
Development of a model for evaluating the effects of ... · Development of a model for evaluating...
11
© 2007 The Authors Journal compilation © 2007 The Royal Entomological Society Agricultural and Forest Entomology (2007), 9, 189–199 DOI:10.1111/j.1461-9563.2007.00335.x Introduction In the last few years, significant locust and grasshopper prob- lems have been reported in many parts of the world, includ- ing major outbreaks in Central Asia ( Latchininsky and Gapparov, 2000), parts of the U.S.A. (Lorentzen, 2002) and north Africa and the Sahel (Enserink, 2004; FAO, 2005). Biopesticides based on the mitosporic fungus, Metarhizium anisopliae var. acridum (= Metarhizium flavoviride Gams and Rozsypal; Driver et al., 2000), have shown considerable potential for biological control of locust and grasshoppers and have been tested extensively throughout Africa, Australia and parts of Europe and Latin America (Thomas et al., 2000; Lomer et al., 2001; Thomas and Kooyman, 2004). Indeed, 2005–06 saw operational use of M. anisopliae var. acridum Development of a model for evaluating the effects of environmental temperature and thermal behaviour on biological control of locusts and grasshoppers using pathogens Justine I. Klass, Simon Blanford and Matthew B. Thomas NERC Centre for Population Biology, Imperial College at Silwood Park, Ascot, Berks., SL5 7PY, U.K. Abstract 1 Recent years have seen an upsurge in locust and grasshopper populations in many parts of the world. Environmentally sustainable approaches to locust and grass- hopper control may be possible through the use of biopesticides based on ento- mopathogenic fungi. Unfortunately, the performance of these biopesticides is highly variable with environmental temperature and host thermoregulatory behaviour critically determining the pattern and extent of mortality after applica- tions. Here, we present a temperature-dependent model that enables us to predict the field performance of Metarhizium anisopliae var. acridum, the key fungal pathogen used in locust biopesticides. 2 The model was constructed using mortality rate data generated across a range of temperatures in the laboratory and is driven by environmental temperature data linked through host body temperature models. 3 Model predictions were validated against empirical field data obtained for five species, Locustana pardalina, Oedaleus senegalensis, Zonocerus variegatus, Nomadacris septemfasciata and Chortoicetes terminifera. Mortality predictions were accurate to a 2-day error in every 10 days. This level of resolution is satis- factory to guide operational use of the biopesticide. 4 The model was subsequently used for a prospective evaluation of the perform- ance of M. anisopliae var. acridum against two additional pest species, Dociostaurus maroccanus and Calliptamus italicus in Spain. Results suggest that this pathogen would work reasonably well against these species as long as early instars are targeted. 5 The model could provide a useful tool to assist in interpreting effectiveness of con- trol operations, develop improved application strategies to optimize the perform- ance of the biopesticide and identify appropriate target species and environments. Keywords Body temperature model, Calliptamus italicus, Chortoicetes terminifera, Dociostaurus maroccanus, environmental temperature, Locustana pardalina, Metarhizium anisopliae var. acridum, Nomadacris septemfasciata, Oedaleus sen- egalensis, thermoregulation, Zonocerus variegatus. Correspondence:Matthew B. Thomas, CSIRO Entomology, GPO Box 1700, Canberra ACT 2601, Australia. Tel.: +61(02) 62464252; fax: +61(02) 62464362; e-mail: [email protected]
-
Upload
hoangthuan -
Category
Documents
-
view
216 -
download
1
Transcript of Development of a model for evaluating the effects of ... · Development of a model for evaluating...

© 2007 The Authors
Journal compilation © 2007 The Royal Entomological Society
Agricultural and Forest Entomology (2007), 9, 189–199 DOI:10.1111/j.1461-9563.2007.00335.x
Introduction
In the last few years, significant locust and grasshopper prob-lems have been reported in many parts of the world, includ-ing major outbreaks in Central Asia ( Latchininsky and Gapparov, 2000 ), parts of the U.S.A. ( Lorentzen, 2002 ) and
north Africa and the Sahel ( Enserink, 2004; FAO, 2005 ). Biopesticides based on the mitosporic fungus, Metarhizium anisopliae var. acridum (= Metarhizium flavoviride Gams and Rozsypal; Driver et al. , 2000 ), have shown considerable potential for biological control of locust and grasshoppers and have been tested extensively throughout Africa, Australia and parts of Europe and Latin America ( Thomas et al. , 2000; Lomer et al. , 2001; Thomas and Kooyman, 2004 ). Indeed, 2005 – 06 saw operational use of M. anisopliae var. acridum
Development of a model for evaluating the effects of environmental temperature and thermal behaviour on biological control of locusts and grasshoppers using pathogens
Justine I. Klass , Simon Blanford and Matthew B. Thomas NERC Centre for Population Biology, Imperial College at Silwood Park, Ascot, Berks., SL5 7PY, U.K .
Abstract 1 Recent years have seen an upsurge in locust and grasshopper populations in many parts of the world. Environmentally sustainable approaches to locust and grass-hopper control may be possible through the use of biopesticides based on ento-mopathogenic fungi. Unfortunately, the performance of these biopesticides is highly variable with environmental temperature and host thermoregulatory behaviour critically determining the pattern and extent of mortality after applica-tions. Here, we present a temperature-dependent model that enables us to predict the field performance of Metarhizium anisopliae var. acridum , the key fungal pathogen used in locust biopesticides.
2 The model was constructed using mortality rate data generated across a range of temperatures in the laboratory and is driven by environmental temperature data linked through host body temperature models.
3 Model predictions were validated against empirical field data obtained for five species, Locustana pardalina , Oedaleus senegalensis , Zonocerus variegatus , Nomadacris septemfasciata and Chortoicetes terminifera. Mortality predictions were accurate to a 2-day error in every 10 days. This level of resolution is satis-factory to guide operational use of the biopesticide.
4 The model was subsequently used for a prospective evaluation of the perform-ance of M. anisopliae var. acridum against two additional pest species, Dociostaurus maroccanus and Calliptamus italicus in Spain. Results suggest that this pathogen would work reasonably well against these species as long as early instars are targeted.
5 The model could provide a useful tool to assist in interpreting effectiveness of con-trol operations, develop improved application strategies to optimize the perform-ance of the biopesticide and identify appropriate target species and environments.
Keywords Body temperature model , Calliptamus italicus, Chortoicetes terminifera , Dociostaurus maroccanus , environmental temperature , Locustana pardalina , Metarhizium anisopliae var. acridum , Nomadacris septemfasciata , Oedaleus sen-egalensis , thermoregulation , Zonocerus variegatus.
Correspondence: Matthew B. Thomas, CSIRO Entomology, GPO Box 1700, Canberra ACT 2601, Australia. Tel.: +61(02) 62464252; fax: +61(02) 62464362 ; e-mail: [email protected]

190 J. I. Klass, S. Blanford and M. B. Thomas
© 2007 The Authors
Journal compilation © 2007 The Royal Entomological Society, Agricultural and Forest Entomology, 9, 189–199
in Australia (> 60 000 ha treated to control the Australian plague locust in Australia; APLC, 2006 ), plus the first operational-scale trials against the Desert locust in Africa ( FAO, 2005 ). Unfortunately, although these new biopesti-cides hold considerable promise, their use is complicated by a high degree of variability in performance, a feature that characterizes many biopesticides and, to some extent, bio-control agents in general ( Lisansky, 1997 ).
Numerous studies of M. anisopliae var. acridum have dem-onstrated that host thermal biology and diurnal temperature range are key factors determining efficacy ( Inglis et al. , 1997 , 1999; Blanford and Thomas, 1999a, b; Arthurs and Thomas, 2000; Blanford et al. , 2000 ). For example, under ideal tem-perature conditions of 25 – 32 °C, the time to death of locusts after treatment can be within 7 days ( Thomas and Jenkins, 1997; Bateman et al. , 1998; Lomer et al. , 2001 ). However, conditions in the field are not constant and temperatures may fluctuate widely. Moreover, the majority of pest acridids are active behavioural thermoregulators ( Chappell and Whitman, 1990 ). That is, they employ a suite of behaviours to maintain a preferred body temperature, relatively independent of am-bient temperatures during the day. For many species from semi-arid regions, this preferred temperature is approxi-mately 38 – 40 °C ( Chappell and Whitman, 1990; Carruthers et al. , 1992; Lactin and Johnson, 1996a, b, 1998; Blanford and Thomas, 1999a, 2000 ). In addition, several species have also been shown to mount a behavioural fever response to infection, which further enhances survival ( Inglis et al. , 1996; Blanford et al. , 1998; Blanford and Thomas, 1999a, 2000; Elliot et al. , 2002; Ouedraogo et al. , 2003 ). Given that the upper limit for growth of M. anisopliae var. acridum is in the range 35 – 40 °C ( Ouedraogo et al. , 1997; Thomas and Jenkins, 1997 ), such body temperatures severely restrict pathogen development, leading to a substantial delay in fungus-induced mortality ( Inglis et al. , 1997; Blanford and Thomas, 1999a, b; Arthurs and Thomas, 2000; Blanford et al. , 2000; Ouedraogo et al. , 2004 ).
The speed of kill of the biopesticide is therefore critically determined by daily temperature fluctuations and the degree to which periods of pathogen growth at night, when hosts cannot thermoregulate, balance periods of minimal growth due to thermoregulation during the day. If conditions at night remain at approximately 20 – 25 °C, as is typical of the Sahel for example ( Blanford et al. , 1998 ), then pathogen growth may be rapid and mortality may still be relatively quick ( Lomer et al. , 1997b; Langewald et al. , 1999 ). However, if temperatures fall below approximately 15 °C, pathogen growth at night will be slow, exacerbating the delay still fur-ther, such that mortality rate is slowed considerably ( Arthurs and Thomas, 2000) .
The practical significance of this variability is that stand-ard control procedures in which chemical products are sprayed and efficacy is assumed are not appropriate for Metarhizium -based biopesticides. Rather, successful deploy-ment of the biopesticide requires the development of novel use strategies where expected performance parameters are defined in advance and decisions on where and when to spray adjusted accordingly ( Thomas et al. , 2000; Thomas, 2000; Lomer et al. , 2001 ). In the absence of such use strategies, the
conventional ‘spray and pray’ tactics will probably compound the inherent variability and seriously undermine the adoption of the biopesticide technology into new integrated control practices. This argument is supported, in part, by the current situation in South Africa where, although a locust biopesti-cide was registered in 1997, adoption of the technology has been negligible ( Thomas et al. , 2000; Enserink, 2004 ).
We present a model that captures the effects of environ-mental temperature and host thermal biology on the in vivo growth of M. anisopliae var. acridum , and enables prediction of the speed of kill of the pathogen after infection. The aim is to use this model to predict the likely efficacy of biopesti-cide applications against different orthopteran targets in dif-ferent environments, and to use this information to assist in defining effective strategies for implementing biological con-trol of locusts and grasshoppers.
Methods
Model overview
In the present study, it is assumed that a locust/grasshopper population becomes infected by M. anisopliae var. acridum after a standard biopesticide application. The application is assumed to be efficient such that all targets become infected with a lethal dose shortly after spraying. In reality, the effi-cacy of a spray application can be quite variable but the main objective here is to understand variability in time to death af-ter infection, and not to explore factors such as formulation, volume application rate, wind speed, temperature and opera-tor error, which can affect the proportion of the population coming into contact with the biopesticide in the first place.
The primary input into the model is ambient temperature data from the field. These ambient data are then used to drive body temperature models for each key species to give the actual temperature the pathogen is exposed to once inside the infected host. The body temperatures then drive a pathogen development model, which accumulates pathogen growth over a series of hourly steps until a point is reached at which either 50 or 90% mortality of the infected population is ex-pected. This enables the effects of different environmental conditions (i.e. variation in temperature and day length over time and space) on standard measures of efficacy (i.e. LT 50 and LT 90 ) for a range of target species to be explored. The model is validated against historical field trial data and then used in a prospective manner to explore expected pathogen performance against novel targets.
The model is written in Visual Basic, run from Excel 97 (Microsoft Corp., Redmond, Washington, DC). Further de-tails of model assumptions and parameter estimation are given below.
Model assumptions
As indicated above, because a massive inundation of spores after a spray application is considered, certain ecological fac-tors such as disease transmission and age structure of the host population, which might otherwise be important in studying

Predicting success of locust and grasshopper biocontrol 191
© 2007 The Authors
Journal compilation © 2007 The Royal Entomological Society, Agricultural and Forest Entomology, 9, 189–199
more natural disease dynamics, are ignored. Dose is also ignored as previous studies ( Thomas and Jenkins, 1997; Arthurs and Thomas, 2001a ; Blanford, S. and Thomas, M.B., 2001 ; Authors’ unpublished data ) indicate this to be of only minor importance in the overall pattern of mortality compared with temperature, at least with respect to performance in the field. Basically, if conditions are optimum for pathogen growth then even a small dose can kill locusts within 1 – 2 weeks. On the other hand, if conditions allow for effective host thermoregulation during the day and are then too cool for growth at night, host survival will be greatly extended no matter what the dose. Interactions with other biotic factors such as predators, which may further influence the overall extent and pattern of mortality in any given field setting ( Thomas et al. , 1998; Arthurs and Thomas, 2001b; Arthurs et al. , 2003 ), are also excluded.
Finally, a key assumption in the model is that certain as-pects of the biology of the host – pathogen interaction are con-served across species. This is important because the basic data used to parameterize the effects of temperature on mortality rate are derived from laboratory-based studies on the desert locust, Schistocerca gregaria ( Arthurs and Thomas, 2001a ). This assumption is valid as extensive studies on, for example, host thermal behaviour ( Chapman, 1965; Stower and Griffiths, 1966; Uvarov, 1977; Kemp, 1986; Inglis et al. , 1996; Lactin and Johnson, 1996a, b, 1998; Blanford and Thomas, 2000) , host immune response ( Gunnarsson, 1988; Gillespie et al. , 2000; Ouedraogo et al. , 2002 , 2003, 2004), behavioural fever ( Boorstein and Ewald, 1987; Blanford et al. , 1998; Elliot et al. , 2002; Bundey et al. , 2003 ) and rate of mortality ( Inglis et al. , 1997 , 1999; Blanford and Thomas, 1999a, b; Price et al. , 1999 ) show considerable similarities between many lo-cust and grasshopper species, including those studied here.
Thermoregulation and host body temperature
Here, we consider seven economically important acridid spe-cies from different geographical locations: the brown locust, Locustana pardalina (Walker), in South Africa; the Senegalese grasshopper, Oedaleus senegalensis (Krauss), in Niger; the variegated grasshopper, Zonocerus variegatus (Linnaeus), in southern Benin; the red locust, Nomadacris septemfasciata (Serville), in Zambia; the Australian plague locust, Chortoicetes terminifera (Walker), in Australia; and the Moroccan locust, Dociostaurus maroccanus (Thunberg) and Italian grasshopper, Calliptamus italicus (Linnaeus), in Spain. For each of these, thermal behaviour was character-ized by measuring body temperatures in relation to ambient field temperatures. Novel measurements of body tempera-tures were taken for L. pardalina in the Karoo, South Africa during February to April 2000, for D. maroccanus and C. italicus in southern Spain during May to June 2000 and 2001, and for N. septemfasciata during February to March 2001 in the Kafue flats in Zambia. Data for O. senegalensis and Z. variegatus were reanalysed from previous published studies ( Blanford et al. , 1998, 2000 ).
The method of measuring body temperature and environ-mental variables was as previously described by Blanford et al. (1998) . In brief, body temperatures were recorded using
a thermocouple connected to a hand-held, single input, fast response, digital thermometer (Omega Engineering Ltd, U.K.). Individual locusts and grasshoppers were captured in a small sweep net or by hand. A small hole was made in the insect’s thorax using the tip of a 0.22-mm diameter hypoder-mic needle, and a 0.125-mm diameter copper constantan thermocouple was inserted to a depth of 2 mm. Body tem-perature readings were taken within 5 – 8 s of capture when the temperature reading had stabilized. Insects held for more than 8 s were discarded because the body temperature might have been affected by handling. Samples were taken over a number of days throughout different hours of the day, start-ing before sunset and working through until 2 – 3 h after sun-set. Data points were therefore representative of body temperatures over a 24-h period. Data were collected in the same manner for all species.
Concurrent with measuring body temperature, measure-ments of ambient temperatures were recorded for a range of microhabitats (soil surface and in vegetation at heights of 1, 15 and 40 cm) at the field site. These data were collected using 3-mm diameter thermisters at 1-min intervals using two Squirrel data loggers (Grant Instruments Ltd, U.K.) that were downloaded regularly. Data from the different habitats were averaged to represent mean temperatures across the mi-crohabitats, matched to body temperatures and used to derive species-specific body temperature models (see below).
Modelling body temperatures
The relationship between body temperature ( Tb ) and ambient temperature ( Ta ) was found to be nonlinear for all species except Z. variegatus ( Fig. 1). Nonlinear relationships are characteristic of behavioural thermoregulators. Zonocerus variegatus is normothermic and therefore showed a linear relationship [ Tb = m ( Ta ) + c].
A range of nonlinear functions, such as logistic regression ( Kemp, 1986 ), polynomial regression ( Carruthers et al. , 1992; Blanford et al. , 1998; Blanford and Thomas, 2000) and sigmoid functions ( Samietz and Köhler, 1998 ) have been used to characterize the ambient-body temperature relation-ship. Polynomial regressions tend to become highly inaccu-rate when predicting body temperatures outside the range of temperatures used to parameterize the model, resulting in inaccurate predictions at the upper or lower end of the curve ( Campbell and Madden, 1990 ). The logistic function is an improvement over the polynomial regression but, although good for making predictions at high temperatures (i.e. when ambient temperature exceeds body temperature), it becomes increasingly inaccurate at lower temperatures ( Samietz and Köhler, 1998 ). The sigmoid function, as proposed by Samietz and Köhler (1998) , overcomes problems of the logistic func-tion at lower temperatures and was therefore used in this study to model body temperatures during daylight hours. The sigmoid function takes the form:
−= +
+
max
inf
1
s
l
T TaTb Ta
Ta
T
[1]

192 J. I. Klass, S. Blanford and M. B. Thomas
© 2007 The Authors
Journal compilation © 2007 The Royal Entomological Society, Agricultural and Forest Entomology, 9, 189–199
Figure 1 Distribution of body temperatures (Tb) against ambient temperatures (Ta) for (A) Locustana pardalina, (B) Dociostaurus maroccanus, (C) Calliptamus italicus, (D) Oedaleus senegalensis, (E) Nomadacris septemfasciata, (F) Chortoicetes terminifera and (G) Zonocerus variegatus. The dots indicate individual body temperature measurements. The broken line shows a null model where Tb = Ta. Best-fit body regression curves for individual species were derived using the parameters in Tables 1 and 2.
(a)
Ambient Temperature (°C)
Ambient Temperature (°C)
Body
Tem
pera
ture
(°C
) Bo
dy T
empe
ratu
re (°
C)
Body
Tem
pera
ture
(°C
) Bo
dy T
empe
ratu
re (°
C)
20
10
30
40
50
10
20
30
40
50
10
20
30
40
50
10
20
30
40
50
10 20 30 4010
20
30
40
50
50
10 20 30 40 50 10 20 30 40 50
10 20 30 40 50 10 20 30 40 50
10 20 30 40 50 10 20 30 40 50
10
20
30
40
50
10
20
30
40
50
(b)
(c) (d)
(e) (f)
(g)

Predicting success of locust and grasshopper biocontrol 193
© 2007 The Authors
Journal compilation © 2007 The Royal Entomological Society, Agricultural and Forest Entomology, 9, 189–199
where Tb is body temperature; Ta is mean ambient tempera-ture at the time the body temperature was recorded; T max is the asymptotic maximum body temperature; T infl is the ambient temperature at the inflection point of the sigmoid curve and s is the slope parameter. T max , T infl and s are param-eter constants estimated, in this study, through standard non-linear procedures (Levenberg – Marquardt Estimation Method) in SPSS for Windows, version 11 (SPSS Inc., Chicago, Illinois).
Behavioural thermoregulation is not possible during the night. Therefore, for the hours between sunset and sunrise, body temperatures are given by:
T b = T a + 1 [2]
Parameter estimates for body temperature models (linear and nonlinear) for all seven species are presented in Tables 1 and 2.
Rate of pathogen development
To describe the basic effects of temperature on pathogen growth (and hence mortality rate), the nonlinear model pro-posed by Lactin et al. (1995 ) was used. This model success-fully captures temperature-dependent rates for a range of processes in arthropods across the full range of environmen-tal temperatures, and takes the form:
r ( T ) = e � T - e [ � k - ( k - T ) / � ] + � [3]
where r ( T ) is the growth rate (per day) at temperature T , p is the Q 10 value for a critical enzyme-catalysed biochemical re-action, k represents the thermal maximum threshold, � the temperature range over which thermal breakdown becomes an over-riding influence. � allows for estimation of a devel-opmental threshold at low temperatures by allowing the curve to intersect the abscissa at suboptimal temperatures. p , k , � and � are fitted parameters. Curves were fitted by iterative nonlinear regression guided by partial derivatives of the de-pendent variable (Levenberg – Marquardt Estimation Method, SPSS, version 10 for Windows) on mortality rates (1/survival time).
Mortality rates of adult desert locusts, S. gregaria , were determined at five constant temperatures (15, 20, 25, 30, 35 ± 1 °C) after infection with M. anisopliae var. acridum
(isolate IMI 330189) at a dose rate of 1 × 10 4 conidia/insect ( Arthurs and Thomas, 2001a ). No pathogen-induced mortality was assumed at temperature extremes of 40 and 10 °C because data from in vitro studies indicate that path-ogen growth approaches zero at these points ( Ouedraogo et al. , 1997; Thomas and Jenkins, 1997 ). Survival rates ex-pressed as 1/time (in days) to achieve either 50% or 90% mortality were used to parameterize Eq. 3 . Best fit parame-ters are given in Table 3. Pathogen growth rate (approxi-mately 1/survival time) was constrained to 0, such that growth rate does not become negative above or below the maximum or minimum temperature for growth, respec-tively. Nonlinear model effects were minimized by integrat-ing constant-temperature developmental rates over fluctuating temperature regimes at hourly intervals, as pro-posed by Xu (1996) . Thus:
= =∫0( ) ( *) ( ) / 24
ys rT r T r T [4]
where s ( rT ) is the accumulated development over the time interval [0, y ], r ( T *) is the hourly rate of development at a particular temperature. Accumulated fungal development s , is initially defined as zero and is assumed equal to 1 at the completion of the development process. Therefore, the pre-dicted development time ( rT *) is given by:
s ( r T * ) = 1 [5]
Accumulated development over time is approximated by: −
=
= ∑
1
0
( ) ( *)n
i
s rT t r T [6]
where n = y/t , t = hourly time interval and the rate of devel-opment ( rT *) is estimated from the average temperature occurring during hour ( t ) and accumulated over time until s ( rT ) = 1. To capture the effects of thermal behaviour, pre-dicted body temperatures (Eq. 1 ) are substituted for ambient temperature.
Initial investigations using this model found that predic-tions for time to reach 50% or 90% mortality under constant temperature conditions were good, with small deviations be-tween observed and predicted values (using the weighted least squares index (WLSI) method described by Richmond and Bacheler (1989) :
Table 1 Parameter estimates (with asymptotic standard errors) for the sigmoid body temperature-ambient temperature model applied to six acridid species ( Locustana pardalina , Dociostaurus maroccanus , Chortoicetes terminifera , Oedaleus senegalensis , Nomadacris septemfasciata and Calliptamus italicus )
Species n
Model parameters
T max ( ± SE) T infl ( ± SE) s ( ± SE) r 2
Locustana pardalina 1759 41.706 (0.167) 26.751 (0.251) – 4.061 (0.158) 0.77 Dociostaurus maroccanus 1199 39.648 (0.162) 27.043 (0.329) – 3.968 (0.151) 0.88 Chortoicetes terminifera 822 39.735 (0.160) 25.099 (0.325) – 3.699 (0.175) 0.84 Oedaleus senegalensis 746 40.384 (0.251) 31.999 (0.483) – 6.824 (0.692) 0.78 Nomadacris septemfasciata 740 36.669 (0.217) 28.926 (0.412) – 7.335 (0.654) 0.80 Calliptamus italicus 929 38.897 (0.152) 31.155 (0.578) – 2.683 (0.192) 0.81
n , Number of body temperature measurements used in fitting the model. Further details are given in the text.

194 J. I. Klass, S. Blanford and M. B. Thomas
© 2007 The Authors
Journal compilation © 2007 The Royal Entomological Society, Agricultural and Forest Entomology, 9, 189–199
−=∑2( )observed predicted
WLSIobserved
[7]
where lower WLSI values (i.e. WLSI = 0) indicate smaller deviations between observed and predicted mortality rates and higher values represent large deviations (i.e. WLSI > 5 represents an error of 7 days in 10 days or 10 days in 20 days, etc.). For example, deviations were smallest when pre-dicting 50% and 90% mortality at intermediate temperatures of 20 – 30 °C (WLSI < 0.06). However, using the model against locust survival data from fluctuating temperature ex-periments revealed much larger errors. For example, under a fixed-point fluctuation of 35/20 °C, the model predicted a slower mortality rate than observed (WLSI = 1.7). Conversely, the model predicted a faster mortality rate than observed un-der a more extreme 40/20 °C fluctuation (WLSI > 7). Initial model predictions against empirical field data also showed the model to generally predict faster speed of kill than actu-ally observed. Given the efforts to correct for differences between rate effects occurring at constant and fluctuating temperatures ( Xu, 1996 ), these errors are not simply a conse-quence of the nonlinear rate functions ( Ratte, 1985 ). Instead, they are better attributed to biological effects of high tem-peratures and fluctuating conditions on the interaction be-tween host and pathogen, which are not captured in the constant temperature data used to parameterize the model. These are discussed below and two correction factors derived to improve the model.
Incorporating delays in pathogen growth in vivo
Several studies have demonstrated that exposure to high tem-peratures and behavioural fever appear to enhance locust/grasshopper survival beyond a simple effect of temperature reducing pathogen growth to zero ( Blanford and Thomas, 1999b, 2000; Elliot et al. , 2002; Ouedraogo et al. , 2003 ). That is, although normal preferred body temperatures around 38 – 40 °C enhance survival by apparently restricting patho-
gen growth, fever temperatures some 3 – 4 °C higher provide additional survival benefits. The mechanisms for this effect remain unclear. A study examining the effects of tempera-ture fluctuations on growth of M. anisopliae var. acridum in vitro , indicated that exposures to 42 °C of more than 4 h resulted in a disproportionately greater reduction in growth than equivalent exposures to 38 °C (E. Watson, unpub-lished PhD Thesis, University of London, 2002 ). This result suggests a detrimental effect of exposure to the higher temperatures on pathogen proliferation, possibly involving cellular/biochemical repair or synthesis and breakdown of heat shock proteins ( Parsell and Lindquist, 1993 ), before cell growth can resume on return to a permissive temperature. In addition, studies on locust immune responses indicate that circulation and phagocytic action of haemocytes is enhanced by thermoregulation and fever ( Ouedraogo et al. , 2003 ). Such enhanced immune action is likely to slow growth of the path-ogen, either by reducing pathogen levels, or at least creating a delay in growth if the pathogen has, initially, to breach haemocytic encapsulations on return to permissive tempera-tures ( Vey and Gotz, 1986 ).
To account for the added effects of thermoregulation and fever, two temperature-dependent delays were incorporated into the 50% and 90% mortality models. Delay 1 for the 50% mortality level states that when locust body temperatures exceed the upper limit for pathogen growth (such that patho-gen growth = 0) for ³ 3 h, an additional 3 h of no growth is assumed. For the 90% mortality model, this period of no growth is increased to 4 h. For Delay 2, it is assumed that if body temperatures fall below 20 °C within 3 h of the end of the first delay, pathogen development remains at zero until temperatures begin to rise and are at or above 20 °C. This delay applies to both the 50% and 90% mortality models. The magnitude of these delays was determined by adjusting the respective models to match observed field mortality data from a trial against brown locust, L. pardalina ( Arthurs and Thomas, 2000) . As such, the delays are phenomenological, but do reflect some of the likely mechanisms referred to above.
Model simulations
Model simulations were run using temperature data collected simultaneously with field operations against the target spe-cies in their respective habitats (i.e. the Karoo, South Africa 1998, for L. pardalina ; southern Niger 1996 and 1997, for O. senegalensis ; southern Benin 1998 for Z. variegatus ; La Serena, Spain 2000 and 2001, for D. maroccanus and C. italicus ; Kafue Flats, Zambia 2001, for N. septemfasciata ;
Table 2 Linear body temperature model parameter estimates for Zonocerus variegatus
Species n
Model parameters
c m r 2
Zonocerus variegatus 1693 5.3091 0.9194 0.70
Parameter estimates are based on a single input variable, mean ambient field temperature.
Table 3 Parameter estimates for temperature-dependent pathogen growth rate in vivo derived from the time taken to reach 50% and 90% mortality across different temperatures at a dose rate of 1 × 10 4 conidia/insect
Mortality level
In vivo pathogen growth parameters ( ± SEM)
p k � � r 2
50% 0.00905 ± 0.0016 42.39639 ± 1.3705 4.30945 ± 1.0371 – 1.08331 ± 0.0280 0.99 90% 0.00938 ± 0.0024 43.13143 ± 1.9539 4.8372 ± 1.5783 – 1.09599 ± 0.0391 0.99

Predicting success of locust and grasshopper biocontrol 195
© 2007 The Authors
Journal compilation © 2007 The Royal Entomological Society, Agricultural and Forest Entomology, 9, 189–199
and Hay, Australia 2000, for C. terminifera ). Ambient tem-peratures were recorded for a range of habitats (soil surface and in vegetation at various heights of 1, 15 and 40 cm) at the field sites. These were taken using 3-mm diameter ther-misters at 1-min intervals using two Squirrel data loggers (Grant Instruments Ltd, U.K.). These data were then aver-aged to give mean hourly temperatures at the field sites, and used to drive the model.
Model predictions were compared with actual mortality data recorded after biopesticide applications of M. anisopliae var. acridum against L. pardalina ( Arthurs and Thomas, 2000) , O. senegalensis ( Langewald et al. , 1999 ), Z. variegatus ( Blanford et al. , 2000 ), N. septemfasciata (Elliot, S.L. and Thomas, M.B., unpublished data , DFID unpublished report) and C. terminifera (D. Hunter, personal communication, Australian Plague Locust Commission, Australia). The accu-racy of the model was assessed using the weighted least squares index (WLSI) method (see Eq. 7 ). Full data on popu-lation mortality after biopesticide applications are not yet available for D. maroccanus and C. italicus , although pre-liminary data confirming susceptibility are available ( Lomer et al. , 1999 ), together with appropriate environmental data and body temperature models. Therefore, for these species, the model was used for prospective analyses to explore the likely effectiveness of the biopesticide.
Results
The brown locust, Locustana pardalina, in the Karoo 1998
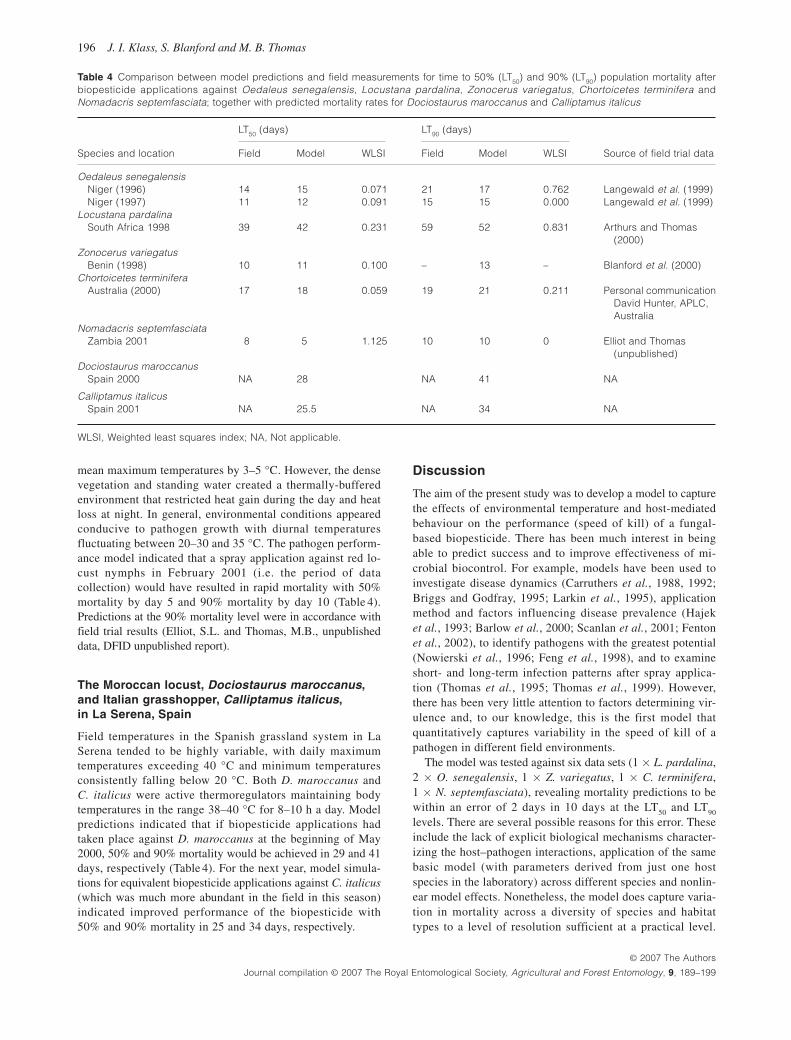
Locustana pardalina inhabits semiarid areas of the Karoo where temperatures can be extreme (high maximum daytime temperatures > 40 °C and relatively cool nights < 15 – 20 °C). During the period of February to March 1998, body tempera-ture measurements indicated locusts were able to maintain body temperatures in the range 38 – 40 °C for 10 h during the day. During the night, body temperatures fell rapidly to below 20 °C. Under these conditions, the model predicted a spray application of the Metarhizium -based biopesticide to act slowly, causing 50% mortality in 42 days and 90% mortality by day 52 ( Table 4). These predictions compare very well with field mortality data after a spray application ( Arthurs and Thomas, 2000) where 50% mortality was observed on day 39 (WLSI = 0.231) and 90% mortality on day 59 (WLSI = 0.831), although it should be recognized that these field data were used to derive the delays in the model and so this represents only a partial validation of the model.
The Senegalese grasshopper, Oedaleus senegalensis, in southern Niger (1996) and 1997
Oedaleus senegalensis is the key grasshopper pest in the Sahelian region during the wet season. Measurements of ambient temperature over July to August in 1996 and 1997 revealed daily maximum temperatures frequently exceeding 35 °C. Body temperature measurements showed O. senegalensis able to maintain body temperatures around 38 °C for 8 h per
day over these periods. Unlike the Karoo, however, night-time ambient temperatures, and hence body temperatures, were generally above 20 °C. Under these conditions, the model predicted much better pathogen performance with 50% and 90% mortality of 15 and 17 days, respectively, for August 1996 and 11 and 15 days, respectively, for August 1997 ( Table 4 ). Comparison with field trial data for the same periods and study sites ( Langewald et al. , 1999 ) showed these model predictions to be generally accurate with a maxi-mum error of 4 days ( Table 4 ).
The variegated grasshopper, Zonocerus variegates, in southern Benin (1998)
Zonocerus variegatus is a grasshopper pest of the humid tropical zone of west and central Africa. Measurements of ambient temperature during March to April in 1998 found daily temperatures oscillating between 22 °C to 36 °C. Body temperatures for Z. variegatus followed a linear distribution, in the range 24 – 46 °C with a mode of 36 °C. Night-time tem-peratures in southern Benin were similar to those recorded in Niger above. Under these conditions, the model predicted that 50% and 90% mortality would be achieved in 11 and 13 days, respectively ( Table 4 ). Comparison with field trial data for the same study period and site ( Blanford et al. , 2000 ) showed model predictions to be accurate, at least at the 50% level, which are the only data that are available ( Table 4 ). However, this rate of mortality is also consistent with other field trial results in this location (Douro-Kpindo et al., 1995; Lomer et al. , 1997a , b).
The Australian plague locust, Chortoicetes terminifera, in Australia (2000)
Chortoicetes terminifera is a locust pest of Eastern and South-western Australia where ambient temperatures during October to November in 2000 were found to oscillate be-tween 9 °C and 41 °C, similar to conditions found in the Karoo, South Africa. During this period, body temperature measurements indicated that C. terminifera were active ther-moregulators. Body temperatures were maintained in the range 38 – 40 °C for 8 h during the day. Under these condi-tions, the model predicted that M. anisopliae var. acridum would cause 50% mortality in 18 days and 90% mortality in 21 days at Hay, Australia ( Table 4 ). Predictions compared fa-vourably with field mortality data after a spray application for the same study period and site (D. Hunter, personal com-munication, Australian Plague Locust Commission) where 50% mortality was observed on day 17 (WLSI = 0.059) and 90% mortality on day 19 (WLSI = 0.211).
The red locust, Nomadacris septemfasciata, in Zambia 2001
Nomadacris septemfasciata nymphs inhabit seasonally flooded dense grassland habitats. Measurements of body temperatures indicated they were active thermoregulators and were able to maintain their body temperatures above the
196 J. I. Klass, S. Blanford and M. B. Thomas
© 2007 The Authors
Journal compilation © 2007 The Royal Entomological Society, Agricultural and Forest Entomology, 9, 189–199
mean maximum temperatures by 3 – 5 °C. However, the dense vegetation and standing water created a thermally-buffered environment that restricted heat gain during the day and heat loss at night. In general, environmental conditions appeared conducive to pathogen growth with diurnal temperatures fluctuating between 20 – 30 and 35 °C. The pathogen perform-ance model indicated that a spray application against red lo-cust nymphs in February 2001 (i.e. the period of data collection) would have resulted in rapid mortality with 50% mortality by day 5 and 90% mortality by day 10 ( Table 4 ). Predictions at the 90% mortality level were in accordance with field trial results (Elliot, S.L. and Thomas, M.B., unpublished data , DFID unpublished report).
The Moroccan locust, Dociostaurus maroccanus, and Italian grasshopper, Calliptamus italicus, in La Serena, Spain
Field temperatures in the Spanish grassland system in La Serena tended to be highly variable, with daily maximum temperatures exceeding 40 °C and minimum temperatures consistently falling below 20 °C. Both D. maroccanus and C. italicus were active thermoregulators maintaining body temperatures in the range 38 – 40 °C for 8 – 10 h a day. Model predictions indicated that if biopesticide applications had taken place against D. maroccanus at the beginning of May 2000, 50% and 90% mortality would be achieved in 29 and 41 days, respectively ( Table 4 ). For the next year, model simula-tions for equivalent biopesticide applications against C. italicus (which was much more abundant in the field in this season) indicated improved performance of the biopesticide with 50% and 90% mortality in 25 and 34 days, respectively.
Discussion
The aim of the present study was to develop a model to capture the effects of environmental temperature and host-mediated behaviour on the performance (speed of kill) of a fungal-based biopesticide. There has been much interest in being able to predict success and to improve effectiveness of mi-crobial biocontrol. For example, models have been used to investigate disease dynamics ( Carruthers et al. , 1988 , 1992; Briggs and Godfray, 1995; Larkin et al. , 1995 ), application method and factors influencing disease prevalence ( Hajek et al. , 1993; Barlow et al. , 2000; Scanlan et al. , 2001; Fenton et al. , 2002 ), to identify pathogens with the greatest potential ( Nowierski et al. , 1996; Feng et al. , 1998 ), and to examine short- and long-term infection patterns after spray applica-tion ( Thomas et al. , 1995; Thomas et al. , 1999 ). However, there has been very little attention to factors determining vir-ulence and, to our knowledge, this is the first model that quantitatively captures variability in the speed of kill of a pathogen in different field environments.
The model was tested against six data sets (1 × L. pardalina , 2 × O. senegalensis , 1 × Z. variegatus , 1 × C. terminifera , 1 × N. septemfasciata ), revealing mortality predictions to be within an error of 2 days in 10 days at the LT 50 and LT 90 levels. There are several possible reasons for this error. These include the lack of explicit biological mechanisms character-izing the host – pathogen interactions, application of the same basic model (with parameters derived from just one host species in the laboratory) across different species and nonlin-ear model effects. Nonetheless, the model does capture varia-tion in mortality across a diversity of species and habitat types to a level of resolution sufficient at a practical level.
Table 4 Comparison between model predictions and field measurements for time to 50% (LT 50 ) and 90% (LT 90 ) population mortality after biopesticide applications against Oedaleus senegalensis , Locustana pardalina , Zonocerus variegatus , Chortoicetes terminifera and Nomadacris septemfasciata ; together with predicted mortality rates for Dociostaurus maroccanus and Calliptamus italicus
Species and location
LT 50 (days) LT 90 (days)
Field Model WLSI Field Model WLSI Source of field trial data
Oedaleus senegalensis Niger (1996) 14 15 0.071 21 17 0.762 Langewald et al. (1999) Niger (1997 ) 11 12 0.091 15 15 0.000 Langewald et al. (1999) Locustana pardalina South Africa 1998 39 42 0.231 59 52 0.831 Arthurs and Thomas
(2000) Zonocerus variegatus Benin (1998) 10 11 0.100 – 13 – Blanford et al. (2000) Chortoicetes terminifera Australia (2000) 17 18 0.059 19 21 0.211 Personal communication
David Hunter, APLC, Australia
Nomadacris septemfasciata Zambia 2001 8 5 1.125 10 10 0 Elliot and Thomas
(unpublished) Dociostaurus maroccanus Spain 2000 NA 28 NA 41 NA
Calliptamus italicus Spain 2001 NA 25.5 NA 34 NA
WLSI, Weighted least squares index; NA, Not applicable.

Predicting success of locust and grasshopper biocontrol 197
© 2007 The Authors
Journal compilation © 2007 The Royal Entomological Society, Agricultural and Forest Entomology, 9, 189–199
That is, in terms of operational locust biocontrol and decid-ing whether it is appropriate to use the biopesticide or not, it is only really necessary to be able to determine whether the biopesticide will work quickly within approximately 10 days, more slowly in around 20 days or much worse at > 30 days. Such predictions would allow locust control officers to decide whether to use the biopesticide and to optimize timing of spraying with respect to locust age. For example, the general aim of preventive locust control is to prevent reproduction and long-range adult dispersal. To achieve this against a popula tion of fourth-instar nymphs would require high levels of mortality within approximately 20 days. Against a popula-tion of first- and second-instar nymphs, on the other hand, then a speed of kill of 30 – 40 days may be sufficient. In this context, the modelling analysis in the current study indicates that species such as N. septemfasciata , Z. variegatus , and possibly O. senegalensis and C. terminifera , might represent good targets that can be treated across the whole of the immature period, whereas species such as L. pardalina would need to be located and sprayed much earlier and might not be suitable for biocontrol at all.
Although the generality of such predictions requires fur-ther analysis, at the very least, the model could provide a use-ful support tool to help monitor and interpret ongoing field trials and control operations. Extending this further, using the model for prospective analysis could be very valuable given the expense of field trials, difficultly in assessing mortality against highly mobile species (especially in small-scale ap-plication typical of preliminary trials), and problems of intro-ducing exotic agents just for testing. That is, having established basic susceptibility, the model enables predictions of likely performance based on relatively simple measures of local climatic conditions and body temperature. These meas-ures can be obtained prior to any introduction or field testing, providing an initial first assessment of whether biocontrol is worth pursuing. For example, our prospective analysis suggests there is scope for control of D. maroccanus and C. italicus , which, as well as being important in the Mediterranean region ( Arias et al. , 1993; Quesada-Moraga, 1998 ), have been major problems in Central Asia in recent years ( Latchininsky and Gapparov, 2000 ). However, speed of kill appears relatively slow. This does not mean that the locusts would not ulti-mately die from disease, or that there would not be important pre- or sublethal effects on reproduction and dispersal ( Arthurs and Thomas, 2000; Elliot et al. , 2005 ) that could contribute to effective control. Nonetheless, it suggests that successful preventive control is likely to depend on targeting early instar nymphs. This not only informs likely use strat-egy, but also emphasizes the need for effective monitoring and surveillance if the biopesticide is to have a role (i.e. if you wait until you see the highly apparent and mobile fourth and fifth instars before acting then it will likely be too late).
Overall, the model presented here is a useful tool that can be used to explore and evaluate the performance of a fungal pathogen currently being used in several locust and grasshop-per biocontrol programmes throughout the world. It has a role in interpreting patterns of mortality after application, provid-ing locust control officers with real-time guidance as to when to expect substantial levels of mortality, and for prospective
analysis to identify candidate targets for biocontrol and opti-mize use strategies. We are currently working to increase the utility of this model by linking with meteorological station data in a Geographic Information Systems framework to ena-ble area-wide applications ( Klass et al. , 2007 ).
Acknowledgements
This work was supported by the European Commission through the Quality of Life and Management of Living Resources Programme implemented under the Fifth Framework Programme (contract number QLRT-1999-01118) Protecting Biodiversity through the Development of Environmentally Sustainable Locust and Grasshopper Control in Europe (ESLOCO).
References
Arias , A. , Alvez , C. , Garcia , F. , Martinez de Velasco , D. , Olivera , J. , Prieto , A. & Santos , R . ( 1993 ) La lucha contra la langosta marro-qui ( Dociostaurus maroccanus Thunb.) en Extremadura durante el decenio 1983 – 92 . Boletin de Sanidad Vegetal , 19 , 425 – 453 .
Arthurs , S.P. & Thomas , M.B . ( 2000 ) Effects of a mycoinsecticide on feeding and fecundity of the brown locust Locustana parda-lina . Biocontrol Science and Technology , 10 , 321 – 329 .
Arthurs , S.P. & Thomas , M.B . ( 2001a ) Effects of dose, pre-mortem host incubation temperature and thermal behaviour on host mor-tality, mycosis and sporulation of Metarhizium anisopliae var. acridum in Schistocerca gregaria . Biocontrol Science and Technology , 11 , 411 – 420 .
Arthurs , S.P. & Thomas , M.B . ( 2001b ) Investigation into behavioural changes in Schistocerca gregaria following infection with a mycoinsecticide: implications for susceptibility to predation . Ecological Entomology , 26 , 227 – 234 .
Arthurs , S.P. , Thomas , M.B. & Langewald , J . ( 2003 ) Field observa-tions of the effects of fenitrothion and Metarhizium anisopliae var. acridum on non-target ground dwelling arthropods in the Sahel . Biological Control , 26 , 333 – 340 .
Australian Plague Locust Commission (APLC) ( 2006 ) Report on Current Control Operations . Available at http://www.affa.gov.au/aplc .
Barlow , N.D. , Caldwell , N.P. , Kean , J.M. & Barron , M.C . ( 2000 ) Modelling the use of NPV for the biological control of Asian gypsy moth Lymantria dispar invading New Zealand . Agricultural and Forest Entomology , 2 , 173 – 184 .
Bateman , R.P. , Douro-Kpindu , O.K. , Kooyman , C. , Lomer , C. & Oambama , Z . ( 1998 ) Some observations on the dose transfer of mycoinsecticide sprays to desert locusts . Crop Protection , 17 , 151 – 158 .
Blanford , S. & Thomas , M.B . ( 1999a ) Role of thermal biology in disease dynamics . Aspects of Applied Biology , 53 , 73 – 82 .
Blanford , S. & Thomas , M.B . ( 1999b ) Host thermal biology: The key to understanding host – pathogen interactions and microbial pest control? Agricultural and Forest Entomology , 1 , 195 – 202 .
Blanford , S. & Thomas , M.B . ( 2000 ) Thermal behaviour of two acri-did species: effects of habitat and season on body temperature and the potential impact on biocontrol with pathogens . Environmental Entomology , 29 , 1060 – 1069 .
Blanford , S. & Thomas , M.B . ( 2001 ) Adult survival, maturation and reproduction of the Desert locust Schistocerca gregaria infected with the fungus Metarhizium anisopliae var. acridum . Journal of Invertebrate Pathology , 78 , 1 – 8 .

198 J. I. Klass, S. Blanford and M. B. Thomas
© 2007 The Authors
Journal compilation © 2007 The Royal Entomological Society, Agricultural and Forest Entomology, 9, 189–199
Blanford , S. , Thomas , M.B. & Langewald , J . ( 1998 ) Behavioural fever in the Senegalese grasshopper, Oedaleus senegalensis , and its implications for biological control using pathogens . Ecological Entomology , 23 , 9 – 14 .
Blanford , S. , Thomas , M.B. & Langewald , J . ( 2000 ) Thermal ecology of Zonocerus variegatus and fungal disease dynamics . Agricul-tural and Forest Entomology , 2 , 3 – 10 .
Boorstein , S.M. & Ewald , P.W . ( 1987 ) Costs and benefits of behav-ioral fever in Melanoplus sanguinipes infected by Nosema acri-dophagus . Physiological Zoology , 60 , 586 – 595 .
Briggs , C.J. & Godfray , H.C.J . ( 1995 ) The dynamics of insect – patho-gen interactions in stage-structured populations . American Natu-ralist , 145 , 855 – 887 .
Bundey , S. , Raymond , S. , Dean , P. , Roberts , S.K. , Dillon , R.J. & Charnley , A.K . ( 2003 ) Eicosanoid involvement in the regulation of behavioral fever in the desert locust, Schistocerca gregaria . Archives of Insect Biochemistry and Physiology , 52 , 183 – 192 .
Campbell , C.L. & Madden , L.V . ( 1990 ) Introduction to Plant Disease Epidemiology . Wiley & Sons, Inc. , U.K.
Carruthers , R.I. , Larkin , T.S. , Firstencel , H. & Feng , Z . ( 1992 ) Influ-ence of thermal ecology on the mycosis of a rangeland grasshopper . Ecology , 73 , 190 – 204 .
Carruthers , R.I. , Larkin , T.S. & Soper , R.S . ( 1988 ) Simulation of insect disease dynamics – an application of SERB to a rangeland eco-system . Simulation , 51 , 101 – 109 .
Chapman , R.F . ( 1965 ) The behaviour of nymphs of Schistocerca gregaria (Forskål) (Orthoptera: Acrididae) in a temperature gradi-ent, with special reference to temperature preference . Behaviour , 24 , 283 – 317 .
Chappell , M.A. & Whitman , D.A . ( 1990 ) Grasshopper thermoregu-lation. Biology of Grasshoppers ( ed . by R. F. Chapman and A. Joern ), pp . 143 – 172 . Wiley Interscience , New York, New York .
Douro-Kpindou , O.K. , Godonou , I. , Houssou , A. , Lomer , C.J. & Shah , P.A . ( 1995 ) Control of Zonocerus variegatus by ultra-low volume application of an oil formulation of Metarhizium flavoviride conidia . Biocontrol Science and Technology , 5 , 131 – 139 .
Driver , F. , Milner , R.J. & Trueman , W.H . ( 2000 ) A taxonomic revi-sion of Metarhizium based on a phylogenetic analysis of rDNA sequence data . Mycological Research , 104 , 134 – 150 .
Elliot , S.L. , Blanford , S. , Horton , C. & Thomas , M.B . ( 2005 ) Im-pacts of fever on locust life history traits: cost or benefit? Biology Letters , 1 , 181 – 184 .
Elliot , S.L. , Blanford , S. & Thomas , M.B . ( 2002 ) Host – pathogen interactions in a varying environment: temperature, behavioural fever and fitness . Proceedings of the Royal Society of London Series B , 269 , 1599 – 1607 .
Enserink , M . ( 2004 ) Can the war on locusts be won? Science , 306 , 1880 – 1882 .
FAO ( 2005 ) Press Release , 28 June . Available at http://www.fao.org/newsroom/en/news/2005/103849/index.html .
Feng , M.-G. , Liu , C.-L. , Xu , J.-H. & Xu , Q . ( 1998 ) Modeling and biological implication of time – dose – mortality data for the ento-mophthoralean fungus, Zoophthora anhuiensis , on the green peach aphid Myzus persicae . Journal of Invertebrate Pathology , 72 , 246 – 251 .
Fenton , A. , Gwynn , R.L. , Gupta , A. , Norman , R. , Fairbairn , J.P. & Hudson , P.J . ( 2002 ) Optimal application strategies for entomo-pathogenic nematodes: integrating theoretical and empirical approaches . Journal of Applied Ecology , 39 , 481 – 492 .
Gillespie , J.P. , Burnett , C.A. & Charnley , K . ( 2000 ) The immune response of the desert locust Schistocerca gregaria during mycosis of the entomopathogenic fungus, Metarhizium anisopliae var acridum . Journal of Insect Physiology , 46 , 429 – 437 .
Gunnarsson , S.G.S . ( 1988 ) Infection of Schistocerca gregaria by the fungus Metarhizium anisopliae : Cellular reactions in the integu-
ment studied by scanning electron and light microscopy . Journal of Invertebrate Pathology , 52 , 9 – 17 .
Hajek , A.E. , Larkin , T.S. , Carruthers , R.I. & Soper , R.S . ( 1993 ) Model-ing the dynamics of Enotmophaga maimaiga (Zygomycetes: Ento-mophthorales) epizootics in gypsy moth (Lepidoptera: Lymantriidae) populations . Environmental Entomology , 22 , 1172 – 1187 .
Inglis , G.D. , Duke , G.M. , Kawchuk , L.M. & Goettel , M.S . ( 1999 ) Influence of oscillating temperatures on the competitive infection and colonization of the migratory grasshopper by Beauveria bassiana and Metarhizium flavoviride . Biological Control , 14 , 111 – 120 .
Inglis , G.D. , Johnson , D.L. , Cheng , K.-L. & Goette , M.S . ( 1997 ) Use of pathogen combinations to overcome the constraints of temperature on entomophagous hyphomycetes against grasshop-pers . Biological Control , 8 , 143 – 152 .
Inglis , D.G. , Johnson , D.L. & Goettel , M.S . ( 1996 ) Effects of tem-perature and thermoregulation on mycosis by Beauveria bassiana in grasshoppers . Biological Control , 7 , 131 – 139 .
Kemp , W.P . ( 1986 ) Thermoregulation in three rangeland grasshop-pers . Canadian Entomologist , 118 , 335 – 343 .
Klass , J.I. , Blanford , S. & Thomas , M.B . ( 2007 ) Use of a geographic information system to explore spatial variation in pathogen viru-lence and the implications for biological control of locusts and grasshoppers . Agricultural and Forest Entomology , in press .
Lactin , D.J. , Holliday , N.J. , Johnson , D.L. & Craigen , R . ( 1995 ) Improved rate model of temperature - dependent development by arthropods . Environmental Entomology , 24 , 68 – 75 .
Lactin , D.J. & Johnson , D.L . ( 1996a ) Convective heat loss and change in body temperature of grasshopper and locust nymphs: relative importance of wind speed, insect size and insect orienta-tion . Journal of Thermal Biology , 23 , 5 – 13 .
Lactin , D.J. & Johnson , D.L . ( 1996b ) Behavioural optimization of body temperature by nymphal grasshoppers ( Melanoplus sanguinipes , Orthoptera: Acrididae) in temperature gradients established using incandescent bulbs . Journal of Thermal Biology , 21 , 231 – 238 .
Lactin , D.J. & Johnson , D.L . ( 1998 ) Environmental, physical, and behavioural determinants of body temperature in grasshopper nymphs (Orthoptera: Acrididae) . Canadian Entomologist , 130 , 551 – 577 .
Langewald , J. , Ouambama , Z. , Mamadou , A . et al . ( 1999 ) Compari-son of an organophosphate insecticide with a mycoinsecticide for the control of Oedaleus senegalensis (Orthoptera: Acrididae) and other Sahelian grasshoppers at an operational scale . Biological Science and Technology , 9 , 199 – 214 .
Larkin , T.S. , Sweeney , A.W. & Carruthers , R.I . ( 1995 ) Simulation of the dynamics of a microsporidian pathogen of mosquitoes . Eco-logical Modelling , 77 , 143 – 165 .
Latchininsky , A.V. & Gapparov , F.A . ( 2000 ) Locust management in Russia, Uzbekistan, Kazakhstan and other CIS Countries, Lessons of Decentralization . Abstract presented at the XXI International Congress of Entomology , 20 – 26 August 2000 . Embrapa , Brazil .
Lisansky , S . ( 1997 ) Microbial insecticides: novelty or necessity? Microbial Biopesticides. BPBC Symposium. Proceedings No. 68 , pp . 3 – 10 . British Crop Protection Council , Farnham, U.K .
Lomer , C.J. , Bateman , R.P. , Dent , D . et al . ( 1999 ) Development of strategies for the incorporation of biological pesticides into the integrated management of locusts and grasshoppers . Agricultural and Forest Entomology , 1 , 71 – 88 .
Lomer , C.J. , Bateman , R.P. , Johnson , D.L. , Langewald , J. & Thomas , M.B . ( 2001 ) Biological control of locusts and grasshoppers . Annual Review of Entomology , 46 , 667 – 702 .
Lomer , C.J. , Prior , C. & Kooyman , C . ( 1997a ) Development of Metarhizium spp. for the control of grasshoppers and locusts . Memoirs of the Entomological Society of Canada , 171 , 265 – 286 .

Predicting success of locust and grasshopper biocontrol 199
© 2007 The Authors
Journal compilation © 2007 The Royal Entomological Society, Agricultural and Forest Entomology, 9, 189–199
Lomer , C.J. , Thomas , M.B. , Douro-Kpindou , O.K. , Gbongboui , C. , Godonou , I. , Langewald , J. & Shah , P.A . ( 1997b ) Control of grasshoppers, particularly Hieroglyphus daganensis , in northern Benin using Metarhizium flavoviride . Memoirs of the Entomologi-cal Society of Canada , 171 , 301 – 311 .
Lorentzen , A . ( 2002 ) Grasshopper infestation plagues West . Washington Post . Available at http://www.washingtonpost.com/wp-dyn/articles/A19438-2002Jul17.html .
Nowierski , R.M. , Zheng , Z. , Jaronski , S. , Delgado , F. & Swearingen , W . ( 1996 ) Analysis and modelling of time – dose – mortality of Melanoplus sanguinipes , Locusta migratoria migratorioides , and Schistocerca gregaria (Orthoptera: Acrididae) from Beauveria , Metarhizium , and Paecilomyces isolates from Madagascar . Jour-nal of Invertebrate Pathology , 67 , 236 – 252 .
Ouedraogo , R.M. , Cusson , M. , Goettel , M.S. & Brodeur , J . ( 2003 ) Enhanced cellular response through thermoregulation in the locust, Locusta migratoria , infected by the fungus Metarhizium anisopliae . Journal of Invertebrate Pathology , 82 , 103 – 109 .
Ouedraogo , A. , Fargues , J. , Goettel , M.S. & Lomer , C.J . ( 1997 ) Effect of temperature on vegetative growth among isolates of Metarhizium anisopliae and M. flavoviride . Mycopathologia , 137 , 37 – 43 .
Ouedraogo , R.M. , Goettel , M.S. & Brodeur , J . ( 2004 ) Behavioral thermoregulation in the migratory locust: a therapy to overcome fungal infection . Oecologia , 138 , 312 – 319 .
Ouedraogo , R.M. , Kamp , A. , Goettel , M.S. , Brodeur , J. & Bidochka , M.J . ( 2002 ) Attenuation of fungal infection in thermoregulating locusts, Locusta migratoria , is accompanied by changes in hemo-lymphal proteins . Journal of Invertebrate Pathology , 81 , 19 – 24 .
Parsell , D.A. & Lindquist , S . ( 1993 ) The function of heat-shock pro-teins in stress tolerance: degradation and reactivation of damaged proteins . Annual Review of Genetics , 27 , 437 – 496 .
Price , R.E. , Muller , E.J. , Brown , H.D. , Uamba , P.D. & Jone , A.A . ( 1999 ) The first trial of a Metarhizium flavoviride myco-insecticide for the control of the red locust in a recognised outbreak area . Insect Science Application , 19 , 323 – 331 .
Quesada-Moraga , E . ( 1998 ) Biología y ecología de la reproducción y desarrollo embrionario de la langosta mediterránea Dociostaurus maroccanus (Thunberg) y su posible interferencia como estrategia de lucha. PhD Thesis, Universidad de Córdoba .
Ratte , H.T . ( 1985 ) Temperature and insect development . Environ-mental Physiology and Biochemistry of Insects ( ed . by K. H. Hoffman ), pp . 33 – 66 . Springer , New York, New York .
Richmond , J.A. & Bacheler , J.E . ( 1989 ) A comparison of two tem-perature-dependent development models for immature stages of the Nantucket pine tip moth (Lepidoptera: Tortricidae) . Journal of Entomological Sciences , 24 , 117 – 123 .
Samietz , J. & Köhler , G . ( 1998 ) The impact of behavioural thermo-regulation on reproductive rates in a grasshopper . Population and Community Ecology for Insect Management and Conservation
( ed . by J. Baumgartner , P. Brandmayr and F. Manly ), pp . 63 – 73 . Balkema , The Netherlands .
Scanlan , J.C. , Grant , W.E. , Hunter , D.M. & Milner , R.J . ( 2001 ) Hab-itat and environmental factors influencing the control of migratory locusts ( Locusta migratoria ) with an entomopathogenic fungus ( Metarhizium anisopliae ) . Ecological Modelling , 136 , 223 – 236 .
Stower , W.J. & Griffiths , J.F . ( 1966 ) The body temperature of the desert locust ( Schistocerca gregaria ) . Entomologica Experimen-talis et Applicata , 9 , 127 – 178 .
Thomas , M.B . ( 2000 ) Development of a mycoinsecticide for biologi-cal control of locusts in Southern Africa . Research Priorities for Migrant Pests of Agriculture in Southern Africa. Proceedings of a DFID/NRI/ARC-PPRI Workshop , Pretoria, South Africa, 24 – 26 March 1999 ( ed . by R.A. Cheke , L.J. Rosenberg and M.E. Kieser ), pp . 173 – 182 . Natural Resources Institute , U.K .
Thomas , M.B. , Blanford , S. , Gbongboui , C. & Lomer , C.J . ( 1998 ) Ex-perimental studies to evaluate spray applications of a mycoinsecticide against the rice grasshopper, Hieroglyphus daganensis , in northern Benin . Entomologia Experimentalis et Applicata , 87 , 93 – 102 .
Thomas , M.B. & Jenkins , N.E . ( 1997 ) Effects of temperature on growth of Metarhizium flavoviride and virulence to the variegated grasshopper, Zonocerus variegatus . Mycological Research , 101 , 1469 – 1474 .
Thomas , M.B. , Klass , J. & Blanford , S . ( 2000 ) The year of the locust . Pesticide Outlook , 11 , 192 – 195 .
Thomas , M.B. & Kooyman , C . ( 2004 ) Locust biopesticides: a tale of two continents . Biocontrol News and Information , 25 , 47N – 51N .
Thomas , M.B. , Wood , S.N. & Lomer , C.J . ( 1995 ) Biological control of locusts and grasshoppers using a fungal pathogen: the importance of secondary cycling . Proceedings of the Royal Society London Series B , 259 , 265 – 270 .
Thomas , M.B. , Wood , S.N. & Solorzano , V . ( 1999 ) Application of insect-pathogen models to biological control . Theoretical Approaches to Biological Control ( ed . by B.A. Hawkins and H.V. Cornell ), pp . 368 – 384 . Cambridge University Press , U.K .
Uvarov , B . ( 1977 ) Grasshoppers and Locusts , 2 . Centre for Overseas Pest Research, U.K .
Vey , A. & Gotz , A.P . ( 1986 ) Antifungal cellular defence mechanisms in insects . Hemocytic and Humoral Immunity in Arthropods ( ed . by A. P. Gupta ), pp . 89 – 116 . Wiley-Interscience , New York, New York .
Watson , E.L . ( 2002 ) Variability, stability and competitive ability of Metarhizium anisopliae var acridum : implications for the environ-mental impact of locust biocontrol. PhD thesis, Imperial College, London .
Xu , X.-M. ( 1996 ) On estimating non-linear response of fungal development under fluctuating temperatures . Plant Pathology , 45 , 163 – 171 .
Accepted 24 December 2006 First published online 29 March 2007


















