
Coupling of synaptic inputs to local cortical activity...
15
RESEARCH ARTICLE Sensory Processing Coupling of synaptic inputs to local cortical activity differs among neurons and adapts after stimulus onset Nathaniel C. Wright, Mahmood S. Hoseini, Tansel Baran Yasar, and Ralf Wessel Department of Physics, Washington University in St. Louis, St. Louis, Missouri Submitted 30 May 2017; accepted in final form 19 September 2017 Wright NC, Hoseini MS, Yasar TB, Wessel R. Coupling of synaptic inputs to local cortical activity differs among neurons and adapts after stimulus onset. J Neurophysiol 118: 3345–3359, 2017. First published September 20, 2017; doi:10.1152/jn.00398.2017.— Cortical activity contributes significantly to the high variability of sensory responses of interconnected pyramidal neurons, which has crucial implications for sensory coding. Yet, largely because of technical limitations of in vivo intracellular recordings, the coupling of a pyramidal neuron’s synaptic inputs to the local cortical activity has evaded full understanding. Here we obtained excitatory synaptic conductance (g) measurements from putative pyramidal neurons and local field potential (LFP) recordings from adjacent cortical circuits during visual processing in the turtle whole brain ex vivo preparation. We found a range of g-LFP coupling across neurons. Importantly, for a given neuron, g-LFP coupling increased at stimulus onset and then relaxed toward intermediate values during continued visual stimula- tion. A model network with clustered connectivity and synaptic depression reproduced both the diversity and the dynamics of g-LFP coupling. In conclusion, these results establish a rich dependence of single-neuron responses on anatomical, synaptic, and emergent net- work properties. NEW & NOTEWORTHY Cortical neurons are strongly influenced by the networks in which they are embedded. To understand sensory processing, we must identify the nature of this influence and its underlying mechanisms. Here we investigate synaptic inputs to cor- tical neurons, and the nearby local field potential, during visual processing. We find a range of neuron-to-network coupling across cortical neurons. This coupling is dynamically modulated during visual processing via biophysical and emergent network properties. cortex; population coupling; synaptic inputs; local field potential; response variability; correlated variability INTRODUCTION Cortical neuron sensory responses are remarkably variable across trials (Britten et al. 1993; Carandini 2004; Schölvinck et al. 2015). With advances in recording techniques, it has be- come increasingly obvious that single-neuron response vari- ability reflects fluctuations that are shared across large regions of cortex (Lin et al. 2015; Okun et al. 2015; Rabinowitz et al. 2015; Schölvinck et al. 2015). That is, sensory input interacts with intrinsic cortical activity, with global cortical fluctuations influencing single-neuron responses. Appropriately, a recent study has introduced the term “population coupling” to de- scribe this relationship (Okun et al. 2015). This and other studies have shown that population coupling in cortex is remarkably diverse across neurons [likely reflecting connec- tivity (Okun et al. 2015; Pernice et al. 2011)] yet can also change with sensory stimulation (Haider et al. 2016; Tan et al. 2014) and network state (Haider et al. 2016; Lin et al. 2015; Okun et al. 2015). This rich dependence of single-neuron responses on anatomical and emergent network properties appears to represent a fundamental principle of cortical func- tion and is only beginning to be explored. Here we investigate three questions vital to a better understanding of cortical variability and its effects on sensory coding. 1) What is the nature of response variability in cortical microcircuits? 2) How strongly are single-neuron synaptic input fluctuations coupled with those of the local population? 3) To what degree are the dynamics of response variability and population coupling de- termined by the cortical network, and what are the relevant network parameters? While the spike-based approach has yielded many important insights, it has two inherent shortcomings. First, it excludes the vast majority of neurons, which are sparse spiking (Henze et al. 2000; O’Connor et al. 2010; Shoham et al. 2006; Thompson and Best 1989) and therefore yield unreliable statistics for the analysis of correlated variability (Cohen and Kohn 2011) (Fig. 1, A and B). Second, it involves sampling populations of neurons that are visible to the experimentalist but may not represent relevant or complete cortical microcircuits. Patch- clamp recordings represent one solution to these two problems (Shoham et al. 2006). First, the subthreshold inputs to a neuron provide a measure of activity that is agnostic to output spike rate. A second perspective, motivated by anatomical connec- tivity, recognizes the neuron as a network sampling “device” that allows the experimenter to tap into the cortical circuitry itself and infer response properties (e.g., variability) of large populations of neurons (Ikegaya et al. 2004; MacLean et al. 2005; Mokeichev et al. 2007) (Fig. 1, A and B). Yet this approach is relatively rare; it is difficult to obtain stable patch-clamp recordings of cortical sensory responses in vivo, and spatially extended cortical pyramidal neurons confound the interpretation of voltage-clamp data (Armstrong and Gilly 1992; Koch 2004; Spruston et al. 1993). Here we overcome these challenges to address the first two questions above. We recorded subthreshold membrane potential visual responses from cortical putative pyramidal neurons in the turtle eye- Address for reprint requests and other correspondence: N. C. Wright, Georgia Inst. of Technology, Dept. of Biomedical Engineering, 313 Ferst Dr., Rm. 2127, Atlanta, GA 30313 (e-mail: [email protected]). J Neurophysiol 118: 3345–3359, 2017. First published September 20, 2017; doi:10.1152/jn.00398.2017. 3345 0022-3077/17 Copyright © 2017 the American Physiological Society www.jn.org Downloaded from www.physiology.org/journal/jn by ${individualUser.givenNames} ${individualUser.surname} (128.252.035.199) on December 13, 2017. Copyright © 2017 American Physiological Society. All rights reserved.
-
Upload
hoanghuong -
Category
Documents
-
view
224 -
download
0
Transcript of Coupling of synaptic inputs to local cortical activity...

RESEARCH ARTICLE Sensory Processing
Coupling of synaptic inputs to local cortical activity differs among neuronsand adapts after stimulus onset
Nathaniel C. Wright, Mahmood S. Hoseini, Tansel Baran Yasar, and Ralf WesselDepartment of Physics, Washington University in St. Louis, St. Louis, Missouri
Submitted 30 May 2017; accepted in final form 19 September 2017
Wright NC, Hoseini MS, Yasar TB, Wessel R. Coupling ofsynaptic inputs to local cortical activity differs among neurons andadapts after stimulus onset. J Neurophysiol 118: 3345–3359, 2017.First published September 20, 2017; doi:10.1152/jn.00398.2017.—Cortical activity contributes significantly to the high variability ofsensory responses of interconnected pyramidal neurons, which hascrucial implications for sensory coding. Yet, largely because oftechnical limitations of in vivo intracellular recordings, the couplingof a pyramidal neuron’s synaptic inputs to the local cortical activityhas evaded full understanding. Here we obtained excitatory synapticconductance (g) measurements from putative pyramidal neurons andlocal field potential (LFP) recordings from adjacent cortical circuitsduring visual processing in the turtle whole brain ex vivo preparation.We found a range of g-LFP coupling across neurons. Importantly, fora given neuron, g-LFP coupling increased at stimulus onset and thenrelaxed toward intermediate values during continued visual stimula-tion. A model network with clustered connectivity and synapticdepression reproduced both the diversity and the dynamics of g-LFPcoupling. In conclusion, these results establish a rich dependence ofsingle-neuron responses on anatomical, synaptic, and emergent net-work properties.
NEW & NOTEWORTHY Cortical neurons are strongly influencedby the networks in which they are embedded. To understand sensoryprocessing, we must identify the nature of this influence and itsunderlying mechanisms. Here we investigate synaptic inputs to cor-tical neurons, and the nearby local field potential, during visualprocessing. We find a range of neuron-to-network coupling acrosscortical neurons. This coupling is dynamically modulated duringvisual processing via biophysical and emergent network properties.
cortex; population coupling; synaptic inputs; local field potential;response variability; correlated variability
INTRODUCTION
Cortical neuron sensory responses are remarkably variableacross trials (Britten et al. 1993; Carandini 2004; Schölvinck etal. 2015). With advances in recording techniques, it has be-come increasingly obvious that single-neuron response vari-ability reflects fluctuations that are shared across large regionsof cortex (Lin et al. 2015; Okun et al. 2015; Rabinowitz et al.2015; Schölvinck et al. 2015). That is, sensory input interactswith intrinsic cortical activity, with global cortical fluctuationsinfluencing single-neuron responses. Appropriately, a recent
study has introduced the term “population coupling” to de-scribe this relationship (Okun et al. 2015). This and otherstudies have shown that population coupling in cortex isremarkably diverse across neurons [likely reflecting connec-tivity (Okun et al. 2015; Pernice et al. 2011)] yet can alsochange with sensory stimulation (Haider et al. 2016; Tan et al.2014) and network state (Haider et al. 2016; Lin et al. 2015;Okun et al. 2015). This rich dependence of single-neuronresponses on anatomical and emergent network propertiesappears to represent a fundamental principle of cortical func-tion and is only beginning to be explored. Here we investigatethree questions vital to a better understanding of corticalvariability and its effects on sensory coding. 1) What is thenature of response variability in cortical microcircuits? 2) Howstrongly are single-neuron synaptic input fluctuations coupledwith those of the local population? 3) To what degree are thedynamics of response variability and population coupling de-termined by the cortical network, and what are the relevantnetwork parameters?
While the spike-based approach has yielded many importantinsights, it has two inherent shortcomings. First, it excludes thevast majority of neurons, which are sparse spiking (Henze et al.2000; O’Connor et al. 2010; Shoham et al. 2006; Thompsonand Best 1989) and therefore yield unreliable statistics for theanalysis of correlated variability (Cohen and Kohn 2011) (Fig.1, A and B). Second, it involves sampling populations ofneurons that are visible to the experimentalist but may notrepresent relevant or complete cortical microcircuits. Patch-clamp recordings represent one solution to these two problems(Shoham et al. 2006). First, the subthreshold inputs to a neuronprovide a measure of activity that is agnostic to output spikerate. A second perspective, motivated by anatomical connec-tivity, recognizes the neuron as a network sampling “device”that allows the experimenter to tap into the cortical circuitryitself and infer response properties (e.g., variability) of largepopulations of neurons (Ikegaya et al. 2004; MacLean et al.2005; Mokeichev et al. 2007) (Fig. 1, A and B). Yet thisapproach is relatively rare; it is difficult to obtain stablepatch-clamp recordings of cortical sensory responses in vivo,and spatially extended cortical pyramidal neurons confound theinterpretation of voltage-clamp data (Armstrong and Gilly1992; Koch 2004; Spruston et al. 1993). Here we overcomethese challenges to address the first two questions above. Werecorded subthreshold membrane potential visual responsesfrom cortical putative pyramidal neurons in the turtle eye-
Address for reprint requests and other correspondence: N. C. Wright,Georgia Inst. of Technology, Dept. of Biomedical Engineering, 313 Ferst Dr.,Rm. 2127, Atlanta, GA 30313 (e-mail: [email protected]).
J Neurophysiol 118: 3345–3359, 2017.First published September 20, 2017; doi:10.1152/jn.00398.2017.
33450022-3077/17 Copyright © 2017 the American Physiological Societywww.jn.org
Downloaded from www.physiology.org/journal/jn by ${individualUser.givenNames} ${individualUser.surname} (128.252.035.199) on December 13, 2017.Copyright © 2017 American Physiological Society. All rights reserved.

attached whole brain ex vivo preparation (Fig. 1C) and used arecently developed algorithm to estimate the excitatory synap-tic conductance (g) from membrane potential (Yasar et al.2016) (Fig. 1C). This inferred conductance provided a mem-brane potential-independent measure of spiking activity in thepool of excitatory presynaptic neurons. We then quantified theresponse variability in g and calculated population coupling,i.e., the correlated variability for g and the nearby local fieldpotential (LFP). We found that visual stimulation evokedsignificant increases in g and LFP variability, which waspredominantly additive in nature. Across the population ofcells, g-LFP correlated variability (CC) was highly variableand transiently increased with visual stimulation.
We addressed the third question by implementing a drivennetwork of leaky integrate-and-fire neurons with clusteredconnectivity. This model qualitatively reproduced the empiri-cal results and suggests a dependence on spatial clustering,network spike rate oscillations, and synaptic adaptation.
Together, our results provide a clearer picture of the sub-threshold dynamics underlying suprathreshold response vari-ability and population coupling in cortex. Moreover, they
implicate specific anatomical and emergent network propertiesthat shape cortical variability and population coupling duringsensory processing.
MATERIALS AND METHODS
Surgery. All procedures were approved by Washington Univer-sity’s Institutional Animal Care and Use Committees and conform tothe National Institutes of Health Guide for the Care and Use ofLaboratory Animals. Fourteen adult red-eared sliders (Trachemysscripta elegans, 150–1,000 g) were used for this study. Turtles wereanesthetized with propofol (2 mg/kg) and then decapitated. Dissectionproceeded as described previously (Crockett et al. 2015; Saha et al.2011; Shew et al. 2015). In brief, immediately after decapitation, thebrain was excised from the skull with right eye intact and bathed incold extracellular saline (in mM: 85 NaCl, 2 KCl, 2 MgCl2·6H2O, 20dextrose, 3 CaCl2·2H2O, 45 NaHCO3). The dura was removed fromthe left cortex and right optic nerve and the right eye hemisected toexpose the retina. The rostral tip of the olfactory bulb was removed,exposing the ventricle that spans the olfactory bulb and cortex. A cutwas made along the midline from the rostral end of the remainingolfactory bulb to the caudal end of the cortex. The preparation wasthen transferred to a perfusing chamber (Warner RC-27LD recording
Fig. 1. Individual neurons subsample the cortex, and the membrane potential provides a spike rate-independent measure of cortical sensory responses. A: corticalneurons are primarily sparse-spiking units (low-opacity circles), and each neuron subsamples the cortex by receiving synaptic inputs from a large, biologicallyrelevant subpopulation. B: while high spike-rate neurons (high-opacity rasters) alone provide reliable statistics for analysis of spiking responses, the subthresholdactivity of a randomly selected neuron (e.g., red voltage trace corresponding to red rasters) contains information about the time course of presynaptic spikingactivity. C, left: we recorded the membrane potentials (V) of cortical putative pyramidal neurons as well as the nearby local field potential (LFP) while presentingmovies to the retina in the turtle eye-attached whole brain ex vivo preparation. Right: we used an algorithm (see MATERIALS AND METHODS) to infer the excitatorysynaptic conductance (g) from V, which gave a more detailed view of synaptic activity (inset). We investigated the nature of the variability in g and its couplingwith that of the simultaneously recorded LFP.
3346 VARIABILITY AND ADAPTATION OF SYNAPTIC INPUT COUPLING
J Neurophysiol • doi:10.1152/jn.00398.2017 • www.jn.org
Downloaded from www.physiology.org/journal/jn by ${individualUser.givenNames} ${individualUser.surname} (128.252.035.199) on December 13, 2017.Copyright © 2017 American Physiological Society. All rights reserved.

chamber mounted to PM-7D platform) and placed directly on a glasscoverslip surrounded by Sylgard. A final cut was made to the cortex(orthogonal to the previous and stopping short of the border betweenmedial and lateral cortex), allowing the cortex to be pinned flat, withventricular surface exposed. Multiple perfusion lines delivered extra-cellular saline, adjusted to pH 7.4 at room temperature, to the brainand retina in the recording chamber.
Intracellular recordings. We performed whole cell current-clamprecordings from 39 cells in 14 preparations. In some cases, werecorded simultaneously from pairs of nearby neurons. Patch pipettes(4–8 M�) were pulled from borosilicate glass and filled with astandard electrode solution (in mM: 124 KMeSO4, 2.3 CaCl2·2H2O,1.2 MgCl2, 10 HEPES, 5 EGTA) adjusted to pH 7.4 at roomtemperature. The visual cortex was identified as described previously(Shew et al. 2015). Cells were targeted for patching with a differentialinterference contrast microscope (Olympus). The pipette tip wasadvanced through the ependymal surface and subcellular layer andinto the cellular layer [which is composed primarily of pyramidalneurons (Connors and Kriegstein 1986)]. All recorded cells werelocated in the cellular layer, within ~150 �m of the subcellular layer.Of the 39 recorded cells, 21 were located within 300 �m of anextracellular recording electrode (also positioned in the cellular layer).Intracellular activity was collected with an Axoclamp 900A amplifier,digitized by a data acquisition panel (National Instruments PCIe-6321), and recorded with a custom LabVIEW program (National Instru-ments), sampling at 10 kHz. We used stepwise current injection toevoke action potentials in patched neurons and distinguished putativepyramidal cells from putative nonpyramidal cells on the basis ofvisual inspection of spike width, afterhyperpolarization, spike-heightadaptation, and interspike interval adaptation (Connors and Kriegstein1986). We present here results for putative pyramidal neurons.
Extracellular recordings. We performed extracellular recordings at12 recording sites, while simultaneously recording the membranepotential from one or more nearby neurons, in seven turtles. We usedtungsten microelectrodes (MicroProbes, heat-treated tapered tip), with~0.5-M� impedance. Electrodes were slowly advanced through tissueunder visual guidance with a manipulator (Narishige) during moni-toring for spiking activity with custom acquisition software (NationalInstruments). Extracellular activity was collected with an A-M Sys-tems model 1800 amplifier, digitized (NI PCIe-6231), and recordedwith custom software (National Instruments), sampling at 10 kHz.
Visual stimulation. The visual stimulation protocol has been de-scribed previously (Clawson et al. 2017; Wright et al. 2017; Wright andWessel 2017). Briefly, visual stimuli were presented with a projector(Aaxa Technologies, P4X Pico Projector) combined with a system oflenses (Edmund Optics) to project images generated by a custom soft-ware package directly onto the retina. The stimulus was a sustained grayscreen, a naturalistic movie [“catcam” (Betsch et al. 2004)], a motion-enhanced movie (Nishimoto and Gallant 2011), or a phase-shuffledversion of the same movie (courtesy Jack Gallant and Woodrow Shew).In all cases, the stimulus was triggered with a custom LabVIEW program(National Instruments).
For each cell and extracellular recording site, we selected one of thefour stimuli listed above to present across all trials. The preparationwas in complete darkness before and after each stimulus presentation.Stimuli lasted either 10 s or 20 s and were shown at least 12 times,with at least 30 s between the end of one presentation and thebeginning of the next.
Processing of intracellular and extracellular voltage recordings.Raw data traces were downsampled to 1,000 Hz. We used an algo-rithm to detect spikes in the membrane potential, and the values in a50-ms window centered on the maximum of each spike were replacedvia interpolation. Finally, we applied a 100-Hz low-pass Butterworthfilter. We used a sine-wave removal algorithm to minimize 60-Hz linenoise in extracellular recordings. This algorithm uses the method ofleast squares to estimate the phase and amplitude of the 60-Hz sine
wave in the ongoing LFP. A sine wave of 3.4-s duration (with theresulting phase and amplitude) is then subtracted from the full trace.
Data included in analysis. For each extracellular recording site, weused visual inspection to determine the quality of the recordings. Ingeneral, we excluded recording sites from consideration if voltagetraces displayed excessive 60-Hz line noise, low-frequency noise(likely reflecting a damaged electrode), or on average small responseamplitudes relative to baseline.
For intracellular recordings, we also excluded some trials and cells.To include a given trial, we required the membrane potential to remainat or above the calculated inhibitory reversal potential from thebeginning of the ongoing epoch to the end of the steady-state epoch.The inhibitory reversal potential was calculated with the chlorideconcentrations in the intracellular and extracellular solutions, butbecause of partial transfer of intracellular solution to the cell interior,it was possible for the recorded membrane potential to drop below thisvalue. This causes the conductance estimation algorithm (see below)to return a singularity. Rather than reset the inhibitory reversalpotential to the minimum membrane potential value for such a trial,we took the more conservative approach of excluding the trial fromconsideration. We also excluded trials with excessive low-frequencyartifacts or membrane potential drift. Finally, we considered only cellswith 12 or more retained trials for analysis.
In some cases, an extracellular electrode remained at a singlerecording site while we performed whole cell recordings either simul-taneously or sequentially from multiple nearby cells. To calculate CCfor a cell with the nearby LFP (see Correlated variability), weobtained the average LFP response (used to calculate residual traces)from those trials in which both the intracellular and extracellularvoltage were recorded and retained.
Inferred excitatory conductance. The algorithm for obtaining anestimated excitatory synaptic conductance (g) from membrane poten-tial V for single trials has been described in detail and validatedpreviously (Yasar et al. 2016). Briefly, the algorithm approximates asolution to the underdetermined equation
0 � CdV�t�
dt� gl�V�t� � El� � ge�t��V�t� � Ee� � gi�t�
�V�t� � Ei�where C is the assumed membrane capacitance, V(t) is the measuredmembrane potential as a function of time, Ee (Ei) is the assumedexcitatory (inhibitory) reversal potential, El is the assumed leakreversal potential, gl is the assumed leak conductance, and ge(t) [gi(t)]is the unknown excitatory (inhibitory) synaptic conductance. Toestimate ge(t), we first introduce a mathematical construct �(t), whichis defined according to
0 � CdV�t�
dt� gl�V�t� � El� � ��t��V�t� � Ei�
For each recording, we solve this equation for �(t). This attributesall membrane potential fluctuations to a single (unrealistic) inhibitoryconductance. As such, �(t) contains negative values and rapid down-ward fluctuations that are due to the influence of excitatory currents onthe membrane potential. Because conductance cannot have negativevalues, we then set the negative values in �(t) equal to zero, resultingin ��t� [previously called “nonnegative �(t)”]. Next, we use linearinterpolation to smooth out the rapid fluctuations in ��t�. The outputof this smoothing process is �(t), a smoother and therefore morerealistic estimation of the inhibitory synaptic conductance. Finally, wesubstitute �(t) into the equation
0 � CdV�t�
dt� gl�V�t� � El� � g�t��V�t� � Ee� � ��t��V�t� � Ei�
to obtain an estimation of the excitatory synaptic conductance (g). Ingeneral, this algorithm sacrifices knowledge about the inhibitory
3347VARIABILITY AND ADAPTATION OF SYNAPTIC INPUT COUPLING
J Neurophysiol • doi:10.1152/jn.00398.2017 • www.jn.org
Downloaded from www.physiology.org/journal/jn by ${individualUser.givenNames} ${individualUser.surname} (128.252.035.199) on December 13, 2017.Copyright © 2017 American Physiological Society. All rights reserved.

conductance to gain a better estimation of the excitatory conductance.Furthermore, it capitalizes on the fact that excitatory currents arefaster than—and therefore tend to interrupt—inhibitory currents. Itshould also be noted that this approach does not correct for thelow-pass filtering of (primarily thalamocortical) distal synaptic inputs(Smith et al. 1980) or (likely relatively infrequent) dendritic sodiumspikes (Larkum et al. 2008).
We have made several improvements to the algorithm since intro-ducing it. The original algorithm reliably estimated excitatory con-ductances with simulated membrane potentials. A recorded membranepotential, however, will contain high-frequency noise, which can beremoved by filtering (with, e.g., a 100-Hz Butterworth low-pass filter).This filtering process also leads to a smoother � �t�. As mentionedabove, detecting fast fluctuations in this signal is a critical step in theestimation process, and the algorithm’s performance was thus com-promised by the filter (as evidenced by its application to filtered, noisysimulated membrane potentials). We therefore revised the criteria fordetecting and replacing rapid fluctuations in ��t� (see Yasar et al. 2016for previous criteria). First, after calculating ��t�, we obtained the timeseries d�� �t��⁄dt. We then determined each time t= at which d���t��⁄dt crossed a threshold of one negative standard deviation. Thisthreshold optimized the algorithm’s performance when applied tonoisy simulated data. Finally, we linearly connected the local maximaof � �t� immediately prior and posterior to t=.
Applying the algorithm to a membrane potential recording alsorequires estimating the resting membrane potential for that trial. Anunrealistic choice will lead to spurious waveforms in the estimatedconductance. We estimated the resting membrane potential for eachtrial by calculating the median membrane potential value during thequiescent period in that trial. To isolate this quiescent period, we firstremoved a window of activity starting at stimulus onset and ending 6s after stimulus offset. This resulted in either a 14-s or 24-s trace of“spontaneous” activity that was on average quiescent relative to thatin the removed window. We then used an algorithm to detect spon-taneous “bursts” of activity lasting at least 1 s within the remainingtrace and removed these bursts. Finally, we took the median value(which is more robust to outliers than the mean) of the resulting traceto be the resting membrane potential (or El) for the correspondingvisual stimulation trial.
We used the following values for algorithm parameters: C � 1 nF,Ee � 90 mV, Ei � �80 mV, gl � 10 nS.
Coefficient of variation. The coefficient of variation (CV) is ascaled measure of variability: the standard deviation divided by themean. Using the set of all cells (n � 39), we calculated CV as afunction of time [CV(t)] for the inferred excitatory conductance. First,we applied a 100-ms “box filter” to each g trace: for each time step,we replaced the value of the trace with the average value in a 100-mswindow starting at that time step. We then advanced the window 10ms and repeated the process for the full length of the trace. Then, foreach cell, we calculated the across-trial standard deviation and meanof the filtered traces as a function of time. This was done for the entirepopulation, resulting in 39 (mean, standard deviation) ordered pairsfor each time step. Next, for each time step we fit the set of means tothe set of standard deviations by linear regression. The slope (standarderror) of this fit was the CV (SE) for the time step. To determine thesignificance of a change in CV across epochs, we compared the set ofall CV values for one epoch to that from the other (e.g., the 100 valuesfrom the ongoing epoch and the 100 values from the transient) withthe Wilcoxon signed-rank test. We report the time-averaged CV valuein a given epoch as the mean � 1 standard deviation.
Additive variability. To calculate the relative contribution of addi-tive variability to single-trial responses for a given cell, we began by“box-filtering” each trace, as described above for the calculation ofCV (except that here we used nonoverlapping 50-ms windows). Then,for each epoch, we regressed the resulting trace onto an across-trialaverage trace. This average trace was calculated from all trials for that
cell, excluding the individual trial in question. This yielded a single-trial R2 value for each epoch, for that trial. We repeated this for alltrials and calculated the across-trial median R2 value for each epoch.We repeated this for all cells.
Correlated variability. For each single-trial time series X, theresidual (Xr or deviation from the average activity) was found bysubtracting the across-trial average time series from the single-trialtime series:
Xr � X � �X�trials
Residuals were then separated into three epochs: the ongoing epoch(defined to be the 1 s before the onset of visual stimulation), thetransient epoch (200–1,200 ms after stimulus onset), and the steady-state epoch (1,400–2,400 ms after stimulus onset). For each gr-LFPr
pair, the Pearson correlation between residuals was then calculated foreach epoch and trial. The results were averaged across all trials,resulting in the trial-averaged correlated variability (CC) for each pairand epoch:
CCepoch � �cov�grepoch, LFPr
epoch� ⁄ [var(grepoch)var�LFPr
epoch�]1⁄2�trials
Significance tests for each pair and the population of pairs wereapplied as described in Statistical analysis.
Power spectral analysis. For each trial and signal, we extracted a4.4-s window of activity (with epoch windows and gaps betweenepochs as described above, plus 500-ms windows on each end toavoid filtering artifacts in the ongoing and steady-state epochs) andcalculated the residual time series as described above. For eachresidual trace, we performed wavelet analysis in MATLAB withsoftware provided by C. Torrence and G. Compo (Torrence andCompo 1998) (available at http://paos.colorado.edu/research/wave-lets/). This resulted in a power time series for each cell, for multiplefrequencies. For each frequency below 100 Hz, we averaged the timeseries across each epoch to obtain the average power at each fre-quency for each epoch. We then averaged across trials to obtainPepoch. For each cell, we also obtained the relative power spectrum(rPepoch) for the transient and steady-state epochs, defined to be thetrial-averaged evoked spectrum divided by the trial-averaged ongoingspectrum:
rPepoch � Pepoch ⁄ Pongoing
For each frequency, we calculated the bootstrap interval for therelative power as described in Statistical analysis.
Network models. To investigate the biophysical mechanisms un-derlying our experimental results, we implemented a series of modelnetworks composed of 800 excitatory and 200 inhibitory single-compartment leaky integrate-and-fire neurons. In the model thatreproduced our principal experimental results, excitatory-excitatoryconnections had clustered connectivity (Bujan et al. 2015; Litwin-Kumar and Doiron 2012; Watts and Strogatz 1998) (with 3% con-nection probability), and all other connections were random (with 3%excitatory-inhibitory and 20% inhibitory-excitatory and inhibitory-inhibitory connection probability). To implement the clustered excit-atory-excitatory connectivity, we began by constructing a “ring net-work” of 800 excitatory nodes. Each node in the network wasconnected to its 24 nearest neighbors (reflecting 3% connectionprobability). The weight of each of these connections was drawn froma beta distribution with average value 1.0. Finally, 1% of theseconnections were randomly rewired. That is, for each nonzero con-nection between a presynaptic and a postsynaptic node a differentpostsynaptic node was randomly selected from the excitatory network,with a probability of 1%.
The dynamics of the membrane potential (V) of each node evolvedaccording to
3348 VARIABILITY AND ADAPTATION OF SYNAPTIC INPUT COUPLING
J Neurophysiol • doi:10.1152/jn.00398.2017 • www.jn.org
Downloaded from www.physiology.org/journal/jn by ${individualUser.givenNames} ${individualUser.surname} (128.252.035.199) on December 13, 2017.Copyright © 2017 American Physiological Society. All rights reserved.

�m
dV
dt� �
1
C�gL�V�t� � EL� � Isyn�t�
where the membrane time constant �m � 50 ms (25 ms) for excitatory(inhibitory) nodes, the membrane capacitance C � 0.4 nF (0.2 nF) forexcitatory (inhibitory) nodes, and the leak conductance gL � 10 nS (5nS) for excitatory (inhibitory) nodes. The leak reversal potential EL
for each node was a random value between �70 and �60 mV, drawnfrom a continuous uniform distribution (to model the variability inresting membrane potentials observed across neurons in the experi-mental data). The reversal potentials for the synaptic current Isyn(t)were EGABA � �68 mV and EAMPA � 50 mV. The spike thresholdfor each neuron was �40 mV. A neuron reset to �59 mV afterspiking and was refractory for 2 ms (excitatory) and 1 ms (inhibitory).
The synaptic conductance gYX(t) for each synapse type (betweenpresynaptic neurons of type X and postsynaptic neurons of type Y) hadthree relevant time constants: delay (�LX, that is, the lag betweenpresynaptic spike time and beginning of the conductance waveform),rise time (�RYX), and decay time (�DYX). After a presynaptic spike attime 0, the synaptic conductance dynamics were described by a fastexponential rise and a slower exponential decay, or an “alpha func-tion”:
gYX�t� � gYX0 �m
�DYX � �RYX��exp�
t � �LX
�DYX� � exp�
t � �LX
�RYX�
where gYX0 is the maximum synaptic conductance and time constants
(in ms) are �LE � 1.5, �REE � 0.2, �DEE � 1.0, �RIE � 0.2, �DIE �1.0, �LI � 1.5, �RII � 1.5, �DII � 6.0, �REI � 2.25, �DEI � 6.0.Maximum conductance values (in nS) were gEE
0 � 3.0, gIE0 � 6.0,
gEI0 � 30, gII
0 � 30. Thus inhibitory synapses were in general strongerand slower than excitatory synapses.
In response to a presynaptic spike in neuron j at time tjspk, the
weight (Wij) of a synapse connecting neurons j and i depressed andrecovered toward the initial value (Wij
0) according to
dWij
dt� �
Wij�t��depress
�t � tjspk� �
Wij0 � Wij�t��recover
with depression time constant �depress � 300 ms and recovery timeconstant �recover � 2,500 ms. Intracortical synapses were subject todepression for the entire simulation, but synaptic depression was onlyapplied to external inputs after the stimulus onset (i.e., after theincrease in external input rate, see below). The synaptic weight matrixwas reset to Wij
0 at the start of each trial.All excitatory and inhibitory neurons received Poisson external
inputs. During “ongoing” activity, the external input rate to eachneuron was 65 Hz, which was sufficiently high to cause intracorticalspiking. The ongoing external input was unique across cells and trials.The stimulus was modeled as a gradual increase to 375 Hz; the inputrate was increased by 77.5 Hz at stimulus onset and by an additional77.5 Hz every 50 ms for 200 ms. This gradual increase provided morerealistic excitatory conductances than a single step function stimulusbut did not qualitatively impact the results. The stimulus was com-posed of two components: one that was unique across cells and trialsand one that was unique across cells but identical across trials,multiplied by proportionality constants 0.25 and 0.75, respectively.Thus for the poststimulus external drive to each neuron 25% of thevariance was explained by an input that was unique to each trial, and75% was explained by an input that was identical across trials. Thevariables for external inputs had the same parameters as for excita-tory-excitatory connections, and maximum conductances weregE � 6.0 nS. There was no synaptic depression for external inputsduring the ongoing epoch; the external drive during this window wassimply used to generate stimulus-independent intracortical spiking,and was thus treated as the “hidden” source triggering intrinsic events,as observed in experiment.
We repeated 20 trials for a single model network (defined byWij
0 ). Each trial was 4.4 s in duration, with stimulus onset at 1.7 s,and the time step was 0.05 ms. The ongoing epoch was defined tobe 1,200 ms to 200 ms before stimulus onset, the transient epoch0 ms to 1,000 ms after stimulus onset, and the steady-state epoch1,200 ms to 2,200 ms after stimulus onset. The additional 500 msat the beginning and end of each trial ensured there were nofiltering artifacts in the ongoing and steady-state epochs.
We modeled the LFP as the sum of all synaptic currents (similar toAtallah and Scanziani 2009; Destexhe 1998) to 100 neighboringneurons, multiplied by a factor of �1 (to mimic the change in polaritybetween voltages measured intracellularly and extracellularly). Thecontribution of each neuron to the LFP was not distance dependent.We then randomly selected 40 neurons from this subpopulation of 100neurons and used the excitatory synaptic conductances to generate 40g-LFP pairs for gr-LFPr CC analysis. For CV and R2 analysis, we used40 neurons randomly selected from the full population of 800 excit-atory neurons.
Statistical analysis. All statistical tests were performed withPython 2.7.
When asking whether a parameter of interest changed significantlyacross epochs for a population (e.g., whether the population-averagedCC for 21 g-LFP pairs changed significantly from the ongoing totransient epoch), we applied the Wilcoxon signed-rank test, whichreturns a P value for the two-sided test that the two related pairedsamples [representing, e.g., the 21 (CCongoing, CCtransient) pairedvalues] are drawn from the same distribution. This test was imple-mented with scipy.stats.wilcoxon (documentation and referencesavailable at https://docs.scipy.org/doc/scipy/reference/generated/scipy.stats.wilcoxon.html). When making multiple comparisons (e.g.,ongoing vs. transient, transient vs. steady state, and ongoing vs. steadystate), we divided the thresholds for significant P values by the totalnumber of comparisons.
To test whether a trial-averaged parameter of interest for one cell orelectrode (e.g., CC, averaged over 15 trials for 1 cell) changedsignificantly from one epoch to another, we used a bootstrap compar-ison test. For each epoch of interest, we calculated the �97.5%confidence intervals for the average value by bootstrapping (that is,resampling with replacement). If the bootstrap intervals for the twoepochs did not overlap, we reported that the two sets of values weresignificantly different (P � 0.05).
When calculating correlations between a pair of signals in which atleast one is slowly varying, it is possible for broad autocorrelations tointroduce spurious cross-correlations. This should be dealt with eitherby removing the broad autocorrelations (e.g., by “prewhitening” thesignals) or by accounting for their contribution to the cross-correla-tion. To avoid changing the temporal structure of the visual responses,we chose the latter approach. First, for each epoch and g-LFP pair, werandomly shuffled the trial order for one of the channels. We thencalculated the trial-average correlation of residuals (CCshuff) and thebootstrap interval for this shuffled data. The CC value for each pairand epoch was determined to be significant (with P � 0.05) if thebootstrap intervals for CC and CCshuff data did not overlap. Weindicate a significant CC value with a filled dot in the CC trajectory.Finally, for a given epoch, we compared the sets of CC and CCshuff
values for the population of g-LFP pairs with the Wilcoxon signed-rank test (as described above for across-epoch comparisons of CC).The population average for unshuffled data was determined to besignificant for P � 0.05. We repeated this second test using bootstrapintervals rather than the signed-rank test, with similar results (data notshown).
RESULTS
Visual stimulation evokes significant increases in corticalactivity. To quantify the response variability of synaptic inputsand its coupling with that of the local population, we recorded
3349VARIABILITY AND ADAPTATION OF SYNAPTIC INPUT COUPLING
J Neurophysiol • doi:10.1152/jn.00398.2017 • www.jn.org
Downloaded from www.physiology.org/journal/jn by ${individualUser.givenNames} ${individualUser.surname} (128.252.035.199) on December 13, 2017.Copyright © 2017 American Physiological Society. All rights reserved.

the membrane potential (V) from 39 putative pyramidal neu-rons in visual cortex of the turtle ex vivo eye-attached wholebrain preparation during visual stimulation of the retina (Fig.1C; see MATERIALS AND METHODS). V, however, is not a straight-forward readout of synaptic activity but rather represents anonlinear integration of excitatory and inhibitory inputs. Wehave recently developed and validated an algorithm to estimatethe excitatory synaptic conductance (g) from V (Yasar et al.2016), and here we applied this method to recordings ofongoing and visually evoked activity (Fig. 1C; see MATERIALS
AND METHODS). For 21 of these neurons, we also recorded thenearby LFP, which has been shown to be a reliable estimator oflocal synaptic activity (Haider et al. 2016).
Ongoing activity in turtle visual cortex was relatively qui-escent, typically with infrequent postsynaptic potentials at thelevel of the membrane potential and little baseline LFP activity(Fig. 2). On a minority of trials, this quiescent activity wasinterrupted by spontaneous “bursts” of activity lasting up tohundreds of milliseconds that were qualitatively similar tovisual responses (Fig. 2, A and D). Visual stimulation evokedbarrages of postsynaptic potentials and large fluctuations in thenearby LFP (Fig. 1C, Fig. 2A). The power spectra of theseevoked membrane potential and LFP fluctuations containedprominent peaks in the 10–100 Hz range (indicating oscilla-tory cortical activity), with peak location and amplitude vary-ing drastically across trials (Hoseini et al. 2017). This resultedin relatively smooth across-trial average relative power spectra
(i.e., power spectra for the transient and steady-state epochsdivided by that for the ongoing epoch) for g and LFP (Fig. 2B).On average, power in the 1–100 Hz frequency range increasedby orders of magnitude for both g and LFP (population-averaged relative power �rP� � 3,632.7 � 3,538.0, mean �SE, Fig. 2, B and C; �rPLFP� � 1,902.9 � 1,350.7, data notshown, transient). Response amplitudes (Fig. 2A) and power(Fig. 2, B and C) decreased from transient to steady state,despite continued visual stimulation (�rP� � 1,251.9 � 962.5,steady state; P � 6.06 � 10�8 for transient vs. steady statecomparison, Wilcoxon signed-rank test; �rPLFP� � 557.9 �449.1, steady state; P � 1.2 � 10�3 for transient vs. steadystate comparison, Wilcoxon signed-rank test).
Visual responses are highly variable across trials. For agiven cell and nearby LFP, the across-trial average response toa given stimulus displayed clear temporal structure (Fig. 2A).Still, responses were highly variable across stimulus presenta-tions; single-trial fluctuations were large relative to the meanresponse, with the across-trial variability increasing along withthe across-trial average activity (Fig. 2, A and D). To determinethe relationship between the variability and the average re-sponse of g, we calculated the scaled variability, or coefficientof variation (CV), as a function of time, for the population ofall cells (see MATERIALS AND METHODS). While variability ofevoked activity was larger than that of ongoing activity (Fig.2D), CV decreased after stimulus onset and slowly recovered
Fig. 2. Visual stimulation evokes increases in synaptic activity, and responses are highly variable across trials. A: inferred excitatory synaptic conductance (g,red) and measured LFP (black) for 3 trials (low opacity) and average across 32 trials (high opacity). Colors indicate ongoing (yellow), transient (blue), andsteady-state (green) epochs (see RESULTS and MATERIALS AND METHODS). a.u., Arbitrary units. B: relative power spectra (mean � bootstrap intervals; seeMATERIALS AND METHODS) for g (top) and LFP (bottom) for transient (blue) and steady-state (green) epochs for example pair in A. C: total relative power (1–100Hz) for 39 cells. Each blue (green) dot represents the across-trial mean relative power for 1 cell during the transient (steady state) epoch. High-opacity linesconnecting dots indicate significant change across epochs (P � 0.05, bootstrap comparison test; see MATERIALS AND METHODS). Asterisks above line connectingepochs indicate P � 1 � 10�3 (Wilcoxon signed-rank test). D: close-up view of ongoing (left) and evoked (right) synaptic activity. E: coefficient of variation(CV) as a function of time, calculated for 39 cells (mean � SE). Dashed line indicates CV � 1.0.
3350 VARIABILITY AND ADAPTATION OF SYNAPTIC INPUT COUPLING
J Neurophysiol • doi:10.1152/jn.00398.2017 • www.jn.org
Downloaded from www.physiology.org/journal/jn by ${individualUser.givenNames} ${individualUser.surname} (128.252.035.199) on December 13, 2017.Copyright © 2017 American Physiological Society. All rights reserved.

(Fig. 2E). Using the windows of activity defined above, wefound that this initial decrease was significant (time-averagedcoefficient of variation �CV� � 1.83 � 0.13 ongoing, 0.22 �0.04 transient, mean � SD, P � 1.74 � 10�16 for ongoing vs.transient comparison, Wilcoxon signed-rank test). Further-more, �CV� increased significantly from transient to steadystate (�CV� � 0.36 � 0.06 steady state, P � 1.74 � 10�16 fortransient vs. steady state comparison, Wilcoxon signed-ranktest) but remained significantly smaller than during ongoingactivity (P � 1.74 � 10�16 for ongoing vs. steady statecomparison, Wilcoxon signed-rank test).
Additive variability dominates single-trial responses. Hav-ing established the presence of significant across-trial responsevariability, we next sought to determine the relative contribu-tion of “additive variability” to single-trial responses. In thecontext of response time series, additive variability refers tosingle-trial deviations from a scaled version of the across-trialaverage response. These within-trial fluctuations could dimin-ish the ability of cortical neurons to reliably encode sequencesof stimuli (e.g., movie frames). In contrast, slower fluctuationsin neural excitability (leading to a rescaling of the averageresponse) may be less harmful.
To quantify this additive component, we first binned eachsingle-trial inferred excitatory synaptic conductance (sum-ming over 50-ms bins, resulting in g) and then calculated theacross-trial average binned conductance (�g�trials, Fig. 3A;
see MATERIALS AND METHODS). Given the linear relationshipbetween synaptic conductance and presynaptic spiking, thisis akin to binning spike counts in the presynaptic popula-tion. Finally, we regressed g onto �g�trials for each trial andtook the across-trial median explained variance (R2) foreach cell and epoch. This measure can be thought of as aproxy for response reliability, with R2 � 1 indicating apurely scaled version of the average response and R2 � 0indicating purely additive variability. By visual inspection, itwas evident that additive variability contributed to visualresponses (Fig. 3A). For example, while a typical response wassomewhat “enveloped” by the average time course, responsesalso tended to possess small, random fluctuations about themean, or in some instances larger deviations away from themean (Fig. 3A, trial 3, steady-state epoch). The responsereliability measure (R2) supported this observation and indi-cated that for a given cell the across-trial average was arelatively poor predictor of the single-trial response (see ex-ample cell in Fig. 3B); across the population, the averageresponse explained only 28.1 � 13.9% of the variance inindividual trials for the transient epoch (across-cell averageexplained variance �R2� � 0.28 � 0.14; Fig. 3C). The ex-plained variance was even lower during the steady state(�R2� � 0.17 � 0.15; Fig. 3C), decreasing significantly fromthat of the transient epoch (P � 1.5 � 10�3 for transient vs.steady state comparison, Wilcoxon signed-rank test). In sum-
Fig. 3. Single-trial variability is a mix of additive and multiplicative gain. A: inferred excitatory synaptic conductance, integrated over a 50-ms sliding window(with no overlap) for individual trials (low opacity), and across-trial average (high opacity) (see MATERIALS AND METHODS). B: across-trial distribution of R2 valuesfor the cell represented in A, calculated via linear regression of single-trial response onto average response for ongoing (left), transient (center), and steady-state(right) epochs. Red vertical lines indicate median values. Black scale bar on right indicates 10%. C: across-trial median R2 values for 39 cells, for each epoch.Each dot represents the across-trial median R2 value for 1 cell during the indicated epoch. High-opacity lines connecting dots indicate significant change acrossepochs (P � 0.05, bootstrap comparison test; see MATERIALS AND METHODS). Asterisks above lines connecting epochs indicate results of comparisons viaWilcoxon signed-rank test (**3.3 � 10�4 P � 3.3 � 10�3, ***P � 3.3 � 10�4).
3351VARIABILITY AND ADAPTATION OF SYNAPTIC INPUT COUPLING
J Neurophysiol • doi:10.1152/jn.00398.2017 • www.jn.org
Downloaded from www.physiology.org/journal/jn by ${individualUser.givenNames} ${individualUser.surname} (128.252.035.199) on December 13, 2017.Copyright © 2017 American Physiological Society. All rights reserved.

mary, single-trial responses were in general dominated byadditive variability, with the relative contribution increasingover the duration of the response.
Population coupling transiently increases after visualstimulation. Single-neuron response variability of this magni-tude has the potential to profoundly influence sensory coding,provided it is significantly coupled across a population ofneurons (Abbott and Dayan 1999; Averbeck et al. 2006;Shadlen and Newsome 1998). This is particularly relevant forthe steady-state response, which was more variable than theearly response (Fig. 3). We quantified this “population cou-pling” (Haider et al. 2016; Okun et al. 2015) for 21 cells bycalculating the single-trial residual responses for the estimatedconductance (gr, the single-trial time series with the across-trialaverage time series subtracted) and the nearby LFP (resultingin LFPr; Fig. 4A) and calculating the Pearson correlationcoefficient for residual pairs for each trial and epoch (similar toTan et al. 2014; Wright et al. 2017; Yu and Ferster 2010; seeMATERIALS AND METHODS).
For a given stimulus condition, the trial-averaged correlationcoefficient (CC) was broadly distributed across the population(Fig. 4B). During ongoing activity, CC was significantly non-zero for 7 of 21 pairs (P � 0.05, comparison to shuffled databy Wilcoxon signed-rank test; see MATERIALS AND METHODS).With visual stimulation, the population of pairs became moreanticorrelated (Fig. 4B); CC amplitudes increased significantlyfor 10 pairs (P � 0.017, across-epoch bootstrap comparison),and the population average decreased significantly (such thatthe amplitude increased; �CC� � 0.009 � 0.04 ongoing, P �0.50 for comparison to shuffled data; �CC� � �0.07 � 0.04transient, P � 1.1 � 10�4 for comparison to shuffled data;
P � 1.9 � 10�4 for ongoing vs. transient comparison, Wil-coxon signed-rank test; Fig. 4, B and C, top). During thistransient epoch, CC was significantly nonzero for 14 pairs(P � 0.05, comparison to shuffled data). This elevated level ofcoordination soon relaxed: from transient to steady state, CCamplitudes significantly decreased for five pairs (P � 0.05,across-epoch bootstrap comparison), such that CC was signif-icantly nonzero for seven pairs (P � 0.05, comparison toshuffled data) and the population average increased signifi-cantly toward zero (�CC� � �0.02 � 0.05 steady state, P �0.005 for transient vs. steady state comparison, Wilcoxonsigned-rank test) to values that were not significant across thepopulation (P � 0.11 for comparison to shuffled data, Fig. 4,B and C, bottom). Finally, as suggested by the crossing oflines in Fig. 4B, CC values in one epoch were in general notstrongly predictive of those in any other epoch (R2 � 0.01,P � 0.66, ongoing vs. transient comparison; R2 � 0.01, P �0.65, ongoing vs. steady state comparison; R2 � 0.33, P �0.007, transient vs. steady state comparison; linearregression).
These results suggest that the across-trial variability inevoked synaptic inputs to an individual neuron is, on average,coupled to that of other neurons in a nearby population in theearly response phase. Moreover, the strength of this coupling ishighly variable across cells. Coupling is not static, however;while response reliability decreases from the early to the latephase of the visual response (Fig. 3), the coupling strength doesas well (Fig. 4, B and C), suggesting that large single-trialfluctuations in the late response are more effectively “averagedout” across a large population. Finally, the fact that the cou-pling of a neuron in one epoch was not in general predictive of
Fig. 4. Correlated variability of synaptic input and LFP transiently increases with visual stimulation. A: residual traces for g (red) and LFP (black) for multipletrials. B: across-trial average Pearson correlation coefficient for g and LFP residual traces for 21 g-LFP pairs. Each dot indicates the across-trial average CC valuefor a given g-LFP pair. Filled dots indicate significant average values (P � 0.05, bootstrap comparison to shuffled data; see MATERIALS AND METHODS).High-opacity lines connecting dots indicate significant change across epochs (P � 0.05, bootstrap comparison test; see MATERIALS AND METHODS). Asterisks abovelines connecting epochs indicate results of comparisons via Wilcoxon signed-rank test (*3.3 � 10�3 P � 1.7 � 10�2, ***P � 3.3 � 10�4). C, top: distributionof change in across-trial average CC values (multiplied by �1) from ongoing to transient, for 21 g-LFP pairs (with results for shuffled data shown in gray). Blackscale bar indicates 10%. Bottom: same, but for transient to steady state.
3352 VARIABILITY AND ADAPTATION OF SYNAPTIC INPUT COUPLING
J Neurophysiol • doi:10.1152/jn.00398.2017 • www.jn.org
Downloaded from www.physiology.org/journal/jn by ${individualUser.givenNames} ${individualUser.surname} (128.252.035.199) on December 13, 2017.Copyright © 2017 American Physiological Society. All rights reserved.

that in another epoch suggests that variables other than con-nectivity determined population coupling during ongoing andvisually evoked activity.
Network properties shape response variability and g-LFPcorrelated variability. Cortical response variability and popu-lation coupling are likely shaped by three general sources:bottom-up sensory drive, recurrent (intracortical) activity, andtop-down (“brain state”) modulation (Rabinowitz et al. 2015).We sought to determine the relative contribution of feedfor-ward drive and recurrent activity to the phenomena we ob-served in experiment [i.e., the dynamics of scaled variability(Fig. 2E), response reliability (Fig. 3), and population coupling(Fig. 4)] and identify the biophysical mechanisms involved. Tothis end, we implemented a simple model cortical networksubject to external drive and analyzed the resulting excitatorysynaptic conductances and “local field potentials” (Fig. 5).
Our model was similar to that described previously (Wrightet al. 2017). The network consisted of 800 excitatory and 200inhibitory leaky integrate-and-fire neurons (see MATERIALS AND
METHODS). Excitatory-to-excitatory connections had clusteredconnectivity (3%), and all other connections were random (3%excitatory-to-inhibitory, 20% inhibitory-to-excitatory and in-hibitory-to-inhibitory). The clustered connectivity not onlyapproximates the space-dependent connectivity of the cortex(Perin et al. 2011; Song et al. 2005) but has been shown toinfluence spatiotemporal network activity patterns (Litwin-Kumar and Doiron 2012; Wright et al. 2017), response vari-ability (Litwin-Kumar and Doiron 2012), and correlated vari-ability (Litwin-Kumar and Doiron 2012; Wright et al. 2017).Nonzero synaptic weights were drawn from a beta distributionwith mean value 1.0, which approximated the heterogeneousnature of synaptic strengths across cortex (Cossell et al. 2015).All neurons received Poisson process external inputs, and thestimulus was modeled as an increase in external input rate. Theexternal drive was unique across neurons and trials during theongoing epoch. After stimulus onset, the external drive was amix of two components: one that was unique across neurons,but identical across trials (with proportionality constant 0.75),and one that was unique across both neurons and trials (withproportionality constant 0.25). Because visual stimulation re-liably evoked strong LFP oscillations in experiment (Fig. 2A;see Hoseini et al. 2017), we selected a set of synaptic rise anddecay times that were consistent with network spike rateoscillations in response to strong external drive (Fig. 5B). Eachsynapse depressed and slowly recovered in response to apresynaptic spike. We modeled the LFP as the sum of allsynaptic currents (similar to Atallah and Scanziani 2009;Destexhe 1998) to a subpopulation of 100 neighboring excit-atory neurons. We selected 40 neurons from the geometriccenter of this population for analysis of excitatory conduc-tances (Fig. 5B; see MATERIALS AND METHODS for additionalmodel details).
We first asked how the dynamics of scaled variability wereshaped by feedforward and recurrent inputs. As in experiment,g and LFP varied considerably across trials (Fig. 6A), despitethe stimulus being primarily the same across trials (seeMATERIALS AND METHODS). We calculated CV for excitatorysynaptic conductances and found that the CV dynamics weredetermined by both the statistics of the external drive andnetwork properties. When external drive during the ongoingepoch was sufficiently strong to cause sparse network spiking,
the CV for the total excitatory synaptic conductance to networkneurons hovered near 1.0 (time-averaged coefficient of varia-tion �CV� � 0.95 � 0.25, mean � SD for the ongoing epoch;Fig. 6B). This value greatly exceeded that of the external inputsalone (�CV� � 0.15 � 0.09; Fig. 6B). The difference arisesfrom the highly variable distribution of nonzero synapticweights (Fig. 6C, top). With stimulus onset, the CV forexternal inputs decreased by design (to �CV� � 0.004 � 0.01for the transient epoch), and CV for total excitatory conduc-tance initially did as well (�CV� � 0.40 � 0.16 for the transient
Fig. 5. Model overview. A: we implemented a model network of 800 excitatory(circles) and 200 inhibitory (triangles) leaky integrate-and-fire neurons, allsubject to Poisson process external inputs (green). Excitatory-excitatory con-nections had clustered connectivity, and all other connections were random. Asubset of 100 neighboring neurons (gray region) was used to define the LFP(see MATERIALS AND METHODS) B: network parameters were tuned such thatspike rate oscillations in the inhibitory (blue) and excitatory (black) popula-tions emerged in response to strong external drive. The LFP was modeled asthe sum of synaptic currents to a subset of 100 neighboring neurons (grayregion and single trial in black below). We selected neurons near the geometriccenter of this subset and analyzed the excitatory synaptic conductances (singletrial in red below, corresponding to neuron indicated by red arrow).
3353VARIABILITY AND ADAPTATION OF SYNAPTIC INPUT COUPLING
J Neurophysiol • doi:10.1152/jn.00398.2017 • www.jn.org
Downloaded from www.physiology.org/journal/jn by ${individualUser.givenNames} ${individualUser.surname} (128.252.035.199) on December 13, 2017.Copyright © 2017 American Physiological Society. All rights reserved.
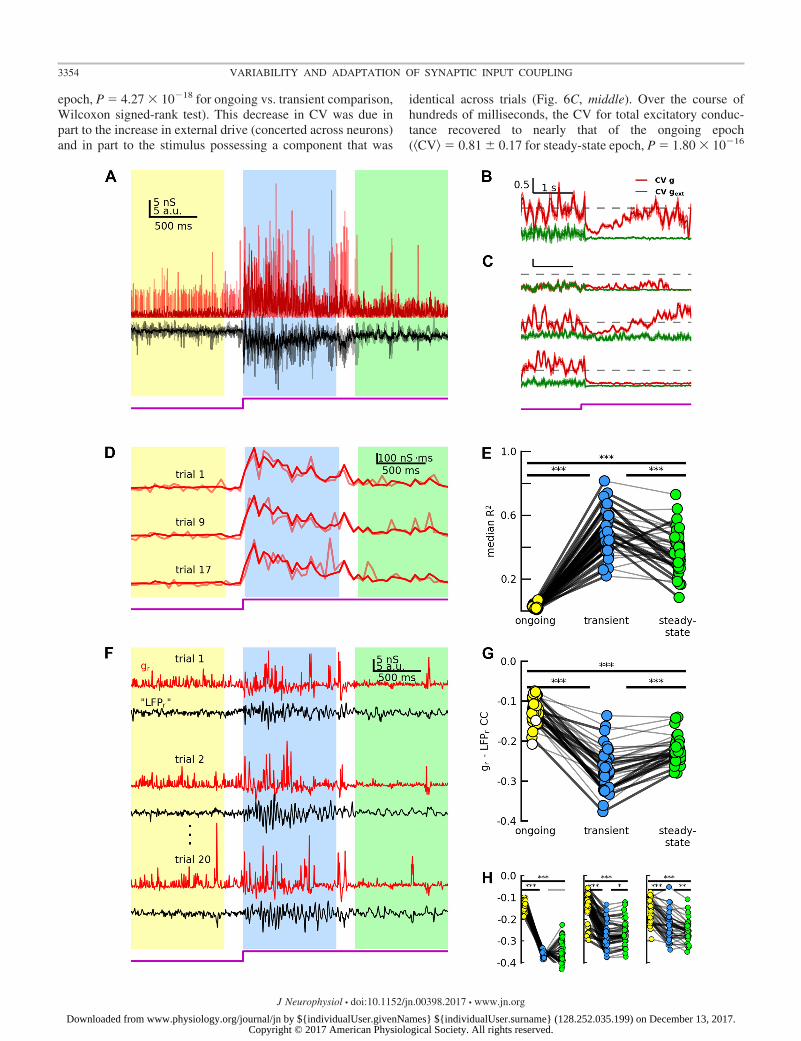
epoch, P � 4.27 � 10�18 for ongoing vs. transient comparison,Wilcoxon signed-rank test). This decrease in CV was due inpart to the increase in external drive (concerted across neurons)and in part to the stimulus possessing a component that was
identical across trials (Fig. 6C, middle). Over the course ofhundreds of milliseconds, the CV for total excitatory conduc-tance recovered to nearly that of the ongoing epoch(�CV� � 0.81 � 0.17 for steady-state epoch, P � 1.80 � 10�16
3354 VARIABILITY AND ADAPTATION OF SYNAPTIC INPUT COUPLING
J Neurophysiol • doi:10.1152/jn.00398.2017 • www.jn.org
Downloaded from www.physiology.org/journal/jn by ${individualUser.givenNames} ${individualUser.surname} (128.252.035.199) on December 13, 2017.Copyright © 2017 American Physiological Society. All rights reserved.

for transient vs. steady state comparison, P � 1.1 � 10�4 forongoing vs. steady state comparison, Wilcoxon signed-ranktest), which was an exaggeration of the empirical scaledvariability dynamic observed here (Fig. 2E) and elsewhere(Churchland et al. 2010). Synaptic depression mediated thisrecovery (Fig. 6C, bottom). Thus CV values and dynamicsdepended on the distribution of synaptic weights, the across-trial variability of external inputs, and synaptic adaptation.
We next investigated the relative contributions of feedfor-ward drive and intracortical activity to the dynamics of additivevariability we observed in experiment (Fig. 3A) and what mech-anisms might be involved. Specifically, we controlled “feedfor-ward” variability by setting R2 � 0.75 for the external driveduring the transient and steady-state epochs (see MATERIALS AND
METHODS). Despite this static feedforward variability, the modelqualitatively reproduced the dynamics of additive variability intransient and steady-state activity. As in experiment, individualtrials contained large additive fluctuations (Fig. 6D, compare withFig. 3A) and response reliability significantly decreased fromtransient to steady state (�R2� � 0.50 � 0.13 transient, �R2� �0.37 � 0.13 steady state, P � 7.6 � 10�6 for transient vs. steadystate comparison, Wilcoxon signed-rank test; Fig. 6E, comparewith Fig. 3C). This decrease was not related to synaptic depres-sion (data not shown), suggesting single-trial “errors” com-pounded over the duration of the response. Notably, the percent-age of single-trial variance explained by the average response ineither epoch was smaller than the 75% predicted by the stimulus.This surplus variability was therefore due to the only other sourceof randomness in the model: the state of the intracortical synapsesat stimulus onset (due to the variable external drive and intracor-tical synaptic depression during the ongoing epoch; see MATERIALS
AND METHODS). Together, these results suggest that recurrent cor-tical activity—and its sensitivity to conditions at stimulus onset—contributed significantly to the additive variability dynamics weobserved in experiment.
Next, we investigated the intracortical mechanisms thatshaped cortical population coupling distributions. As in exper-iment, we calculated correlated variability for g-LFP pairs (thatis, gr-LFPr CC, the Pearson correlation coefficient of residuals;Fig. 6F). The synaptic weight distribution strongly influencedgr-LFPr CC distributions. For each epoch, CC was broadlydistributed across the population (Fig. 6G). While some vari-ability is to be expected from such a sparsely connectednetwork, CC distributions were far less variable in a network
with binary synapses (but the same average synaptic weight;Fig. 6H, left).
Finally, we asked whether intracortical mechanisms couldexplain the changes in population coupling across epochs (Fig.4B). We found that the dynamics of gr-LFPr CC did indeeddepend on a variety of network parameters and emergentnetwork properties, such as oscillations. We recently used asimilar network to demonstrate the effects of coordinatedspiking on membrane potential correlated variability (Wright etal. 2017). Briefly, when the external drive triggers networkspike rate oscillations, high-frequency (20–100 Hz) membranepotential fluctuations become more correlated. Synaptic adap-tation subsequently reduces these correlations by modulatingthe network oscillations. Here, we find that this coordinationdynamic is also manifested as an increase in g-LFP correlatedvariability from the ongoing to the transient epoch(�CC� � �0.12 � 0.03 ongoing; �CC� � �0.27 � 0.05 tran-sient; P � 3.57 � 10�8 for ongoing vs. transient comparison,Wilcoxon signed-rank test; Fig. 6G). Synaptic depression withslow recovery (see MATERIALS AND METHODS) diminished net-work activity levels and, crucially, abolished large-scale coor-dinated spiking (Fig. 5B). This had the effect of drasticallyreducing gr-LFPr CC amplitudes from transient to steady state(�CC� � �0.22 � 0.03 steady state; P � 1.1 � 10�7 fortransient vs. steady state comparison, Wilcoxon signed-ranktest; Fig. 6G), despite continued network activity (Fig. 5B, Fig.6A). When either synaptic depression was removed (Fig. 6H,center) or the network was tuned to remain asynchronous (Fig.6H, right; see MATERIALS AND METHODS), changes in �CC� weremuch smaller across epochs and did not qualitatively match theexperimental results. As such, these results implicate emergentnetwork oscillations—and the corresponding relevant anatom-ical network properties (i.e., synaptic time constants and syn-aptic depression)—in the determination of population couplingdynamics.
Taken together, these model results suggest that 1) corticalproperties are sufficient to qualitatively reproduce the experi-mentally observed response variability and population cou-pling of synaptic inputs; 2) the salient cortical propertiesinclude synaptic clustering, time constants, and depression; and3) these properties modulate gr-LFPr CC distributions anddynamics in large part via their roles in generating and mod-ulating emergent cortical phenomena.
Fig. 6. A model network qualitatively reproduces the experimental results. A: excitatory synaptic conductance for 1 model neuron (g, red) and nearby LFP (black)for 3 trials (low opacity) and average across 20 trials (high opacity). Colors indicate ongoing (yellow), transient (blue), and steady-state (green) epochs. Examplecell is located at the geometric center of the pool defining the LFP (see RESULTS and MATERIALS AND METHODS). B: coefficient of variation (CV) as a functionof time (mean � SE), calculated using 40 cells randomly selected from the network, for total excitatory synaptic conductance (red) and for external excitatoryconductance (green). Dashed line indicates CV � 1.0. C: CV for alternate model versions. Top: network with binary synaptic weights. Middle: network subjectto unique stimulus on each trial. Bottom: network without synaptic adaptation. Scale bar and dashed line same as in B. D: excitatory synaptic conductance for1 model neuron, integrated over a 50-ms sliding window (with no overlap) for individual trials (low opacity), and across-trial average (high opacity; seeMATERIALS AND METHODS). E: across-trial median R2 values for 40 cells randomly-selected from network, for each epoch. Each dot represents the across-trialmedian R2 value for 1 cell during the indicated epoch. High-opacity lines connecting dots indicate significant change across epochs (P � 0.05, bootstrap comparisontest; see MATERIALS AND METHODS). Asterisks above lines connecting epochs indicate results of comparisons via Wilcoxon signed-rank test (***P � 3.3 � 10�4). F:residual traces for g (red) for 1 model neuron and nearby LFP (black) for multiple trials. G: across-trial average Pearson correlation coefficient for g and LFP residualtraces for 40 g-LFP pairs (where 40 cells were selected from geometric center of pool defining LFP). Each dot indicates the across-trial average CC value for a giveng-LFP pair. Filled dots indicate significant average values (P 0.05, bootstrap comparison to shuffled data; see MATERIALS AND METHODS). Filled dots indicate significantaverage values. High-opacity lines connecting dots indicate significant change across epochs (P � 0.05, bootstrap comparison test; see MATERIALS AND METHODS).Asterisks above lines connecting epochs indicate results of comparisons via Wilcoxon signed-rank test (***P � 3.3 � 10�4). H: same as in G, for alternate modelversions. Left: network with binary synapses (i.e., synaptic weights either 1 or 0). Center: network without synaptic depression. Right: asynchronous network (*3.3 �10�3 P � 1.7 � 10�2, **3.3 � 10�4 P � 3.3 � 10�3).
3355VARIABILITY AND ADAPTATION OF SYNAPTIC INPUT COUPLING
J Neurophysiol • doi:10.1152/jn.00398.2017 • www.jn.org
Downloaded from www.physiology.org/journal/jn by ${individualUser.givenNames} ${individualUser.surname} (128.252.035.199) on December 13, 2017.Copyright © 2017 American Physiological Society. All rights reserved.

DISCUSSION
To obtain a spike rate-independent measure of single-neuronacross-trial response variability, and to measure the neuron’scoupling with local population activity, we simultaneouslyrecorded the membrane potential from putative pyramidalneurons and the nearby LFP in the turtle visual cortex duringongoing and stimulus-modulated activity (Fig. 1). We esti-mated the excitatory synaptic conductance (g) from the mem-brane potential and quantified the across-trial variability in gand correlated variability with the LFP. We discovered thatvisual responses were highly variable across trials and that bothadditive and multiplicative gain contributed to the responsevariability. Importantly, we found a range of neuron-to-net-work coupling across cortical neurons. The results of a modelinvestigation suggest that this coupling is dynamically modu-lated during visual processing via biophysical and emergentnetwork properties.
Studies spanning several decades have described a remark-able degree of variability in the sensory-evoked spiking re-sponses of cortical neurons (Britten et al. 1993; Carandini2004; Schölvinck et al. 2015). Recent work suggests that thisvariability is shaped by the cortex itself. First, cortical vari-ability surpasses that of the inputs from thalamus (Schölvincket al. 2015). Second, scaled variability decreases with stimulusonset across a variety of cortical areas and behavioral states,suggesting that the phenomenon is a property of large, recur-rent networks (Churchland et al. 2010). Third, single-neuronresponse variability can be modeled as a mix of multiplicativeand additive gain due to global cortical activity (Goris et al.2014; Lin et al. 2015; Rabinowitz et al. 2015). Our work,which incorporates a novel conductance estimation method,generally strengthens this “cortico-centric” view of responsevariability, and our network model identifies specific corticalproperties that likely shape cortical response variability.
For instance, we found that individual neurons receivedexcitatory synaptic inputs that were extremely variable acrossstimulus presentations (Fig. 2, A and D, Fig. 3). This variabilitywas predominantly additive in nature (i.e., the fluctuationsoccurred on short timescales relative to the duration of thestimulus), and the contribution from this additive variabilityincreased from transient to steady state (Fig. 3). What deter-mined the level of variability and its dynamics during theresponse? There are at least three possible candidates: bot-tom-up inputs (i.e., sensory drive), recurrent intracortical ac-tivity, and fluctuations in brain state (Rabinowitz et al. 2015).To identify the potential contribution from feedforward andrecurrent inputs, we implemented a simple model network withknown feedforward variability (that was constant across thetransient and steady-state epochs). This model qualitativelyreproduced the empirical results (Fig. 6, D and E) and suggeststhat small deviations in initial conditions (e.g., intracorticalsynaptic strengths) lead to a modest initial amplification ofthalamic response variability, with single-trial error com-pounding over the duration of the response. In other words,recurrent activity likely contributed significantly to the R2
dynamics we observed in experiment. Such chaotic dynamicsare a hallmark of balanced networks (Shadlen and Newsome1998; van Vreeswijk and Sompolinsky 1996). While at firstglance this seems extremely disadvantageous to sensory cod-ing, the balanced regime has other advantages, including fast
responses to changes in external stimuli (van Vreeswijk andSompolinsky 1996), effective signal propagation (Vogels andAbbott 2005), and maximized information capacity (Shew etal. 2011).
Previous work has shown that (spike based) populationcoupling is broadly distributed across cells, which may reflectthe degree to which a given neuron samples the local popula-tion (Okun et al. 2015) and the structure of that connectivity(Pernice et al. 2011). In agreement with this, we found thatg-LFP correlated variability was broadly distributed acrosscells for a given stimulus condition (Fig. 4B). Our modelresults reinforce the hypothesis that this across-cell variabilityis related to the underlying connectivity: CC values werebroadly distributed for relatively realistic, heavy-tailed synap-tic weight distributions (Fig. 6G) but narrowly distributed forbinary synapses (Fig. 6H, left). These weight distributions alsoshaped the dynamics of scaled variability (Fig. 6, B and C, top).These results suggest that cortical connectivity patterns aremanifested in the response variability and coordinated variabil-ity of synaptic activity. Given the relevance of synaptic activityto spiking (Doiron et al. 2016; Litwin-Kumar et al. 2011;Lyamzin et al. 2015), this likely reflects the correspondingresponse properties of population spiking observed elsewhere(Okun et al. 2015).
Anatomical connectivity was not the only relevant variable;we found that g-LFP correlated variability amplitudes signifi-cantly increased with visual stimulation (Fig. 4, B and C, top),in agreement with previous work (Haider et al. 2016). Inter-estingly, CC amplitudes decreased after the early responsephase (Fig. 4, B and C, bottom), despite persistent synaptic andlocal population activity (Fig. 2, A–C, Fig. 4A). Was thisdynamic imposed by the external inputs, or was the corticalnetwork itself capable of exhibiting multiple population cou-pling “states”? Because of the limitations of our experimentalapproach, we cannot rule out the former explanation. Still, ourmodel results support the latter hypothesis. Specifically, weconstrained the model network to reproduce two aspects ofempirical visual responses: LFP oscillations (Fig. 5B, bottom,Fig. 1B, bottom) and evoked activity that was more coordinatedacross the network in the early response and more locallycoordinated in the later response (Fig. 5B, bottom; see Shew etal. 2015). These response properties are not themselves trivi-ally related to local population coupling, yet imposing theseconstraints recovered the gr-LFPr CC dynamic (Fig. 6G). Thatis, the coupling dynamics at one scale may be concomitantwith networkwide state fluctuations. Such a relationship wouldbe consistent with the observations that spontaneous fluctua-tions in cortical state can influence g-LFP (Haider et al. 2016)and spike-spike (Okun et al. 2015; Schölvinck et al. 2015)population coupling and further suggests that variations in thecortical state (i.e., the excitatory-inhibitory balance) acrossrecording sessions and experiments could be responsible forthe general reordering of CC values across epochs (Fig. 4B).Taken together, our results advance a “cortico-centric” view ofpopulation coupling by identifying specific features of cortex(e.g., synaptic time constants and synaptic adaptation) capableof influencing population coupling dynamics via emergentnetwork phenomena.
The population coupling values we observed were in generalsmall, even during the early response phase (Fig. 4B), which isroughly in line with previous observations in mouse (Haider et
3356 VARIABILITY AND ADAPTATION OF SYNAPTIC INPUT COUPLING
J Neurophysiol • doi:10.1152/jn.00398.2017 • www.jn.org
Downloaded from www.physiology.org/journal/jn by ${individualUser.givenNames} ${individualUser.surname} (128.252.035.199) on December 13, 2017.Copyright © 2017 American Physiological Society. All rights reserved.

al. 2016) and monkey (Tan et al. 2014) V1. In other words,most of the response variability in synaptic inputs to a givenneuron was not explained by fluctuations in the local networkdefined by the reach of the nearby extracellular electrode. Thislikely reflects the “constellation-like” connectivity of cortex, inwhich a given neuron is driven by a sparse subset of the localpopulation (as well as more distant neurons). In line with this,we have recently shown that during visual responses prespikedepolarizations (Wright and Wessel 2017) and high-frequencysubthreshold oscillations (Wright et al. 2017) are only weaklyshared between pairs of nearby neurons. This suggests that theLFP is unlikely to isolate individual cortical microcircuits,making the population-averaged g-LFP CC value in a givenepoch difficult to interpret on its own. Nevertheless, changes ing-LFP CC across stimulus conditions likely track shiftingnetwork coordination levels. Thus the CC dynamics we ob-served here suggest that networkwide coordination is strongerduring the early response and returns to baseline within 1 s ofstimulus onset. This is supported by our network modelingresults (Fig. 6) and our previous studies of large-scale corticalactivity in this same preparation (Clawson et al. 2017; Shew etal. 2015). According to one view of population coding, thisdecrease in coupling in the late response may benefit corticalfunction: while steady-state activity is less reliable than that inthe early response (Fig. 2E, Fig. 3), these later fluctuations aremore private and therefore tend to average out across a neuralensemble (Zohary et al. 1994). Our model results suggest thatthis transition is mediated by adaptation (i.e., synaptic depres-sion), which is consistent with the emerging view that corticaladaptation serves as much more than a modulator of activitylevels but is in addition a “knob” for fine-tuning a variety offunctionalities (Benucci et al. 2013; Gutnisky and Dragoi2008; Ollerenshaw et al. 2014; Shew et al. 2015; Whitmire andStanley 2016; Wright et al. 2017; Zheng et al. 2015). In otherwords, adaptation modulates both the overall level (i.e., totalspike rate) and the regime (i.e., spatiotemporal neuronal cor-relations patterns) of cortical activity.
There exist several useful algorithms for estimating proper-ties of synaptic activity from membrane potential recordings(Bédard et al. 2012; Berg and Ditlevsen 2013; Borg-Graham etal. 1998; Lankarany et al. 2013; Paninski et al. 2012; Puggioniet al. 2017; Rudolph and Destexhe 2003; Wehr and Zador2003). The analysis performed here requires knowledge of thesynaptic conductance time series on individual trials, which afew cleverly crafted algorithms can provide (Bédard et al.2012; Berg and Ditlevsen 2013; Lankarany et al. 2013; Pan-inski et al. 2012). Each of these algorithms has a particular setof strengths, weaknesses, and underlying assumptions. Forinstance, one that uses a Bayesian approach requires priorknowledge of the statistics of presynaptic firing (Paninski et al.2012), and several do not appear to resolve fluctuations thatoccur at timescales on the order of 10 ms (Berg and Ditlevsen2013; Lankarany et al. 2013; Paninski et al. 2012). Another faroutperforms our algorithm in the estimation of inhibitoryconductances (Bédard et al. 2012), but excitatory conductancesare less reliable (compare Fig. 4 in Yasar et al. 2016 and Fig.8 in Bédard et al. 2012). Here we apply an algorithm thatcomplements these approaches and is particularly well suitedfor this study; it makes no assumptions about the statistics ofnetwork activity and allows us to calculate population coupling
for excitatory conductances with relatively high temporal res-olution.
One major limitation of our experimental work is the lack ofdirect measurements of inhibitory synaptic conductances,which are a key component of single-neuron and networkwideresponse properties. Inhibition represents a significant propor-tion of the total synaptic input to a given neuron (Haider et al.2013) and tends to be correlated across pairs of neurons(Hasenstaub et al. 2005), and the relative timing of excitatoryand inhibitory currents may determine precise spike timing(Haider and McCormick 2009; Hasenstaub et al. 2005; Nowaket al. 1997) and feature selectivity (Wilent and Contreras2005). Furthermore, the inhibitory population is known to playa vital role in such emergent network phenomena as spike rateoscillations (Brunel and Wang 2003), and the excitation-inhi-bition balance may represent a fundamental aspect of thecortical code (Denève and Machens 2016). In future studies, itwill be straightforward to modify our experimental approachand conductance estimation algorithm to investigate inhibition.More importantly, this approach can be combined with multi-whole cell recording to simultaneously infer excitatory con-ductances in one cell, and inhibitory in another, similar tostudies of evoked activity in rat somatosensory cortex (Okunand Lampl 2008) and spontaneous activity in rat hippocampus(Atallah and Scanziani 2009) and mouse thalamocortical slice(Graupner and Reyes 2013). This would be particularly usefulin areas such as visual cortex, where responses can be complexand highly variable across trials (thus limiting the utility ofrecording excitation and inhibition from one cell on alternatingtrials).
The turtle ex vivo preparation is well suited to obtainingwhole cell recordings of cortical visual responses (Crockett etal. 2015; Wright et al. 2017; Wright and Wessel 2017), butthere are important differences between turtle and mammalianvisual cortex that should be taken into account when seeking togeneralize these results. Most notably, the turtle cortex hasthree rather than six layers, and the single “cellular layer” maybe comparable to L5 and L6 of mammalian visual cortex(Reiner 1993). In fact, the cytoarchitecture of turtle visualcortex is perhaps more reminiscent of mammalian olfactorycortex or hippocampus (Fournier et al. 2015). Nevertheless, theturtle visual thalamocortical system possesses anatomical andphenomenological properties that make it useful for investigat-ing cortical function generally (Shepherd 2011) [e.g., diversemorphological and electrophysiological cortical cell types(Connors and Kriegstein 1986; Crockett et al. 2015; Mancillaand Ulinski 2001), feedforward inhibition (Mancilla and Ulin-ski 2001), and cortical oscillations (Clawson et al. 2017;Hoseini et al. 2017; Shew et al. 2015)]. Thus we anticipate thatcortical response variability and population coupling in mam-malian visual cortex are in general shaped by the variablesidentified here and that specific results (e.g., population cou-pling distributions) may vary considerably across cortical lay-ers (as would be predicted from the strong dependence onconnectivity demonstrated by our model, Fig. 6).
Taken together, our results demonstrate the highly variablenature of visually evoked synaptic inputs (and therefore spa-tiotemporal spike patterns) in cortical microcircuits. Further-more, these results suggest that several properties of thisvariability are largely determined intracortically and identifyspecific, highly relevant cortical parameters. Importantly, these
3357VARIABILITY AND ADAPTATION OF SYNAPTIC INPUT COUPLING
J Neurophysiol • doi:10.1152/jn.00398.2017 • www.jn.org
Downloaded from www.physiology.org/journal/jn by ${individualUser.givenNames} ${individualUser.surname} (128.252.035.199) on December 13, 2017.Copyright © 2017 American Physiological Society. All rights reserved.

cortical properties together lead to an adapted network statethat is in many ways ideal for sensory processing. As such, thiswork contributes to a clearer picture of the effects of anatom-ical and emergent network properties on single-neuron sensoryresponses and networkwide function.
ACKNOWLEDGMENTS
We thank Woodrow Shew for assistance with the design of the visualstimulus. We credit the conception and design of the “motion-enhanced”stimulus to Jack Gallant.
GRANTS
This research was supported by Whitehall Foundation Grant 20121221 (R.Wessel) and National Science Foundation CRCNS Grant 1308159 (R. Wes-sel).
DISCLOSURES
No conflicts of interest, financial or otherwise, are declared by the authors.
AUTHOR CONTRIBUTIONS
N.C.W., M.S.H., T.B.Y., and R.W. conceived and designed research;N.C.W. performed experiments; N.C.W. analyzed data; N.C.W., M.S.H.,T.B.Y., and R.W. interpreted results of experiments; N.C.W. prepared figures;N.C.W. and R.W. drafted manuscript; N.C.W., M.S.H., and R.W. edited andrevised manuscript; N.C.W., M.S.H., T.B.Y., and R.W. approved final versionof manuscript.
REFERENCES
Abbott LF, Dayan P. The effect of correlated variability on the accuracy ofa population code. Neural Comput 11: 91–101, 1999. doi:10.1162/089976699300016827.
Armstrong CM, Gilly WF. Access resistance and space clamp problemsassociated with whole-cell patch clamping. Methods Enzymol 207: 100–122, 1992. doi:10.1016/0076-6879(92)07007-B.
Atallah BV, Scanziani M. Instantaneous modulation of gamma oscillationfrequency by balancing excitation with inhibition. Neuron 62: 566–577,2009. doi:10.1016/j.neuron.2009.04.027.
Averbeck BB, Latham PE, Pouget A. Neural correlations, population codingand computation. Nat Rev Neurosci 7: 358–366, 2006. doi:10.1038/nrn1888.
Bédard C, Béhuret S, Deleuze C, Bal T, Destexhe A. Oversampling methodto extract excitatory and inhibitory conductances from single-trial mem-brane potential recordings. J Neurosci Methods 210: 3–14, 2012. doi:10.1016/j.jneumeth.2011.09.010.
Benucci A, Saleem AB, Carandini M. Adaptation maintains populationhomeostasis in primary visual cortex. Nat Neurosci 16: 724–729, 2013.doi:10.1038/nn.3382.
Berg RW, Ditlevsen S. Synaptic inhibition and excitation estimated via thetime constant of membrane potential fluctuations. J Neurophysiol 110:1021–1034, 2013. doi:10.1152/jn.00006.2013.
Betsch BY, Einhäuser W, Körding KP, König P. The world from a cat’sperspective—statistics of natural videos. Biol Cybern 90: 41–50, 2004.doi:10.1007/s00422-003-0434-6.
Borg-Graham LJ, Monier C, Frégnac Y. Visual input evokes transient andstrong shunting inhibition in visual cortical neurons. Nature 393: 369–373,1998. doi:10.1038/30735.
Britten KH, Shadlen MN, Newsome WT, Movshon JA. Responses ofneurons in macaque MT to stochastic motion signals. Vis Neurosci 10:1157–1169, 1993. doi:10.1017/S0952523800010269.
Brunel N, Wang XJ. What determines the frequency of fast network oscil-lations with irregular neural discharges? I. Synaptic dynamics and excita-tion-inhibition balance. J Neurophysiol 90: 415–430, 2003. doi:10.1152/jn.01095.2002.
Bujan AF, Aertsen A, Kumar A. Role of input correlations in shaping thevariability and noise correlations of evoked activity in the neocortex. JNeurosci 35: 8611–8625, 2015. doi:10.1523/JNEUROSCI.4536-14.2015.
Carandini M. Amplification of trial-to-trial response variability by neurons invisual cortex. PLoS Biol 2: E264, 2004. doi:10.1371/journal.pbio.0020264.
Churchland MM, Yu BM, Cunningham JP, Sugrue LP, Cohen MR,Corrado GS, Newsome WT, Clark AM, Hosseini P, Scott BB, BradleyDC, Smith MA, Kohn A, Movshon JA, Armstrong KM, Moore T,Chang SW, Snyder LH, Lisberger SG, Priebe NJ, Finn IM, Ferster D,Ryu SI, Santhanam G, Sahani M, Shenoy KV. Stimulus onset quenchesneural variability: a widespread cortical phenomenon. Nat Neurosci 13:369–378, 2010. doi:10.1038/nn.2501.
Clawson WP, Wright NC, Wessel R, Shew WL. Adaptation towards scale-free dynamics improves cortical stimulus discrimination at the cost ofreduced detection. PLOS Comput Biol 13: e1005574, 2017. doi:10.1371/journal.pcbi.1005574.
Cohen MR, Kohn A. Measuring and interpreting neuronal correlations. NatNeurosci 14: 811–819, 2011. doi:10.1038/nn.2842.
Connors BW, Kriegstein AR. Cellular physiology of the turtle visual cortex:distinctive properties of pyramidal and stellate neurons. J Neurosci 6:164–177, 1986.
Cossell L, Iacaruso MF, Muir DR, Houlton R, Sader EN, Ko H, Hofer SB,Mrsic-Flogel TD. Functional organization of excitatory synaptic strength inprimary visual cortex. Nature 518: 399–403, 2015. doi:10.1038/nature14182.
Crockett T, Wright N, Thornquist S, Ariel M, Wessel R. Turtle dorsalcortex pyramidal neurons comprise two distinct cell types with indistin-guishable visual responses. PLoS One 10: e0144012, 2015. doi:10.1371/journal.pone.0144012.
Denève S, Machens CK. Efficient codes and balanced networks. Nat Neurosci19: 375–382, 2016. doi:10.1038/nn.4243.
Destexhe A. Spike-and-wave oscillations based on the properties of GABAB
receptors. J Neurosci 18: 9099–9111, 1998.Doiron B, Litwin-Kumar A, Rosenbaum R, Ocker GK, Josic K. The
mechanics of state-dependent neural correlations. Nat Neurosci 19: 383–393, 2016. doi:10.1038/nn.4242.
Fournier J, Müller CM, Laurent G. Looking for the roots of cortical sensorycomputation in three-layered cortices. Curr Opin Neurobiol 31: 119–126,2015. doi:10.1016/j.conb.2014.09.006.
Goris RL, Movshon JA, Simoncelli EP. Partitioning neuronal variability. NatNeurosci 17: 858–865, 2014. doi:10.1038/nn.3711.
Graupner M, Reyes AD. Synaptic input correlations leading to membranepotential decorrelation of spontaneous activity in cortex. J Neurosci 33:15075–15085, 2013. doi:10.1523/JNEUROSCI.0347-13.2013.
Gutnisky DA, Dragoi V. Adaptive coding of visual information in neuralpopulations. Nature 452: 220–224, 2008. doi:10.1038/nature06563.
Haider B, Häusser M, Carandini M. Inhibition dominates sensory responsesin the awake cortex. Nature 493: 97–100, 2013. doi:10.1038/nature11665.
Haider B, McCormick DA. Rapid neocortical dynamics: cellular and networkmechanisms. Neuron 62: 171–189, 2009. doi:10.1016/j.neuron.2009.04.008.
Haider B, Schulz DP, Carandini M. Millisecond coupling of local fieldpotentials to synaptic currents in the awake visual cortex. Neuron 90: 35–42,2016. doi:10.1016/j.neuron.2016.02.034.
Hasenstaub A, Shu Y, Haider B, Kraushaar U, Duque A, McCormick DA.Inhibitory postsynaptic potentials carry synchronized frequency informationin active cortical networks. Neuron 47: 423–435, 2005. doi:10.1016/j.neuron.2005.06.016.
Henze DA, Borhegyi Z, Csicsvari J, Mamiya A, Harris KD, Buzsáki G.Intracellular features predicted by extracellular recordings in the hippocam-pus in vivo. J Neurophysiol 84: 390–400, 2000.
Hoseini MS, Pobst J, Wright N, Clawson W, Shew W, Wessel R. Inducedcortical oscillations in turtle cortex are coherent at the mesoscale of popu-lation activity, but not at the microscale of the membrane potential ofneurons. J Neurophysiol 118: 2579–2591, 2017. doi:10.1152/jn.00375.2017.
Ikegaya Y, Aaron G, Cossart R, Aronov D, Lampl I, Ferster D, Yuste R.Synfire chains and cortical songs: temporal modules of cortical activity.Science 304: 559–564, 2004. doi:10.1126/science.1093173.
Koch C. Biophysics of Computation. Oxford, UK: Oxford Univ. Press, 2004.Lankarany M, Zhu WP, Swamy MN, Toyoizumi T. Inferring trial-to-trial
excitatory and inhibitory synaptic inputs from membrane potential usingGaussian mixture Kalman filtering. Front Comput Neurosci 7: 109, 2013.doi:10.3389/fncom.2013.00109.
Larkum ME, Watanabe S, Lasser-Ross N, Rhodes P, Ross WN. Dendriticproperties of turtle pyramidal neurons. J Neurophysiol 99: 683–694, 2008.doi:10.1152/jn.01076.2007.
3358 VARIABILITY AND ADAPTATION OF SYNAPTIC INPUT COUPLING
J Neurophysiol • doi:10.1152/jn.00398.2017 • www.jn.org
Downloaded from www.physiology.org/journal/jn by ${individualUser.givenNames} ${individualUser.surname} (128.252.035.199) on December 13, 2017.Copyright © 2017 American Physiological Society. All rights reserved.

Lin IC, Okun M, Carandini M, Harris KD. The nature of shared corticalvariability. Neuron 87: 644–656, 2015. doi:10.1016/j.neuron.2015.06.035.
Litwin-Kumar A, Doiron B. Slow dynamics and high variability in balancedcortical networks with clustered connections. Nat Neurosci 15: 1498–1505,2012. doi:10.1038/nn.3220.
Litwin-Kumar A, Oswald AM, Urban NN, Doiron B. Balanced synapticinput shapes the correlation between neural spike trains. PLOS Comput Biol7: e1002305, 2011. doi:10.1371/journal.pcbi.1002305.
Lyamzin DR, Barnes SJ, Donato R, Garcia-Lazaro JA, Keck T, LesicaNA. Nonlinear transfer of signal and noise correlations in cortical networks.J Neurosci 35: 8065–8080, 2015. doi:10.1523/JNEUROSCI.4738-14.2015.
MacLean JN, Watson BO, Aaron GB, Yuste R. Internal dynamics determinethe cortical response to thalamic stimulation. Neuron 48: 811–823, 2005.doi:10.1016/j.neuron.2005.09.035.
Mancilla JG, Ulinski PS. Role of GABAA-mediated inhibition in controllingthe responses of regular spiking cells in turtle visual cortex. Vis Neurosci 18:9–24, 2001. doi:10.1017/S0952523801181022.
Mokeichev A, Okun M, Barak O, Katz Y, Ben-Shahar O, Lampl I.Stochastic emergence of repeating cortical motifs in spontaneous membranepotential fluctuations in vivo. Neuron 53: 413–425, 2007. doi:10.1016/j.neuron.2007.01.017.
Nishimoto S, Gallant JL. A three-dimensional spatiotemporal receptive fieldmodel explains responses of area MT neurons to naturalistic movies. JNeurosci 31: 14551–14564, 2011. doi:10.1523/JNEUROSCI.6801-10.2011.
Nowak LG, Sanchez-Vives MV, McCormick DA. Influence of low and highfrequency inputs on spike timing in visual cortical neurons. Cereb Cortex 7:487–501, 1997. doi:10.1093/cercor/7.6.487.
O’Connor DH, Peron SP, Huber D, Svoboda K. Neural activity in barrelcortex underlying vibrissa-based object localization in mice. Neuron 67:1048–1061, 2010. doi:10.1016/j.neuron.2010.08.026.
Okun M, Lampl I. Instantaneous correlation of excitation and inhibitionduring ongoing and sensory-evoked activities. Nat Neurosci 11: 535–537,2008. doi:10.1038/nn.2105.
Okun M, Steinmetz N, Cossell L, Iacaruso MF, Ko H, Barthó P, Moore T,Hofer SB, Mrsic-Flogel TD, Carandini M, Harris KD. Diverse couplingof neurons to populations in sensory cortex. Nature 521: 511–515, 2015.doi:10.1038/nature14273.
Ollerenshaw DR, Zheng HJ, Millard DC, Wang Q, Stanley GB. Theadaptive trade-off between detection and discrimination in cortical repre-sentations and behavior. Neuron 81: 1152–1164, 2014. doi:10.1016/j.neuron.2014.01.025.
Paninski L, Vidne M, DePasquale B, Ferreira DG. Inferring synaptic inputsgiven a noisy voltage trace via sequential Monte Carlo methods. J ComputNeurosci 33: 1–19, 2012. doi:10.1007/s10827-011-0371-7.
Perin R, Berger TK, Markram H. A synaptic organizing principle forcortical neuronal groups. Proc Natl Acad Sci USA 108: 5419–5424, 2011.doi:10.1073/pnas.1016051108.
Pernice V, Staude B, Cardanobile S, Rotter S. How structure determinescorrelations in neuronal networks. PLOS Comput Biol 7: e1002059, 2011.doi:10.1371/journal.pcbi.1002059.
Puggioni P, Jelitai M, Duguid I, van Rossum MC. Extraction of synapticinput properties in vivo. Neural Comput 29: 1745–1768, 2017. doi:10.1162/NECO_a_00975.
Rabinowitz NC, Goris RL, Cohen M, Simoncelli EP. Attention stabilizes theshared gain of V4 populations. eLife 4: e08998, 2015. doi:10.7554/eLife.08998.
Reiner A. Neurotransmitter organization and connections of turtle cortex:implications for the evolution of mammalian isocortex. Comp BiochemPhysiol Comp Physiol 104: 735–748, 1993. doi:10.1016/0300-9629(93)90149-X.
Rudolph M, Destexhe A. Characterization of subthreshold voltage fluctua-tions in neuronal membranes. Neural Comput 15: 2577–2618, 2003. doi:10.1162/089976603322385081.
Saha D, Morton D, Ariel M, Wessel R. Response properties of visual neuronsin the turtle nucleus isthmi. J Comp Physiol A Neuroethol Sens NeuralBehav Physiol 197: 153–165, 2011. doi:10.1007/s00359-010-0596-3.
Schölvinck ML, Saleem AB, Benucci A, Harris KD, Carandini M. Corticalstate determines global variability and correlations in visual cortex. JNeurosci 35: 170–178, 2015. doi:10.1523/JNEUROSCI.4994-13.2015.
Shadlen MN, Newsome WT. The variable discharge of cortical neurons:implications for connectivity, computation, and information coding. J Neu-rosci 18: 3870–3896, 1998.
Shepherd GM. The microcircuit concept applied to cortical evolution: fromthree-layer to six-layer cortex. Front Neuroanat 5: 30, 2011. doi:10.3389/fnana.2011.00030.
Shew WL, Clawson WP, Pobst J, Karimipanah Y, Wright NC, Wessel R.Adaptation to sensory input tunes visual cortex to criticality. Nat Phys 11:659–663, 2015. doi:10.1038/nphys3370.
Shew WL, Yang H, Yu S, Roy R, Plenz D. Information capacity andtransmission are maximized in balanced cortical networks with neuronalavalanches. J Neurosci 31: 55–63, 2011. doi:10.1523/JNEUROSCI.4637-10.2011.
Shoham S, O’Connor DH, Segev R. How silent is the brain: is there a“dark matter” problem in neuroscience? J Comp Physiol A NeuroetholSens Neural Behav Physiol 192: 777–784, 2006. doi:10.1007/s00359-006-0117-6.
Smith LM, Ebner FF, Colonnier M. The thalamocortical projection inPseudemys turtles: a quantitative electron microscopic study. J Comp Neurol190: 445–461, 1980. doi:10.1002/cne.901900304.
Song S, Sjöström PJ, Reigl M, Nelson S, Chklovskii DB. Highly nonrandomfeatures of synaptic connectivity in local cortical circuits. PLoS Biol 3: e68,2005. (Erratum. PLoS Biol 3: e350, 2005). doi:10.1371/journal.pbio.0030068.
Spruston N, Jaffe DB, Williams SH, Johnston D. Voltage- and space-clamperrors associated with the measurement of electrotonically remote synapticevents. J Neurophysiol 70: 781–802, 1993.
Tan AY, Chen Y, Scholl B, Seidemann E, Priebe NJ. Sensory stimulationshifts visual cortex from synchronous to asynchronous states. Nature 509:226–229, 2014. doi:10.1038/nature13159.
Thompson LT, Best PJ. Place cells and silent cells in the hippocampus offreely-behaving rats. J Neurosci 9: 2382–2390, 1989.
Torrence C, Compo GP. A practical guide to wavelet analysis. Bull AmMeteorol Soc 79: 61–78, 1998. doi:10.1175/1520-0477(1998)079�0061:APGTWA�2.0.CO;2.
van Vreeswijk C, Sompolinsky H. Chaos in neuronal networks with balancedexcitatory and inhibitory activity. Science 274: 1724–1726, 1996. doi:10.1126/science.274.5293.1724.
Vogels TP, Abbott LF. Signal propagation and logic gating in networks ofintegrate-and-fire neurons. J Neurosci 25: 10786–10795, 2005. doi:10.1523/JNEUROSCI.3508-05.2005.
Watts DJ, Strogatz SH. Collective dynamics of “small-world” networks.Nature 393: 440–442, 1998. doi:10.1038/30918.
Wehr M, Zador AM. Balanced inhibition underlies tuning and sharpens spiketiming in auditory cortex. Nature 426: 442–446, 2003. doi:10.1038/nature02116.
Whitmire CJ, Stanley GB. Rapid sensory adaptation redux: a circuit per-spective. Neuron 92: 298–315, 2016. doi:10.1016/j.neuron.2016.09.046.
Wilent WB, Contreras D. Dynamics of excitation and inhibition underlyingstimulus selectivity in rat somatosensory cortex. Nat Neurosci 8: 1364–1370, 2005. doi:10.1038/nn1545.
Wright NC, Hoseini MS, Wessel R. Adaptation modulates correlated sub-threshold response variability in visual cortex. J Neurophysiol 118: 1257–1269, 2017. doi:10.1152/jn.00124.2017.
Wright NC, Wessel R. Network activity influences the subthreshold andspiking visual responses of pyramidal neurons in the three-layer turtlecortex. J Neurophysiol 118: 2142–2155, 2017. doi:10.1152/jn.00340.2017.
Yasar TB, Wright NC, Wessel R. Inferring presynaptic population spikingfrom single-trial membrane potential recordings. J Neurosci Methods 259:13–21, 2016. doi:10.1016/j.jneumeth.2015.11.019.
Yu J, Ferster D. Membrane potential synchrony in primary visual cortexduring sensory stimulation. Neuron 68: 1187–1201, 2010. doi:10.1016/j.neuron.2010.11.027.
Zheng HJ, Wang Q, Stanley GB. Adaptive shaping of cortical responseselectivity in the vibrissa pathway. J Neurophysiol 113: 3850–3865, 2015.doi:10.1152/jn.00978.2014.
Zohary E, Shadlen MN, Newsome WT. Correlated neuronal discharge rateand its implications for psychophysical performance. Nature 370: 140–143,1994. (Erratum. Nature 371: 358, 1994. doi:10.1038/370140a0.
3359VARIABILITY AND ADAPTATION OF SYNAPTIC INPUT COUPLING
J Neurophysiol • doi:10.1152/jn.00398.2017 • www.jn.org
Downloaded from www.physiology.org/journal/jn by ${individualUser.givenNames} ${individualUser.surname} (128.252.035.199) on December 13, 2017.Copyright © 2017 American Physiological Society. All rights reserved.















![Astrocytes, neurons, synapses: a tripartite view on cortical circuit … · 2018. 5. 1. · synaptic behavior [7, 30]. In addition to contacting neu-rons, astrocytes are interconnected](https://static.fdocuments.in/doc/165x107/60f3a9b5cc1e622054756bab/astrocytes-neurons-synapses-a-tripartite-view-on-cortical-circuit-2018-5-1.jpg)


