Leiden, The Netherlands: Backhuys Publishers, 339–356.
Sauter M, Hager A. 1989. The mycorrhizal fungus Amanita muscaria induces chitinase activity in roots and in suspension-cultured cells of its host Picea abies. Planta 179: 61–66.
Schwacke R, Hager A. 1992. Fungal elicitors induce a transient release of active oxygen species from cultured spruce cells that is dependent on Ca2+ and protein-kinase activity. Planta 187: 136–141.
Selosse MA, Le Tacon F. 1998. The land flora: a phototroph-fungus partnership? Trends in Ecology & Evolution 13: 15–20.
Smith SA, Read D. 1997. Mycorrhizal symbiosis, 2nd edn. London, UK: Academic Press.
Spellman PT, Sherlock G, Zhang MQ, Iyer VR, Anders K, Eisen MB, Brown PO, Botstein D, Futcher B. 1998. Comprehensive identification of cell cycle-regulated genes of the yeast Saccharomyces cerevisiae by microarray hybridization. Molecular Biology of the Cell 9: 3273–3297.
Strauss EJ, Falkow S. 1997. Microbial pathogenesis: genomics and beyond. Science 276: 707–712.
Thomson J, Melville LH, Peterson RL. 1989. Interaction between the ectomycorrhizal fungus Pisolithus tinctorius and root hairs of Picea mariana (Pinaceae). American Journal of Botany 76: 632–636.
Voiblet C, Duplessis S, Encelot N, Martin F. 2001. Identification of symbiosis-regulated genes in Eucalyptus globulus-Pisolithus tinctorius ectomycorrhiza by differential hybridization of arrayed cDNAs. Plant Journal 25: 181–191.
Wang R, Guegler K, LaBrie ST, Crawford NM. 2000. Genomic analysis of a nutrient response in Arabidopsis reveals diverse expression patterns and novel metabolic and potential regulatory genes induced by nitrate. Plant Cell 12: 1491–1510.
Yang YO, Shah J, Klessig DF. 1997. Signal perception and transduction in defense responses. Genes and Development 11: 1621–1639.
NPH169.fm Page 154 Tuesday, May 29, 2001 5:12 PM


















