
core.ac.uk · Ancylostomiasis is one of the major public health problems in tropical countries. The...
146
Characterisation of Antigens of Hookworm Parasites ( SUMMARY ) A DISSERTATION SUBMITTED TO THE ALIGARH MUSLIM UNIVERSITY, ALIGARH FOR THE DEGREE OF DOCTOR OF PHILOSOPHY IN BIOCHEMISTRY BY SHAIINI MOHAN M. Sc. (Biochem. ),M. Phil. BIOCHEMISTRY DIVISION CENTRAL DRUG RESEARCH INSTITUTE LUGKNOW-226 00I AUGUST-1990
Transcript of core.ac.uk · Ancylostomiasis is one of the major public health problems in tropical countries. The...

Characterisation of Antigens of
Hookworm Parasites
( SUMMARY )
A DISSERTATION SUBMITTED
TO THE
ALIGARH MUSLIM UNIVERSITY, ALIGARH
FOR THE DEGREE OF DOCTOR OF PHILOSOPHY
IN
BIOCHEMISTRY
BY
SHAIINI MOHAN M. Sc. (Biochem. ) ,M. Phil.
BIOCHEMISTRY DIVISION
CENTRAL DRUG RESEARCH INSTITUTE LUGKNOW-226 00I
AUGUST-1990

Ancylostomiasis is one of the major public health problems
in tropical countries. The world incidence of hookworm infect
ions are 1000 million with the death rate of 55 thousand per
year. The currently available anthelmintics are not very
effective. This clearly emphasises the need to intensify
the control measures in hookworm endemic areas . An alternate
approach for the permanent control of the disease would be
to develop immunoprophylactic measures against hookworm
paras i tes . However, the existing knowledge about the molecular
immunology of hookworm infection is st i l l in i ts infancy.
Hookworm parasites evoke functional protective immunity and
some reports are available regarding the protective potential
of live parasit ic stages as well as of somatic extracts and
excretory-secretory products from adult and larval stages,
however, not much is known about the molecules/antigens which
are involved in evoking these protective responses. A basic
knowledge about the total antigenic composition of the para
sites is essential not only for understanding the host parasite
relationships but also for the identification of antigenic t a r
gets of protective immunity. • We have, therefore, directed
our studies towards the characterisation of antigens from adult
and larval stages of human hookworm parasi te ; Ancylostoma
ceylanicum, which has earl ier been used for the screenings
of anthelmintic compounds by several workers. Total protein
and antigenic composition of adult and larval stages of
A. ceylanicum were studied employing SDS-polyacrylamide gel

electrophoresis (SDS-PAGE) and immiinoelectrophoretic techniques
using hyperimmune rabbit sera. The levels of parasite specific
antibodies in immune sera were determined by enzyme linked
immunosorbent assay (ELISA). Similar techniques were used
for antigenic analysis of the excretory-secretory products
from adult A. ceylanicum. The protective potential of somatic
extracts from adult and larvae as wellas the excretory-secretory
products from adult A.ceylanicum was also studied by immunising
the naive hamsters with these antigens (somatic extracts from
adult and larvae and excretory-secretory products from adults)
followed by challenge infection.
The somatic extracts from adult (AcA) and larval stages
(AcL) of A. ceylanicum were separated on SDS-polyacrylamide
gradient gel electrophoresis (SDS-PAGE), which showed the
presence of 47 protein bands m adults and 43 in Larvae m
the molecular weight range of 10-170 kD. Most ot thr f)rr)tfin
bands were found to be common between these two stages
(adult and la rvae) , however, 8 (130, 125, 42, iO, 20, 17,
16.5 and 10 kD) and 4 (170, 160, 153 and 85 kD) protein
bands specific to adult and larval stages respectively were
observed. The quantitation of parasite specific antibodies
in immune rabbi t sera was done by enzyme linked immunosorbent
assay and reciprocal antibody t i te rs of 256,000 and 128, 000
were obtained for rabbit anti-adult and rabbit anti-larval
sera respectively using AcA antigen at optimum concentration
of 0.25 ug/well. Both these sera showed the reciprocal anti-

body t i t e r s of 256,000 with AcL at optimum concentration
of 0.50 ug/well.
The antigenic analysis of somatic extracts from adult
and larval stages of A.ceylanicum was init ially done by Immuno
electrophoresis ( lEP), which revealed 10-12 precipitin lines
in AcA with rabbit anti-AcA serum and 6-7 precipitin bands
in AcL with rabbit anti-AcL serum. Further characterisation
of these somatic antigens was carried out employing crossed
Immunoelectrophoresis (CIE) which showed the presence of
34 antigenic components in AcA and 19 antigens in AcL. Out
of 34 AcA antigens, 25 were anodic and 9 cathodic while 17
anodic and 2 cathodic antigens were observed in AcL. In order
to study the cross-reactive or common as well as stage/parasite
specific antigens of A.ceylanicum, crossed Immunoelectrophoresis
employing heterologous sera and crossed line immunoelectro-
phoresis (CLIE) were used. When AcA and AcL were reacted
with heterologous immune rabbit sera in CIE, most of the
antigens were found common or cross-reactive between the
two stages, however, 12 antigens specific to adult and 5 anti
gens specific to larval stage were observed. The subsequent
analysis of CLIE also revealed the same number of common
or cross-reactive as well as stage specific antigens in AcA
and AcL. In order to analyse A. ceylanicum antigens having
cross-react ivi ty with rat hookworm - Nippostrongylus brasi l iensis ,
CIE using immune rabbi t sera raised against adult (NbA) and
infective larval (NbL) somatic extracts of N, brasil iensis and

CLIE using NbA and NbL in the in te rmedia te ge l , were done.
Most of the antigens (22-26) were found to be AcA spec i f i c ,
only 12 and 8 antigens of AcA were found to be common/cross
r eac t i ve with NbA and NbL r e s p e c t i v e l y . On analysing the
c r o s s - r e a c t i v e or common antigens of AcL with NbL and NbA,
13 common or c ross r e a c t i v e and 6 AcL speci f ic antigens were
o b s e r v e d .
Another set of antigens analysed in t h e p resen t s tudy
i s the e x c r e t o r y - s e c r e t o r y (E-S) p roduc t s from adult
A. ceylanicum . The condit ions for the p repara t ion of E-S
p roduc t s were opt imised with r e s pec t to medium as well as
the incubation t i m e . Out of t h r e e different media; RPMI-1640,
DMEM and R inge r ' s solution t r i e d , R inge r ' s solution proved
to be the bes t as the mot i l i ty of the worms was maximum
in t h i s medium and the worms could be maintained upto 6h
without changing the medium. The amount of pro te in re leased
was found to be 95±15 ug/100 adults in RPMI-1640 and DMEM
as compared to 125±25 ug/100 adult worms in R inge r ' s solut ion.
Immunoelectrophoret ic ana lys i s using r a b b i t anti-AcA serum
showed the presence of 5-7 p r e c i p i t i n bands in E-S products
p r e p a r e d in t h e s e media . As t h e r e was not any significant
difference in the amount of pro te in r e l ea sed as well as the
antigenic p a t t e r n s of E-S p roduc t s p r e p a r e d in t h r e e media ,
R inge r ' s solution (containing s a l t s and glucose) was chosen
to obtain the E-S p roduc t s from A.ceylanicum adu l t s because
t h i s i s much s imple r and more economical in comparison to
o the r two media (RPMI-1640 and DMEM).

A polyvalent hyperimmune rabbit serum was raised against
A. ceylanicum adult E-S products and the t i ter of parasite
specific antibodies in immune rabbit sera was determined
by enzyme linked immunosorbent assay using optimum concentrat
ion of 0.50 ug E-S antigen/well. A reciprocal antibody t i ter
of 256,000 N*as obtained with rabbit anti-E-S serum. The
E-S antigens were also tested with the immune rabbit sera
raised against somatic extracts of A.ceylanicum in ELISA and
a t i ter value of 128,000 was obtained for both rabbit anti-
adult and rabbit anti- larval sera . Crossed Immunoelectropho
res is (CIE) using rabbit anti-E-S serum revealed 16 antigens
in A. ceylanicum E-S products, out of which 13 were anodic
and 3 moved towards the cathode. E-S products have also
been found to share antigenic determinants with the somatic
extracts from adult and larval stages. lEP analysis of E-S
products showed the presence of 6 and 4 precipitin bands
with rabbit anti-AcA and rabbit anti-AcL sera respectively.
On further analysing the E-S products by CIE using rabbit
sera raised against somatic extracts 12 and 7 antigens of
E-S products were found to share antigenic determinants with
AcA and AcL respect ively . Tandem crossed immunoelectropho-
resis (TCIE) of E-S products with AcA and AcL also revealed
almost the same number of common/cross-reactive antigens
between the E-S products and the somatic ex t rac ts . The pre
sence of host serum proteins in the E-S products was also
analysed by CIE using rabbit serum raised against normal

6
hamster serum, and 6 antigens in the E-S products appeared
to be of host origin.
The protective potential of somatic extracts from adult
and larval stages as well as excretory-secretory products
from adult A.ceylanicum was tested by immunizing the hamsters
with these antigens and evaluating the immunized hamsters
for protective immunity. The sera obtained from immunized
hamsters, before challenge, showed significant levels of para
site specific antibodies. The immunodiffusion and immuno-
electrophoretic analysis revealed the presence of precipitin
antibodies in immune hamster sera and 7 precipitin bands
in adult somatic extracts and 4 in both larval somatic extracts
and E-S products. The hamsters immunized with larval somatic
extracts showed maximum protection (93% reduction in worm
burden) while 75% and 63% protection was observed in hamsters
immunized with excretory-secretory products and adult somatic
extracts respect ively.
Therefore, in the present study we have been able to
characterise the protein and antigenic components of adult
and larval stages of A.ceylanicum using hyperimmune rabbit
sera having high t i te rs of parasite specific antibodies and
certain biochemical and immunochemical techniques. The common
or cross-reactive antigens between the two life stages of
A. ceylanicuiD and also between the two hookworm parasites
(A.ceylanicum and N.braslliensls) as well as stage and parasite

specilic antigens have also been identified. We have been
able to prepare the excretory-secretory products of A.ceylanicum
adults using Ringer's solution which is much simpler and
a more economical medium. Antigenic characterisation of E-S
products revealed less complex nature of these products as
compared to the a-dult SQma.tic extracts» The protective potea-
t ials of the somatic extracts from adult and larval stages
and also of E-S products from adult A. cey lanicum was also
studied. The larval somatic ext rac ts , though contain less
number of antigens, showed maximum protection followed by
excretory-secretory products and adult somatic extracts .
These studies provide basic information about the antigenic
compositions of gomatic extracts and excretory-secretory pro
ducts of A. ceylatnicum as well as the comparative efficacy
of these antigens in inducing protective immunity against this
paras i te . However, further studies on molecular characterisa
tion of these antigens are certainly required for identifying
the antigenic targets of protective immunity.

Characterisation of Antigens of
Hookworm Parasites
A DISSERTATION SUBMITTED
TO THE
ALIGARH MUSLIM UNIVERSITY, ALIGARH
FOR THE DEGREE OF DOCTOR OF PHILOSOPHY
IN
BIOCHEMISTRY
BY
5Hi4LINI MOHAN M. Sc. (Biochem. ) ,M. Phil.
BIOCHEMISTRY DIVISION
CENTRAL DRUG RESEARCH INSTITUTE
LUGKNOW-226 001 AUGUST-1990

T4022
- 7 AUG 19S2
V
cm ,D-200^
. - r C ''
-~.^.^'-
c«?-,CJ^^ -n tA

ACKNOblLEVGEUENTS
I takd thd piiviiZQH to KZcoid my thanks to Vn.. A.M.Siddiqui,
Pn.o{ie.Mofi, dtpaitmant o^ Biochzmi^tfty, Migaih Uu^tim Univzi^ity,
AZigafih (^o^i hi^ gaidanct and hzlp.
I exp^e4-6 thz deep -ien^e oi$ gratitude, to P- .lM- ) W.A.
Kaa-iihal, Stiznti^t, ViviMon o($ Biothzmiit^y and V^. V.C. KaU'6ha-£,
Sciznti^t, Viviiion 0($ Hicfiobiology, CVRl, Lucfenow, ^oi thzii
az^thztic. 4ugge4-t-con4, in^piiing gixidanaz and {,Kult{^al tfiiticium
through u)hich thi^ paramount ta-ik could acqulKZ thz p^e^ew-t
ihapz.
Uy gtatz^ainz^^ -6 ai'do duz touoaA-d^ Vn.. B.N. Vhawan,Vi'iZctoi,
CVRl, Vi. Pizm Sagai, Hzad, VzpafitmZnt 0($ Biochzmi4>tiy and thz
collzaguZ'ii 0($ thz dzpan-trnznti o($ Biochzmi'i^tfLy and Vaia-ftitotogy
who havz hzipzd me duiing thz zompZztion Oj$ thi'i> di^iy^zitation.
I am indzbtzd to Council o{^ Scizntif^ic and Inda-iit'ilal
Rz^za^ch, Wew Vzlhi (Jo-t financial Mppo^t. Thz Irmaculatz typing
oi Mfi. K.L. Gupta -c4 al^o acknooolzdgzd.
Lastly, but not thz Iza^t, I havz no itioid^ to zx.piZM my
humblz giatitudz, pKoiound fizgaid^ and lovz to my paiznt^ ^on.
thzin i^oibzaiancz and a^^zctionatz blzMing^ which uiziz all
thz uoay.
ShcUinJUofutifr

C O N T E N T S
Preface
Page No.
i - i i
Abbrevia t ions . . . . . . . . . i i i
Chapter I
Introduction and Review of L i t e ra tu re . . . . . . 1 - 2 1
Chapter II
Mater ia ls and Methods . . . . . . 2 2 - 3 2
Chapter III
Results and Discussion . . . . . .
Section I
Antigenic ana lys i s of somatic e x t r a c t s from adul t and infect ive l a r v a l s tages of Ancylostoma ceylanicum . . . 3 3 - 7 5
Section II
Excre to ry -Sec re to ry p roduc t s from adult
A.ceylanicum 76 - 106
Chapter IV
Summary and Conclusion . . . . . . 107 - 114
Bibliography . . . . . . . . . 1 1 5 - 1 2 5
* * * * * * * *
*

( i )
P r e f a c e
Almost every individual in the tropical regions of
the world is suffering from parasit ic infections of one kind
or the other . According to some recent repor ts , millions
of people are affected by differnt helminth parasites including
hookworms which account for 1000 million cases annually.
Heavy burdens of hookworm parasi tes cause severe anaemia
which may sometime prove fatal and can also lead to in
tellectual and growth retardation. Recently, chemotherapy
targetted to heavily infected individuals has come out as
a real is t ic approach, however, the permanent control measures
of the disease are s t i l l lacking. Hence, there is an urgent
need to evolve an alternate strategy for controlling the hook
worm disease. The existing knowledge about the molecular
immunology of hookworm infection is very poor and characteri
sation of hookworm antigens may provide valuable leads about
the antigenic moelcules of the possible immunoprophylactic
and immunodiagnostic significance.
The studies reported here are directed towards the
characterisation of somatic as well as excretory-secretory
antigens of Ancylostoma ceylanicum - a human hookworm, which
has been used for chemotherapeutic screening of potential
anthelmintics in our Institute. The antigens from adult and
infective larval stages of A. ceylanicum were also analysed
in order to identify the common/cross-reactive and stage/
parasite specific antigens of A. ceylanicum. The potential
of these antigens (somatic extracts from adult and larval

( i i)
stages as well as of excretory-secretory products from adult
A.ceylanicum) in inducing protective immunity in experimental
animals to challenge infection was investigated and the larval
somatic extracts showed significant degree of protection. The
information obtained from these findings may be of great impor
tance for the identification of antigens of possible protective
potential.

A B B R E V I A T I O N S
( i i i )
AcA
AcL
CIE
CLIE
ELISA
E-S
Ig
lEP
ID
"2°2
ug
NbA
Nbl,
OPD
SDS-PAGE
TEMED
TIE
A.ceylanicum adul t somatic e x t r a c t
A.ceylanicum l a r v a l somatic e x t r a c t
Crossed-immuno e l e c t r o p h o r e s i s
Crossed l ine Immunoelectrophoresis
Enzyme l inked immunosorbent assay
E x c r e t o r y - s e c r e t o r y
Immunoglobulin
Immunoelectrophoresis
Immunodiffusion
Hydrogen p e r o x i d e
Infective l a r v a l stage
microgram
N . b r a s i l i e n s i s adul t somatic e x t r a c t
N . b r a s i l i e n s i s l a r v a l somatic ex t r ac t
0 -phenylene diamine
SDS-polyacrylamide gel e l e c t r o p h o r e s i s
Tetra e t h y l methylene e thy lene diamine
Tandem-crossed Immunoelectrophoresis

CHAPTER I
INTRODUCTION AND
REVIEW OF LITERATURE

1
Hookworm parasites are among the imoortant intestinal
nematodes whose prevalence and concomitant detrimental effects
on the body functions have made them to be ranked as one
of the important helminthic diseases of man in developing
and underdeveloped countries. Although the infection does
not cause mortality, it saps the vital i ty of the hosts already-
suffering from malnutrition. According to the recent repor ts ,
about 1000 million people may be infected with hookworm
parasites (Hotez ^ al^.. 1987; Crompton, 1989).
"Hookworm disease", in man is mainly caused by the
three species - Necator americanus, Ancylostoma duodenale
and Ancylostoma ceylanicum. Besides the above three human
pathogens, the other parasites pathogenic to livestocks are
A.caninum (dog), A.tubaeformae (ca ts ) , Nippostrongylus
brasil iensis ( r a t s ) , Nematospiroides dubius (mouse), Bunonstomum
trigonocephalum (sheep and goat), B.phelabotomum (cow) and
Gaigeria pachycetis ( sheep) .
Hookworm infection is more prevalent in tropical and
subtropical regions of Asia, Africa, Europe and America. In
Asian continent the most endemic countries are India, Malaysia,
China and Japan. In India, the tropical climate, poor sanitation
and poorer health education favour the growth, multiplication
and dissemination of all parasit ic infections including hook
worms. The morbidity and mortalitv caused by the disease
were first reported in tea plantations of Assam, where approxi
mately 90% of the population harbour this infection. The inci-

dence of the disease is also very high in plain where p^ddy
cultivation is predominant (Raghavan, 1967), while the incidence
is low in Uttar Pradesh and Pondicherry (Yunus, 1977; Singh
e^ al^., 1978).
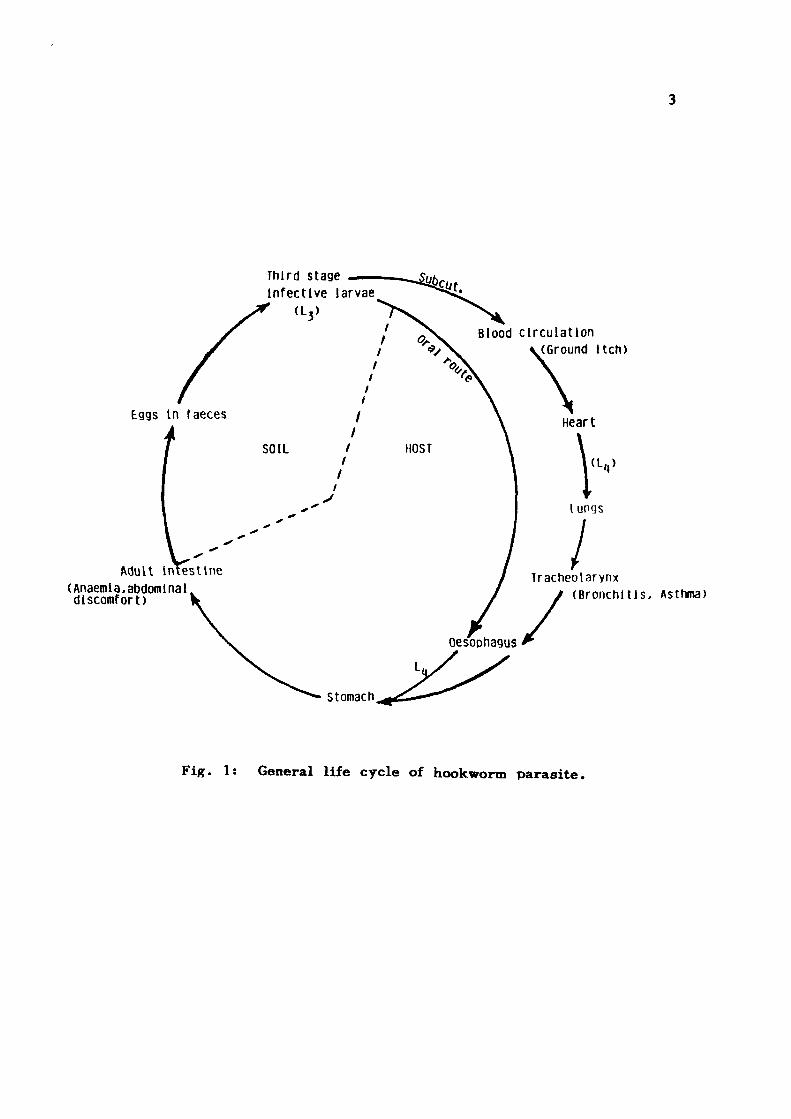
The period from infection to the first appearance of
eggs in the faeces of the host is constant for each species
of hookworm. The ovum undergoe," four moults and second
moult produces an infective stage i . e . the thi rd stage larvae,
L . . This stage after penetrating the skin moves towards lungs
and the th i rd moult takes place. L. larvae with a provisional
buccal capsule, enters the larynx, then moves to pharynx,
oesophageous and finally settles down in the small intestine.
The migratory phase occupies about 7-15% of the prepatent
period. The last moulting occurs in the intestine, the adults
mature, mate and reproduce and thus the cycle continues. Among
human hookworm paras i tes , the shortest prepatent period has
been observed in case of A.ceylanicum (18-26 days) , while
the prepatent period for A.duodenale and N.americanus is longer.
Maximum egg output was observed between 12 and 18 months
of A.duodenale infection in man and the life span of the worms
was 6-7 years (Kendrick, 1934), while that of N.americanus
was reported to be 5-6 years .
There are no specific signs and symptoms in the vast
majority of cases of hookworm infection and their order of
appearance are closely related to the chronology of migration
and development of the paras i tes . The symptoms can con-
Third stage Infective larvae
(L3)
Blood circulation (Ground Itcti)
Eggs In faeces
SOIL
>
Adult Intestine (Anaemia,abdominal, discomfort)
Stomach
Tracheolarynx (Bronchitis, Asthma)
/
Fig . 1: General l i fe cyc le of hookworm paras i te .

veniently be divided into two; those associated with migration
of L and presence of adult worms are primary ones, while
the secondary symptoms are the consequences of physiological,
biochemical and haematological disturbances that follow the
persistence of intestinal infection and constitute what is commonly
known as 'hookworm d i sease ' . A primary heavy infection
will induce ground itch followed by fever, asthmatic wheezing
and a high degree of eosinophilia which will be self limiting.
In later stages, the ancylostomiasis is connected with iron
deficiency anaemia associated with epigastric pain or discomfort
which may be relived by food and mistaken for duodenal ulcer.
The functional consequences of th is disturbed iron-balance
and impaired nutritional status include growth retardation,
reduced working productivity and a potential adverse effect
on learning and cognitive development. Severe intestinal haemo
rrhage due to the infection might lead to a drop in haemoglobin
(Hb) level as low as 2 g/100 g of blood and in erythrocyte
count to below 1000/ml. Chronic and acute, both kinds of
anaemic conditions are observed in the patients of hookworm
infection. Numerous secondary signs and symptoms attributable
to anaemia and hypoproteinaemia accompany chronic hookworm
infection. The most important of these are associated with
progressive cardiac insufficiency and congestive failure. A
wide range of reproductive disorders have also been attributed
to the effects of hookworm infection.

Chemotheraoy based on efficacious anthelmintic drugs
like-Mebendazole, Thiabendazole, Bephinium, Levamisole ,
Tetrachloroethylene etc . remains the starting point of a modern
control programme against hookworms (Crompton, 1989). However,
these drugs are effective against the adult stage of the para
site and repeated doses cause various types of side effects.
Hence, there is an urgent need to evolve an alternate strategy
which should be effective against the infective larval stage
and can check the infection as soon as it occurs. A realistic
approach would be to diagnose the disease at an early stage
as well as to develop immunoproohylactic measures, which
require a good understanding of the molecular immunology of
hookworm infections. These studies will certainly provide
the basis for rational vaccine development as well as for immuno-
diagnosis of the disease. Accordingly, a survey of literature
was made along these l ines.
REVIEW OF LITERATURE
The efforts to understand the molecular and immuno
logical processes underlying hookworm infection have greatly
been limited due to the lack of suitable animal models. Most
of the JjT vivo studies for examining the complete life cycle
of Ancylostnma hookworms require the use of dogs or cats,
of course with the exceotion of some studies which were done
in primates. Most of the studies have been done using these

animal models but the expense and the time required to maintain
the parasites m these laboratory hosts has hampered the studies
on hookworm paras i tes . An important development has been
the establishment of human hookworms in laboratory rodents
so that controlled experimental studies can be made on these
paras i tes . The maintenance of A.ceylanicum m golden hamsters
(Carroll and Grove, 1984) and of N.americanus m neonatal
hamsters (Behnke et_ al^., 1986) has opened up new vistas for
laboratory investigations. This achievement has facilitated
work on antigen characterisation, immunity and vaccine develop
ment. The role of surface antigens and excretory-secretorv
products which include certain metabolic enzymes like proteases,
IS st i l l not well establ ished. By using modern techniques,
these antigens can be well studied and it will help m develop
ing better immunoprophylactic and immunodiagnostic measures.
Studies on immunology of hookworm parasites have mainly been
directed towards the two major aspects - one is immunodiagnosis
dealing mainly with serological or host immune responses and
the other aspect is protective immunity.
Immunodiagnosis
The development of immunodiagnostic methods for hook
worm infection has received a l i t t le attention m view of the
relative ease of diagnosis by faecal examination, but, in fact,
the hookworm eggs may be sometimes misunderstood with the
other intestinal helminth paras i te . Hookworm parasites have

been shown to mediate a wide v a r i e t y of serological react ions
in v i t r o , de tec ted by complement fixation (Magaudda Borzi
and Penn is i , 1958), gel diffusion (Singh, 1965), la tex flocculation
(Rombert et_ al^., 1967); haemagglutination (Desowitz, 1967:
Jayawardene and Viiayaratnam, 1968; Ball and Ba r t l e t t , 1969),
immunoprecipitat ion (Yamanaka, 1960; Ball and Ba r t l e t t , 1969;
Ogilvie et^ al^., 1978) and immunofluorescence (de Azevedo et
a l . , 1971; Lewis et_ al^., 1978). A s tudy of immunological r e
actions during exper imenta l infection of N.americanus was done
long back by Ball and Bar t le t t (1969) . Later on, Poulain et_
a l . (1976) s tudied the local sys temic responses in r a t s infected
with N . b r a s i l i e n s i s by determining t h e an t ibody t i t e r s of both
se ra and in tes t ina l mucosa using p a s s i v e haemagglutination t e s t
and found tha t a cons iderab le port ion of a c t i v i t y was associa ted
with IgA f rac t ion . Specific IgA and o the r i so types of ant ibody
to N . b r a s i l i e n s i s antigens have a lso been de tec ted in sera as
well as in tes t ina l secre t ions by seve ra l o the r workers (See
see et^al_., 1976; Sinski and Holmes, 1977; Brown et_ al_., 1981a,b) .
People have t r i e d to quant i ta te the an t ibody t i t e r s during the
course of infection employing var ious immunological techniques
l ike counter current immunoelect roohores is (Bhopale et_ a l . ,
1981; Men on and Bhopale , 1985), enzyme l inked immunosorbent
assay (Ogilvie et^ al^., 1978) and radioimmunoassay (Sinski and
Holmes, 1978). Counter current Immunoelectrophoresis has been
shown to have a ve ry l imited appl ica t ion in serodiagnos is because
of mult iple helminth infections as well as c ros s - r eac t ions among
var ious hookworm s p e c i e s . Radioimmunoassay has successfuUv

been used for quantifying the specific antibody responses in
rats infected with N.brasiliensis and it was found to be of
local IgA type (Sinski and Holmes, 1978). It has also been
reported by the same workers that the local specific IgA anti
bodies were closely associated with immunity against gastro
intestinal nematode. The local antibody responses (IgA, IgM
and IgG levels) in rats parasit ised with N.brasiliensis infection
were investigated by measuring the levels of coproantibodies
(Wedrychowicz et_ al_., 1983), Recently, the usefulness of radio
immunoassay and reverse enzyme immunoassay using excretory-
secretory products from hookworm parasites has been demons
trated in serodiagnosis and prognosis of hookworm infections
(Ganguly et_ al^., 1988).
The hookworm infections have been shown to evoke
immediate type hypersensit ivi ty reactions as demonstrated by
intradermal tests conducted with antigenic preparations from
various hookworm parasites (de Hurtado and Layrisse, 1968).
The sensitivity of skin reaction was found to be 80% or even
more and some results indicated that the intensity of skin
reaction may be related to the intensity of hookworm infection
(Lobel et_ al^., 1968). In support of these findings a positive
relationship has been shown between the skin reaction (diameter
of the wheel formed) and the level of infection (faecal egg
count) by some workers (de Hurtado and Layrisse, 1968; Lobel
et al_. ,1968). A significant correlation between the wheel size
and eosinophillia and presence of signs and symptoms of hook-

worm infection has also been shown by Ishizaki et a l . (1961).
Antigens used for the skin test include protein or polysaccharide
fractions prepared respectively by t rypt ic digestions of larvae
or adult hookworms (Noda, 1953; Sawada et_ al^., 1954; Wei
and Kuo, 1958; Shih et al^., 1959; Prasad and Mathur, 1962)
or by aqueous extraction (Yamanaka, 1960; Ishizaki et_ a l . ,
1961; de Hurtado and Layrisse, 1968; Lobel et a]_., 1968). Skin
test has been found to be positive in man using antigens from
canine hookworm suggesting the cross reactivity among these
antigens (Sawada et_ al^., 1954, 1961; Yamanaka, 1960). It has
been shown that the sera obtained from hookworm or ascariasis
patients also cross-react well with cestode antigens (Correa-
Oliveira et al^., 1988). The demonstration of these cross r e
actions points out the problems of serodiagnosis. The heat
shock proteins have been shown to have better prospects for
a species specific diagnostic test (Newport et_ al^., 1988).
Immunoprophy laxis
Though substantial evidence is there for the presence
of functional protective immunity during hookworm infection,
but the molecular basis of the protective immune responses
has not been well defined. The strongest evidence for the
acquired immune responses to hookworm infection comes from
an early demonstration that the infective larvae of A.caninum
elicit immunity in dog (Herricke, 1928; McCoy, 1931; Foster,
1935). Different mechanisms have been documented for the
active evasion of the hos t ' s immune responses in many parasitic

10
infections and it is supposed that hookworms also employ similar
strategies for evading or suppressing the host protective res
ponses (Hotez et al_., 1987). The protective immunity to animal
hookworm parasi tes has been investigated in some detail (Miller,
1971; Carroll and Grove, 1984) and has been found to be asso
ciated with both serological (Ball and Bartlett , 1969; Ogilvie
et a l . , 1978) and cellular responses (Ogilvie and Jones, 1973;
Taylor and Turton, 1976; Maxwell et_ a]_., 1987). However, a
strong correlation between these phenomena and host protective
immunity has yet to be conclusively establ ished.
Although, a number of investigations have been con
ducted on the systemic immune responses of host to hookworm
infection (Kassai, 1982) and the possible role of local anti
bodies in conferring immunity to the parasites (Poulain et_ a l . ,
1976; Sinski and Holmes, 1977, 1978; Wedrychowicz et a]_.,
1983a, b , 1984a,b), very l i t t le information is available about
the antigens important in the stimulation of intestinal antibody
responses. It has been suggested that IgA antibodies are in
duced by antigens irrelevant to parasite survival and these
antibodies are supposed to play important role in establishing
host-parasi te relationship rather than evoking protective immunity
(Befus and Bienennstock, 1982). The most striking immunological
property of helminth infections is a capacity to provoke high
levels of IgE and reagin type antibodies in their hosts . Studies
concerning the function of IgE in parasit ic infections began
on the firm basis with the work on intestinal anaphylaxis

11
in N.brasiliensis infected rats (Urquhart et_ a]_., 1965; Jarrett
and Bazin, 1977). Both parasite specific and non-SDecifically
potentiated IgE responses have been reported during N.brasiliensis
infection m rats (Ogilvie, 1967; Orr and Blair, 1969; Bout
et a l . , 1977). However, subsequent studies established that
non-specific IgE does not protect against allergic reactions
as the passive transfer of immunity could be achieved using
antisera devoid of anaphylactic activity (Jones et_ al^., 1970;
Leid and Williams, 1974; Jarret t et_ a]_., 1980).
Various mechanisms have been proposed for the induction
of protective immunity in hosts; these are antibody mediated
immunity, cell-mediated immunity, and lastly the local inflamma
tory responses (Jarret t and Urquhart, 1971; Ogilvie and Jones,
1973; Love et a l . , 1974). Immunity to mouse hookworm N.dubius
is believed to have two stages: antibodies damage the worms
which are then predisposed to antibody independent expulsion
from the gastrointestinal tract (Prowse et_ al^., 1978). This
self cure phenomenon seems to be achieved by a combination
of all the three mechanisms proposed earl ier for protective
immunity. However, passive transfer of protective immunity
to mice with immune sera against N.dubius infection, demons
trated that serum antibody mediate protective immunity (Jones
and Robin, 1974; Dobson and Owen, 1978). Later on, it was
suggested that passive transfer of anti-N.dubius serum directly
or indirectly influenced the survival , growth and fecundity
of N.dubius (Dobson, 1982). It has also been found that the

12
pas s ive t r ans fe r of IgG, - fraction of anti-N .dubius «?erum
pro tec ted the mice and resu l t ed in the r e t a rded growth of
the p a r a s i t e s . When combined with adop t ive ly t rans fe r red
immune mesentr ic lymph node c e l l s , enhanced worm expulsion
from the in tes t ine was also obse rved ( P r i t c h a r d e^ a l . , )983).
According to the e a r l i e r r e p o r t s , i t was suggested that worms
p rev ious ly damaged by the host immune responses we>-p found
to be more suscep t ib le to expulsion mechanism as rompar-'-d
to the undamaged worms (Ofrilvie and Love, 1974). However,
in some c a s e s , the worms taken from the hos ts shor t ly before
expulsion and t r ansp lan ted to the o ther hos ts were no more
suscep t ib le to expuls ion (Hopkins et_ al^., 1972; Hopkins and
Zajac, 1976: Wakelin and Wilson, 1979). It has been suggested
tha t worm expulsion was due to the metabolic c r i s i s through
which the worms were going and the findings support the
en te roa l le rg ic ind i r ec t mechanism for worm reiect ion (Kassai
et a l , , 1987). Recent ly , some work has been done regarding
the expuls ion phenomenon of N . b r a s i l i e n s i s and free r ad i ca l s
(oxygen r a d i c a l ) a re sa id to be involved in the expulsion of
p r imary infections (Smith and Bryant , 1986; Smith, 1989).
Hookworm p a r a s i t e s have a lso been shown to evoke
cel l mediated immune responses (Ali et_ a^. , 1986). Infection
with A .duodenale has been found to be assoc ia ted with marked
cell mediated immunity as manifested by increased T-cel l counts,
increased inhib i t ion of leucocyte migration and increased lympho-
b las t t ransformat ion . An inve r se r e l a t i o n s h i p was proven between

13
the degree of cel l mediated immune responses and the oa ras i t e
load . Fu r the rmore , some o the r group of workers have also
r e p o r t e d the p r o b a b l e role of cel l mediated immunity m vacc i
nated an imals . According to t he se worke r s a delayed hyper
sens] t ]v i tv react ion has been obse rved m pups vaccinated witbi
i r r a d i a t e d infect ive l a rvae of A .caninum and t h i s skin reaction
has h<-f"ti reoor ted to cor re la te with th*- jnhibi t ion ot rhif msH'"*
tiop oi leucocytes in the presence of larvae or adult worm
antigen (Gautum et_ aJ^., 1986).
A successful induction of p ro t ec t i ve immunity with
soluble e x t r a c t s from adul t s of A.caninum and A.ceylanicum
has been shown by the reduction in worm burden bv 59% and
741 r e s p e c t i v e l y (Thorson, 1956; Car ro l l and Grove, 1984, 1986).
Dogs immunised with soluble e x t r a c t s p r e p a r e d from adult worms
have been found to be p a r t i a l l y r e s i s t an t to challenge infection
with L^ of A.ceylanicum (Car ro l l and Grove, 1985). Double
dose of i r r a d i a t e d L- l a rvae of A.caninum gave 90% protection
to subsequent chal lenge (Mi l le r , 1965, 1971). The route ol
adminis t ra t ion i s also an important f ac to r . in t raper i toneal
or subcutaneous route has been shown to be les^ effective as
compared to oral adminis t ra t ion which gave 100% reduction in
worm burden (GuDta and Kat iya r , 1985). Menon and Bhonale
(1985) have vaccinated the golden hamste r s with i r r a d i a t e d
larvcie of A.ceylanicum and obtained 95% reduction in worm
burden . However, such vaccines p r e p a r e d from i r r a d i a t e d
at tenuated l a rvae cannot be employed in human beings because

14
of some e th i c l e r e a sons , such as the danger of vaccinated host
becoming infec ted .
Characterization of antigens
Attempts to analyse the somatic e x t r a c t s from adult
and l a r v a l s tages of A.caninum and t h e i r c ross r e a c t i v i t v with
sera obtained from N.americanus infected pa t i en t s were made
long back by Williams (1970), using immunoelectrophoret ic t ech
nique . The l a r v a l antigens could be sepa ra ted into 18 p rec ip i t in
bands whereas the adul t antigenic p repa ra t ion showed the p r e
sence of 13 bands with homologous immune r a b b i t serum. Only
one < ro^s r eac t ive component wa'? observod bftwron A.raniniim
and N.americanus employing pat ient s e r a , whereas the e x t r a c t s
from N.americanus adul t worms have been r epor t ed to show
four p r e c i p i t i n bands (Will iams, 1970). In contrast to these
f indings , Desowitz (1962) , employing Ouchterlony gel diffusion
t e s t has shown more r e l a t i o n s h i p between the same stage of
different spec i e s than between the di f ferent s tages of the same
s p e c i e s . Later on, a compara t ive ana lys i s of p ro te ins of
A.ceylanicum and A . b r a s i l i e n s i s was done employing disc gel
e l e c t r o p h o r e s i s and an over a l l s im i l a r i t y between the protein
pa t te rn of the two p a r a s i t e s was obse rved (Yoshida et^ a l . ,
1977), Antigens of 250, 60, 45 and 30 kD have been found
to be common between the adul t and l a r v a l s tages of N.dubius
(Hurlev et^ a]_. , 1980; Mitchell and Munaz, 1983; Mitchel] and
Cru i se , 1984; Monroy et^ £!.• » 1985; Monroy and Dobson, 1* 86;
Adams et a l . , 1987). Affinity pur i f ied fraction of adult worms

15
containing 55 and 20 kD antigen has been shown to induce oro
tection against infection with N.dubius (Monroy and Dobson,
1987). Charac te r iza t ion of antig-ens of N . b r a s i l i e n s i s adult
worm have also been done using monoclonal an t ibod ies (Bohn
and Konig, 1985). The antigenic pa t te rn of different d^veloo-
mental as well as the stage specif ic antigens of N.americanus
have been s tud ied using immune sera from human (natural host)
and hamste rs (Carr and P r i t c h a r d , 1987).
The antigens of nematode p a r a s i t e s have bc^n shown
to be excre ted or secre ted from the internaJ ce l lu la r and gJan
dular -structures during in v i t r o maintenance ot parasite->. Ih'=*se
e x c r e t o r y - s e c r e t o r y (L-S) antigens a re supposed to be Les"
heterogenous as compared to somatic e x t r a c t s and have been
found to play important ro le m diagnosis as well as m pro tec t ive
immunity. The role of F-S produc ts from hookworms m inducing
immunit ; *as repor ted by Otto and Kerr (1939j about a half
century ago. The e x c r e t o r y - s e c r e t o r y products of rat hookworm
pa ra s i t e N.muris have been shown to induce p ro t ec t i ve a n t i
bodies (Thorson, 1951). Later on, the p ro t ec t i ve role of
E-S products from a number of hookworm p a r a s i t e s was s tudied
by many workers and significant degree of protect ion was o b s e r
ved in p a r a s i t i s e d hos ts (Clegg and Smith, 1978; Iain et a l . ,
1979; Wedrychowicz et_ al_. , 1983). The immunogenicity of P-S
products from hookworm p a r a s i t e s was compared with that of
somatic antigens and it was suggested by many workers that
E-S produc ts were more immunogenic (Kat iyar et a l . , 1972 ; Goviia

16
et_ a]^. , 1973; Murray et_ al_., 1979). The E-S antigens of human
hookworm N.americanus have been analysed emplovine; immune-
blotting technique using serum from infected hamsters as well
as from the patients suffering from hookworm infection (Carr
and Pr i tchard, 1986, 1987). Ey (1988) has identified and charac
terized the E-S products from exsheathed L- of Heligmosoides
polygyrus (N.dubius) and has also studied the role of E-S
products m protection. Besides, hookworms the E~S antigens
of other nematodes like T.canis (Sugane et_ al^., 1985), B.malayi
(Kaushal et_ al^., 1982, 1984), W.bancrofti (Maizels et al^., 1986),
S.cervi (Malhotra et_ al_., 1987) and Strongylus stercoralis(Brindlev
et al.,1988) have been successfully characterised using immuno-
electrophoretic and other immunochemical techniques.
The sex specific antigens present m the secretionh
of N.dubius adults have also been studied (Adams et aj^., 1988).
The E-S products of adult N.dubius have been shown to reduce
the ratio of male worms to female worms. A highly immuno
genic molecule of 66 KD has been found to be present in the
secretions and also on the surface of males only and has been
supposed to induce protective immunity specific to male worms,
thus resulting in the less chances for mating and so the reduct
ion in worm burden. The evidence for the cuticular contri
bution to E-S products comes from an early demonstration that
N.americanus sheds significant amount of i ts surface antigen
into the medium during jji vitro incubation (Pritchard et a l . ,
1985). The adsorption of E-S antigens onto the surface has

17
been shown in case of T.spiral is (Silberstein and Despommier,
1985). Rats immunized with E-S products from Strongylus
stercoralis adults showed reduction in faecal ego; count as well
as adult worm burden (Mimori et_ al^., 1987), thereby suggestini?
the protective role of E-S products.
Surface antigens have also been shown to play a key
role in the induction and expression of host immune responses
(OgiJvie et ai^., 1980; Maizels et al^., 1982, 1983). The cuticle
of helminths including hookworms have been shown to be prone
to damage by antibody and complement and by many of the
antibody dependent cellular cytotoxicity reactions (Kazura and
Grove, 1978; Mackenzie et_ al_., 1978; Kazura and Aikawa, 1980;
Ogilvie et a l . , 1980). The stage specific surface antigen of
N.brasiliensis have been investigated using surface labelling
125 with 1, solubilisation with detergent followed by immuno-
precipitation and SDS-polyacrylamide gel electrophoretic analysis
(Maizels and Ogilvie, 1980; Ogilvie and Phi l l ip , 1980). The
cross reactions between the surface proteins of various nematodes,
which differ completely in their molecular weight characteris t ics ,
have also been documented by several investigators. Maizels
et a l . (1982) suggested that carbohydrate side chains could
be responsible for the cross-react ivi ty . The cuticular antigens
from N.americanus adults were studied by Pritchard et a l .
(1986), using sera from infected hamsters as well as human.
A 67 kD antigen of N.americanus has been reported to be pro
duced in vitro also in measurable amounts (Carr and Pritchard,

18
1987) suggesting t h e r e b y the involvement of cut icular antigen
m inducing immune r e s p o n s e s . The surface antigens of
N.americanus have been shown to be h igh ly immunogenic and
have been found to be collagenous in nature which i s the mam
s t ruc tu ra l component of p a r a s i t e cut ic le (Keymer and Bundv,
1989). It has a lso been suggested recent ly bv P r i t cha rd et
a l . (1988a) tha t natural immune response against surface soeoi t ic
epitopp'- could be boosted with purif ied surface antigens TO
induro i sufficiently strong and p'^rsistr>nt r e s p o n s e s . It has
also been suggested tha t small collagenous molelcules and a s s o
ciated d i su l f ide bonds hold toge ther the cut ic le of N.americanus
( P r i t c h a r d et a l . , 1988b).

19
SCOPE AND PLAN OF WORK
Hookworm disease is a major public health problem
affecting millions of people and causing profound morbidity
and mortality. Chemotherapy has emerged as the only proce
dure for temporary cure of the disease. However, repeated
doses of anthelmintic drugs are toxic and cause various types
of side effects. Therefore, the knowledge about molecular
immunology of hookworm infection is of urgent need as it
may provide certain leads for development of vaccines.
Several hookworm species are known to infect men
and l ivestocks. Due to the strong host specificity, Necator
americanus and Ancylostoma duodenale; the two human hookworms
are less amenable for the s tudies. Another hookworm parasite
Ancylostoma ceylanicum, (which infects man besides cats and
dogs), being maintained in golden hamsters (Mesocricetus auratus)
has extensively been used for chemotherapeutic and immuno
logical s tudies. Many efforts have been made for studying
immune responses and protective notential of hookworm para
s i tes , but the antigens responsible for evoking these r e s
ponses are not well defined. The characterisation of antigens
will provide the basic information not only for understanding
the host-parasi te relationship but also for identitying the
antigenic targets of protective immunity. Hence, in the present
study an endeavour has been made to analyse the somatic
and excretory-secretory antigens of A. ceylanicum. The present
dissertation deals with the following aspects:

20
I. Antigenic analysis of somatic extracts from adult and
infective larval stages of Ancylostoma ceylanicnm
1 . Protein pattern of adult and larval stages
of A.ceylanicum.
2. Immunisation of rabbi ts with somatic extracts and
quantitation of parasite specific antibodies in immune
sera .
3 . Antigenic analysis of adult and larval stages of
A.ceylanicum using immune rabbit sera .
4 Identification of common or cross reactive as well
as stage/parasite specific antigens of adult and larval
stages of A.ceylanicum.
5. Immunisation of hamsters with adult and larval somatic
extracts of A.ceylanicum and evaluation of immunised
hamsters for protective immunity.
II. Antigenic analysis of excretory-secretory products of adult
A.ceylanicum
1. Short term jji^ vi t ro maintenance of A.ceylanicum adult
worms and preparation of excretory-secretory products.
2. Immunisation of rabbi ts with excretory-secretory pro
ducts and quantitation of parasite specific antibodies
in immune sera .
3. Antigenic analysis of excretory-secretory products
using immune rabbit sera .

21
4. Analysis of common or cross reactive antigens between
excretory-secretory products and somatic extracts
from adult and larval stages.
5. Immunisation of hamsters with excretory-secretory
products and evaluation of immunised hamsters for
protective immunity.

CHAPTER II
MATERIALS AND METHODS

22
PARASITES
Ancylostoma ceylanicum
Ancylostoma ceylanicum infection was or ig ina l ly procured
from Sarabha i Research Cent re , Baroda, Ind ia . The infection
was maintained in golden hamste r s (Mesocricetus aura tus) weigh
ing 60-80 g by giving an inoculum of 60-65 infective l a rvae
( L - ) . Infected hams te r s were k i l l ed 21 days post infection
and the adul t worms were col lected from the mucous laver
of small i n t e s t i ne , p a r t i c u l a r l y the jejunal port ion by cutting
i t longi tudinal ly throughout i t s l eng th . The worms were then
washed thoroughly with isotonic sal ine containing ant ibiot ic
mixture (penic i l l in 100 U/ml + qtreptomycin 80 \)g/ml) to rprnovp
the adher ing in tes t ina l d e b r i s and bac t e r i a l contamination.
The clean worms were used immediately for antigenic p r epa ranon
or fcepT frozen a1 -20 C until u sed .
infect ive l a rvae of A. ceylanicum were collected trom
the coprocul tures of infected h a m s t e r s , 20-30 days post infect
ion according to the method of Ray et_ al_. (1972). The faecal
pe l l e t s were emulsified with d i s t i l l e d water to a th ick pas te
which was then sp read over the centre of a moist Whatman
No.l f i l t e r p a p e r , kept in p e t r i d i s h e s and incubated in a humid
chamber at 27±2 C for about 5-6 d a y s . The l a rvae thus col lected
were suspended in d i s t i l l e d water containing an t ibo i t i c s and
washed thoroughly to remove extraneous mater ia l and the imour i -
t i e s of faecal ma t t e r . The clean l a rvae were then subjected
to mi l l ipore f i l t ra t ion (5 um) ui o r d e r to remiove the bacter ia l

23
contamination. The bacteria free larvae were recovered by
low speed centrifugation and kept frozen at -20 C until used.
Nipp>ostrongylus brasiliensis
Nippostrongylus brasi l iensis infection obtained from
Wellcome Laboratories of Tropical Medicine, London, were main
tained in albino ra ts (UF strain, 25-35 g ) . Rats infected
with an inoculum of 2000 L- of N. brasi l iensis were kil led,
7-10 days post infection. The intestines were cut longitudinally
throughout the whole length and the clean adults , sticking
to the mucosal layer of jejunum, were obtained by baermanni-
sation of the suspension through muslm cloth. The adults
thus obtained were washed with lukewarm isotonic saline contain
ing antibiotic mixture and stored frozen at -20 C until used.
Infective larvae of N.brasiliensis were collected essent
ially according to the method of Keeling (1960). Coprocultures
were made from rats infected with N.brasi l iensis , 10-15 days
post infection. The paste of faecal pel le ts , made with isotonic
saline was mixed with granular charcoal in a ratio of 1:3.
The paste was then spread over the centre of moist filter
papers kept m petr i dishes and incubated at 27±2°C in a humid
chamber. The larvae developed after 4-5 days and migrated
towards the edge of the fil ter paper on attaining the infective
larval stage. The motile larvae were harvested by trimming
the edges of fil ter papers and baermannisation of the larval
suspension. The larvae were washed repeatedly with isotonic

14
sal ine containing an t ib io t ic mixture (pen ic i l l in 100 U/ml and
s t reptomycin 80 ug/ml) and s to red frozen at -20 C until used .
CHEMICALS USED
Acrylamide , aga rose , b i s a c r y l a m i d e , bovine serum
albumin, Coomassie Br i l l i an t Blue (R-250) , goat a n t i - r a b b i t
Ig p e r o x i d a s e and TEMED were procured from Sigma Chemical
Company , St . Louis , MO, U.S.A. The powdered media DMEM
and RPMI-1640 were obtained from Gibco Labora to r i e s , New
York, U.S.A. Ammonium pe r su l f a t e , beta mercaptoethanol were
t h e p roduc t s of E .Merck , A.G. Darmstadt , West Germany. F r eund ' s
Complete and Incomplete adjuvants were obtained from Difro,
Detro i t , MI, USA. All o the r chemicals used were of analvt ica]
grade and t r i p l e glass d i s t i l l e d water was used for a l l the
e x p e r i m e n t s .
PREPARATION OF MEDIA
Rosewell Park Memorial Institute 1640 (RPMI-1640):
The powdered medium supplemented with L-glutamine
(10.4 g) was d i s so lved in approx imate ly 900 ml of water and
then 5.94 g HEPES was added and the final volume was made
upto 960 ml . The medium was s t e r i l i s e d by f i l te r ing through
0.22 urn membrane f i l t e r s . P r i o r to use 4.0 ml of 5% s t e r i l e
sodium bicarbonate was added pe r 96 ml of medium.
Dullbeco's Minium Essential Medium (DMEM):
The powdered medium with E a r l e ' s sa l t and L-glutamine
was d i s so lved in approx ima te lv 900 ml of double d i s t i l l ed

25
water. Sodium bicarbonate (3.7 g) was then added and the
volume of the solution was made upto one l i t r e . The medium
was steril ized as described for RPMI-1640.
Ringer's Solution:
Weighed quantities of sodium chloride (9 g) , potassium
chloride (0.42 g) , glucose (0.25 g) and Ca.n]2 (0.42 g) were
dissolved in a minimum amount of water. After raising the
volume upto 900 ml, separately dissolved Tris (1.21 g) was
added and the volume was made upto one l i t r e . This was
then steri l ised as described for RPMI-1640.
PREPARATION OF ANTIGENS
Somatic antigens
Soluble antigenic extracts from adult and larval stages
of A.ceylanicum and N.brasil iensis were prepared essentially
according to the method described by Kaushal et al_. (1982)
for the preparation of filarial antigens. Briefly, lyophilised
adults/infective larvae were ground to fine powder in a pestle
and mortar and suspended in isotonic saline (0.15 M NaCl).
The suspension was sonicated 4 times for 1 min each with
an interval of 1 min using an ultrasonic cell disruptor (Heat
systems, Ultrasonics Inc. Ltd. NYW-220 F) at 20 Kc/sec, and
extracted on ice for 4-5 h, with continuous gentle st irr ing.
The extracts were then centrifuged at 10,000xg for 30 min
and the supernatants thus obtained were used as a source of

26
somatic antigens. These antigenic extracts were kept frozen
at -20 C in small aliquots until used.
Excretory-secretory antigens
Fresh, motile worms obtained from the infected hamsters
were thoroughly washed with isotonic steri le saline containing
a mixture of antibiotics (penicillin and streptomycin). Separate
batches of 100 adult worms were incubated at 37 C in 20 ml
of three different media (Ringer's solution, DMEM and RPMI-
1640) containing penicillin (100 U/ml) and streptomycin (80
ug/ml). The media collected at the end of incubation period
were lyophil ised, dialysed and used as a source of excretory
secretory antigens.
Immune rabbit sera
Polyvalent hyperimmune sera were raised in albino
rabbi ts (weighing 1.0-1.5 kg) against somatic extracts prepared
from adult and larval stages of A.ceylanicum and N.brasi l iensis .
Rabbits were immunised intramuscularly with an emulsion of
somatic extracts (1 mg protein) made in Freund's complete
adjuvant. One month after the first injection, three subsequent
injections were given at one week interval and the rabbits
were bled from the marginal ear vein, one week after the
fourth injection. After tha t , immunisation and bleeding of
the rabbi ts was done alternatively at weekly intervals t i l l
the hyperimmune serum was obtained. The collected blood
was allowed to clot by keeping at 37°C for 1 h and then left
overnight at 4 C. The serum was collected by centrifuging
at 800xg (10 min) and stored at -20 C in small aliquots.

27
Polyvalent hyperimmune serum afi;ainst e x c r e t o r y - s e c r e t o r v
p roduc ts from A.ceylanicum adu l t s was a lso r a i s ed in r a b b i t s
( 1 . 0 - 1 . 5 kg) by giving 0.5 mg prote in emulsified in Freund' s
complete ad juvant . The immunisation and bleeding schedule
followed was same as d e s c r i b e d for somatic an t igens .
For ra i s ing an t ibod ies against normal hamster serum,
r a b b i t ' were immunised with 1.0 mg prote in ot normal hamster
>eruni eraiusified with Freund ' s complete ddiuvanx: a?- dfescrit-ed
lor somatic an t igens . The serum obta ined was s tored in small
a i iquots at -20 C until used .
PREPARATION OF GAMMA GLOBULIN FRACTION
Gamma globulin fraction from immune r abb i t sera was
p r e p a r e d by adding ammonium sulfate (45% sa tura t ion) to the
serum with continuous gentle s t i r r i n g at room tempera ture for
30 min and then overn ight at 4 C. The p r e c i p i t a t e obtained
was recovered by centrifugation at 10,000xg for 30 min and
d i s so lved in minimum amount of isotonic s a l i n e . The ammonium
sulfate was removed by d ia lys ing against t h r e e changes of
isotonic s a l i ne . The gamma globulin fraction was centrifuged
at 800xg for 15 min and s tored at -20 C in small a i i q u o t s .
IMMUNISATION OF HAMSTERS
Golden hamste rs (Mesocricetus a u r a t u s ) , weighing M~
80 g were immunised with somatic e x t r a c t s (100 ug prote in)
ot A«^.^_l£nicum adult and l a rvae and E-S products (SO ug orutein)
ot A.ceylanicum adul t p a r a s i t e emulsif ied with F r e u n d ' s complete

28
ad iuvan t . The immunisation was c a r r i e d out by giving a total
of five injections subcutaneously at 15 days i n t e r v a l s . Hamsters
were b led one week af ter each injection and the sera collected
were s to red at -20 C unti l u sed . One week after the 5th in
jection the hams te r s were challenB;ed with 60 L^. The animals
were k i l l e d 21 days post chal lenge and the worm burden was
recorded and compared to unimmunised infected con t ro l s .
ENZYME LINKED IMMUNOSORBENT ASSAY
Enzyme l inked immunosorbent assav (ELISA) was p e r
formed according to VoUer et_ al_. (1974) . Br ie f ly , 100 ul
(0.015 ug - 1.0 ug) of A.ceylanicum adult somat ic / l a rva l somatic/
adult e x c r e t o r y - s e c r e t o r y antigens d i lu ted in phospha te buffered
sal ine (0.01 M, pH 7.2) were added to each well of a mic ro-
t i t r e ELISA p la te and incubated for 1 h at 37 C and then at
4°C ove rn igh t . The p la t e was washed with PBS-Tween ( p h o s
pha te buffered sal ine containing 0.05% Tween-20) to remove
the unbound ant igen. Two hundred mic ro l i t e r s of 1% BSA (made
in O.OIM PBS, pH 7.2) were added to the wel ls and the p la te
was incubated for 1 h at 37 C in o r d e r to block the uncoated
s i t e s . The p la t e was washed t h r i c e with PBS-Tween. Different
d i lu t ions of serum (100 ul) were then added to the wells and
p la te was kept at 37 C for 2 h . After washing t h r e e t imes
with PBS-Tween the p la te was incubated at 37 C for 1 h with
100 ul of pe rox idase l abe l l ed conjugates of goat a n t i - r a b b i t
Ig ( 1: 1000) / rabbi t an t i -hams te r Ig (1 :500, p r epa red in our
l a b ) . After t u r t h e r washing 100 ul of subs t r a t e solution

29
(1 mg/ml 0-phenylene diamine in c i t r a t e buffer 0.05 M, pH
5 .2 , containing 1 u l /ml of hydrogen pe rox ide ) was added to
each we l l . The react ion was s topped af ter 10 min with 50
ul of 5N H^SO. and the r e s u l t s were r eco rded as extinction
at 492 nm. The amount of colour change (OD) wil l be p r o p o r
t ional to the amount of ant ibody in the t e s t serum.
SDS-POLYACRYLAMIDE GEL ELECTROPHORESIS
SDS-polyacrylamide gel e l e c t r o p h o r e s i s (SDS-PAGE)
of A.ceylanicum adul t and l a r v a l somatic e x t r a c t s was performed
according to the method of Laemmli (1970) using Bio-rad s lab
gel e Jec t rophores i s appa ra tus {Bio-rad Labora to r i e s , U . S . A . ; .
A separa t ing gel of 10-20% acry lamide concentration was made
with the he lp of a gradient m i x e r . The composition of 10%
acry lamide gel mixture was : 5 ml of s tock acry lamide (30%):
b i s ac ry lamide (0.8%) mix tu re , 3.75 ml of 1.5 M Tris-HCl
(pH 8 . 8 ) , 0.15 ml of 10% SDS, 0.05 ml of 10% ammonium per
su l fa te , 0.005 ml of t e t r ame thy le thy lene diamine (TEMED) in
the to ta l volume of 15 ml . The 20% acrv lamide gel mixture
contained 10 ml of s tock a c r y l a m i d e : b i s ac ry lamide and 1.8
ml of g l y c e r o l . The r e s t of the components were added to
the mixture in t h e same amount as d e s c r i b e d above for 10%
ac ry lamide gel m i x t u r e . The volume was made upto 15 ml
wxth d i s t i l l e d w a t e r . The s tacking gel (3%) contained 1.0
ml of ac ry lamide stock solution (30% a c r y l a m i d e : 1.0% bis acry la
m i d e ) , 2.5 mi of 0.5 M Tris-HC] (pH 6 . 8 ) , 0.1 mi of 10% SDS,
2.5 ml of 60% suc rose , 0.05 ml of 10% ammonium per sulfa te ,

30
0.005 ml of TEMED in a to ta l volume of 10 ml. The samples
were p r e p a r e d by mixing equal volume of the antigen ex t r ac t s
(100-150 ug/20 ul) with SDS sample buffer (containing 10 ml
g lyce ro l , 5 ml of be t a -mercap toe thano l , 23 ml of 10% SDS,
1 ml ot 0.1% bromophenol b lue , 8.32 ml of 0.5 M Tris HCl,
pH b . 8 , in a to ta l volume of 50 ml) in 1:1 ra t io and keeping
for 5 min in a boil ing water b a t h .
E l ec t rophore s i s was c a r r i e d out at a constant current
of 25 mA per gel for about 4-5 h t i l l the t racking dye reached
to 1 cm above to the bottom of the ge l . After e l e c t r o p h o r e s i s
the gel was s ta ined with 0.25% Coomassie Br i l l iant Blue (R-
250; made in 50% methanol p lus 10% acet ic ac id) and desta ined
with 30% methanol p lus 7% acet ic a c i d .
IMMUNOELECTROPHORESIS
Immunoelectrophoresis ( lEP) was performed according
to the method of Graber and Williams (1953) . The somatic
e x t r a c t s and E-S p roduc t s from A.ceylanicum were separa ted
e l e c t r o p h o r e t i c a l l y in 1.5 mm th i ck l a y e r of 1% (w/v) agarose
made m b a r b i t a l buffer (20.0 mM d ie thy l b a r b i t u r i c ac id ,
80.0 rriM T r i s , 2.0 mM sodium a z i d e , 0.8 mM calrmm lact^ti",
pH 8.b( toy i n at 100 v o l t s . After eiectrophoresi-^ nne aidT«<-.
were Kept over night in a moist chamber and the separa ted
antigens were allowed to react with immune r abb i t s e r a . The
p la tes were then washed with isotonic sal ine for 30 min and
p r e s s e d under s eve ra l shee t s of f i l t e r paper for 10 min. This
p rocedure of a l t e rna te washing and p ress ing was repea ted t h r i ce

31
and the p l a t e s were washed with d i s t i l l e d water before the
final p r e s s i n g . After a i r d ry ing the p l a t e s were s tained with
0.25% (w/v ) Coomassie Br i l l ian t Blue (R-250) made in 45% ethanol
p lus 10% acet ic acid and des ta ined with t h e same.
CROSSED IMMUNOELECTROPHORESIS
Crossed immunoelect rophores is (CIE) was performed
according to the method of Axelsen et_ al_. (1973). The antigens
were f i r s t s epa ra t ed e l e c t r o p h o r e t i c a l l y in the f i r s t dimension
m 1.5 mm t h i c k l a y e r of 1% (w /v ) agarose at 10 V/cm. A longi
tudinal s t r i p (1x7 cm) containing t h e sepa ra ted antigens was
cut and t r a n s f e r r e d to t h e another p l a t e (7x9 cm) . Anodic
(5x7x0.12 cm) and ca thodic (2x7x0.12 cm) gels containing immune
serum (5-20%) were then poured on the p l a t e . An in termediate
gel s t r i p of 1x7x0.13 cm without an t ibody was also poured
m between t h e f i r s t and t h e second dimension g e l s . The an t i
gens were then al lowed to move e l e c t r o p h o r e t i c a l l y m second
dimension for 18-20 h at r i gh t angle to the f i r s t dimension
gel (2-3 V/cm) . After e l e c t r o p h o r e s i s the p la tes were washed
and s ta ined in t h e usual way as d e s c r i b e d for lEP.
CROSSED LINE IMMUNOELECTROPHORESIS
Crossed l ine immunoelectrophoresis (CLIE) is the modi
fication of CIE and was c a r r i e d out in a manner s imi lar to
tha t of CIE except tha t m CLIE the t e s t antigen (protein :
50-100 ug) was added m the in te rmedia te ge l . After the e lect ro
ph o re s i s the p l a t e s were washed and s ta ined in a way s imi lar

32
to that of lEP. On staininf?, the antigenic peaks which give
lines at their bases are considered to be common between the
two antigen ext rac ts .
TANDEM CROSSED IMMUNOELECTROPHORESIS (TCIE)
This is the modification of crossed immunoelectrophoresis
in which the two antigen samples were mixed together and
placed in one application hole. The electrophoresis was carried
out in the same way as for CIE. The common antigens from
the two samples fuse to give an increase in the heights of
the peaks. The control plate contained only the standard
antigen. The washing and staining procedures were exactly
the same as described earl ier for lEP.
PROTEIN ESTIMATION
Protein contents of the somatic extracts from adult
and infective larvae and excretory-secretory products were
estimated according to the colorimetric method of Lowry et
a l . (1951).

CHAPTER III
RESULTS AND DISCUSSION

SECTION I
ANTIGENIC ANALYSIS OF SOMATIC EXTRACTS
FROM ADULT AND INFECTIVE LARVAL STAGES
OF ANCYLOSTOMA CEYLANICUM

33
The somatic antigens from the adult and larval stages
of hookworm parasi tes are capable of inducing various types
of serological reactions but the antigens involved in these
responses have not been well defined. Characterization of
parasite antigens is of particular significance not only for
understanding the host parasite relationship but also for identi
fying the immunologically important components of the parasi tes.
Due to the differences in the habitat and nutritional require
ments, the protein and antigenic compositions of different
parasites as well as of different developmental stages of a
parasite may be different. Therefore, a basic knowledge
about the total protein and antigenic make up of different
stages of a parasite is a necessary pre-requisi te and may
provide useful leads about the antigens having role in pro
tective immunity. Accordingly, in the present study, detailed
investigations were carried out to analyse the protein and
antigenic mosaic of adult and infective larval stages of
A. ceylanicum employing certain biochemical and immunochemical
techniques.
PROTEIN PATTERNS OF ADULT AND LARVAL STAGES OF
A. CEYLANICUM
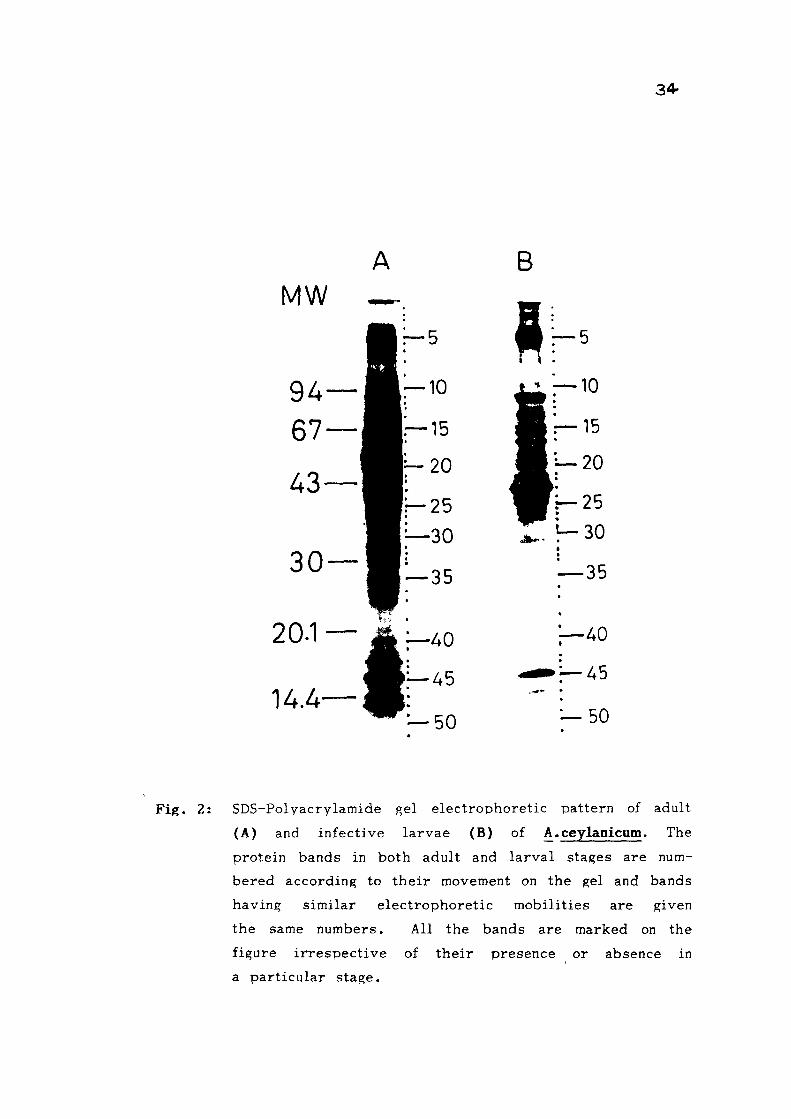
Somatic extracts from adult and infective larvae of
A. ceylanicum were analysed on SDS-PAGE and their protein
patterns are shown in Fig. 2. The molecular weight of various
proteins present in these extracts was determined by running
xn paralleJ the standard proteins of known molecular weights.
34-
A B MW
94 67
43-
30
r~5
r-10
[—IS
i -20
i -25
— 35
f—AO
i—45
'—50
J/k*"'
r -10
'r-15
20
i—25 30
— 35
40
i—45
•—50
Fig. 2: SDS-Polvacrylamide gel electrophoretic pattern of adult
(A) and infective larvae (B) of A. ceylanicum. The
protein bands in both adult and larval stages are num
bered according to their movement on the gel and bands
having similar electrophoretic mobilities are given
the same numbers. All the bands are marked on the
figure irrespective of their presence or absence in
a particular stage.

35
The protein bands were numbered according to their electro-
phoretic mobilities and the bands having identical electro-
phoretic mobilities in both the stages were given the same
number (Table 1 ) . Fourty seven protein bands in Ac A and
43 protein bands in AcL were observed in the molecular
weight range of 10-170 kD. Most of the protein bands detected
in L_ were also present in adult somatic extracts with minor
differences in thei r intensit ies, however, 8 protein bands
specific to adults and 4 bands specific to larvae were observed.
ANTIGENIC ANALYSIS OF ADULT AND LARVAL STAGES OF
A.CEYLAmCUM
( i ) Quaotitation of parasite specific antibodies in immune
rabbit sera by enzyme linked, immunosorbent assay
The antibody t i t e r s of immune rabbit sera raised against
the somatic extracts from adult and larval stages were deter
mined by enzyme linked immunosorbent assay (ELISA). The
assay system was standardised in terms of optimum concentration
of antigen, antibody-conjugate, o-phenylene diamine (OPD)
and hydrogen peroxide (H_0_). Out of different dilutions
of goat ant i-rabbit immunoglobulin-peroxidase conjugate tested,
1:1000 dilution showed optimum resul ts . This dilution of
the antibody conjugate was used in further experiments. Different
concentrations of OPD and H_0_ were also t r ied and 1 mg/ml
of OPD and 1 ul/ml of H^O was found to give optimum ELISA
specific ac t iv i ty . The optimum concentrations of AcA and

36
Table 1: Comparat ive ana lys i s of p ro te ins of aduJt and in
fect ive l a rvae of A. ceylanicum.
Protein band number
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
Apparent molecular mass (kD)
170.0
160.0
153.0
145.0
140.0
130.0
125.0
120.0
100.0
94.0
88.0
85.0
80.0
73.0
66.0
64.0
60.0
55.0
47.0
45.0
43.0
42.0
40.0
39.0
38.0
37.0
36.0
35.0
34.0
33.0
32.0
31.0
Adult
-
-
-
++
+ + +
++
+
+
++
++
+ + +
-
+
+ + +
++
+ + 4
+
+ + +
+ +
+ + +
+ +
+ +
+
+
+ +
+ +
+ +
±
+
+
+ +
+ +
Larvae
+
++
+
+
++++
-
-
+
+
+
+ + +
+
+
++
++
++
++
+
+
+
+
-
++++
+++
+++
+
+
+
+
+
+
+
( c o n t d . . . . ]

37
33 30.0
34 29.0
35 27.0
36 26.0
37 25.0
38 22.0
39 20.0
40 19.0
41 18.0
42 17.5
43 17.0
44 16.5
45 16.0
46 15.0
47 14.0
48 13.0
49 12.0
50 U.O
51 10.0
+
+
+ +
++
++
+
+
++
+
+++
+
+++
+++
+
++
+++
+
+
+
—
±
+
+
±
+
-
++
+
+
-
-
+++
±
++
+
+
+
_
The adul t and infect ive l a rvae of A.ceylanicum were solubi l ized
in 1 X SDS-sample buffer (containing 5 ml g lyce ro l , 2.5 ml
^ -mercap toe thano l , 2.5% SDS, 0.001% bromophenol b lue , 0.0625M
Tr i s HCl, pH 6 . 8 , in a to ta l volume of 50 ml) by keeping
in a boil ing water ba th for 5 min and sepa ra ted on 10-20%
SDS-polyacrylamide gradient g e l . The prote in bands were num
bered according to t h e i r molecular w e i g h t s .
++++ = ve ry in tense , +++ = in tense , ++ - modera te , + = weak,
± = faint

38
AcLwere determined by using different concentration of ant i
gens (0.015 to 1 ug/well) at fixed dilutions of immune rabbit
serum (1:500, 1:1000, 1:2000) and it was found to be 0.25
ug/well for Ac A and 0.50 ug/well for AcL. No significant
increase in ELISA specific activity was observed by increasing
the antigen concentration upto 1 ug/well, however, on decreasing
the antigen concentration, decrease in ELISA specific activity
<ff'As noticed (Big. 3,4) .
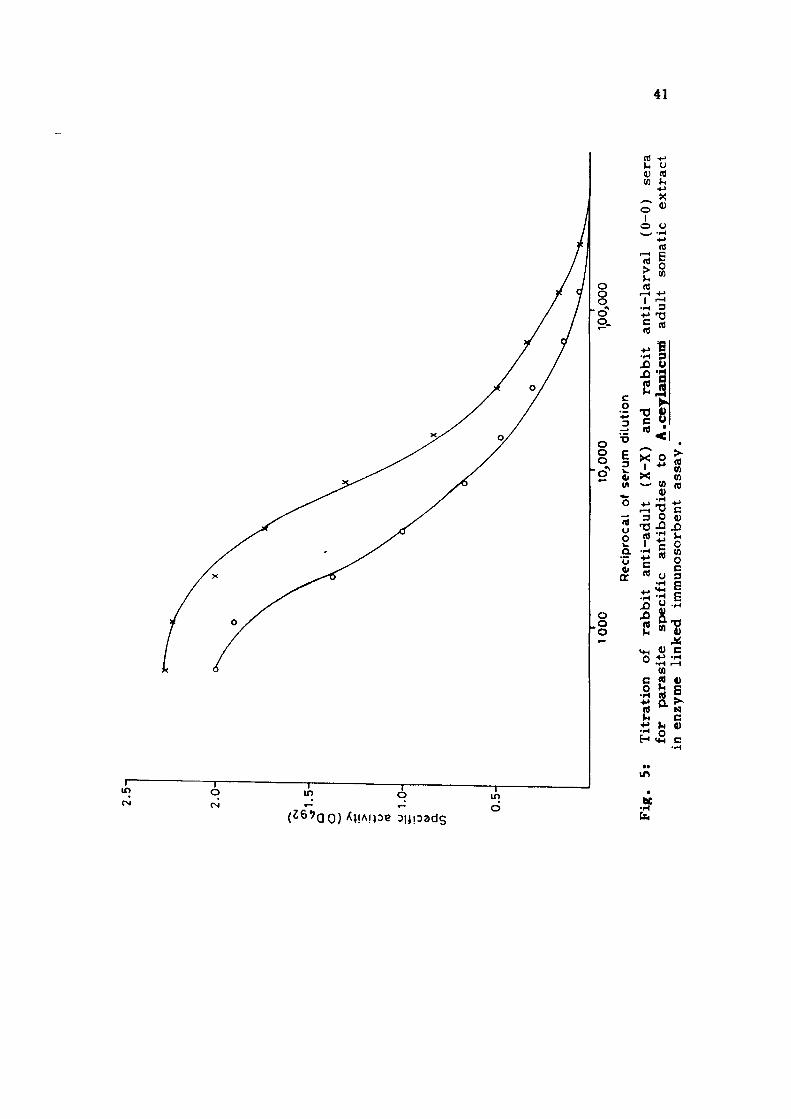
After optimising the assay system, the antibody t i ters
of parasite specific antibodies in immune rabbit sera were
determined and the titration curves are shown in Fig. 5 and
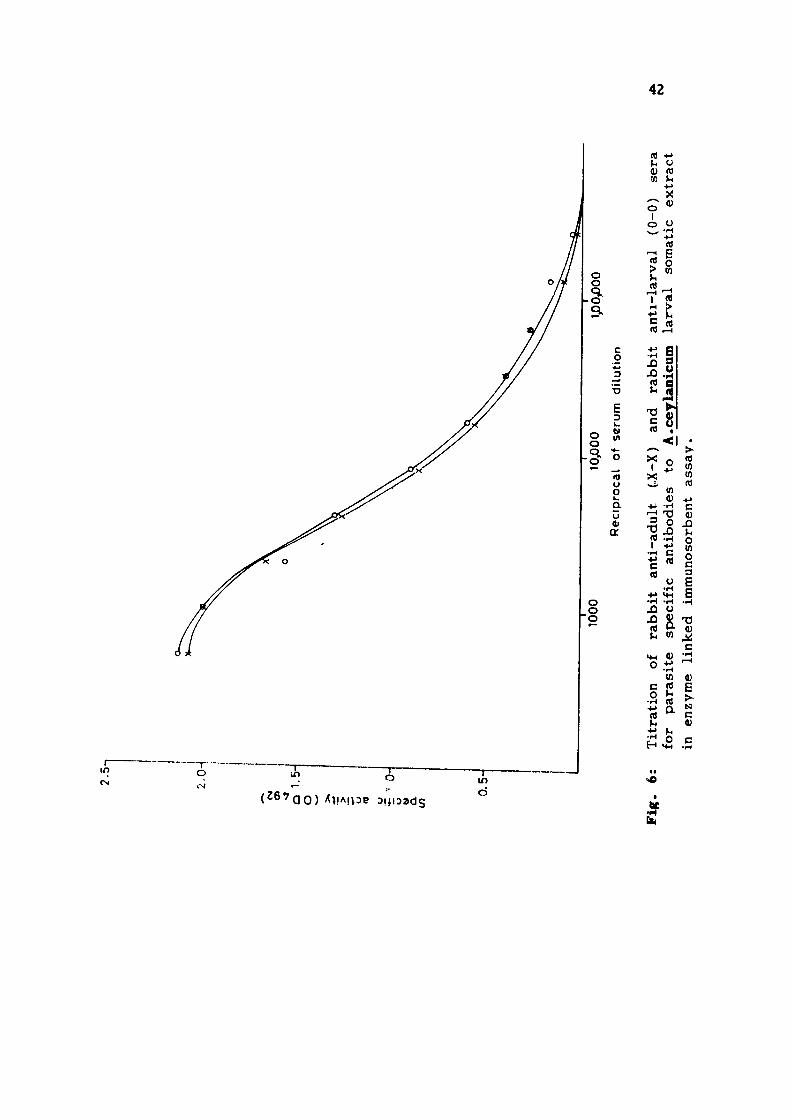
6. The titration curves obtained with AcA using homologous
(rabbit anti-AcA) and heterologous (rabbit anti-AcL) immune
rabbit sera showed linearity upto a serum dilution of 1/32,000
(Fig. 5) . The reciprocal antibody t i t e r s of 256,000 and 128,000
were obtained with homolgous and heterologous sera respect
ively . The titration curves obtained with AcL using rabbit
anti-AcL (homologous) and rabbi t anti-AcA (heterologous) sera
are shown in Fig. 6. Both the titration curves were linear
upto the serum dilution of 1/32,000 and a reciprocal antibody
t i t e r of 256,000 was obtained with both homologous and hetero
logous immune rabbi t sera .
( i i ) Antigenic pattern
Immunoelectrophoretic analysis: Antigenic characterization
of the somatic extracts was initially done bv immunoeie-ctro

39
^
0.20 O.AO 0.60 0.80
Ant igen concentrat ion ( / j q / w e l l )
• • 1
1 0
Fig. 3: Effect of different concentrations of adult somatic extract of A.ceylanicum on the specific activity in enzyme linked immunosorbent assay using 1:500 (X-X), 1:1000 (0-0) and 1:2000 (£»-A ) dilutions of rabbi t , anti-A.ceylanicum adult serum.
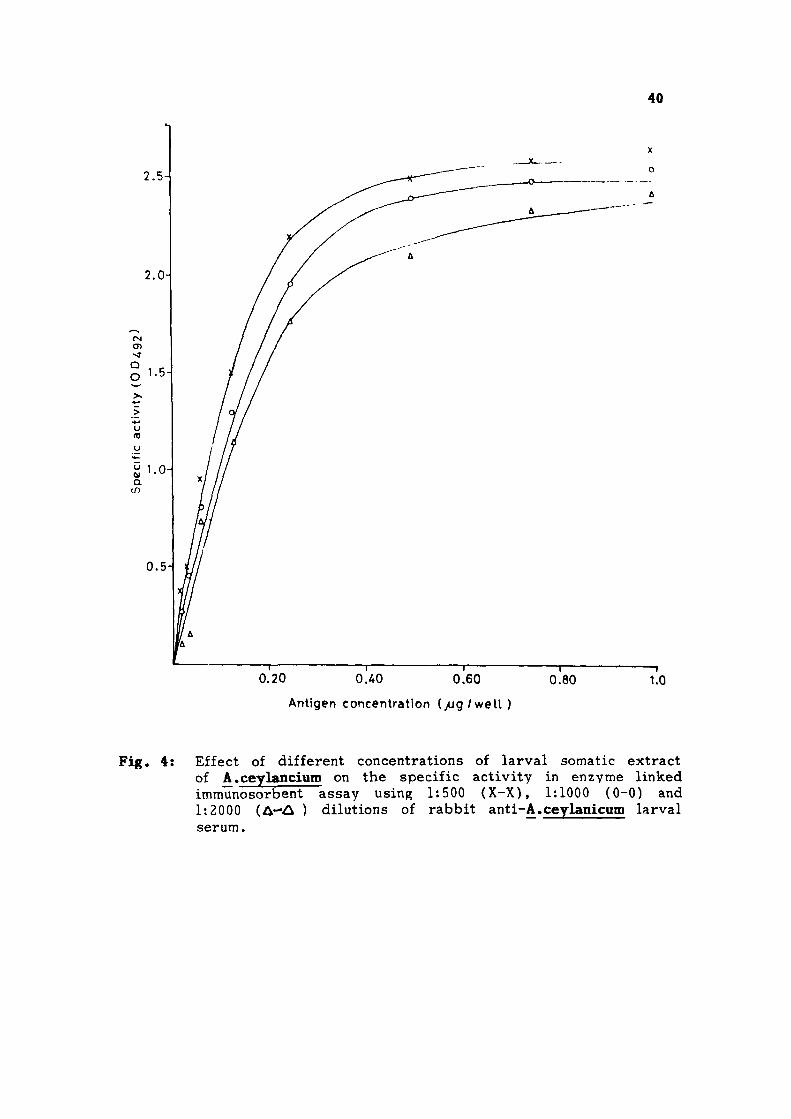
40
0.20 O.AO 0.60
Antigen concentration ( / jg /we l l )
0.80 1
1.0
Fig. 4: Effect of different concentrations of larval somatic extract of A.ceylancium on the specific activity in enzyme linked immunosorbent assay using 1:500 (X-X), 1:1000 (0-0) and 1:2000 (A—A ) dilutions of rabbit anti-A.ceylanicum larval serum.
41
o o o
•\ o o . T —
o o o
o o o
c o 3
•6 E 3
o -7^
{J
0 9.
cr
rt •1-'
0 «
0 0 >^ - H
nl ^ H •<-'
' ':i •H 3 •^ "TJ C (TJ
rt "
* 1 •rt a J3 0 n t 18
•
?5 rt w 0) rt
4) • M ; « - ^
3 0 0) 73 -5 JD
1 ^ 0
•M « 0
rt 0 D
*- 5 1 - « "
sh X
^ V C 0 +J -H '-' -H 1—1
(a
C «l 0)
(4 B •^ 0 c
(Z6!^QQj/^jiAjpe oijiDads
i n
«c
42
ir>
CM
J^^
^ ^ v o
^. 6 • " •
( Z 6 ' 7 Q O ) ^iiAnop Diioadg
I • in
c3
O If o o
o o o o ^
o o o
c o 3
E 3 61 in
« • -
o _ <0 u o a u
cr
(B •(-> u u 0) nl en u
•t->
X o <" 1
••->
n)
rt 0 > to
- i «) 3
73 Sr 5 o
^ >-
X 2 (0
^ - ^
•H -H -H ^ y
c
to 0) c «i E 0 1 s,
nl ^ C u a
•
b4

43
phoresis ( lEP). The antigenic extracts from adult and larval
stages were separated electrophoretically and then allowed
to react with hyperimmune rabbit sera . Ten tn twelve preci
pitin lines were observed in AcA with rabbit anti-AcA serum,
while antigenic preparation from infective larvae showed the
presence of only 6-7 precipitin bands with rabbit anti-larval
serum (Fig. 7 ) .
Crossed immunoelcctrophoretic analysis (CIE): Antigenic com
ponents of adult and larval stages of A.ceylanicum were further
separated more clearly using two dimensional Immunoelectro
phoresis , i . e . crossed Immunoelectrophoresis (CIE). Different
concentrations of serum (gamma-globulin fraction) were tr ied
in order to get bet ter resolution of antigenic components.
Antigenic peaks became much stronger and sharper with increas
ing the antibody concentration, however, with higher per
centage of serum, slight decrease in the height of antigenic
peaks was observed which may be due to shift in the zone
of equivalence.
When somatic antigenic preparation from adult worms
(AcA) was allowed to react with i t s homolgous serum (20%-
gamma-globulin fraction of rabbit anti-adult serum), 34 antigenic
peaks could be observed, 25 moving towards the anode and
9 towards the cathode. Fig. 8a shows the protein stained
CIE pattern of AcA while i ts diagrammatic representation is
shown in Fig. 8b. Each peak represents a single antigen
and the faint antigens are shown by the broken l ines. Almost

4-4-
Fig. 7: Immunoelectrophoretic patterns of somatic extracts from
adult and infective larval stages of A.ceylanicum adult
somatic extract (A) and larval somatic extract (B).
1 = Rabbit antiadult serum; 2 = Rabbit anti-larval
serum.

4-5'
Fig. 8: Crossed immunoelectrophoretic pattern of A.ceylanicum
adult somatic extract antigen (a) Protein stained pattern,
(b) Diagrammatic representation. Solid lines represent
strong antigenic peaks . Arrows indicate the point
of application of antigen. Anode ( + ) is to the right
in the first dimension and to the top in second dimen
sion gel. Antibodies: 20% gamma-globulin fraction
of antiserum raised against somatic extracts from adult
A. ceylanicum.

46
similar antigenic pattern was observed using 15% and 20% gamma
globulin fraction, however, with a lower antibody concentration
of 10%, 3-4 antigenic peaks (numbering 5,7,11 and 15) d i s
appeared completely and the intensity of some of the antigenic
peaks declined. Reduction in the number of cathodic preci
pitin peaks was also observed with 10% serum concentration.
When larval antigenic preparation (AcL) was run with
rabbit anti-AcL serum (10% gamma globulin fraction), 19 anti-f
genie peaks could be observed. The antigenic pattern of
the somatic extracts from AcL, as revealed by CIE, i s depicted
in Fig. 9. Out of 19 antigenic peaks 17 were anodic and
2 were cathodic. One of the two cathodic antigens (AcL-
18) was slow migrating and precipitated in the first dimension
gel only. Using a lower concentration of gamma-globulin fraction
(5%), some of the faint peaks numbering 1, 3 and 11 d i s
appeared completely and a decrease in the intensity of some
of the antigenic peaks (numbering 4 and 8) was also observed.
Table 2 summarises the results obtained by lEP and
CIE analysis of AcA and AcL using the homologous rabbit
sera.
IDENTIFICATION OF COMMON OR CROSS-REACTIVE AND STAGE/
PARASITE SPECIFIC ANTIGENS
Antigenic similarit ies do exist among different stages
of a parasite as well as between two different paras i tes ,
hence studies were also conducted to identify the common

Fig. 9: Crossed immunoelectrophoretic pattern of A.ceylanicum
larval scrmtic extract (a) Protein stained pattern, (b) Dia
grammatic representation. Solid lines represent strong
antigenic peaks . Arrows indicate the point of appl i
cation of antigen. Anode ( + ) is to the right in the
first dimension and to the top in second dimension
gel. Antibodies: 10% gamma-globulin fraction of anti
serum raised against somatic extracts from larval
A.ceylanicum.

48
• n
H u
(0 0) bf rt
+ j en
1—1
«! > yi
1—1
T( C rt
•»-> r-H 3
T) «)
M H
O
(0 •H tn >-• — 1
rt c rt o
• H
c t) bf
• H 4 J
c <
• • N
4) F H
Xi J^ H
E 3 U i) (n
•H
c <
c i) b£
•H +-> C <
m 0) 3 O
• H C
Si u d)
H
c rt •H 4)
O 73 O
•rt U5
C <
< u <
I 1 • H ••J c rt
-4-> • H J2 J3 A
Oii
J U < 1 1
•H •fJ
c (t
+-> • H J3 X) (6
K
< <
u <
1 o u • » ->
o a;
f-H
0) o c 3 E E
)—i
w • H w (U H
o XI
a
in r-CO r-H
< < I
•H c
J3
u <
I
c (0
•H XI Si nt
OS
<
I
u <
0 05 C -H 3 m E Ji 1 o
to i o "
U 0)
• » - »
u nl U
+ j
X (U
u • H + J
rt E o (0
+ J r-H
3 TJ
O nt U
• • - »
X (U
u • H
•*-> nl E 0 n
r—(
n! > rt rt r^
B 9 U
1 •< ft)
u
B 9 U •H B
43 •< ft)
u <l <l
< o <

49
or cross-reactive antigens between the adult and larval stages
of A.ceylanicum and also between the two hookworm parasites
(A.ceylanicum and N.bras i l iensis) .
(i) Common/cross-reactive and stage specific antigens
The common/cross-reactive antigens between the adult
(AcA) and larval stages (AcL) of A.ceylanicum were analysed
by doing CIE of AcA and AcL using immune rabbit sera raised
against these stages. When the somatic extracts from adult
worms were allowed to react with rabbit anti-AcL serum in
CIE, 22 antigenic peaks were observed (Fig. 10b) while CIE
analysis of A.ceylanicum larval antigen (AcL) with rabbit
anti-AcA serum showed the presence of 14 antigens (F ig . l ib ) .
On comparing these results with CIE patterns of AcA (Fig.8)
and AcL (Fig. 9) with the homologous immune rabbit sera,
12 and 5 antigens appeared to be specific to adult and larval
stages respectively (Table 3) .
The presence of common or cross-reactive and stage-
specific antigens between adult and larval stages was further
confirmed by doing cross-line Immunoelectrophoresis (CLIE).
Fig. 10c shows the CLIE pattern of adult antigen in which
the larval antigen was introduced in the intermediate gel
s t r i p . On comparing the CLIE pattern of AcA (Fig.10c) with
i ts CIE pattern (Fig. 10a), most of the adult antigens appeared
to be common to the larval stage as indicated by the presence
of lines at the bases of these antigens. However, the antigens
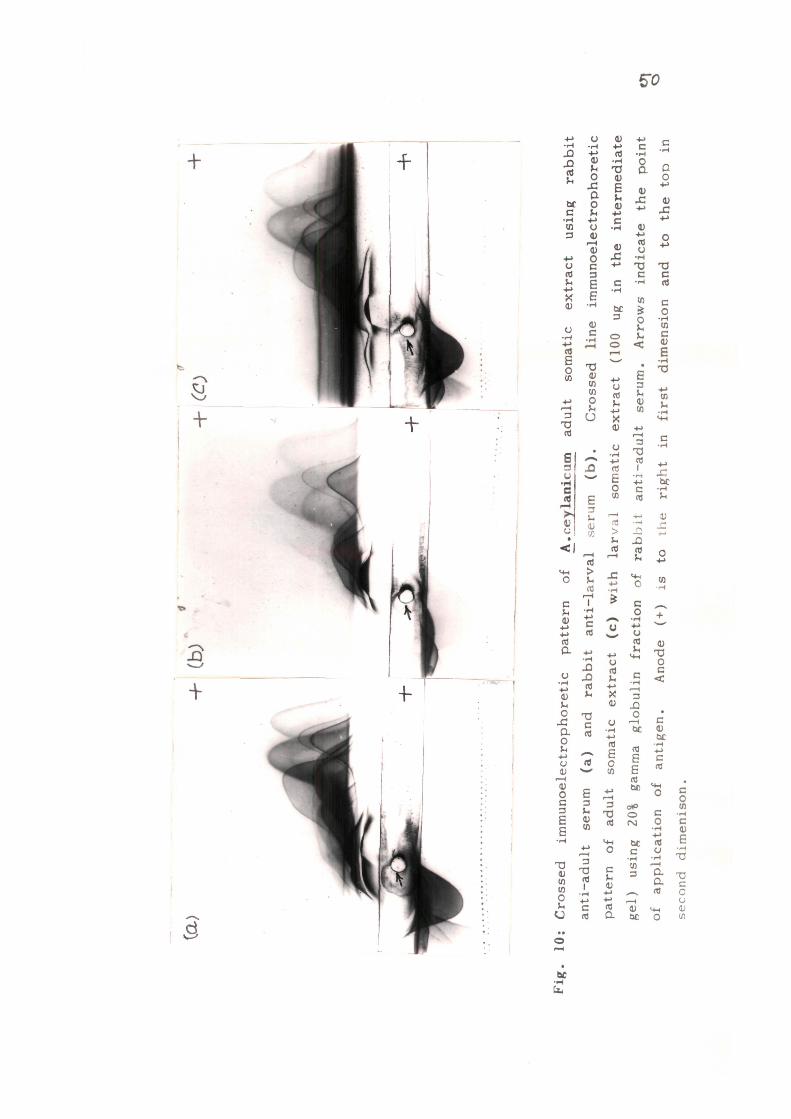
STO
n) U
c •H 01 D
O A
-t-> X
E o CO
3
u o
a o o 4}
1—I 4) O C D E E
a; c
t) in (0 o
0) ••->
ei -
c u
•» ->
+-•
a
0)
o x: a o o i)
<u o c 3 E E
E •3
>
c
Xi
in )H
t3 c
(U E u 9)
J3
3
O O
o n)
+->
o a 0)
o o +->
J3
nj o T3 c
o u
<
E 3 U en
3
« (
c w
C rt
c o
•iH U) c <u E
-H
en
be
ifl flj
Si +-> H
c o
o
X 0)
0) E o en
E 3 U 0)
en
3 J2 O
•—I «
E E rtj 61)
T3 O
^H be O c
o c
<
c 4)
+-> c
c o
c 0 in
•H c t) E
(A in 0 u
3
(0 I H
•» ->
c (TJ
c (U
•(-J
m
a
3
bC
a a (0
-a c o u u (A
s
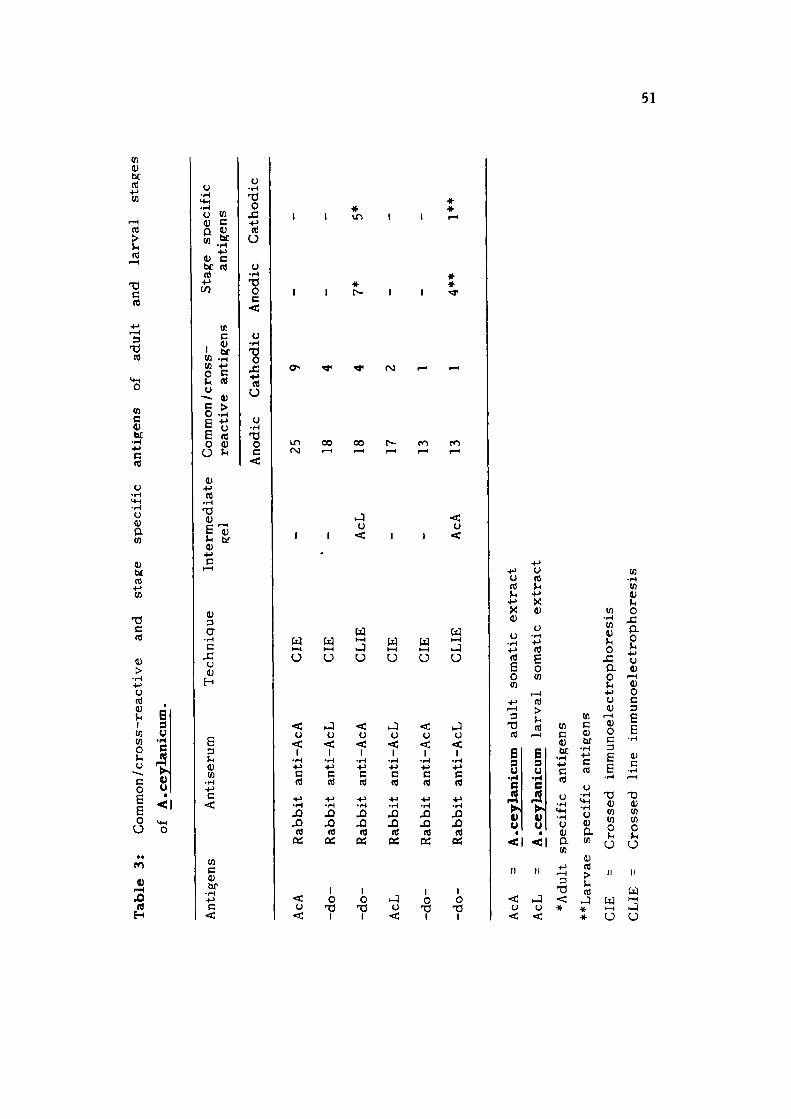
51
> (0
T3 c n!
nt
w c 4) bt •H c
o
0 -
be
c
u n) (U ^ I tn en O ) H
o c o E 6 o o
A)
E2
B 9 U
•H
o
c u
< l
o
o tn
- tt) a (A
be
<U C bc n) n)
I W CO O *H
u B O E E o u
c 0) bf •H •< ->
B (Tt
0) >
•H • U U n) (1)
m •H
(U ^ E 0) 1^ br
3 CT
• H
c
o
H
E
3
CO
c <
to B (U b f
•H
c <
u •r-l
O X! ••->
rt U
73 O c <
o • 4 ->
n) U
O B <
• in
* t ^
*
V
i n 00 00
u <
< u <
1 o
T3 1
1 O
T3 1
J O
<
< I <
w 1—t
o M •—1
u
u t—4
b j
u M 1—1
u u t—(
o
w •—1
hJ u
< u <
1 1 • H •t->
B rt
+ j • H
J3 J3 (0 «
J u <
1 1 • H •<->
B rt
••-> • H
^ J3 n)
CH
< o
< 1 1
• H + J
B rt
••-' •H
x> J3 (« «
J u <
1 1 •H 4 J
B rt
4-1 • H
X) ^ nt «
< o < 1 1
•rt •» ->
B rt
•4-> • H
XI XI n) «
J o < 1 1
•H •«-> B (d
+ j • H
-D XI n)
OS
I I 0 o 1 I
U
m
a)
nt E o CO
3 T3 (0
(J n)
+-> X
E o en
>
s 9 u •H
a ^ >> 0)
u
B 9 U •a 43 • 0) u
<l <l
< u <
tn E <l) bc
• H + J B
o
a to
3
< •Il
ea
o x; a o
•t->
o
to (U u o x: 0-o u
• 4 - "
o 0)
I—( 0) o B 3
to E (1) br H
B
1—1
0 B 3
E E
E E
•fH
1 T3 ID 0) 0)
U C
a to 0)
CO CO
o u
CO CO
O O U
W w
o u

52
AcA 2, 3, 18, 19, 21, 24, 25 and 30-34 which did not have
lines at the bases appeared to be specific to adult stage.
CLIE of AcL was done by adding AcA in the intermediate
gel s t r i p . On comparing the CLIE (Fig. l i e ) with CIE (Fig.
11a) patterns of AcL, most of the larval antigens were found
common to the adult stage, however, 5 antigens (AcL-2,13,
16-18) specific to AcL were also observed. The results of
stage specific as well as common/cross-reactive antigens of
the two stages are summarised in Table 3.
( i i ) Common/cross-reactive and parasite specific antigens
A.ceylanicum antigens having cross-reactivity with
the rat hookworm parasite N.brasil iensis were also analysed
using heterologous immune rabbit sera (raised against adult
and infective larval somatic extracts of N.bras i l iensis) . CIE
of AcA with rabbi t anti-NbA serum revealed the presence
of 12 cross-reactive antigens between the adult stages of
the two parasites (Fig. 12a). When the AcA antigen was
allowed to react with rabbit anti-NbL serum, 8 cross-reactive
antigens between the two stages of both the parasites were
found (Fig. 12b). Common/cross-reactive as well as parasite
specific antigens of AcA were further analysed using CLIE.
Fig. 13a shows the CLIE pattern of AcA with NbA in the
intermediate gel s t r i p . On comparing the CIE pattern (Fig.8)
of AcA with i ts CLIE pattern (Fig. 13a), 22 antigens (AcA
1-3, 6, 9, 10, 12-14, 18, 19, 21, 25-34) were found to be
AcA specific. Fig. 13b shows the CLIE pattern of AcA using

o
: ^
+
o
\
\. + + . >
1—1
ti > u n)
1—1 •H • « ->
c
• H J3 Xi
B •H CO D
O A $ 1
i)
a •fH • ! - >
nj E o (fl
>-H
n) > ti
a 3 U
•H 0
>> 4) u • <
O
c u 4}
•t->
(0 a u
u o
a o u o 0)
r—1 0) o c 3 6 E
<4H
O
c u
a u
- H
(U u o
Si a o u
•*->
u 0)
1—I i) o c 3 E E
• f H
c •iH 1—I
•T3 4) (A (0 o
u
, ^^ Xi
E 3 ^H
V «
<—( 3
73 rt 1
•iH
c <fl
XI
c nJ
E 3 ^4
0)
ClC c • H (A 3
1-H
bt
0) • • J n)
• H
E
c • H
0)
-t->
c
tac 3
o o 1—1
^^ • ! - >
u n)
(J •rH • 4 - "
E 0 'n
•>->
1—1
3
• H
5
u >•«'
o R) tH
• ( - >
X
u • J
m E o (0
1—1
> (fl
1 nt u • H
1—1
a 0. n)
VtH
o
c • H
a • » ->
0) + J
rt o • H 73 c • H
tn ^ o
<
E 3
0)
1—1
n3 > ^H
<« 1
•r-t
c «J
-t-" M
Xi XI
<4H
0
c 0 H
+ J
o
c .—1
3 Xi 0
E E bc
c*> O
1 c 4) E
•H
XI
73 c 0 O 0) V)
c
a O
• 1 - '
u Xi - t - l
o •»->
TJ c
c o iH CO
c 0)
E • i H
TD
i H •-t-t
c • H
••->
be
(U XI
o
tn H
-f
0) T I O C <
• c i) be H
c n)
V M
o c 0 rt
c o
5

m + \h) +
n— H- -f-
Fig. 12: Crossed immunoelectrophoretic pattern of A.ceylanicum
adult somatic extract using 15% gamma globulin fraction
from rabbit anti-N.brasil iensis adult serum (a) and
rabbi t anti-N.brasil iensis larval serum ( b ) . Arrows
indicate the point of application of antigen. Anode
( + ) is to the right in first dimension and to the top
in second dimension.

es-
Fig. 13: Crossed line immunoelectrophoretic pattern of A.ceylanicum
adult somatic extract with N.brasil iensis adult somatic
antigen (a) and N.brasiliensis larval somatic antigen
(b) (100 ug of each in the intermediate gel) using
20% gamma globulin fraction of rabbit anit-AcA serum.
Arrows indicate the point of application of antigen.
Anode ( + ) is to the right in first dimension and to
the top in second dimension.

56
NbL in the in te rmedia te gel s t r i p . Only 8 antigens were
found common between AcA and NbL. Antigens numbering 1-
3 , 10-25, 27, 29-34 were found to be AcA spec i f i c . Table
4 d e p i c t s t h e summary of t he se r e s u l t s .
Common/cross- react ive antigens between the l a r v a l
s tages of A.ceylanicum and N . b r a s i l i e n s i s were a lso ana lysed .
CIE ana lys i s of AcL with r a b b i t ant i-NbL and r a b b i t a n t l -
NbA sera r evea l ed 13 antigens ( F i g . 1 4 a , b ) . On comparing
t he se r e s u l t s wi th CIE pa t t e rn of AcL with homologous ( r a b b i t
ant i -AcL) serum, 6 antigens appea red to be AcL spec i f i c . The
speci f ic and common antigens were fu r the r analysed by doing
CLIE. On comparing CIE ( F i g . 9) and CLIE (F ig . 15a) of
AcL, 13 antigens appea red to be cross- reac t ive /common between
the two p a r a s i t e s whi le antigens numbering 12-14, 16, 17 and
19 were found to be AcL s p e c i f i c . CLIE pa t t e rn of AcL with
NbA i s shown in F i g . 15b. On comparing the CLIE (F ig . 15b) ,
of AcL with i t s CIE (F ig . 9) most of the antigens were found
common between AcL and NbA. Antigens numbering 2, 13,14,
16, 17 and 19 appea red to be AcL s p e c i f i c . The r e s u l t s of
common or c r o s s - r e a c t i v e as well a s speci f ic antigens of AcL
as compared to NbL and NbA are summarised in Table 5 .
IMMUNIZATION OF HAMSTERS WITH SOMATIC EXTRACTS FROM
ADULT AND LARVAL STAGES AND EVALUATION OF IMMUNISED
HAMSTERS FOR PROTECTIVE IMMUNITY
Hamsters were immunised with the somatic e x t r a c t s
from adul t and l a r v a l s tages of A.ceylanicum at 15 d a y s i n t e rva l .
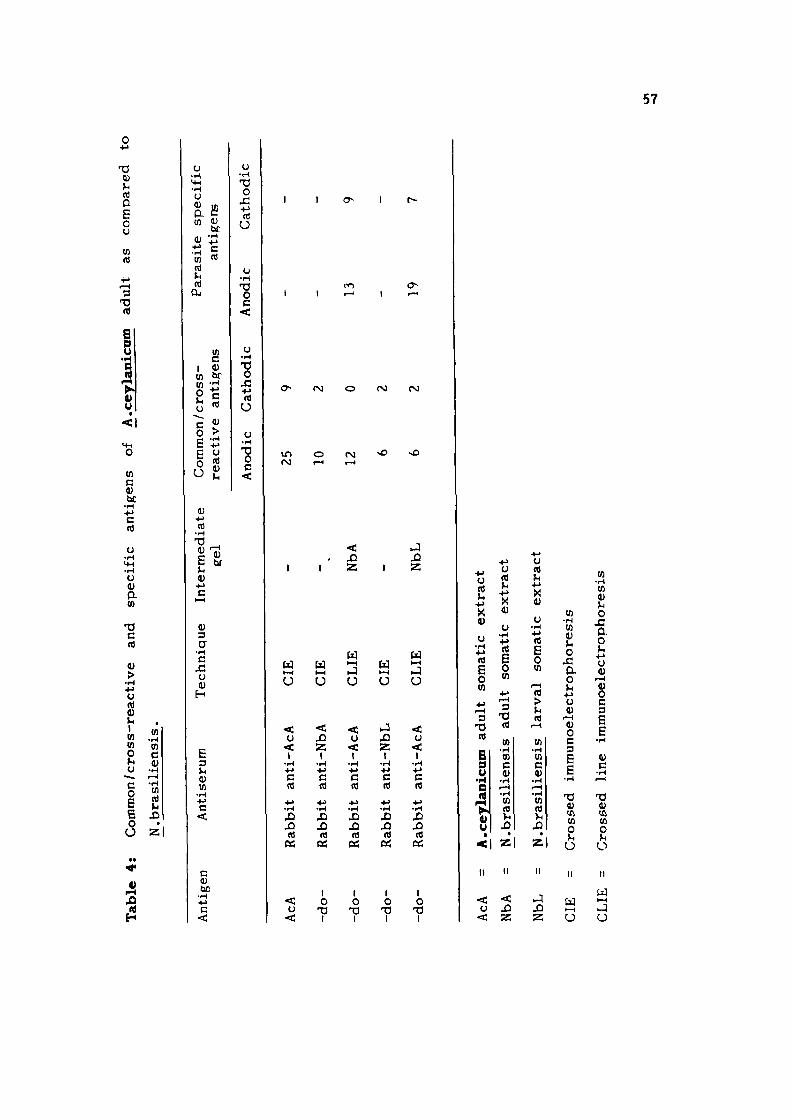
57
73 to U
a E o u
(fl n)
D T! («
B s u
• H
I 0)
u
(0
c 0) be
•H c nt
u (1)
a (0
' 0 c n)
0)
> • H o n) 0) v< I (0 (n o »^ u
c o B E o o
E2
(0 •r-l (0 c 0)
in n)
U
•t! = « "J
! a,
c I 4) to tiT
o (fl
E X2
o ™
•H
(U
(U 3 D"
• H c o t)
H
E D
0) tfi
c <
c i) bC
• H •t-> c
<;
o •r-( T l O
•4->
(fl
o
(J •H TJ O C <
T3 0
J3
(fl O
c <
(?
I r-l
fM
IT) (M
(M
Z z
w • — t
u u l-H
u
w • — 1
>- o
w h-l
u
u l-H I - ]
u
< u <
1 1 •H • f j
c <fl
4 J • H XI XI (fl ec
< X3 z
1 1 •H • f j
c (fl
••-» • H XI X) (fl «
< u <
1 1 •H + J
c (fl
•*-• • H X) XI (fl oi
J X) z
1 1
+ J
c (fl
•»-> •1 -4
XI X3 (fl «
< o <
1 I • H -tJ
c (fl
•IJ • H XI X5 (fl
Otf
u «fl
•t->
X
(fl
E 0 n
u (fl
X
(fl E o 0)
T3 rt
o (fl
•<-»
X 0)
(fl E o
(fl > Vi (fl
< o <
1 o
XI
1 o
13 1
1
o T) 1
1 O
n 1
(0
B s u •H
1 ^ 4) U
(» •H tn c 0)
•rt rH •H (0 ? )H XI
to •H tn c 0)
• H l-H •H tfi (fl V* XI
< l z l
< < o XI
< z
tn •rt (fl o; 0
x: 0-o V4 u V
4) 0 c 3 E E
tn 0
(0 4) ; H
o X! a 0 u
•(->
u 4)
l-H 4) 0 c D E E
• r t
4) C
X) 4J
tn o
• J M • - ( XI S J z o o
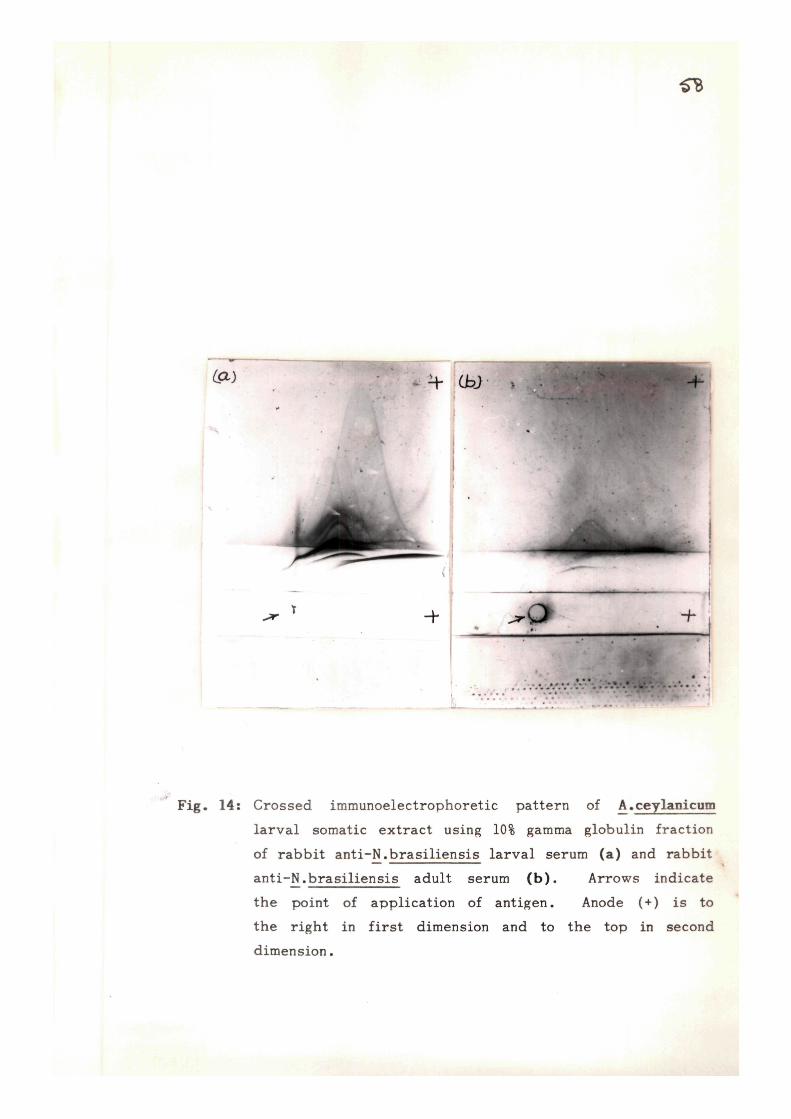
^
ta)
i s
- H- (bJ >
4-
iiiMHMi
a 4-
Fig. 14: Crossed immunoelectrophoretic pattern of A. cey lanicum
larval somatic extract using 10% gamma globulin fraction
of rabbi t anti-N.brasil iensis larval serum (a) and rabbit
( b ) . Arrows indicate
Anode ( + ) is to
anti-N.brasil iensis adult serum
the point of application of antigen.
the right in first dimension and to the top in second
dimension.
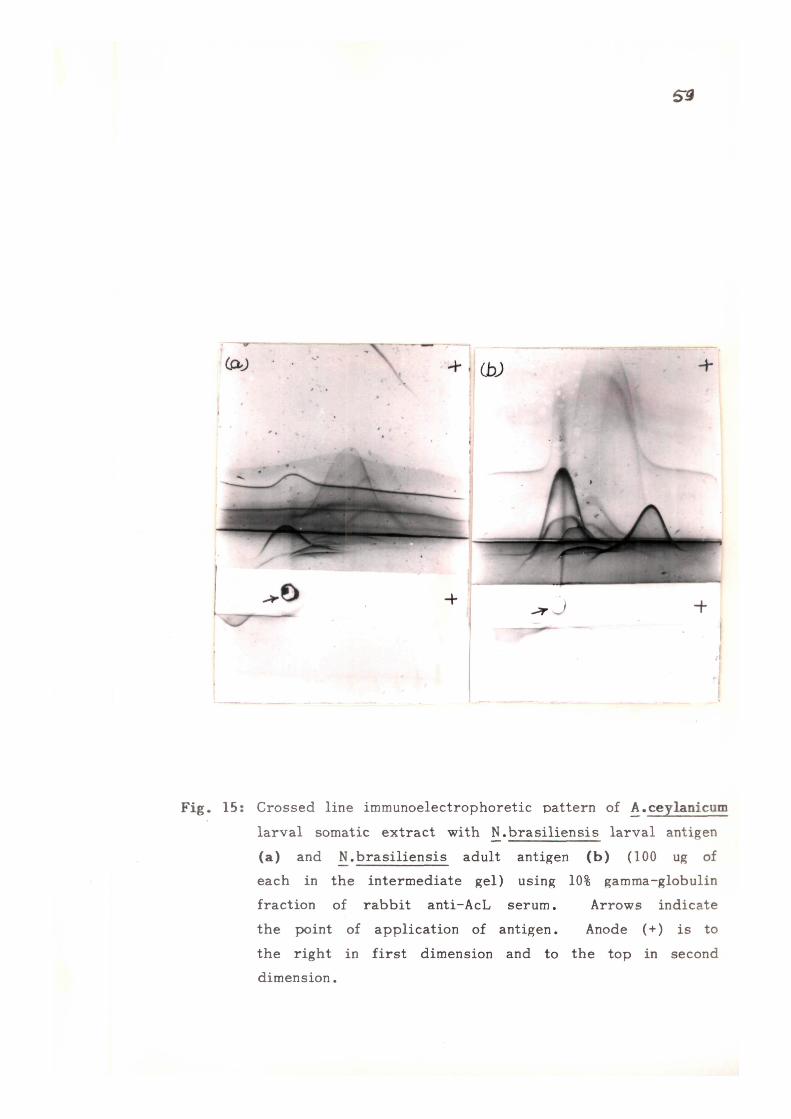
53
Fig. 15: Crossed line immunoelectrophoretic pattern of A.ceylanicum
larval somatic extract with N.brasil iensis larval antigen
(a) and N.brasil iensis adult antigen (b) (100 ug of
each in the intermediate gel) using 10% gamma-globulin
fraction of rabbi t anti-AcL serum. Arrows indicate
the point of application of antigen. Anode ( + ) is to
the right in f irst dimension and to the top in second
dimension.

60
(fi c 0)
•H .—I •H 01
JO
Z l
u 0) a tfl
OJ •M • H m
a,
-^ (U >
• H
•»-> u ni (L ^ 1 (fi tn 0
U
V) c V
• H -t->
c «)
w c t) bf
• H •t->
c rt c 0 E E o o
Tl 0 x: U
u •H T3 O C <
u • H T3 0
XI -t-"
n! U
o • H
1 c <
I r-l
•H Xi m r-l E l i
0)
3 cr
•H C
XI u d)
H
E 3 0) (0
c <
c 0) be
• H
c
< o
<
fVJ (N3
T3 < XI X I
I Z I 2
w 1—1
o u t—1
u
w 1—1
l-J u
u 1—1
u
w t-H
h j u
J u < 1 • H
+-" c rt
•<-» •rt -Q X3 rt a;
J XI 2 1 • H + J
c nJ
4-" • H
X) XJ rt
OJ
J o < 1 1
+ j
c rt
•»-> •rt ^ X3 m o;
< XI 2 1
• H • M c rt
+ j • H
X3 X5 n) «
J u < 1
• H 4->
c rt ^ • H
X3 XI n)
OS
1 o
T l 1
1 O
T3 1
1 0
T l 1
1 O
TJ
u
0)
E o m
>
o
<
o Rl u
•» ->
X 0)
rt E o (0
2
u n)
4-> X 0)
u •H
n) E o m
3
t-~t
B 9 U
•H
1 >> 0) u
m • H (0 c V
• H
• H
{* >H
XI
to • H (0 c 01
• H r-( •H
(fl ^ X!
21 21
o X3 0.
o ^4
-t-" o
I—I dj o c 3 E E
t 3 (U (A 0)
o ( J
< XI 2
tn •H to <D U O x: a o u
+ J o lU
i-H <u o c 3 E E 0) c
•V
tn in o u u
II
w W '-:!
O u

61
The sera obtained one week after every injection were tested
in enzyme linked immunosorbent assay (ELISA) and immuno
diffusion (ID). The antibody t i te rs of immune hamster sera,
expressed as geometrical mean reciprocal t i ter (GMRT) are
given in Table 6. The parasite specific antibody could not
be detected in sera obtained after 1st injection, however,
significant levels of antibodies were detected in sera from
2nd injection onwards. The GMRT values obtained for hamster
anti-AcA and hamster anti-AcL serum after the 2nd injection
were 65.97 and 57.48 respect ively. The GMRT values of
these sera were significantly increased to 527.80 for hamster
anti-AcA and 303.14 for hamster anti-AcL following the 4th
injection. No significant change m the antibody t i ters of
these sera was observed after 5th injection. The immuno
diffusion analysis of these immune hamster sera showed that
the precipitin bands first appeared after the 3rd injection
and 3 precipitin bands in AcA and 2 precipitin bands in AcL
were observed. The number of precipitin bands was increased
to 4 and 3 with AcA and AcL respectively after the fourth
injection and it remained constant after 5th injection. The
results of ID are summarised in Table 7. The hyperimmune
hamster anti-AcA and hamster anti-AcL sera obtained after
the 5th injection were tested in lEP, which showed the p re
sence of 7 and 4 precipitin bands in AcA and AcL respectively
(Fig. 16).
The hyperimmune hamsters were evaluated for protective
immunity by challenging them with an oral dose of 60 L-

01 (0
c 0)
U
o (0 o c D E E
c •H
E > N c 0)
JD
rt u (U (0
; (U
w E «J
V c a E E
(0
0)
t3 O
• H
c <
E2
c o
^ • ^ LTl 4)
• * - >
>-O
•H
c <
C O
c
C
o
E
m •H c <
c (V M
• H • ( - >
c <
00 (M
o
00
00
rj<
r-N i n
en o en
ID
O IT)
i n o
00
r~ o
• i n >o
00 •«i<
• r i n
Q Z
Q 2
u < I
c nl
U (U
• » - >
E <«
• J u < I
•H +J
c
U <u (A E n)
X
< <
+ J
o nJ
X (U
o •H + J
rt E 0 «
• < - '
r—1 3
B s u -g ^
u < II
< u <
• f j <J nt 4-> X 0)
u • f t 4-> n) E o (0
»—1 nl
> nl
•—1
5 a u •H
>>
<
II
J o <
(0
(U ••-> (D
E nl
X
i> >
•H M-l
E 0 u
t) C
•H
rt
0
H
o
a> "•J • H ••->
1—t
nl u O U & u
c nJ
E .—( nl U
• H
0) E o C)G
(fl nl
T3 0)
0)
a X ' CJ t-J
o CJ <
nl 73 c
en n)
^ < •H O
>> U T ) O
o '^ :2x -t-> u C nl
< i)
0)
o 0) +-> (U
T3
T3 O
•H +-> c nl
o 2
II
Q 2
62

63
C
m 3
S s
H u
rt » •(->
X (1)
o • H - t J
<B
E 0 m
r—(
> u ta
r—<
T) c rt
• * j
3 73 nl
>+-i
0
to • H
(0
>-r - H
n) c rt
c o •H Ul D
• H
T) 0 C D
E E
• rt >-( 0) m
u i)
••-> OT
E
(U c 3
E E
E2
Cfi Ti C rt
XI
j :3 •(->
>
c o •H o <u 'f~
a
o tt)
a
o
>
•H
c o
^ .2.
c o
c
c o
tn -^
tn •H +-> c <
c i) be
<
E 3 U 0) (0
< o <
1 • H 4->
c (6
y, 4)
•»->
CD
E n a:
< u <
E 3
^ 0) cn
J u < 1 1
+-> c m u <D
•(-> CO
E rt ac
.-) o <
c o
o
o
4)
a)
4) C O
T3
(U
& (A
tt) +J to E
X!
tt)
H
•V 4J N
•H c 3 E E
•H
U CO
tt)
CO E rt
Xi
O
rt X
X o rt tt)
c tt) E
•rt tt) a X 4)
tt) tt)
JC
c rt • tt) cn
e g rt c
tt) ^H o tt) g
C tt) tt) J :
•H
O CO tt) j 3 3 <j rt
tt) rt > 4) ^
X -H H &
B a u t
^ •> 0 u
B s u •H
a
V u 1
<l
u a u •M X tt)
(0 E o CO
3
rt
<
<l
I tt)
rt E o CO
> U
o

^4-
A
Fig. 16: Immunoelectrophoretic patterns of somatic extracts from
adult and infective larval stages of A.ceylanicum using
. immune hamster serum. A = adult somatic extract;
B = larval somatic extract . 1 = Hamster anti-adult
serum; 2 = Hamster anti-larval serum.

65
S 9
E o u
o re ;H
X 0)
o
re >
re
T3 c re
3
re
0
c <u o a
> •H +-> o (U o
00
1)
E2
c 0)
c o
o
^ c
0) !H U 3
<U a.
T3 0) c
XI O (D
be m n p <u r '-I J-i - — I o re ^ X !
o o u
E D 2
C 0) txp
•H -M C
<
CO O O I
o o
o o I
CO CO
I
i n +1 o +1
i n +1
(U c o
TJ c re
05 u 0)
E re
x :
< u <
o <
0) X
re
^ -o w
re + j • ^ w
• H
-0.5 S E
13 re
• 1 - '
•—{
3 m 73 Jj re >
m <^ 0 J
o _ ^ c •H . •
O <u
o + T3 C
tie ^ re 3 . ,
o c ? 0 0 ) + ^ .—1 r—1
re j - i X X :g • M O
!* <U 0 X
T) -^ x : <u _ <J to 5 "1
•H O (U
g o x
- H
a , i n .N ^ c
^ •a 0) c
to + j c >H ^ E <a '^ -1-^
-t->
jn ^ 0) E <u >H re <u 0)
•X. ^ ^
+-> o re
X
(J
re E o
r—1
3 T) re
B 9 U
•H B (0
i-H
> V u • <
II
< u <
+ - i
o re u
4->
0)
(J • H
re E o Vi
re >
re 1—1
B 9 U
-3 ^ •> V u • <
II
J o <
en - M
c 0)
E • H
1)
a X 0) 0) 0)
X
<4H
0
c re 0) E
<u X -f-»
0)
^ re
0) 3
.—1
re > 0)
X H

66
at one week after the 5th injection. The worm burden was
seen on autopsy at 21 days post challenge and percent reduction
in worm burden gave the degree protection. The hamsters
immunised with adult somatic extracts gave 63% reduction m
worm burden while 93% reduction in worm burden was observed
in hamsters immunised with larval somatic extracts (Table
8 ) .
DISCUSSION
Ancylostoma ceylanicum (a human hookworm parasite
maintained in golden hamsters) has extensively been used
for chemotherapeutic and immunological studies by several
workers (Misra et_ al_., 1981; Katiyar et al^., 1982; 1984; Kamath
et al^., 1985; Carroll and Grove, 1985; Gupta and Katiyar
1985). However, no information is available about i ts total
antigenic composition as well as about the antigenic molecules
involved in inducing protective immune responses. In the
present study an effort has been made for analysing qualita
tively , the protein and antigenic make up of adult and infective
larval stages of A. ceylanicum employing SDS-polyacrylamide
gel electrophoretic, crossed immunoelectrophoretic and crossed
line immunoelectrophoretic techniques. The levels of parasite
specific antibodies in sera raised against the two life stages
(adult and larvae) of A. ceylanicum have been determined

67
employing enzyrfle linked immunosorbent assay technique. The
protective potential of these somatic extracts has also been
evaluated. Since both male and female worms have been
shown to be equally effective in conferring protective immunity
(Gupta and Katiyar, 1985), the antigenic preparations made
from both female and male worms has been used in the present
study.
The protein composition of adult and larval stages
of A.ceylanicum was studied by SDS-PAGE, which provides
a high resolution of protein bands according to their molecular
weight and the best separation of proteins was obtained using
10-20% gradient of acrylamide gel. Lower percentage of acryl-
amide in the gel showed poor resolution as most of the pro
teins present in these stages were of low molecular weight
proteins. This technique also enables a comparison of protein
patterns of different parasites preparations on the same gel.
SDS-PAGE has earl ier been used for studying the protein
patterns of many nematode parasi tes v iz . Ascaris (Zayatas,
1978), D.viteae (Neilson, 1978; Prusse et_ al^., 1982; Lucius
et a]^., 1983); hookworm (Wange et_ al^., 1980), Trichinella
(Bany et_ al_., 1976; Efremove and Ermolin, 1981); Angiostrongylus
cantonensis (Kum and Ko, 1985) and S.cervi (Malhotra et
a]^., 1986). In the present study, SDS-PAGE helped in the
separation of 47 and 43 protein bands in adult and larval
stages respect ively, of which larval stage exhibited only
13 major protein bands as compared to 28 observed in adult

68
stage. These results revealed a less complex nature of larval
somatic ex t rac ts . The different life stages of some other
nematode parasi tes have also been shown to have approxi
mately the similar number of protein bands (Kaushal et a l . ,
1982; Prusse et a l . , 1982; Malhotra et_ al_., 1986). However,
earl ier studies of Yoshida e^ al^. (1977) have shown the p re
sence of only 13-15 protein bands in A.brasiliensis and
A.ceylanicum adults by disc gel electrophoretic analysis .
In the present study SDS-PAGE analysis revealed
some interesting qualitative and quantitative differences between
the two stages of A.ceylanicum. Protein bands numbering
1, 2, 3 and 12 (170, 160, 153 and 85 kD) were found to be
specific to the larval stage while protein bands numbering
6, 7, 22, 33, 39, 43, 42 and 51 (130, 125, 42, 30, 20, 17,
16.5 and 10 kD) appeared to be adult specific, as they were
only present in the larvae and adults of A.ceylanicum. A
comparison of stage specific proteins indicated that most
of the adult specific proteins (except protein bands 6 and
7) were below 50 kD, while 3 out of 4 larvae specific proteins
were above 150 kD. Carr and Pri tchard (1987) have also
shown low molecular weight (19 kD) protein specific to
N.americanus adult . A 74 kD protein specific to larvae of
N.americanus was also observed by the same workers .
In the present study, the levels of parasite specific
antibodies in immune rabbit sera were determined by enzyme
linked immunosorbent assay (ELISA) which has extensively

69
been used for quantitation of antibodies against a number
of helminth parasi tes including hookworms (Oeilvie et_ a l . ,
1978; Tandon et_ al^., 1981, 1983; Novoselska et_ aJ . , 1981;
Wedrychowicz et al^., 1984; East et_ al_., 1988; Roach et_ a l . ,
1988;Ey, 1988; Malhotra, 1988). The assay conditions were
optimised in order to get the optimum resul t s . The per
oxidase labelled goat ant i-rabbit Ig conjugate was used to
measure the total antibody responses in the immune rabbit
sera . The rabbit anti-adult and rabbit anti-larval serum
showed high t i te rs of parasite specific antibodies against
both adult and larval somatic antigens thereby showing a
high level of cross-react ivi ty between the two stages of
A.ceylanicum. This technique has been used successfully
to detect the antibody levels against N.brasiliensis(Wedrychowicz
et ad., 1984) and N.dubius (East et_ al^., 1988). ELISA has
also been used to detect the cross reactivity between the
two helminth parasi tes T.trichuria and T.muris (Roach et
al^., 1988).
Immunoelectrophoretic techniques have generally been
used to study the antigenic compositions of several parasites
and have also proved to be useful in the identification of
common or cross-reactive as well as stage and parasite specific
antigens (Williams, 1970; Neilson, 1978; Kaushal et_ al_., 1982;
Lobos and Weiss, 1985; Cavalan et_ a]_., 1985; Malhotra et
al^., 1986). In the present study the techniques of immuno-
electrophoresis (lEP) and crossed immunoelectrophoresis (CIE)

70
have been employed to analyse the antigenic patterns of adult
and infective larval stage of A.ceylanicum. The antigenic
analysis by lEP revealed less number of precipitin bands
in the somatic extracts from the larval stage of th is parasite
while the somatic extracts from the adult stage showed more
precipitin bands. On the contrary, Williams (1970) has shown
more complex pattern for infective larval stage of A. caninum
than for adults using the same technique of Immunoelectro
phores is . This technique has also been used to analyse
the antigens of some cestodes v iz . Hymenolepis nana and
H.diminuta and showed the presence of 14 and 9 precipitin
arcs with their respective sera (Manas et_ al_., 1985). Employ
ing the same technique of lEP the antigens of several helminth
parasi tes and also the cross-reactive antigens among these
have been analysed (Choi and Lee, 1979).
Antigenic characterisation by CIE also revealed lower
number of antigens in the infective larval stage as compared
to adults of A.ceylanicum. This technique helped in better
separation of antigens from these stages. Various concentrations
of gamma-globulin fraction from hyperimmune rabbit sera
were tried in order to get a sharp and balanced pattern
of the antigenic peaks and a 20% concentration of gamma
globulin fraction was found to be optimum for resolving the
antigenic pattern of adults of A.ceylanicum, while a 10%
roncf^ntration of gamma-globulin fraction was required
for the separation of larval antigens. On lowering the anti-

71
serum concentration further, the faint antigenic peaks could
not be seen clearly while the higher concentrations resulted
in poor resolution of some of the closely placed antigenic
peaks. The CIE technique showed the presence of 34 antigens
in adults and 19 in larvae of A.ceylanicum. Out of the two
cathodic antigens of infective larvae of A. ceylanicum, one
was slow migrating and precipitated in the first dimension
gel only. This could be due to the presence of a low charge
on the antigen. CIE technique has also been used for antigenic
analysis of many helminth parasites including hookworm para
s i t e s . Twenty six antigenic components have been visualised
by CIE in the extracts of N.brasil iensis (Cavalan et a l . ,
1985). Kaushal et al - (1982) have demonstrated the presence
of 35 antigens in the extracts from B.malayi adults using
the hyperimmune rabbit serum. At least 21 antigens in the
isolates of 0.volvulus (Lobos and Weiss, 1985) and 22-24
antigens in S.cervi adults (Malhotra et_ a
72
certain helminth paras i tes , like tapeworms, whipworms have
been identified (Milner et_ ail_., 1987; Correa-Oliveira et a l . ,
1988; Roach et al_., 1988). Cross protection has also been
observed between A.ceylanicum and N.brasiliensis (Gupta
and Katiyar, 1985). In view of these findings an analysis
of the common or cross-reactive as well as specific antigens
between different stages and also between different parasites
would be important for understanding the mechanisms involved
in protective immune responses. In the present study we
have analysed the common/cross-reactive as well as stage
specific antigens displayed by two developmental stages of
A.ceylanicum. Common or shared antigens exist between the
two stages as evident by the reactivity of somatic extracts
from these stages with heterologous sera in CIE as well as
by CLIE analaysis. CIE analysis of AcA and AcL with sera
against both these stages (adult and larvae) showed the p re
sence of 12 (AcA) antigens specific to adult and 5 (AcL)
antigens specific to larval stage. CLIE analysis also revealed
same number of stage specific (12 adult specific and 5 larvae
specific) antigens of A.ceylanicum. The presence of common
antigens between the adult and the larval stages of A.caninum
has been reported earl ier employing immunoelectrophoretic
technique (Williams, 1970). Common or cross-reactive antigens
between different stages as well as stage specific antigens
have been identified for other parasi tes including hookworms
viz . D.viteae (Neilson, 1978); B.malayi (Kaushal et aJ_.,

73
1982); T.spi ra l is (Phil l ip et_ al^., 1980; Parkhouse and Clark,
1983; Parkhouse and Ortegga, 1984); S.cervi (Malhotra et
a l . , 1986; Kaushal £t_ al^., 1987); N.americanus (Carr and
Pr i tchard, 1987); N.dubius (Adams et_ aJ^., 1988); P.knowlesi
(Singh, 1990).
In the present study an effort has also been made
to analyse the common or cross-reactive antigens between
the two hookworm parasi tes ; A.ceylanicum and N.brasi l iensis .
Antigenic cross-reactivi ty between the adult and larval stages
of A.ceylanicum and N.brasil iensis was studied using CIE
and CLIE. These studies showed that out of a total of 34
antigens present in A.ceylanicum adul ts , only 12 and 8 were
found to be common -or cross reactive respectively with adult
(NbA) and larval (NbL) stages of N.brasi l iensis . However,
the larval stage of A.ceylanicum showed 13 (out of 19) common
with both adult and larvae of N.brasi l iensis . These studies
have also revealed A.ceylanicum parasite specific antigens.
Presence of cross-react ivi ty among different helminth parasites
has been shown ear l ier and the cross reactive/common as
well as parasite specific antigens have been identified (Williams
1970; Rao et_ al^., 1980; Turner e; al^., 1980; Maizels et aJ^.,
1983; Gottstein et a l . , 1986; Kennedy et_ al^., 1987; Roach
et_ a]_., 1988).
Most of the studies conducted so far have shown
that protective immunity to hookworm parasi tes can be induced
In experimental animals by live parasi t ic stages and HJSO

74
by immunising with somatic extracts from adults and larvae
(Foster, 1935; Miller, 1971, 1979; Carroll and Grove, 1985;
Gupta and Katiyar, 1985). In the present study the protective
potential of A.ceylanicum antigens has been evaluated by
immunising the hamsters with somatic ext rac ts . The sera
obtained from the hamsters immunised with AcA and AcL,
before challenge, showed significant levels of parasite specific
antibodies. The immunodiffusion and lEP analysis revealed
the presence of precipitin antibodies in these immune sera
as well as the number of AcA and AcL antigens recognised
by these sera . The immunised hamsters when challenged
with live infective larvae showed significant resistence to
challenge infection. The infective larval stage proved to
be better in inducing protective immunity as it exhibited
over 90% protection (reduction in worm burden) as compared
to 63% obtained in the hamsters immunised with adult stage.
Carroll and Grove (1985) have observed 74% reduction in
worm burden in dogs immunised with the somatic extracts
from adult A.ceylanicum About 76% reduction in worm burden
was obtained in hamsters immunised with live infective larvae
of A.ceylanicum (Gupta and Katiyar, 1985).
Therefore, in the present study we have been able
to obtain basic information about the protein and antigenic
composition of adult and larval stages of A.ceylanicum.
These studies have revealed less complex nature of infective
larvae as compared to the adults as evident by the less

75
number of proteins and antigens in the larval stage. Certain
common or cross-reactive antigens between the two different
stages (adult and larvae) of A.ceylanicum and between the
two hookworm parasi tes (A.ceylanicum and N.brasilieisns)
have also been identified. These studies also showed the
presence of certain stage and parasite specific antigens of
A.ceylanicum as indicated by CLIE, analysis . The level
of cross reactivity between the two life stages has also been
studied by quantitating the parasite specific antibodies in
immune rabbit sera by ELISA. On evaluating the protective
potential of adult and larval somatic extracts , it has been
found that the larval stage is capable of exhibiting high
degree of protection, however, i t certainly needs further
evaluation in order to determine the possible mechanism of
protective immunity and also to identify the antigenic molecules
involved in inducing the protective immune responses.

SECTION II
EXCRETORY-SECRETORY PRODUCTS FROM
ADULT ANCYLOSTOMA CEYLANICUM

76
The excretory-secretory (E-S) products, released
during HI vi tro incubation of live paras i tes , are known to
be less complex in nature than somatic antigens and have
been shown to be of potential importance not only for establish
ing specific diagnostic techniques (Maizels et al^., 1984, 1986;
Brindley et al^., 1988; Marrero et al^., 1988) but also in induc
ing protective immunity against certain helminth parasites
(Thorson, 1951; Jenkins and Wakelin, 1983; Sugane and Oshima,
1983; Maizels et al^.. 1984; Gamble, 1985; Urban and Romanowski,
1985; Silberstein and Despommier, 1985; Badley et ad., 1987;
Mimori et a l . , 1987; Ey, 1988). Although a few reports are
available showing the protective potential of E-S products
released by the hookworm parasi tes (Katiyar et al^., 1972;
Govila et_al_., 1973; Murray et al^., 1979; Ey, 1988a,b), l i t t le
is known about the antigens present in these products. This
may be due to the difficulties in obtaining sufficient quantities
of E-S products . In the present studies we have optimised
the conditions for the preparation of E-S products from the
adults of Ancylostoma ceylanicum and analysed them employing
certain immunoelectrophoretic techniques. The protective
potential of E-S antigens was also studied.
PREPARATION OF E-S PRODUCTS
The conditions for the preparation of E-S products
from adult A.ceylanicum parasites were standardised. Different
media (RPMI-1640, DMEM and Ringer's solution) were tried

77
for short term j ^ vitro maintenance of A.ceylanicum adults
and the quantities of E-S products obtained in different media
are shown in Table 9. Among the three media used. Ringer's
solution proved to be the best for the preparation of E-S
products as the motility of worms was found to be maximum
in th is medium and the worms could be maintained up to
6 h without the change of medium. Analysis of E-S products
showed the release of 125±25 ug protein per 100 adult worms
in Ringer' s solution, while the protein released in the other
two media (RPMI-1640 and DMEM) was 95±15 ug per 100 adult
worms. Immunoelectrophoretic analysis of the E-S products
prepared in different media, using immune rabbit serum raised
against adult soma.tic antigen (AcA), also revealed almost
the same number of precipitin bands in all the three E>S
preparations i . e . 5-6 precipitin bands in RPMI-1640 and DMEM
and 6-7 in the Ringer's solution (Table 9 ) . Hence, in further
experiments, the preparation of E-S products was done using
Ringer's solution. In order to determine the optimum time
for the collection of E-S products, the adult worms were
maintained iji vi t ro and the E-S products were collected at
different hours by changing the medium at 2 h intervals .
The maximum amount of protein (125 ug/100 adult worms)
was released by the worms during the initial two hours.
After that , a decrease in the amount of protein released was
observed and i t became one fifth during 6-8 h period of
in vitro incubation (Table 1 0 ) . ^ ^ came sluggish
Ace No,

78
0) (0
(1) I—I
s
B 9 u
4) U
u a
T( O U
a
o 4-" i) u u t) <J) I
!>. o
4-> (U VH
u X w
Xi
E2
13 0) E
c U i)
(U
E 3
z
a, w
0) C
D-J3
c +J O c o o o
T3 S bc 4J ? 3
(0
E o
C -•H (0 <U rt •!->
E 3
•H
<u
+1
in
o
3 +J •rt +J (0
c
u o E 0)
(0 OH
(A O
I I
+1
E 3
•H TJ
<1)
c <u (A 10
w E 3 E
o o 0)
J3
in CM +1 in
C O
O
i)
c 0) E
• H
^ (U P-X (U
+J c 0) v< 4)
MH •H tJ
4) 4)
S H 0
c m 4} E 4}
J3 •*->
v >H nj
en + j 1—(
3
4)
4J XI H
CU W 1—1
c • H
4) 05 3
03 (fl ^
E 3 V, 4) Oi
< O < • H •» ->
c a
XI J3 (0 «
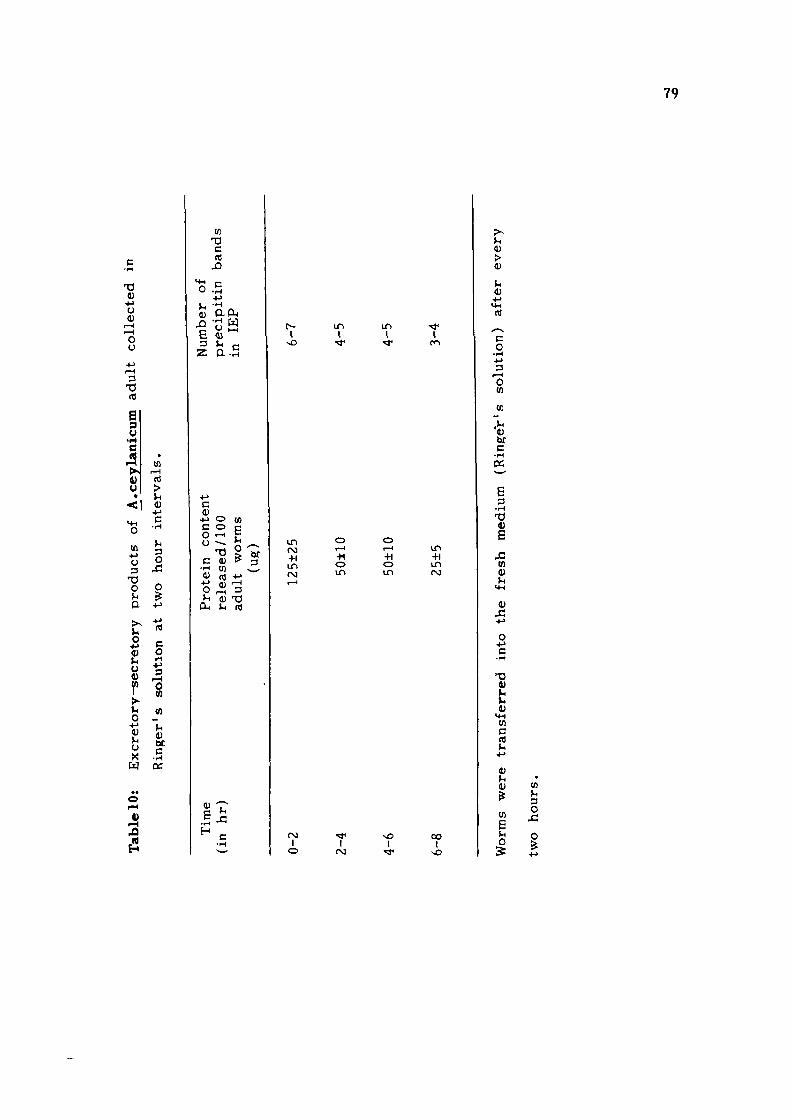
79
Tl 0}
• M
o (U 1—1 1—<
0 u +-> r—I 3
T) rt
B 9 U
•H C
^ >> u u •
< 4-1 o
o
T) 0 ^H
a
0 V
1 >-u o
u o
• (fl r—(
rt > >H
ii •»-> c •H
3 0
xi 0 ^
+ J
•t->
c o H
p .—1 s (0
0)
c
.a
OS
en T) c
XI
S o-o.
E « 3 >H _
Z D-.S
E O
c 0)
+J O 0) c o o o
'^ ft C (U l>
•H to
IT I i n I.
i n
+1 in (M
+1 o in
+1 o in
in +1 in (NJ
(NJ
O 1 1
(VI
1 1
0 0
1
(U
>
u 4)
c o
o tn
<u bf c
•H 06
E 3
• H
- O 4}
E
(n
0)
0) O
•(->
c
0} •4-1 (0 c (\)
(U
u
(fl E o
tn
3 O
O

80
after 6 h of incubation and could not regain their motility
even after transferring them to the fresh medium. Hence,
an incubation period of 6 h was used further for the collection
of E-S products. Fig. 17 depicts the lEP analysis of E-S
products collected at every two hours. The number of bands
was maximum ( i . e . 6-7 precipitin bands) during the initial
two hours and the number gradually decreased to 4-5 in the
E-S products collected during 2-4 h and 4-6 h of incubation.
Only 3-4 precipitin bands could be observed in the medium
collected during 6-8 h of incubation.
IMMUNISATION OF RABBITS AND QUANTITATION OF PARASITE
SPECIFIC ANTIBODY BT ENZTME LINKED IMMUNOSORBENT ASSAY
The rabbi ts were immunized with E-S products (pre
pared in Ringer's solution) and sera obtained were tested
for parasite specific antibodies by enzyme linked immunosorbent
(ELJISA). The assay conditions such as the optimum concent
rations of OPD, H-0_ and optimum dilution of antibody conju
gate have already been described in Section I. In order
to determine the optimum antigen concentration different concent
rations of E-S antigens (0.015 ug to 1 ug/well) were tested
at three fixed serum dilutions of rabbit anti-E-S serum (1:500,
1:1000, 1:2000) and it was found to be 0.50 ug/well. After
that , there was not significant increase in ELISA specific
activity on increasing the antigen concentration to 1 ug/well
(Fig. 18). After determining the optimum concentration of

81
Fig* 17 • Immunoelectrophoretic patterns of A.ceylanicum E-S
products collected at 2 hr ( a ) , 4 h ( b ) , 6 h (c) and
8 h (d) using rabbi t anti- / \cAserum.

82
0.20 O.AO 0.60 0.80
Antigen concentration ( / jg /we l l )
1
1.0
Fig. 18: Effect of different concentrations of excretory-secretory products from A.ceylanicam adults on the specific activity in enzyme linked immunosorbent assay using 1:500 (X-X); 1:1000 (0-0) and 1:2000 (A-A ) dilutions of rabbit anti-E-S serum.

83
E-S antigen, the antibody t i te rs of parasite specific antibodies
in all the three immune rabbit sera (rabbi t anti-E-S, rabbit
anti-Ac A and rabbit anti-AcL) were determined and the t i t r a
tion curves are shown in Fig. 19. All the three titration
curves of E-S products obtained with rabbi t anti-E-S and
rabbit anti-adult and rabbit anti-larval sera were parallel
and showed linearity upto a serum dilution of 1/16000. A
reciprocal ELISA t i te r value of 256,000 was obtained for rabbit
anti-E-S, while rabbit anti-AcA and rabbit anti-AcL sera showed
an antibody t i te r of 128,000.
ANTIGENIC ANALYSIS OF E-S PRODUCTS
The E-S products preparation in Ringer's solution
were analysed by immunoelectrophoresis (lEP) using rabbit
anti-E-S serum which showed the presence of 8 precipitin
bands in E-S products (Fig. 20). The E-S products prepared
in other two media (RPMI-1640 and DMEM) were alao analysed
in lEP with rabbi t anti-E-S serum and almost the same number
of precipit in bands (7-8) were observed with slight differences
in the intensities of certain precipitin bands (Fig. 20).
The E-S products were further analysed by crossed
immunoelectrophoresis (CIE). The CIE pattern of E-S products
using rabbi t anti-E-S serum is shown in Fig. 21a and the
diagrammatic representation of the same is given in Fig.21b.
Strong antigenic peaks are shown by solid lines whereas the
broken lines indicate that the faint antigenic peaks. Each

84
8 o
o o
o
o - o o
c o 2 •5
E
o
"ifl u O w
a ij cr
1 ^
S -P " 0
rj -H C
'2 4' = £ ^ E
/ - . 4) +jT3
? 1 ^ ^ « 5
0.^ W5 (U
W O >.
c n! <iJ
n) C > if>
•*^ •*-• ^
(Z6VaO) '^H'^IPP DijrDads •H
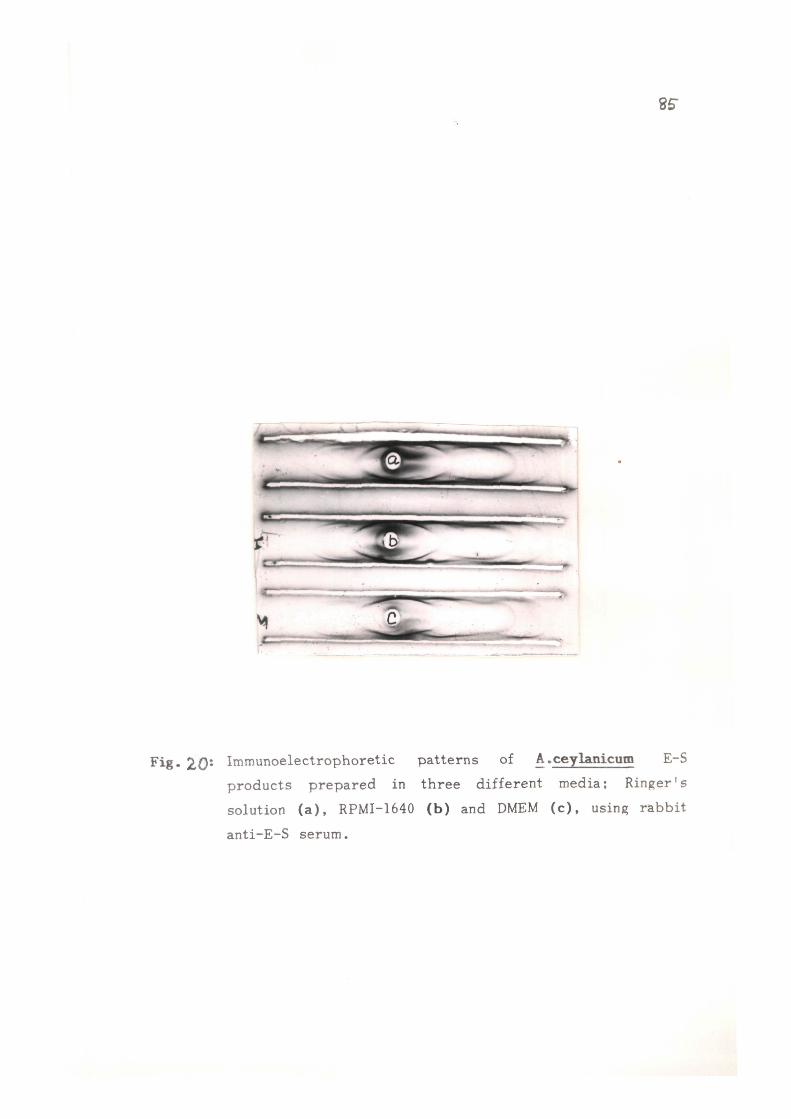
85-
Fig. 20" Immunoelectrophoretic patterns of A.ceylanicum E-S
products prepared in three different media; Ringer's
solution ( a ) , RPMI-1640 (b) and DMEM (c) , using rabbit
anti-E-S serum.

2€
1
•
'
• -
V
J ^ y
- ^ —^
y
Fig. 21: Crossed immunoelectrophoretic pattern of A.ceylanicum
adult E-S products using 10% rabbit anti-E-S serum
(a) Protein stained pattern, (b) Diagrammatic represent
ation. Solid lines represent strong antigenic peaks.
Arrows indicate the point of application of antigen.
Anode ( + ) is to the right in first dimension and to
the top in second dimension.

87
antigen is denoted by a single peak and CIE revealed 16
antigens in A. ceylanicum E-S products, 13 moving towards
anode and 3 towards cathode.
Common antigens between the E-S products and the somatic
extracts
In order to identify the common antigens between
the E-S products and the somatic extract from adult and larval
stages of A. ceylanicum, the E-S products were analysed in
lEP with sera raised against the somatic antigens. Six and
4 precipitin bands were observed in E-S products with rabbit
anti-AcA and rabbit anti-AcL serum respectively (Fig. 22),
Almost the same number of antigens were detected in somatic
extracts from adult and larvae, when tested in lEP against
the rabbit anti-E-S serum (data not shown). The E-S products
were further analysed by CIE with sera raised against somatic
antigens. CIE analysis of E-S products with rabbit anti-adult
serum showed the presence of 12 antigens, out of which 11
antigens were anodic in nature and one antigen moved towards
the cathode (Fig. 23). When rabbi t anti- larval serum was
employed in CIE and 6-7 antigenic neaks were
observed in E-S products and all of them appeared to be
anodic (Fig. 24). The results of lEP and CIE analysis of
E-S products with immune rabbit sera are summarised in Table
11. The common antigenic determinants between the E-S pro
ducts and somatic extracts from adult and larval stages of

8S
Fig. 22: Immunoelectrophoretic patterns of A.ceylanicum adult
E-S products with rabbit anti-E-S serum (1) , rabbit
anti-adult serum (2) and rabbit anti-larval serum (3) .
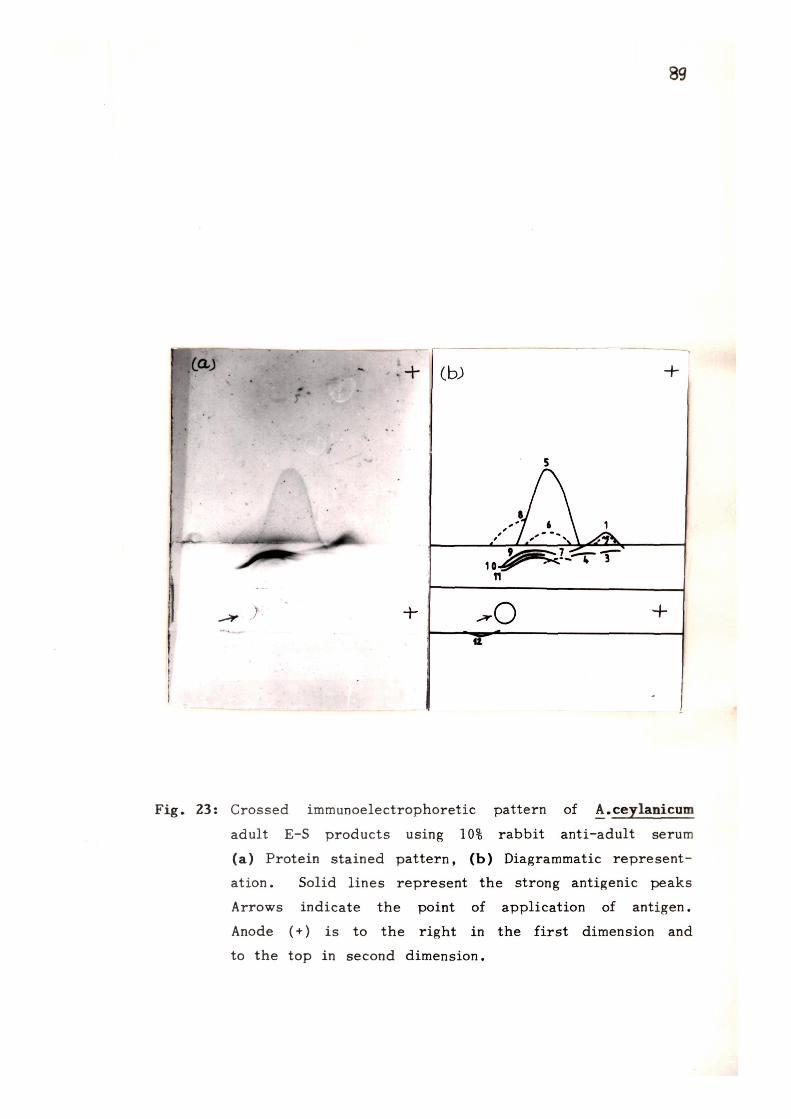
89
Fig. 23: Crossed immunoelectrophoretic pattern of A.ceylanicum
adult E-S products using 10% rabbit anti-adult serum
(a) Protein stained pattern, (b) Diagrammatic represent
ation. Solid lines represent the strong antigenic peaks
Arrows indicate the point of application of antigen.
Anode ( + ) is to the right in the first dimension and
to the top in second dimension.
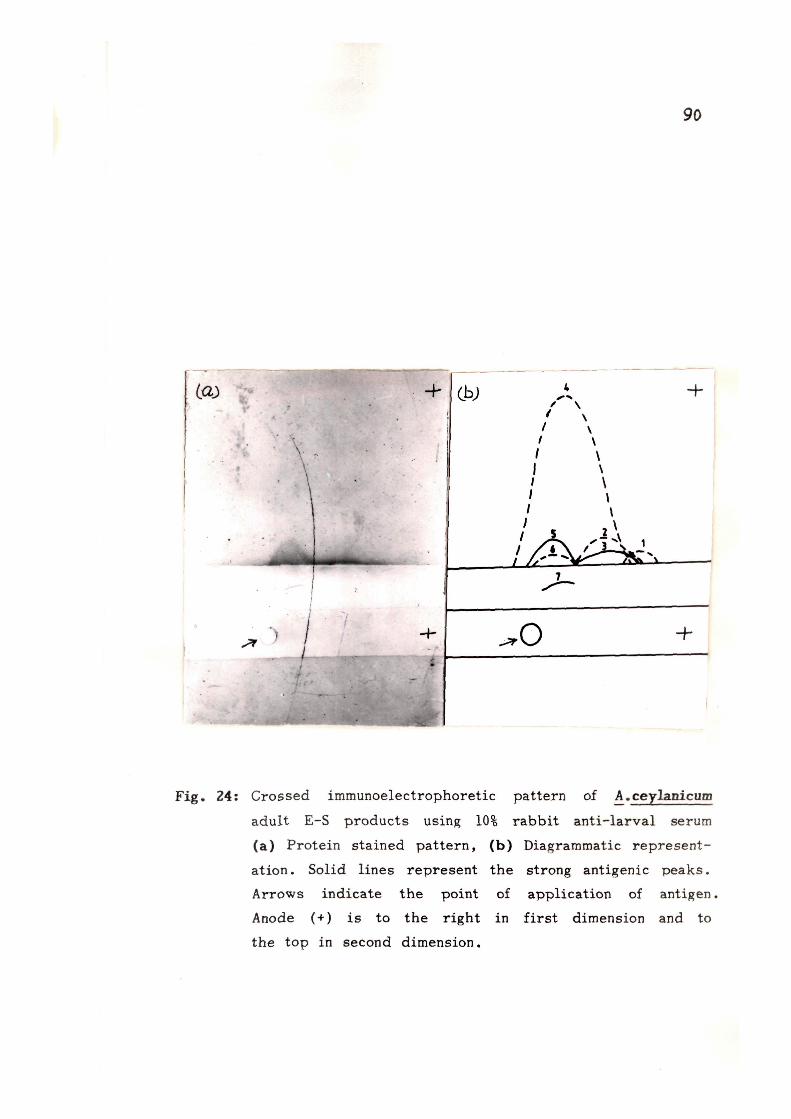
90
Fig. 24: Crossed immunoelectrophoretic pattern of A.ceylanicum
adult E-S products using 10% rabbit anti-larval serum
(a) Protein stained pattern, (b) Diagrammatic represent
ation. Solid lines represent the strong antigenic peaks.
Arrows indicate the point of application of antigen.
Anode ( + ) is to the right in first dimension and to
the top in second dimension.

91
3
nl
B s
<H U a
i 4) U
•»-> u 3
0
a I
to
1—I
c
c • H
c <
ni 0)
a tn
tJ c rt
X!
•H
0 o
0
0)
B 3 z
E 3
in •rt c <
B «i OC
• H
c <c
••
V I - t jQ ,« H
<u 3
• H
c ,n o 0)
H
u •H
C
••J
m O
CO I—1
o •H
c
t/3 I
w
(0 •H tn
u o
X!
0 o c u 3 -*->
1 <" I—( 0)
l-H t ^
E 3 u (U to
Ul 1 1 1
• H + J c A
• H
^ 2 rt oe;
e 3 v< 0) to
< • < j
.A • p
c «)
• H J3 X) n)
oe;
E 3 U V to
J u < 1 • H
4->
c rt'
• H J3 JD (d
OS
E 3 >H 0) to
Vi 1
W 1 •1-1 4 J
c ni
•H ja Xi Rt
CHS
E 3 U 4) to
< U
< 1 • H
+ J c nl
•H Xi XI (6 cc
E 3 U 1) (0
J o < 1 1
•H + J
c n)
•H J3 ^ «)
OS
I w
0 c 3 H E
•H
0) to to O U
U
to • H to (1)
0 X! a o u +> u 9) t)
I I r
•»->
u .•« )4
*> X 4)
u •1-1 ••J
«) E 0 to
1—1 3
T3 R)
B 9 U
1 •< V u •
< II
< V <
••J u nt U
•»->
X 4)
U • H ••J
(« E 0 to
r—1
n) > nl
r - t
B a u • H
J •^ 0) u •
1 < II
J o <
3
e o >H
«*H
to •«-> u 3
T3 o u a.
>^
0 •*->
4) o 4J to 1 > u 0
4^ t) u V X
1 w II
t/2 1
w

92
A.ceylanicum were further analysed by doing Tandem crossed
immunoelectrophoresis (TIE) of E-S products with adult (AcA)
and larval (AcL) somatic extracts and the TIE patterns are
shown in Fig. 25. On comparing the TIE pattern of E-S pro
ducts plus AcA with CIE pattern of E-S antigens (Fig.2^0)
11 antigens were found to be common with adult stage (Fig.
25 b ) . TIE analysis of E-S products plus AcL is shown in
Fig. 25a, which revealed the presence of 6 common antigens
between the E-S products and the larval somatic ext rac ts .
Presence of host serum proteins in E-S products
In order to detect the presence of host proteins in
E-S products hyperimmune rabbit serum was raised against
normal hamster serum (NHS). When E-S products were reacted
with rabbi t anti-NHS serum in CIE, 6 antigenic peaks were
observed (Fig. 26), suggesting thereby the presence of host
proteins in the excretory-secretory products of A.ceylanicum
adults .
IMMUNISATION OF HAMSTERS WITH EXCRETORY-SECRETORY PRO
DUCTS AND EVALUATION OF IMMUNISED HAMSTERS FOR PRO
TECTIVE IMMUNITY
In view of potential importance of E-S products of
parasit ic nematodes including hookworm parasi tes in develop
ment of protective immunity, hamsters were immunised with
E-S antigens from adult A.ceylanicum followed by subsequent
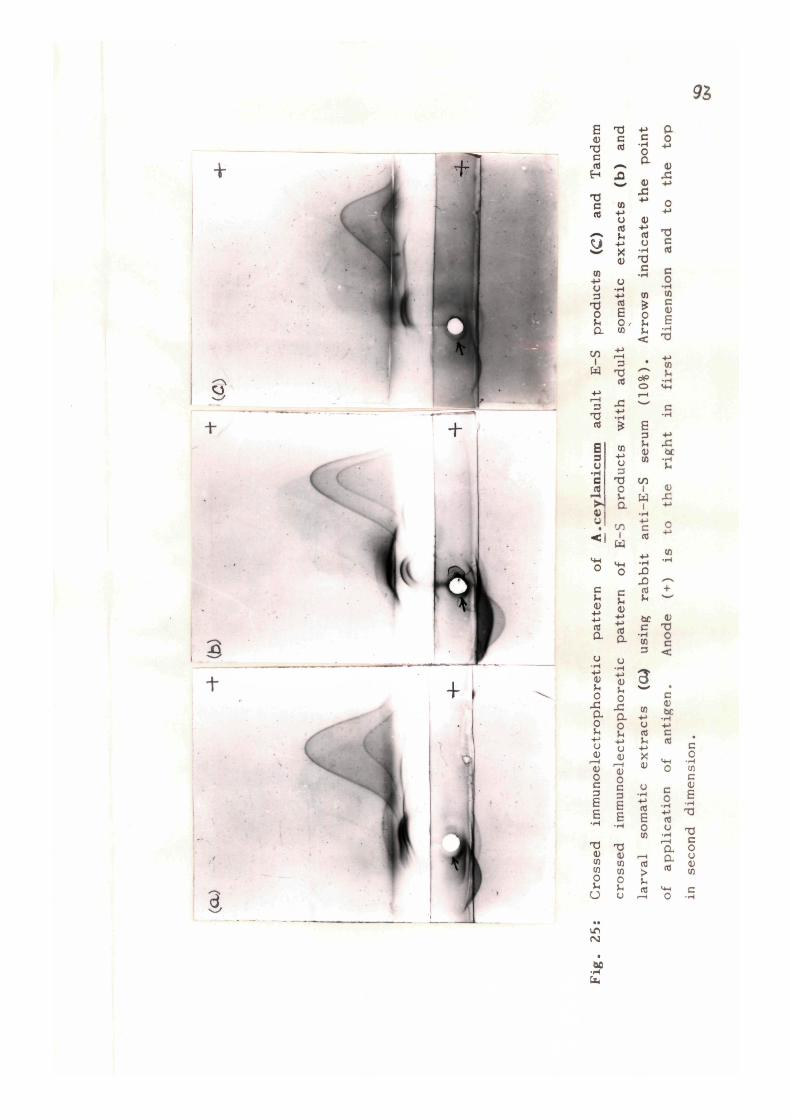
95
< . ^
t
V
^
E T3 c 0)
c n)
u CD
+-> o 3
t! O
a V2
I
w
3
nt
B s u
u
c u 0)
m
a
^ o x; a o u + j
o 0)
.—( (U o c 3 E E
T3 (U
(n O
O
IT)
no c nt
1i ^ c o o ^
J3
u
•t->
X 0)
3
u 3
T ) O J-.
a I
w <4-4
o
c u t)
+->
a
nt o
C
CO
o
<
O
E 3 u V 10
C/1 I
w I
c
nj +
C T3
to c => <
0)
o X! a o u +-> o v
I — I
(U o c 3 E E
<D (0 U] o u u
0)
E o m
>
c «)
c o
•H to c i) E
CO
x: bC
4) X
C 0)
in ac u
c
c o
s a
c o
•H CO c 1) E
•H Tl 73 c o o i) CO
O -H
• H

4-
i ' ^
Fig. 26: Crossed immunoelectrophoretic pattern of A.ceylanicum
E-S products using rabbit anti-normal hamster serum.
Arrows indicate the point of application of antigen.
Anode ( + ) is to the right in the first dimension and
to the top in second dimension.

95
challenge infection. The immunised hamsters were bled 15
days after the first injection. Further immunisation and
bleeding was done alternately at weekly intervals and sera
obtained after each injection were tested by enzyme linked
immunosorbent assay (ELISA) for the presence of parasite
specific antibodies. The antibody t i t e r s of hamster anti-
E-S sera (obtained after every injection) are expressed as
geometrical mean reciprocal t i te r (GMRT) and are given In
Table 12. No antibody could be detected in the serum obtained
after 1st injection. A GMRT value of 50.00 was obtained
after the 2nd injection which increased to GMRT values of
200.00 and 303.14 after the 4th and 5th injections respectively.
The presence of precipitin antibodies in immune hamster sera
was analysed by immunodiffusion (ID). No precipitin bands
could be observed in sera taken after first injection while
one and 2 precipitin bands were observed in ID after the
2nd and the 3rd injections respect ively . The number of preci
pitin bands increased to 3 after the 4th injection and remained
constant after that ( i . e . after 5th injection). The results
of ID analysis are summarised in Table 13. The immune hamster
serum obtained after the 5th injection was theln tested in
lEP and 4 precipit in bands were observed in E-S products
(Fig. 2 7). The immune hamsters were then evaluated for
protective immunity by challenging them with an oral dose
of 60 L- one week after the fifth injection. The percent
reduction in worm burden at 21 days post challenge showed

96
nl (0 (fl n)
c V
O (n o c 3 E E
T3 (U
c
E N
c
>
u
(n E nl
J5
9) c 3 E E
0)
O JD •H +J B
<
(M
n) H
c o
4J ft)
c o
•H
u J3
>
H OS S o
0)
u
I <
T3 ^
C
o •H
o 0)
c
B
o
•*
o
o o
• o o (M
i n
c c •H
c
E 3 0) (0
• H -t-> c <
B 0) M
•H
B
<
O o o i n
Q Z
I
I
B <
<U • » - '
cn E m X
4) >
E o u
T3 0) B
J3 O
0)
I W
(0 •(->
u 3 73 0 h 0.
>-u o
•t-> 0) >H
o 0) cn 1 > -u o ••-> 0)
o X W
II
V5 1
W
nt u 0 U
a •H
u 4) U
B
0)
E i-H n) u
•H
0) E 0
(0 «)
73 V (0 (0 4J
a X
• (0 B 0}
• H + J B n)
C/2 1
U
0) JC ^ nl
(0
- t j • H
^ 4) TJ ^
•!-> •H •t-»
0) N
• H B 3
>- E 73 0
J3 • H
-(-> B n)
0) X
E • H
(0 ^4
m E n)
H Xi
• V
r—1 JD n)
-t->
u 0)
+ J 4}
13
+ j
o B
^ TJ 0
J3 •H + J B < II
Q 2

97
•g
3 T3
E o u
u
3
0
a C/2
I
• H (fl t>-
—) nt c n)
c o
E 3 U 0) en
t/2 1
W
3
O C 3
e B
c
u 0)
-t-" tn E n)
X !
en
c
o d)
a O
0) JD
E 3
z
E 3 U V (A
•rt c <
c
• H 4-> C
<
c o
> 0>
c o
+» u > 0)
c
I—I i)
T( B
1—( t-H
•!->
in t-H
•;3 o 4)
c" •H
c 0
•H 4-* o 0)
E 3 U i) (0
{/3 I
w I
•H •(-> c rt
0)
(fl E M
I
w
(0 V< 4)
•t-> (0
E n)
u n)
u n) 0)
B O
U 0)
43 o nt
(U
0)
c o
1)
(A
0) ••-» (Q
E n)
J3
c 0)
E •H
K X 0)
0) 0)
u
c 4)
E
x: (U Vi cd
D
>
(0 4) 3
.—I nt >
(U N
•H B 3 E E
B a u
•H
I H V u <l 3
T) nt
E o u
u 3
T ) O
> •
U
o (U u u 0) (A I >~ u o
u u X w
II
(U

92>
Fig. 27: Immunoelectrophoretic pattern of E-S products from
A.ceylanicum adults using hamster anti-E-S serum.

100
the degree of protection. Seventy five percent reduction
in worm burden could be observed as compared to the un-
immunised infected controls (Table 14).
DISCUSSION
The release of antigenic molecules (excretory-secretory
products) during jii^ vitro incubation has been well documented
for a wide variety of nematode parasi tes v i z . , T. spiral is
(Despommier and Muller, 1976; Parkhouse and Clark, 1983;
Silberstein and Despommier, 1985; Gamble, 1985; Capo et a l . ,
1986), T.canis and^ T.cati (Sugane and Oshima, 1983; Maizels
et al^., 1984; Badley 1987a,b; Kennedy et al^., 1987a), T.muris
(Jenkins and Wakelin, 1983), A.suum and A.lumbricoides (Urban
and Romanowski, 1985; Kennedy and Qureshi, 1986; Kennedy
e ^ a l . , 1987b), N.americanus (Carr and Pr i tchard , 1986, 1987)
and H. poly gyrus (Ey 1988a,b).
The JL£ vi t ro released or the excretory-secretory
(E-S) products of parasit ic nematodes including hookworms
have been shown to be more immunogenic as compared to soma
tic antigens (Thorson, 1951; Campbell, 1956; Chipman, 1957;
Katiyar et al^., 1972; Govila et al^., 1973; Poulain et al^.,
1976; Murray et al^., 1979; Silberstein and Despommier, 1984;
Mimori et al_., 1987). In spite of their less complex nature,
relatively l i t t le effort has been made to evaluate critically

101
the potential usefulness of E-S antigens in inducing protective
imnnunity to hookworm parasi tes and also to analyse the anti
gens present in these products.
In the present study, we have optimised the conditions
tor preparing the E-S products from adult hookworm parasite
A.ceylanicum and have analysed the E-S products using hyper
immune rabbit sera employing certain immunochemical techniques.
The conditions for the short term iji vitro maintenance of
adult A.ceylanicum and preparation of E-S products were stand
ardised in order to get the maximum viabil i ty of the worms
during in vitro incubation and also the maximum yield of
E-S products. The time for ^ vi t ro incubation was also
optimised and was- found to be 6 h . The motility of the
worms started decreasing when the worms were kept for the
longer period. The parasite could not be maintained beyond
8 h even after transferring them into the fresh medium. A
period of 6 h for jji vi tro incubation of worms was chosen
in order to avoid the contamination of E-S products with
the products from the dead and disintegrating parasi tes .
Since there was not any significant difference in the amount
of protein released as well as the antigenic pattern of E-S
products prepared in the three media (Ringer's solution,
RPMI-1640, DMEM), Ringer's solution has been chosen to prepare
the E-S products because this is much simpler (containing
only salts and glucose) and more economical medium as compared
to RPMI-1640 and DMEM. The amount of protein released

102
by A.ceylanicum adults in 6 h (125±25 ug/100 adults) was
in accordance with other findings by Rotmans et ajl_. (1981),
who have also shown the release of 100 ug E-S products per
100 worms of Schistosoma mansoni by maintaining them in
M 199. Minute quantities of E-S products have been shown
to be released by the larvae of T.canis (0.8 ug/100 larvae/day)
and H.polygyrus ( 20 ng/100 larvae/day) during in vitro incu
bation (Badley et_ a l . , 1987a; Ey, 1988). The human filarial
parasi te: Brugia malayi has been shown to release 18 ug of
E-S products/100 adult worms (Kaushal et al_., 1982). However,
considerably high amount of E-S products (12-15 mg/100 adult
worms) were released by the bovine filarial parasite Setaria
cervi (Malhotra et al_., 1987).
The t i t e r s of parasite specific antibodies in rabbit
anti-E-S serum were quantitated by ELISA. This test has
been used ear l ier for detecting the antibodies against nematode
parasites including hookworms (Wedrychowicz et al^., 1984;
Roach e^al^. ,1988; East et_ al., 1988). The rabbit sera raised
against E-S products gave quite a high antibody t i ter and
the t i ter with anti-E-S serum was one dilution higher when
compared to anti-adult and anti- larval sera. The highti tered
hyperimmune rabbi t anti-E-S serum was used to analyse the
antigens present in E-S products of A. ceylanicum adults by
immunoelectrophoretic techniques. The antigenic analysis by
lEP revealed the presence of 8 precipitin bands in A.ceylanicum
E-S products. Almost same number of antigens (7 precipitin

103
bands) have been reported in the E-S products of S.nyansoni
(Rotmans et al^., 1981), A.cantonensis (Techasoponmani and
Sirisinha, 1980) and S.cervi (Malhotra et al^., 1987) using
lEP. NKU et al^. (1981) could detect at least 4 precipitin
lines in E-S products of 0 . volvulus mf with patient serum
in lEP. Antigenic characterization by CIE resulted in better
resolution of A. ceylanicum E-S products. Out of different
serum concentrations t r i ed , 10% concentration of immune rabbit
serum was found optimum for the better separation of antigenic
peaks by CIE. On decreasing the antiserum concentration,
the faint antigenic peaks disappeared while the use of higher
serum concentration resulted in the poor resolution of certain
antigenic peaks . A total of 16 antigens were identified in
the E-S products of adult A.ceylanicum out of which 8 were
found to be major. The E-S products of A.ceylanicum appeared
to be less complex in comparison to the somatic antigenic
preparation from adult stage as it revealed the presence of
34 antigens in CIE. The E-S products of A.ceylanicum may
contain some heterogenous antigens as indicated by the doubling
of antigenic peaks 11, 12. This could be due to a number
of reasons viz; the presence of isoenzymes genetic variants
of the same protein, or protein complexes and their monomers
or the presence of molecular heterogeneity deriving from
different degree of glycosylation.
The E-S products appeared to share antigenic determi
nants with the somatic antigens from adult and infective Jarval

104
stages of A.ceylanicum as shown by the reactivity of E-S
products with rabbit sera raised against the somatic antigens.
Twelve (nos.2-5, 7-13, 16) and 7 (nos. 1-3, 5, 8-10) antigens
from a total of 16 antigens of E-S products were recogined
bv the rabbit anti-adult and rabbit anti-larval serum respect
ively. The presence of common antigenic determinants between
the E-S products and somatic extracts from adult and larvae
of A.ceylanicum was further confirmed by doing Tandem crossed
immunoelectrophoresis, which also showed almost the same
number of common antigens. Antigen sharing between E-S pro
ducts and the somatic antigens has been observed for a number
of paras i tes . Kaushal et_ al_. (1982) have reported that most
of the E-S antigens_ of B.malayi adults were common to those
present in the somatic extracts of adult worms. Antigenic
sharing has also been reported between the somatic extracts
and E-S prodoucts of L.carinii (Ishii 1980) , F_. hepatica
(Klimenko, 1977), A.cantonensis (Techasoponmani and Sirismha,
J980), S.vulgaris (Wynn et_ a^., 198i) and Trichuris muris
(Jenkins and Wakelin, 1977; 1983). Commonness of antigens
between the somatic extracts from adult and microfilariae
and E-S products of female adult S.cervi has also been ^hown
employing immunoelectrophoretic technique (Malhotra e£ al_.,
1987).
Parasitic worms are known to absorb the host oroteins
on their surface to evade the immunologically hostile environ
ment of the host (Smithers et a l . , 1969). The adults of

105
A. ceylanicum worms reside in the submucosal layer of small
intestine and are voracious blood suckers, there is a high
probabili ty of absorption of serum proteins bv the worms
and these proteins would subsequently be released during
in vitro maintenance ot the worms. Hyperimmune rabbit serum
against normal hamster serum (NHS) were prepared to detect
the presence of host serum proteins in E-S products.
CIE of A. ceylanicum E-S products with rabbit anti-normal
hamster serum showed the presence of 6 precipitin peaks
thereby indicating the presence of host proteins in E-S products.
The presence of host proteins in the E-S products of different
parasites have been reported by several workers (Kaushal
et^al^., 1982; Maizels et_ a]_., 1984; Harnett et a^., 1986: Malhotra
et a l . , 1987).
There is substantial evidence indicating the protective
potentials of E-S antigens of some nematode parasites including
hookworms (Thorson, 1951; Katiyar et_ al^., 1972; Govila et_
al^., 1973; Poulain et_ a]_., 1976; Sugane and Oshima, 1983;
Jenkins and Wakelin, 1983; Maizels et_ al^., 1984; Badley e^
al_., 1987a,b; Ey, 1988). In the present study, the E-S anti
gens of A. ceylanicum adults have been found to stimulate a
significant level of protective immunity as the hamsters immu
nised with E-S products showed 75% reduction in worm burden.
Wedrychowicz et_ al^. (1984) have reported 69% protection bv
E-S products from adult N.brasi l iensis . The antigens obtained
from cultures of A.duodenale (Jain et_ al^., 1979), T.earns (Sugane

106
and Oshima, 1983; MaizeLs e; al^., 1984; Badley et al^-. 1987a,b)
and T.muris (Jenkins and Wakelin, 1983) have also been shown
to induce part ial resistence against infection with the respective
paras i tes .
In conclusion, the excretory-secretory (E-S) products
from A.ceylanicum adults could be obtained by short term
in vitro maintenance of adult worms in Ringer's solution which
is much simpler and economical medium. The rabbit anti-
E-S sprurri having high t i te rs ot parasite specific antibodies,
showed the presence of 16 antigens in the E-S products. Twelve
and 7 antigens of E-S products shared antigenic determinants
with adult and larval somatic extracts respect ively. Some
of the antigens of ' E-S products are derived from the host
serum proteins. The protective potential of E-S products
has also been evaluated and the hamsters immunised with
E-S products showed significant level of protection. A logical
extension of these studies would be to characterise the E-S
products on molecular basis in order to identify the protective
antigens.

CHAPTER lY
SUMMARY AND CONCLUSION

107
Ancylostomiasis is one of the major public health problems
in tropical countries. The world incidence of hookworm infect
ions are 1000 million with the death rate of 55 thousand per
year. The currently available anthelmintics are not very
effective. This clearly emphasises the need to intensify
the control measures in hookworm endemic areas . An alternate
approach for the permanent control of the disease would be
to develop immunoprophylactic measures against hookworm
paras i tes . However, the existing knowledge about the molecular
immunology of hookworm infection is s t i l l in i ts infancy.
Hookworm parasi tes evoke functional protective immunity and
some reports are available regarding the protective potential
of live parasit ic stages as well as of somatic extracts and
excretory-secretory products from adult and larval stages,
however, not much is known about the molecules/antigens which
are involved in evoking these protective responses. A basic
knowledge about the total antigenic composition of the para
sites is essential not only for understanding the host parasite
relationships but also for the identification of antigenic ta r
gets of protective immunity. We have, therefore, directed
our studies towards the characterisation of antigens from adult
and larval stages of human hookworm parasi te ; Ancylostoma
ceylanicum, which has earl ier been used for the screening
ot anthelmintic compounds by several workers. TotaJ proteir)
and antigenic composition of adult and larval stages of
A. ceylanicum were studied employing SDS-polyacrylamide gel

108
electrophoresis (SDS-PAGE) and immunoelectrophoretic techniques
using hyperimmune rabbit sera . The levels of parasite specific
antibodies in immune sera were determined by enzyme linked
immunosorbent assay (ELISA). Similar techniques were used
for antigenic analysis of the excretory-secretory products
from adult A.ceylanicum. The protective potential of somatic
extracts from adult and larvae as wellas the excretory-secretory
products from adult A.ceylanicum was also studied by immunising
the naive hamsters with these antigens (somatic extracts from
adult and larvae and excretory-secretory products from adults)
toilowed b \ challenge infection.
The somatic extracts from adult (AcA) and larval stages
(AcL) of A.ceylanicum were separated on SDS-polyacrylamide
gradient gel electrophoresis (SDS-PAGE), which showed the
presence of 47 protein bands in adults and 43 in larvae in
the molecular weight range of 10-170 kD. Most of the protein
bands were found to be common between these two stages
(adult and la rvae) , however, 8 (130, 125, 42, 30, 20, 17,
16.5 and 10 kD) and 4 (170, 160, 153 and 85 kD) protein
bands specific to adult and larval stages respectively were
observed. The quantitation of parasite specific antibodies
in immune rabbit sera was done by enzyme linked immunosorbent
assay and reciprocal antibody t i t e r s of 256,000 and 128, 000
were obtained for rabbit anti-adult and rabbit anti-larval
sera respectively using AcA antigen at optimum concentration
of 0.25 ug/well. Both these sera showed the reciprocal anti-

109
body t i t e r s of 256,000 with AcL at optimum concentration
of 0.50 ug/well.
The antigenic analysis of somatic extracts from adult
and larval stages of A.ceylanicum was initially done by Immuno
electrophoresis ( lEP), which revealed 10-12 precipitin lines
in AcA with rabbi t anti-AcA serum and 6-7 precipitin bands
in AcL with rabbit anti-AcL serum. Further characterisation
of these somatic antigens was carried out employing crossed
Immunoelectrophoresis (CIE) which showed the presence of
34 antigenic components in AcA and 19 antigens in AcL. Out
of 34 AcA antigens, 25 were anodic and 9 cathodic while 17
anodic and 2 cathodic antigens were observed in AcL. In order
to study the cross-reactive or common as well as stage/parasite
specific antigens of A.ceylanicum, crossed Immunoelectrophoresis
employing heterologous sera and crossed line immunoelectro-
phoresis (CLIE) were used. When AcA and AcL were reacted
with heterologous immune rabbit sera in CIE, most of the
antigens were found common or cross-reactive between the
two stages, however, 12 antigens specific to adult and 5 anti
gens specific to larval stage were observed. The subsequent
analysis of CLIE also revealed the same number of common
or cross-reactive as well as stage specific antigens in AcA
and AcL. In order to analyse A.ceylanicum antigens having
cross-reactivi ty with rat hookworm - Nippostrongylus brasi l iensis ,
CIE using immune rabbit sera raised against adult (NbA) and
infective larval (NbL) somatic extracts of N.brasiliensis and

110
CLIE using NbA and NbL in the in te rmedia te ge l , were done.
Most of the antigens (22-26) were found to be AcA spec i f i c ,
only 12 and 8 antigens of AcA were found to be common/cross
r e a c t i v e wi th NbA and NbL r e s p e c t i v e l y . On analysing the
c r o s s - r e a c t i v e or common antigens of AcL with NbL and NbA,
13 common or c ross r e a c t i v e and 6 AcL specif ic antigens were
o b s e r v e d .
Another set of antigens analysed in the p resen t study
i s t he e x c r e t o r y - s e c r e t o r y (E-S) p roduc t s from adult
A. cey lanicum. The condit ions for t h e p repa ra t ion of E-S
p roduc t s were opt imised with r e s p e c t to medium as well as
the incubation t i m e . Out of t h r e e different media; RPMI-1640,
DMEM and R i n g e r ' s solution t r i e d . R inge r ' s solution proved
to be the bes t as the mot i l i ty of t h e worms was maximum
in t h i s medium and t h e worms could be maintained upto 6h
without changing t h e medium. The amount of prote in re leased
was found to be 95±15 ug/100 adults in RPMI-1640 and DMEM
as compared to 125±25 ug/100 adult worms in R inge r ' s solut ion.
Immunoelectrophoret ic ana lys i s using r a b b i t anti-AcA serum
showed the presence of 5-7 p r e c i p i t i n bands in E-S products
p r epa red in t h e s e media . As t h e r e was not any significant
difference in t h e amount of pro te in r e l ea sed as well as the
antigenic p a t t e r n s of E-S p roduc t s p r e p a r e d in t h r e e media .
R inge r ' s solution (containing s a l t s and glucose) was chosen
to obtain t h e E-S p roduc t s from A»cey lanicum adu l t s because
t h i s i s much s imple r and more economical in comparison to
o the r two media (RPMI-1640 and DMEM).

I l l
A polyvalent hyperimmune rabbit serum was raised dgainst
A.ceylanicum adult E-S products and the t i ter of parasite
specific antibodies in immune rabbi t sera was determined
by enzyme linked immunosorbent assay using optimum concentrat
ion of 0.50 ug E-S antigen/well. A reciprocal antibody ti ter
of 256,000 vras obtained with rabbit anti-E-S serum. The
E-S antigens were also tested with the immune rabbit sera
raised against somatic extracts of A.ceylanicum in ELISA and
a t i ter value of 128,000 was obtained for both rabbit anti-
adult and rabbit anti-larval sera . Crossed Immunoelectropho
resis (CIE) using rabbit anti-E-S serum revealed 16 antigens
m A.ceylanicum E-S products, out of which 13 were anodic
and 3 moved towards the cathode. E-S products have also
been found to share antigenic determinants with the somatic
extracts from adult and larval stages. lEP analysis of E-S
products showed the presence of 6 and 4 precipitin bands
with rabbit anti-AcA and rabbit anti-AcL sera respectively.
On further analysing the E-S products by CIE using rabbit
sera raised against somatic extracts 12 and 7 antigens of
E-S products were found to share antigenic determinants with
AcA and AcL respect ively. Tandem crossed Immunoelectropho
resis (TCIE) of E-S products with AcA and AcL also revealed
almost the same number of common/cross-reactive antigens
between the E-S products and the somatic ext rac ts . The pre
sence of host serum proteins in the E-S products was also
analysed by CIE using rabbit serum raised against normaJ

112
A polyvalent hyperimmune rabbit serum was raised against
A.ceylanicum adult E-S products and the t i ter of parasite
specific antibodies in immune rabbi t sera was determined
by enzyme linked immunosorbent assay using optimum concentrat
ion of 0.50 ug E-S antigen/well. A reciprocal antibody t i ter
of 256,000 v/as obtained with rabbit anti-E-S serum. The
E-S antigens were also tested with the immune rabbit sera
raised against somatic extracts of A.ceylanicum in ELISA and
a t i te r value of 128,000 was obtained for both rabbit anti-
adult and rabbit anti- larval sera . Crossed Immunoelectropho
res is (CIE) using rabbit anti-E-S serum revealed 16 antigens
in A.ceylanicum E-S products, out of which 13 were anodic
and 3 moved towards the cathode. E-S products have also
been found to share antigenic determinants with the somatic
extracts from adult and larval stages. lEP analysis of E-S
products showed the presence of 6 and 4 precipitin bands
with rabbi t anti-AcA and rabbit anti-AcL sera respectively.
On further analysing the E-S products by CIE using rabbit
sera raised against somatic extracts 12 and 7 antigens of
E-S products were found to share antigenic determinants with
AcA and AcL respect ively. Tandem crossed Immunoelectropho
resis (TCIE) of E-S products with AcA and AcL also revealed
almost the same number of common/cross-reactive antigens
between the E-S products and the somatic ext rac ts . The pre
sence of host serum proteins in the E-S products was also
analysed by CIE using rabbit serum raised against normal

113
hamster serum, and 6 antigens in the E-S products appeared
to be of host origin.
The protective potential of somatic extracts from adult
and larval stages as well as excretory-secretory products
trom ddult A.ceylanlcum was tested by immunizing the hamsters
with these antigens and evaluating the immunized hamsters
for protective immunity. The sera obtained from immunized
hamsters, before challenge, showed significant levels of para
site specific antibodies. The immunodiffusion and immuno-
electrophoretic analysis revealed the presence of precipitin
antibodies in immune hamster sera and 7 precipitin bands
in adult somatic extracts and 4 in both larval somatic extracts
and E-S products. -The hamsters immunized with larval somatic
extracts showed maximum protection (93% reduction in worm
burden) while 75% and 63% protection was observed in hamsters
immunized with excretory-secretory products and adult somatic
extracts respect ively.
Therefore, in the present study we have been able to
characterise the protein and antigenic components of adult
and larval stages of A.ceylanicum using hyperimmune rabbit
sera having high t i t e r s of parasite specific antibodies and
certain Diochemicai and Immunochemical techniques. The common
or cross-reactive antigens between the two life stages of
A.ceylanicum and also between the two hookworm parasites
(A.ceylanicum and N.brasiliensis) as well as stage and parasite

114
specific antigens have also been identified. We have been
able to prepare the excretory-secretory products of A.ceylanicum
adults using Ringer's solution which is much simpler and
a more economical medium. Antigenic characterisation of E-S
products revealed less complex nature of these products as
compared to the adult somatic ext rac ts . The protective poten
t ials of the somatic extracts from adult and larval stages
and also of E-S products from adult A. ceylanicum was also
studied. The larval somatic ext rac ts , though contain less
number of antigens, showed maximum protection followed by
excretory-secretory products and adult somatic extracts .
These studies provide basic information about the antigenic
compositions of somatic extracts and excretory-secretory pro
ducts of A. ceylanicum as well as the comparative efficacy
of these antigens in inducing protective immunity against this
paras i te . However, further studies on molecular characterisa
tion of these antigens are certainly required for identifying
the antigenic targets of protective immunity.

BIBLIOGRAPHY

115
Adams, J . H . , Monrov , F . G . , Eas t , I . J . and Dobson, C. (1987). Immunol. Cell Bio l . 65, 393.
Adams, J . H . , Eas t , I . J . , Monroy, F .G . and Dobson, C. (1988). Int . J . P a r a s i t o l . 18, 999.
All , M.M., Hegazi, M.M., Abdel-Magied, S .A. , El imekdv, S.A. and El Ganayani, G.A. (1986) . J . Egyp t . Soc. P a r a s i t o l . 16, 303.
Au, A .C .S . and Ko, R.C. (1979). Z. P a r a s i t e n k d e , 59 161.
Axelsen, N .H . , Krol l , J . and Weeke, B. (1973) . Scand. J . Immunol. 2 (Suppl . 1 ) .
Badley , J . E . , Gr ieve , R . B . , Bowman, D.D. , Glickman, L .T . and Rockey, J . H . (1987a) . J . P a r a s i t o l . 73, 593.
Badley , J . E . , Gr i eve , R . B . , Rockey, J . H . and Glickman, L . T . (1987b) . Pa ra s i t e Immunol. 9, 133.
Bany, J . , Bankowski , A. and Kazinuerezuk, J . (1976) . P roc . 4th In t . Conf. T r i c h . , 26-28th Aug. Univ. P r e s s New Eng . , 95 .
Bal l , P . A . J , and B a r t l e t t , A. (1969) . T r a n s . Roy. Soc. T r o p . Med. Hyg. 63, 362.
Befus, D. and Bienenstock, J . (1982) . P r o r r e s s in Allergy 3 1 . 76.
Behnke, J . M . , Pau l , V. and Ra jaseka r i ah , G.R. (1986) . T rans . Roy. Soc. T r o p . Med. Hyg. 80, 146.
Bhopale , M.K., Menon, S. and Kamath, V.R. (1981) . Bull . Haff. In s t t . 9, 15.
Bohn, A. and Konig, W. (1985) . Z. P a r a s i t e n k d . 71 , 663.
Bout, D . , Conlien, Y. , Dessaint , J . P . and Capron, A. (1977). INSERM Symposium 72, 7 1 .
Br ind ley , P . J . , Gam, A .A. , P e a r c e , E . J . , Po index te r , R.W. and Neva, F .A. (1988) . Mol. Biochem. P a r a s i t o l . , 28, 171.
Brown, P . J . , Poulain, J . and P e r y , P . (1981a) Vet. Immunol. Immunopathol. 2 , 343.
Brown, P . J . , Poulain, J . and P e r y , P . (1981b). Vet. Immunol. Immunopathol. 2, 475.

116
Campbel l , C.H. (1955) . J . P a r a s i t o l . 4 1 , 483.
Capo, v . , S i l b e r s t e i n , D. and Despommier, D.D. (1986). J . P a r a s i t o l . 72, 931 .
C a r r , A. and P r i t c h a r d , D . I . (1986) . Mol. Biochem. Pa ra s i t o l . 19, 251.
C a r r , A. and P r i t c h a r d , D . I . (1987) . Pa ra s i t e Immunol. 9, 219.
C a r r o l l , S.M. and Grove, D . I . (1984) . J . Inf. Dis . 150, 284.
C a r r o l l , S.M. and Grove, D . I . (1985) . E x p . P a r a s i t o l . 60, 263.
C a r r o l l , S.M. and Grove, D . I . (1986) . T r a n s . Roy. Soc. T r o p . Med. Hyg. 80, 406.
Cavalan, S . , Rhalem, A. , Luffau, G. and P e r y , P . (1985) . Ann. Rech. Vet. 16, 305.
Chipman, P . B . (1957) . J . P a r a s i t o l . 43 , 593.
Choi , W.Y. and Lee', O.R. (1979) . Kor. J . P a r a s i t o l . 17, 147.
Clegg, J . A. and Smith, N.A. (1978) . Adv. P a r a s i t o l . 16, 165.
Co r r ea -Ol ive i r a , R . , Dusse, L . M . S . , Viana, I . R . C . , Col ley, D.G. , Garva lho , O.S. and Gazzinel l i , G. (1988). Am. J , T r o p . Med. Hyg. 38, 348.
Cromptom, D.W.T. (1989) . Paras i to logy Today, 5, 1.
de Azevedo, J . F . , Vie i ra , R.A. and Rombert , P . C . (1971). Anais da Escola Nacional de Saude Publ ica e de Medicina T r o p i c a l . 5, 115.
de Hurtado, I . and L a y r i s s e , M. (1968) . Am. J . T rop . Med. Hyg. 17, 72.
Desowitz, R .S . (1962) . Ann. T r o p . Med. P a r a s i t . 51,161.
Desowitz, R . S . (1967) . Seminar on Helminthiasis and Eosinop h i l i c Meningi t is , Noumea, New Caledonia , Abstracted in Helminthological A b s t r a c t s , 38, 2959.
Despommier, D.D. and Muller , M. (1976) . J . P a r a s i t o l . 62, 775.

117
Dobson, C. (1982) . Int . J . P a r a s i t o l . 12, 207.
Dobson, C. and Owen, M.E. (1978) . In t . J . P a r a s i t o l . 8, 359.
Eas t , I . J . , Washington, E . A . , Br ind ley , P . J . , Monroy, F.G. and Scott-Young, N. (1988) . E x p . P a r a s i t o l . 66, 7.
Ef remove, E . E . and Ermolin, G.A. (1980) . P roc . 5th Int . Cong. T r i c h . 1st to 5th Sep t . 47.
E r i k s e n , L. (1982) . Nord. Vet. Med. 34, 188.
Ey , P . L . (1988a) . E x p . P a r a s i t o l . 67, 210.
Ey , P . L . (1988b) . E x p . P a r a s i t o l . 65, 232.
Koster , A.O. (1935) . Am. J . Hyg. 22, 65 .
Gamble, H.R. (1985) . J . P a r a s i t o l . 71, 680.
Ganguly, N.K. , Mahajan, R . C . , Sehgal , R . , She t ty , P . and Di lawar i , J . B . (1988) . J . Cl in . Microbiol . 26, 739.
Gautum, O . P . , Ahmed, M.U. and B a n ^ r j e e , D .P . (1986). 10, 283.
Got ts te in , B . , Tsang, V.C.W. and Schantz , P.M. (1986). Am. J . T r o p , Med. Hyg. 35, 308.
Govila , P . , Kat iyar , J . C . and Sen, A.B. (1973) . Ind. J . E x p . Biol . 1 1 , . 529.
G r a b e r , P . et and Will iams, C.A. (1953) . Biochem. B iophys . Acta 10, 193.
Gupta, S. and Ka t iya r , J . C . (1985) . Helminthologia 22, 181.
Harnet t , W., Meghji, M., Worms, M.J . and Pa rkhouse , R.M.E. (1986) . P a r a s i t o l . 93 , 317.
Her r i ck , C.A. (1928) . Am. J . Hyg. 8a, 125.
Hopkins, C .A. , Subramaniam, G. and Sta l land , H. (1972). P a r a s i t o l . , 64, 4 0 1 .
Hopkins , C.A. and Zajac, A. (1976) . P a r a s i t o l . 73 , 73 .
Hotez, P . J . , Le Trang N. and Cerami , A. (1987) . Paras i tology Today, 3, 247.
Hurley, J . C , Day, K . P . and Mi tche l l , G .F . (1980) . Aust. J . E x p . Biol . Med. Sc i . 58, 231 .

118
I s h i i , A. (1970) . J a p . J . E x p . Med. 40, 39.
I s h i z a k i , T . , Kutsumi, H. , Kumada, M., Komiya, Y., Araki , H . , Takayama, H. , Okada, K. and Nozaki, S. (1961). J a p . J . P a r a s i t o l . 10, 211 .
J a in , P . . Sawhnery , S. and Vinayak, V.K. (1979) . Bull . PGI Med. Educ . and Res . 13, 131.
J a r r e t t , E . E . E . and Urquaha r t , G.M. (1971) . Int . Rev. Trop . Med. 4, 53 .
J a r r e t t , E . E . E . and Bazin, H. (1977) . Cl in . E x p . Immunol. 30, 330.
J a r r e t t , E . , Mackenzie, S. and Bennich, H. (1980) . Nature 283, 302.
J ayewerdene , G. and Vijayaratnam, Y. (1968) . Ceylon. J . Med. Sc i . 17, 17.
J e n k i n s , N.S . and Wakelin, D. (1977) . P a r a s i t o l . 74, 153.
J e n k i n s , N.S . and Wakelin, D. (1983) . P a r a s i t o l . 86, 73 .
Jones , V . E . , E d w a r d s , A . J . and Ogi lv ie , B.M. (1970) . Immunol. 18, 621 .
Jones , C .E . and Rubin, R. (1974). E x p . P a r a s i t o l . 35, 434.
Kamath, V.R. , Bhopale , M.K. and Bhide , M.B. (1985). Hel-minthologia 22, 203.
Kassa i , T. (1982) . Handbook of N . b r a s i l i e n s i s (nematode) . Common Wealth Agricul tural Bureaux, Akademiai Kiado, Budapes t .
Kassa i , T . , T a k a s , C. and Red, P . (1987) . J . P a r a s i t o l . 73, 345.
Kat iyar , J . C , Govila , P . and Sen, A.B. (1972) . Ind. J . E x p . Biol . 10, 201 .
Kat iyar , J . C , Visen, P . K . S . , Misra , A, Gupta, S. and Bhadur i , A . P . (1984) . Acta Trop ica , 4 1 , 279.
Ka t iya r , J . C , Visen, P . K . S . , Gupta, S . , Sen, A . B . , Dubey, S.K. and Sharma, S. (1982) . E x p e r i e n t i a , 38, 457.
Kaushal , N .A. , Hussain , R . , Nash, T . E . and Ottesen, E.A. (1982) . J . Immunol. 129, 338.
Kaushal , N .A . , Hussain, R. and Ot tesen, E.A. (1984). Clin. E x p . Immunol. 56, 567.

119
Kaushal , N .A . , Kaushal , D.C. and Ghatak, S. (1987). Immunol. Inves t . 16, 139.
Kazura, J .W. and Grove, D . I . (1978) . Nature, 274, 588.
Kazura, J .W. and Aikawa, M. (1980) . J . Immunol. 124, 355.
Keeling, J . E . D . (1960) . Ann. T r o p . Med. P a r a s i t o l . 54, 182.
Kendr ick , J . F . (1934) . Am. J . T r o p . Med. Hyg. 15, 363.
Kennedy, M.W., Maizels , R ,M. , Meghj i ,M. , Young, L . , Qureshi , F . and Smith, H.V. (1987a) . P a r a s i t e Immunol. 9, 407.
Kennedy, M.W., Quresh i , P . , Haswel l -E lk ins , M. and E lk ins , D.B. (1987b) . Cl in . E x p . Immunol. 67, 20.
Keymer, A. and Bundy, D. (1989) . Nature, 337, 114.
Klimenko, V.V. (1980) . Med. P a r a z i t o l . P a r a z i t . Bolezni, 49, 37.
Kornishina , M.V. (1977) . im N.E . Baumana, 126 ^ 9 5 ,
Kum, W.W.S. and Ko, R .C. (1985) . Z. P a r a s i t e n k d e , 71, 789.
Laemmli, U.K. (1970) . Nature, 227, 680.
Lee, T .D .G . , Gren ic i s , R.K. and Wakelin, D. (1982) . P a r a s i t o l . 84, 381 .
Le id , R.W. and Will iams, J . F . (1974) . Immunol. 27, 195.
Lewis , E . A . , Salimonu, L . S . and Osunkoga, B.O. (1978). Af. J . Med. Sc i . 17, 197.
Lobel , A .O . , Kagan, I .G. and Kaiser , R .L . (1968) . Am. J . Ep idemio l . 87, 58.
Lobos, E. and Weiss , N. (1985) . P a r a s i t . Immunol. 7, 333.
Love, R . J . , Kel ly , J . D . and Dineen, J . K . (1974) . Int . J . P a r a s i t . 4, 183.
Lowry, O.H. , Rosebrough, N .Y. , F a r r , A.L. and Randall , R . J . (1951) . J . Biol . Chem. 193, 265.
Lucius , R. , Raute rberg , E.W. and Diesfeld, H . J . (1983) . Trop . Med. P a r a s i t o l . 34, 133.
Mackenzie, C D . , P res ton , P .M. and Ogi lv ie , B.M. (1978) . Nature, 276, 826.

120
Magaudda-Borzi , L. and Penn i s i , L. (1958) . Rivista I tal iana d ' l g i ene 18, 3 .
Maizels , R.M. and Ogi lv ie , B.M. (1980) . Abstract of Session 8A. 4th Internat ional Congress of Immunology.
Maizels , R .M. , P h i l l i p , M. and Ogi lv ie , B.M. (1982) . Immunol. Rev. 61 , 109.
Maizels , R .M. , Meghi]i , M. and Ogi lv ie , B.M. (1983) . Immunol. 48, 107.
Maizels , R .M. , Par tono , F . , Oemijat i , S . , Denham, D.A. and Ogi lv ie , B.M. (1983) . P a r a s i t o l . 87, 249.
Maizels , R .M. , DeSavigny, D and Ogi lv ie , B.M. (1984) . Pa ra s i t e Immunol., 6, 23 .
Maizels , R .M. , P h i l l i p , M., Dasgupta, A. and Pa r ton i , F . (1984) . Pa ra s i t e Immunol. 6, 185.
Maizels , R . , Burke , J . , Sutanto, I . , Purnomo and Partono, F . (1986) . Mol. Biochem. P a r a s i t o l . 19, 27.
Malhotra , A. , Kaushal , N.A. and Ghatak , S. (1986) . Immunol. Inves t . 15, 505._
Malhotra , A. (1988) . P h . D . t h e s i s . "Immunochemical s tudies on bovine f i l a r i a l p a r a s i t e - Setar ia c e r v i . Univers i ty of Lucknow, Lucknow.
Malhotra , A . , Kaushal , N .A. , Kaushal , D.C. and Ghatak, S. (1987) . T r o p . Med. P a r a s i t . 38, 106.
Manas-Almendros, I . , Campos-Bueno, M., Lozano-Maldonado, J . and Gonzalez-Castro , J . (1985) . Revista Iber ica de P a r a s i t o l . 45 , 159.
Maxwell , C , Hussain, R . , Nutman, T . B . , Po index te r , R.W., L i t t l e , M.D. , Schad, G.A. and Ottesen, E.A. (1987). Am. J . T r o p . Med. Hyg. 37, 126.
McCoy, O.R. (1931) . Am. J . Hyg. 14, 268.
Menon, S. and Bhopale , M.K. (1985) . J . Helminthol . 59, 287,
Mi l le r , T.A. (1965) . J . P a r a s i t o l . 51 , 200.
Mi l le r , T.A. (1971) . Adv. P a r a s i t o l . 9, 153.
Mi l le r , T.A. (1979) . Adv. Pa ra s i t o l 17, 315.
Mimori, T , , Tanaka, M. and Tada , T . (1987) . E x p . P a r a s i t o l . 64, 342.

121
Misra , A. , Visen, P . K . S . and Ka t iya r , J . C . (1981) . J . Hel-min tho l . 55, 273.
Milner, A . , Beall, J . A. and Orwat , A. (1987) . Paras i te Immunol. , 9, 615.
Mi tche l l , G . F . and C r u i s s e , K.M. (1984) . Aus. J . E x p . Biol . Med. Sc i . 62, 523.
Mi tche l l , G .F . and Munaz, J . J . (1983) . Aus. J . E x p . Biol . Med. Sc i . 6 1 , 425.
Monroy, F . G . , B r i n d l e y , P . J . and Dobson, C. (1985) . Aus. J . E x p . Bio l . Med. Sc i . 63 , 423.
Monroy, F .G . and Dobson, C. (1986) . Aust. J . E x p . Biol . Med. Sc i . 64, 335.
Monroy, F .G . and Dobson, C. (1987) . Immunol. Cell Biol . 65, 223.
Murray, M., Robinson, P . B . , Gierson, C. and Crawford, R.A. (1979) . Acta Tropica 36, 297.
Neilson, J . T . M . (1975) . J . P a r a s i t o l . 6 1 , 785.
Neilson, J . T . M . (197'8). E x p . P a r a s i t o l . 44, 225.
Newport, G. , Cu lppe r , J . and Agabian, N. (1988) . Paras i tology Today, 4, 306.
Ngu, J . L . , Ndumbe, P . M . , T i t an j i , V. and Rose, L. (1981). P a r a s i t o l . 3 1 , 165.
Noda, M. (1953) . Osaka City Med. Cent re , 2, 203.
Novoselska, L . , Zhelev , Z.H. and Daskalova, M. (1981). Scientific Works of the Higher Medical Ins t i tu te P leven , 1, 34.
Ogi lv ie , B.M. (1967) . Immunol. 12, 113.
Ogi lv ie , B.M. and Jones , V .E . (1973) . P rog . Allergy 17, 93 .
Ogi lv ie , B.M. and Love, R . J . (1974) . T r a n s . Rev. 19, 147.
Ogi lv ie , , B .M. , B a r t l e t t , A . , Godfrey, R . C . , Turbin , J . A . , Worms, M.J . and Yeater , R.A. (1978) . T rans . Roy. Soc. T r o p . Med. Hyg. 72, 66.
Ogi lv ie , B.M. and P h i l l i p , M. (1980) . Mol. Cells and P a r a s i t e s in Immunol. (Academic P r e s s , NY), 163.

122
Orr , T . S . C . and B l a i r , A .M.J .N. (1969) . Lite Sex. 8, 1073.
Otto, G .F . and Ker r , K.B. (1939) . Am. J . Hyg. 29, 2S.
Pa rkhouse , R.M.E. and C l a r k , N.W.T. (1983) . Mol. Biochem. P a r a s i t o l . 9, 319.
Pa rkhouse , R.M.E. and O r t e g g a - P i e r r e s , G. (1984) . P a r a s i t o l . 88, 623.
P h i l l i p , M., Pa rkhouse , R.M.E. and Ogi lv ie , B.N. (1980). P r o c . 5th In t e rna t t . Conf. T r i c h i n e l l o s i s . 1-5 Sept . Netherl a n d s .
Poulain, J . , Luffau, G. and PerVi P- (1976a) . Ann. Immunol. 127C, 209.
Poulain, J . , Luffau, G. and P e r y , P . (1976b) . Ann. Immunol. 127C, 215.
P r a s a d , B.G. and Mathur, G.B. (1962) . Ind . J . Med. Res. 50, 1.
P r i t c h a r d , D . I . , Wil l iams, D . J . L . , Behnke, J .M. and Lee, T.D.G. (1983) . Immunol. 49, 353.
P r i t c h a r d , D . I . , Ali', N.M.H. and Behnke, J .M. (1984) . Immuno l . 51 , 633.
P r i t c h a r d , D . I . , Crawford, C . R . , Duce, I .R . and Behnke, J .M. (1985) . P a r a s i t e Immunol. 7, 575.
P r i t c h a r d , D . I . , Behnke, J . M . , C a r r , A. and Wells , C. (1986). Pa ra s i t e Immunol. 8, 359.
P r i t c h a r d , D . I . , McKean, P .G . and Rogan, M.T. (1988a) . Mol. Biochem. P a r a s i t o l . 28, 275.
P r i t c h a r d , D . I . , McKean, P .G . and Rogan, M.T. (1988b) . Paras i to logy Today, 4 , 239.
P rowse , S . J . , Mi tche l l , G . F . , Ey , P . L . and Jenk in , C.R. (1978) . Aus. J . E x p . Bio l . Med. Sc i . 56, 561.
P r u s s e , A. , Diefeld, H . J . and Vollmer, S. (1982) . J . Hel-min tho l . 46, 357.
Raghavan, P . (1967) . J . Pos tg rad . Med. 13, 187.
Rao, Y .V.B.G. , Mehta, K., Subrahamanyam, B. and Rao, C.K. (1980) . Ind. J . Med, Res . 72, 47.
Ray, D.K. , Bhopale , K.K. and S r i v a s t a v a , V.B. (1972). J . Helminthol . 46, 357.

123
Roach, T . I . A . , Wakelin, D. , E l se , K . J . and Bundy, D.A.P, (1988) . Pa ra s i t e Immunol. 10, 279.
Rombert , P . , V ie i ra , V. and de Azevedo, J . F . (1967). 0 Medico Lisbon 803, 3 .
Rotmans, J . P . , Vander Voort, M . J . , Looze, M., Mooize, G.W. and n e e d i e r , A.M. (1981) . E x p . P a r a s i t o l . 52, 171.
Sawada, P . , Sano, M. and Ueno, T. (1954a) . Gumma J . Med. S c i . , 3 , 2 1 .
Sawada, T . , Suzuki , I . , Oka, T. and Sano, M. (1954b) . Gunma T. Med. Sc i . 3 , 29.
Sawada, T . , Kono, M., Sato, S. and Oikawa, S. (1961). J a p . J . P a r a s i t o l . 10, 171.
See s ee , F . M . , Wescott, R .B . and Gorham, J . R . (1976) . Exp . P a r a s i t o l . 39, 214.
Shih-Huei , G. , Chen-Huang, S . , Shao-Hsun, G. and Hsi-Cheng, C. (1959) . Chinese Med. J . , 78, 267.
S i l b e r s t e i n , D.S. and Despommier, D.D. (1984) . J . Immunol. 132, 888.
S i l b e r s t e i n , D.S. and Despommier, D.D. (1985) . J . P a r a s i t o l . 71, 516.
Singh, M. (1965) . Med. J . Malaya 20, 167.
Singh, N. (1990) . P h . D . T h e s i s . ' immunochemical s tudies on malar ia l p a r a s i t e s I' Univers i ty of Kanpur, Kanpur.
Singh, D . S . , Das, A.K. , Bha t ia , V .N. , B i sh t , D.B. and Kumar, V, (1978) . Ind . J . P a r a s i t o l . 2, 5 1 .
S insk i , E. and Holmes, P .H . (1977) . E x p . P a r a s i t o l . 43 , 382.
S insk i , E. and Holmes, P . H . (1978) . J . P a r a s i t o l . 64, 189.
Smith, N.C. (1989) . P a r a s t i o l . Res . 76, 423.
Smith, N.C. and Bryant , C. (1986) . In t . J . P a r a s i t o l . 16, 617.
Smi the r s , S .R. , T e r r y , R . J . and Hockley, D . J . (1969) . P roc . Roy. Soc. 171, 483.
Sugane, K. and Oshima, T. (1983) . Immunol. 50, 113.
Sugane, K., Howell, M.J . and Nicholas , W.L. (1985) . J . Hel-min tho l . 59, 147.

124
Tandon, A . , Saxena, R . P . , Bhat ia , B . , Saxena, K.C. and S r i v a s t a v a , V.K. (1981) . Ind . J . Med. Res, 73 , ( S u o p l . ) , 93 .
Tandon, A. , S r i v a s t a v a , A.K. , Saxena, R . P . , Saxena, R.K. and Saxena, K.C. (1983) . T r a n s . R. Soc. T rop . Med. Hyg. , 77, 439.
Tay lo r , M.M. and Turton, J . A. (1976) . Trop Med. P a r a s i t o l . 27, 89.
Techasoponmani, R.O. and S i r i s i n h a , S. (1980) . P a r a s i t o l . 80, 457.
Thorson, R .E . (1951) . J . P a r a s i t o l . 3 7 ( S u p p l . ) , 18.
Thorson, R .E . (1956) . J . P a r a s i t o l . 42, 501.
Turner , K . J . , F i s h e r , E .H. and McWilliam, A.S. (1980). Aust. J . E x p . Biol . Med. Sc i . 58, 249.
Urban, T . F . and Romanowski, R.D. (1985) . E x p . P a r a s i t o l . 60, 245.
Urquhar t , G.M., Mulligan, W., Ead i e , R.M. and Jennings, F.W. (1965) . ExD. P a r a s i t o l . 17, 210.
Vol ler , A . , B idwel l , D . , . Huld, G. and Engval l , E. (1974). Bul l . WHO 51 , 209.
Wakelin, D. and Wilson, M.M. (1979) . E x p . P a r a s i t o l . 48, 305.
Wange, F . , Yang, G. and Wang., X. (1980) . Cl in . Med. J . 93, 857.
Wedrychowicz, H. , Maclean, J .M. and Holmes, P .H. (1983a), Pa ra s i t e Immunol. 5, 277.
Wedrychowicz, H. , Maclean, J .M. and Holmes, P .H . (1983b) . T r o p . Med. P a r a s i t o l . 34, 207.
Wedrychowicz, H . , Maclean, J .M. and Holmes, P .H . (1984a). Pa ra s i to logy , 89, 145.
Wedrychowicz, H. , Maclean, J .M. and Holmes, P .H. (1984b) . E x p . P a r a s i t o l . 57, 25 .
Wei, P .H. and Kuo, N.K. (1958) . Cl in . Med. J . 76, 556.
Wil l iams, J . C . (1970) . E x p . P a r a s i t o l . 28, 226.
Wynne, E . , Slocombe, J . O . D . and Wilkie , B.N. (1981). Can. J . Comp. Med. 45, 259.

125
Yamanaka, M. (1960) . Bull . Osaka Med. Sch. 6, 25 .
Yoshida, Y. , Kondo, K., Okada, S . , Kurimoto, H. , Oda, K., Shimada, Y . , Shiota , J . , I k a i , T. and Kizaki , K. (1977). J a p . J . P a r a s i t o l . 26, 168.
Yunus, M., Zaheer , M. and Sinha, S.N. (1977) . J . Ind. Med. A s s o c , 69, 241.
Zayatas , R.G. (1978) . Biol . Nauki. 5, 94.


















