
Current Topics in Medicinal Chemistry 2004, 4 403-429 403 ... · 404 Current Topics in Medicinal...
28
Current Topics in Medicinal Chemistry 2004, 4, 403-429 403 1568-0266/04 $45.00+.00 © 2004 Bentham Science Publishers Ltd. Structural Features of Angiotensin-I Converting Enzyme Catalytic Sites: Conformational Studies in Solution, Homology Models and Comparison with Other Zinc Metallopeptidases Georgios A. Spyroulias *,1 , Athanassios S. Galanis 1 , George Pairas 1 , Evy Manessi-Zoupa 2 and Paul Cordopatis *,1 Departments of Pharmacy 1 and Chemistry 2 , University of Patras, GR-26504, Patras, GREECE Abstract: Angiotensin-I Converting Enzyme (ACE) is a Zinc Metallopeptidase of which the three-dimensional stucture was unknown until recently, when the Xray structure of testis isoform (C-terminal domain of somatic) was determined. ACE plays an important role in the regulation of blood pressure due to its action in the frame of the Renin-Angiotensin System. Efforts for the specific inhibition of the catalytic function of this enzyme have been made on the basis of the X- ray structures of other enzymes with analogous efficacy in the hydrolytic cleavage of peptide substrate terminal fragments. Angiotensin-I Converting Enzyme bears the sequence and topology characteristics of the well-known gluzincins, a sub-family of zincins metallopeptidases and these similarities are exploited in order to reveal common structural elements among these enzymes. 3D homology models are also built using the X-ray structure of Thermolysin as template and peptide models that represent the amino acid sequence of the ACE’s two catalytic, zinc-containing sites are designed and synthesized. Conformational analysis of the zinc-free and zinc-bound peptides through high resolution 1 H NMR Spectroscopy provides new insights into the solution structure of ACE catalytic centers. Structural properties of these peptides could provide valuable information towards the design and preparation of new potent ACE inhibitors. 1. INTRODUCTION 1.1. Angiotensin-I Converting Enzyme and Hypertension Angiotensin-I Converting Enzyme (ACE), isolated in the mid 50’s, is a Zinc Metallopeptidase and one of the major components of the so-called Renin-Angiotensin System (RAS) [1-3]. Renin is responsible for the liberation of Angiotensin I (AI) in blood, after renin’s catalytic action on the angiotensinogen. ACE possesses a crucial role in the regulation of blood pressure since it catalyzes the cleavage of the C-terminal His-Leu dipeptide of the rather inactive decapeptide Angiotensin I (AI), in the vasopressor octapeptide Angiotensin II (AII) [4-6] (Fig. (1)). However, ACE impact is not only focused on the generation of Angiotensin II but also extended to inactivation of the vasodilator peptides Bradykinin (BK) and Kallidin [7-9]. ACE is encountered in two distinct forms in humans, the somatic and the testis form. These differ from the structural point of view, mainly in size and number of catalytic sites [10,11]. According to its function, ACE is classified among the peptidyl dipeptidases of zinc metallopeptidases super- family due to its ability to remove C-terminal dipeptide [5] from substrates. ACE can also exhibit activity of an endopeptidase against substrates such as Substance P, Cholecystokinin and Luliberin (LHRH), peptides with amidated C-end [9,12,13]. As far as the ACE role in blood pressure is concerned, the inhibition of ACE enzymatic activity against AI was considered as one of the major challenges against hypertensive disease and congestive heart failure [14]. Therapy, today and after extensive research for *Address correspondence to these authors at Department of Pharmacy, University of Patras, GR-265 04, Greece; Tel: +30 2610 997 721; Fax: +30 2610 997 714; e-mail: [email protected] and [email protected] the last 30 years has been achieved through inhibitors based on the pioneering work of Ferreira S.H. [15] and Ondetti M.A. [16,17]. These researchers showed that the venom of a Brazilian pit viper contained a factor that greatly enhanced the smooth-muscle-relaxing action of the nonapeptide BK which also inhibiting ACE. All ACE inhibitors were prepared in the absence of the ACE’s three-dimensional structure and bear two main characteristics: (i) designed on the basis of venom peptide extracting structural information for the enzyme catalytic site from the crystal structure of Carboxypeptidase A (CPA) [18], and (ii) high biological activity strongly coupled with enhanced zinc binding ability [19]. 1.2. ACE Inhibitors and their Impact in Medicine and Pharmacology ACE inhibitors are considered among the most potent antihypertensive drugs and apart their major action, exhibit beneficial lateral effects in the prevention of cardiovascular disease in various classes of hypertensive patients. Additionally ACE inhibitors have been proven more effective than other hypertensive substances in reducing proteinuria and retarding the progression of renal damage in patients with various types of nephropathy. These features are probably among the reasons that two (International Society of Hypertension-World Health Organization, ISH- WHO; Canadian Society of Hypertension) of the three health organisations (the third is the British Hypertension Society) affiliated with hypertension, recommend ACE inhibitors in the first-line of antihypertensive drug treatment, after the results of the first trials of this type of inhibitors became available at 1996. An alternative treatment of hypertension is focused on the blockade of AII receptors, AT 1 R and AT 2 R (94% amino
Transcript of Current Topics in Medicinal Chemistry 2004, 4 403-429 403 ... · 404 Current Topics in Medicinal...

Current Topics in Medicinal Chemistry 2004, 4, 403-429 403
1568-0266/04 $45.00+.00 © 2004 Bentham Science Publishers Ltd.
Structural Features of Angiotensin-I Converting Enzyme Catalytic Sites:Conformational Studies in Solution, Homology Models and Comparisonwith Other Zinc Metallopeptidases
Georgios A. Spyroulias*,1, Athanassios S. Galanis1, George Pairas1, Evy Manessi-Zoupa2 and PaulCordopatis*,1
Departments of Pharmacy1 and Chemistry2, University of Patras, GR-26504, Patras, GREECE
Abstract: Angiotensin-I Converting Enzyme (ACE) is a Zinc Metallopeptidase of which the three-dimensional stucturewas unknown until recently, when the Xray structure of testis isoform (C-terminal domain of somatic) was determined.ACE plays an important role in the regulation of blood pressure due to its action in the frame of the Renin-AngiotensinSystem. Efforts for the specific inhibition of the catalytic function of this enzyme have been made on the basis of the X-ray structures of other enzymes with analogous efficacy in the hydrolytic cleavage of peptide substrate terminalfragments. Angiotensin-I Converting Enzyme bears the sequence and topology characteristics of the well-knowngluzincins, a sub-family of zincins metallopeptidases and these similarities are exploited in order to reveal commonstructural elements among these enzymes. 3D homology models are also built using the X-ray structure of Thermolysin astemplate and peptide models that represent the amino acid sequence of the ACE’s two catalytic, zinc-containing sites aredesigned and synthesized. Conformational analysis of the zinc-free and zinc-bound peptides through high resolution 1HNMR Spectroscopy provides new insights into the solution structure of ACE catalytic centers. Structural properties ofthese peptides could provide valuable information towards the design and preparation of new potent ACE inhibitors.
1. INTRODUCTION
1.1. Angiotensin-I Converting Enzyme and Hypertension
Angiotensin-I Converting Enzyme (ACE), isolated in themid 50’s, is a Zinc Metallopeptidase and one of the majorcomponents of the so-called Renin-Angiotensin System(RAS) [1-3]. Renin is responsible for the liberation ofAngiotensin I (AI) in blood, after renin’s catalytic action onthe angiotensinogen. ACE possesses a crucial role in theregulation of blood pressure since it catalyzes the cleavage ofthe C-terminal His-Leu dipeptide of the rather inactivedecapeptide Angiotensin I (AI), in the vasopressoroctapeptide Angiotensin II (AII) [4-6] (Fig. (1)). However,ACE impact is not only focused on the generation ofAngiotensin II but also extended to inactivation of thevasodilator peptides Bradykinin (BK) and Kallidin [7-9].ACE is encountered in two distinct forms in humans, thesomatic and the testis form. These differ from the structuralpoint of view, mainly in size and number of catalytic sites[10,11]. According to its function, ACE is classified amongthe peptidyl dipeptidases of zinc metallopeptidases super-family due to its ability to remove C-terminal dipeptide [5]from substrates. ACE can also exhibit activity of anendopeptidase against substrates such as Substance P,Cholecystokinin and Luliberin (LHRH), peptides withamidated C-end [9,12,13]. As far as the ACE role in bloodpressure is concerned, the inhibition of ACE enzymaticactivity against AI was considered as one of the majorchallenges against hypertensive disease and congestive heartfailure [14]. Therapy, today and after extensive research for
*Address correspondence to these authors at Department of Pharmacy,University of Patras, GR-265 04, Greece; Tel: +30 2610 997 721; Fax: +302610 997 714; e-mail: [email protected] and [email protected]
the last 30 years has been achieved through inhibitors basedon the pioneering work of Ferreira S.H. [15] and OndettiM.A. [16,17]. These researchers showed that the venom of aBrazilian pit viper contained a factor that greatly enhancedthe smooth-muscle-relaxing action of the nonapeptide BKwhich also inhibiting ACE. All ACE inhibitors wereprepared in the absence of the ACE’s three-dimensionalstructure and bear two main characteristics: (i) designedon the basis of venom peptide extracting structuralinformation for the enzyme catalytic site from the crystalstructure of Carboxypeptidase A (CPA) [18], and (ii) highbiological activity strongly coupled with enhanced zincbinding ability [19].
1.2. ACE Inhibitors and their Impact in Medicine andPharmacology
ACE inhibitors are considered among the most potentantihypertensive drugs and apart their major action, exhibitbeneficial lateral effects in the prevention of cardiovasculardisease in various classes of hypertensive patients.Additionally ACE inhibitors have been proven moreeffective than other hypertensive substances in reducingproteinuria and retarding the progression of renal damage inpatients with various types of nephropathy. These featuresare probably among the reasons that two (InternationalSociety of Hypertension-World Health Organization, ISH-WHO; Canadian Society of Hypertension) of the three healthorganisations (the third is the British Hypertension Society)affiliated with hypertension, recommend ACE inhibitors inthe first-line of antihypertensive drug treatment, after theresults of the first trials of this type of inhibitors becameavailable at 1996.
An alternative treatment of hypertension is focused onthe blockade of AII receptors, AT1R and AT2R (94% amino

404 Current Topics in Medicinal Chemistry, 2004, Vol. 4, No. 4 Spyroulias et al.
acid sequence identity), with appropriate antagonists. Theclass of AT1R which are currently under continuousdevelopment and trials, exhibit at the moment only one mainthe advantage over ACE inhibitors, which is the absence ofcough as a side effect. Interestingly, only ISH-WHO hasrecommended AT1R antagonists as first line antihypertensivedrugs, and this is probably due to the absence of anypublished long-term trial results. For all these reasons, thedesign and preparation of new potent ACE inhibitors stillremains one of the main challenges in the intersection of thefields of chemistry, pharmacology and medicine.
1.3. Zinc Catalytic Sites and their Characteristics
Zinc sites in metalloenzymes and related biomoleculesare classified according to their ligands and coordinationgeometry into three types of zinc binding sites [20,21]: (i)the catalytic, (ii) the cocatalytic and (iii) the structural.Hydrolases like ACE possess a catalytic zinc site whichusually coordinates with nitrogen, oxygen and sulphurdonors of His, Glu, Asp and Cys residues while His is mostfrequently encountered in the coordination sphere of zincmetal ion. Water is also a zinc ligand in catalytic sites and isactivated for ionisation, polarisation, or displacement by theidentity and arrangement of ligands coordinated with zinc[22]. The zinc coordination number for this kind of siteshas been found to be four or five and the donor atoms ofresidues define a distorted-tetrahedral or trigonal-bipyramidalcoordination geometry. Ionisation and/or polarisation of theactivated H2O is assisted by the base form of an active site-residue or in some cases by a “second-shell” residuethat yields hydroxide ions at neutral pH while waterdisplacement results in Lewis acid catalysis on the part of thecatalytic zinc metal.
The structure of the zinc catalytic sites comprises: (i) thezinc-bound residues, (ii) the characteristic sequence of theamino acids around those ligated to the metal, (iii) the shortor long amino acid “spacers” among the three (in some casesfour) protein ligands, and (iv) the conformational features of
the “spacers”. Zinc sites are also characterised by secondaryinteractions with neighbouring amino acids that position inspace and conformational features strongly depend on theoverall protein folding and three-dimensional structure.These features are critical for the structure-functionrelationship of this class of metalloenzymes and dictate theirclassification into various families.
The characteristic amino acid sequences, which containthe potential three zinc ligands in the zinc metallopeptidasefamily, comprise the binding motif sequences that are adiagnostic tool in enzyme classification. The first two of theprotein ligands are found in the first three-, four- or five-residue binding motif, while the third is found in a secondcharacteristic motif. These residue-ligands are generallyseparated either by short amino acid “spacers” among thefirst, second and third ligands, or by a short spacer betweenthe two first ligands and by a large spacer between thesecond and third, or fourth ligand, should one exist. Themagnitude of a short spacer could vary from one to threeamino acids between the first two ligands. On the other hand,the long spacers usually found in various metallopeptidasesubfamilies could vary from 5 to over 100 amino acids. Thelength of the spacer between the two first ligands belongingto the same binding motif often characterizes the secondarystructure of this protein fragment. For example, a three-residue spacer is characteristic of a α-helix conformationwhile a one-residue spacer indicates a β sheet conformation.
1.4. Focus of this Article
This article aims to provide new structural insights intoACE, an enzyme whose role in hypertension has stimulatedover the last 35 years extensive and continuous efforttowards designing its potential inhibitors, even without themost important tool in the hands of biochemists,enzymologists and drug designers: the ACE three-dimensional structure. What follows is an attempt to reviewthe latest progress in structural biology of zincmetallopeptidases and extract structural information through:
Fig. (1). Schematic representation of the Renin-Angiotensin system and the role of the two enzymes, Renin and Angiotensin ConvertingEnzyme (ACE) on regulation of blood pressure through the generation and release of Angiotensin II vassopressor peptide.

Structural Features of Angiotensin-I Converting Enzyme Catalytic Sites Current Topics in Medicinal Chemistry, 2004, Vol. 4, No. 4 405
(i) an investigation of ACE sequence and structuresimilarities/differences among representative members ofmetallopeptidase family, (ii) homologous modeling based onknown enzyme structure, and (iii) a solution conformationalanalysis through 1H NMR spectroscopy on 36-residuesynthetic peptides whose sequences represent the two ACEcatalytic sites and their zinc-binding properties.
2. ZINC METALLOPEPTIDASES CHARACTER-ISTICS AND CLASSIFICATION
The super-family of zinc-containing bio-macromoleculeshas been enormously extended over the last years, with thesame occurring in the frame of the zinc metalloproteases/peptidases family [23,24] (Fig. (2)). An attempt to classifypreviously unknown biopolymers with distinct motifs andzinc-binding affinity at the beginning of the 90’s ledresearchers to identify diagnostic motifs within thepolypeptide amino acid sequences through sequence andtopology comparison [45]. Using this methodology, not onlysequence but also structural similarities have been identifiedamong various zinc metalloproteases and thus enzymegroups into certain categories [46-52]. In 1992 Jiang andBond [48] compare the sequences around the diagnostic,
zinc-binding motif, HEXXH (X = any amino acid residue) inorder to classify the known zinc metalloproteases into fivefamilies, where each one has its prototype: (i) Thermolysin(TLN) [27,53-55] (ii) Astacin (AST) [34], (iii) Serratia [56-58], (iv) Matrixin [59,60] and (v) Reprolysin/Adamalysin[24,49,52,61]. Determination of the zinc metalloproteases3D crystal structure sheds new light on the structural featuresof these enzymes and new terms, such as zincins, metzincins,aspzincins, gluzincins and inverzincins, have been used inorder to discriminate between them [24,51,52] (see Fig. (2)).In addition to ACE conformational features extracted throughtheoretical and experimental studies, an update of the sequenceand topology characteristics of zinc metalloproteases/peptidases through the structural analysis of known 3Dstructures is presented.
2.1. Zincins, Metzincins and Gluzincins: Where ACEbelongs?
Overall, the zinc metallopeptidases can be divided intotwo categories according to the sequence of the first zincbinding motif: (i) the zinc enzymes with the characteristicHEXXH motif, where the two histidines are the potentialprotein ligands, and (ii) the zinc enzymes without HEXXH
Fig. (2). Classification of Zinc Metalloproteases according to MEROPS Protease Database (July 2002; http://www.merops.co.uk/merops/index.htm). Data concerning protein source and zinc ligands, amino acid spacer between the binding motifs, accession number and codes forsequence and coordinates are also given. The X-ray structures of the active site for some representative metalloenzymes are presented. Thezinc protein ligands are labelled. The number of amino acids (aa) refers to the spacer length between the protein zinc ligands. X stands forany amino acid while, in bold letters, His, Glu and Met residues are noted.

406 Current Topics in Medicinal Chemistry, 2004, Vol. 4, No. 4 Spyroulias et al.
motif. The presence of HEXXH defines the so-called zincinsfamily [49]. The amino acid sequence and topology close tothe second binding motif or the motif sequence itself dividesthe zincins family into metzincins [49,52] and gluzincins [24]subfamilies (Fig. (2)). According to Bode et al. [49] thecrystal structure determination of AST (pdb: 1ast [34]) andAdamalysin II (pdb: 1iag [61]; see also Fig. (3)) revealedthat these two proteinases exhibit low sequence similaritybut significant topological equivalence. Their main feature isthe almost identical zinc environment with a methionine-containing turn and, according to these findings, Bode hassuggested that these enzymes could be termed as metzincins.The third zinc ligand in the vast majority of metzincins is ahistidine residue, while in some members of this family atyrosyl residue has been identified as the fourth coordinatedprotein ligand [49,52]. However, the X-ray crystal structuresof Snapalysin (M7 family, pdb : 1kuh [31], Fig. (3)) andPeptidyl-Lys Metalloendopeptidase (M35 family, pdb: 1g12[62], Fig. (3)) revealed that an aspartate residue is the thirdprotein ligand, although these enzymes belong to the MA(M)clan and have been termed aspzincins [62].
The term gluzincins was introduced in the mid 90’s byHooper N. M. for the zincins whose third zinc ligand is aglutamic acid [24] found in the consensus binding motif
sequence EXXXX. TLN is considered the prototype memberof this category of zinc metallopeptidases, whose first X-raycrystal structure was solved almost 30 years ago [27,53-55](see also Fig. (2), (3)). The last residue of the EXXXXsequence for TLN (pdb: 1lnd) [27]) and Neprilysin (orNeutral Endopeptidase; NEP) (pdb: 1dmt [28]) crystalstructures has been found to be an aspartatic acid which isalso found in the same position of the proposed secondbinding motif of ACE (EAIGD). According to the sequenceidentity of the key-residues in the two potential bindingmotif sequences, ACE is classified among the gluzincinssuperfamily and specifically in the M2 family of MA(E) clanof zinc metalloproteases (Table (1)).
In 1994, the gluzincin term, according to Hooper,spanned six families of the metalloprotease MA clan, M1,M2, M3, M4, M13 and M27 with representative membersAminopeptidase A (EC 3.4.11.7), ACE (EC 3.4.15.1),Thimet Oligopeptidase (EC 3.4.24.15), TLN (EC 3.4.24.27),NEP (EC 3.4.24.11), and Botulinum neurotoxin A (EC 3.4.24.69) respectively. Until mid 2002, the gluzincin termencompasses eighteen metallopeptidase families, accordingto the Protease Database (http: // www . merops . co.uk/merops/merops.htm), while there are 9 3D X-ray structures out of 99assigned peptidase sequences classified in gluzincins,
Fig. (3). 3D X-ray structures of the zinc-containing active sites of the two superfamilies, metzincins MA(M) (left) and gluzincins MA(E)(right), of zinc metalloproteases, available until July 2002. Representative models, from each clan, which possesses at least one characteristicenzyme with available 3D structure, are presented and pdb codes are given. Prototype clan members with no 3D structure determined so farare also listed. Active site helices are shown in blue/gold ribbon representation while non-active helices are depicted in grey/light grey.

Structural Features of Angiotensin-I Converting Enzyme Catalytic Sites Current Topics in Medicinal Chemistry, 2004, Vol. 4, No. 4 407
Table 1. Amino Acid Composition of Zinc Binding Motif Sequences for Representative Members of All Known Families of MA(E)Clan, Gluzincins, Zinc Metallopeptidases. Horizontal Black Shaded Boxes Indicate the Family of the MA(E) Clan. Dark-gray Columns Indicate the Metal Binding Ligand which have Either been Identified Through the Available X-rayStructure or Proposed in Analogy to other Enzymes. PDB Code, when Available, is Given (Last Column).
Sequence1 MA(E) - M2 PDB REF
ACEN H390 EMG H394 ---23--- E418 AIGD
ACEC H390 EMG H394 ---23--- E418 AIGD 1O8A [148-9]
MA(E) – M1
LTA4 Hydrolase H295 EIS H299 ---18--- E318 GHTV 1hs6 [25]
M3A
Neurolysin H474 EFG H478 ---24--- E503 VPSQ 1i1i [26]
M3B
Oligopeptidase F H387 ETG H476 ---23--- E415 IAST
M4
Thermolysin H142 ELT H146 ---19--- E166 AISD 1lnd [27]
M5
Mycolysin H201 EAG H205 ---33--- E239 GFAD
M9A
Microbial collagenase H401 EYV H405 ---23--- E429 GIAE
M9B
Collagenase colA (H) H414 EYT H418 ---27--- E446 GGAE
M13
Neprilysin H583 EIT H587 ---58--- E646 NIAD 1dmt [28]
M26
IgA1 protease H1494 EMT H1498 - - - 1 9 - - -- - - 1 9 - - - EE 1 5 1 81 5 1 8 F F A K F F A K 2
M27
Botulinum neurotoxin H222 ELI H226 ---34--- E261 LRTF 3bta [29]
M30
Hyicolysin H246 EYQ H250 ---44--- E269 AFAM
M32
Carboxypeptidase Pfu H276 EMG H280 - - - 2 4 - - -- - - 2 4 - - - EE 3 0 53 0 5 S Q S R S Q S R 3 1k9x [150]
M34
Anthrax Lethal factor H719 EFG H723 ---44--- E768 FFAE 1j7n [30]
M36
Fungalysin H429 EYT H432 - - - 2 5 - - -- - - 2 5 - - - EE 4 5 84 5 8 G W S D G W S D 4
M41
FtsH endopeptidase H414 E_G H418 - - - 5 7 - - -- - - 5 7 - - - EE 4 7 64 7 6 E I I Y E I I Y 5
M47

408 Current Topics in Medicinal Chemistry, 2004, Vol. 4, No. 4 Spyroulias et al.
Table 1. (Contd….)
Sequence1 MA(E) PDB REF
PRSM1 metallopeptidase H204 ELG H208 -------- ---- ----6
M48_
CAAX prenyl protease H335 ELG H339 ---75--- E415 FQAD
M48_
HtpX endopeptidase H139 E_S H143 - - - 7 8 - - -- - - 7 8 - - - EE 2 2 22 2 2 F H A DF H A D 4
M60
Enhacin H248 ELG H252 - - - 1 4 - - -- - - 1 4 - - - EE 2 6 72 6 7 V W N NV W N N 7
M61
Glycyl aminopeptidase H248 ELG H252 - - - 2 9 - - -- - - 2 9 - - - EE 2 8 22 8 2 G Q T QG Q T Q 8
Consensus Sequence H EXX H E XXXX1 Residue numbering is according to the sequence record deposited at Swiss-Prot (http://www.expasy.ch/) when no 3D structure is available. With peptidases for which
coordinates have been deposited at Protein Data Bank (http://www.rcsb.org/pdb/) the residue numbering follows the PDB record. Numbers between the 2nd and 3rd protein ligandindicate the magnitude of amino acid spacer.
2 Tentative assignment – E 1518 is the 1 st conserved glutamate in all known sequences of M26 family. The 2nd conserved E is found after 90 intervening residues in a pentapeptidethat possesses the sequence EGNSI.
3 Tentative assignment – E305 is found in the consensus sequence of HESQSX (where X = R or L) of M32 family. This E has been assigned as 3rd zinc ligand according to theconserved residues of the motif and to the preceding histidyl residue, which is also found in the same position in ACE sequences.
4 Tentative assignment – E 458 has been assigned as 3rd zinc ligand since the 4th residue of the consensus sequence EXXXD is an Aspartic acid as has also been found in the sameposition in M4, M5, M13, M48A and ACE sequences.
5 Tentative assignment – E476 is the only conserved glutamate in all known sequences of M41 family.6 No assignment is attempted since only one sequence for M47 family is available. First E residue is found after 19 intervening residues in the sequence EWPGG.7 Tentative assignment – E267 is the 1st conserved glutamate in all known sequences of M60 family in the consensus sequence EXWZN (where X = I or V and Z= N or T). The 2nd
conserved E is found after 50 intervening residues in the non-consensus motif sequence ERNIA.8 Tentative assignment – E282 is the only conserved glutamate in all known sequences of M61 family in the consensus sequence EGXTZ (where X = T, F or Q and Z= S or Q).
yielding representative 3D models for six of the families(M1, M3, M4, M13, M27 and M34; see Fig. (3)).
The catalytic, zinc-containing, sites extracted from theavailable X-ray structures of representative members of zincmetalloproteases metzincins, MA(M), and gluzincins, MA(E)clans are illustrated in Fig. (3).
2.2. Sequence and Structure Characteristics: BindingMotifs and Active Sites of Zincins, Metzincins andGluzincins
Enzymes from each of the above mentionedmetallopeptidase clans and families are grouped belowaccording to three main characteristics concerning thecharacteristic sequence of: (i) the first zinc binding motif, (ii)the second zinc binding motif and (iii) the amino acid spacerbetween the two binding motifs and its magnitude.
The amino acid composition of their binding motifsequences and the number of residues which constitute thespacer for gluzincin sequences for which both binding motifshave been distinguished and characterized is illustrated atTable (1) while analogous data for metzincins are presentedat Table (2).
According to Hooper N. M. [24] the consensus sequenceof gluzincins is HEXXH-spacer-EXXXX and the magnitudeof the spacer varies between 18 and >80 amino acids. Exceptfor the three zinc ligands and the glutamic acid that is present
in all zincins’ first binding motif there is no other conservedamino acid in the two binding motifs. However, among the18 gluzincins families (plus three subfamilies), in 7 familiesthe fourth residue of the second, the “glutamate” bindingmotif, is an aspartic acid, as also happens in the ACEsequence. Of these families, only two, MA(E)-M4 and -M13,possess sequences for which X-ray crystal structures areavailable (Fig. (3)). Representative members of these twofamilies are TLN and NEP respectively, exhibiting theHEXTH and EXIXD consensus zinc binding motif sequences,which exhibit significant similarities with the HEMGH andEAIGD sequences of ACE. Additionally, TLN’s active sitepossesses a 19 amino acid spacer that is comparable to thatof the 23 amino acid spacer attributed to ACE’s active sitepolypeptide, while NEP possesses a spacer consisting of 58amino acids.
Despite the low overall sequence identity betweenmetzincins and gluzincins peptidases, such as for AST andTLN respectively, significant topological similarities in theiractive site have been implied by sequence comparison[24,29]. Crystal structure determination of metzincinsrepresentatives in the early 90’s, such as Adamalysin [pdb:1iag, 61] and AST [pdb: 1ast, 34], revealed that the firstbinding consensus zincins sequence HEXXH constitutes ahelix, which has also been found in the X-ray structure ofTLN [49,27,53-55]. Crystal structures of metzincins andgluzincins peptidases solved over the last decade verify the

Structural Features of Angiotensin-I Converting Enzyme Catalytic Sites Current Topics in Medicinal Chemistry, 2004, Vol. 4, No. 4 409
assumption that the HEXXH first binding sequence for allzincins is part of a helix, the so-called ‘active site helix’. Thesecond binding motif of metzincins and gluzincins variesstructurally as implied by sequence comparison betweenvarious peptidases of the two MA(M) and MA(E) clans. Thethird protein zinc ligand is found in a helical fragment for allgluzincins structures solved, but this is not the case withmetzincins peptidases. The latter possess a histidine oraspartate residue as a zinc third ligand, which has not arisenfrom a second helix but is located in an open coil segment.No matter what the amino acid composition, the length or theconformation of the spacer between the two binding motifsin gluzincins (Fig. (3)), the EXXXX sequence alwayscomprises a second ‘active site helix’. Consequently, theactive site environment in gluzincins is characterised by two‘active site helices’, which constitute the main component ofthe zinc catalytic pocket. Additionally, the major structuralfeature that characterises the conformation of the 18-58amino acids spacer is the α-helix, especially when themagnitude of this spacer is longer than 20 residues.
Summarising the information drawn from the analysis ofzincins structure available so far are one should note that:(i) Topological equivalence may occur even without
sequence equivalence, (ii) conformation of first and secondbinding zinc motif sequence for both metzincins andgluzincins is remarkably similar among members of eachfamily even when sequence identity of active site is low (15-25%) and intervening residues differ in nature and length.
2.3. Functional and Structural Relationship of ACE withother Zinc Metallopeptidases
Structural and catalytic properties of ACE have beendiscussed in the literature in comparison either with a zincin,the gluzincin metallopeptidase TLN, or with a non-zincinmetallopeptidase the CPA [63-69]. ACE is a dipeptidylcarboxypeptidase while CPA is an exopeptidase [63,64] andTLN is an endopeptidase [66,69].
Carboxypeptidase A (see Fig. (1)) has a HXXE as a firstbinding motif sequence and HXX as a second. CPA firstbinding motif is a four-residue motif in contrast to the five-residue zincins motif and possesses the glutamate and thehistidine as second and third protein ligand. In contrast, thefirst zincins motif sequence bears the two histidines and theglutamate follows as the third ligand sited at the secondbinding motif. None of the CPA binding motifs comprises ahelical fragment while the amino acid spacer between them
Table 2. Amino Acid Composition of Zinc Binding Motif Sequences for Representative Members of MA(M) Clan Families,Metzincins, Zinc Metallopeptidases which Posses at Least One Member with known Xray Crystal Structure. HorizontalBlack Shaded Boxes Indicate the Family of the MA(M) Clan. Dark-gray Columns Indicate the Metal Binding LigandAccording to Xray Structure. The Light-gray Column Indicates the Three-residue Sequence of the Characteristic ‘Met-turn’ while Tyrosyl Residue in bold Characters Indicates the Assigned Fourth Protein Zinc Ligand. PDB Code, whenAvailable, is Given (Last Column).
Sequence1 MA(M) – M7 PDB REF
Snapalysin H83 ETG H87 ----5--- D93 HYSG --5- M103SG 1kuh [31]
M8
Leishmanolysin H264 EMA H268 ---67--- H334 IKMR --6- M345AP 1lml [32]
M10A
Collagenase 12 H218 ELG H222 ----5--- H228 STDI --3- M236YP 1fbl [33]
M10B
Serralysin2 H176 EIG H180 ----5--- H186 PGDY -23- M214SY 1srp [58]
M12A
Astacin2 H92 EAM H96 ----5--- H102 EHTR -40- M147HY 1ast [34]
M12B
Adamalysin H142 ELG H146 ----5--- H152 DGKD --9- M166RP 1iag [61]
M35
Peptidyl-Lysendopeptidase
H117 ESS H121 ----9--- D130 YAYG ---- ----3 1g12 [62]
Consensus Seq. H EXX H H/D XXXX MXX1 Residue numbering is according to the record with the structure coordinates (PDB record) which have been deposited at Protein Data Bank (http://www.rcsb.org/pdb/). Numbers
between the 2nd and 3rd protein ligand, as well as before the “Met-turn” Methionine, indicate the magnitude of amino acid spacer.2 Tyrosine in the Met-turn sequence is noted with bold characters and indicates the 4th zinc ligand.3 No Met-turn motif has been identified for this peptidase either in identified sequence or in X-ray structure. Last residue in sequence (P81054; Swiss-Prot :
http://www.expasy.ch/) and in X-ray structure is Ser167.

410 Current Topics in Medicinal Chemistry, 2004, Vol. 4, No. 4 Spyroulias et al.
varies from 108 to 135 amino acids among the variouscarboxypeptidases sequences.
2.3.1. Carboxypeptidase A.
Research on the inhibition of ACE’s catalytic actionstarted in the early 1970s [15-17], and has been based onfunction similarities of ACE with other enzymes with known3D structures [70-72]. One of these enzymes was CPA (M14family of MC clan), the structure of which was determinedby X-ray crystallography (pdb: 1yme, Fig. (4)) [18,36] andits active site and catalytic mechanism has been extensivelyinvestigated. [63,73]. CPA possesses a zinc-binding motifwith the characteristic HXXE sequence, and is able to cleavea single amino acid from the carboxy-terminal end of apeptide substrate, in contrast with ACE, which hydrolysesthe COOH-terminal dipeptides. Ondetti and Cushman, whowere recently honored for their pioneering work andcontinuous effort in the design and study of potent andspecific ACE inhibitors [19], proposed a late 1970’s
hypothetical model of ACE based on the active center ofCPA. (see Fig. (4C)) [72]. They also suggested that thecatalytic mechanism of ACE is similar to that of CPA. Basedon these studies several research groups designed andsynthesized numerous potential ACE inhibitors, arriving atthe first pharmacologically promising antihypertensivecompounds that bind at ACE’s active sites [19].
Analysis of 3D structure of CPA catalytic cavity [73]revealed that a positively charged residue (noted by “+” inFig (4C)), Arg145, extends its side-chain NH3 groupstowards the metal site and has the role of forming ionicbonds with the negatively charged carboxyl group of thesubstrate’s C-terminal (Scheme (1A)). As an analogy to that,a positively charged residue is believed to exist at ACEactive site and participate in the catalytic mechanism of thehydrolytic dipeptide cleavage [72]. Also, in CPA, the zincion, which polarizes the carbonyl group of the scissilepeptide bond, has been found in a substrate’s single-residuefar from Arg145, which interact with the substrate’s COO-
Fig. (4). (A ) X-ray crystal structure of CPA (pdb: 1yme; top) and TLN (pdb: 1lnd; bottom). (B) Conformation of structural and catalyticalimportant residues of the zinc active site of CPA (B, top) and TLN (B, bottom). Side-chains of residues are presented in ball and stick. (C)Cartoon representation of active site of CPA based on its X-ray structure and a CPA-based hypothetical model of ACE active site wherevarious known inhibitors are accommodated. Sub-sites noted as S1, S2, S1’ and S2’ are cavities or areas in an enzyme’s active site whereamino acid groups interact with adjacent side-chains of the substrate’s (peptide or inhibitors) groups in a molecular recognition and complex-formation procedure.

Structural Features of Angiotensin-I Converting Enzyme Catalytic Sites Current Topics in Medicinal Chemistry, 2004, Vol. 4, No. 4 411
terminal group. On the other hand, at the ACE catalytic site,zinc metal ion has to be found in a position at a distance of adipeptide-residue, which itself must be far from theenzyme’s positively charged residue. Furthermore, Arg127,Glu270 orient their side-chains towards a zinc-catalytic site(Fig. (4B) & Scheme (1A)) and are implicated in CPAcatalytic mechanism. They have the task of polarizing thescissile carbonyl bond, while the Tyr248 OH group ishydrogen bonded to the terminal carboxylate of the substrate[63] (Scheme (1A)).
2.3.2. Thermolysin
Thermolysin is a thermostable extracellular gluzincinendopeptidase which belongs to the M4 family of MA(E)zinc metallopeptidase clan and has been isolated fromBacillus thermoproteolyticus. TLN catalyses the hydrolyticcleavage of the peptide bond specifically on the imino sideof large hydrophobic residues, and especially leucine,isoleucine and phenylalanine [74] His142, His146 andGlu166 together with a solvent water are found coordinatedwith the zinc ion in a distorted tetrahedral coordination [27].Glu143 and Asp170 are positioned in the so-called “secondcoordination shell” of the zinc cation and both belong to theconsensus HEXXH and EXIXD sequences of the first andsecond binding zinc motifs, respectively (Fig (4B)). Glu143is highly possibly hydrogen-bonded with its side-chaincarboxylate to the coordinated H2O, while Asp170, which isconserved in various gluzincins including ACE, is considereda structurally and/or functionally important residue (Scheme(1B)). The Asp170 structural role arises from the fact that itscharged side chain forms a salt link with the imidazole ringof His142, the first protein zinc-ligand [11,27,53]. Accordingto extensive structural studies of enzyme-inhibitorcomplexes, the catalytic role of the outer shell residues
His231, Tyr157 and Asp226 in the enzyme’s function hasbeen also suggested, Fig. (4B) [75-80].
The peptide substrate forms a Michaelis complex(Scheme (1B); 2nd step) with an enzyme’s active site and itscarbonyl oxygen is accommodated among His231, Tyr157and the coordinated H2O, which however becomes slightlydisplaced from its original position after peptide binding.Coordinated H2O is then sited closer to Glu143, in a positionfavorable for its polarization, which is also assisted by zinccation. Glu143 and metal ion therefore enhance thenucleophilicity of the H2O and promote its attack on thecarbonyl carbon. Glu143 accept a proton which in turn isdonated to the substrate’s peptide bond nitrogen, forming agem-diolate intermediate (Scheme (1B); 3rd step). At thisstage the zinc metal is in a five-coordination state and thepeptide carbon possesses a tetrahedral geometry. Thegeometry of the metal site and the intermediate has beenidentified through transition-state analogue inhibitors [75-80]. The formation of the intermediate product is believed tobe assisted by a hydrogen bond network where the side-chains of His231, Tyr157 and the carbonyl oxygen ofsubstrate are involved. The role of Asp266 is focused on thestabilization of the positive charge required for catalysisthrough a salt-bridge between its negative carboxylate side-chain and the protonated imidazole of His231 [74,80].Finally, the peptide C-N bond is cleaved and the protonatedproduct is released, while Glu143 is proposed to abstract thesecond water proton, shuttling it to the amine nitrogen(Scheme (1B); 4th step).
2.3.3. Angiotensin-I Converting Enzyme and Inhibitors
The discovery of two active sites in somatic ACE hasprovoked many assays to establish functional or structural
Scheme 1. Proposed catalytic mechanisms for the (A) Carboxypeptidase A and (B) Thermolysin catalyzed cleavage of peptides.

412 Current Topics in Medicinal Chemistry, 2004, Vol. 4, No. 4 Spyroulias et al.
difference in these two highly homologous enzyme regions.Studies of an ACE somatic isoform fragment that containsonly the N-terminal active site has been performed with theaim of identifying the functional differences or the selectiveinhibition between the two catalytic centers of ACE. Forexample, whereas the most important peptide substrates suchas BK and AI [4,9, 81,82] are hydrolyzed at both sites, whilesome others, like LHRH or Angiotensin1-7, are cleavedpreferentially only by N-domain active site [83-86].Additionally, it has been reported that the phosphiniccompound RXP407 is an N-selective inhibitor of ACE, ableto differentiate between the two active sites [87,88], insimilarity with some other peptide inhibitors (BBPs) whichhave been reported recently to selectively inhibit C-domainactive sites. Furthermore, it has been reported that ACEcatalytic activity requires monovalent anions such as Cl- formaximal activity and that the extent of activation is substrateand chloride dependent [89-91]. Data suggest that the C-domain active site is more sensitive to chloride concentrationfor the hydrolysis of some substrates [81,92].
The catalytic action of ACE has long been discussed inconcert with that of CPA and/or TLN [74,80], whose 3Dcrystal structure is available. In the absence of an ACEstructure, speculations are made on the basis of sequencehomology, or topology similarities with the above mentionedenzymes and structure-function correlation studies ofmutated enzymes. Between ACE and CPA/TLN, there isundoubtedly low sequence identity while the knownstructures of CPA and TLN differ considerably. Moreover,ACE binding motif and spacer resemble those of TLN whileboth differ remarkably from those of CPA. ACE bindingmotif sequences consist of the residues HEMGH.….EAIGDand between them 23 intervening residues have beenidentified in analogy to the sequences of TLN,HELTH…..EAISD separated by 19 residues. However, aprofound analysis of the TLN and CPA zinc environment intheir 3D structures suggest that there are some commonelements. For example CPA Glu270 is relative to TLNGlu143 (see Fig. (4A) and (4B)), and in analogy toGlu362/960 of the two ACE active sites considered vital forenzyme activity [93]. On the other hand the two residuessuggested as the proton donors in the catalytic mechanismsof CPA and TLN, Tyr248 and His231 respectively, are notfound in a comparable position in their structures. Thisstructural variation has promoted the aspect that there was noabsolute requirement for the involvement of a histidine or atyrosine in zinc neutral proteases hydrolytic action.Nevertheless, recently a histidyl residue, His1089 has beensuggested to possess a similar role to that of TLN His231.Additionally, sequence comparison of the amino acid spacerthat separates the second and the third zinc ligand in TLNand ACE reveals three and two tyrosines (368/965, 369 and372/970; somatic form numbering) for the two ACE catalyticsites analogous to Tyr157 in TLN.
Since until recently ACE structure remained obscure, thebinding affinity of potential inhibitors to ACE and structureactivity relationships have not been totally illuminated.Extensive studies have been performed towards a detailedunderstanding its active centers specificity. Data indicate thatthe majority of ACE inhibitors exhibit specificity on
interaction with mainly three subsites or pockets at the ACEactive sites, named S1, S1' and S2 (Fig. (4C)). Substratescontaining phenyl group (R3) that interacts with the ACE S1subsite have proved to be significant for the binding propertiesof inhibitors [94-97] like antihypertensive compoundsenalapril, lisinopril, trandolapril, fosinopril etc. (Fig. (4C)).Furthermore, S1' subsite has been found to have a weakerbinding affinity toward the peptide substrate, while it maynot be large enough to accommodate peptide substrates withextended, long, side-chains [98,99]. Additionally, mostsubstrates that specifically inhibit ACE active sites bearproline or proline-like analogues at their carboxy terminus,which interact with the S2' binding pocket [72,100](captopril, enalapril, lisinopril etc.; see Fig. (4C)). Anotherfeature that might be essential in a potential enzyme inhibitoris its free C-terminal carboxylate group [101]. According tothe zinc-binding group, inhibitors or potential inhibitors canbe classified in three categories: (i) those with a sulfydrylgroup, such as captopril [72,100,102], (ii) those with acarboxyl group, bound to zinc ion as enalapril [102],lisinopril [97, 103] etc and (iii) inhibitors with a phosphonategroup, like fosinopril [104]. The structures of the abovementioned compounds accommodated at the proposedhypothetical ACE catalytic cavity are presented in Fig. (4).
2.4. Structural Insights to ACE Active Sites ThroughSite-directed Mutagenesis
Attempts to extract conformational characteristics essentialfor an enzyme’s function have been also focused on thesubstitution of residues, which in analogy to other peptidasesare believed to be actively involved in ACE’s catalyticmechanism. Such residues should be sited close to the zincsite (Table (1)) and their role elucidated through site directedmutagenesis and structure-function relationship studies.
When the two histidines of the first potential ACE zinc-binding motif have been mutually substituted by other aminoacids, it is revealed that enzyme [11,92,103,105,106]completely abolishes its activity. These data indicate thatthese two histidyl residues are essential for ACE catalyticactivity. Moreover, these amino acids are believed to be thetwo first protein zinc-ligands. Further site-directedmutagenesis experiments yield a GluàAsp substitution,where the replaced glutamate residue is that found in thepossible two-histidyl binding motif with the sequenceHEMGH, which has resulted in suppression of enzymecatalytic activity [92]. Aspartate residue retains the negativecharge in the sequence, which, however, has been displacedof approximately 1.4 Å. This glutamate is involved in a basicattack of the substrate peptide bond and its role is ofcrucial importance to the catalytic efficacy of ACE. Thisstrongly supports the role postulated for Glu143 of theHELTH in TLN [86].
As far as the nature and properties of the second ACEpossible binding motif are concerned, the glutamate residuein EAIGD sequence is conserved in many other zincmetalloproteases which highly possibly plays the role of thethird protein zinc ligand. This glutamate characterizes thegluzincins and is, indeed, coordinated with the zinc metal ionaccording to the X-ray structures solved for other membersof this family. When an aspartic residue or a valine replaces

Structural Features of Angiotensin-I Converting Enzyme Catalytic Sites Current Topics in Medicinal Chemistry, 2004, Vol. 4, No. 4 413
this glutamic acid, ACE catalytic activity has been decreasedby more than two orders of magnitude, or has beencompletely extinguished, respectively [11]. Finally, mutationstudies performed by the same research group with asparticacid in EAIGD motif as target residue, which is replaced bya glutamic, or an alanine residue, indicate a specificfunctional role of its carboxylate side-chain. Asp170 role hasbeen proposed to as the salt link formation with theimidazole ring of the first potential zinc coordinated histidine[11], which is observed in TLN X-ray structure [27,53]. Asimilar role in ACE function is expected for Asp393/991(somatic isoform numbering).
Site directed mutagenesis studies have been alsoperformed in order to probe the nature of a basic amino acid,adjacent to an enzyme’s active site, which is believed to binda chloride ion. Concentration of chloride ions has beenreported among the crucial parameters that adjust ACEcatalytic activity [90,107] and Arg1098 [108] has beenreported critical for the chloride dependence of ACEcatalytic activity and binds the anion. However, this is incontrast with the results obtained through earlier chemicalmodification studies on lysyl residues [109], which hadproposed active involvement in chloride and othermonovalent anion binding [109,110].
Furthermore, substitution of His1089 of human somaticform, by an Ala or Leu, has prompted the researcher topropose that this histidyl residue stabilizes the transition statecomplex through hydrogen bonding with the tetrahedralintermediate product [111]. A similar role has been alsoattributed to His231 in the catalytic mechanism of TLN[74,80]. Finally, mutation studies of human and rabbit testisACE isoform have demonstrated a functional role of Tyr200and Tyr236, respectively [112-113] in analogy to the Tyr198in CPA.
3. 3D HOMOLOGY MODELS OF ACE ACTIVE SITES
Sequence alignment [114] of ACE and other gluzincinactive site fragments for which an X-ray crystal structure hasbeen determined is performed and presented in Fig. (5A).The structure of TLN active site (pdb: 1lnd [27]) has beenchosen as a template in order to generate 3D homologymodels of the 36-residue peptide that represent theACEN[His361-Ala396] and ACEC[His959-Ala994] zincactive sites, as shown in Fig. (5B) [115]. TLN zinc sitefulfils the following criteria: (i) high sequence identity withACE binding motifs, (ii) similar amino acid spacermagnitude, (iii) topological and conformational similaritiesresulting from theoretical prediction of ACE secondarystructure at NPSA server (Fig. (5C)). Despite the lowprimary structure similarity (<25%), the above mentionedfeatures indicate remarkable topological analogy.
3D homology ACE peptide models indicate that thebackbone folds in two helical fragments, one at eachterminus, where the two binding motifs are sited. Therefore,the two histidyl side-chains of the HEMGH motif areseparated by a α-helix turn and their side-chains are parallel.Therefore, they adopt the desired geometry in order todonate the imidazole nitrogens to the zinc coordinationsphere. The oxygen atom of the third zinc protein ligand thatof glutamate is also found in favourable geometry for zinc
coordination. The aspartate residue of EAISD motif orientsits side-chain parallel to that of glutamate and towards thefirst histidine imidazole ring.
The two antiparallel terminal helices in ACE homologymodels are in great agreement with the “two active helicessite” model adopted according to the two helices observed inthe catalytic cavities of all the 9 X-ray gluzincin crystalstructures available hitherto. These features appearindependently of the active site amino acid composition andmagnitude. Additionally, theoretical prediction of secondarystructure of ACE structure in Network Protein SequenceAnalysis [116] through GOR protocols [117-119] revealstwo major helices, one towards the N-terminus and the othertowards the C-terminus, both of which include the HEMGHand EIAGD bonding motifs.
Having analysed all the above data we wish to make aneffort to elucidate experimentally the structural features ofACE active sites. This attempt is performed through conform-ational analysis in solution, using NMR spectroscopy, of thesynthetic 36-residue ACE model peptides. The results ofNMR data analysis for apo- and zinc-bound forms ofACEN[His361-Ala396] peptide and NMR solution structureof the free peptide are presented below.
4. SYNTHESIS OF THE TWO ACE Zn-SITES – WHATCOULD BE THE BENEFIT?
X-ray crystallography and NMR spectroscopy are thetwo experimental methods, which could yield a high-resolution three-dimensional structure of biopolymers.However, both of these techniques have to overcome somelimitations. X-ray crystallography has the drawback ofsingle-crystal and heavy-atom derivative preparation. On theother hand, NMR bypasses the time-consuming stage ofcrystallization but is restricted by the size of the molecule.Larger NMR structures of biomolecules or biomolecularcomplexes reaches the limit of 50-60 kDa while sporadic arethe examples that complete sequence specific resonanceassignment has been performed for proteins with more than400 amino acid. But even in that case, NMR studies becomefeasible only after the development and the successfulapplications of molecular biology and biotechnologytechniques which permits the selective labeling anddeuteration in concert with 15N and 13C labeling. Untilrecently, ACE structure had not been elucidated, neither insolid-state, probably due to unsuccessful attempts to obtainhigh-quality crystals, nor in solution since its molecularweight exceeds 80 and 140 kDa, for testis and somaticisoform respectively. On the other hand, solid-phasesynthesis of peptides and polypeptides is able to producepeptides or polypeptides with any given sequence-bearingprotein or even non-protein amino acids in satisfactorily highyields and purity [120]. Additionally, synthetic peptides ofmetal-binding sequences are widely used in order to probethe structure, the metal-binding affinity and transportpathways of heavy metals to target proteins, since theirstructure closely resembles the native active site of ametallobiomolecule [121-123].
Zinc metal is essential for ACE function and substratebinding and for this reason our approach to investigate theconformational features of ACE catalytic cavity begins with

414 Current Topics in Medicinal Chemistry, 2004, Vol. 4, No. 4 Spyroulias et al.
the solid-phase synthesis of 36-residue peptide whose aminoacid composition and sequence represent the ACE active sitefragment. ACE zinc-binding sequence comprises a 29-residue fragment which contains the three proposed proteinligands; His361/959, His365/963 and Glu389/987 for thetwo zinc-sites, found at HEMGH and EAIGD (somaticisoform numbering). 36-residue synthetic peptides representthe ACEN[His361-Ala396] and ACEC[His959-Ala994] zinc-
binding sequences (ACEN and ACEC refer to zinc sitestowards N- and C- terminus respectively; Fig. (5C)).
The amino acid sequence of the above ACEN and ACEC
peptides was built “step by step” on the acid-sensitive 2-chlorotrityl chloride resin (substitution 0.6 mmol/g) applyingthe Fmoc strategy [124-126]. Final purification was achievedby semipreparative HPLC on a RP C-18 support (Phase SepC-18 S10 ODS2) eluted with a linear gradient 20% to 60%
Fig. (5). (A) Sequence alignment of the polypeptide fragments comprising the zinc active sites of ACE and representative gluzinicins ofwhich 3D X-ray structures are available. (B) TLN zinc-containing active site polypeptide (32 residue X-ray, pdb: 1lnd) and 3D homologymodels of ACEC/ACEN 36-residue peptides. Side chains of zinc ligands and F/Y, M/L, A/S and E/R variable ACEC/ACEN residues areshown in ball and stick representation for all models while the four different residues in ACEC and ACEN sequences are also illustrated. (C)Prediction of secondary structure at NPSA server through GOR protocols.

Structural Features of Angiotensin-I Converting Enzyme Catalytic Sites Current Topics in Medicinal Chemistry, 2004, Vol. 4, No. 4 415
acetonitrile (0.1% TFA) over 30 min at a 2 ml/min flow rate[127]. The final products were determined to be at least 96%pure by analytical HPLC. The overall yield was 48% forboth 36-residue constructs (Scheme 2).
These peptides share 89% sequence identity and are themain fragment of the ACE catalytic sites region (see Table(3)). Active site reconstitution has been performed throughaddition of ZnCl2 in a peptide solution and metal bindingproperties are monitored in solution.
5. CONFORMATIONAL STUDY OF ACEN
CATALYTIC SITE IN SOLUTION
5.1. 1H NMR Spectroscopy of ACEN[His361-Ala396] 36-residue Zinc-Free Peptide
5.1.1. NOE and Secondary Structure
The complete spin-system of 35 out of 36 residues andsequential assignment has been accomplished through the
combined analysis of the TOSCY [128,129] (Fig. (6A-B))and NOESY [130,131] spectra. His1 is the only amino acidwhere backbone and aliphatic chain protons have not beenidentified. However, the characteristic cross peak betweenthe non-exchangeable protons of His1 imidazole ring hasbeen assigned. No HN proton resonance was identified forGlu2. Assignment and chemical shifts are given in Table (4).
Fig. (6C) shows the short and medium range NOEobserved for the backbone and CβH protons in the ACEN
NOESY maps. There are three regions with diagnosticconnectivities for helix conformation, such as HN-HN(i,i +2), Hα-HN(i,i + 2), Hα-HN(i,i + 3) and Hα-Hβ(i,i + 3), thatwere observed. Met3-Tyr12, Val17-Gly22 and Gly26-Ala36define these regions in both peptides. On the other hand Hα-HN(i,i + 4) NOE are observed only in the first of the above-mentioned regions. This is due to signal overlapping whichdoes not allow unambiguous assignment of such type ofNOE for the Val17-Ala36 region.
Scheme 2. Flow chart for solid phase, step-by-step, synthesis and purification procedure for the ACE 36-residue constructs (HOBt, 1-hydroxybenzotriazole; DIC, N,N’-diisopropylcarbodiimide; Pip, pideridine; EDT, 1,2-ethanedithiol, TFA, trifluoroacetic acid).

416 Current Topics in Medicinal Chemistry, 2004, Vol. 4, No. 4 Spyroulias et al.
5.1.2. 3JHNHα Coupling Constants
Spin-spin coupling constant values could also be of greatimportance for the diagnosis of the secondary polypeptidestructure. The observed J depends on the average of the Js ofeach available conformation multiplied by its statisticalweight, and α- and 310- helical structures exhibit experimentalJ values in the range of 4.8 to 5.6 [132], somewhat largerthan the ideal values of 3.9 and 4.2 Hz respectively. On theother hand, extended structures such as the parallel or anti-parallel β sheet give rise to J values larger than 8.0 Hz. Most(14 out of 18 3JHNHα values) of the J coupling constantsmeasured in ACE peptides exhibits values below 6.2 Hz (seeFig. (6C)) [133,134].
5.1.3. Chemical Shift Index
The strong relationship between the backboneconformation and chemical shift Hα values serves as a strongindicator for the assignment of secondary structure in anypolypeptide sequence [135,136]. Chemical shift differenceanalysis between the observed Hα shift values and thecorresponding random coil values [137] is presented in Fig.(6D) and provides strong evidence for the conformationalpreference of the majority of amino acids towards the helical
configuration. There are three distinct regions where the∆δHα values are negative suggesting helical character (Ile6-Lys13, Val17-Gly22 and Gly26-Val34) and three shorterregions where the ∆δHα values are positive suggestingextended conformation (Glu2-His5, Asp14-Pro16 andAla23-Pro25). The most negative ∆δHα value is measured inboth peptides for Val17 (~ 0.4 ppm) while Asn24 exhibitsthe larger positive ∆δHα value (~ 0.3 ppm) (Fig. (6D)).
An overall evaluation of the observed NOE, 3JHNHα and∆δHα values, implies that in three fragments the skeleton ofthe 36-residue ACEN active site model peptide adopts a well-defined α-helical structure even in the absence of zinc metal[138]. The longer helices are anticipated for their C- and N-termini, and a smaller one for the intermediate region. Thetwo prolines, at positions 17 and 25, act as helix-stopresidues between the three helices. The second binding motifEAIGD seems to be part of the C-terminal helix while nosafe conclusion could be extracted for the first binding motif,HEMGH, which comprise the N-terminal pentapeptide.Taking into account the flexibility of any terminal peptidefragment and the fact that resonances for all protons for His1and HN proton of Glu2 have not been observed theconformation of this pentapeptide should be the average over
Table 3. Amino Acid Composition of the Active Sites’ 44 Residue Polypeptide Containing One or Two Zinc Binding Motifs ofANGIOTENSIN-I Converting Enzyme Known Sequences. Dark-gray Columns Indicate the Metal Binding LigandAccording to the SWISS-PROT Record.
Sequence 1st B. M.1 Spacer 2nd B. M.1 AA2
ACE_HUMAN3 VAH H390 EMG H394 IQYYLQYKDLPVSLRRGANPGFH E418 AIGD VLALS 1306
“ --- H988 --- H992 ---FM-------A--E------- E1016 ---- -----
ACE_MOUSE3 TVH H395 EMG H399 VQYYMQYKDLHVSLRRGANPGFH E423 AIGD VLALS 1312
“ IA- H993 --- H997 I--F------P-TF-E------- E1021 ---- IM---
ACE_CHICK3 TVH H288 EMG H292 VQYYLQYKDQPVSFRGGANPGFH E316 AIGD VLSLS 1193
“ --- H886 --- H890 ---F---M-Q-I---D------- E913 ---- -MA--
ACE_RABIT3,4 TVH H395 EMG H399 VQYYLQYKDQPVSLRR- ANPGFH E422 AIGD VLALS 1310
“ V-- H992 --- H996 I--FM----L--A--EG------ E1020 ---- -----
ACE_RAT3 TVH H396 EMG H400 VQYYLQYKDLHVSLRRGANPGFH E424 AIGD VLALS 1310
“ IA- H994 --- H998 I--FM-----P-TF--------- E1022 ---- -----
ACE_DROME5 TVH H367 ELG H371 IQYFLQYQHQPFVYRTGANPGFH E395 AVGD VLSLS 615
ACE_HAEIE6 --- H367 -M- H371 ----------------------- E395 ---- ----- 615
ACET_HUMAN VAH H414 EMG H418 IQYFMQYKDLPVALREGANPGFH E442 AIGD VLALS 732
ACET_MOUSE I-- H413 --- H417 ------------TF--------- E441 ---- IM--- 732
ACET_RABIT VV- H419 --- H423 ----------------------- E447 ---- ----- 737
Cons. Seq. XXH H EXG H XQYXXQYXXXXXXXRXGANPGFH E AXGD XXXLS1 Amino acids belong to 1st and 2nd binding motif. Numbering is according to the sequence record deposited at Swiss-Prot (http://www.expasy.ch/).2 Number of amino acids found in the sequence.3 Somatic ACE isoforms possess two zinc-containing active sites, while testis isoform contains only one.4 A residue is missing at the position of conserved Gly residue in all the other ACE sequences and noted by - 5 Drosophila melanogaster (Fruit fly).

Structural Features of Angiotensin-I Converting Enzyme Catalytic Sites Current Topics in Medicinal Chemistry, 2004, Vol. 4, No. 4 417
various conformers. However, the positive ∆δHα values arerelatively large for Glu2 and His5 (>0.10 ppm) and close tothe random coil values for Met3 and Gly4. The J values,feasibly measured for Met3 and His5, found ≤ 6.2 Hz forACEC, the same holding for Met3. These data arecontroversial with ∆δHα values, but are in great agreementwith the NOE sequential connectivities illustrated at Fig.(6C). According to the observed NOE, almost all the helix-diagnostic connectivities of Hα-HN(i,i + 2), Hα-HN(i,i + 3),Hα-Hβ(i,i + 3), and Hα-HN(i,i + 4) type, involving theMet3-Gly4-His5 amino acids, have been observed inNOESY maps. These data suggest that these three residuesof the N-terminal binding motif are in α-helical structure andpossibly comprise the initial turn of the first 10-12-residuehelix of ACEN/C.
5.2. High Resolution NMR Solution Structures ofACEN[His361-Ala396] 36-residue Zinc-Free Peptide
DYANA [139,140] structure calculations have beenperformed using 22.0 NOE-derived distance constraints perresidue (>15 meaningful) together with 20 H-bond distanceconstraints (two distance limits for each H-bond) and 18 ϕ
constraints. The resulting DYANA family of 30 structureshas rmsd values (calculated for residues 3-33) of 0.57 ± 0.24Å and 1.25 ± 0.21 Å respectively, for backbone and heavyatoms. The target function lies in the range 0.39-0.45 Å2
(0.43 ± 0.018 Å2). The final REM [141,142] family exhibitspairwise rmsd values for the 30 structures 0.57 ± 0.24 Å,1.27 ± 0.21 Å and to the mean structure 0.40 ± 0.18 Å, 0.89± 0.11 Å for backbone and heavy atoms respectively (Fig.(6E)). Restraint violations and structural and energeticstatistics for the ACEN[His361-Ala396] 36-residue Zinc-Freepeptide are reported at Table (5).
The ACEN polypeptide chain (Fig. (7)) is characterizedby the high content of helical structure, which is distributedin three fragments; the α-helical N- and C- terminal togetherwith the 310-helix observed in the center of the intermediatefragment between the two other helices. Two turns of thepeptide skeleton, the first after the end of the N-terminalhelix in the region of Asp14-Pro16 and the second after theintermediate helix in the region of Asn24-Pro25 creates a U-shaped cavity in the middle of the peptide sequence. This U-turn structure is comprised of the residues Lys13-Pro25 andbrings the two terminal helices to a distance, which varies
Fig. (6). 1H-1H 2D TOCSY 600-MHz NMR of ACEN (A & B) and ACEC (F & G) fingerprint region of H_-HN protons (H2O/TFE-d2
34%/66% v/v, at pH=5.0, T= 298 K). The number of the amino acid in the ACE sequences, to which the H_-HN and side-chain protonconnectivities belong, is noted. (C) Sequential connectivities for ACEN are presented together with the predicted secondary structureelements. 3JHNH_ coupling constants are illustrated by arrows (↓ for values in the range 4.2 –5.6, ↑ for values equal or above 8.0 Hz) or byfilled circles (values in the range of 6.0-7.0 Hz). (D) ∆δH_ chemical shift difference (in ppm) from random coil values according to CSI. TheNMR data were acquired on a Bruker AVANCE 600 spectrometer. (E) Number of meaningful NOE constraints per residue for ACEN.White, grey, and dark grey bars respectively represent intraresidue, sequential, and medium-range connectivities. Long-range connectivitieshave not been observed. Schematic presentation of the predicted secondary structures according to the sequential connectivities is shown atthe bottom of panel (C) and according to CSI at the top of panel (D).

418 Current Topics in Medicinal Chemistry, 2004, Vol. 4, No. 4 Spyroulias et al.
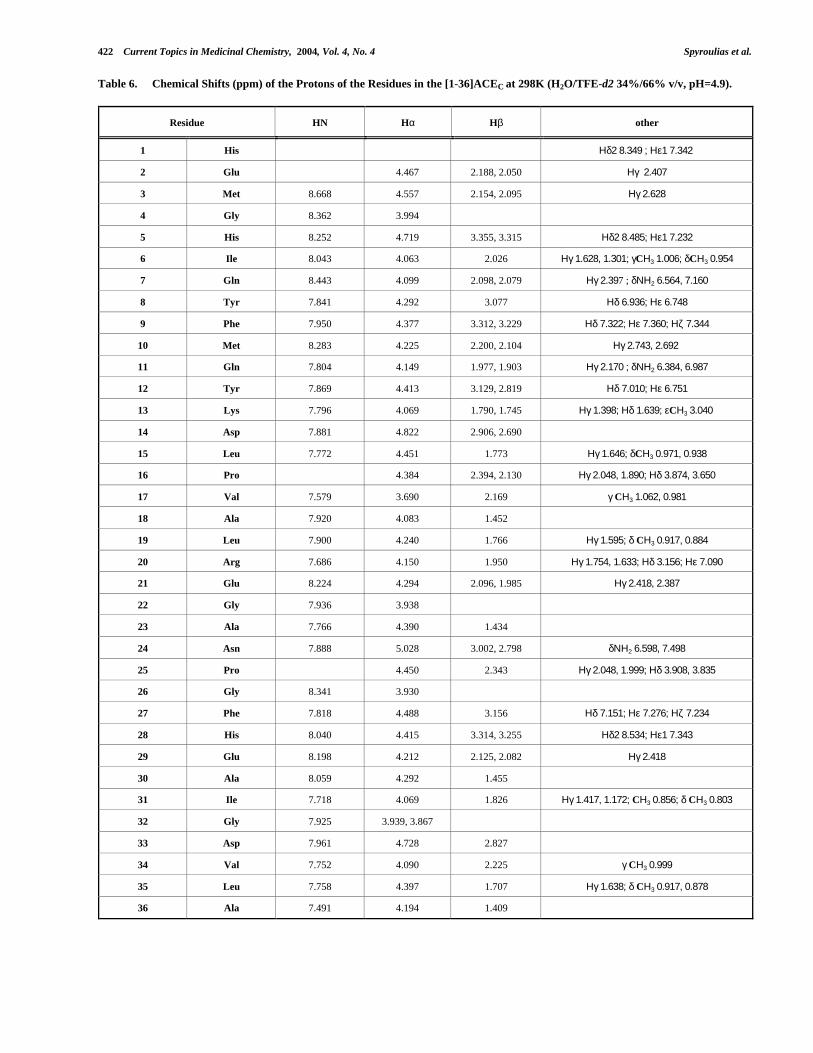
Table 4. Chemical Shifts (ppm) of the Protons of the Residues in the [1-36]ACEN at 298K (H2O/TFE-d2 34%/66% v/v, pH=4.9).
Residue HN Hα Hβ other
1 His Ηδ2 8.263; Ηε1 7.298
2 Glu 4.457 2.184, 2.040 Ηγ 2.389
3 Met 8.692 4.539 2.150, 2.094 Ηγ 2.618
4 Gly 8.385 3.994
5 His 8.214 4.743 3.370, 3.323 Ηδ2 8.457; Ηε1 7.246
6 Ile 8.032 4.071 2.039 Ηγ 1.628, 1.313; γCΗ3 1.010; δCΗ3 0.963
7 Gln 8.511 4.029 2.115, 2.081 Ηγ 2.392 ; δΝΗ2 6.554, 7.141
8 Tyr 7.785 4.268 3.088 Ηδ 6.948; Ηε 6.754
9 Tyr 7.823 4.311 3.253, 3.158 Ηδ 7.177; Ηε 6.881
10 Leu 8.202 4.052 1.945, 1.874 Ηγ 1.526; δCΗ3 0.930
11 Gln 7.780 4.123 1.985, 1.890 Ηγ 2.191, 2.138 ; δΝΗ2 6.389, 6.975
12 Tyr 7.850 4.426 3.135, 2.819 Ηδ 7.015; Ηε 6.775
13 Lys 7.830 4.082 1.795, 1.751 Ηγ 1.398; Ηδ 1.624; εCΗ3 3.032
14 Asp 7.915 4.809 2.875, 2.657
15 Leu 7.778 4.532 1.779 Ηγ 1.597; δCΗ3 0.970, 0.943
16 Pro 4.450 2.316, 2.105 Ηγ 2.041, 1.967; Ηδ 3.862, 3.653
17 Val 7.688 3.815 2.179 γCΗ3 1.053, 0.999
18 Ser 7.999 4.240 3.980, 3.924
19 Leu 7.796 4.345 1.733 Ηγ 1.650; δCΗ3 0.936, 0.891
20 Arg 7.754 4.207 1.929 Ηγ 1.748, 1.628 ; Ηδ 3.169; Ηε 7.063
21 Arg 8.016 4.278 1.895, 1.835 Ηγ 1.744, 1.647 ; Ηδ 3.174; Ηε 7.153
22 Gly 7.953 3.959
23 Ala 7.780 4.380 1.416
24 Asn 7.956 5.020 2.931, 2.762 δΝΗ2 6.613, 7.443
25 Pro 4.444 2.387, 2.319 Ηγ 2.054, 1.990; Ηδ 3.900, 3.804
26 Gly 8.331 3.931
27 Phe 7.816 4.512 3.153, 3.117 Ηδ 7.180; Ηε 7.299; Ηζ 7.249
28 His 8.053 4.458 3.306, 3.223 Ηδ2 8.493; Ηε1 7.303
29 Glu 8.244 4.228 2.123, 2.063 Ηγ 2.394
30 Ala 8.097 4.323 1.457
31 Ile 7.722 4.091 1.849 Ηγ 1.453, 1.196; γCΗ3 0.883; δCΗ3 0.830
32 Gly 7.952 3.954, 3.863
33 Asp 7.960 4.728 2.810
34 Val 7.759 4.099 2.226 γCΗ3 0.990
35 Leu 7.795 4.400 1.700 Ηγ 1.629; δCΗ3 0.918, 0.880
36 Ala 7.476 4.179 1.402

Structural Features of Angiotensin-I Converting Enzyme Catalytic Sites Current Topics in Medicinal Chemistry, 2004, Vol. 4, No. 4 419
Table 5. Statistical Analysis for the REM a and <REM> a Structures of [1-36]ACEN.
REM <REM>
RMS Violations per Experimental Distance Constraints (Å) b
intraresidue (128) 0.0213 ± 0.0030 0.0215
sequential (253) 0.0258 ± 0.0018 0.0311
medium-range (173) 0.0265 ± 0.0015 0.0237
total (544) 0.0251 ± 0.0013 0.0269
Average Number of Violations per Structure
intraresidue 7.00 ± 1.84 10.0
sequential 17.67 ± 2.05 17.0
medium-range 14.07 ± 1.39 14.0
total 38.73 ± 2.94 41.0
average no. of NOE violations > than 0.3 Å 0.000 ± 0.00 00.0
largest residual NOE distance violation (Å) 0.203 0.259
average distance penalty function (Å2) 0.415 ± 0.03 0.466
Statistics of Other Structural Constraints
ϕ constraints from 3JHNHα (18)
RMS violations per ϕ constraint 0.0627 ± 0.24
average no. of ϕ violations per structure 0.0667 ± 0.25
largest residual ϕ violations 0.0062
average torsion penalty function (kJ mol-1) 0.0008 ± 0.003 0.0004
AMBER energy (kJ mol-1) -1078.44 ± 59.8 -1107.12a REM indicates the energy-minimized family of 40 structures and <REM> the mean energy-minimized structure.
b Numbers in parenthesis indicate the number of meaningful upper distance limits per class.
Fig. (7). Backbone representation of: (A) Family of 30 ACEN REM models and (B) mean REM structure.
420 Current Topics in Medicinal Chemistry, 2004, Vol. 4, No. 4 Spyroulias et al.
from 13.5 to 24.5 Å (between the C_ atoms of Gln7 andHis28, the carbon atom of -COO- and the nitrogen atom of -NH3, respectively), one above the other, while their dipolesform an angle of almost 90°. The helical fragmentscomprised by 12 (Glu2-Lys13) and 8 (Phe27-Val34) residuesfor the N- and C- termini, respectively, and by 3 residues(Arg20-Gly22) for the intermediate [138].
The structure of the C-terminal fragment has beendetermined with higher resolution than that of the N-terminal, due to the higher number of NOE and angleconstraints. No proton resonances have been identified forthe backbone and side-chain aliphatic protons of His1 andamide proton of Glu2, probably due to the conformationalaveraging and/or exchange with solvents of labile NHprotons. The two prolines, in position 16 and 25 of thesequence, found in the intervening residues between thehelices probably interrupt the helicity of the backbone assuggested also by sequential NOE and ∆δHα values. Addit-ionally, next to Pro25 is found Gly26 and these two residuescomprise a notorious residue-pair with helix-destabilizingability, since both one characterized by the lowest helixpropensity among the 20 natural amino acids [143,144].
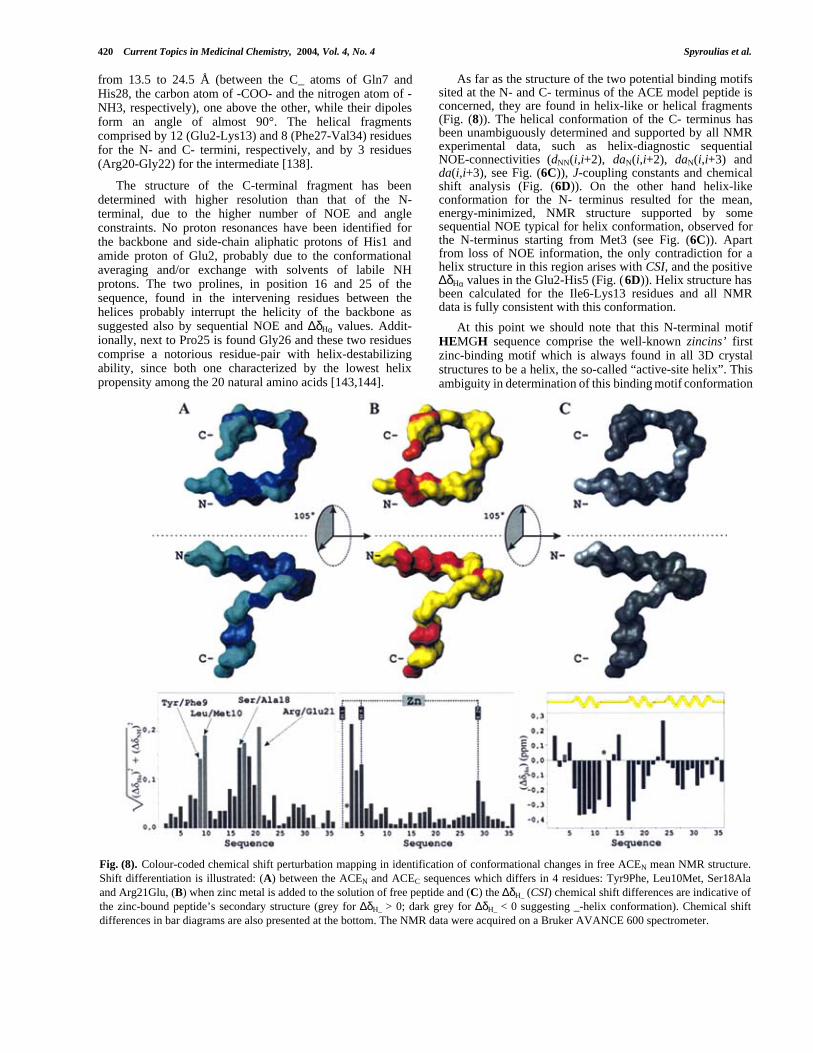
As far as the structure of the two potential binding motifssited at the N- and C- terminus of the ACE model peptide isconcerned, they are found in helix-like or helical fragments(Fig. (8)). The helical conformation of the C- terminus hasbeen unambiguously determined and supported by all NMRexperimental data, such as helix-diagnostic sequentialNOE-connectivities (dNN(i,i+2), daN(i,i+2), daN(i,i+3) andda(i,i+3), see Fig. (6C)), J-coupling constants and chemicalshift analysis (Fig. (6D)). On the other hand helix-likeconformation for the N- terminus resulted for the mean,energy-minimized, NMR structure supported by somesequential NOE typical for helix conformation, observed forthe N-terminus starting from Met3 (see Fig. (6C)). Apartfrom loss of NOE information, the only contradiction for ahelix structure in this region arises with CSI, and the positive∆δHα values in the Glu2-His5 (Fig. (6D)). Helix structure hasbeen calculated for the Ile6-Lys13 residues and all NMRdata is fully consistent with this conformation.
At this point we should note that this N-terminal motifHEMGH sequence comprise the well-known zincins’ firstzinc-binding motif which is always found in all 3D crystalstructures to be a helix, the so-called “active-site helix”. Thisambiguity in determination of this binding motif conformation
Fig. (8). Colour-coded chemical shift perturbation mapping in identification of conformational changes in free ACEN mean NMR structure.Shift differentiation is illustrated: (A) between the ACEN and ACEC sequences which differs in 4 residues: Tyr9Phe, Leu10Met, Ser18Alaand Arg21Glu, (B) when zinc metal is added to the solution of free peptide and (C) the ∆δH_ (CSI) chemical shift differences are indicative ofthe zinc-bound peptide’s secondary structure (grey for ∆δH_ > 0; dark grey for ∆δH_ < 0 suggesting _-helix conformation). Chemical shiftdifferences in bar diagrams are also presented at the bottom. The NMR data were acquired on a Bruker AVANCE 600 spectrometer.

Structural Features of Angiotensin-I Converting Enzyme Catalytic Sites Current Topics in Medicinal Chemistry, 2004, Vol. 4, No. 4 421
strongly depends on the fact that the first histidine of themotif is the first peptide residue. However, even in this casethe mean NMR solution structure of the free peptides fitswell with the “two active-site helices” model suggested forgluzincins. Both binding motifs are in helix or helix-likeconformation. The side-chains of the two histidines areparallel, the same occurring with side-chains of the thirdprotein zinc-ligand, the glutamate, and the aspartate fourresidues after glutamate towards the C-terminus, whoseconformation plays a crucial role in the catalytic activity ofsome gluzincins.
5.3. Analysis of the Tyr9Phe, Leu10Met, Ser18Ala andArg21Glu Differentiation Between ACEN and ACEC
Through Chemical Shift Perturbation Mapping
The two ACE 36-residue peptides designed andsynthesized differ, as actually happens in native sequence, infour residues (89% sequence identity), found in positions 9,10, 18 and 21. The aromatic and aliphatic character of aminoacids in positions 9 and 10 are in general maintainedbetween the two sequences, due to Tyr/Phe and Leu/Metsubstitution. On the other hand, the polar Ser18 which isconsidered a “surface” residue is replaced by the non-polar,“internal”, Ala. Additionally, the basic, positively chargedArg21 has been replaced by the acidic, negatively charged,glutamic acid without perturbing the hydrophilic propertiesat this point of the sequence. These residue variationsbetween the two ACE peptides do not seem to provoke anynoticeable conformational change in the secondary structureof the studied biopolymers, since according to NMR data(sequential connectivities and ∆δHα values) the helicalcharacter of the fragments where the four residues reside isnot disturbed.
However, comparative analysis of proton resonancesthrough Chemical Shift Perturbation Mapping [145], provides∆δHα -HN values between 0.15 and 0.20 ppm. These differencesare mapped in the backbone of the 3D average NMRstructure of ACEN peptide and presented in different colorsat the top and center of Fig. (8A). Chemical shift changesaround Tyr/Phe9 and Leu/Met10 are moderate and slightlyexceed 0.05 ppm, while the relative changes for the peptidefragment Leu15-Arg/Glu21 fluctuate between 0.07 and 0.17ppm. Additionally, Asn24 present a chemical shift differencevalue of around 0.075 ppm. Differences around 0.05 ppmhave also been estimated for the His28-Ala30 tripeptide.Plots of the Hα and HN shift differences are also presentedat the bottom of Fig. (8A). Assignment and chemical shiftsfor the 36-residue peptide ACEC are given in Table (6).
5.4. 1H NMR Spectroscopy, Zn(II) Binding Propertiesand Conformational Features of ACEN[His361-Ala396]36-residue Peptide
5.4.1. NOE and Secondary Structure – Differences withFree Peptides
Color-coded chemical shift changes, upon Zn(II) addition,are presented in Fig. (8B) while chemical shift differenceanalysis between the observed Hα shift values and theircorresponding random coil values are presented in Fig. (8C).Zinc binding properties of ACE peptides are monitored
through 1H 1D and 2D NMR Spectroscopy and many protonresonances in the NH region shift considerably and theirhalf-width is considerably increased when Zn(II) is added(Fig. (9A) & (9D)). Additionally, the previously degeneratedresonance of the two geminal Gly Hα protons has been splitinto two well-defined resonances (Fig. (9B) & (9C). Thisresonance different-iation for the previously degenerated Hαglycine protons suggests a metal-coupled structural changeof the N-terminal motif and indicates a different magneticenvironment for those nuclei in metal-peptides from those intheir free state. NMR data suggest similar elements ofsecondary structure between free and zinc-bound peptides[146].
The Hα values of residue fragment with negative orpositive ∆δHα values in free peptides exhibit the same natureand magnitude of deviation from their corresponding randomcoil values. In the presence of Zn(II) the majority of residues(23 residues) still exhibit negative deviation (larger than 0.05ppm) from random coil values (Fig. (8C)). ∆δHα values inACEN-Zn peptide define three possibly helical regions : (i) a7-residue fragment close to the peptide N-terminus (Ile6-Lys13), (ii) another 6-residue fragment in the middle of thesequence (Val17-Arg21), and (iii) a third comprising of 10residues close to peptide C-terminus (Gly26-Val34). Thesedata strongly suggest that in the zinc-bound peptide, thesecond proposed binding motif (EAIGD) retains its helicalconformation, also observed for the free peptides. As far asthe two histidyl motif is concerned, no definite conclusioncould be reached [146]. In the TLN [27] crystal structure,that binding motif, which possesses similar sequence to thatof ACE, has been found in helical conformation.
However, helix-diagnostic dαN(i + 3) type connectivitybetween Gly4 and Gln7, present in the NOESY spectrum offree peptides disappears when Zn(II) is added (Fig. (9B)).Nevertheless, typical helix dαN(i + 4) and dαN(i + 3) connect-ivities between Ile6-Met10 and Gln7-Met10 respectively, infree peptides are still present after zinc addition [146]. Thesedata strongly suggest that the HEMGH motif conformationmight undergo a conformational transition from helix-like orpartial helix to non-helix structure when zinc coordinateswith the donor atoms of the peptide ligands. This couldprobably be due to the fact that the first peptide zinc ligand isalso the first residue in the peptide sequence. Upon the lackof other residue(s), the His1 backbone and side-chain possessa remarkably high degree of conformational freedom. Thus,when zinc binds the peptide bonds of the pentapeptidebinding sequence HEMGH, it could be accommodated in arandom, open coil segment. Such conformation justifies theabsence of the typical sequential NOE for a helix and hasresulted after preliminary DYANA structure calculation forACEN 36-residue peptide. On the other hand, no changes forthe fragment Ile6-Lys13 are implied by NMR data.Moreover, the same type as for free peptides sequential NOEhas been detected and structure calculation indicates that thefirst HEMGH open coil structure in ACEN follows a helicalfragment which is extended from Ile6 to Tyr12/Lys13.Another difference between free and zinc-bound peptides,which has also been identified through sequential connect-ivities diagram and preliminary structure calculations, is theabsence of the helical fragment in the area Ser18-Arg21 in
422 Current Topics in Medicinal Chemistry, 2004, Vol. 4, No. 4 Spyroulias et al.
Table 6. Chemical Shifts (ppm) of the Protons of the Residues in the [1-36]ACEC at 298K (H2O/TFE-d2 34%/66% v/v, pH=4.9).
Residue HN Hα Hβ other
1 His Ηδ2 8.349 ; Ηε1 7.342
2 Glu 4.467 2.188, 2.050 Ηγ 2.407
3 Met 8.668 4.557 2.154, 2.095 Ηγ 2.628
4 Gly 8.362 3.994
5 His 8.252 4.719 3.355, 3.315 Ηδ2 8.485; Ηε1 7.232
6 Ile 8.043 4.063 2.026 Ηγ 1.628, 1.301; γCΗ3 1.006; δCΗ3 0.954
7 Gln 8.443 4.099 2.098, 2.079 Ηγ 2.397 ; δΝΗ2 6.564, 7.160
8 Tyr 7.841 4.292 3.077 Ηδ 6.936; Ηε 6.748
9 Phe 7.950 4.377 3.312, 3.229 Ηδ 7.322; Ηε 7.360; Ηζ 7.344
10 Met 8.283 4.225 2.200, 2.104 Ηγ 2.743, 2.692
11 Gln 7.804 4.149 1.977, 1.903 Ηγ 2.170 ; δΝΗ2 6.384, 6.987
12 Tyr 7.869 4.413 3.129, 2.819 Ηδ 7.010; Ηε 6.751
13 Lys 7.796 4.069 1.790, 1.745 Ηγ 1.398; Ηδ 1.639; εCΗ3 3.040
14 Asp 7.881 4.822 2.906, 2.690
15 Leu 7.772 4.451 1.773 Ηγ 1.646; δCΗ3 0.971, 0.938
16 Pro 4.384 2.394, 2.130 Ηγ 2.048, 1.890; Ηδ 3.874, 3.650
17 Val 7.579 3.690 2.169 γ CΗ3 1.062, 0.981
18 Ala 7.920 4.083 1.452
19 Leu 7.900 4.240 1.766 Ηγ 1.595; δ CΗ3 0.917, 0.884
20 Arg 7.686 4.150 1.950 Ηγ 1.754, 1.633; Ηδ 3.156; Ηε 7.090
21 Glu 8.224 4.294 2.096, 1.985 Ηγ 2.418, 2.387
22 Gly 7.936 3.938
23 Ala 7.766 4.390 1.434
24 Asn 7.888 5.028 3.002, 2.798 δΝΗ2 6.598, 7.498
25 Pro 4.450 2.343 Ηγ 2.048, 1.999; Ηδ 3.908, 3.835
26 Gly 8.341 3.930
27 Phe 7.818 4.488 3.156 Ηδ 7.151; Ηε 7.276; Ηζ 7.234
28 His 8.040 4.415 3.314, 3.255 Ηδ2 8.534; Ηε1 7.343
29 Glu 8.198 4.212 2.125, 2.082 Ηγ 2.418
30 Ala 8.059 4.292 1.455
31 Ile 7.718 4.069 1.826 Ηγ 1.417, 1.172; CΗ3 0.856; δ CΗ3 0.803
32 Gly 7.925 3.939, 3.867
33 Asp 7.961 4.728 2.827
34 Val 7.752 4.090 2.225 γ CΗ3 0.999
35 Leu 7.758 4.397 1.707 Ηγ 1.638; δ CΗ3 0.917, 0.878
36 Ala 7.491 4.194 1.409
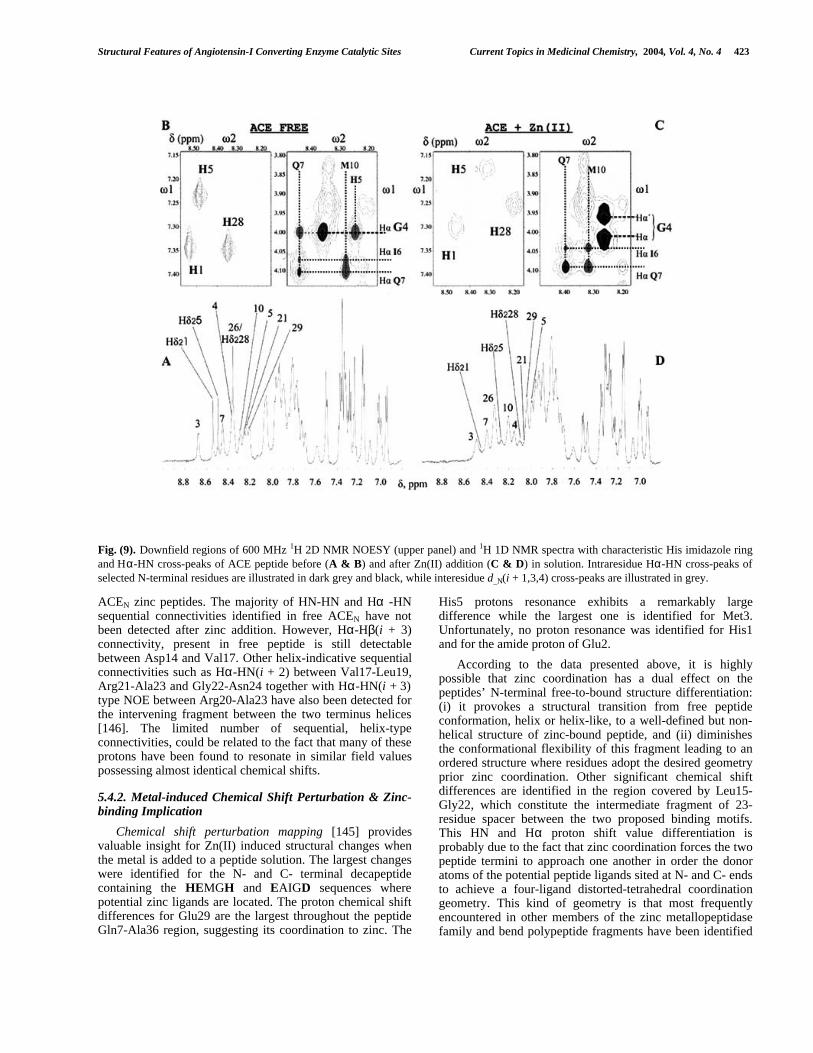
Structural Features of Angiotensin-I Converting Enzyme Catalytic Sites Current Topics in Medicinal Chemistry, 2004, Vol. 4, No. 4 423
Fig. (9). Downfield regions of 600 MHz 1H 2D NMR NOESY (upper panel) and 1H 1D NMR spectra with characteristic His imidazole ringand Hα-HN cross-peaks of ACE peptide before (A & B) and after Zn(II) addition (C & D) in solution. Intraresidue Hα-HN cross-peaks ofselected N-terminal residues are illustrated in dark grey and black, while interesidue d_N(i + 1,3,4) cross-peaks are illustrated in grey.
ACEN zinc peptides. The majority of HN-HN and Hα -HNsequential connectivities identified in free ACEN have notbeen detected after zinc addition. However, Hα-Hβ(i + 3)connectivity, present in free peptide is still detectablebetween Asp14 and Val17. Other helix-indicative sequentialconnectivities such as Hα-HN(i + 2) between Val17-Leu19,Arg21-Ala23 and Gly22-Asn24 together with Hα-HN(i + 3)type NOE between Arg20-Ala23 have also been detected forthe intervening fragment between the two terminus helices[146]. The limited number of sequential, helix-typeconnectivities, could be related to the fact that many of theseprotons have been found to resonate in similar field valuespossessing almost identical chemical shifts.
5.4.2. Metal-induced Chemical Shift Perturbation & Zinc-binding Implication
Chemical shift perturbation mapping [145] providesvaluable insight for Zn(II) induced structural changes whenthe metal is added to a peptide solution. The largest changeswere identified for the N- and C- terminal decapeptidecontaining the HEMGH and EAIGD sequences wherepotential zinc ligands are located. The proton chemical shiftdifferences for Glu29 are the largest throughout the peptideGln7-Ala36 region, suggesting its coordination to zinc. The
His5 protons resonance exhibits a remarkably largedifference while the largest one is identified for Met3.Unfortunately, no proton resonance was identified for His1and for the amide proton of Glu2.
According to the data presented above, it is highlypossible that zinc coordination has a dual effect on thepeptides’ N-terminal free-to-bound structure differentiation:(i) it provokes a structural transition from free peptideconformation, helix or helix-like, to a well-defined but non-helical structure of zinc-bound peptide, and (ii) diminishesthe conformational flexibility of this fragment leading to anordered structure where residues adopt the desired geometryprior zinc coordination. Other significant chemical shiftdifferences are identified in the region covered by Leu15-Gly22, which constitute the intermediate fragment of 23-residue spacer between the two proposed binding motifs.This HN and Hα proton shift value differentiation isprobably due to the fact that zinc coordination forces the twopeptide termini to approach one another in order the donoratoms of the potential peptide ligands sited at N- and C- endsto achieve a four-ligand distorted-tetrahedral coordinationgeometry. This kind of geometry is that most frequentlyencountered in other members of the zinc metallopeptidasefamily and bend polypeptide fragments have been identified

424 Current Topics in Medicinal Chemistry, 2004, Vol. 4, No. 4 Spyroulias et al.
in other gluzincs, such as TLN. Such a model is consistentwith a bend peptide skeleton of a U-like shape, as has beenobserved in the case of TLN X-ray crystal structure [146].
6. CONCLUSIONS
6.1. Common and Variable Structural Features BetweenFree ACE Peptides, 3D Homology ACE Models andCatalytic Sites of Known Gluzincin Structures
Having gone through theoretical and experimental studiesin order to elucidate the solution structure of the ACE zinc-containing active sites various valuable structuralinformation for these biologically interesting catalyticcentres has been extracted:
i. Despite the low sequence similarities among knownsequences of gluzincins, all the available X-ray crystalstructures possess a characteristic “two helix active-site”, no matter what the length of the inter-helicalamino acid spacer (Fig. (3)).
ii. Theoretical study of the secondary structure of the 36-residue active site model ACE peptides predictshelical conformation for the N- and C- terminus,where the two zinc-binding motif sequences are sited(HEMGH and EAIGD, respectively, Fig. (5C)).
iii. 3D homology models of these 36-residue peptidespossess two helices, one at each peptide terminus,suggesting that the peptide bonds could beaccommodated in conformation with high helicaldegree (Fig. (5B)). These helices are found to form anangle similar to that of TLN helices (Fig. (10)).
iv. Experimental study of the free peptides in solutionthrough high resolution 1H NMR spectroscopy,followed by structure calculation using NMR-derivedstructural constraints (NOEs, 3JHNHα) in concert withCSI suggest helix conformation for the N- and C-terminus (Fig. (6C-E), (7) and (8)).
v. Minor conformational differences between free andzinc-bound peptides could be identified for thesecondary structure of the first binding motifsequence, constituting the first pentapeptide fragment.The conformational freedom of the first peptide zinc-ligand, His1, could be responsible for this structuralvariation, for which however the helical structure hasalso been predicted (Fig. (5C)).
vi. Remarkable backbone conformational similaritiescould be identified even between the free modelpeptides studied in solution and the structure ofTLN’s active site in solid state. Among them the twohelices in homologous regions of the polypeptidechains such as the binding motifs (the N- and C-ACEN peptide termini) are of immense interest (Fig.(10), upper panel).
vii. The NMR solution structure of the free ACEN peptide,the 3D homology ACE-Zn models and the TLN activesite structure exhibit striking similarities. Data suggestthat even in the absence of metal ion the synthesisedpolypeptide folds in a tertiary structure where thehelical content is comparable to that of the model and
TLN and the two helices approach each other. TheNMR structure of the ACEN free peptide reveals thatthe potential zinc ligands could bind the metal withoutseverely disturbing the elements of its tertiary andsecondary structure. The conformation of the aminoacid spacer (310 helix fragment) and the relativeorientation of the two helices could be substantiallydifferentiated upon zinc binding to the polypeptide,and the peptide structure could further resemble thatof the models and TLN.
viii. The conformation of the 23-residue spacer couldeither retain or perturb the 310 helix conformation ofthe Arg20-Gly22 fragment observed in free peptideNMR structure. The small negative value exhibited byLeu19 (see Fig. (8C) bottom) could be consideredwithin the error of the CSI method and if that happenswould indicate a helix-stop signal for this peptidefragment. Nevertheless, a non-helix, coil-like structureis consistent with TLN active site and helix structurecould be also consistent with the Neurolysin structure,which possess a spacer similar to ACE 24-residuewith a small helical fragment (see also Fig. (3)).
ix. One of the main structure-property related differencesbetween the TLN and ACE active centre is the lack ofany positively-charged residue in the TLNpolypeptide fragment, which comprise its zinc-containing catalytic site (Fig. (10), lower panel).Electrostatic potentials not only in the surface but alsoin the substrate channel and cavity are crucial for themolecular recognition and complex formation processbetween enzyme and substrate.
6.2. Outlook and Perspectives
According to the above data, free and zinc peptidesexhibit conformational features highly similar to the zinc-containing active sites of various known zinc metallo-peptidases that belong to the same super-family as ACE, thegluzincins. The structural calculation based on NMR-derivedconstraints for the two ACE-Zn peptides is under way andthe models of both zinc-containing ACE catalytic sites willbecome available in the form of 36-residue peptides. Thesemodels could be used in enzyme-substrate complexsimulation/docking studies in an effort to understand thefactors governing the association of the two molecules.
The structural information acquired for ACE active sitesthrough NMR spectroscopy and structure calculation indicatethat the multidimensional process based on design, solid-phase synthesis and conformational analysis of ACE metal-binding sequences seems to be able to provide a suitablemaquette of the actual ACE catalytic sites. Valuable inform-ation could be acquired and would be exploited in structure-based design of biologically interesting substances againsthypertension, until the three-dimensional structure of theentire Angiotensin-I Converting Enzyme becomes available.
Note added in proof
While this paper was in press, the crystal structures of humantesticular [148] and Drosophila [149] ACE were reported.Additionally, shortly before the release of ACE model, the

Structural Features of Angiotensin-I Converting Enzyme Catalytic Sites Current Topics in Medicinal Chemistry, 2004, Vol. 4, No. 4 425
Xray structure of another gluzincin metalloprotease, that ofCarboxypeptidase Pyrococcus Furiosus (M32 clan), hadbeen determined [150].
ACKNOWLEDGEMENTS.
University of Patras for a K. Karatheodoris Research Grant(P.C., G.A.S.) and General Secretariat of Research andTechnology of Greece-Pened 99 Program (G.P., E.M.-Z.)are acknowledged for financial support. We also thank
EC’s Access to Research Infrastructures Action of theImproving Human Potential Program (PARABIO, ContractNo. HPRI-CT-1999-00009) for further support (G.A.S., P.C.and A.S.G.).
REFERENCES[1] Inagami, T. The renin–angiotensin system. Essays Biochem. 1994,
28, 147–164.[2] Ondetti, M. A.; Cushman, D. W. Enzymes of the renin-angiotensin
system and their inhibitors. Annu. Rev. Biochem. 1982, 51, 283-308.
Fig. (10). Comparison between the ribbon representation of the 3D structures of zinc active sites of (A) Thermolysin, (B), ACEC (C) ACEN
3D homology models, and (D) NMR structure is presented in upper panel. The helix angles calculated in terms of helix dipoles are alsogiven. The distribution of the electrostatic potential on the surface of the models is also presented for the peptides in lower panel. Figureswere generated with the program MOLMOL [147].

426 Current Topics in Medicinal Chemistry, 2004, Vol. 4, No. 4 Spyroulias et al.
[3] Roks, A.; Buikema, H.; Pinto, Y. M.; van Gilst, W. H. The renin-angiotensin system and vascular function. The role of angiotensinII, angiotensin-converting enzyme, and alternative conversion ofangiotensin I. Heart Vessels 1997, 12, 119-124.
[4] Skeggs, L. T.; Kahn, J. R.; Shumway, N. P. Preparation andfunction of the hypertension converting enzyme. J. Exp. Med. 1956,103, 295-299.
[5] Soffer, R. L. Angiotensin-converting enzyme and the regulation ofvasoactive peptides. Annu. Rev. Biochem. 1976;45, 73-94.
[6] Ehlers, M. R.; Riordan, J. F. Angiotensin-converting enzyme: newconcepts concerning its biological role. Biochemistry 1989, 28,5311-5318.
[7] Dorer, F. E.; Kahn, J. R.; Lentz, K. E.; Levine, M.; Skeggs, L. T.Hydrolysis of bradykinin by angiotensin-converting enzyme. Circ.Res. 1974, 24, 824–827.
[8] Kokkonen, J. O.; Lindstedt, K. A.; Kuoppala, A.; Kovanen, P. T.Kinin-degrading pathways in the human heart. Trends Cardiovasc.Med. 2000, 10, 42-45.
[9] Yang, H. Y. Y.; Erdös, E. G.; Levin, V. A dipeptidylcarboxypeptidase that converts angiotensin I and inactivatesbradykinin. Biochem. Biophys. Acta 1970, 214, 374-376.
[10] Soubrier, F.; Alhenc-Gelas, F.; Hubert, C.; Allegrini, J.; John, M.;Tregear, G.; Corvol, P. Two putative active centers in humanangiotensin I-converting enzyme revealed by molecular cloning.Proc. Natl. Acad. Sci. U.S.A. 1988, 85, 9386-9390.
[11] Williams, T. A.; Corvol, P.; Soubrier, F. Identification of two activesite residues in human angiotensin I-converting enzyme. J. Biol.Chem. 1994, 269, 29430-29434.
[12] Skidgel, R. A.; Erdös, E. G. Novel activity of human angiotensin Iconverting enzyme: release of the NH2- and COOH-terminaltripeptides from the luteinizing hormone-releasing hormone. Proc.Natl. Acad. Sci. U.S.A. 1985, 82, 1025-1029.
[13] Ehlers, M. R.; Riordan, J. F. Angiotensin-converting enzyme: zinc-and inhibitor-binding stoichiometries of the somatic and testisisozymes. Biochemistry 1991, 30, 7118-7126.
[14] Gavras, H. Angiotensin converting enzyme inhibition and its impacton cardiovascular disease. Circulation 1990, 81, 381-388.
[15] Ferreira, S. H.; Bartelt, D. C.; Greene, L. J. Isolation of bradykinin-potentiating peptides from Bothrops jararaca venom . Biochemistry1970, 9, 2583-2593.
[16] Ondetti, M. A.; Williams, N. J.; Sabo, E. F.; Pluscec, J.; Weaver, E.R.; Kocy, O. Angiotensin-converting enzyme inhibitors from thevenom of Bothrops jararaca. Isolation, elucidation of structure, andsynthesis. Biochemistry 1971, 10, 4033-4039.
[17] Cheung, H. S.; Cushman, D. W. Inhibition of homogenousangiotensin-converting enzyme of rabbit lung by synthetic venompeptides of Bothrops jararaca. Biochim. Biophys. Acta 1973, 293,451-463.
[18] Steitz, T. A.; Ludwig, M. L.; Quiocho, F. A.; Lipscomb, W. N. Thestructure of carboxypeptidase A. V. Studies of enzyme-substrateand enzyme-inhibitor complexes at 6 Å resolution. J. Biol. Chem.1967, 242, 4662-4668.
[19] Cushman, D. W.; Ondetti, M. A. Design of angiotensin convertingenzyme inhibitors. Nat. Med. 1999, 5, 1110-1113.
[20] Vallee, B. L.; Auld, D. S. Zinc Coordination, Function, andStructure of Zinc Enzymes and Other Proteins. Biochemistry 1990,29, 5647-5659.
[21] Vallee, B. L.; Auld, D. S. New Perspective on Zinc Biochemistry:Cocatalytic Sites in Multi-Zinc Enzymes. Biochemistry 1993, 32,6493-6500.
[22] Vallee, B. L; Auld, D. S. Active-site zinc ligands and activated H2Oof zinc enzymes. Proc. Natl. Acad. Sci. U.S.A. 1990, 87, 220-224.
[23] Auld, D. S. Zinc sites in metalloenzymes and related proteins. InHandbook on Metallopreoteins; Bertini, I.; Sigel, A.; Sigel, H., Eds,Marcel Dekker, 2001, pp. 881-959.
[24] Hooper, N. M. Families of zinc metalloproteases. FEBS Lett. 1994,354, 1-6.
[25] Thunnissen, M. M. G. M.; Nordlund, P. N.; Haeggstrom, J. Z.Crystal Structure of Human Leukotriene A4 Hydrolase, aBifunctional Enzyme in Inflammation. Nat. Struct. Biol. 2001, 8,131-135.
[26] Brown, C. K.; Madauss, K.; Lian, W.; Beck, M. R.; Tolbert, W. D.;Rodgers, D. W. Structure of Neurolysin Reveals a Deep Channelthat Limits Substrate Access. Proc. Nat. Acad. Sci.U.S.A. 2001, 98,3127-3132.
[27] Holland, D. R.; Hausrath, A. C.; Juers, D.; Matthews, B. W.Structural analysis of zinc substitutions in the active site ofthermolysin. Protein Sci. 1995, 4, 1955-1965.
[28] Oefner, C.; D'Arcy, A.; Hennig, M.; Winkler, F. K.; Dale, G. E.Structure of Human Neutral Endopeptidase (Neprilysin) Complexedwith Phosphoramidon. J. Mol. Biol. 2000, 296, 341-349
[29] Lacy, D. B.; Tepp, W.; Cohen, A. C.; Dasgupta, B. R.; Stevens, R.C. Crystal Structure of Botulinum Neurotoxin Type a andImplications for Toxicity. Nat. Struct. Biol. 1998, 5, 898-902.
[30] Pannifer, A. D.; Wong, T. Y.; Schwarzenbacher, R.; Renatus, M.;Petosa, C.; Bienkowska, J.; Lacy, D. B.; Collier, R. J.; Park, S.;Leppla, S. H.; Hanna, P.; Liddington, R. C. Crystal Structure of theAnthrax Lethal Factor. Nature 2001, 414, 229-233
[31] Kurisu, G.; Kinoshita, T.; Sugimoto, A.; Nagara, A.; Kai, Y.; Kasai,N.; Harada, S. Structure of the zinc endoprotease fromStreptomyces caespitosus. J. Biochem. 1997, 121, 304-308.
[32] Schlagenhauf, E.; Etges, R.; Metcalf, P. The crystal structure of theLeishmania major surface proteinase leishmanolysin (gp63).Structure 1998, 6, 1035-1046.
[33] Li, J.; Brick, P.; O'Hare, M. C.; Skarzynski, T.; Lloyd, L. F.; Curry,V. A.; Clark, I. M.; Bigg, H. F.; Hazleman, B. L.; Cawston, T. E.;Blow, D. M. Structure of full-length porcine synovial collagenasereveals a C-terminal domain containing a calcium-linked, four-bladed beta-propeller. Structure 1995, 3, 541-549.
[34] Bode, W.; Gomis-Ruth, F. X.; Huber, R.; Zwilling, R.; Stocker, W.Structure of astacin and implications for activation of astacins andzinc-ligation of collagenases. Nature 1992, 358, 164-167.
[35] Mcauley, K. E.; Jia-Xing, Y.; Dodson, E. J.; Lehmbeck, J.;Ostergaard, P. R.; Wilson, K. S. A Quick Solution: Ab InitioStructure Determination of a 19 kDa Metalloproteinase UsingAcorn Acta Crystallogr., Sect. D 2001, 57, 1571-1578.
[36] Greenblatt, H. M.; Feinberg, H.; Tucker, P. A.; Shoham, G.Carboxypeptidase A: native, zinc-removed and mercury-replacedforms. Acta Crystallogr., Sect. D. 1998, 54, 289-305.
[37] 1lbu is the pdb code for the “Refined Structure of a Zn2+ D-Alanyl-D-Alanine Carboxypeptidase of S. Albus G at 1.8 AngstromsResolution” (Wery, J.-P.; Charlier, P.; Dideberg, O.) which refers tothe original enzyme structure with pdb code 0zgp : Dideberg, O.;Charlier, P.; Dive, G.; Joris, B.; Frere, J. M.; Ghuysen, J. M.Structure of a Zn2+-containing D-alanyl-D-alanine-cleavingcarboxypeptidase at 2.5 A resolution. Nature 1982 299, 469-470.
[38] Taylor, A. B.; Smith, B. S.; Kitada, S.; Kojima, K.; Miyaura, H.;Otwinowski, Z.; Ito, A.; Deisenhofer, J. Crystal Structures ofMitochondrial Processing Peptidase Reveal the Mode for SpecificCleavage of Import Signal Sequences. Structure 2001, 9, 615-625.
[39] Burley, S. K.; David, P. R.; Taylor, A.; Lipscomb, W. N. Molecularstructure of leucine aminopeptidase at 2.7-A resolution. Proc. Natl.Acad. Sci. U.S.A. 1990, 87, 6878-6882.
[40] Roderick, S. L.; Matthews, B. W. Structure of the cobalt-dependentmethionine aminopeptidase from Escherichia coli: a new type ofproteolytic enzyme. Biochemistry 1993, 32, 3907-3912.
[41] Rowsell, S.; Pauptit, R. A.; Tucker, A. D.; Melton, R. G.; Blow, D.M.; Brick, P. Crystal structure of carboxypeptidase G2, a bacterialenzyme with applications in cancer therapy. Structure 1997, 5, 337-347.
[42] Chevrier, B.; Schalk, C.; D'Orchymont, H.; Rondeau, J. M.; Moras,D.; Tarnus, C. Crystal structure of Aeromonas proteolyticaaminopeptidase: a prototypical member of the co-catalytic zincenzyme family. Structure 1994, 2, 283-291.
[43] Pearson, M. A.; Michel, L. O.; Hausinger, R. P.; Karplus, P. A.Structures of Cys319 variants and acetohydroxamate-inhibitedKlebsiella aerogenes urease. Biochemistry 1997, 36, 8164-8172.
[44] Fritsche, E.; Paschos, A.; Beisel, H.-G.; Bock, A.; Huber, R. CrystalStructure of the Hydrogenase Maturating Endopeptidase Hybd fromEscherichia Coli. J. Mol. Biol. 1999, 288, 989-998.
[45] Laszlo, P.; Blundell, T. L. Sequences and topology. Curr. Opin.Struct. Biol. 1995, 5, 357-359
[46] Jongeneel, C. V.; Bouvier, J.; Bairoch, A. A unique signatureidentifies a family of zinc-dependent metallopeptidases. FEBS Lett.1989, 242, 211-214.
[47] Vallee, B. L.; Auld, D. S. Short and long spacer sequences andother structural features of zinc binding sites in zinc enzymes.FEBS Lett. 1989, 257, 138-140.
[48] Jiang, W.; Bond, J. S. Families of metalloendopeptidases and theirrelationships. FEBS Lett. 1992, 312, 110-114.

Structural Features of Angiotensin-I Converting Enzyme Catalytic Sites Current Topics in Medicinal Chemistry, 2004, Vol. 4, No. 4 427
[49] Bode, W.; Gomis-Rüth, F.-X.; Stöckler, W. Astacins, serralysins,snake venom and matrix metalloproteinases exhibit identical zinc-binding environments (HEXXHXXGXXH and Met-turn) andtopologies and should be grouped into a common family, the`metzincins'. FEBS Lett. 1993, 331, 134-140.
[50] Rawlings, N. D.; Barrett, A. J. Evolutionary families of peptidases.Biochem. J. 1993, 290, 205-218.
[51] Blundell, T. L. Metalloproteinase superfamilies and drug design.Nat. Struct. Biol. 1994, 1, 73-75.
[52] Bode, W.; Stöckler, W. Structural features of a superfamily of zinc-endopeptidases: the metzincins. Curr. Opin.. Struct. Biol. 1995, 5,383-390.
[53] Colman, P. M; Jansonius, J. N.; Matthews, B. W. The structure ofthermolysin : an electron density map at 2-3 A resolution. J. Mol.Biol. 1972, 70, 701-24.
[54] Matthews B.W., Weaver L.H., Kester W.R. The conformation ofthermolysin. J. Biol. Chem. 1974, 249, 8030-8044.
[55] Holmes MA, Matthews BW. Structure of thermolysin refined at 1.6A resolution. J. Mol. Biol. 1982, 160, 623-639.
[56] Baumann U. Crystal structure of the 50 kDa metallo protease fromSerratia marcescens. J. Mol. Biol. 1994, 242, 244-251.
[57] Katsuya, Y.; Hamada, K.; Hata, Y.; Tanaka, N.; Sato, M., Katsube,Y.; Kakiuchi, K.; Miyata, K. Preliminary X-ray studies on Serratiaprotease. J. Biochem. 1985, 98, 1139-1142.
[58] pdb code 1srp is linked to the reference : Hamada, K.; Hiramatsu,H.; Katsuya, Y.; Hata, Y.; Matsuura, Y.; Katsube, Y. StructuralAnalysis of Serratia Protease. Acta Crystallogr., Sect. A 1993, 49,153. However, this citation seems to be a misprint, as it does notexist to the corresponding journal. A relative article to serralysinstructure should be the following : Katsuya, Y.; Hamada, K.; Hata,Y.; Tanaka, N.; Sato, M.; Katsube, Y.; Kakiuchi, K.; Miyata, K.;Preliminary X-ray studies on Serratia protease. J. Biochem. 198598, 1139-1142.
[59] Spurlino, J. C.; Smallwood, A. M.; Carlton, D. D.; Banks, T. M.;Vavra, K. J.; Johnson, J. S.; Cook, E. R.; Falvo, J.; Wahl, R. C.;Pulvino, T. A.; Wendoloski, J. J.; Smith, D. L. 1.56 Å structure ofmature truncated human fibroblast collagenase. Proteins 1994, 19,98-109.
[60] Browner M. F.; Smith W. W.; Castelhano A. L. Matrilysin-inhibitorcomplexes : common themes among metalloproteases.Biochemistry 1995, 34, 6602-6610.
[61] Gomis-Ruth, F. X.; Kress, L. F.; Bode, W. First structure of a snakevenom metalloproteinase: a prototype for matrixmetalloproteinases/collagenases. EMBO J. 1993, 12, 4151-4157.
[62] Hori, T.; Kumasaka, T.; Yamamoto, M.; Nonaka, T.; Tanaka, N.;Hashimoto, Y.; Ueki, T.; Takio, K. Structure of a New 'Aspzincin'Metalloendopeptidase from Grifola Frondosa: Implications for theCatalytic Mechanism and Substrate Specificity Based on SeveralDifferent Crystal Forms. Acta Crystallogr., Sect. D 2001, 57, 361-368.
[63] Christianson, D. W.; David, P. R.; Lipscomb, W. N. Mechanism ofcarboxypeptidase A: hydration of a ketonic substrate analogue.Proc. Natl. Acad. Sci. U.S.A. 1987, 84, 1512-1515.
[64] Christianson, D. W.; Lipscomb, W. B. Carboxypeptidase-A. Acc.Chem. Res. 1989, 22, 62-69.
[65] Osterman, A. L.; Grishin, N. V.; Smulevitch, S. V.; Matz, M. V.;Zagnitko, O. P.; Revina, L. P.; Stepanov, V. M. Primary structure ofcarboxypeptidase T: Delineation of functionally relevant features inZn-carboxypeptidase family. J. Protein Chem. 1992, 11, 561-570.
[66] Lipscomb, W. N.; Sträter N. Recent Advances in Zinc Enzymology.Chem. Rev. 1996, 96, 2375-2433.
[67] Makarova, K. S.; Grishin, N. V. The Zn-peptidase superfamily:functional convergence after evolutionary divergence. J. Mol. Biol.1999, 292, 11-17.
[68] Greenblatt, H. M.; Feinberg, H.; Tucker, P. A.; Shoham, G.Carboxypeptidase A: native, zinc-removed and mercury-replacedforms. Acta Crystallogr., Sect. D 1998, 54, 289-305.
[69] Blundell T. L. Metalloproteinase superfamilies and drug design.Nat. Struct. Biol. 1992, 2, 73-75.
[70] Cushman, D. W.; Pluscec, J.; Williams, N. J.; Weaver, E. R.; Sabo,E. F.; Kocy, O.; Cheung, H. S.; Ondetti, M. A. Inhibition ofangiotensin-coverting enzyme by analogs of peptides fromBothrops jararaca venom. Experientia 1973, 29, 1032-1035.
[71] Elisseeva, Y. E.; Orekhovich, V. N.; Pavlikhina, L. V.; Alexeenko,L. P. Carboxycathepsin--a key regulatory component of two
physiological systems involved in regulation of blood pressure.Clin. Chim. Acta. 1971, 31, 413-419.
[72] Ondetti, M. A.; Rubin, B.; Cushman, D. W. Design of specificinhibitors of angiotensin-converting enzyme: new class of orallyactive antihypertensive agents. Science 1977, 196, 441-444.
[73] Quiocho, F. A.; Lipscombe, W. N. Carboxypeptidase A: a proteinand an enzyme. Adv. Protein Chem. 1971, 25, 1-78.
[74] Pelmenschikov, V.; Blomberg, M. R. A.; Siegbahn, P. E. M. Atheoretical study of the mechanism for peptide hydrolysis bythermolysin. J. Biol. Inorg. Chem. 2002, 7, 284-298.
[75] Kester, W. R.; Matthews, B. W. Crystallographic study of thebinding of dipeptide inhibitors to thermolysin: implications for themechanism of catalysis. Biochemistry 1977, 16, 2506-2516.
[76] Holmes, M. A.; Matthews, B. W. Binding of hydroxamic acidinhibitors to crystalline thermolysin suggests a pentacoordinate zincintermediate in catalysis. Biochemistry 1981, 20, 6912-6920.
[77] Hangauer, D. G.; Monzingo A. F.; Matthews, B. W. An interactivecomputer graphics study of thermolysin-catalyzed peptide cleavageand inhibition by N-carboxymethyl dipeptides. Biochemistry 1984,23, 5730-5741.
[78] Holden, H. M.; Tronrud, D. E.; Monzingo, A. F.; Weaver, L. H.;Matthews, B. W. Slow- and fast-binding inhibitors of thermolysindisplay different modes of binding: crystallographic analysis ofextended phosphonamidate transition-state analogues. Biochemistry1987, 26, 8542-8553.
[79] Tronrud, D. E.; Holden, H. M.; Matthews, B. W. Structures of twothermolysin-inhibitor complexes that differ by a single hydrogenbond. Science 1987, 30, 571-574.
[80] Matthews, B. W. Structural Basis of the Action of Thermolysin andRelated Zinc Peptidases. Acc. Chem. Res. 1988, 21, 333-340.
[81] Jaspard, E.; Wei, L.; Alhenc-Gelas, F. Differences in the propertiesand enzymatic specificities of the two active sites of angiotensin I-converting enzyme (kininase II). Studies with bradykinin and othernatural peptides. J. Biol. Chem. 1993, 268, 9496-9503.
[82] Erdös, E. G.; Deddish, P. A.; Marcic, B. M. Potentiation ofBradykinin Actions by ACE Inhibitors. Trends Endocrinol. Metab.1999, 10, 223-229.
[83] Skidgel, R. A.; Erdos, E. G. Novel activity of human angiotensin Iconverting enzyme: release of the NH2- and COOH-terminaltripeptides from the luteinizing hormone-releasing hormone. Proc.Natl. Acad. Sci. U. S. A. 1985, 82, 1025-1029.
[84] Ehlers, M. R.; Riordan, J. F. Angiotensin-converting enzyme: zinc-and inhibitor-binding stoichiometries of the somatic and testisisozymes. Biochemistry 1991, 30, 7118-7126.
[85] Casarini, D. E.; Plavinik, F. L.; Zanella, M. T.; Marson, O.;Krieger, J. E.; Hirata, I.Y.; Stella, R. C. Angiotensin convertingenzymes from human urine of mild hypertensive untreated patientsresemble the N-terminal fragment of human angiotensin I-converting enzyme. Int. J. Biochem. Cell Biol. 2001, 33, 75-85.
[86] Deddish, P. A.; Marcic, B.; Jackman, H. L.; Wang, H. Z.; Skidgel,R. A.; Erdös, E. G. N-domain-specific substrate and C-domaininhibitors of angiotensin-converting enzyme: angiotensin-(1-7) andketo-ACE. Hypertension 1998, 31, 912-917.
[87] Dive, V.; Cotton, J.; Yiotakis, A.; Michaud, A.; Vassiliou, S.;Jiracek, J.; Vazeux, G.; Chauvet, M. T.; Cuniasse, P.; Corvol, P.RXP 407, a phosphinic peptide, is a potent inhibitor of angiotensin Iconverting enzyme able to differentiate between its two active sites.Proc. Natl. Acad. Sci. U. S. A. 1999, 96, 4330-4335.
[88] Junot, C.; Gonzales, M. F.; Ezan, E.; Cotton, J.; Vazeux, G.;Michaud, A.; Azizi, M.; Vassiliou, S.; Yiotakis, A.; Corvol, P.;Dive, V. RXP 407, a selective inhibitor of the N-domain ofangiotensin I-converting enzyme, blocks in vivo the degradation ofhemoregulatory peptide acetyl-Ser-Asp-Lys-Pro with no effect onangiotensin I hydrolysis. J. Pharmacol. Exp. Ther. 2001, 297, 606-611.
[89] Corvol, P.; Williams, T. A. In Handbook of Proteolytic Enzymes;Barrett, A. J.; Rawlings, N. D.; Woessner, J. F., Eds.; AcademicPress: New York, 1998; pp 1066-1076.
[90] Shapiro, R.; Holmquist, B.; Riordan, J. F. Anion activation ofangiotensin converting enzyme: dependence on nature of substrate.Biochemistry 1983, 22, 3850-3857.
[91] Lamango, N. S.; Sajid, M.; Isaac, R. E. The endopeptidase activityand the activation by Cl- of angiotensin-converting enzyme isevolutionarily conserved: purification and properties of an

428 Current Topics in Medicinal Chemistry, 2004, Vol. 4, No. 4 Spyroulias et al.
angiotensin-converting enzyme from the housefly, Muscadomestica. Biochem. J. 1996, 314, 639-646.
[92] Wei, L.; Alhenc-Gelas, F.; Corvol, P.; Clauser, E. The twoHomologous Domais of Human Angiotensin I-converting EnzymeAre Both Catalically Active. J. Biol. Chem. 1991, 266, 9002-9006.
[93] Kester, W. R.; Matthews, B. W. Comparison of the structures ofcarboxypeptidase A and thermolysin. J. Biol. Chem. 1977, 252,7704-7710.
[94] Mayer, M.; Meyer, B. Mapping the active site of angiotensin-converting enzyme by Transferred NOE Spectroscopy. J. Med.Chem 2000, 43, 2093-2097.
[95] Bünning, P.; Holmquist, B.; Riordan, J. F. Substrate specificity andkinetic characteristics of angiotensin converting enzyme.Biochemistry 1983, 22, 103-110.
[96] Patchett, A. A.; Cordes, E. H. The design and properties of N-carboxyalkyldipeptide inhibitors of angiotensin-converting enzyme.Adv. Enzymol. Relat. Areas Mol. Biol. 1985, 57, 1-84.
[97] Patchett, A. A.; Harris, E.; Tristram, E. W.; Wyvratt, M. J.; Wu, M.T.; Taub, D. Peterson, E. R.; Ikeler, T.; ten Broeke, J.; Payne, L. G.;Ondeyka, D. L.; Thorsett, E. D.; Greenlee, W. J.; Lohr, N. S.;Hoffsommer, R. D.; Joshua, H.; Ruyle, W. V.; Rothrock, J. W.;Aster, S. D.; Maycock, A. L.; Robinson, F. M.; Hirschmann, R.;Sweet, C. S.; Ulm, E. H.; Gross, D. M.; Vassil, T. C.; Stone, C. A.A new class of angiotensin-converting enzyme inhibitors. Nature1980, 288, 280-283.
[98] Moore, V. A.; Irwin, W. J.; Timmins, P.; Chong, S.; Dando, S. A.;Morrison, R. A. A rapid screening system to determine drugaffinities for the intestinal dipeptide transporter 1 : systemcharacterisation. Inter. J. Pharm. 2000, 210, 15-27.
[99] Moore, V. A.; Irwin, W. J.; Timmins, P.; Chong, S.; Dando, S. A.;Morrison, R. A. A rapid screening system to determine drugaffinities for the intestinal dipeptide transporter 2: affinities of ACEinhibitors. Inter. J. Pharm. 2000, 210, 29-44.
[100] Cushman, D. W.; Cheung, H. S.; Sabo, E. F.; Ondetti, M. A. Designof potent competitive inhibitors of angiotensin-converting enzyme.Carboxyalkanoyl and mercaptoalkanoyl amino acids Biochemistry1977, 16, 5484-5491.
[101] Petrillo, E. W.; Ondetti, M. A. Angiotensin-converting enzymeinhibitors: medicinal chemistry and biological actions. Med. Res.Rev. 1982, 2, 1-41.
[102] Cushman, D. W.; Cheung, H. S.; Sabo, E. F.; Ondetti, M. A.Development and design of specific inhibitors of angiotensin-converting enzyme. Am. J. Cardiol. 1982, 49, 1390-1394.
[103] Wu, M. T.; Douglas, A. W.; Ondeyka, D. L.; Payne, L. G.; Ikeler,T. J. Joshua, H.; Patchett, A. A.; Synthesis of N2-[(S)-1-carboxy-3-phenylpropyl]-L-lysyl-L-proline (lisinopril). J. Pharm. Sci. 1985,74, 352-354.
[104] Krapcho, J.; Turk, C.; Cushman, D. W.; Powell, J. R.; DeForrest, J.M.; Spitzmiller, E. R.; Karanewsky, D. S.; Duggan, M.; Rovnsak,G.; Schwartz, J.; Natarajan, S.; Godfrey, J. D.; Ryono, D. E.;Neubeck, R.; Atwal, K. S.; Petrillo, E. W. Jr. Angiotensin-converting enzyme inhibitors. Mercaptan, carboxyalkyl dipeptide,and phosphinic acid inhibitors incorporating 4-substituted prolines.J. Med. Chem. 1988, 31, 1148-1160.
[105] Michaud, A.; Chauvet, M. T.; Corvol, P. N-domain selectivity ofangiotensin I-converting enzyme as assessed by structure-functionstudies of its highly selective substrate, N-acetyl-seryl-aspartyl-lysyl-proline. Biochem. Pharmacol. 1999, 57, 611-618.
[106] Araujo, M. C.; Melo, R. L.; Cesari, M. H.; Juliano, M. A.; Juliano,L.; Carmona, A. K. Peptidase specificity characterization of C- andN-terminal catalytic sites of angiotensin I-converting enzyme.Biochemistry 2000, 39, 8519-8525.
[107] Ehlers, M. R. W.; Kirsch, R. E. Catalysis of Angiotensin IHydrolysis by Human Angiotensin-Converting Enzyme : Effect ofChloride and pH. Biochemistry, 1988, 27, 5538-5544.
[108] Liu, X.; Fernandez, M.; Wouters, M. A.; Heyberger, S.; Husain, A.Arg(1098) is critical for the chloride dependence of humanangiotensin I-converting enzyme C-domain catalytic activity. J.Biol. Chem. 2001, 276, 33518-33525.
[109] Bünning, P.; Holmquist, B.; Riordan, J. F. Functional residues atthe active site of angiotensin converting enzyme. Biochem. Biophys.Res. Commun. 1978, 83, 1442-1449.
[110] Shapiro, R.; Riordan, J. F. Critical Lysine Residue at the ChlorideBinding Site of Angiotensin Converting Enzyme. Biochemistry1983, 22, 5315-5321.
[111] Fernandez, M.; Liu, X.; Wouters, M. A.; Heyberger, S.; Husain, A.Angiotensin I-converting enzyme transition state stabilization byHis1089: evidence for a catalytic mechanism distinct from othergluzincin metalloproteinases. J. Biol. Chem. 2001, 276, 4998-5004.
[112] Chen, Y. N.; Ehlers, M. R.; Riordan, J. F. The functional role oftyrosine-200 in human testis angiotensin-converting enzyme.Biochem. Biophys. Res. Commun. 1992, 184, 306-309.
[113] Sen, I.; Kasturi, S.; Abdul Jabbar, M.; Sen, G. C. Mutations in twospecific residues of testicular angiotensin-converting enzymechange its catalytic properties. J. Biol. Chem. 1993, 268, 25748-25754.
[114] Thompson, J. D.; Gibson, T. J.; Plewniak, F.; Jeanmougin, F.;Higgins, D. G. The CLUSTAL_X windows interface: flexiblestrategies for multiple sequence alignment aided by quality analysistools. Nucleic Acids Res. 1997, 25, 4876-4882.
[115] Martí-Renom, M. A.; Stuart, A. C.; Fiser, A.; Sánchez, R.; Melo,F.; ali, A. Comparative protein structure modeling of genes andgenomes. Annu. Rev. Biophys. Biomol. Struct. 2000, 29, 291-325.
[116] Combet, C.; Blanchet, C.; Geourjon, C.; Deléage, G. NPS@:Network Protein Sequence Analysis. TIBS 2000, 25, 147-150.
[117] Garnier, J.; Osguthorpe, D. J.; Robson, B. Analysis of the accuracyand implications of simple methods for predicting the secondarystructure of globular proteins. J. Mol. Biol. 1978, 120, 97-120.
[118] Gibrat, J. F.; Garnier, J.; Robson, B. Further developments ofprotein secondary structure prediction using information theory.New parameters and consideration of residue pairs. J. Mol. Biol.1987, 198, 425-443.
[119] Garnier, J.; Gibrat, J. F.; Robson, B. GOR method for predictingprotein secondary structure from amino acid sequence. MethodsEnzymol. 1996, 266, 540-553.
[120] Merrifield R. B. Solid Phase Synthesis. I. The synthesis of aTetrapeptide. J. Am. Chem. Soc. 1963, 85, 2149-2154.
[121] Veglia, G.; Porcelli, F.; DeSilva, T.; Prantner, A.; Opella, S.J. Thestructure of the metal-binding motiff GMTCAAC is similar in an18-residue linear peptide and the mercury binding protein Merp. J.Am. Chem. Soc. 2000, 122, 2389-2390.
[122] Opella, S. J.; DeSilva, T. M.; Veglia, G. Structural biology ofmetal-binding sequence. Curr. Opin. Chem. Biol. 2002, 6, 217-223.
[123] DeSilva, T. M.; Veglia, T.; Porcelli, F.; Prantner, A. M.; Opella, S.J. Selectivity in heavy metal-binding to peptides and proteins.Biopolymers 2002, 64, 189-197.
[124] Carpino, L. A.; Han, G. Y.; 9-Fluorenylmethoxycarbonyl function,a new base-sensitive amino-protecting group J. Am. Chem. Soc.1970, 92, 5748-5749.
[125] Barlos, K.; Gatos, D.; Kallitsis, J.; Papaphotiou, G.; Sotiriou, P.;Wenquing, Y.; Shafer, W. Darstellung geshützter peptid-fragmenteunter einsatz substituierter triphenylmethyl-harze, Tetraedron Lett.1989, 30, 3943-3946.
[126] Papazacharias, S.; Pairas, G.; Kouimtzoglou, E.; Dermitzaki, E.;Manessi-Zoupa, E.; Gravanis, A.; Cordopatis, P. Synthesis andBiological Evaluation of New h/rCRH Peptide Analogs. In Peptides2000 ; Martinez, J.; Fehrentz, J.-A., Eds, EDK, 2001, pp. 933-934.
[127] King, D. S.; Fields, C. G.; Fields G. B. A cleavage method whichminimizes side reactions following Fmoc solid phase peptidesynthesis. Int. J. Pept. Protein Res. 1990, 36, 255-266
[128] Braunschweiler, L.; Ernst, R. R. Coherence transfer by isotropicmixing : application to proton correlation spectroscopy. J. Magn.Reson. 1983, 53, 521-528.
[129] Bax, A.; Davis, D. G. MLEV-17-based two-dimensionalhomonuclear magnetization transfer spectroscopy. J. Magn. Reson.1985, 65, 355-360.
[130] Marion, D.; Wüthrich, K. Application of phase sensitive two-dimensional correlated spectroscopy (COSY) for measurements of1H-1H spin-spin coupling constants in proteins. Biochem. Biophys.Res. Commun. 1983, 113, 967-974.
[131] Jeener, J., Meier, B. H., Bachmann, P.; Ernst R.R. Investigation ofexchange processes by two-dimensional NMR spectroscopy. J.Chem. Phys. 1979, 71, 4546-4553.
[132] Wüthrich, K. In NMR of Proteins and Nucleic Acids; Wiley: NewYork, 1986.
[133] Wang, Y.; Nip, A.M.; Wishart D. S. A simple method toquantitatively measure polypeptide JHNHα coupling constants fromTOCSY or NOESY spectra. J. Biomol. NMR 1997, 10, 373-382.
[134] Smith, L. J.; Bolin, K. A.; Schwalbe, H.; MacArthur, M. W.;Thorton, J. M.; Dobson, C. M. Analysis of main chain torsion
S
>

Structural Features of Angiotensin-I Converting Enzyme Catalytic Sites Current Topics in Medicinal Chemistry, 2004, Vol. 4, No. 4 429
angles in proteins: prediction of NMR coupling constants for nativeand random coil conformations. J. Mol. Biol. 1996, 255, 494-506.
[135] Wishart, D.; Sykes, B.; Richards, F. Relationship between nuclearmagnetic resonance chemical shift and protein secondary structure.J. Mol. Biol. 1991, 222, 311-333.
[136] Wishart, D.; Sykes, B.; Richards, F. The chemical shift index: a fastand simple method for the assignment of protein secondarystructure through NMR spectroscopy. Biochemistry 1992, 31, 1647-1651.
[137] Merutka, G.; Dyson, H. J.; Wright, P. E. 'Random coil' 1H chemicalshifts obtained as a function of temperature and trifluoroethanolconcentration for the peptide series GGXGG. J. Biomol. NMR1995, 5, 14-24.
[138] Galanis, A. S.; Spyroulias, G. A.; Pairas, G.; Manessi-Zoupa, E.;Cordopatis, P. NMR solution structures of apoform ACEN andACEC active sites model peptides. In preparation.
[139] Güntert, P.; Braun, W.; Wüthrich, K. Efficient computation ofthree-dimensional protein structures in solution from nuclearmagnetic resonance data using the program DIANA and thesupporting programs CALIBA, HABAS and GLOMSA. J. Mol.Biol. 1991, 217, 517-530.
[140] Güntert, P.; Mumenthaler, C.; Wüthrich, K. Torsion angle dynamicsfor NMR structure calculation with the new program DYANA. J.Mol. Biol. 1997, 273, 283-298.
[141] Pearlman, D. A.; Case, D. A.; Caldwell, J. W.; Ross, W. S.;Cheatham, T. E.; Ferguson, D. M.; Seibel, G. L.; Singh, U. C.;Weiner, P. K.; Kollman, P. A. AMBER 5.0. University ofCalifornia, San Francisco, 1997.
[142] Pearlman, D. A.; Case, D. A.; Caldwell, G. C.; Siebel, G. L.; Singh,U. C.; Weiner, P.; Kollman, P. A. AMBER 4.0. University ofCalifornia, San Francisco, 1991.
[143] Shortle, D.; Clarke, N. Alpha helix propensity of amino acids.Science 1993, 260, 1637-1640.
[144] Shortle, D.; Clarke, N. Alpha helix propensity of amino acids.Science 1993, 262, 917-918.
[145] Zuiderweg, E. R. P. Mapping Protein-Protein Interactions inSolution by NMR Spectroscopy. Biochemistry, 2002, 41, 1-7.
[146] Galanis, A. S.; Spyroulias, G. A.; Pierattelli, R.; Tzakos, A.;Troganis, A.; Gerothanassis, I. P.; Pairas, G.; Manessi-Zoupa, E.;Cordopatis, P. Zinc-Binding in Peptide Models of ACE Active SitesStudied Through 1H NMR & Chemical Shift Perturbation Mapping.Biopolymers, 2003, 69, 244-252.
[147] Koradi, R.; Billeter, M.; Wüthrich, K. MOLMOL: a program fordisplay and analysis of macromolecular structures. J. Mol. Graph.1996, 14, 51-55.
[148] Natesh, R.; Schwager, S. L. U.; Sturrock, E. D.; Acharya, K. R.Crystal structure of the human angiotensin-converting enzyme-lisinopril complex. Nature 2003, 421, 551-554
[149] Kim, H. M.; Shin, D. R.; Yoo, O. J.; Lee, H.; Lee, J. O. Crystalstructure of Drosophila angiotensin I-converting enzyme bound tocaptopril and lisinopril. FEBS Lett. 2003, 538, 65-70
[150] Arndt, J. W.; Hao, B.; Ramakrishnan, V.; Cheng, T.; Chan, S. I.;Chan, M. K. Crystal structure of a novel carboxypeptidase from thehyperthermophilic archaeon Pyrococcus furiosus. Structure 2002,215-224.



















