
Detecting DNA damage at a single-molecule level by atomic...
11
Detecting DNA damage at a single-molecule level by atomic force microscopy Yong Jiang 1 , and Piotr E. Marszalek 2 1 School of Chemistry and Chemical Engineering, Southeast University, Jiangning, Nanjing, Jiangsu, 211189, China 2 Center for Biologically Inspired Materials and Material Systems and Department of Mechanical Engineering and Materials Science, Duke University, Durham, NC 27708, USA DNA damage can be caused daily by as many as one million molecular lesions per cell by normal metabolic activities and environmental factors. The uncorrected lesions in critical genes (such as tumor suppressor genes) can inhibit a cell's ability to carry out its regular function and accordingly increase the possibility of tumor formation. Detecting DNA damage is the first step towards understanding the complex damage repair process. Hereby, we introduce single-molecule methods to detect DNA damage using atomic force microscope (AFM) imaging and single-molecule force spectroscopy (SMFS). These new techniques allow us to visualize and manipulate single DNA under nearly in vivo conditions in real time. First, the majority of most DNA damage affects the primary structure of the double helix. The lesions cause a change in DNA mechanics which can be measured easily by SMFS. Second, a variety of repair strategies have evolved to restore lost genetic information. Specific DNA repair enzymes or antibodies bind at or near the site of damage, recruiting other molecules to bind and triggering the actual repair to take place. These processes can also be visualized directly, and the lesions can be quantified at a single-molecule level by AFM imaging of these repair enzymes or antibodies. The article highlights research progress in the detection of DNA damages using AFM imaging and SMFS methods. Multiple examples are presented on visualizing and manipulating DNA at a single-molecule level. Keywords DNA damage; Single molecule; Atomic force microscopy (AFM), Force spectroscopy, Imaging 1. Introduction DNA is being damaged at a rate of 10 3 to 10 6 lesions per cell per day [1] due to inside metabolic processes and outside environmental factors. Even though these lesions only take 10 -6 of the human genome which is approximately 10 9 base pairs, uncorrected lesions in critical genes such as tumor suppressor genes can block cells and prevent them from carrying out regular functions and increase the possibility of tumor formation. In most cases of DNA damage, the bases are chemically modified by non-native chemical bonds or bulky adducts. These chemical modifications can in turn alter the spatial configuration of the structure of the DNA double helix. Such alterations can be detected by the specific DNA enzymes. Once damage is localized, specific DNA repair enzymes bind at or near the site of damage, summoning other enzymes to bind and form a complex that triggers the following repair to take place. Detecting DNA damage is the first step towards understanding the complex damage repair process. DNA damage detection is typically a multi-step process involving many enzymatic reactions, various labeling methodologies and the separation of DNA molecules [2]. Many methods and techniques have been developed to study DNA damage quantitatively in bulk material [3-8]. Among them, gel electrophoresis [3, 4] is a well-established bulk analysis method, it is frequently used in DNA damage studies because it allows separation of intact and damaged plasmids by its size or geometry. High sensitivity methods include immunoassays, which use specific antibodies in response to DNA damage [5] and HPLC coupled with mass spectrometry [5, 6]. Within the last ten or so years, single-cell gel electrophoresis, or the comet assay, was established as one of the standard methods for quantifying DNA damage [7, 8]. Since DNA alterations vary from one DNA molecule to another, it would be advantageous to detect various lesions in individual molecules. The last decade witnessed a rapid development of new single-molecule techniques which were useful in detecting DNA damage [9]. Atomic force microscopy (AFM) has been utilized for manipulating and imaging of individual DNA since 1989 [10-26]. It is one of the leading techniques in single molecule or nanoscale science because it can visualize individual DNA, proteins and DNA-protein complexes under physiological conditions in real time. However, AFM studies of DNA were mostly focused on the biological structure and dynamic processes of DNA, [10-17] and relatively limited application was found in DNA damage and repair research [18-26]. For example, Wang et al investigated the interaction between MutS and DNA in the mismatch repair system [18]. Chen et al used AFM cantilevers with carbon nanotube probes to visualize human 8-oxoguanine DNA glycosylase scanning DNA in the search for damage [19]. Murakami et al imaged supercoiled plasmids damaged by high doses of gamma radiation by AFM and compared the results with that obtained by gel electrophoresis of damaged DNA [24]. Pang et al quantified single-strand breaks (SSB) and double-strand breaks (DSB) in supercoiled DNA plasmid induced by high doses of various types of ionizing radiation [25, 26]. We recently exploited the AFM imaging of individual DNA molecules, alone as well as in complexes with a suite of DNA enzymes or antibodies, to directly quantify DNA damage caused by UV and gamma radiation [27-29]. We found that high doses of short wave ultraviolet (UVC) and medium wave ultraviolet (UVB) light not only produce Microscopy: Science, Technology, Applications and Education A. Méndez-Vilas and J. Díaz (Eds.) ©FORMATEX 2010 459 ______________________________________________
Transcript of Detecting DNA damage at a single-molecule level by atomic...

Detecting DNA damage at a single-molecule level by atomic force
microscopy
Yong Jiang1 , and Piotr E. Marszalek
2
1School of Chemistry and Chemical Engineering, Southeast University, Jiangning, Nanjing, Jiangsu, 211189, China
2Center for Biologically Inspired Materials and Material Systems and Department of Mechanical Engineering and
Materials Science, Duke University, Durham, NC 27708, USA
DNA damage can be caused daily by as many as one million molecular lesions per cell by normal metabolic activities and
environmental factors. The uncorrected lesions in critical genes (such as tumor suppressor genes) can inhibit a cell's
ability to carry out its regular function and accordingly increase the possibility of tumor formation. Detecting DNA
damage is the first step towards understanding the complex damage repair process. Hereby, we introduce single-molecule
methods to detect DNA damage using atomic force microscope (AFM) imaging and single-molecule force spectroscopy
(SMFS). These new techniques allow us to visualize and manipulate single DNA under nearly in vivo conditions in real
time. First, the majority of most DNA damage affects the primary structure of the double helix. The lesions cause a
change in DNA mechanics which can be measured easily by SMFS. Second, a variety of repair strategies have evolved to
restore lost genetic information. Specific DNA repair enzymes or antibodies bind at or near the site of damage, recruiting
other molecules to bind and triggering the actual repair to take place. These processes can also be visualized directly, and
the lesions can be quantified at a single-molecule level by AFM imaging of these repair enzymes or antibodies. The article
highlights research progress in the detection of DNA damages using AFM imaging and SMFS methods. Multiple
examples are presented on visualizing and manipulating DNA at a single-molecule level.
Keywords DNA damage; Single molecule; Atomic force microscopy (AFM), Force spectroscopy, Imaging
1. Introduction
DNA is being damaged at a rate of 103 to 10
6 lesions per cell per day [1] due to inside metabolic processes and outside
environmental factors. Even though these lesions only take 10-6
of the human genome which is approximately 109 base
pairs, uncorrected lesions in critical genes such as tumor suppressor genes can block cells and prevent them from
carrying out regular functions and increase the possibility of tumor formation. In most cases of DNA damage, the bases
are chemically modified by non-native chemical bonds or bulky adducts. These chemical modifications can in turn
alter the spatial configuration of the structure of the DNA double helix. Such alterations can be detected by the specific
DNA enzymes. Once damage is localized, specific DNA repair enzymes bind at or near the site of damage, summoning
other enzymes to bind and form a complex that triggers the following repair to take place.
Detecting DNA damage is the first step towards understanding the complex damage repair process. DNA damage
detection is typically a multi-step process involving many enzymatic reactions, various labeling methodologies and the
separation of DNA molecules [2]. Many methods and techniques have been developed to study DNA damage
quantitatively in bulk material [3-8]. Among them, gel electrophoresis [3, 4] is a well-established bulk analysis method,
it is frequently used in DNA damage studies because it allows separation of intact and damaged plasmids by its size or
geometry. High sensitivity methods include immunoassays, which use specific antibodies in response to DNA damage
[5] and HPLC coupled with mass spectrometry [5, 6]. Within the last ten or so years, single-cell gel electrophoresis, or
the comet assay, was established as one of the standard methods for quantifying DNA damage [7, 8].
Since DNA alterations vary from one DNA molecule to another, it would be advantageous to detect various lesions
in individual molecules. The last decade witnessed a rapid development of new single-molecule techniques which
were useful in detecting DNA damage [9]. Atomic force microscopy (AFM) has been utilized for manipulating and
imaging of individual DNA since 1989 [10-26]. It is one of the leading techniques in single molecule or nanoscale
science because it can visualize individual DNA, proteins and DNA-protein complexes under physiological conditions
in real time. However, AFM studies of DNA were mostly focused on the biological structure and dynamic processes of
DNA, [10-17] and relatively limited application was found in DNA damage and repair research [18-26]. For example,
Wang et al investigated the interaction between MutS and DNA in the mismatch repair system [18]. Chen et al used
AFM cantilevers with carbon nanotube probes to visualize human 8-oxoguanine DNA glycosylase scanning DNA in the
search for damage [19]. Murakami et al imaged supercoiled plasmids damaged by high doses of gamma radiation by
AFM and compared the results with that obtained by gel electrophoresis of damaged DNA [24]. Pang et al quantified
single-strand breaks (SSB) and double-strand breaks (DSB) in supercoiled DNA plasmid induced by high doses of
various types of ionizing radiation [25, 26].
We recently exploited the AFM imaging of individual DNA molecules, alone as well as in complexes with a suite of
DNA enzymes or antibodies, to directly quantify DNA damage caused by UV and gamma radiation [27-29]. We found
that high doses of short wave ultraviolet (UVC) and medium wave ultraviolet (UVB) light not only produce
Microscopy: Science, Technology, Applications and Education A. Méndez-Vilas and J. Díaz (Eds.)
©FORMATEX 2010 459
______________________________________________

cyclobutane pyrimidine dimers (CPDs), but also cause significant DNA degradation. At very low UVB doses, the
relationship between the number of CPDs and UVB dose is almost linear [27]. We also examined long-wave
ultraviolet (UVA) damage to DNA at a single-molecule level and found that UVA (365 nm) produces a significant
amount of abasic sites and CPDs. Our results strongly suggest that CPDs can be produced directly by UVA [28].
Besides, the result of the detection and quantification of gamma radiation damage show that the number of SSB per
DNA molecule is linearly proportional to the plasmid size and inversely correlated to the DNA concentration [29]. All
these results show that AFM imaging is a direct and quantitative approach to detect DNA damage. Its accuracy and
reliability are validated through the comparison with traditional agarose gel electrophoresis.
A clear relationship between DNA damage and the nano mechanics of DNA double helixes by AFM single-molecule
force spectroscopy (SMFS) was also established [30]. The results show that the width of the B-S transition shortens in
a UV dose-dependent manner below the standard 70% of the contour length and becomes less cooperative. In λ-phage
DNA the second, high-force transition is also reduced in a dose-dependent manner. Thus, SMFS can also be used to
detect and investigate DNA lesions. Taken together these observations lay a foundation for the future use of SMFS in
nanoscale DNA damage diagnostics.
In the article we highlight these new assays for the detection of DNA damage at a single-molecule level using AFM
imaging and SMFS. We also describe in detail how different kinds of DNA damage were detected by AFM imaging
and SMFS. Multiple examples are presented on visualizing and manipulating DNA at a single-molecule level.
2. Experimental Section
2.1 Materials
pUC18, 2686 base pairs (bp), was isolated from E. coli and purified using the QIAFILTER plasmid maxi kit (QIAGEN
Inc). λ-phage DNA (D3779) was purchased from Sigma–Aldrich Co. DNA endonucleases that might be used for UV
damage detection were summarized in Table 1. E. coli Photolyase was bought from TREVIGEN.
Table 1 DNA endonucleases for damage detection.
Enzyme or Antibody
for damage detection Supplier
Specific lesions these enzymes
can detected
Enzyme’s
activity
E. coli endonuclease
IV
New England
Biolabs
apurinic/apyrimidinic site base
paired with adenine 100%
5,6-dihydrothymine <10%
T4 endonuclease V
New England
Biolabs and
Epicentre
cyclobutane pyrimidine dimer 100%
apurinic/apyrimidinic site base
paired with adenine 100%
E. coli endonuclease
III Trevigen
thymine glycol 100%
apurinic/apyrimidinic site base
paired with adenine 100%
5,6-dihydrothymine <10%
E. coli Fpg
Formamidopyrimidine
DNA Glycosylase
Trevigen
8 oxoguanine base paired with
a cytosine or guanine 100%
apurinic/apyrimidinic site base
paired with adenine <10%
5,6-dihydrothymine <10%
2,6-diamino-4-hydroxy-5-N-
methylformamidopyrimidine
2.2 Enzyme and antibody treatment
For DNA endonuclease treatment, 10 µg/ml UV irradiated supercoiled pUC18 plasmids were incubated with different
kinds of enzymes as shown in Table 1 to a total volume of 40 µl at 37 °C for 30-60 minutes in their preferred reaction
buffer as suggested by the enzyme provider. After specific enzyme incubation, the complex was diluted by DNA
imaging buffer containing 10 mM Tris HCl, 1 mM EDTA, and 100 mM NaCl to the final DNA concentration of 0.5-1
µg/ml and then deposited on mica for AFM imaging. For photolyase treatment, 10 µg/ml DNA was incubated with 2
µg/ml photolyase in 1x REC Buffer 14 containing 20 mM Tris-HCl pH=7.8, 1 mM EDTA, 1 mM DTT, and 50 mM
NaCl at a total volume of 20 µl. The incubation was performed for 30 min at room temperature in darkness to prevent
photolyase from repairing damage and disassociating from damage sites. The solution was then diluted with the
addition of 10 mM Tris HCl, 1 mM EDTA, and 5-10 mM MgCl2 buffer to a final DNA concentration of 0.5-1 µg/ml.
The sample was then deposited on mica immediately for AFM imaging.
Microscopy: Science, Technology, Applications and Education A. Méndez-Vilas and J. Díaz (Eds.)
460 ©FORMATEX 2010
______________________________________________

2.3 AFM imaging
For experiments in which the imaging buffer contains Mg2+
, a freshly cleaved mica was used as DNA substrate directly,
while the imaging buffer did not have Mg2+
, 1-(3-Aminopropyl)-silatrane-functionalyzed mica (APS-mica) was used for
the binding of DNA molecules. APS-mica was prepared as described by Shlyakhtenko et al [31]. A drop of 10-50 µl of
DNA solution (0.5-1 µg/ml) was deposited on the APS-mica surface at room temperature for 3 min. The sample was
rinsed and air-dried before imaging. AFM images were taken using a Nanoscope IIIa MultiMode Scanning Probe
Microscope (Veeco Instruments Inc) using Tapping mode with an E scanner. RTESP probes (Veeco) were used for
imaging in air. The spring constant of AFM cantilevers was 20-80 N/m and their resonance frequency was 275-316
kHz. NP-S probes (Cantilever C) were used for imaging in solution. The spring constant of the cantilever was 0.32-
0.64 N/m and their resonance frequency was 56-75 kHz. The All images were collected at a scan rate of 2-3 Hz, a scan
resolution of 512×512 pixels, and scan sizes of 1-5 µm.
2.4 Quantifying and verification of DNA damage from AFM images
The number of lesions was quantified by a Poisson distribution f(n; λ)= λn×exp(-λ)/n! [32, 33], based on percentages of
supercoiled and relaxed DNA. The average number of lesions per molecule, λ, can be obtained from λ=-ln[f(λ; 0)],
where f(λ; 0) is the fraction of supercoiled (damage n=0) plasmids. Using this method we can directly detect and
quantify damage to supercoiled DNA, by visualizing and counting individual intact and damaged plasmids captured by
AFM imaging.
2.5 AFM-based SMFS
For SMFS measurements, we used nonspecific adsorption of DNA to fresh gold substrates [34] and untreated silicon
nitride AFM probes (MLCT from Veeco). The cantilevers had a nominal spring constant of 10 mN/m and an actual
spring constant between 10-20 mN/m, as determined by using the energy equipartition theorem [35]. DNA pulling
experiments were carried out on λ-phage DNA after exposing the molecules to various doses of UV radiation. 100 ml
of a DNA solution (100 mg/ml) suspended in PBS/Tris EDTA buffer was deposited onto a gold substrate, incubated for
2–3 h, and then gently rinsed 3-5 times with the buffer solution.
3. Detect DNA damage by AFM Imaging
3.1 Detect DNA SSB and DSB by AFM imaging
Supercoiled DNA is a good model for studying DNA damage because it is highly sensitive to structural modifications
such as SSB and DSB [36, 37]. One SSB will relax its superhelical structure to a circular form. Similarly, one DSB
will open the relaxed circular plasmid to a linear form. More DSBs will fragment the plasmid to linear duplexes of
shorter lengths as compared to the original length. All these topological or length variations of DNA are easy to resolve
by visualizing individual intact and damaged plasmids in AFM [20, 21, 24-26].
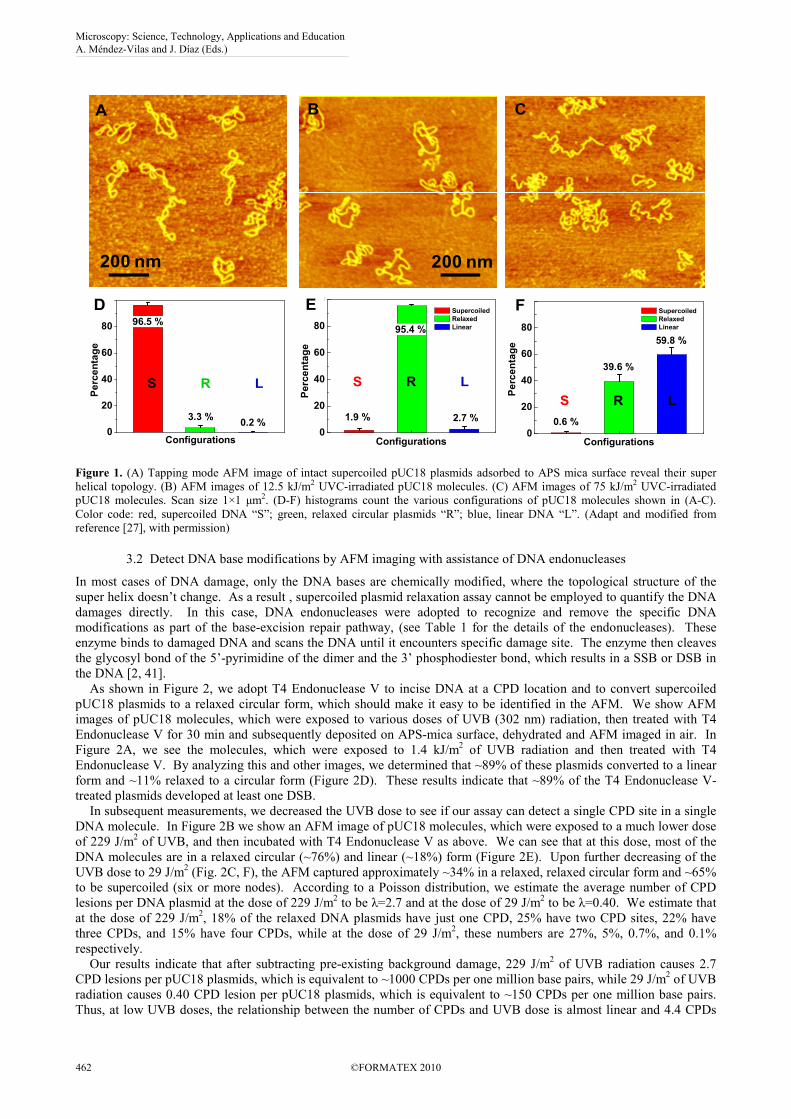
Shown in Figure 1A, is a typical AFM image of pUC18 plasmid deposited on the APS-mica from a buffer solution.
Neither the binding to the mica nor the AFM imaging process significantly affect the DNA structure which remains in
the plectonemic supercoiled configuration. We followed the approaches which were developed earlier for electron
microscopy and AFM imaging of supercoiled plasmids [36, 38, 39], and counted the number of nodes in each DNA
molecule as the number of visible crossover points in AFM images [38]. For a given plasmid size, the number of
supercoiled nodes typically vary somewhat among different molecules, and also strongly depends on the ionic
conditions in the buffer and deposition conditions [40]. In the case of pUC18 relaxation assay, we used the same buffer
with constant NaCl concentration of 100 mM in all experiments. For intact pUC18 we found the average number of
nodes is eight, and this is consistent with expected value for a supercoiled pUC18 DNA [16, 36-38]. We considered the
molecules with a node number greater than five as intact, whereas molecules with the node number equal to or less than
five indicated relaxed DNA. By this criterion, our pUC18 DNA samples contain more than 95±1% (mean±strandard
deviation) of intact supercoiled DNA and this high percentage of intact DNA is important for a reliable quantification of
DNA damage as shown in Figure 1D. Less than 5±1% of the plasmids show less than six nodes. We assume that these
plasmids represent a background of damaged DNA in our sample of “intact” pUC18 after DNA purification processes.
Figure 1B and 1C show AFM images of pUC18 plasmids, which were exposed to UVC (254 nm). At the dose of 12.5
kJ/m2, almost all supercoiled molecules relaxed, as shown in figure 1B and 1E, with 95±1% of molecules having at
least one SSB (Figure 1E). At the dose of 75 kJ/m2, 60±6% of the plasmids became linear, indicating that these
molecules developed DSBs (Figure 1C and 1F). We conclude that AFM imaging can capture even single SSB lesion in
single DNA molecules.
Microscopy: Science, Technology, Applications and Education A. Méndez-Vilas and J. Díaz (Eds.)
©FORMATEX 2010 461
______________________________________________
Percentage0
20
40
60
80
100
Percentage
Configurations
96.5 %
3.3 %0.2 %
percentage0
20
40
60
80
100
Supercoiled
Relaxed
Linear
Percentage
Configurations
59.8 %
39.6 %
0.6 %
percentage0
20
40
60
80
100
Supercoiled
Relaxed
Linear
Percentage
1.9 %
95.4 %
2.7 %
Configurations
CB
FE
S R L
200 nm
A
D
200 nm
S R L
S R L
Figure 1. (A) Tapping mode AFM image of intact supercoiled pUC18 plasmids adsorbed to APS mica surface reveal their super
helical topology. (B) AFM images of 12.5 kJ/m2 UVC-irradiated pUC18 molecules. (C) AFM images of 75 kJ/m2 UVC-irradiated
pUC18 molecules. Scan size 1×1 µm2. (D-F) histograms count the various configurations of pUC18 molecules shown in (A-C).
Color code: red, supercoiled DNA “S”; green, relaxed circular plasmids “R”; blue, linear DNA “L”. (Adapt and modified from
reference [27], with permission)
3.2 Detect DNA base modifications by AFM imaging with assistance of DNA endonucleases
In most cases of DNA damage, only the DNA bases are chemically modified, where the topological structure of the
super helix doesn’t change. As a result , supercoiled plasmid relaxation assay cannot be employed to quantify the DNA
damages directly. In this case, DNA endonucleases were adopted to recognize and remove the specific DNA
modifications as part of the base-excision repair pathway, (see Table 1 for the details of the endonucleases). These
enzyme binds to damaged DNA and scans the DNA until it encounters specific damage site. The enzyme then cleaves
the glycosyl bond of the 5’-pyrimidine of the dimer and the 3’ phosphodiester bond, which results in a SSB or DSB in
the DNA [2, 41].
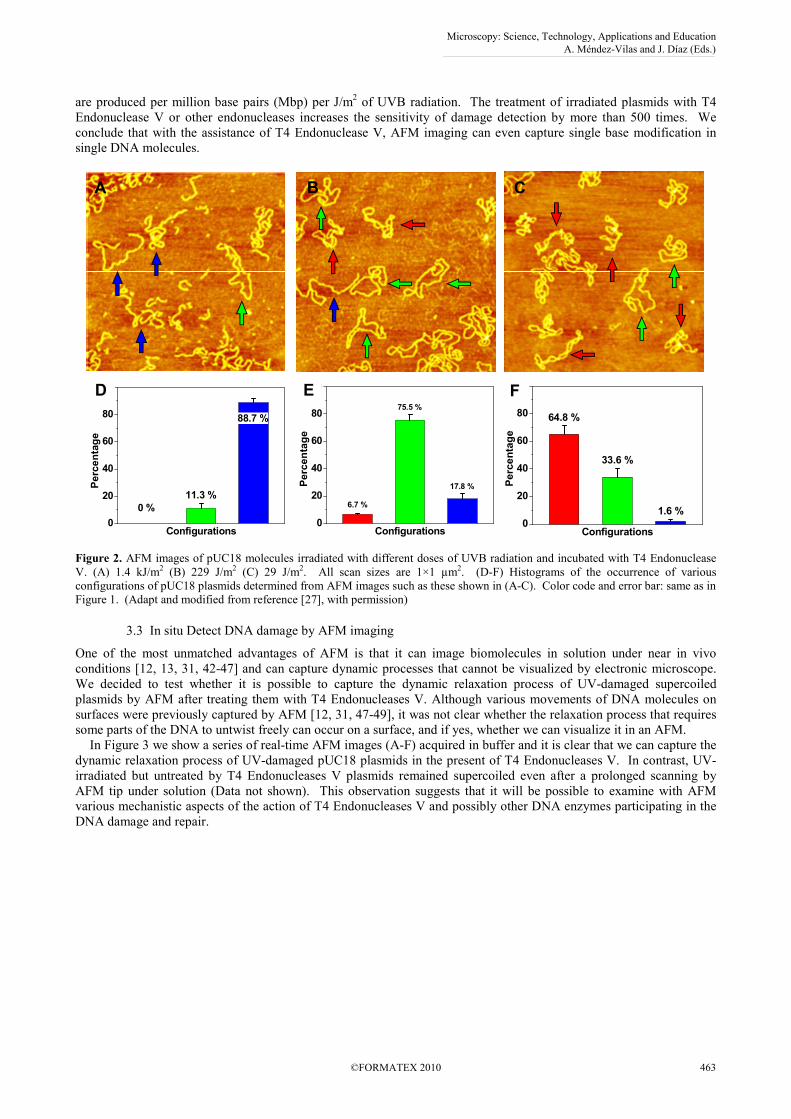
As shown in Figure 2, we adopt T4 Endonuclease V to incise DNA at a CPD location and to convert supercoiled
pUC18 plasmids to a relaxed circular form, which should make it easy to be identified in the AFM. We show AFM
images of pUC18 molecules, which were exposed to various doses of UVB (302 nm) radiation, then treated with T4
Endonuclease V for 30 min and subsequently deposited on APS-mica surface, dehydrated and AFM imaged in air. In
Figure 2A, we see the molecules, which were exposed to 1.4 kJ/m2 of UVB radiation and then treated with T4
Endonuclease V. By analyzing this and other images, we determined that ~89% of these plasmids converted to a linear
form and ~11% relaxed to a circular form (Figure 2D). These results indicate that ~89% of the T4 Endonuclease V-
treated plasmids developed at least one DSB.
In subsequent measurements, we decreased the UVB dose to see if our assay can detect a single CPD site in a single
DNA molecule. In Figure 2B we show an AFM image of pUC18 molecules, which were exposed to a much lower dose
of 229 J/m2 of UVB, and then incubated with T4 Endonuclease V as above. We can see that at this dose, most of the
DNA molecules are in a relaxed circular (~76%) and linear (~18%) form (Figure 2E). Upon further decreasing of the
UVB dose to 29 J/m2 (Fig. 2C, F), the AFM captured approximately ~34% in a relaxed, relaxed circular form and ~65%
to be supercoiled (six or more nodes). According to a Poisson distribution, we estimate the average number of CPD
lesions per DNA plasmid at the dose of 229 J/m2 to be λ=2.7 and at the dose of 29 J/m
2 to be λ=0.40. We estimate that
at the dose of 229 J/m2, 18% of the relaxed DNA plasmids have just one CPD, 25% have two CPD sites, 22% have
three CPDs, and 15% have four CPDs, while at the dose of 29 J/m2, these numbers are 27%, 5%, 0.7%, and 0.1%
respectively.
Our results indicate that after subtracting pre-existing background damage, 229 J/m2 of UVB radiation causes 2.7
CPD lesions per pUC18 plasmids, which is equivalent to ~1000 CPDs per one million base pairs, while 29 J/m2 of UVB
radiation causes 0.40 CPD lesion per pUC18 plasmids, which is equivalent to ~150 CPDs per one million base pairs.
Thus, at low UVB doses, the relationship between the number of CPDs and UVB dose is almost linear and 4.4 CPDs
Microscopy: Science, Technology, Applications and Education A. Méndez-Vilas and J. Díaz (Eds.)
462 ©FORMATEX 2010
______________________________________________
are produced per million base pairs (Mbp) per J/m2 of UVB radiation. The treatment of irradiated plasmids with T4
Endonuclease V or other endonucleases increases the sensitivity of damage detection by more than 500 times. We
conclude that with the assistance of T4 Endonuclease V, AFM imaging can even capture single base modification in
single DNA molecules.
Percentage0
20
40
60
80
100
Percentage
Configurations
88.7 %
11.3 %
0 %
Percentage0
20
40
60
80
100
Percentage
Configurations
64.8 %
33.6 %
1.6 %
C
F
Percentage0
20
40
60
80
100
Percentage
Configurations
75.5 %
6.7 %
17.8 %
BA
ED
Figure 2. AFM images of pUC18 molecules irradiated with different doses of UVB radiation and incubated with T4 Endonuclease
V. (A) 1.4 kJ/m2 (B) 229 J/m2 (C) 29 J/m2. All scan sizes are 1×1 µm2. (D-F) Histograms of the occurrence of various
configurations of pUC18 plasmids determined from AFM images such as these shown in (A-C). Color code and error bar: same as in
Figure 1. (Adapt and modified from reference [27], with permission)
3.3 In situ Detect DNA damage by AFM imaging
One of the most unmatched advantages of AFM is that it can image biomolecules in solution under near in vivo
conditions [12, 13, 31, 42-47] and can capture dynamic processes that cannot be visualized by electronic microscope.
We decided to test whether it is possible to capture the dynamic relaxation process of UV-damaged supercoiled
plasmids by AFM after treating them with T4 Endonucleases V. Although various movements of DNA molecules on
surfaces were previously captured by AFM [12, 31, 47-49], it was not clear whether the relaxation process that requires
some parts of the DNA to untwist freely can occur on a surface, and if yes, whether we can visualize it in an AFM.
In Figure 3 we show a series of real-time AFM images (A-F) acquired in buffer and it is clear that we can capture the
dynamic relaxation process of UV-damaged pUC18 plasmids in the present of T4 Endonucleases V. In contrast, UV-
irradiated but untreated by T4 Endonucleases V plasmids remained supercoiled even after a prolonged scanning by
AFM tip under solution (Data not shown). This observation suggests that it will be possible to examine with AFM
various mechanistic aspects of the action of T4 Endonucleases V and possibly other DNA enzymes participating in the
DNA damage and repair.
Microscopy: Science, Technology, Applications and Education A. Méndez-Vilas and J. Díaz (Eds.)
©FORMATEX 2010 463
______________________________________________

A B C
D E F
A B C
D E F
Figure 3. (A). Real-time AFM imaging in solution captures the relaxation of UV-damaged pUC18 plasmids by T4 Endonuclease V.
pUC18 plasmids were irradiated at 1.4 KJ/m2 of UVB and mixed on ice with T4 Endonuclease V. The sample was transferred to the
AFM instrument and imaged immediately in solution, at room temperature. The images (A-F) were captured at 27, 32, 41, 59, 63,
and 68 minutes after the AFM tip had been engaged and the first image had been acquired.
3.4 Sensitivity, accuracy and advantages of AFM assay
Based on this supercoiled plasmid relaxation assay, we can detect single lesion (SSB, DSB or base modifications) per
DNA molecule. Since one SSB is able to relax a whole supercoiled plasmid, increasing the size of the plasmid should
also increase the sensitivity of the damage detection. To test this hypothesis we exposed pNEBR-R1 supercoiled
plasmids (10,338 bp) with 29 J/m2 of UVB radiation and treated them with T4 Endonuclease V. After imaging these
plasmids in the AFM, we find that 55±3% of pNEBR-R1 plasmids are in the relaxed circular form (at least one
pyrimidine dimer) and 14±3%, are already in the linear form (at least 2 damage sites in close proximity on the opposite
strands). From the percentage of supercoiled molecules, we estimate that pNEBR-R1 plasmids developed at least
1.12±0.035 CPD/plasmid (λ=1.12), i.e. 108±13 CPD/Mbp. Thus, increasing the size of the supercoiled plasmid by 3.8-
fold resulted in a >2.8-fold (2.8=1.12/0.40) increase in damage detection sensitivity, which is consistent with our
predictions. Table 2 Comparison of DNA detection sensitivity by gel electrophoresis and by
AFM
Gel Electrophoresis AFM
Minimum amount
of DNA ~ 400 pg per lane [50] ~ 1 pg (total)
Sensitivity ~ 25 pg per band [50], (equivalent to 8.5×106
pUC18 molecules)
Single
molecules (Adapt from reference [27], with permission)
To test the accuracy of the AFM assay, we compared our results with those obtained by well-established methods
such as gel electrophoresis assay. The quantitative results by AFM imaging and gel electrophoresis on same sample are
consistent, which indicates that AFM is an accurate tool to quantify DNA damages. It is also interesting to compare our
results with the results obtained by traditional methods by other research group. We can estimate the number of
CPDs/Mbp obtained by other researchers by extrapolating their results to our dose of 229 J/m2 of UVB. For example,
Setlow and Carrier [51], who extracted radio-labeled DNA from H. influenzae and irradiated it at 280 nm and 4 kJ/m2,
estimated the number of CPDs/100bp to be 2.06. Assuming that the number of CPDs scales linearly also between 229
Microscopy: Science, Technology, Applications and Education A. Méndez-Vilas and J. Díaz (Eds.)
464 ©FORMATEX 2010
______________________________________________
J/m2 and 4000 J/m
2, we find that the Setlow and Carrier’s data correspond to 1180 CPDs/Mbp at 229 J/m
2, which is
similar to our own result (~1000 CPDs/Mbp).
Over the last ten years or so AFM has been making steady progress in demonstrating its unique advantages in
nanoscale biology research. In our opinion the two unique features of AFM that make it particularly suitable for
examining DNA damage and repair are: a) the ability to examine individual DNA molecules and DNA protein
complexes under nearly in vivo conditions, b) extremely small amounts of DNA and protein material needed for the
observation. To test this second feature we coated 0.1 µl of a pUC18 plasmid solution with the DNA concentration of
10 pg/µl on the mica surface and imaged the surface at a big scan size of 5×5 µm2. We can still collect enough DNA
molecules for DNA damage quantification. Thus, our measurements can be performed on the amount of total DNA of
1pg, which is ~400 times less than what is presently needed for the most sensitive gel electrophoresis assays [50] and
what amounts to ~1/5th of the DNA in a single cell. It may be advantageous to combine the power of gel
electrophoresis with AFM imaging together in some studies under extreme conditions. Gel electrophoresis would
separate the damaged DNA into descrete bands and the DNA extracted from a particular band would provide enough
material to be further examined by high resolution AFM imaging.
3.5 Detect DNA damage by AFM imaging using repair enzyme or antibodies binding assay
0 1 2 3 4 5 60
10
20
30
40
50
Percentage (%)
0.7 photolyases/DNA
Photolyases/DNA0 1 2 3 4 5 6
0
10
20
30
Percentage (%)
Photolyases/DNA
Average: 2.5 Photolyases/DNA
BA
DC
200 nm
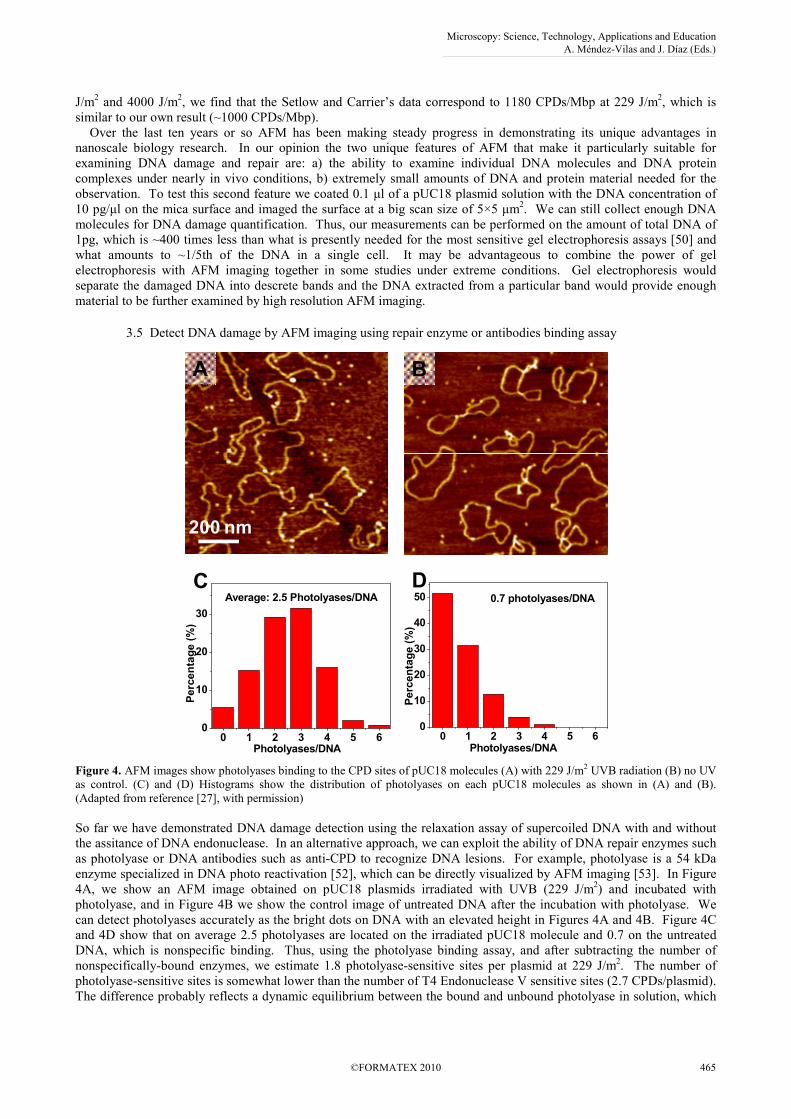
Figure 4. AFM images show photolyases binding to the CPD sites of pUC18 molecules (A) with 229 J/m2 UVB radiation (B) no UV
as control. (C) and (D) Histograms show the distribution of photolyases on each pUC18 molecules as shown in (A) and (B).
(Adapted from reference [27], with permission)
So far we have demonstrated DNA damage detection using the relaxation assay of supercoiled DNA with and without
the assitance of DNA endonuclease. In an alternative approach, we can exploit the ability of DNA repair enzymes such
as photolyase or DNA antibodies such as anti-CPD to recognize DNA lesions. For example, photolyase is a 54 kDa
enzyme specialized in DNA photo reactivation [52], which can be directly visualized by AFM imaging [53]. In Figure
4A, we show an AFM image obtained on pUC18 plasmids irradiated with UVB (229 J/m2) and incubated with
photolyase, and in Figure 4B we show the control image of untreated DNA after the incubation with photolyase. We
can detect photolyases accurately as the bright dots on DNA with an elevated height in Figures 4A and 4B. Figure 4C
and 4D show that on average 2.5 photolyases are located on the irradiated pUC18 molecule and 0.7 on the untreated
DNA, which is nonspecific binding. Thus, using the photolyase binding assay, and after subtracting the number of
nonspecifically-bound enzymes, we estimate 1.8 photolyase-sensitive sites per plasmid at 229 J/m2. The number of
photolyase-sensitive sites is somewhat lower than the number of T4 Endonuclease V sensitive sites (2.7 CPDs/plasmid).
The difference probably reflects a dynamic equilibrium between the bound and unbound photolyase in solution, which
Microscopy: Science, Technology, Applications and Education A. Méndez-Vilas and J. Díaz (Eds.)
©FORMATEX 2010 465
______________________________________________

leaves some CPD sites on DNA unoccupied. In addition, while T4 Endonuclease V makes an incision at all CPD sites,
photolyase that we used detects a subset of all CPDs, namely the cis-syn CPD [2].
4. Detect DNA damage by AFM-based SMFS
Besides AFM imaging, SMFS can also be used to detect DNA damage. Single DNA molecule was picked up by the
AFM tip and stretched vertically in solution to determine it force-extension relationship. Figure 5 shows the result of
six independent SMFS measurements on λ phage DNA after different doses of UV radiation (from 0 to 52.5 kJ/m2).
The red trace in Figure 5 shows the typical force-extension curve of intact double stranded DNA (no UV radiation).
The elastic properties of DNA have two unique features [34, 54, 55]. At a critical force of about 60-90 pN, a long
plateau amounting to 70% of the original contour length appears in the force-extension relationship; a second plateau
appears at higher forces of 150-450 pN; relaxing traces reveal patterns which are characterized by different degree of
hysteresis.
UV-irradiation time
0 min 15 min30 min45 min60 min75 min0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.6 0.8 1.0 1.2 1.4 1.6 1.8 2.0
0
200
400
600
800
Normalized Extension
Force / pN
0 kJ/m2
42.0 kJ/m2
52.5 kJ/m2
10.5 kJ/m2
21.0 kJ/m2
31.5 kJ/m2
UV
Figure 5. Force spectrograms of UV-irradiated λ-phage DNA with different doses: 0 kJ/m2, red line; 10.5 kJ/m2, green line with
square; 21 kJ/m2, blue line with diamond; 31.5 kJ/m2, orange line with hexagon; 42 kJ/m2, purple line with circle; and 52.5 kJ/m2,
black line with triangle. The traces represent measurements on different molecules. Inset: Schematic map of λ-phage DNA forming
pyrimidine dimers after UV radiation. (Adapted and modified from reference [30], with permission)
The force spectrograms of UV-treated individual DNA molecules in Figure 5 show significant differences in the
elasticity as compared to the untreated DNA (red line). The B-S transition was shortened by UV treatment below 70%
of the original contour length measured for the untreated DNA. Moreover, the shortening of the plateau is dose-
dependent. Interestingly, the second transition is also decreased in the dose-dependent manner. The shortening of the
B-S transition originates from the extensive formation of CPDs and 6-4 lesions, which results in a partial loss of the
double helical structure along the DNA backbone. cis-syn CPDs are known to induce bending of 20-30° and unwinding
of regular B-DNA by 10° [56, 57] and 6-4 lesions cause much more severe bends or kinks of 44° [58]. These bending
and unwinding caused by DNA damages weakened the base stacking and base pairing structure and DNA double helix
was partially melted [59]. This effect could be responsible for the observed shortening of the overstretching transition
in the UV-irradiated DNA. In addition, the normalized initial contour lengths before the onset of the B-S transition
increase in a dose dependent manner, which supports our hypothesis of UV-induced unwinding of the double helix.
Similarly, the same effects could also cause a shortening of the width of the second transition. The B-S transition of
UV irradiated DNA at the dose of 10.5 kJ/m2 (green line with square trace) is ~ 9% shorter compared to that of intact
DNA. Douki et al. reported that the number of CPD is 30/10,000 bases at the dose of 10.5 kJ/m2 [60]. It means ~0.3 %
of all nucleotides form CPDs. As exposure time is increased, B-S transition length shortens, suggesting an increase in
the number of pyrimidine dimers. Strikingly, the shortening of the width of the B-S transition saturates at the dose of
~40 kJ/m2 (purple line with circle trace) which is exactly the same dose at which Douki et al. observed the saturation of
Microscopy: Science, Technology, Applications and Education A. Méndez-Vilas and J. Díaz (Eds.)
466 ©FORMATEX 2010
______________________________________________

the number of UV-induced lesions. This coincidence proves that the shortening of the B-S transition is indeed caused
by the formation of CPD lesions.
We established the relationships between UV damage and the mechanics of DNA double helixes at a single-molecule
level. The width of the B-S transition shortens in a dose-dependent manner below the standard 70% of the contour
length and becomes less cooperative. The results show that SMFS, which provided many insights into the mechanics of
intact DNA, can also be used to study DNA lesions. This research is the first direct measurement that determines the
relationship between DNA nanomechanics and damage, and lays a foundation for the future use of SMFS in DNA
damage and repair.
5. Summary
AFM, as one of the leading techniques in single molecule or nanoscale science, can be used to detect DNA damage
quantitatively and accurately. Different from the traditional techniques, this damage detection method based on AFM
imaging is performed at a single-molecule level with higher sensitivity and consumes less DNA materials than bulk
methods. Besides, SMFS can also be used to investigate DNA damages by measuring the DNA nanomechanic change
caused by damage, which provide unique method in DNA diagnostics.
Acknowledgements We thank Filip Marszalek for his help with editing the manuscript. This work was supported by grants from
the National Science Foundation (MCB-0450835) and National Institutes of Health (1R21-GM071197) to P.E.M and by the start-up
fund from Southeast University to Y.J.
References
[1] Lodish H, Berk A, Matsudaira P, Kaiser CA, Krieger M, Scott MP, Zipursky SL, Darnell J. Molecular Biology of the Cell. 5th
ed. New York, NY: WH Freeman; 2004.
[2] Friedberg EC, Walker GC, Siede W. DNA Repair and Mutagenesis. 1995; ASM Press, Washington, D.C.
[3] Sutherland BM, Bennett PV, Sidorkina O, Laval J. Clustered DNA damages induced in isolated DNA and in human cells by
low doses of ionizing radiation. PNAS. 2000; 97(1): 103-108.
[4] Sutherland BM, Georgakilas AG, Bennett PV, Laval J, Sutherland JC. Quantifying clustered DNA damage induction and
repair by gel electrophoresis, electronic imaging and number average length analysis. Mutation Research/Fundamental and
Molecular Mechanisms of Mutagenesis. 2003; 531(1-2): 93-107.
[5] Besaratinia A, Synold TW, Chen HH, Chang C, Xi BX, Riggs AD, Pfeifer GP. DNA lesions induced by UV A1 and B
radiation in human cells: Comparative analyses in the overall genome and in the p53 tumor suppressor gene. Proceedings of the
National Academy of Sciences of the United States of America. 2005; 102(29): 10058-10063.
[6] Mouret S, Baudouin C, Charveron M, Favier A, Cadet J, Douki T. Cyclobutane pyrimidine dimers are predominant DNA
lesions in whole human skin exposed to UVA radiation. Proceedings of the National Academy of Sciences of the United States
of America. 2006; 103(37): 13765-13770.
[7] Collins AR. The comet assay for DNA damage and repair - Principles, applications, and limitations. Mol. Biotechnol. 2004;
26(3): 249-261.
[8] Brendler-Schwaab S, Hartmann A, Pfuhler S, Speit G. The in vivo comet assay: use and status in genotoxicity testing.
Mutagenesis. 2005; 20(4): 245-254.
[9] Filippova EM, Monteleone DC, Trunk JG, Sutherland BM, Quake SR, Sutherland JC. Quantifying double-strand breaks and
clustered damages in DNA by single-molecule laser fluorescence sizing. Biophysical Journal. 2003; 84(2): 1281-1290.
[10] Bustamante C Rivetti C. Visualizing protein-nucleic acid interactions on a large scale with the scanning force microscope.
Annual Review of Biophysics and Biomolecular Structure. 1996; 25(395-429.
[11] Colton RJ, Baselt DR, Dufrene YF, Green JBD, Lee GU. Scanning probe microscopy. Current Opinion in Chemical Biology.
1997; 1(3): 370-377.
[12] Guthold M, Bezanilla M, Erie DA, Jenkins B, Hansma HG, Bustamante C. Following the Assembly of Rna-Polymerase DNA
Complexes in Aqueous-Solutions with the Scanning Force Microscope. Proceedings of the National Academy of Sciences of the
United States of America. 1994; 91(26): 12927-12931.
[13] Hansma HG. Surface biology of DNA by atomic force microscopy. Annual Review of Physical Chemistry. 2001; 52(71-92.
[14] Hansma HG, Kasuya K, Oroudjev E. Atomic force microscopy imaging and pulling of nucleic acids. Current Opinion in
Structural Biology. 2004; 14(3): 380-385.
[15] Lindsay SM, Thundat T, Nagahara L, Knipping U, Rill RL. Images of the DNA Double Helix in Water. Science. 1989;
244(4908): 1063-1064.
[16] Lyubchenko YL Shlyakhtenko LS. Visualization of supercoiled DNA with atomic force microscopy in situ. Proceedings of the
National Academy of Sciences of the United States of America. 1997; 94(2): 496-501.
[17] Shao ZF, Mou J, Czajkowsky DM, Yang J, Yuan JY. Biological atomic force microscopy: What is achieved and what is
needed. Advances in Physics. 1996; 45(1): 1-86.
[18] Wang H, Yang Y, Schofield MJ, Du CW, Fridman Y, Lee SD, Larson ED, Drummond JT, Alani E, Hsieh P, et al. DNA
bending and unbending by MutS govern mismatch recognition and specificity. Proceedings of the National Academy of
Sciences of the United States of America. 2003; 100(25): 14822-14827.
[19] Chen LW, Haushalter KA, Lieber CM, Verdine GL. Direct visualization of a DNA glycosylase searching for damage.
Chemistry & Biology. 2002; 9(3): 345-350.
Microscopy: Science, Technology, Applications and Education A. Méndez-Vilas and J. Díaz (Eds.)
©FORMATEX 2010 467
______________________________________________

[20] Boichot S, Fromm M, Cunniffe S, O'Neill P, Labrune JC, Chambaudet A, Delain E, Le Cam E. Investigation of radiation
damage in DNA by using atomic force microscopy. Radiation Protection Dosimetry. 2002; 99(1-4): 143-145.
[21] Psonka K, Brons S, Heiss M, Gudowska-Nowak E, Taucher-Scholz G. Induction of DNA damage by heavy ions measured by
atomic force microscopy. Journal of Physics-Condensed Matter. 2005; 17(18): S1443-S1446.
[22] Lysetska M, Knoll A, Boehringer D, Hey T, Krauss G, Krausch G. UV light-damaged DNA and its interaction with human
replication protein A: an atomic force microscopy study. Nucleic Acids Research. 2002; 30(12): 2686-2691.
[23] Moreno-Herrero F, de Jager M, Dekker NH, Kanaar R, Wyman C, Dekker C. Mesoscale conformational changes in the DNA-
repair complex Rad50/Mre11/Nbs1 upon binding DNA. Nature. 2005; 437(7057): 440-443.
[24] Murakami M, Hirokawa H, Hayata I. Analysis of radiation damage of DNA by atomic force microscopy in comparison with
agarose gel electrophoresis studies. J. Biochem. Biophys. Methods. 2000; 44(1-2): 31-40.
[25] Pang D, Berman BL, Chasovskikh S, Rodgers JE, Dritschilo A. Investigation of neutron-induced damage in DNA by atomic
force microscopy: Experimental evidence of clustered DNA lesions. Radiation Research. 1998; 150(6): 612-618.
[26] Pang D, Rodgers JE, Berman BL, Chasovskikh S, Dritschilo A. Spatial distribution of radiation-induced double-strand breaks
in plasmid DNA as resolved by atomic force microscopy. Radiation Research. 2005; 164(6): 755-765.
[27] Jiang Y, Ke CH, Mieczkowski PA, Marszalek PE. Detecting ultraviolet damage in single DNA molecules by atomic force
Microscopy. Biophysical Journal. 2007; 93(5): 1758-1767.
[28] Jiang Y, Rabbi M, Kim M, Ke C, Lee W, Clark RL, Mieczkowski PA, Marszalek PE. UVA Generates Pyrimidine Dimers in
DNA Directly. Biophysical Journal. 2009; 96(3): 1151-1158.
[29] Ke C, Jiang Y, Mieczkowski PA, Muramoto GG, Chute JP, Marszalek PE. Nanoscale detection of ionizing radiation damage to
DNA by atomic force microscopy. Small. 2008; 4(2): 288-294.
[30] Lee G, Rabbi M, Clark RL, Marszalek PE. Nanomechanical fingerprints of UV damage to DNA. Small. 2007; 3(5): 809-813.
[31] Shlyakhtenko LS, Gall AA, Filonov A, Cerovac Z, Lushnikov A, Lyubchenko YL. Silatrane-based surface chemistry for
immobilization of DNA, protein-DNA complexes and other biological materials. Ultramicroscopy. 2003; 97(1-4): 279-287.
[32] Lobachevsky PN, Karagiannis TC, Martin RF. Plasmid DNA breakage by decay of DNA-associated auger electron emitters:
Approaches to analysis of experimental data. Radiat. Res. 2004; 162(1): 84-95.
[33] Sachs RK, Ponomarev AL, Hahnfeldt P, Hlatky LR. Locations of radiation-produced DNA double strand breaks along
chromosomes: a stochastic cluster process formalism. Math. Biosci. 1999; 159(2): 165-187.
[34] Rief M, Clausen-Schaumann H, Gaub HE. Sequence-dependent mechanics of single DNA molecules. Nature Structural
Biology. 1999; 6(4): 346-349.
[35] Florin EL, Rief M, Lehmann H, Ludwig M, Dornmair C, Moy VT, Gaub HE. SENSING SPECIFIC MOLECULAR-
INTERACTIONS WITH THE ATOMIC-FORCE MICROSCOPE. Biosensors & Bioelectronics. 1995; 10(9-10): 895-901.
[36] Cherny DI Jovin TM. Electron and scanning force microscopy studies of alterations in supercoiled DNA tertiary structure. J.
Mol. Biol. 2001; 313(2): 295-307.
[37] Vologodskii AV Cozzarelli NR. Conformational and Thermodynamic Properties of Supercoiled DNA. Annu. Rev. Biophys.
Biomolec. Struct. 1994; 23(609-643.
[38] Bussiek M, Mucke N, Langowski J. Polylysine-coated mica can be used to observe systematic changes in the supercoiled DNA
conformation by scanning force microscopy in solution. Nucleic Acids Research. 2003; 31(22): e137.
[39] Sperrazza JM, Register IIIJC, Griffith J. Electron microscopy can be used to measure DNA supertwisting. Gene. 1984; 31(1-3):
17-22.
[40] Lushnikov AY, Brown BA, Oussatcheva EA, Potaman VN, Sinden RR, Lyubchenko YL. Interaction of the Z alpha domain of
human ADAR1 with a negatively supercoiled plasmid visualized by atomic force microscopy. Nucleic Acids Research. 2004;
32(15): 4704-4712.
[41] Gruskin EA Lloyd RS. The DNA Scanning Mechanism of T4 Endonuclease-V - Effect of Nacl Concentration on Processive
Nicking Activity. Journal of Biological Chemistry. 1986; 261(21): 9607-9613.
[42] Pope LH, Davies MC, Laughton CA, Roberts CJ, Tendler SJB, Williams PM. Intercalation-induced changes in DNA
supercoiling observed in real-time by atomic force microscopy. Analytica Chimica Acta. 1999; 400(27-32.
[43] Ono MY Spain EM. Dynamics of DNA condensates at the solid-liquid interface by atomic force microscopy. Journal of the
American Chemical Society. 1999; 121(32): 7330-7334.
[44] Bussiek M, Toth K, Brun N, Langowski J. DNA-loop formation on nucleosomes shown by in situ scanning force microscopy
of supercoiled DNA. Journal of Molecular Biology. 2005; 345(4): 695-706.
[45] Kasas S, Thomson NH, Smith BL, Hansma HG, Zhu XS, Guthold M, Bustamante C, Kool ET, Kashlev M, Hansma PK.
Escherichia coli RNA polymerase activity observed using atomic force microscopy. Biochemistry. 1997; 36(3): 461-468.
[46] Lyubchenko YL. DNA structure and dynamics - An atomic force microscopy study. Cell Biochemistry and Biophysics. 2004;
41(1): 75-98.
[47] Abdelhady HG, Allen S, Davies MC, Roberts CJ, Tendler SJB, Williams PM. Direct real-time molecular scale visualisation of
the degradation of condensed DNA complexes exposed to DNase I. Nucleic Acids Research. 2003; 31(14): 4001-4005.
[48] Bezanilla M, Drake B, Nudler E, Kashlev M, Hansma PK, Hansma HG. Motion and Enzymatic Degradation of DNA in the
Atomic-Force Microscope. Biophysical Journal. 1994; 67(6): 2454-2459.
[49] Ellis JS, Abdelhady HG, Allen S, Davies MC, Roberts CJ, Tendler SJB, Williams PM. Direct atomic force microscopy
observations of monovalent ion induced binding of DNA to mica. Journal of Microscopy-Oxford. 2004; 215(297-301.
[50] http://www.clarechemical.com/gold.htm.
[51] Setlow RB Carrier WL. Pyrimidine Dimers in Ultraviolet-irradiated DNA's Journal of Molecular Biology. 1966; 17(237-254.
[52] Sancar A. Structure and function of DNA photolyase and cryptochrome blue-light photoreceptors. Chemical Reviews. 2003;
103(6): 2203-2237.
[53] van Noort J, Orsini F, Eker A, Wyman C, de Grooth B, Greve J. DNA bending by photolyase in specific and non-specific
complexes studied by atomic force microscopy. Nucleic Acids Research. 1999; 27(19): 3875-3880.
Microscopy: Science, Technology, Applications and Education A. Méndez-Vilas and J. Díaz (Eds.)
468 ©FORMATEX 2010
______________________________________________

[54] Smith SB, Cui YJ, Bustamante C. Overstretching B-DNA: The elastic response of individual double-stranded and single-
stranded DNA molecules. Science. 1996; 271(5250): 795-799.
[55] Cluzel P, Lebrun A, Heller C, Lavery R, Viovy JL, Chatenay D, Caron F. DNA: An extensible molecule. Science. 1996;
271(5250): 792-794.
[56] Park H, Zhang KJ, Ren YJ, Nadji S, Sinha N, Taylor JS, Kang CH. Crystal structure of a DNA decamer containing a cis-syn
thymine dimer. Proceedings of the National Academy of Sciences of the United States of America. 2002; 99(25): 15965-15970.
[57] Kim JK, Patel D, Choi BS. Contrasting Structural Impacts Induced by Cis-Syn Cyclobutane Dimer and (6-4)-Adduct in DNA
Duplex Decamers - Implication Ln Mutagenesis and Repair Activity. Photochemistry and Photobiology. 1995; 62(1): 44-50.
[58] Sinha RP Hader DP. UV-induced DNA damage and repair: a review. Photochemical & Photobiological Sciences. 2002; 1(4):
225-236.
[59] Rouzina I, Pant K, Karpel RL, Williams MC. Theory of electrostatically regulated binding of T4 gene 32 protein to single- and
double-stranded DNA. Biophysical Journal. 2005; 89(3): 1941-1956.
[60] Douki T, Court M, Sauvaigo S, Odin F, Cadet J. Formation of the main UV-induced thymine dimeric lesions within isolated
and cellular DNA as measured by high performance liquid chromatography-tandem mass spectrometry. Journal of Biological
Chemistry. 2000; 275(16): 11678-11685.
Microscopy: Science, Technology, Applications and Education A. Méndez-Vilas and J. Díaz (Eds.)
©FORMATEX 2010 469
______________________________________________


















