
Copeia, 1996(4), pp. 927-937 - southeastern.edu
12
Ultrastructure of the Spermathecae of Necturus beyeri (Amphibia: Proteidae) in Relation to Its Breeding Season Author(s): David M. Sever and Henry L. Bart, Jr. Source: Copeia, Vol. 1996, No. 4 (Dec. 27, 1996), pp. 927-937 Published by: American Society of Ichthyologists and Herpetologists Stable URL: http://www.jstor.org/stable/1447655 . Accessed: 27/12/2010 15:11 Your use of the JSTOR archive indicates your acceptance of JSTOR's Terms and Conditions of Use, available at . http://www.jstor.org/page/info/about/policies/terms.jsp. JSTOR's Terms and Conditions of Use provides, in part, that unless you have obtained prior permission, you may not download an entire issue of a journal or multiple copies of articles, and you may use content in the JSTOR archive only for your personal, non-commercial use. Please contact the publisher regarding any further use of this work. Publisher contact information may be obtained at . http://www.jstor.org/action/showPublisher?publisherCode=asih. . Each copy of any part of a JSTOR transmission must contain the same copyright notice that appears on the screen or printed page of such transmission. JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range of content in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new forms of scholarship. For more information about JSTOR, please contact [email protected]. American Society of Ichthyologists and Herpetologists is collaborating with JSTOR to digitize, preserve and extend access to Copeia. http://www.jstor.org
Transcript of Copeia, 1996(4), pp. 927-937 - southeastern.edu

Ultrastructure of the Spermathecae of Necturus beyeri (Amphibia: Proteidae) in Relation toIts Breeding SeasonAuthor(s): David M. Sever and Henry L. Bart, Jr.Source: Copeia, Vol. 1996, No. 4 (Dec. 27, 1996), pp. 927-937Published by: American Society of Ichthyologists and HerpetologistsStable URL: http://www.jstor.org/stable/1447655 .Accessed: 27/12/2010 15:11
Your use of the JSTOR archive indicates your acceptance of JSTOR's Terms and Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp. JSTOR's Terms and Conditions of Use provides, in part, that unlessyou have obtained prior permission, you may not download an entire issue of a journal or multiple copies of articles, and youmay use content in the JSTOR archive only for your personal, non-commercial use.
Please contact the publisher regarding any further use of this work. Publisher contact information may be obtained at .http://www.jstor.org/action/showPublisher?publisherCode=asih. .
Each copy of any part of a JSTOR transmission must contain the same copyright notice that appears on the screen or printedpage of such transmission.
JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
American Society of Ichthyologists and Herpetologists is collaborating with JSTOR to digitize, preserve andextend access to Copeia.
http://www.jstor.org

Copeia, 1996(4), pp. 927-937
Ultrastructure of the Spermathecae of Necturus beyeri (Amphibia: Proteidae) in Relation to Its Breeding Season
DAVID M. SEVER AND HENRY L. BART JR.
Sperm occurred in the spermathecae of all female Necturus beyeri collected in a Louisiana stream from Dec. through May, including those taken from nests after oviposition in May. The spermathecae possessed two types of epithelial cells: dark cells, with secretory vacuoles 1-2 jm dia; and light cells, with secretory vacuoles 0.5-1.0 im dia. The vacuoles of both types of cells contained glyco- proteins that were secreted into the lumen and bathed sperm during storage. The vacuoles were most abundant early in the breeding season, and light cells in individuals examined after oviposition were completely depleted whereas dark cells retained vacuoles only along the luminal border. If the secretions help maintain sperm viability, the depletion of secretory product could limit the
storage of remnant sperm during the summer and fall months. However, no evidence of sperm degradation was found, and the fate of sperm remaining in the spermathecae following fertilization is unknown. Necturus beyeri can store
sperm for at least six months prior to oviposition.
THE Proteidae consists of five species of branchiated, aquatic salamanders in the
genus Necturus from eastern North America and Proteus anguinus from the Adriatic coast of northeastern Italy and Croatia (Frost, 1985). Like other salamanders in the suborder Sala- mandroidea, cloacal glands in males produce spermatophores and cloacal glands, called sper- mathecae, in females store sperm (Sever, 1991 a, 1992b). The actual transfer of a spermatophore and fertilization has never been observed in the Proteidae. Salthe (1967) suggested that sper- matophore transfer in proteids is via cloacal ap- position. Eggs apparently are fertilized by re- lease of sperm from the spermathecae as the eggs pass through the cloaca (Jordan, 1893; Boisseau and Joly, 1975).
Ultrastructure of sperm storage in the the spermathecae of female salamanders has been described in the Ambystomatidae (Sever and Kloepfer, 1993; Sever, 1995; Sever et al., 1995), Plethodontidae (Pool and Hoage, 1973; Davitt and Larsen, 1988a; Sever and Brunette, 1993), and Salamandridae (Dent, 1970; Brizzi et al., 1995; Sever et al., 1996), but no reports exist for the Proteidae. In this paper, we report on aspects of female sperm storage during the breeding season of Necturus beyeri in a popula- tion in which certain other aspects of the re- production were studied by Shoop (1965), and we describe the cytology of the spermathecae of N. beyeri during the breeding season.
MATERIALS AND METHODS
This study was based on 17 specimens of N. beyeri collected 13 Dec. 1993 to 8 Jan. 1995
from Talisheek Creek between the Gulf, Mo- bile, and Ohio Railroad crossing at Talisheek and the LA Hwy 41 crossing, Saint Tammany Parish, Louisiana (Table 1). Specimens collect- ed in May were postoviposition females re- moved from nests. Numbers of embryos in each nest at the time of guardian removal are given in Table 1. We collected other specimens with modified Thiel-Turkay traps baited with earth- worms (Manning, 1986). We sacrificed these specimens before oviposition.
We sacrificed specimens 0-2 days after col- lection by immersion in 10% MS-222. We ex- cised cloacal regions and preserved them in a 1:1 solution of 2.5% glutaraldehyde in Millon- ig's phosphate buffer at pH 7.4 and 3.7% form- aldehyde buffered to pH 7.2 with monobasic and dibasic phosphate (NBF). We preserved car- casses in NBF and cataloged them into the Tu- lane Museum of Natural History (TU) herpe- tology collection (Table 1).
After fixing cloacal tissue in glutaraldehyde: formalin, we subsequently prepared it by par- affin infiltration for light microscopy (LM), or for embedding in epoxy resin for semithin (LM) or ultrathin sections for transmission electron microscopy (TEM). For the paraffin method, we rinsed the tissue in water after fixation, dehy- drated it in ethanol, and cleared it in Histosol (National Diagnostics, Inc., Manville, NJ) prior to embedment in paraffin. We cut sagittal sec- tions (10 ,Lm) with a rotary microtome, affixed them to albuminized slides, and stained alter- nate slides with hematoxylin-eosin (HE) for gen- eral cytology or alcian blue at pH 2.5 (AB) for glycosaminoglycans with a counterstain of pe- riodic acid and Schiffs reagent (PAS) for "neu-
? 1996 by the American Society of Ichthyologists and Herpetologists
COPEIA, 1996, NO. 4
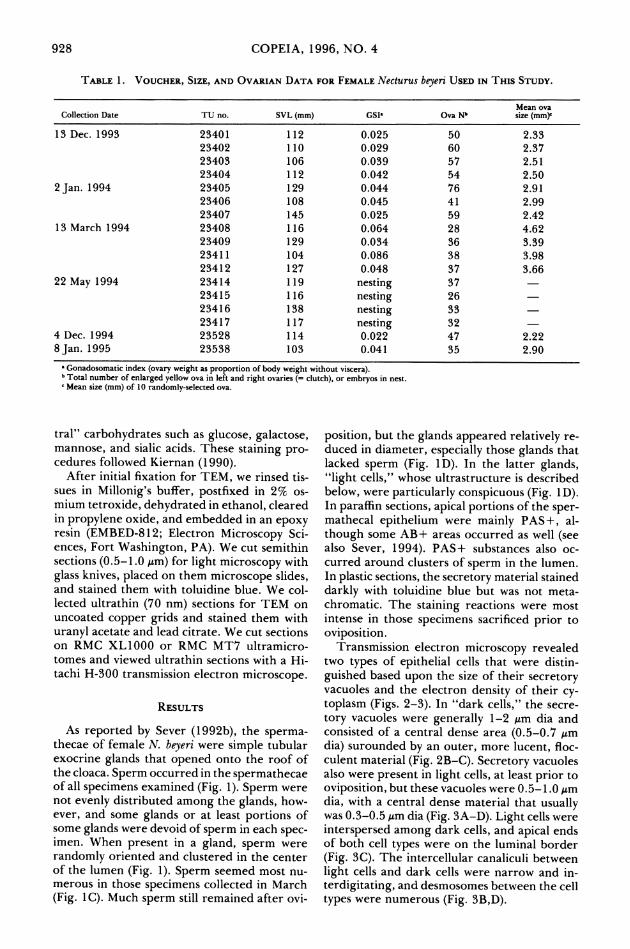
TABLE 1. VOUCHER, SIZE, AND OVARIAN DATA FOR FEMALE Necturus beyeri USED IN THIS STUDY.
Mean ova Collection Date TU no. SVL (mm) GSI' Ova Nb size (mm)
13 Dec. 1993 23401 112 0.025 50 2.33 23402 110 0.029 60 2.37 23403 106 0.039 57 2.51 23404 112 0.042 54 2.50
2 Jan. 1994 23405 129 0.044 76 2.91 23406 108 0.045 41 2.99 23407 145 0.025 59 2.42
13 March 1994 23408 116 0.064 28 4.62 23409 129 0.034 36 3.39 23411 104 0.086 38 3.98 23412 127 0.048 37 3.66
22 May 1994 23414 119 nesting 37 23415 116 nesting 26 23416 138 nesting 33 23417 117 nesting 32
4 Dec. 1994 23528 114 0.022 47 2.22 8 Jan. 1995 23538 103 0.041 35 2.90
* Gonadosomatic index (ovary weight as proportion of body weight without viscera). b Total number of enlarged yellow ova in left and right ovaries (= clutch), or embryos in nest. c Mean size (mm) of 10 randomly-selected ova.
tral" carbohydrates such as glucose, galactose, mannose, and sialic acids. These staining pro- cedures followed Kiernan (1990).
After initial fixation for TEM, we rinsed tis- sues in Millonig's buffer, postfixed in 2% os- mium tetroxide, dehydrated in ethanol, cleared in propylene oxide, and embedded in an epoxy resin (EMBED-812; Electron Microscopy Sci- ences, Fort Washington, PA). We cut semithin sections (0.5-1.0 ium) for light microscopy with glass knives, placed on them microscope slides, and stained them with toluidine blue. We col- lected ultrathin (70 nm) sections for TEM on uncoated copper grids and stained them with uranyl acetate and lead citrate. We cut sections on RMC XL1000 or RMC MT7 ultramicro- tomes and viewed ultrathin sections with a Hi- tachi H-300 transmission electron microscope.
RESULTS
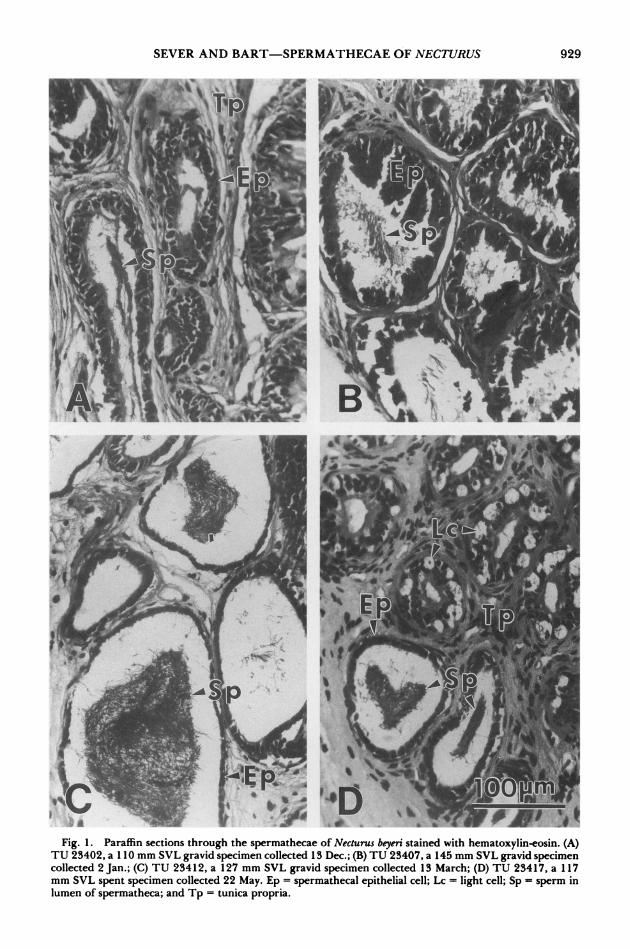
As reported by Sever (1992b), the sperma- thecae of female N. beyeri were simple tubular exocrine glands that opened onto the roof of the cloaca. Sperm occurred in the spermathecae of all specimens examined (Fig. 1). Sperm were not evenly distributed among the glands, how- ever, and some glands or at least portions of some glands were devoid of sperm in each spec- imen. When present in a gland, sperm were randomly oriented and clustered in the center of the lumen (Fig. 1). Sperm seemed most nu- merous in those specimens collected in March (Fig. 1C). Much sperm still remained after ovi-
position, but the glands appeared relatively re- duced in diameter, especially those glands that lacked sperm (Fig. 1D). In the latter glands, "light cells," whose ultrastructure is described below, were particularly conspicuous (Fig. 1D). In paraffin sections, apical portions of the sper- mathecal epithelium were mainly PAS+, al- though some AB+ areas occurred as well (see also Sever, 1994). PAS+ substances also oc- curred around clusters of sperm in the lumen. In plastic sections, the secretory material stained darkly with toluidine blue but was not meta- chromatic. The staining reactions were most intense in those specimens sacrificed prior to oviposition.
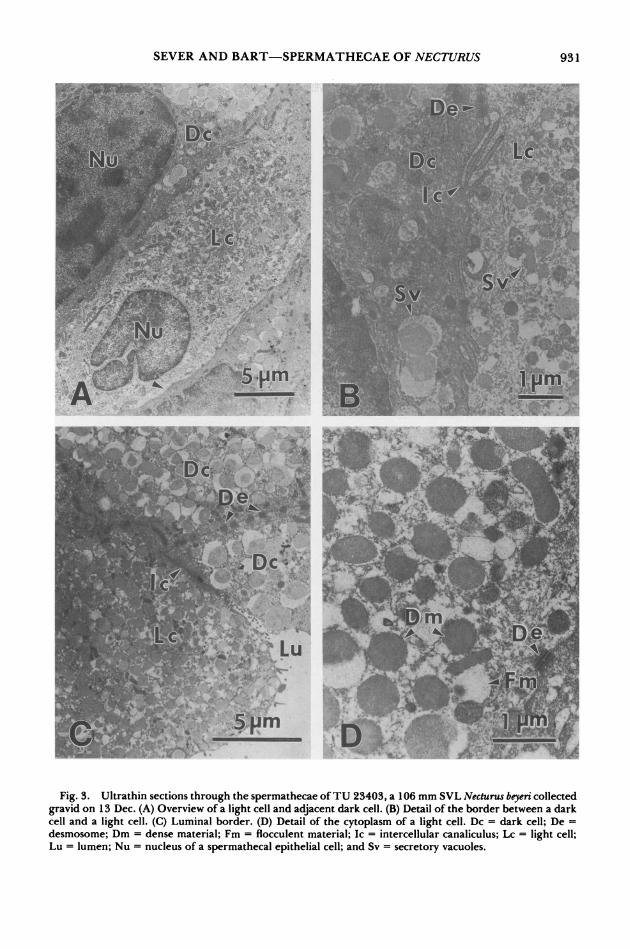
Transmission electron microscopy revealed two types of epithelial cells that were distin- guished based upon the size of their secretory vacuoles and the electron density of their cy- toplasm (Figs. 2-3). In "dark cells," the secre- tory vacuoles were generally 1-2 ,m dia and consisted of a central dense area (0.5-0.7 ,um dia) surounded by an outer, more lucent, floc- culent material (Fig. 2B-C). Secretory vacuoles also were present in light cells, at least prior to oviposition, but these vacuoles were 0.5-1.0 ,m dia, with a central dense material that usually was 0.3-0.5 ,m dia (Fig. 3A-D). Light cells were interspersed among dark cells, and apical ends of both cell types were on the luminal border (Fig. 3C). The intercellular canaliculi between light cells and dark cells were narrow and in- terdigitating, and desmosomes between the cell types were numerous (Fig. 3B,D).
928
SEVER AND BART-SPERMATHECAE OF NECTURUS
Fi. 1' ' s 0 f Fig. 1. Paraffin sections through the spermathecae of Necturus beyeri stained with hematoxylin-eosin. (A)
TU 23402, a 110 mm SVL gravid specimen collected 13 Dec.; (B) TU 23407, a 145 mm SVL gravid specimen collected 2 Jan.; (C) TU 23412, a 127 mm SVL gravid specimen collected 13 March; (D) TU 23417, a 117 mm SVL spent specimen collected 22 May. Ep = spermathecal epithelial cell; Lc = light cell; Sp = sperm in lumen of spermatheca; and Tp = tunica propria.
929
\ t^l . .fl .
lA4 i

COPEIA, 1996, NO. 4
Fig. 2. Spermathecae of TU 23403, a 106 mm SVL Necturus beyeri collected gravid on 13 Dec. (A) Plastic semithin section stained with toluidine blue. (B) Ultrathin section of a dark cell. (C) Ultrathin section showing detail of secretory vacuoles and organelles involved in synthesis of the product. Cv = condensing vacuoles; De = desmosome; Dm = dense material; Ep = spermathecal epithelial cells; Fm = flocculent material; Go =
Golgi apparatus; Ic = intercellular canaliculus; Lu = lumen; Mf = microfilaments; Nu = nucleus of a sper- mathecal epithelial cell; Rer = rough endoplasmic reticulum; Sv = secretory vacuoles; and Tp = tunica propria.
930
SEVER AND BART-SPERMATHECAE OF NECTURUS
i > yS; ^. {t h 3aJSf1'-^-..~. ~-'
Jte^"*>^' ^*;'C ~~~~~~~~~~::2
" r
A, .' . :" 4.
r?
5I m
0,:. {~.:.~ ~ .. :,-..
,,
-?,,..
~~. :P Lu P- . ;-
.,.~.~ ...:~,,,~ .... ~? L
~ '" :'~
';~ ,?,~,' ,,,
Fig. 3. Ultrathin sections through the spermathecae of TU 23403, a 106 mm SVL Necturus beyeri collected gravid on 13 Dec. (A) Overview of a light cell and adjacent dark cell. (B) Detail of the border between a dark cell and a light cell. (C) Luminal border. (D) Detail of the cytoplasm of a light cell. Dc = dark cell; De = desmosome; Dm = dense material; Fm = flocculent material; Ic = intercellular canaliculus; Lc = light cell; Lu = lumen; Nu = nucleus of a spermathecal epithelial cell; and Sv = secretory vacuoles.
931
.;s o..,
,~:'':'~,.?.t "~. x~ + L?~'." ... ...~-~~ . / .D" ' '~.;-t,.-
'
'D -'' * , '
'. 4~'. " T, ?; ? .. ... :,

COPEIA, 1996, NO. 4
The secretory vacuoles in the light and dark cells probably were responsible for the PAS+ reaction in the cytoplasm since the PAS+ re- action was most intense around the luminal bor- der where the secretory vacuoles were most nu- merous. Therefore, release of the product like- ly was responsible for the PAS + reaction around sperm in the lumen. Golgi bodies and rough endoplasmic rectiulum (Rer) were associated with the vacuoles in the cytoplasm (Fig. 2C). A PAS+ reaction combined with these synthetic organelles indicates the product contained gly- coproteins (Kiernan, 1990).
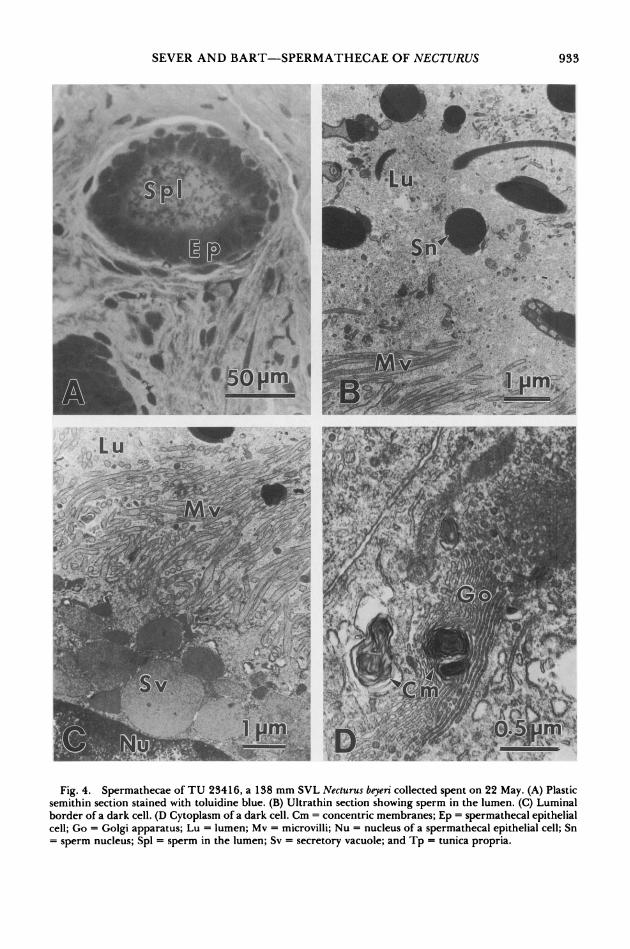
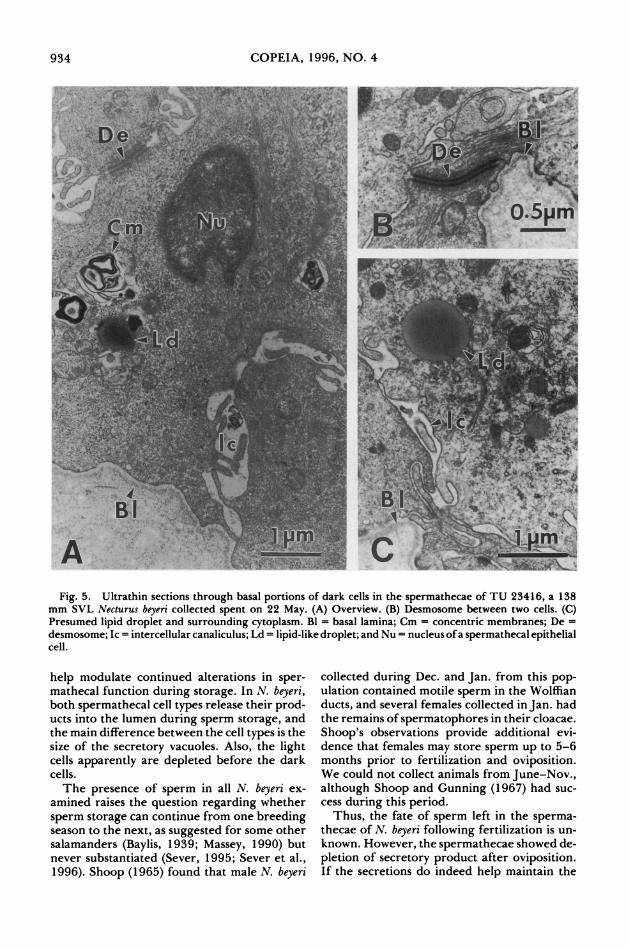
In specimens collected in May, shortly after oviposition, sperm still were numerous in some spermathecae (Figs. 1D, 4A-B). The luminal sperm appeared normal in cytology (Fig. 4B). Instead of a wide supranuclear area filled with secretory vacuoles, the apical cytoplasm of spec- imens collected after oviposition contained rel- atively few vacuoles (Fig. 4C). Also, the secre- tory vacuoles appeared more uniform in density (Fig. 4C) than those observed in specimens sac- rificed prior to oviposition. Microvilli seemed relatively more elongate after oviposition (Fig. 4B-C). No evidence of spermiophagy by the spermathecal epithelium was noted, but struc- tures consisting of concentric membranes were common especially basally in the cytoplasm of specimens sacrificed after oviposition (Figs. 4D, 5A). These concentric membrane structures of- ten were associated with Golgi bodies (Fig. 4D). Intercellular canaliculi were wider basally fol- lowing oviposition (Fig. 5A), and desmosomes were frequent at the basal borders of adjacent cells (Fig. 5B). Basal globules that appear to be lipid droplets (Sever and Kloepfer, 1993) were seen in individuals sacrificed after oviposition (Fig. 5C).
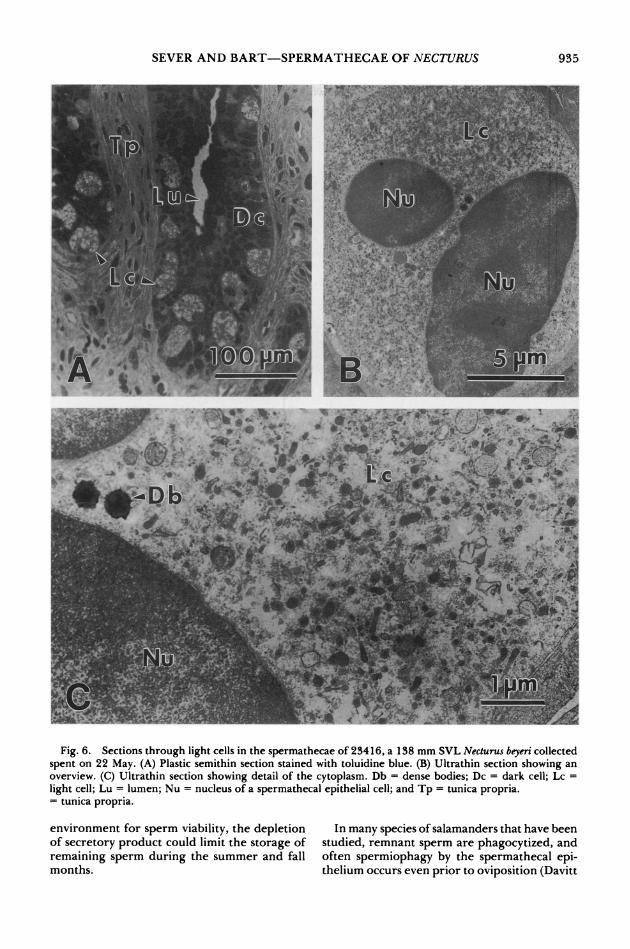
Light cells seemed especially depleted after oviposition. Secretory vacuoles and organelles involved in synthetic activity were not notice- able in these light cells (Fig. 6). The most con- spicuous structures were small dense bodies (about 0.3 ,tm) that occasionally were seen in the cytoplasm (Fig. 6C).
DISCUSSION
Secretory vacuoles in the plethodontid Eu- rycea cirrigera (Sever, 1991b, 1992a) and in the salamandrid Notophthalmus viridescens (Sever et al., 1996) are of uniform density throughout the period of production. Secretory vacuoles consisting of a central dense area and a sur- rounding flocculent material have been noted in the salamandrid Salamandrina terdigitata (Brizzi et al. 1989, 1995) and in the ambysto-
matids Ambystoma opacum (Sever and Kloepfer, 1993) and A. tigrinum (Sever, 1995). In S. ter- digitata and A. opacum, both the dense material and the flocculent material are exported into the lumen to bathe the sperm during storage (Brizzi et al., 1995; Sever and Klopefer, 1993). In A. tigrinum, however, Sever (1995) believed that the flocculent material results from disso- ciation of the compacted granules, and the vac- uole is then released into the lumen via exo-
cytosis. The process of release of the secretory material was not determined in N. beyeri, but the quantity of secretory vacuoles along the api- cal border decreased during sperm storage, and the release of the product probably is merocrine as reported in other salamanders (Brizzi et al., 1995; Sever, 1991a, 1995).
The secretion bathed the sperm during stor- age in N. beyeri, as reported in salamandrids (Brizzi et al., 1995, Sever et al., 1996) and in A. opacum (Sever and Kloepfer, 1993). Since sperm are inactive during storage (Hardy and Dent, 1986), Sever and Kloepfer (1993) proposed that this type of secretion helps maintain the envi- ronment for sperm viability and quiescence. In A. tigrinum, however, the secretions are not re- leased until oviposition and may serve to flush the sperm from the spermathecae (Sever, 1995). This role for these secretions, hypothesized by Sever (1995) to be the ancestral function of spermathecal secretions, is associated with short- term sperm storage (several days or less). Nec- turus beyeri can store sperm for at least six months prior to oviposition (Dec.-May), and the secre- tions apparently were released slowly through- out the period of storage, since some secretory vacuoles still remained (at least in the dark cells) after oviposition. Thus, we hypothesize that the secretions provide the environment for keeping the sperm viable but inactive during long-term storage in N. beyeri.
We found two distinct types of spermathecal epithelial cells, dark cells and light cells, in N. beyeri. Only one other study reported more than one type of epithelial cell in spermathecae of salamanders. Davitt and Larsen (1990), in an abstract, stated that an unspecified "plethodon- tid salamander" has five types of cells in its sper- mathecae. Their cell type II consists of an elec- tron dense core surrounded by more lucent ar- eas, much like the secretory vacuoles we found in N. beyeri. The other cell types found by Davitt and Larsen (1990) contain vacuoles that differ in size and shape as well as the density of the product, which is uniform in each cell type. Dav- itt and Larson (1990) proposed that some of the secretions are involved in sperm activation whereas others, including those in cell type II,
932
SEVER AND BART-SPERMATHECAE OF NECTURUS
L' :u ' ' : ..... Lu .- '~ .
',. .:,- .''~'
? - . . ... . ... s ~ ~ ,;f, ~ ;,~
f~~~V~~f>:.> ,c) ~~~~-e
I - z/ ' i "'" .... ~'", '.~i.' ."~S~
'.", . r '~,. ', ,, jJf';n~
1 m u m & , ~ ~ ~ ~ ~ ~ , v ( 4 t~ - ; - '
I? "
C ; - ' . . , '~ '~'~';'"'~ ~'~~: .~..-:?-:i.,"~~~~~~.
i.f ! .. : ~ .....'' .'- 'r ,! A
'~ ,D ,;?,,' ::"","~,Q? ' :,~r " ?=~;. ,' ? ' , , - u i: . ~ :.. ~,.?.- :. ?'?4''~
Fig. 4. Spermathecae of TU 23416, a 138 mm SVL Necturus beyeri collected spent on 22 May. (A) Plastic semithin section stained with toluidine blue. (B) Ultrathin section showing sperm in the lumen. (C) Luminal border of a dark cell. (D Cytoplasm of a dark cell. Cm = concentric membranes; Ep = spermathecal epithelial cell; Go = Golgi apparatus; Lu = lumen; Mv = microvilli; Nu = nucleus of a spermathecal epithelial cell; Sn = sperm nucleus; Spl = sperm in the lumen; Sv = secretory vacuole; and Tp = tunica propria.
933
COPEIA, 1996, NO. 4
Fig. 5. Ultrathin sections through basal portions of dark cells in the spermathecae of TU 23416, a 138 mm SVL Necturus beyeri collected spent on 22 May. (A) Overview. (B) Desmosome between two cells. (C) Presumed lipid droplet and surrounding cytoplasm. Bl = basal lamina; Cm = concentric membranes; De = desmosome; Ic = intercellular canaliculus; Ld = lipid-like droplet; and Nu = nucleus of a spermathecal epithelial cell.
help modulate continued alterations in sper- mathecal function during storage. In N. beyeri, both spermathecal cell types release their prod- ucts into the lumen during sperm storage, and the main difference between the cell types is the size of the secretory vacuoles. Also, the light cells apparently are depleted before the dark cells.
The presence of sperm in all N. beyeri ex- amined raises the question regarding whether
sperm storage can continue from one breeding season to the next, as suggested for some other salamanders (Baylis, 1939; Massey, 1990) but never substantiated (Sever, 1995; Sever et al., 1996). Shoop (1965) found that male N. beyeri
collected during Dec. and Jan. from this pop- ulation contained motile sperm in the Wolffian ducts, and several females collected in Jan. had the remains of spermatophores in their cloacae. Shoop's observations provide additional evi- dence that females may store sperm up to 5-6 months prior to fertilization and oviposition. We could not collect animals from June-Nov., although Shoop and Gunning (1967) had suc- cess during this period.
Thus, the fate of sperm left in the sperma- thecae of N. beyeri following fertilization is un- known. However, the spermathecae showed de- pletion of secretory product after oviposition. If the secretions do indeed help maintain the
934
SEVER AND BART-SPERMATHECAE OF NECTURUS
.4js~ #j.#'?. 0~ L3~~~~C ~ ;r ~.!e~u,L~
1? iB
Fig. 6. Sections through light cells in the spermathecae of 23416, a 138 mm SVL Necturus beyeri collected spent on 22 May. (A) Plastic semithin section stained with toluidine blue. (B) Ultrathin section showing an overview. (C) Ultrathin section showing detail of the cytoplasm. Db = dense bodies; Dc = dark cell; Lc = light cell; Lu = lumen; Nu = nucleus of a spermathecal epithelial cell; and Tp = tunica propria. = tunica propria.
environment for sperm viability, the depletion of secretory product could limit the storage of remaining sperm during the summer and fall months.
In many species of salamanders that have been studied, remnant sperm are phagocytized, and often spermiophagy by the spermathecal epi- thelium occurs even prior to oviposition (Davitt
935

COPEIA, 1996, NO. 4
and Larson, 1988b; Sever, 1992a; Sever and Kloepfer, 1993). No evidence of spermiophagy was found in N. beyeri, a situation also noted in A. tigrinum (Sever, 1995). Perhaps the sperm remaining in the spermathecae simply "leak out" gradually or are expelled by myoepithelial con- tractions, as reported for sperm remaining in the vas deferens of A. macrodactylum (Zalisko and Larsen, 1989). A complete understanding of the entire annual cycle of sperm storage in fe- male N. beyeri awaits the study of specimens with known breeding histories from June-Nov.
ACKNOWLEDGMENTS
Laboratory work at Saint Mary's College was supported by National Science Foundation grant DEB 9024918 to DMS. We thank D. Miller and E. Moen for their help in preparing tissue sam- ples. This is publication number 9 from the Saint Mary's College Electron Microscopy Fa- cility.
LITERATURE CITED
BAYLIS, H. A. 1939. Delayed reproduction in the spotted salamander. Proc. Zool. Lond. 109A:243- 246.
BOISSEAU, C., ANDJ.JOLY. 1975. Transport and sur- vival of spermatozoa in female Amphibia, p. 94- 104. In: The biology of spermatozoa: transport, survival, and fertilizing ability. E. S. E. Hafez and C. G. Thibault (eds.). Karger, Basel, Switzerland.
BRIZZI, R., G. DELFINO, AND C. CALLONI. 1989. Fe- male cloacal anatomy in the spectacled salamander, Salamandrina terdigitata (Amphibia: Salamandri- dae). Herpetologica 45:310-322.
, M. G. SELMI, AND D. M. SEVER. 1995. The spermathecae of Salamandrina terdigitata (Am- phibia: Salamandridae): patterns of sperm storage and degradation. J. Morphol. 223:21-33.
DAVITT, C. M., AND J. H. LARSEN JR. 1988a. Scan- ning electron microscopy of the spermatheca of Plethodon larselli (Amphibia: Plethodontidae): changes in the surface morphology of the sper- mathecal tubule prior to ovulation. Scanning Mi- crosc. 2:1805-1812.
, AND . 1988b. Phagocytosis of stored spermatozoa and cytoplasmic droplets by the sper- mathecal epithelium of the female salamander Rhy- acotriton olympicus. Am. Zool. 28:30A.
,AND . 1990. Morphology of the prin- ciple cell types of the plethodontid salamander sper- matheca following treatment with gonadotropin. Ibid. 30:38A.
DENT, J. N. 1970. The ultrastructure of the sper- matheca in the red spotted newt. J. Morphol. 132: 397-424.
FROST, D. R. (ED). 1985. Amphibian species of the world: a taxonomic and geographic reference. Al- len Press, Inc., and Association of Systematic Col- lections, Lawrence, KS.
HARDY, M. P., ANDJ. N. DENT. 1986. Transport of sperm within the cloaca of the female red-spotted newt. J. Morphol. 190:259-270.
JORDAN, E. 0. 1893. The habits and development of the newt (Diemyctylus viridescens). Ibid. 8:269-366.
KIERNAN, J. A. 1990. Histological and histochemical methods: theory and practice. 2d ed. Pergamon Press, Oxford.
MANNING, R. B. 1986. A small trap for collecting crustaceans in shallow water. Proc. Biol. Soc. Wash. 99:266-268.
MASSEY, A. 1990. Notes on the reproductive ecology of red-spotted newts (Notophthalmus viridescens). J. Herpetol. 24:106-107.
POOL, T. B., AND T. R. HOAGE. 1973. The ultra- structure of secretion in the spermatheca of the salamander, Manculus quadridigitatus (Holbrook). Tissue Cell 5:303-313.
SALTHE, S. N. 1967. Courtship patterns and the phy- logeny of the urodeles. Copeia 1967:100-117.
SEVER, D. M. 199 la. Comparative anatomy and phy- logeny of the cloacae of salamanders (Amphibia: Caudata). I. Evolution at the family level. Herpe- tologica 47:165-193.
. 199 b. Sperm storage and degradation in the spermathecae of the salamander Eurycea cirri- gera (Green). J. Morphol. 210:71-84.
. 1992a. Spermiophagy by the spermathecal epithelium of the salamander Eurycea cirrigera. Ibid. 212:281-290.
1992b. Comparative anatomy and phylogeny of the cloacae of salamanders (Amphibia: Caudata). V. Proteidae. Herpetologica 48:318-329.
1994. Observations on regionalization of se- cretory activity in the spermathecae of salamanders and comments on phylogeny of sperm storage in female salamanders. Ibid. 50:383-397.
. 1995. Spermathecae of Ambystoma tigrinum (Amphibia Caudata): development and a role for the secretion. J. Herpetol. 29:243-255.
,AND N. S. BRUNETTE. 1993. Regionalization of eccrine and spermiophagic activity in the sper- mathecae of the salamander Eurycea cirrigera (Am- phibia: Plethodontidae).J. Morphol. 217:161-170.
,AND N. M. KLOEPFER. 1993. Spermathecal cytology of Ambystoma opacum (Amphibia: Ambys- tomatidae) and the phylogeny of sperm storage or- gans in female salamanders. Ibid. 217:115-127.
,J. D. KRENZ, K. M.JOHNSON, AND L. C. RANIA. 1995. Morphology and evolutionary implications of the annual cycle of secretion and sperm storage in spermathecae of the salamander Ambystoma opa- cum (Amphibia: Ambystomatidae). Ibid. 223:35-46.
, L. C. RANIA, AND J. D. KRENZ. 1996. The annual cycle of sperm storage in the spermathecae of the red-spotted newt, Notophthalmus viridescens (Amphiba: Salamandridae). Ibid. 227:145-170.
SnooP, C. R. 1965. Aspects of reproduction in Lou- isiana Necturus populations. Amer. Midi. Nat. 74: 357-367.
, AND G. E. GUNNING. 1967. Seasonal activity and movements of Necturus in Louisiana. Copeia 1967:732-737.
ZALISKO, E. J., AND J. H. LARSEN JR. 1989. Fate of
936

SEVER AND BART-SPERMATHECAE OF NECTURUS SEVER AND BART-SPERMATHECAE OF NECTURUS
unused sperm in post-breeding male Ambystoma ma- crodactylum columbianum. J. Herpetol. 23:463-464.
(DMS) DEPARTMENT OF BIOLOGY, SAINT MARY'S COLLEGE, NOTRE DAME, INDIANA 46556; AND (HLB) TULANE UNIVERSITY MUSEUM OF
unused sperm in post-breeding male Ambystoma ma- crodactylum columbianum. J. Herpetol. 23:463-464.
(DMS) DEPARTMENT OF BIOLOGY, SAINT MARY'S COLLEGE, NOTRE DAME, INDIANA 46556; AND (HLB) TULANE UNIVERSITY MUSEUM OF
NATURAL HISTORY, BELLE CHASSE, LOUI- SIANA 70037. Send reprint requests to DMS. Submitted: 8 Sept. 1995. Accepted: 23 Jan. 1996. Section editor: J. R. Spotila.
NATURAL HISTORY, BELLE CHASSE, LOUI- SIANA 70037. Send reprint requests to DMS. Submitted: 8 Sept. 1995. Accepted: 23 Jan. 1996. Section editor: J. R. Spotila.
Copeia, 1996(4), pp. 937-944
Gymnotus bahianus sp. nov., a New Gymnotid Fish from Eastern Brazil (Teleostei: Ostariophysi: Gymnotiformes),
with Evidence for the Monophyly of the Genus
RICARDO CAMPOS-DA-PAZ AND WILSON J. E. M. COSTA
Gymnotus bahianus sp. nov. is described from the Rio Almada basin, near Ilheus, Bahia State, Brazil. It is currently known only from that area, repre- senting the third record of a gymnotid species in eastern Brazilian coastal plain basins, north of the Rio Doce basin, Espirito Santo State. Gymnotus bahianus shares with G. cylindricus LaMonte, G. inaequilabiatus (Valenciennes), and G. pantherinus (Steindachner), a pattern of small dark rounded spots over the body, which distinguish these species from their congeners. The new species is further distinguished by a unique combination of morphological features, including depth of the body and head, length of the head, pectoral fin and maxillary bone, and the number of perforated scales along lateral line. The phylogenetic rela- tionships of the new species are still unknown. New evidence for the monophyly of Gymnotus is presented.
Gymnotus bahianus sp. nov. e descrita da bacia do rio Almada, vizinhancas de Ilheus, Estado da Bahia, Brasil. Atualmente, e conhecida apenas daquelas areas, consistindo o terceiro registro de uma especie de Gymnotidae para as bacias costeiras do leste do Brasil, ao norte da bacia do rio Doce, Estado do Espirito Santo. Gymnotus bahianus e similar a G. cylindricus LaMonte, G. inaequilabiatus (Valenciennes) e G. pantherinus (Steindachner), diferindo das demais especies do genero por seu padrio de colorido, o qual consiste de pequenas manchas escuras e arredondadas distribuidas sobre o corpo. A nova espicie e tambem distinta com base numa combinacao unica de caracteristicas morfologicas, com- preendendo as alturas do corpo e da cabeca, comprimentos da cabeca, nadadeira peitoral e osso maxilar, e numero de escamas perfuradas da linha lateral. As relaqies filogeneticas de G. bahianus sio ainda desconhecidas. Nova evidencia para o monofiletismo de Gymnotus e apresentada.
Copeia, 1996(4), pp. 937-944
Gymnotus bahianus sp. nov., a New Gymnotid Fish from Eastern Brazil (Teleostei: Ostariophysi: Gymnotiformes),
with Evidence for the Monophyly of the Genus
RICARDO CAMPOS-DA-PAZ AND WILSON J. E. M. COSTA
Gymnotus bahianus sp. nov. is described from the Rio Almada basin, near Ilheus, Bahia State, Brazil. It is currently known only from that area, repre- senting the third record of a gymnotid species in eastern Brazilian coastal plain basins, north of the Rio Doce basin, Espirito Santo State. Gymnotus bahianus shares with G. cylindricus LaMonte, G. inaequilabiatus (Valenciennes), and G. pantherinus (Steindachner), a pattern of small dark rounded spots over the body, which distinguish these species from their congeners. The new species is further distinguished by a unique combination of morphological features, including depth of the body and head, length of the head, pectoral fin and maxillary bone, and the number of perforated scales along lateral line. The phylogenetic rela- tionships of the new species are still unknown. New evidence for the monophyly of Gymnotus is presented.
Gymnotus bahianus sp. nov. e descrita da bacia do rio Almada, vizinhancas de Ilheus, Estado da Bahia, Brasil. Atualmente, e conhecida apenas daquelas areas, consistindo o terceiro registro de uma especie de Gymnotidae para as bacias costeiras do leste do Brasil, ao norte da bacia do rio Doce, Estado do Espirito Santo. Gymnotus bahianus e similar a G. cylindricus LaMonte, G. inaequilabiatus (Valenciennes) e G. pantherinus (Steindachner), diferindo das demais especies do genero por seu padrio de colorido, o qual consiste de pequenas manchas escuras e arredondadas distribuidas sobre o corpo. A nova espicie e tambem distinta com base numa combinacao unica de caracteristicas morfologicas, com- preendendo as alturas do corpo e da cabeca, comprimentos da cabeca, nadadeira peitoral e osso maxilar, e numero de escamas perfuradas da linha lateral. As relaqies filogeneticas de G. bahianus sio ainda desconhecidas. Nova evidencia para o monofiletismo de Gymnotus e apresentada.
FISHES of the ostariophysan order Gymno- tiformes attracted the attention of early
ichthyologists mainly because of their very spe- cialized morphology (e.g., Kaup, 1856). Sub-
sequently, Lissmann (1951, 1958) demonstrat- ed that gymnotiform species, in addition to the electric eel Electrophorus electricus (Linnaeus), possess electrogenic and electroreceptive abili- ties, resulting in a change in the focus of interest to the electrophysiology of the order. As a re-
FISHES of the ostariophysan order Gymno- tiformes attracted the attention of early
ichthyologists mainly because of their very spe- cialized morphology (e.g., Kaup, 1856). Sub-
sequently, Lissmann (1951, 1958) demonstrat- ed that gymnotiform species, in addition to the electric eel Electrophorus electricus (Linnaeus), possess electrogenic and electroreceptive abili- ties, resulting in a change in the focus of interest to the electrophysiology of the order. As a re-
sult, an increasing amount of information con- cerning gymnotiform electrophysiology has be- come available (e.g., Heiligenberg and Bastian, 1980; Kramer et al., 1981), but taxonomic problems at various levels preclude an objective investigation of those observed patterns. Ellis' monograph (1913) and the recently published study of Mago-Leccia (1994) are the most com- prehensive published studies on gymnotiform taxonomy, but our understanding of the species
sult, an increasing amount of information con- cerning gymnotiform electrophysiology has be- come available (e.g., Heiligenberg and Bastian, 1980; Kramer et al., 1981), but taxonomic problems at various levels preclude an objective investigation of those observed patterns. Ellis' monograph (1913) and the recently published study of Mago-Leccia (1994) are the most com- prehensive published studies on gymnotiform taxonomy, but our understanding of the species
? 1996 by the American Society of Ichthyologists and Herpetologists ? 1996 by the American Society of Ichthyologists and Herpetologists
937 937


















