
Developing Synthetic Seeds for Propagation of Australian ... · Developing Synthetic Seeds ... for...
48
Developing Synthetic Seeds for Clonal Propagation of Australian Plants Somatic embryogenesis as a precursor to synthetic seeds A report for the Rural Industries Research and Development Corporation by Dr M Panaia September 2006 RIRDC Publication No 06/058 RIRDC Project No KPW-2A
Transcript of Developing Synthetic Seeds for Propagation of Australian ... · Developing Synthetic Seeds ... for...

Developing Synthetic Seeds for Clonal Propagation
of Australian Plants Somatic embryogenesis as a precursor to synthetic seeds
A report for the Rural Industries Research and Development Corporation
by Dr M Panaia
September 2006
RIRDC Publication No 06/058 RIRDC Project No KPW-2A

ii
© 2006 Rural Industries Research and Development Corporation. All rights reserved. ISBN 1 74151 321 9 ISSN 1440-6845 Developing Synthetic Seeds for Clonal Propagation of Australian plants - Somatic embryogenesis as a precursor to synthetic seeds Publication No. 06/058 Project No. KPW-2A The information contained in this publication is intended for general use to assist public knowledge and discussion and to help improve the development of sustainable industries. The information should not be relied upon for the purpose of a particular matter. Specialist and/or appropriate legal advice should be obtained before any action or decision is taken on the basis of any material in this document. The Commonwealth of Australia, Rural Industries Research and Development Corporation, the authors or contributors do not assume liability of any kind whatsoever resulting from any person's use or reliance upon the content of this document. This publication is copyright. However, RIRDC encourages wide dissemination of its research, providing the Corporation is clearly acknowledged. For any other enquiries concerning reproduction, contact the Publications Manager on phone 02 6272 3186. Researcher Contact Details Dr M Panaia Botanic Gardens & Parks Authority Fraser Avenue, West Perth, WA 6005 Phone: 08 9480 3622 Fax: 08 9480 3641 Email: [email protected]
In submitting this report, the researcher has agreed to RIRDC publishing this material in its edited form. RIRDC Contact Details Rural Industries Research and Development Corporation Level 2, 15 National Circuit, Barton ACT 2600 PO Box 4776 KINGSTON ACT 2604 Phone: 02 6272 4819 Fax: 02 6272 5877 Email: [email protected]. Web: http://www.rirdc.gov.au Published in September 2006 Printed on environmentally friendly paper by Canprint

iii
Foreword Many Australian plants produce poor quality seeds or seeds that are deeply dormant. Successful plant establishment by cuttings can often be difficult and sometimes impossible. Research into new propagation technology is essential for meeting the present and future demands for the efficient mass-propagation of Australian plants to be used in land restoration and for horticultural utilisation. Somatic embryogenesis has a high potential as a clonal propagation method and it has been estimated that 60,000 plants can be obtained from one gram of parent tissue. The aim of this study is to deliver a superior, mass propagation method for native plant taxa that cannot normally be produced by conventional techniques. This study undertakes a detailed examination of the effects of various factors including plant growth regulators and media nutrients on somatic embryogenesis for several species important for the horticultural and rehabilitation industries. The report identifies, evaluates and provides detailed information on Baloskion tetraphyllum (Koala Fern), Macropidia fuliginosa (Black Kangaroo Paw), Stirlingia latifolia (Blue Boy), and Lepidosperma squamatum (a sedge). The first three species are highly desirable in the horticultural industry and both Stirlingia and Lepidosperma are vital species for the rehabilitation of disturbed sites including disused mining land. This project was funded from RIRDC core funds that are provided by the Australian Government. This report is an addition to RIRDC’s diverse range of over 1500 research publications. It forms part of our Wildflower and Native Plants R&D program, which aims to improve the profitability, productivity and sustainability of the Australian wildflower and native plant industry. Most of our publications are available for viewing, downloading or purchasing online through our website: • downloads at www.rirdc.gov.au/fullreports/index.html • purchases at www.rirdc.gov.au/eshop Peter O’Brien Managing Director Rural Industries Research and Development Corporation

iv
Acknowledgments This project was conducted with financial support from the Rural Industries Research and Development Corporation, Botanic Gardens & Parks Authority and Yates Botanicals (Elite Propagation).
Abbreviations ABA abscisic acid ANOVA Analysis of Variance BM basal medium conversion the development of entire plantlets from somatic embryos GA gibberellic acid GB Gamborg (media salt) IAA indole-3-acetic acid K kinetin BGPA Botanic Gardens & Parks Authority MS Murashige & Skoog PFPP photosynthetic photon flux density PGR plant growth regulator(s) SE somatic embryos (se) standard error of treatment means sdw sterile deionised water 2,4-D 2,4-dichlorophenoxyacetic acid TDZ thidiazuron μM microMole per litre μM m-2 s-1 microMoles of photons per square metre per second Z zeatin

v
Contents Foreword ............................................................................................................................................... iii Acknowledgments................................................................................................................................. iv Abbreviations........................................................................................................................................ iv Executive Summary ............................................................................................................................ vii
Background ....................................................................................................................................... vii Objectives.......................................................................................................................................... vii Methodology ..................................................................................................................................... vii Results/Key Findings ....................................................................................................................... viii Implications for relevant stakeholders................................................................................................ ix Recommendations ............................................................................................................................... x
1. Introduction .......................................................................................................................... 1 1.1 What is Somatic Embryogenesis? .......................................................................................... 1 1.2 Overview of Target Species ................................................................................................... 2 1.3 Objectives............................................................................................................................... 2
2. Materials and Methods ........................................................................................................ 3 2.1 Collection of Seed and Plant Material.................................................................................... 3 2.2 Generation of B. tetraphyllum coleoptiles.............................................................................. 3 2.3 Seed sterilisation and excision of zygotic embryos for M. fuliginosa, S. latifolia and
L. squamatum ......................................................................................................................... 3 2.4 Basal medium and common culture conditions...................................................................... 4 2.5 Observations recorded............................................................................................................ 4 2.6 Examination of various plant growth regulators for the stimulation of somatic
embryogenesis in the target species ....................................................................................... 4 2.6.1 Evaluation of 2,4-D to stimulate primary SE in B. tetraphyllum ....................................... 4 2.6.2 Investigation of primary SE as the explant source for secondary somatic embryogenesis
in B. tetraphyllum ............................................................................................................... 4 2.6.3 ABA and its effect on imparting desiccation tolerance to SE of B. tetraphyllum .............. 4 2.6.4 Effect of heat treatment in stimulating germination of M. fuliginosa seed ........................ 5 2.6.5 Influence of 2,4-D on the stimulation of SE for M. fuliginosa using coleoptiles and
excised zygotic embryos .................................................................................................... 5 2.6.6 Proliferation of secondary somatic embryogenesis in M. fuliginosa ................................. 5 2.6.7 Evaluation of 2,4-D on somatic embryogenesis of S. latifolia........................................... 5 2.6.8 Investigation of different colour selections of S. latifolia .................................................. 6 2.6.9 Investigation of four different selections of S. latifolia on Gamborg medium................... 6 2.6.10 Primary somatic embryogenesis of L. squamatum............................................................. 6 2.6.11 Secondary somatic embryogenesis of L. squamatum......................................................... 6
2.7 Desiccation tolerance of L. squamatum SE when treated with ABA..................................... 7 2.8 Statistical Analysis ................................................................................................................. 7
3. Results ................................................................................................................................... 8 3.1 Initiation of somatic embryogenesis for B. tetraphyllum utilising 2,4-D............................... 8 3.2 Primary SE as the explant source for stimulation of secondary SE for B. tetraphyllum...... 10 3.3 ABA and its effect on desiccation tolerance of B. tetraphyllum SE..................................... 11 3.4 Effect of heat treatment on germination of M. fuliginosa seed ............................................ 12 3.5 Influence of 2,4-D on germination and production of SE for M. fuliginosa ........................ 13 3.6 Secondary somatic embryogenesis in M. fuliginosa ............................................................ 14

vi
3.7 Effect of 2,4-D on somatic embryogenesis of S. latifolia .................................................... 18 3.7.1 Comparison of four colour selections of S. latifolia utilising shoot material................... 19 3.7.2 Comparison of basal material of four selections of S. latifolia ........................................ 20 3.7.3 Effect of TDZ and IAA on somatic embryogenesis for S. latifolia.................................. 20 3.7.4 Effects of TDZ and IAA on somatic embryogenesis for S. latifolia when using basal
material as the explant source .......................................................................................... 25 3.7.5 Influence of Gamborg salts on the stimulation of SE for S. latifolia ............................... 25
3.8 Effect of 2,4-D on somatic embryogenesis of L. squamatum .............................................. 27 3.9 Secondary somatic embryogenesis for L. squamatum ......................................................... 29 3.10 Effect of ABA on desiccation tolerance of L. squamatum SE ............................................. 31
4. Discussion............................................................................................................................ 33 4.1 Somatic embryogenesis for B. tetraphyllum ........................................................................ 33 4.2 Response of M. fuliginosa to heat treatment and 2,4-D ....................................................... 34 4.3 Effect of 2,4-D on somatic embryogenesis of S. latifolia .................................................... 35 4.4 L. squamatum and its response to plant growth regulators .................................................. 36
Conclusion............................................................................................................................................ 37 References: ........................................................................................................................................ 38

vii
Executive Summary Background Many desirable Australian plants produce poor quality seeds or seeds that are deeply dormant. Successful plant establishment can often be difficult and sometimes impossible. Research into new propagation technology is essential for meeting the present and future demands for land restoration and horticultural utilisation. Somatic embryogenesis represents a major technology for the low cost, rapid, large-scale production of plant species that respond poorly to conventional methods of propagation. Somatic embryogenesis was first described for carrot (Steward et al. 1958) and has the potential to produce up to 60,000 plants from one gram of parent tissue (Senaratna 1992). It is a process by which somatic (i.e. non-sexual) cells undergo a developmental sequence, similar to that seen in zygotic embryos, without the need for sexual reproduction. Under normal circumstances it is necessary for the pollen (male) to make contact with the ovary (female) of the flower to fertilise the egg and thus produce a seed. In somatic embryogenesis, a variety of plant growth regulators and environmental conditions are applied in vitro to a tissue source (such as shoot material), which causes the cell to produce an embryo that eventually develops into an entire plant. The embryo is formed without genetic recombination and is therefore a clone of the parent plant. The term “synthetic” or “artificial seed” is used to describe somatic embryos (SE) either in a hydrated or desiccated form, with or without encapsulation. The production of synthetic seeds has enormous potential in the agricultural, horticultural and mining industries and the basis for synthetic seed technology is the phenomenon of somatic embryogenesis. During the study, a detailed examination of various factors including plant growth regulators and media nutrients on somatic embryogenesis for Baloskion tetraphyllum (Koala Fern), Macropidia fuliginosa (Black Kangaroo Paw), Stirlingia latifolia (Blue Boy), and Lepidosperma squamatum (a sedge) were investigated. The first three species are highly desirable in the horticultural industry and Stirlingia and Lepidosperma are vital species for the rehabilitation of disturbed landscapes. Objectives The objective of the project was to develop a process for the large-scale production of SE for selected Australian native species to be used as synthetic seeds for the rehabilitation of disturbed landscapes and for horticultural utilisation. More specifically, the project examined the following stages in the development of SE for the above species (1) initiation of SE from primary explants (2) proliferation of embryogenic cultures (3) conversion of SE into functioning plantlets and (4) preliminary investigation into the desiccation ability of SE as a pre-requisite to encapsulation and production of synthetic seeds. Methodology While theoretically every cell has the potential to be “totipotent” (develop into an entire plant), the signalling process is not common for all species. The success of this technique is dependent on the tissue source, physiological status, environmental conditions and the cells response to the exogenous application of plant growth regulators. To that end, specific protocols must be developed for each species of interest.

viii
Trials were undertaken to evaluate the impact of selected plant growth regulators (e.g. auxins, cytokinins and thidiazuron) on the ability of different tissue sources (shoot explants, basal portions and zygotic embryos) to express somatic embryogenic potential. Nutritional conditions for embryo induction including different basal salts and variation in the carbohydrate source were also investigated. Once SE had been successfully stimulated, several experiments were undertaken to improve their ability to withstand desiccation (as a precursor to coating, storage, transportation to site and mechanised sowing) by including abscisic acid (ABA) in the media. Results/Key Findings Somatic embryogenesis was successfully achieved for B. tetraphyllum, M. fuliginosa, S. latifolia, and L. squamatum. Conversion of SE into functioning plantlets was successful for M. fuliginosa, S. latifolia and B. tetraphyllum with plants successfully transferred to soil (70-100% survival) without the addition of plant growth regulators and minimum acclimatisation (1-2 weeks in the glasshouse). As is the case with all species investigated, the stimulation of SE appears to be closely linked to the age of the explant and the younger the material used as the explant, the more successful the process became. Desiccation of SE for L. squamatum and B. tetraphyllum demonstrated encouraging results. A number of SE displayed germination events (after being dried for 2-3 hours) including re-hydration and production of root hairs. These preliminary results clearly demonstrate that drying (desiccation) of SE is possible pending further research to optimise this protocol. The key findings of this study are summarised below:
1. Somatic embryogenesis was successfully achieved for Baloskion tetraphyllum (Koala Fern) with more than 1,500 SE produced in 6 weeks from 0.02 g of parent material. This has the potential to produce ~75,000 SE from 1 g of original parent material.
2. Somatic embryogenesis for Macropidia fuliginosa (Black Kangaroo Paw) varied depending
on the tissue source:
• 196 SE were produced after 6 weeks from 0.05 g of coleoptile explants. This could potentially result in ~3,900 SE from 1 g of parent material. • 562 SE were stimulated when using callus as the explant source. • In excess of 3,200 secondary SE were produced after 6 weeks when using primary SE as the explant source. It is estimated that up to ~26,000 SE could potentially be produced from 1 g of parent material using this protocol.
3. Somatic embryogenesis was achieved for Stirlingia latifolia (Blue Boy) with 53 SE
produced on ½ MS + 1 μM 2,4-D when using callus as the explant source after 6 weeks. Callus that remained in culture for a further 6 weeks continued to produce SE. At the end of 12 weeks, there were a total of 278 SE produced from ~1.235g of callus.
4. Significant differences in the response of S. latifolia selections to various plant growth
regulators were observed. During experiments investigating 2,4-D and TDZ on shoot explants, only Selection 1 (Deep Red) produced entire plantlets (~ 449) after an extensive treatment regime as follows: callus produced on ½ MS + 10 μM 2,4-D was transferred to ½ MS + 5 μM TDZ (for shoot development) and then ½ MS + 5 μM IAA (for root development).
5. Primary SE were produced for three selections of S. latifolia (Deep Red, Dark Red and CY)
when cultured on TDZ, IAA and a combination of these two plant growth regulators. Deep Red responded with the highest number of 170 SE from ~ 0.3 g of shoot material which equates to a possible 566 SE from 1 g of original parent material.

ix
6. There were significant numbers of shoot meristems produced that could be excised and rooted separately for S. latifolia (organogenesis). Selection 4 (CY) had the highest number with 454 shoot meristems produced in 6 weeks from ~0.03 g of shoot material cultured on ½ MS + 10 μM TDZ and 5 μM IAA.
7. SE were successfully stimulated for Lepidosperma squamatum using zygotic embryos as the
explant source. A total of ~470 SE were produced from ~0.05 g which is estimated could produce ~9,400 SE from 1 g of zygotic tissue.
8. Proliferation of secondary somatic embryogenesis was achieved for Lepidosperma
squamatum with a ~ 3-fold increase when primary SE were subcultured onto fresh media.
9. Callus stimulated the production of secondary SE when cultured on a variety of media. The best treatment for Lepidosperma squamatum was a combination of ½ MS + 1 μM 2,4-D, followed by transfer to ½ MS + 5 μM TDZ with a total of 148 secondary SE from ~1.2 g of callus.
10. Conversion of SE into functioning plantlets was achieved for M. fuliginosa, S. latifolia and
B. tetraphyllum with plants being transferred to soil with 70-100% survival.
11. Desiccation of SE for B. tetraphyllum demonstrated encouraging results with ~ 50 % displaying germination events (after being dried for 2-3 hours) including re-hydration and the production of root hairs when cultured on ½ MS + 20 μM ABA for one week prior to drying.
12. Desiccation of L. squamatum primary SE was also successful with the best treatment
being ½ MS + 50 μM ABA for one week prior to the drying treatment. Although the number of rehydrated SE is small, these preliminary results clearly demonstrate that drying (desiccation) of SE is possible pending optimisation of this protocol.
Implications for relevant stakeholders The mining of native ecosystems (including the successful restoration of such sites) and the development of horticultural industries largely depend on the development of an efficient and cost effective propagation technique that will provide acceptable levels of biodiversity replacement. The estimated cost of producing a single plant under normal micropropagation methods can vary from less than $1 to over $5 depending on the species and how difficult it is to initiate into tissue culture. In general, 15-20 plantlets can be produced in one tissue culture vessel compared to approximately 200 SE per petri dish. Somatic embryogenesis is therefore at least 10 times more efficient, even with non-optimal protocols, and, as such, the cost can conservatively be estimated at 10 to 50 cents per plant. This is a significant saving over current production costs. Mining in biodiverse regions or highly diverse floras such as the southern parts of Western Australia often means that significant components of the biome are not effectively rehabilitated. For example, southern rushes (Restionaceae), sedges (Cyperaceae) and native heaths (Ericaceae) represent up to 30 % of pre-mined diversity and biomass, yet current rehabilitation technology returns barely 1 – 2 % (Willyams 2005a). In addition, the horticultural potential of unique Australian plants has not yet been fully exploited. The Black Kangaroo Paw (Macropidia fuliginosa) and Koala Fern (Baloskion tetraphyllum) are in heavy demand in the horticultural industry, but are slow to multiply in sufficient numbers to satisfy this demand. A considerable amount of native plant material is still wild sourced, which leads to significant problems in quality and guaranteed supply as well as the environmental damage and loss of biodiversity. The use of somatic embryogenesis as a technique to deliver a low cost, superior, mass propagation method for these native plant taxa will effectively reduce the cost of production, reduce loss of biodiversity, increase the efficiency of rehabilitation programmes and boost the supply of these species to the horticultural industry.

x
Recommendations This project has established a sound foundation to continue work on these species to improve the process of somatic embryogenesis. It was clear that significant variation occurred between different tissue types and this phenomenon requires further investigation to optimise protocols and consistently produce large numbers of SE with a wide range of selected genotypes. One of the challenges associated with somatic embryogenesis is that germination and growth of plantlets is often not as vigorous as that of their zygotic counterparts. This is, in part, due to the lack of storage reserves in the SE. As with other characteristics of somatic embryogenesis, the amendment of the initiation media through the addition of plant growth regulators may increase the storage proteins in SE, however, this is yet to be determined for the species investigated in this study and it is recommended that this be a priority research area in the future. Once synchronous production of SE has been established, it is important that SE be dried for storage purposes without the loss of viability. While preliminary desiccation experiments were encouraging, follow-up research is required investigating factors such as heat shock, non-lethal chilling or osmotic stress to promote desiccation tolerance. Low levels of ABA are present at the onset of reserve accumulation in zygotic seeds, but rise with the onset of desiccation tolerance. ABA is thought to be involved in the accumulation of storage reserves and it is important therefore that its role in promoting desiccation tolerance does not interfere with the deposition of storage reserves. Investigations are therefore required to determine the levels and timing of application to induce desiccation tolerance without negatively influencing reserve accumulation. One of the final stages is to establish a protocol for the encapsulation of SE to create synthetic seeds. Encapsulation provides protection from mechanical damage as well as delivering plant growth regulators, nutrients and other chemical or biological components required for rapid germination and plant establishment. This aspect of the development of synthetic seeds remains to be done. However, the present study has provided significant advances in production of SE from recalcitrant species that will facilitate progression to synthetic seed production.

1
1. Introduction In vitro propagation has been extensively developed over the past 40 years and is used as a method of clonal plant propagation. The technique utilises the ability of a single plant cell to regenerate into an entire plant (totipotency) when provided with adequate conditions for growth and development (Vasil and Thorpe 1994). This is especially advantageous when parent material is rare, when there is a lack of seed or suitable growing tips are not available for conventional vegetative propagation. Advances in plant propagation techniques have made it possible to propagate previously recalcitrant Australian plant species but, in some vital cases, have not been universally successful with species belonging to the Restionaceae, Cyperaceae (Meney and Dixon 1995), Proteaceae and Haemodoraceae. 1.1 What is Somatic Embryogenesis? Somatic embryogenesis represents a major technology for the rapid large-scale propagation of important plant species where conventional micropropagation techniques have not been successful and provides the basis for producing artificial seeds (Vajrabhaya 1988). It is a process by which somatic (non-sexual) cells undergo a developmental sequence, similar to that seen in zygotic embryos, without the need for fusion of gametes (pollen and ovary) (Cervelli and Senaratna 1995). In contrast to organogenesis, where cells undergo changes leading to the production of a unipolar structure (either a root or a shoot), somatic embryogenesis results in a bipolar structure with both root and shoot meristems and an independent vascular system. The switch from a non-sexual cell to embryo formation is stimulated by a variety of chemicals or plant growth regulators (PGR) and environmental conditions. There are several advantages associated with this technique. For example, during conventional micropropagation techniques, explants must be dis-infested to remove bacterial and fungal contamination and micro-shoots are produced which must then be rooted in a separate procedure (Cervelli and Senaratna 1995). Rooting of native species is often problematic, requiring extensive investigations into the types, levels and duration of exposure to auxins to stimulate effective rooting (Taji and Williams 1996). An acclimatisation period (up to 4 weeks) on PGR-free medium is then usually required before plantlets can be successfully transferred to soil. In contrast, SE are bipolar structures (as described above) and as such there is great potential for a reduction in labour, time and cost (Cervelli and Senaratna 1995). The potential therefore exists for the production of higher numbers of plants, compared to conventional methods, with morphological and cytological uniformity and this is particularly important for commercial applications and germplasm preservation. It is possible for thousands of propagules to be produced by somatic embryogenesis from a very small amount of plant tissue [i.e. an estimated 60,000 units per gram of tissue (Senaratna et al. 1990). A number of PGR, particularly auxins, have been demonstrated to stimulate somatic embryogenesis (Zimmerman 1993). It is known that auxins are involved in the regulation of elongation and growth in young stems and coleoptiles and can also affect root initiation and lateral bud dormancy (Taiz and Zeiger 1991). Auxins are often required for the induction of callus from explants (George 1996) and the synthetic auxin analogue 2,4-D is commonly utilised for this purpose. In general, once cells have fully differentiated and matured they do not divide again during the life of the plant (Taiz and Zeiger 1991). However, plant cells that retain a nucleus at maturity can be forced by exogenous events (such as the application of PGR) to divide (Taiz and Zeiger 1991). Cytokinins are noted for their ability to stimulate cell division and are usually found in young, rapidly dividing cells of the shoot and root apical meristems. Cytokinins are also known to affect the rate of protein synthesis (Taiz and Zeiger 1991) that in turn influences cell division. Thidiazuron (TDZ) is a highly phyto-active phenylurea compound that has been shown to stimulate somatic embryogenesis by mimicing the actions of both cytokinins and auxins combined (Murthy et al. 1995; Hutchinson and Saxena 1996) and was therefore chosen for investigation in this study.

2
1.2 Overview of Target Species The following provides a brief description of each species investigated in this study. Baloskion tetraphyllum (J.J.H. de Labillardiere) B.G. Briggs & L.A.S. Johnson (Restionaceae) is confined to eastern Australia and extends from North Queensland to Tasmania and South Australia. It grows in sand or peaty soil in swamps and along stream banks and is an obligate seeder, regenerating from seed mostly after fire with seed maturation taking 6-8 months (Meney and Pate 1999). Koala Fern (common name) is prized as a cut flower and is either wild picked or propagated by seed (when available). Propagation is both labour intensive and time consuming, whilst wild picking can have detrimental conservation outcomes. Macropidia fuliginosa (Hook.) Druce. Commonly called the Black Kangaroo Paw, is a striking plant that has become extremely popular in the horticultural industry, so much so, that supply is unable to keep up with demand. It is a rhizomatous, perennial herb ranging in height from 0.2 to 1.8m. It has magnificent green/black flowers from August through to December. It is found in the south west of Western Australia including the Avon wheatbelt, the Geraldton Sandplains and the Swan Coastal Plain. Stirlingia latifolia (R.Br.) Steud. Is an erect, lignotuberous shrub ranging from 0.2 to 1.5m high. Flowers vary in colour from yellow, through brown and red and occur from August to October. It is commonly called Blue Boy due to the “blue/grey” colour of the striking foliage. It is typically found on white, grey, yellow-brown or black sand, sometimes with lateritic gravel. It is distributed throughout jarrah forests in the south west of Western Australia, including the Esperance Plains, the Geraldton sandplains, the Swan Coastal Plain and the Warren region. Lepidosperma squamatum R.Br. (previously L. angustatum) (Cyperaceae) is widespread and common in a variety of soils and habitats from heath and shrubland to woodland. The species occurs from the north of Geraldton to Israelite Bay (Wheeler 2002) and is of high value for revegetation of disused mining sites within Western Australia requiring large numbers for successful rehabilitation. 1.3 Objectives The objective of the project is to develop a process for the large-scale production of SE for Australian native species to be used as synthetic seeds for the rehabilitation of disturbed landscapes and for horticultural utilisation. More specifically, the project will examine the following stages in the development of SE for the above species (1) initiation of SE from primary explants (2) proliferation of embryogenic cultures (3) conversion of SE into functioning plantlets and (4) a preliminary investigation into the desiccation ability of SE as a pre-requisite to encapsulation and production of synthetic seeds.

3
2. Materials and Methods The following materials and methods are common to all plant species investigated and experiments in this study unless otherwise stated. 2.1 Collection of Seed and Plant Material Tissue cultured plantlets and seed for the target species were obtained from a variety of sources (Table 1). Cultures were maintained on 1/2 strength Murashige & Skoog (MS)(Murashige and Skoog 1962)medium free of plant growth regulators (PGR) for a minimum of 4 weeks before treatments were applied. Table 1: Sources of tissue cultured plant material and seed. Family Species Name Source Restionaceae
Baloskion tetraphyllum (J.J.H. de Labillardiere) B.G. Briggs & L.A.S. Johnson
Wildseed Tasmania, Tasmania (seed). Ellison Horticultural Pty Ltd, NSW (seed).
Cyperaceae Lepidosperma squamatum R.Br.
Alcoa Australia (Tissue cultured plant material and seed.)
Proteaceae Stirlingia latifolia (R.Br.) Steud.
Botanic Gardens & Parks Authority (Tissue culture plant material and seed.)
Haemodoraceae Macropidia fuliginosa (Hook.) Druce.
Botanic Gardens & Parks Authority – seed. Yates Botanicals (Tissue culture plant material.)
2.2 Generation of B. tetraphyllum coleoptiles Seeds of B. tetraphyllum were extracted from their outer culms by abrasion between two pieces of sandpaper. The seed and culm mixture was passed through a 60 µm wire sieve to separate the “naked” seed from debris. Seeds were surface sterilised in 1.0 % (w/v) calcium hypochlorite [Ca(OCl)2] for 15 min followed by rinsing 3 times in sterile deionised water (sdw) and transferred to 1/2 MS medium solidified with 6 g L-1 agar and 20 g L-1 sucrose, pH 6.0 (adjusted prior to autoclaving) for germination. Observations were made daily until coleoptiles had reached a minimum length of ~10 mm and then excised into ~2 mm sections for transfer to treatments. 2.3 Seed sterilisation and excision of zygotic embryos for M. fuliginosa, S. latifolia and L. squamatum Seeds were surface sterilised in 1.0 % (w/v) Ca(OCl)2 for 5/5/5 mins on/off/on under vacuum and rinsed 3 times in sdw before aseptically removing the seed coat. The intact endosperm, containing the embryo, was further sterilised with 1.0 % (w/v) Ca(OCl)2 for 5 mins, rinsed 3 times in sdw and imbibed overnight in sdw. Zygotic embryos of M. fuliginosa and L. squamatum were aseptically excised, while the entire endosperm for S. latifolia were transferred to the appropriate treatment medium.

4
2.4 Basal medium and common culture conditions Basal medium (BM) consisted of 1/2 strength MS salts supplemented with 20 g L-1 sucrose, 6 g L-1 agar, pH 6.0 (adjusted prior to autoclaving) with the following additional components: 500 μM Myo-inositol, 4 μM Niacin, 3 μM Thiamine HCl and 2.5 μM Pyridoxine HCl. All non-heat labile PGR, mineral salts, organics and 2,4-D were added to the media prior to autoclaving, whilst heat labile PGR (IAA, zeatin, ABA and TDZ) were filter sterilised and added after autoclaving. Explants were cultured in petri dishes containing ~25 ml of treatment medium and observations were recorded weekly. In general, cultures were maintained in the dark at 22-25 ˚C for a minimum of 6 weeks. Light incubation (when required) consisted of 16 h at ~ 30 µM m-2 s-1 PPFD and 8 h dark. Experiments comprised 10 explants per petri dish (unless otherwise stated) with a minimum of 3 replicates per treatment. 2.5 Observations recorded The numbers of SE were recorded on a weekly basis. Callus was classified by width and given a numeric score as follows: 1 = 1-3 mm, 2 = 4-6 mm, 3 = 7-9 mm, 4 = ≥10 mm. The number of shoot meristems was recorded weekly. Swelling of the explant (as an indicator of response) was assessed as absent (0) or present (1). Abundance of root hair was classified using a hedonic scale of 1 to 4 with 1 representing the first visible evidence of root hair and 4 representing total coverage of the explant/callus/SE with root hair. Conversion of SE into plantlets was calculated based on the total number of well-developed plantlets obtained in each treatment in relation to the original number of SE. 2.6 Examination of various plant growth regulators for the stimulation of somatic embryogenesis in the target species Explant sterilisation, preparation, BM and culture conditions were as previously described for all experimental work unless otherwise specified. 2.6.1 Evaluation of 2,4-D to stimulate primary SE in B. tetraphyllum Coleoptiles (~ 2 mm) were initiated, excised and transferred to the following treatments to stimulate SE: ½ MS (control), ½ MS + 1 μM 2,4-D or ½ MS + 10 μM 2,4-D. 2.6.2 Investigation of primary SE as the explant source for secondary somatic embryogenesis in B. tetraphyllum Primary SE initiated in 2.6.1 were transferred to ½ MS (control), ½ MS + 1 μM 2,4-D or ½ MS + 10 μM 2,4-D and maintained in the dark for five weeks. The numbers of secondary SE were recorded on a weekly basis. 2.6.3 ABA and its effect on imparting desiccation tolerance to SE of B. tetraphyllum Primary SE of B. tetraphyllum were subcultured onto various ABA concentrations to determine whether ABA imparted desiccation tolerance. Treatments included: ½ MS (not dried), ½ MS (dried), ½ MS + 1 μM ABA or ½ MS + 20 μM ABA. SE were maintained on the treatments for one week and then dried in a laminar flow for 3 hours prior to being transferred to ½ MS + 3 μM GA for a further 5 weeks. Observations were made daily to determine whether any SE had survived the drying process. The experiment was repeated with higher levels of ABA: ½ MS (control), ½ MS + 20 μM ABA, ½ MS + 50 μM ABA and ½ MS + 100 μM ABA.

5
2.6.4 Effect of heat treatment in stimulating germination of M. fuliginosa seed To obtain coleoptiles to be used as the explant source for experimentation, it was necessary to germinate M. fuliginosa seed. Heat treatment consisted of 3 hours at 100oC, while controls received no heat treatment (100 seed per treatment). Seeds were allowed to cool for 30 mins prior to being surface sterilised for 15 mins with 2 % (w/v) Ca(OCl)2 with constant agitation, and then rinsed in sdw 3 times. All seeds were left to imbibe 24 hrs in sdw and subsequently transferred to water agar for germination in the dark. 2.6.5 Influence of 2,4-D on the stimulation of SE for M. fuliginosa using coleoptiles and excised zygotic embryos Coleoptiles (~ 2 mm) generated in 2.6.4 were transferred to ½ MS (control), ½ MS + 1 μM 2,4-D or ½ MS + 10 μM 2,4-D and maintained in the dark for 6 weeks. In addition, 21 zygotic embryos (for each treatment) were excised from M. fuliginosa seed and subcultured onto the same three media to determine whether this tissue source was competent to produce SE. 2.6.6 Proliferation of secondary somatic embryogenesis in M. fuliginosa Primary SE initiated on ½ MS + 1 μM 2,4-D were subcultured onto the following treatments to stimulate conversion into plantlets or the production of secondary SE: ½ MS (control), ½ MS + 0.5 μM Z, ½ MS + 0.5 μM GA, or ½ MS + 1 μM 2,4-D. There were 6 replicates for each treatment with 10 primary SE per replication. Seeds of M. fuliginosa were surface sterilised in 2 % w/v Ca(OCl)2 for 15 mins, rinsed with sdw, imbibed for 24 hrs and then zygotic embryos aseptically extracted. Ten embryos were transferred to either: ½ MS (control), ½ MS + 1 μM 2,4-D or ½ MS + 10 μM 2,4-D. Primary SE developed in the above experiment were subsequently transferred to ½ MS (for conversion into plantlets) or ½ MS + 1 μM 2,4-D for the stimulation of secondary SE. Callus developed in the above experiment was used as the explant source and transferred to ½ MS (to stimulate conversion into plantlets) or ½ MS + 1 μM 2,4-D for secondary SE. 2.6.7 Evaluation of 2,4-D on somatic embryogenesis of S. latifolia Seeds were surface sterilised in 1 % (w/v) Ca(OCl)2 for 5 mins, rinsed 3 times in sdw and imbibed in sdw for 24 hrs. The exterior coat was removed and the endosperm transferred to the following treatments: ½ MS (control), ½ MS + 1 μM 2,4-D and ½ MS + 10 μM 2,4-D. There were 5 endosperms per plate with 5 replicates per treatment. Experimental units were maintained in the dark. Callus produced in this experiment was subsequently transferred to ½ MS (control), ½ MS + 1 μM 2,4-D and ½ MS + 10 μM 2,4-D for further development.

6
2.6.8 Investigation of different colour selections of S. latifolia 2.6.8.1 Comparison of shoot material of Deep Red, Dark Red, Coral and CY Stirlingia selections when treated with ½ MS and 2,4-D. Shoot material (~ 2 mm) of four different selections of S. latifolia was aseptically excised and transferred to the following media to induce somatic embryogenesis: ½ MS (control), ½ MS + 1 μM 2,4-D and ½ MS + 10 μM 2,4-D. 2.6.8.2 Comparison of basal material of Deep Red, Dark Red, Coral and CY Stirlingia selections when treated with ½ MS and 2,4-D. Basal material of four selections of S. latifolia were aseptically excised and transferred to the following media to induce somatic embryogenesis: ½ MS (control), ½ MS + 1 μM 2,4-D and ½ MS + 10 μM 2,4-D. 2.6.8.3 Comparison of shoot material of Deep Red, Dark Red, Coral and CY Stirlingia selections when treated with ½ MS supplemented with TDZ and IAA. Shoot material (~ 2 mm) of four different selections of S. latifolia was aseptically excised and transferred to the following media to induce somatic embryogenesis: ½ MS (control), ½ MS + 10 μM TDZ, ½ MS + 5 μM IAA and ½ MS + 10 μM TDZ + 5 μM IAA. 2.6.8.3 Comparison of basal material of Deep Red, Dark Red, Coral and CY Stirlingia selections when cultured on ½ MS with TDZ and IAA. Basal material of four selections of S. latifolia were aseptically excised and transferred to the following media to induce somatic embryogenesis: ½ MS (control), ½ MS + 10 μM TDZ, ½ MS + 5 μM IAA and ½ MS + 10 μM TDZ + 5 μM IAA. 2.6.9 Investigation of four different selections of S. latifolia on Gamborg
medium Shoot material (~ 2 mm) of four different selections of S. latifolia (Deep Red, Dark Red, Coral and CY) was aseptically excised and transferred to the following media to induce somatic embryogenesis: ½ GB (control), ½ GB + 1 μM 2,4-D, ½ GB + 10 μM 2,4-D and ½ GB + 100 μM 2,4-D. 2.6.10 Primary somatic embryogenesis of L. squamatum Seeds of L. squamatum were surface sterilised as previously described for other species and zygotic embryos aseptically excised and transferred to the following media for stimulation of SE: ½ MS (control), ½ MS + 1 μM 2,4-D and ½ MS + 10 μM 2,4-D. 2.6.11 Secondary somatic embryogenesis of L. squamatum Primary SE of L. squamatum from the above experiment (2.6.10) were cultured onto the following treatments to determine whether secondary somatic embryogenesis was possible: ½ MS (control), ½ MS + 1 μM 2,4-D and ½ MS + 10 μM 2,4-D.

7
2.7 Desiccation tolerance of L. squamatum SE when treated with
ABA A preliminary experiment using SE of L. squamatum was undertaken using the following media: ½ MS or ½ MS + 20 μM ABA for one week prior to drying in the laminar flow. This experiment was repeated with an increase in the concentrations of ABA, namely: ½ MS, ½ MS + 20 μM ABA, ½ MS + 50 μM ABA and ½ MS + 100 μM ABA to determine whether the higher concentrations of ABA increased the survival of SE. 2.8 Statistical Analysis Data were statistically analysed using ANOVA (one-way or balanced) where appropriate. Mean comparisons between treatments were made using Fisher’s Protected LSD with 95 % confidence intervals.

8
3. Results 3.1 Initiation of somatic embryogenesis for B. tetraphyllum
utilising 2,4-D. Stimulation of primary SE was achieved using young (5-7 day old), white coleoptiles of B. tetraphyllum cultured on ½ MS supplemented with either 1 μM or 10 μM 2,4-D (Figure 1). The best treatment ½ MS + 1 μM 2,4-D produced significantly (p<0.05) more SE than 10 μM 2,4-D with twice the number of SE produced (Figure 2).
Figure 1: Mean number of SE produced after 6 weeks for each coleoptile explant (~ 2 mm) of B. tetraphyllum on two concentrations of 2,4-D. (Vertical bars represent standard error of the means.)
Figure 2: Total number of B. tetraphyllum SE produced after 6 weeks of culture on either ½ MS + 1 μM or 10 μM 2,4-D using a total of 30 white coleoptiles per treatment (weighing ~ 0.02g) as the explant source.

9
A B
C
Plate 1: Stages of somatic embryogenesis in Baloskion tetraphyllum. A: Numerous somatic embryos that have developed on ~2 mm section of coleoptile (bar = 0.5mm). B: A single germinating somatic embryo with evidence of root hair production (bar = 5mm). C: Plantlets developed from somatic embryos successfully transferred to soil (bar = 200mm).

10
3.2 Primary SE as the explant source for stimulation of secondary SE for B. tetraphyllum Primary SE were utilised as the explant source for this experiment. Stimulation of secondary somatic embryogenesis was successfully achieved with the best treatment ½ MS + 1 μM 2,4-D (mean 58.53), followed by ½ MS + 10 μM 2,4-D (25.23 SE) per explant (p<0.05) (Figure 3). The total number of secondary SE produced in five weeks is shown below (Figure 4).
Figure 3: Mean number of secondary SE produced after 5 weeks when using primary SE of B. tetraphyllum as the explant source. (Vertical bars represent standard error of the means.)
Figure 4: Total number of secondary SE produced after 5 weeks for B. tetraphyllum utilising primary SE (~ 3 SE/explant) as the tissue source on ½ MS (control) or ½ MS supplemented with 1 μM or 10 μM 2,4-D. There was a significant difference (p<0.05) in the number of primary and secondary SE produced for B. tetraphyllum (Figure 5) depending on the source of explant material. Primary SE were produced using young coleoptiles and secondary SE were produced using primary SE as the tissue source.

11
Figure 5: Primary somatic embryogenesis (using young, white coleoptiles as the explant source) versus secondary somatic embryogenesis (using primary SE as the explant source) for B. tetraphyllum. (Vertical bars represent standard error of the means.) 3.3 ABA and its effect on desiccation tolerance of B. tetraphyllum SE Primary SE of B. tetraphyllum were subcultured onto various concentrations of ABA for one week prior to drying to determine whether ABA imparted desiccation tolerance. SE were then dried for 3 hours prior to being transferred to ½ MS + 3 μM GA for a further 5 weeks. A number of SE survived the drying process (Figure 6) showing evidence of re-hydration and production of root hairs.
Figure 6: The percentage of B. tetraphyllum SE that survived drying (3 hours) in the laminar flow after treatment with various concentrations of ABA for one week. Measurements were taken after 5 weeks incubation.

12
The experiment was repeated with higher levels of ABA. The controls (no drying) had the highest survival rate (to be expected) followed by ½ MS + 20 μM ABA (Figure 7).
Figure 7: The percentage of B. tetraphyllum SE that survived 3 hours drying in the laminar flow after treatment with various concentrations of ABA. Measurements were taken after 5 weeks incubation. 3.4 Effect of heat treatment on germination of M. fuliginosa seed A heat treatment was investigated to aid the germination of M. fuliginosa seeds to stimulate coleoptiles. There was a significant difference (p<0.05) in the number of seeds that germinated with heat (7%) and without heat (0%) (data not shown). Coleoptiles were then utilised to stimulate SE with a significant difference in the number of SE produced on ½ MS + 1 µM 2,4-D compared to the control and ½ MS + 10 µM 2,4-D (Figure 8).
Figure 8: Effect of 2,4-D on the stimulation of primary SE after 6 weeks culture when using coleoptiles of M. fuliginosa.

13
3.5 Influence of 2,4-D on germination and production of SE for M. fuliginosa There were a total of 21 zygotic embryos extracted from their outer seed coats for M. fuliginosa with 17 embryos germinating on ½ MS alone. The inclusion of 2,4-D in the media completely inhibited germination (Figure 9).
Figure 9: Total number of extracted embryos of M. fuliginosa and the total number of germinating embryos cultured on the control (1/2 MS) and two concentrations of 2,4-D. Coleoptiles produced on the above controls (Figure 9) were subsequently transferred to ½ MS with 1 or 10 µM 2,4-D to stimulate SE (Figure 10), whilst callus produced (from 1 or 10 µM 2,4-D, data not shown) was transferred to ½ MS alone to determine whether it was embryogenic. The total number of SE was significantly different (p<0.05) with ½ MS + 1 µM 2,4-D being the best treatment (Figure 11).
Figure 10: Mean response per explant of swelling, callus and root hair for M. fuliginosa when using coleoptiles or callus previously formed. (Vertical bars represent the standard error of the means.)

14
Figure 11: Total number of SE produced from coleoptiles or callus of M. fuliginosa on ½ MS supplemented with 1 or 10 μM 2,4-D after 6 weeks incubation. 3.6 Secondary somatic embryogenesis in M. fuliginosa Primary SE developed in experiment 3.5 were transferred to the following treatments for conversion into plantlets and/or stimulation of secondary SE. Treatment ½ MS + 1 μM 2,4-D successfully produced secondary SE (Figure 12). The control (1/2 MS) and the other treatments (Z and GA) did not differ significantly in their ability to “convert” SE into functioning plantlets (Figure 12).
Figure 12: Effect on stimulation of root hair, the production of secondary SE, germinating SE and the production of callus of M. fuliginosa when using primary SE as the explant source. (Vertical bars represent standard error of the means.)

15
. A B
C D
E F
Plate 2: Overview of somatic embryogenesis in Macropidia fuliginosa. A: Many somatic embryos clumped together on callus (bar = 0.5mm). B: A single somatic embryo (bar = 0.1mm). C: Evidence of root hair development (bar = 0.1mm). D: Production of multiple roots and root hair (bar = 1mm). E: A single somatic embryo converted into a functioning plantlet (bar = 2mm). F: Successful transfer to soil with flowering (bar = 7cm).

16
Zygotic embryos for M. fuliginosa were aseptically excised and transferred to various treatments. The stimulation of direct SE was low, however, there was a significant difference with ½ MS + 1 μM 2,4-D producing more SE than the other treatments (Figure 13).
Figure 13: A number of zygotic embryos germinated on ½ MS alone for M. fuliginosa after 6 weeks, with the remaining treatments producing a small number of SE. (Vertical bars represent the standard error of the means.) Yellow callus produced in the above experiment (Figure 13) on ½ MS + 10 μM 2,4-D was used as the primary tissue source and transferred to ½ MS alone to stimulate conversion into plantlets or secondary somatic embryogenesis. There was no evidence of conversion, but a significant number of secondary SE were produced (Figure 14).
Figure 14: Mean production of callus, secondary SE and root hair following 6 weeks incubation for M. fuliginosa when utilising yellow callus as the tissue source. (Vertical bars represent the standard error of the means.)

17
Primary SE from the above experiment (Figure 14) were used as the tissue source for this experiment. SE transferred to ½ MS alone converted into entire plantlets with the production of root hairs and those transferred to ½ MS + 1 μM 2,4-D produced secondary SE (Figure 15) with a total of ~3,200 secondary SE produced in 6 weeks.
Figure 15: Conversion of M. fuliginosa primary SE on ½ MS alone after 6 weeks compared to the production of secondary SE when cultured on ½ MS + 1 μM 2,4-D for 6 weeks. (Vertical bars represent standard error of the means.) All SE transferred to 1/ 2 MS converted into plantlets and were successfully transferred to soil (Figure 16). A multiplication factor of ~5.8 is inferred as observations showed that what appeared to be a single SE actually produced 6 plantlets.
Figure 16: Total number of plantlets produced (conversion) from primary SE of M. fuliginosa when transferred to ½ MS alone. There were a total of 175 plantlets transferred to soil from the original 30 SE.

18
3.7 Effect of 2,4-D on somatic embryogenesis of S. latifolia Endosperm were aseptically excised and used as the tissue source for the following experiment. There was evidence of swelling and germination of S. latifolia endosperm. Callus was produced on both concentrations of 2,4-D (Figure 17) over 7 weeks.
Figure 17: Evidence of swelling, callus and germination of endosperm for S. latifolia when cultured on ½ MS alone, or ½ MS supplemented with 1 or 10 μM 2,4-D after 7 weeks incubation. (Vertical bars represent the standard error of the means.) Callus produced in the above experiment (Figure 17) was subsequently transferred to ½ MS alone for conversion into plantlets. All callus appeared embryogenic as significant numbers of plants were produced (Figure 18) and successfully transferred to soil.
Figure 18: Conversion of callus into entire plantlets of S. latifolia once transferred to ½ MS alone (at 6 weeks). Treatments in brackets were the pre-treatments prior to transfer to ½ MS PGR free medium. A number of callus explants were left for a further 5 weeks in culture and continued to produce SE with significantly (p<0.05) more SE on ½ MS + 1 µM 2,4-D (mean ~11.12 SE) compared to 10 µM 2,4-D (mean ~4.52 SE). The total number of SE produced were 0, 278 and 113 SE respectively (Figure 19) for the control, ½ MS + 1 µM 2,4-D and ½ MS + 10 µM 2,4-D.

19
Figure 19: The total number of SE for S. latifolia following 12 weeks culture on the control (1/2 MS) or ½ MS supplemented with 1 or 10 µM 2,4-D. 3.7.1 Comparison of four colour selections of S. latifolia utilising shoot material There were 4 flower colour selections of S. latifolia available for experimental purposes. Results demonstrated a significant variation in the response of these selections when using shoot material for treatment with 2,4-D with Selections 3 (Coral) and 4 (CY) producing more callus than the other two selections (Figure 20). However, when the callus was transferred to subsequent media to induce somatic embryogenesis, the only selection to produce entire plantlets was Selection 1 (Deep Red) with a total of 449 plantlets produced after an extensive treatment regime as follows: callus was produced on ½ MS + 10 μM 2,4-D from shoot material. The callus was transferred to ½ MS + 5 μM TDZ (for shoot development) and then ½ MS + 5 μM IAA (for root development). As the treatment included both cytokinins and auxins, it is unclear whether the callus was embryogenic or whether shoot and root development had been stimulated separately (organogenesis).
Figure 20: Mean callus and swelling response for S. latifolia selections on two concentrations of 2,4-D when using shoot material as the explant source. Key: 1 = Deep Red, 2 = Dark Red, 3 = Coral, 4 = CY. (Vertical bars represent the standard error of the means.)

20
3.7.2 Comparison of basal material of four selections of S. latifolia In contrast to the above experiment, when using basal material of the four different selections, there was no significant difference between them. The results shown are therefore the mean for all four selections. There were no SE produced for any selection, but evidence of swelling, callus, green shoots and root hair (Figure 21).
Figure 21: Effect of two concentrations of 2,4-D on swelling, callus production, browning, green shoots and root hair of four different selections of S. latifolia when using basal portions as the explant source. (Results are the mean of all Selection data.) (Vertical bars represent the standard error of the means.) 3.7.3 Effect of TDZ and IAA on somatic embryogenesis for S. latifolia TDZ and IAA have been shown to produce SE in some species and were investigated with shoot material from the four different selections of S. latifolia. As there was a significant difference (p<0.05) in the response of the different selections, the results have been presented in separate graphs. There was evidence of swelling, callus and root hair production as well as a number of SE produced for Selection 1 (Deep Red) (Figure 22, Figure 23); Selection 2 (Dark Red) (Figure 24, Figure 25); Selection 3 (Coral) (Figure 26, Figure 27) and Selection 4 (CY) (Figure 28, Figure 29).
Figure 22: Effect of TDZ and IAA on swelling, callus and the number of SE when using shoot material of Selection 1 (Deep Red) S. latifolia. (Vertical bars represent the standard error of the means.)

21
Figure 23: The total number of SE produced for S. latifolia, Selection 1 (Deep Red) after 6 weeks incubation when using shoot material on TDZ and IAA.
Figure 24: Effect of TDZ and IAA on swelling, callus and number of SE when using shoot material of Selection 2 (Dark Red) S. latifolia. (Vertical bars represent the standard error of the means.)

22
A B
C D
E F
G H
Plate 3: Various stages of somatic embryogenesis in Stirlingia latifolia. A: Individual somatic embryos on a shoot explant (bar = 0.3mm). B: Callus formation at the end of the explant (bar = 0.8mm). C: Callus eventually overwhelms the original explant (bar = 1mm). D: Individual somatic embryos that are germinating (bar = 0.5mm). E: Conversion of somatic embryos into plantlets in vitro (bar = 20mm). F: Successful transfer to pots (bar = 70mm). G: Fully grown plant of S. latifolia in its natural environment (bar = 150mm). H: Close up of the flower head (bar = 20mm).

23
The following graph shows the total number of germinating SE (38) and the total number of shoot meristems (321) produced on TDZ and IAA when using shoot material of Selection 2 (Dark Red). (Figure 25).
Figure 25: Total number of germinating SE and shoot meristems produced from shoot material of Selection 2 (Dark Red) of S. latifolia when on IAA alone or combined with TDZ after 6 weeks incubation. The following graph depicts the mean per explant of various observations when using shoot material of Coral selection on TDZ and IAA (Figure 26).
Figure 26: Effect of TDZ and IAA on swelling, callus and production of shoot meristems after 6 weeks incubation when using shoot material of Selection 3 (Coral) S. latifolia. (Vertical bars represent the standard error of the means.) The following graph details the total number of shoot meristems produced on TDZ and IAA when using shoot material of Selection 3 (Coral). There were a significant number (288) shoot meristems stimulated for the best treatment of ½ MS + 10 μM TDZ (Figure 27).
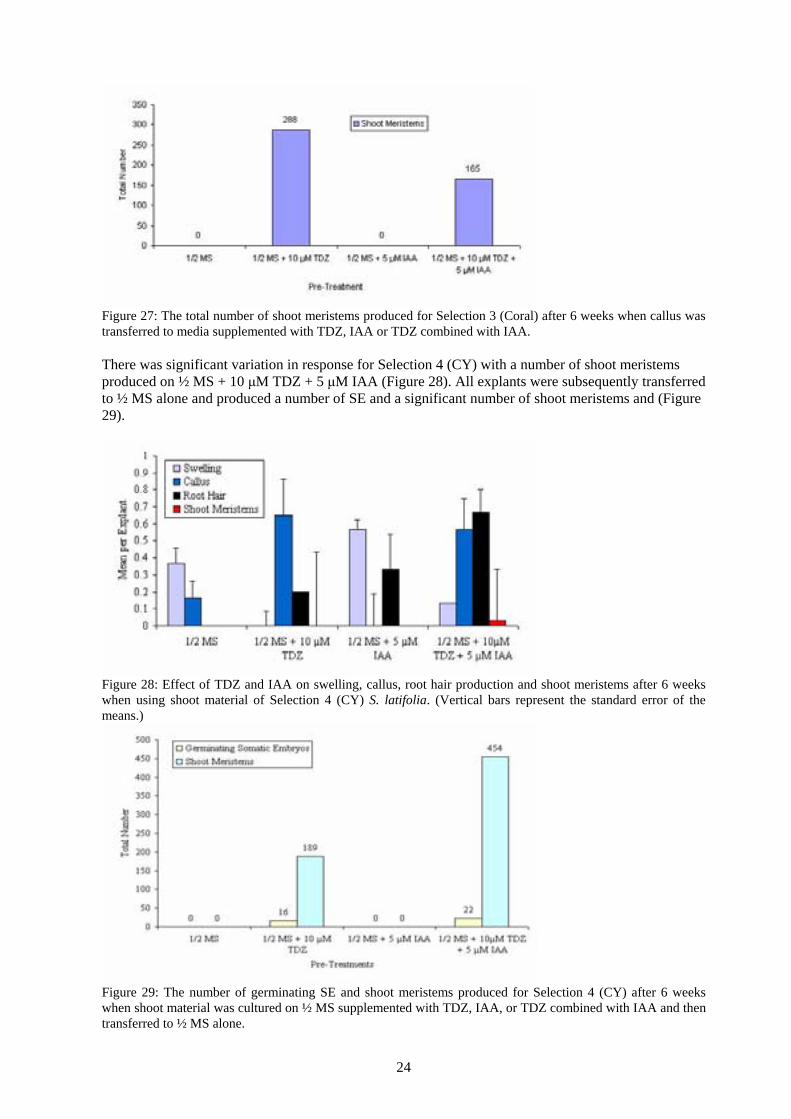
24
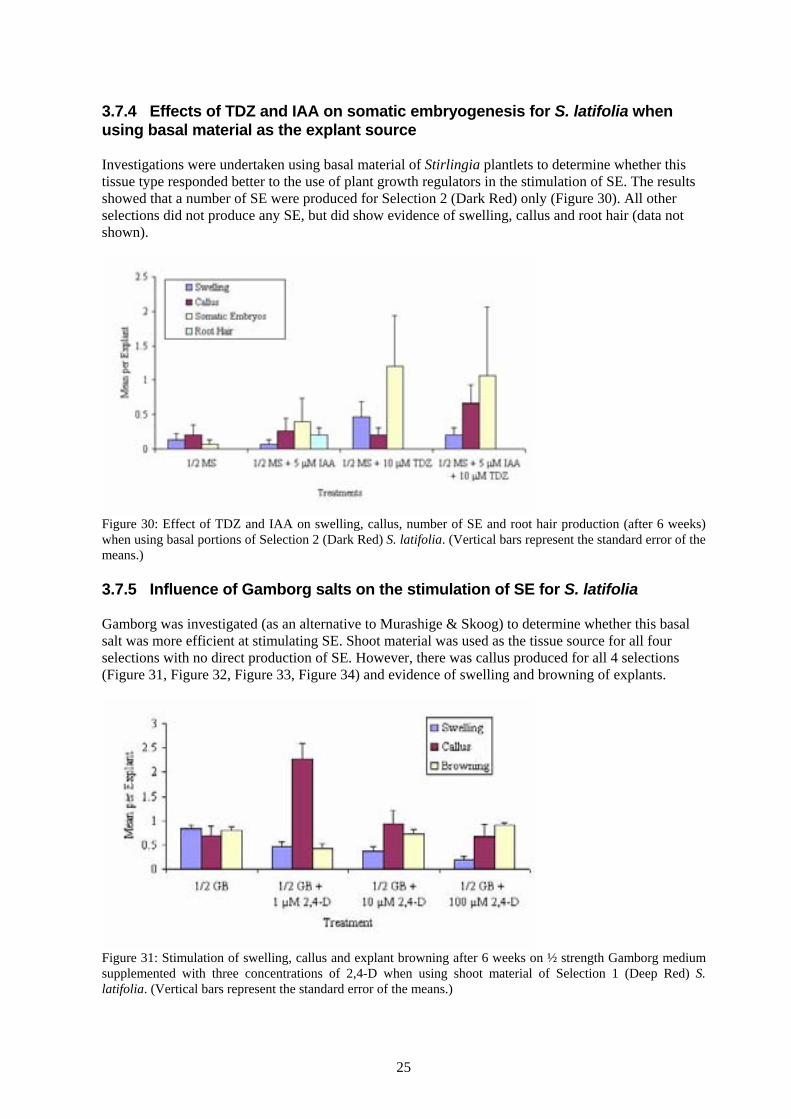
Figure 27: The total number of shoot meristems produced for Selection 3 (Coral) after 6 weeks when callus was transferred to media supplemented with TDZ, IAA or TDZ combined with IAA. There was significant variation in response for Selection 4 (CY) with a number of shoot meristems produced on ½ MS + 10 μM TDZ + 5 μM IAA (Figure 28). All explants were subsequently transferred to ½ MS alone and produced a number of SE and a significant number of shoot meristems and (Figure 29).
Figure 28: Effect of TDZ and IAA on swelling, callus, root hair production and shoot meristems after 6 weeks when using shoot material of Selection 4 (CY) S. latifolia. (Vertical bars represent the standard error of the means.)
Figure 29: The number of germinating SE and shoot meristems produced for Selection 4 (CY) after 6 weeks when shoot material was cultured on ½ MS supplemented with TDZ, IAA, or TDZ combined with IAA and then transferred to ½ MS alone.
25
3.7.4 Effects of TDZ and IAA on somatic embryogenesis for S. latifolia when using basal material as the explant source Investigations were undertaken using basal material of Stirlingia plantlets to determine whether this tissue type responded better to the use of plant growth regulators in the stimulation of SE. The results showed that a number of SE were produced for Selection 2 (Dark Red) only (Figure 30). All other selections did not produce any SE, but did show evidence of swelling, callus and root hair (data not shown).
Figure 30: Effect of TDZ and IAA on swelling, callus, number of SE and root hair production (after 6 weeks) when using basal portions of Selection 2 (Dark Red) S. latifolia. (Vertical bars represent the standard error of the means.) 3.7.5 Influence of Gamborg salts on the stimulation of SE for S. latifolia Gamborg was investigated (as an alternative to Murashige & Skoog) to determine whether this basal salt was more efficient at stimulating SE. Shoot material was used as the tissue source for all four selections with no direct production of SE. However, there was callus produced for all 4 selections (Figure 31, Figure 32, Figure 33, Figure 34) and evidence of swelling and browning of explants.
Figure 31: Stimulation of swelling, callus and explant browning after 6 weeks on ½ strength Gamborg medium supplemented with three concentrations of 2,4-D when using shoot material of Selection 1 (Deep Red) S. latifolia. (Vertical bars represent the standard error of the means.)

26
Figure 32: Effect on swelling, callus and explant browning after 6 weeks when using shoot material of Selection 2 (Dark Red) S. latifolia on ½ strength Gamborg (GB) medium supplemented with 2,4-D. (Vertical bars represent the standard error of the means.)
Figure 33: Gamborg (GB) medium supplemented with three concentrations of 2,4-D and its effect on swelling, callus and explant browning after 6 weeks when using shoot material of Selection 3 (Coral) S. latifolia. (Vertical bars represent the standard error of the means.)

27
Figure 34: Effect on swelling, callus and explant browning when using ½ strength Gamborg (GB) medium supplemented with three concentrations of 2,4-D and utilising shoot material of Selection 4 (CY) S. latifolia. (Vertical bars represent the standard error of the means.) 3.8 Effect of 2,4-D on somatic embryogenesis of L. squamatum There were significantly more SE (p<0.05) produced on ½ MS + 1 μM 2,4-D than the other treatments (Figure 35) when using excised zygotic embryos for L. squamatum. In total, there were ~ 470 SE produced on 1 μM 2,4-D, compared to a total of 292 SE for 10 μM 2,4-D.
Figure 35: Mean number of primary SE produced when utilising excised zygotic embryos of L. squamatum as the tissue source. (Vertical bars represent the standard error of the means.)

28
A B
Plate 4: Somatic embryogenesis in Lepidosperma squamatum. A: Shows a combination of white callus and somatic embryos on the same explant (bar = 1.5mm). B: A close-up of the individual, spherical somatic embryos (bar = 0.3mm).

29
3.9 Secondary somatic embryogenesis for L. squamatum Primary SE were utilised as the explant source to stimulate conversion into plantlets and secondary somatic embryogenesis. As noted in Figure 36, SE on ½ MS alone did not convert into plantlets, but continued to produce more SE as indeed did those SE transferred to 1 µM 2,4-D.
Figure 36: Mean number of secondary SE of L. squamatum produced when using primary SE as the explant source after 6 weeks incubation on ½ MS alone or supplemented with 1 μM 2,4-D. (Vertical bars represent the standard error of the means.)
Figure 37: The total number of secondary SE of L. squamatum produced after 6 weeks culture when using primary SE as the explant source. Excised zygotic embryos were used as the tissue source to promote somatic embryogenesis with significantly fewer SE produced (Figure 38) than the above experiment and only a small amount of callus production.

30
Figure 38: Mean number of SE of L. squamatum and callus production after 6 weeks when using excised zygotic embryos. (Vertical bars represent the standard error of the means.) Callus (10 explants per treatment) produced (in experiment 3.9) were subsequently transferred to a variety of media to promote conversion and/or stimulation of SE with good results. Media supplemented with 5 µM TDZ or 0.5 µM Z produced the highest number of SE with a total of 148 and 111 respectively (Figure 39). Primary somatic embryogenesis versus secondary somatic embryogenesis is detailed in Figure 40. This graph demonstrates that once embryogenic callus is produced, there is abundant production of secondary SE.
Figure 39: Total number of L. squamatum SE produced after 6 weeks incubation on various media when using callus (10 explants per treatment) as the tissue source.

31
Figure 40: Comparison of primary somatic embryogenesis (using excised zygotic embryos) versus secondary somatic embryogenesis (using callus as the tissue source) for L. squamatum. (Vertical bars represent the standard error of the means.) 3.10 Effect of ABA on desiccation tolerance of L. squamatum SE Preliminary experiments investigated the plant growth regulator ABA to determine whether its inclusion in the media would increase desiccation tolerance in primary SE. Embryos were treated with 20 µM ABA for one week prior to drying in the laminar flow for 2 hours. SE treated with ABA were significantly heavier than the controls (Figure 41) following 2 hours drying.
Figure 41: Dry weight (μg) of L. squamatum SE treated with 20 μM ABA for one week verus non-treated (-ABA) embryos. Treated embryos were significantly heavier after 2 hours drying than the controls. (Vertical bars represent the standard error of the means.) Following the desiccation process, dried embryos were transferred to ½ MS PGR-free medium to stimulate germination. Although the numbers of SE that survived were small, there was clear evidence of rehydration and more SE survived when treated with 20 μM ABA (Figure 42) compared to those dried without ABA. The controls (without drying) were significantly better.

32
Figure 42: Mean number of L. squamatum SE alive after 2 hours drying in the laminar flow. There is clear evidence that treatment with 20 μM ABA increased tolerance of SE to desiccation. (Vertical bars represent the standard error of the means.) Continuing the previous experiment, higher concentrations of ABA were investigated to determine whether an increase in the concentration would increase the desiccation tolerance of SE and therefore their survival. Primary SE of L. squamatum were cultured on ABA for one week prior to drying in the laminar flow for 2 hours. The number of SE that survived were minimal, but represent over 56 % survival which is significant (p<0.05) with ½ MS + 50 μM ABA being the best treatment (Figure 43).
Figure 43: Percentage of SE that survived 2 hours of drying in the laminar flow for L. squamatum after treatment with 3 concentrations of ABA for one week.

33
4. Discussion Somatic embryogenesis was successfully achieved for B. tetraphyllum, M. fuliginosa, S. latifolia, and L. squamatum. Conversion of SE into functioning plantlets was also successful for M. fuliginosa, S. latifolia and B. tetraphyllum with plants successfully transferred to soil (70-100% survival) without the addition of plant growth regulators and minimum acclimatisation (1-2 weeks in the glasshouse). As is the case with all species investigated, the stimulation of SE appears to be closely linked to the age of the explant and the younger the material used as the explant, the more successful the process became. Desiccation of SE for L. squamatum and B. tetraphyllum demonstrated encouraging results. A number of SE displayed germination events (after being dried for 2-3 hours) including re-hydration and the production of root hairs. These preliminary results clearly demonstrate that drying (desiccation) of SE is possible, however, further investigation is required to optimise the protocol. The estimated cost of producing a single plant under the normal micropropagation method can vary from less than $1 to over $5 depending on the species and how difficult it is to acclimate into tissue culture. In general, 15-20 plantlets can be produced in one tissue culture vessel compared to approximately 200 SE per petri dish. Somatic embryogenesis is therefore at least 10 times more efficient, even with non-optimal protocols, and, as such, the cost can conservatively be estimated at 10 to 50 cents per plant. 4.1 Somatic embryogenesis for B. tetraphyllum Primary SE were successfully induced from excised coleoptiles of B. tetraphyllum with an average of 34 SE per 2 mm coleoptile explant. In total, there was in excess of 1,500 SE produced after 6 weeks using this method (from 0.02g of coleoptile material) (Figure 2). This has the extraordinary potential to produce ~75,000 SE from 1 g of parent material. Results demonstrated that 2,4-D at low concentrations was crucial for the stimulation of primary SE and is in agreement with other studies that have shown the presence of 2,4-D is essential for somatic embryogenesis of monocotyledonous plants (Kim and Soh 1996; Turner et al. 2000). In addition, secondary somatic embryogenesis (using primary SE as the tissue source) was also successful with over 1,755 SE produced in 5 weeks (Figure 4) from 90 SE, a multiplication rate of 19.5, significantly greater than the usual in vitro multiplication rate of 3 to 4 (using shoot cultures) (Willyams 2005b). Primary SE were transferred to ½ MS PGR-free medium for conversion into plantlets with 100% efficiency and successful transfer to soil. Several desiccation experiments were undertaken with primary SE drying for 3 hours in the laminar flow. Of all the concentrations investigated, 20 μM ABA was significantly better than the other treatments (1/2 MS control, 50 or 100 μM ABA). In one experiment, over 50 % of SE treated with 20 μM ABA for one week survived 3 hours drying in the laminar flow, a significant outcome, with rehydration and the production of root hairs clearly evident. In an effort to increase desiccation tolerance, it was anticipated that inclusion of ABA in the initiation media may be beneficial, however, the inclusion of ABA at the initiation stage completely suppressed somatic embryo development. Several other experiments investigated different carbohydrate (sugar) sources that are known to increase desiccation tolerance (data not shown). Maltose, trehalose and sucrose in one experiment, and sucrose, mannitol and sorbitol were all investigated alone and in combination with each other to impart desiccation tolerance to primary SE. None of these sugars was significantly better than the usual carbohydrate source of sucrose alone and it appeared that the concentration of ABA was significantly more important in desiccation tolerance than the sugar source. Future work should investigate the use of xylitose.

34
4.2 Response of M. fuliginosa to heat treatment and 2,4-D Experiments were undertaken using a heat treatment to germinate M. fuliginosa seeds to obtain coleoptiles as this tissue type had been previously shown to be embryogenically competent in other species. Although germination rates were low (7%) they were significantly better than no heat treatment (0% germination). The coleoptiles that developed were excised and used as the tissue source for the production of primary SE for this species. Zygotic embryos were also investigated to determine whether these could be stimulated to produce SE. The inclusion of 1 and 10 μM 2,4-D in the medium inhibited germination of the embryos and did not stimulate direct somatic embryogenesis. There was, however, production of root hair and white callus, an indication that this tissue type was responsive to this particular auxin at relatively low concentrations. This experiment was repeated with varying results. Zygotic embryos were again cultured on ½ MS alone or with 1 or 10 μM 2,4-D. In this experiment, there was production of callus and direct somatic embryogenesis (Figure 13). However, in this experiment the callus produced on 10 μM 2,4-D was yellow in colour and quite different in appearance from the white callus produced in the previous experiment. The yellow callus was subsequently transferred to ½ MS alone with interesting results. There was significant production of SE with ~27 SE per callus explant (Figure 14) and a total of ~562 SE (from 21 callus explants) produced in 6 weeks. This was in contrast to the previous experiment where white callus did not produce any SE. These SE were then transferred to ½ MS for conversion into entire plantlets with 100% success and ½ MS + 1 μM 2,4-D for the stimulation of secondary SE (Figure 15). There were ~106 SE produced per primary SE and a significant total of ~ 3,200 secondary SE produced in 6 weeks. It is therefore estimated that it may be possible to stimulate up to ~26,000 SE from 1 g of parent material using this protocol. Experiments with coleoptiles as the tissue source were repeated with significant differences in the response (most likely due to the different seed batches) as described below. During the first experiment, SE were successfully stimulated on ½ MS + 1 μM 2,4-D with an average of 6 SE per coleoptile explant (a total of ~196 SE, compared to ~38 SE for ½ MS + 10 μM 2,4-D). The experiment was repeated and the results in this second experiment confirmed that 1 μM 2,4-D produced significantly more SE than 10 μM 2,4-D, but, there were less SE produced than in the previous experiment (a total of 55 SE for 1 μM 2,4-D and 17 SE for 10 μM 2,4-D). White callus produced on ½ MS + 10 μM 2,4-D and transferred to ½ MS alone did not stimulate SE, which is clear evidence that it was not embryogenic. The primary SE that were produced were subsequently transferred to a variety of media to stimulate conversion into entire plantlets or to promote secondary somatic embryogenesis (Figure 12). Successful conversion into plantlets occurred on ½ MS alone, or when the media was supplemented with 0.5 μM Z or 0.5 μM GA. In addition, there was significant production of secondary SE (~22 per explant) from primary SE that had been transferred to ½ MS + 1 μM 2,4-D. In total there was approximately 1,100 secondary SE produced in 6 weeks of culture (from 50 SE) and this equates to ~ 16,500 SE from 1 g of original plant material (in this case). Thirty clumps of M. fuliginosa SE were transferred to ½ MS PGR-free to assess conversion into entire plantlets. From these, 175 plantlets were eventually transferred to soil (Figure 16). The explanation for the significant multiplication in the number of plantlets is that SE are extremely small, and whilst all care is taken to excise one single SE for transfer to treatments, it is almost impossible, and as a result, there are a number of SE (or clumps) excised as an explant instead of one.

35
4.3 Effect of 2,4-D on somatic embryogenesis of S. latifolia S. latifolia seeds are extremely hard and it is necessary to use a vice to crack open the seed to extract the endosperm containing the embryo. Whole endosperms were cultured on ½ MS alone or supplemented with 1 or 10 μM 2,4-D to stimulate direct somatic embryogenesis. Several endosperms on the controls germinated, while the others on 2,4-D produced small amounts of callus (Figure 17). This callus was subsequently transferred to ½ MS and successfully converted into plantlets that were transferred to soil. Although the number of plantlets was low (Figure 18) compared to the above species, this is the first reported evidence of primary somatic embryogenesis for this species. Callus from the above experiment was used as the explant source for stimulation of secondary SE and cultured on ½ MS supplemented with 1 or 10 μM 2,4-D. There were a number of secondary SE produced in this experiment with ~ 11 SE per callus explant for 1 μM 2,4-D and ~ 4 SE per callus explant for 10 μM 2,4-D. Somatic embryogenesis was not as efficient in this species when using callus as the explant source with a total of 278 SE (from an average of 1.235g) for 1 μM 2,4-D and 113 SE for 10 μM 2,4-D. SE production was also much slower in this species (12 weeks). There were four selections of S. latifolia available for experimental purposes, each with a different coloured flower. There was significant variation in the response of these selections when using shoot material to treatment with 2,4-D with Selections 3 (Coral) and 4 (CY) producing more callus than the other two selections (Figure 20). However, when callus was transferred to subsequent media to induce somatic embryogenesis, the only selection to produce entire plantlets was Selection 1 (Deep Red) with a total of 449 plantlets after an extensive treatment regime including both cytokinins and auxins. As both types of PGR were utilised, it is unclear whether callus was embryogenic or whether organogenesis had occurred. Numerous experiments were undertaken to determine whether TDZ and IAA could also stimulate somatic embryogenesis in this species. Both shoot explants (Figure 22 through to Figure 29) and basal portions (Figure 30) were investigated to determine whether one tissue type was more responsive to these plant growth regulators. Selection 4 (CY) was the most responsive when using shoot material producing the highest number of shoot meristems (450) on ½ MS + 10 μM TDZ + 5 μM IAA. Selection 2 (Dark Red) did not respond at all to the plant growth regulators when they were alone, but did respond to the combined treatment with ~170 SE on the combined treatment. The variation in response is clear evidence that “selection” differences play a vital role in the success or otherwise of somatic embryogenesis for this species. In comparison, basal portions of the same selections did not respond as well as shoot explants, which was a little unexpected. It is thought that the basal portion of the plant has actively dividing cells that are more responsive to the exogenous application of plant growth regulators and it was therefore expected that somatic embryogenesis would have been more successful using this tissue type. In fact, only one selection (Selection 2, Dark Red) showed any response with a number of SE produced. Investigation of Gamborg (Figure 31 through to Figure 34) as an alternative medium was undertaken for all four selections of S. latifolia using shoot explants with no production of callus, shoot meristems or SE when using 2,4-D. These results demonstrated that Gamborg medium was not as suitable as Murashige and Skoog medium as a substrate for S. latifolia.

36
4.4 L. squamatum and its response to plant growth regulators A small number of L. squamatum seed became available and due to previous experience with poor germination, it was necessary to use extracted zygotic embryos for the initiation of SE in this species. On average there was ~ 15 SE produced per extracted embryo for ½ MS + 1 μM 2,4-D and ~ 10 SE per extracted embryo for 10 μM 2,4-D (Figure 35). In total there were ~ 470 SE (from 0.05 g) and ~ 292 SE (from 0.05 g) produced respectively for the treatments in 6 weeks. The potential therefore exists to produce ~ 9,400 SE from 1 g of plant material when using the lower concentration of 2,4-D and ~ 5,840 SE from 1 g of plant material for the higher concentration of 2,4-D. Primary SE produced above were utilised as the explant source to stimulate conversion into plantlets and secondary somatic embryogenesis. Rather than convert into plantlets on ½ MS PGR-free medium (as expected) primary SE continued to produce more SE (Figure 36). This was not totally unexpected, as the researcher had observed this phenomenon in other species. It is possible that there was a carry-over effect of the PGR causing the stimulation of SE to continue. SE that were transferred to 1 µM 2,4-D also continued to produce secondary SE. In total there was ~ 317 SE produced (on ½ MS) from 0.16g of plant material and ~ 487 SE produced (on ½ MS + 1 μM 2,4-D) from 0.2173g which potentially equates to ~ 1,981 SE and ~ 2,2123 SE for each treatment respectively from 1 g of parent material. The above experiment was repeated, with excised zygotic embryos used as the explant source to promote somatic embryogenesis with significantly fewer SE produced (Figure 38) than the first experiment and only a small amount of callus produced. However, callus produced in this experiment was subsequently transferred to a variety of media to promote conversion and/or stimulation of SE with very good results. Callus that was transferred to media supplemented with 5 µM TDZ and 0.5 µM Z produced the most number of SE with a total of 148 and 111 respectively for these treatments (Figure 39). All callus appeared to be embryogenic as even callus transferred to ½ MS alone produced a small number of SE. It is evident that somatic embryogenesis for this species is quite variable and dependent on the seed source. Primary somatic embryogenesis versus secondary somatic embryogenesis is detailed in Figure 40 and clearly demonstrates that once embryogenic callus is produced, there is a significant production of secondary SE. Preliminary investigations were undertaken using primary SE to determine whether the inclusion of ABA in the media would increase desiccation tolerance. Primary SE were treated with 20 µM ABA for one week prior to drying in the laminar flow for 2 hours and then being transferred to ½ MS media for germination. SE treated with ABA were significantly heavier than the controls (Figure 41) after 2 hours drying. As expected many SE browned quickly after desiccation and were not able to be rehydrated. However, there were a number of SE that did survive the drying process and those SE treated with 20 µM ABA were significantly more resilient than those without any ABA. Progressing on from the previous experiment, a number of different concentrations of ABA were investigated to determine whether an increase in the concentration of ABA would increase the desiccation tolerance of SE and therefore their survival. Once again, primary SE of L. squamatum were transferred to various concentrations of ABA for one week prior to drying in the laminar flow for 2 hours. Although the number of SE that survived appears minimal, it did in fact represent over 50 % survival which is significant with ½ MS + 50 μM ABA being the best treatment in this case (Figure 43), slightly better than 20 μM ABA previously. This was clear evidence that desiccation of SE is possible, but the concentration and application timing of ABA needed further refinement. The ability to desiccate SE is a significant step towards the production of synthetic seeds.

37
Conclusion Direct somatic embryogenesis from several tissue sources (depending on the species) was successful for B. tetraphyllum, M. fuliginosa, S. latifolia and L. squamatum. The study showed that it was possible to produce thousands of SE after 6 weeks of culture from a limited amount of plant material. This is a significant improvement over conventional methods, where in many cases, the multiplication rate can be as low as 3-4 per month. The estimated cost of producing a single plant under normal micropropagation methods can vary from less than $1 to over $5 depending on the species and how difficult it is to initiate into tissue culture. In general, with a particularly responsive species, it may be possible to produce 15-20 plantlets in one tissue culture vessel compared to approximately 200 SE per petri dish. Somatic embryogenesis is therefore at least 10 times more efficient and, as such, the cost can conservatively be estimated at 10 to 50 cents per plant. This is a significant saving in production costs. In addition, plantlets derived from SE were successfully established in soil with minimal acclimatisation. Desiccation of SE for two species demonstrated encouraging results with a number of SE displaying evidence of rehydration and root hair production. These preliminary results clearly demonstrate that drying (desiccation) of SE is possible, however, optimisation of this protocol requires further research. The protocol investigated has the potential to be commercially viable with an estimated 75,000 SE for B. tetraphyllum and 26,000 SE for M. fuliginosa theoretically possible from 1 g of plant material. The use of somatic embryogenesis as a technique to deliver a mass propagation method for these native plant taxa will effectively reduce production costs, reduce the loss of biodiversity, increase the efficiency of rehabilitation programmes and boost the supply of these species to the horticultural industry.

38
References: Cervelli R.; Senaratna T. Economic aspects of somatic embryogenesis. Dordrecht, The Netherlands:
Kluwer Academic Publishers; 1995:29-64. George E. F. Plant Propagation by Tissue Culture, Parts 1 & 2. Edington, Wilts: Exegetics Ltd;
1996:265-322. Hutchinson M. J.; Saxena P. K. Role of purine metabolism in thidiazuron-induced somatic
embryogenesis of geranium (Pelargonium x hortorum) hypocotyl cultures. Physiol. Plant. 98:517-522; 1996.
Kim J. W.; Soh W. Y. Plant regeneration through somatic embryogenesis from suspension cultures of Allium fistulosum L. Plant Sci. 114:215-220; 1996.
Meney K., A.; Pate J. S., eds. "Australian Rushes. Biology, Identification and Conservation of Restionaceae and Allied Families". University of Western Australia Press, Perth; 1999.
Meney K. A.; Dixon K. W. In vitro propagation of Western Australian Rushes (Restionaceae and related families) by embryo culture. Part 1. In vitro embryo growth. Plant Cell Tiss. Org. Cult. 41:107-113; 1995.
Murashige T.; Skoog F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 15:473-497; 1962.
Murthy B. N. S.; Murch S. J.; Saxena P. K. Thidiazuron-induced somatic embryogenesis in intact seedlings of peanut (Arachis hypogaea) - Endogenous growth regulator levels and significance of cotyledons. Physiol. Plant. 94:268-276; 1995.
Senaratna T. Artificial seeds. Biotechnol. Adv. 10:379-392; 1992. Senaratna T.; McKersie B. D.; Bowley S. R. Artificial seeds of alfalfa (Medicago sativa L). Induction
of desiccation tolerance in somatic embryos. In Vitro Cell. Dev. Biol. 26:85-90; 1990. Steward F. C.; Mapes M. O.; Mears C. Growth and organised development of cultured cells. II.
Organisation in cultures grown from freely suspended cells. Am. J. Bot. 45:653-704; 1958. Taiz L.; Zeiger E., eds. "Plant Physiology". The Benjamin/Cummings Publishing Company, Inc.,
Redwood City, California; 1991. Taji A.; Williams R., eds. "Tissue Culture of Australian Plants". University of New England, Armidale
NSW, Australia; 1996. Turner S. R.; Tan B.; Senaratna T.; Bunn E.; Dixon K. W.; Touchell D. H. Cryopreservation of the
Australian species Macropidia fuliginosa (Haemodoraceae) by vitrification. Cryo-Letters. 21:379-388; 2000.
Vajrabhaya M. Embryogenesis. , Cell and Tissue Culture in Field Improvement. FFTC Book Series; 1988:24-32.
Vasil K. I.; Thorpe T. A., eds. "Plant Cell & Tissue Culture". Kluwer Academic, Dordrecht, The Netherlands; 1994.
Wheeler J., ed. "Flora of the South West : Bunbury, Augusta, Denmark.". Australian Biological Resources Study, Canberra; 2002.
Willyams D. (2005a). Current rehabilitation rates return 1-2 % of the pre-mined biodiversity. Personal Communication.
Willyams D. (2005b). Typical in vitro multiplication rates using shoot cultures is approximately 3-4 in one month. Personal Communication.
Zimmerman J. L. Somatic embryogenesis - A model for early development in higher plants. Plant Cell. 5:1411-1423; 1993.


















