Coordination Chemistry Reviews - REZOLVE SCIENTIFIC · recent therapeutic applications will be...
22
Review Bright lights down under: Metal ion complexes turning the spotlight on metabolic processes at the cellular level q Todd A. Gillam a , Martin J. Sweetman a,b , Christie A. Bader c , Janna L. Morrison d , John D. Hayball b,e , Doug A. Brooks c , Sally E. Plush a,f,⇑ a School of Pharmacy and Medical Sciences, University of South Australia, Adelaide, SA 5000, Australia b Experimental Therapeutics Laboratory, Hanson Institute and Sansom Institute, School of Pharmacy and Medical Sciences, University of South Australia, Adelaide, SA 5000, Australia c Mechanisms in Cell Biology and Disease Research Group, School of Pharmacy and Medical Sciences, Sansom Institute for Health Research, University of South Australia, Adelaide, Australia d Early Origins of Adult Health Research Group, School of Pharmacy and Medical Sciences, Sansom Institute for Health Research, University of South Australia, Adelaide, South Australia 5000, Australia e Robinson Research Institute, Discipline of Obstetrics and Gynaecology, School of Medicine, University of Adelaide, SA 5005, Australia f Future Industries Institute, University of South Australia, Mawson Lakes, SA 5095, Australia article info Article history: Received 29 September 2017 Received in revised form 20 November 2017 Accepted 21 November 2017 Available online 8 December 2017 abstract The widespread uptake and use of emissive metal ion complexes for investigating cellular structure, com- position and function, is evidence of the effectiveness and vast potential for this type of imaging probe. Metal ion complexes provide significant advantages over their organic fluorophore counterparts, includ- ing long emission lifetime, resistance to photobleaching and the capacity to readily modify their periph- eral chemistry to target specific organelles, signalling pathways and individual molecules. This review will discuss recent progress in the development and use of metal ion complexes, specifically for studying metabolic diseases at the cellular level. Advanced metal ion complexes for organelle imaging and the detection of biorelevant species, to elaborate complexes for understanding cellular mechanisms and recent therapeutic applications will be reviewed. To align with the special issue, Coordination Chemistry Reviews: Coordination Chemistry in Australia, the work of Australian researchers actively engaged in this field is featured prominently, along with key developments from the global research community. Crown Copyright Ó 2017 Published by Elsevier B.V. All rights reserved. Contents 1. Introduction ......................................................................................................... 235 2. Organelle specific imaging.............................................................................................. 236 2.1. Investigating lipid distribution profiles of the lipid droplet/liposome ...................................................... 236 2.2. Visualising endoplasmic reticulum, and related processes ............................................................... 238 2.3. Imaging and therapeutic activity of metal ion complexes within mitochondria.............................................. 239 2.4. Investigation of lysosomal function with metal ion complexes ........................................................... 241 2.5. Imaging and therapeutic activity of metal ion complexes within the nucleus ............................................... 242 3. Metal ion complexes with conjugated targeting and functional motifs .......................................................... 242 3.1. Peptide or protein bearing complexes ............................................................................... 243 3.2. Biolabelling with metabolites ...................................................................................... 246 3.3. Carbohydrate appended complexes ................................................................................. 247 3.4. Therapeutic compounds conjugated to metal ion complexes............................................................. 248 4. Responsive complexes ................................................................................................. 249 4.1. Detecting pathological reactive oxygen and nitrogen species ............................................................ 249 https://doi.org/10.1016/j.ccr.2017.11.026 0010-8545/Crown Copyright Ó 2017 Published by Elsevier B.V. All rights reserved. q Author ORCID IDs: 0000-0003-2611-1991 (Todd A. Gillam), 0000-0002-9074-2125 (Martin J. Sweetman), 0000-0002-4653-912X (Christie A. Bader), 0000-0002-8602- 8519 (Janna L. Morrison), 0000-0002-3089-4506 (John D. Hayball), 0000-0001-9098-3626 (Doug A. Brooks), 0000-0002-9999-9154 (Sally E. Plush). ⇑ Corresponding author at: School of Pharmacy and Medical Sciences, University of South Australia, Adelaide, SA 5000, Australia. E-mail address: [email protected] (S.E. Plush). Coordination Chemistry Reviews 375 (2018) 234–255 Contents lists available at ScienceDirect Coordination Chemistry Reviews journal homepage: www.elsevier.com/locate/ccr
Transcript of Coordination Chemistry Reviews - REZOLVE SCIENTIFIC · recent therapeutic applications will be...

Coordination Chemistry Reviews 375 (2018) 234–255
Contents lists available at ScienceDirect
Coordination Chemistry Reviews
journal homepage: www.elsevier .com/ locate /ccr
Review
Bright lights down under: Metal ion complexes turning the spotlight onmetabolic processes at the cellular levelq
https://doi.org/10.1016/j.ccr.2017.11.0260010-8545/Crown Copyright � 2017 Published by Elsevier B.V. All rights reserved.
q Author ORCID IDs: 0000-0003-2611-1991 (Todd A. Gillam), 0000-0002-9074-2125 (Martin J. Sweetman), 0000-0002-4653-912X (Christie A. Bader), 0000-0008519 (Janna L. Morrison), 0000-0002-3089-4506 (John D. Hayball), 0000-0001-9098-3626 (Doug A. Brooks), 0000-0002-9999-9154 (Sally E. Plush).⇑ Corresponding author at: School of Pharmacy and Medical Sciences, University of South Australia, Adelaide, SA 5000, Australia.
E-mail address: [email protected] (S.E. Plush).
Todd A. Gillam a, Martin J. Sweetman a,b, Christie A. Bader c, Janna L. Morrison d, John D. Hayball b,e,Doug A. Brooks c, Sally E. Plush a,f,⇑a School of Pharmacy and Medical Sciences, University of South Australia, Adelaide, SA 5000, Australiab Experimental Therapeutics Laboratory, Hanson Institute and Sansom Institute, School of Pharmacy and Medical Sciences, University of South Australia, Adelaide, SA 5000, AustraliacMechanisms in Cell Biology and Disease Research Group, School of Pharmacy and Medical Sciences, Sansom Institute for Health Research, University of South Australia,Adelaide, Australiad Early Origins of Adult Health Research Group, School of Pharmacy and Medical Sciences, Sansom Institute for Health Research, University of South Australia, Adelaide,South Australia 5000, AustraliaeRobinson Research Institute, Discipline of Obstetrics and Gynaecology, School of Medicine, University of Adelaide, SA 5005, Australiaf Future Industries Institute, University of South Australia, Mawson Lakes, SA 5095, Australia
a r t i c l e i n f o
Article history:Received 29 September 2017Received in revised form 20 November 2017Accepted 21 November 2017Available online 8 December 2017
a b s t r a c t
The widespread uptake and use of emissive metal ion complexes for investigating cellular structure, com-position and function, is evidence of the effectiveness and vast potential for this type of imaging probe.Metal ion complexes provide significant advantages over their organic fluorophore counterparts, includ-ing long emission lifetime, resistance to photobleaching and the capacity to readily modify their periph-eral chemistry to target specific organelles, signalling pathways and individual molecules. This reviewwill discuss recent progress in the development and use of metal ion complexes, specifically for studyingmetabolic diseases at the cellular level. Advanced metal ion complexes for organelle imaging and thedetection of biorelevant species, to elaborate complexes for understanding cellular mechanisms andrecent therapeutic applications will be reviewed. To align with the special issue, CoordinationChemistry Reviews: Coordination Chemistry in Australia, the work of Australian researchers activelyengaged in this field is featured prominently, along with key developments from the global researchcommunity.
Crown Copyright � 2017 Published by Elsevier B.V. All rights reserved.
Contents
1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2352. Organelle specific imaging. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 236
2.1. Investigating lipid distribution profiles of the lipid droplet/liposome . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2362.2. Visualising endoplasmic reticulum, and related processes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2382.3. Imaging and therapeutic activity of metal ion complexes within mitochondria. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2392.4. Investigation of lysosomal function with metal ion complexes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2412.5. Imaging and therapeutic activity of metal ion complexes within the nucleus . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 242
3. Metal ion complexes with conjugated targeting and functional motifs . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 242
3.1. Peptide or protein bearing complexes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2433.2. Biolabelling with metabolites . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2463.3. Carbohydrate appended complexes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2473.4. Therapeutic compounds conjugated to metal ion complexes. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2484. Responsive complexes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 249
4.1. Detecting pathological reactive oxygen and nitrogen species . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2492-8602-

T.A. Gillam et al. / Coordination Chemistry Reviews 375 (2018) 234–255 235
4.2. Biomolecular recognition . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 253
5. Conclusion and future perspectives . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 253Acknowledgments . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 254References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 254
1. Introduction
Australia has a rich history in the research and application ofmetal ion complexes in biology, from the pioneering work of Fran-cis Dwyer in the 1950s, to the current developments of our inter-national research collaboration, showing heterogeneousarrangement of polar lipids within the matrix of cellular lipid dro-plets [1–3]. In recent times, there has been considerable researcheffort directed towards using metal ion complexes to enhancethe understanding of intracellular processes and, in particular, inrelation to disease pathogenesis [4]. The combination of advancedimaging techniques and the synthesis of highly emissive, novelmetal ion complexes is now allowing researchers to visualise andidentify subcellular components in unprecedented detail [5,6].This, coupled with the availability of responsive bio-analyticalmetal ion complexes that selectively report the detection of speci-fic metabolites, is allowing researchers to understand biologicalmechanisms as never before [7]. It is envisaged that understandingthese mechanisms will lead to the development of accurate andearly diagnosis of pathogenic conditions and vital medical break-throughs with the development of targeted therapeutics.
A significant area of research where the use of metal ion com-plexes will provide valuable new insights is in understandingmetabolic processes. The World Health Organisation (WHO) hasidentified that annually there are 40 million deaths, which equatesto 70% of all deaths worldwide are due to noncommunicable dis-eases including cancer, cardiovascular disease, chronic lung diseaseand diabetes [8]. An underlying link between these diseases isaltered cellular metabolism. To address the impact of metabolicdisease in the population, research efforts are focused on discern-ing the mechanistic basis of these pathologies at a molecular level.The detailed understanding of the mechanisms underpinning thepathogenesis of cancer or diabetes, and the effective targeting ofdiseased cells with therapeutic agents, may provide a paradigmshift in how diagnosis and treatment are practiced.
Presently, a range of diagnostic and investigative tools exist,such as fluorescence microscopes and magnetic resonance imaging(MRI) instruments, that allow detection and imaging of diseasedcells and tissue. These techniques require the cells to be appropri-ately identified and labelled, where chemical probes provide bothselective targeting and contrasting features that allow differentia-tion from endogenous biological material. The current key meth-ods for whole body imaging of disease tissues include positronemission tomography (PET) and MRI, both of which provide excel-lent spatial resolution as whole-body imaging platforms. Metal ioncomplexes of gadolinium are particularly effective contrast agentsfor MRI due to their effect on the T1 relaxation times on surround-ing nuclei [9]. At the cellular and subcellular level, fluorescencemicroscopy and flow cytometry are routinely used to image andtrack (in real-time) specifically labelled cells, organelles, biomole-cules and metabolites of interest.
Metal ion complexes are uniquely positioned as imaging agentsfor disease because they possess the necessary physical and chem-ical properties for detailed imaging. The ability to alter the chelat-ing ligands of the metal ion complex can facilitate the selectiveaccumulation of the complex in diseased tissues (such as tumours),cells or organelles of interest [10–12]. In terms of investigating the
fundamental mechanisms of metabolism, fluorescence microscopyis more commonly used than MRI or PET, although the uses varybetween research and clinical settings. To date, fluorescencemicroscopy for cellular imaging has largely utilised emissiveagents based on organic fluorophores, such as small fluorescentmolecules or fluorescently labelled antibodies. Organic fluo-rophores suffer significant limitations however, including photo-bleaching and short emissive lifetimes (nanoseconds). Fluores-cently labelled antibodies offer high substrate labelling specificity,though unfortunately require cell fixation for imaging, which doesnot permit the visualisation of dynamic cellular processes. In con-trast, emissive metal ion complexes exhibit high quantum yields,long-lived visible emission (milliseconds rather than nanoseconds)and are resistant to photo-bleaching [13]. Careful construction canalso yield metal ion complexes with a certain level of selectivity forlabelling specific cells or organelles [4]. Importantly, many emis-sive metal ion complexes that have been developed allow live cellimaging, as such they are particularly applicable to visualising live-dynamic processes which may be altered in disease [14,15].
Emissive metal ion complexes provide an ideal platform for thedevelopment of cellular imaging agents for both fluorescencemicroscopy and flow cytometry [13,16–18]. Fluorescence micro-scopy is particularly useful for observing discrepancies in cellularmorphology, while flow cytometry can rapidly differentiate selec-tively labelled cells from within a mixed population. Metal ioncomplexes have also been developed that function to report onthe presence of key metabolites, providing greater insights intocellular functions. Additionally, many metal ion complexes providean excellent platform for multimodal imaging that may combineFourier transform infrared (FTIR), Raman mapping, PET or MRI[19,20]. Due to the metal centre, these complexes are also usefulanalytes for inductively coupled plasma mass spectrometry (ICP-MS), allowing their cellular uptake to be accurately quantified[21,22]. ICP-MS is regarded as the most reliable method for theaccurate determination of metals, with sensitivity in the partsper trillion range [23]. Fortunately, emissive metal ions used inimaging agents are a rarity within the cellular matrix and can beseparated from an organic matrix by acidic or microwave digestionof biological samples, reducing matrix interferences.
The favourable emissive characteristics (photo-bleaching resis-tance and long emissive lifetimes) and potential biocompatibility(live cell imaging) of metal ion complexes has accelerated theexploitation of this type of probe by biologists. It is now highly rel-evant for chemists to pursue with earnest, the synthesis of metalion complexes furnished with ligands and moieties that will enablehighly specific interactions with relevant biomolecules. Biologistsrequire imaging agents with novel biological specificities and sub-strate specific responses, to allow the visualisation of the intricatesubcellular processes involved in different disease conditions. Dueto the chemical structure and photophysical properties of this classof compounds, metal ion complexes offer great potential for devel-oping selective, sensitive, compatible and versatile imaging agentsto study cellular mechanisms at the molecular level.
Luminescent metal ion complexes are derived from two distinctcategories. Firstly, low-spin transition metal complexes, such as Re(I), Ru(II), Ir(III) and Pt(II) [24,25]. Secondly, complexes of opticallyemissive lanthanoid(III) ions, such as Eu(III) and Tb(III), which are

236 T.A. Gillam et al. / Coordination Chemistry Reviews 375 (2018) 234–255
most commonly applied in cellular imaging applications [24].Near-infrared emitting lanthanoid complexes are also gaining con-siderable interest, however these complexes will not be discussedin this review [26,27]. Emission from low-spin metal ion com-plexes results from charge transfer between the metal centre andhighly conjugated ligands. The charge transfer bands of low-spinmetal ion complexes, namely ligand to metal charge transfer(LMCT) and metal to ligand charge transfer (MLCT) bands producelong-lived phosphorescent emission, a feature that permits sup-pression of background endogenous fluorescence within the cellusing time-gated imaging systems. Low-spin metal ion complexesare regarded as colour-tuneable, as incorporation of electron with-drawing and donating substituents within the metal bound ligandsmodulates the HOMO–LUMO energy gap altering the LMCT andMLCT emission. Lanthanoids exhibit low molar absorptivity dueto the Laporte forbidden f–f transition and as such possess intrin-sically weak quantum yields [28,29]. Fortunately, sensitisingligands based on organic fluorophores with highmolar absorptivitycan facilitate efficient population of the metal ions phosphorescenttriplet state by means of an intersystem crossing (ISC) chargetransfer mechanism. Sensitised lanthanoid(III) complexes produceenhanced triplet state emission with characteristically sharp emis-sive peaks, unique to each specific lanthanoid [20]. Recently, low-spin metal ion complexes have proven to be effective sensitisers oflanthanoid luminescence and as such mixed d–f complexes havebeen developed [30].
This review will discuss recent advances from our own researchgroup, the wider Australian chemistry community and outstandinginternational achievements. Metal ion complexes specific for keyorganelles, complexes targeted to key bioactive compounds andthose that can act as reporters of bio-analyte concentration arereviewed. This review will specifically focus on the impact thatmetal ion complexes have and will have on the study of metabolicdiseases with an emphasis on live cell imaging.
2. Organelle specific imaging
One of the key steps in labelling cells with emissive metal ioncomplexes is to ensure efficient uptake and internalisation of thecomplex. This has been the focus of considerable research effort,to identify specific ligand chemistries that facilitate targeteduptake [31]. In pursuit of developing the tools for the investigationof metabolic disease mechanisms, it is highly useful to develop andutilise metal ion complexes that localise to specific organelles.Labelling of specific organelles is particularly useful for evaluatingmorphological discrepancies within cells, identifying the localpresence of biomolecules of interest and for real-time tracking ofcellular processes. Probes that report behaviour and function ofspecific organelles can also confer significant information aboutcell health and function. Recent examples of metal ion complexesthat localise to specific organelles are discussed in this section,
Fig. 1. Structure of ReZolve-L1TM an
along with the application of these complexes for discerning func-tional information at the molecular level.
2.1. Investigating lipid distribution profiles of the lipid droplet/liposome
A collaborative venture between groups within University ofSouth Australia, Curtin University and the University of Bolognahas resulted in the development of interesting metal ion com-plexes suitable for imaging lipids within live cells. This new classof complexes addressed a deficit in the availability of live cellularimaging agents suited to lipid imaging. This approach leveragesthe knowledge that altered metabolic homeostasis of lipids is amarker of malignancy [32].
A Re(I) tricarbonyl tetrazolo complex bearing a 4-cyanophenyltetrazolato ancillary ligand, henceforth referred to as ReZolve-L1TM (Fig. 1) has been found to interact with polar lipids and to loca-lise within the core matrix of lipid droplets as well as withautolysosomes [10]. The lipid droplet is an organelle traditionallyassociated with the storage of neutral lipids, and as such was con-sidered to be a homogeneous environment with polar lipids onlyresiding in the membrane. Staining with ReZolve-L1TM revealed anon-homogenous localisation pattern within the droplet [3,10].This staining pattern hinted that there may be a specificity of thisprobe for a particular lipid species. A series of lipid overlay exper-iments revealed that this complex exhibits a high affinity for polarlipids such as sphingomyelin and phosphatidylethanolamine overneutral or non-polar lipid species [3]. While there has beenincreasing commentary that lipid droplets are more than justhomogenous pools of neutral lipids, there was little evidence toaccount for the staining observed. Therefore, in conjunction withthe University of Sydney, to account for the staining of ReZolve-L1 in lipid droplets, Raman mapping of the lipid droplets of Droso-phila fat body tissue revealed the presence of a spectral signatureunique to phospholipids (at �1096 cm�1). This supported the pre-mise that lipid droplets are quite heterogeneous, contrary to previ-ous assumptions that polar lipids were arranged at the periphery ofthe lipid droplets only [3].
Facial tricarbonyl metal ion complexes are highly amenable toFTIR investigation within the cellular environment, as the C„Ostretches are unobstructed by endogenous infrared (IR) absorbancewithin the cellular matrix, allowing for their detection in associa-tion with other biologically relevant signals. The localisation ofReZolve-L1TM with polar lipids within lipid droplets could thereforebe further characterised using FTIR studies within 3t3-L1 mam-malian adipocytes [19]. The three C„O stretches of the 4-cyanophenyl Re(I) tricarbonyl tetrazolo complex (at �2027 cm�1
and a doublet signal at 1915/1893), were detected in correlationwith C@O stretch of lipid esters at 1712–1775 cm�1 within thelipid droplet [19]. The FTIR spectra from two regions with highcomplex accumulation were extracted and compared to the
d tetrazolato complex 1 [10].

Fig. 2. Localisation of different lipid classes in H9c2 cardiomyoblasts using fluorescence microscopy. Triacylglycerides and cholesterol esters detected by BODIPY 493/503 (A),free cholesterol detected by Fillipin III (B) and localisation of polar lipids shown by ReZolve-L1 (C) (data currently in preparation for publication).
T.A. Gillam et al. / Coordination Chemistry Reviews 375 (2018) 234–255 237
spectra of pure lipids from eight representative lipid families. CAHstretches of the sampled regions showed correlation with the CAHstretching profile of polar lipids, confirming the selectivity of theaccumulation of ReZolve-L1TM with polar lipid species over neutrallipids.
The development of the ReZolve-L1TM represents the first exam-ple of a transition metal complex with polar lipid specificity thatcan be imaged in live cells. This complex is uniquely positionedas it offers several advantages over conventional fluorescent stainsfor lipid visualisation. Stains including, Oil red, Nile Red and FilipinIII require cell fixation, which can induce lipid depletion of the celland obstruct the visualisation of a dynamic, live system [3]. Fluo-rescent dyes such as BODIPY and LipidTOX are applicable for livecell imaging, but are prone to quenching effects. Presently theuse of the ReZolve-L1TM complex overcomes each of these limita-tions [3].
Lipophilicity is often provided as a rationale for cellular uptakeand localisation into lipid droplets. However, a comparison of thelipophilicity of two other commonly used lipophlilic lipid dropletimaging agents BODIPY 493/503 (log P = 3.5 ± 0.04 [38]) and NileRed (log P = 5.0 [53]) shows that ReZolve-L1TM (log P value = 2.53± 0.08) is at least an order of magnitude less lipophilic. Interest-ingly, slight changes in structure which have minimal impact onlog P value alter cellular localisation, suggesting that while adegree of lipophilicity is required for interactions with lipid envi-
Fig. 3. Re(III) tricarbonyl complexes that exhibit anti-proliferative effects on cancercells [35].
ronments the chemical structure is still key. For example, alter-ations to the structure of the lipophilic phenanthroline backboneor the ancillary tetrazolato ligand result in altered localisation pat-terns (some of which will be discussed in the following sections).This, along with research by Mitra and co-workers suggests thatthe 4-cyanophenyl motif confers a targeting effect to the lipid dro-plet and that a reduced lipophilicity may impart a degree of selec-tivity to particular lipid families [33]. The neutrality of ReZolve-L1TM is also expected to play a role in its staining pattern.
Fig. 2 shows the staining of different lipid families in H9c2 car-diomyoblasts (a myoblastic cell line derived from embryonic BD1Xrat heart tissue) using BODIPY, Filipin and ReZolve-L1TM. It is clearthat staining with ReZolve-L1TM provides superior structural detail,with bright areas correlating to regions of high polar lipid density.
This work challenges conventional assumptions about lipid dro-plet biology, demonstrating that both polar and neutral lipids aredistributed in the core matrix of these organelles. Stimulation ofautophagy within Drosophila larvae results in an accumulation ofReZolve-L1TM within the autophagosome, an organelle known toconcentrate lipids. A platform that enables the visualisation ofautophagy and lipid storage has considerable value in studyingmetabolic processes that exhibit pathological lipid metabolism,prostate cancer for instance, is known to exhibit differences in lipidmetabolism compared to healthy tissue [34].
The Re(I) tetrazolato complexes are highly biocompatible andkinetically stable within the cellular environment [10]. In a collab-oration with the Harris group at the University of Adelaide, thein vivo biostability of the Re(III) tetrazolato complexes has beendemonstrated. This was achieved using epifluorescence micro-scopy in conjunction with X-ray fluorescence investigations of22RV1 cells (human prostate carcinoma derived cell line) with aninternalised Re(I)(CO3)(1,10-phenantroline) tetrazolato complex(Fig. 1, complex 1), furnished with a 4-iodophenyl ancillary ligand[5]. The tetrazolato was shown to remain bound to the Re(I) metalcentre as the X-ray fluorescence spectrum of cellular Re and I werefound to maintain similar cellular distribution [5]. The data to datesuggests that the tetrazolato ligand itself, combined with the resul-tant neutrality may be key to the biocompatibility of thesecomplexes.
Although not applicable to lipid imaging, the in vivo stability ofcoordinating ligands of Re(CO3) complexes has been furtherexplored by the Massi group. Re(III) tricarbonyl complexes baringlabile N-heterocyclic carbene ligands have been found to arrestthe cell cycle at the G2/M checkpoint by inhibition of Aurora-Akinase, resulting in an anti-cancer effect [35]. Re(CO3) complexes(Fig. 3, complexes 2, 3, 4) can inhibit the cell cycle progression ofthree pancreatic cell lines (HPA-II, ASPC1, and CFPAC). The inhibi-tory characteristics are attributed to the presence of the Re(CO)3fragment, which is generated by decomposition of the complex

Fig. 4. Timelaps micrographs of the ReZolve-ER labelling the ER within PNT2 cells; an enlarged inset image in each pannel shows the nuclear phagosome forming at theperiphery of the nucleus over the time course. Scale bar: 10 mm [13]. Copyright � 2016, reprinted by permission of John Wiley & Sons, Inc.
Fig. 5. Passive diffusion allows ReZolve-ERTM to be applied to and removed from the cell. Micrographs: (A) 10 min incubation with ReZolve-ERTM; (B) 3� wash with PBSremoving ReZolve-ERTM; (C) 10 min incubation with ReZolve-ERTM after 1 h treatment with DTT; (D) 3� wash with PBS removing ReZolve-ERTM, (E) 10 min incubation withReZolve-ERTM after 2 h treatment with DTT (Scale bar = 10 lM).
238 T.A. Gillam et al. / Coordination Chemistry Reviews 375 (2018) 234–255
due to the poor stability of the carbene ligand. The important nextsteps for investigating these compounds will be to assess their tox-icity to non-cancerous cells and if they can be successfully used toimage cells, simultaneously to inhibiting proliferation.
2.2. Visualising endoplasmic reticulum, and related processes
The live cell imaging application of neutral Re(I) tetrazolatocomplexes has been further showcased in the development of anendoplasmic reticulum (ER) localised fac-[Re(CO)3(1,10-phenanthroline)4-pyridyltetrazolate] complex, herein referred to asReZolve-ERTM [13]. This complex shows a defined and highly speci-fic labelling of the ER and has been used to visualise (real-time) livecell events at the nuclear membrane of PNT2 cells (human
epithelial prostate derived cell line) (Fig. 4) and ER, which arebelieved to be associated with trafficking into and out of thenucleus [13]. This is an excellent example of how optical imagingagents that are suitable for time-course imaging (resistant to pho-tobleaching) of live cells can reveal fleeting and dynamic processesat the subcellular scale. The ability to visualise nuclear events inreal-time may reveal changes in metabolic activity in states of dis-ease. Visualising these events would be otherwise unachievableusing imaging agents that require cell fixation or that are proneto photobleaching.
The mechanism that drives the localisation of ReZolve-ERTM tothe ER is yet to be fully elucidated, however it is believed thatit’s lipophilicity (as it shares key structural elements withReZolve-L1TM), may be key in its observed staining pattern [13].

Fig. 6. Staining of the ER and lipid droplets by neutral cyclometallated complex 5[40]. Copyright � 2017, image adapted and reprinted by permission of JohnWiley &Sons, Inc.
T.A. Gillam et al. / Coordination Chemistry Reviews 375 (2018) 234–255 239
Uptake of ReZolve-ERTM is reported to occur by passive diffusion[13]. It was also noted that when labelled cells were washed withfresh cell culture media, a weaker intracellular signal was detected.This suggested that the complexes were in fact influenced by a dif-fusion gradient. As such ReZolve-ERTM may be useful as an ‘‘in–out”probe, that may be useful for drug screening. For example, asshown in Fig. 5 wherein the ER is stained with Rezolve-ERTM andimaged, the cell is then washed of the stain, treated with dithio-threitol (DTT) (a compound that induces ER stress), then staineda second time. Endoplasmic stress, is associated with the develop-ment of liver disease and is a marker of several cancers, includingprostate cancer [36,37]. As such complexes which reveal abnormalstructures in the endoplasmic reticulummay offer significant diag-nostic benefit.
We have also achieved staining of the ER in H9c2 cardiomy-oblasts with neutral cyclometalated Ir(III) complexes (Fig. 6, com-plex 5); some lipid droplet staining could also be observed. This isnot unexpected as the ER is the predominant site of de novo lipidsynthesis within the cell and is involved in the formation of lipiddroplets [38]. In contrast to prior reports of cationic luminescentiridium complexes that stain the ER, these neutral ER staining com-plexes were found to be relatively non-toxic and did not perturbthe cellular environment. Cationic iridium complexes have beenassociated with stimulating the efflux of Ca2+ ions from the ER intothe cytosol, resulting in apoptosis [39]. Imaging with cytotoxicagents results in perturbation of the natural cellular environment,skewing the observation of normative cellular structure and func-tion. Therefore, these neutral tetrazolato complexes present anexcellent class of cyclometalated Ir(III) complex for the imagingof live H9c2 cells [40].
2.3. Imaging and therapeutic activity of metal ion complexes withinmitochondria
The essential role of mitochondria in cellular health cannot beunderstated. Mitochondria play a critical role not only in energyproduction but also in calcium ion signal transduction, reactive
oxygen species (ROS) production, enzyme production (essentialto the assembly of haem) and thermogenesis to name but a fewcritical cellular processes [41]. Dysfunction of this organelle consti-tutes a significant detriment to healthy cellular function and meta-bolism. In fact the mitochondria are crucial mediators oftumourigenesis, responding dynamically to meet the elevatedmetabolic demands of malignancy [42]. The ability to observemitochondrial arrangement and function within live cells is there-fore highly valued as an important goal for metabolic imaging. Thissection will review recent metal ion complexes that enable mito-chondrial imaging and complexes that target the mitochondriafor therapeutic applications. Metal ion complexes that are sensitiveto mitochondrial ROS will be discussed in Section 4 of this review,wherein ROS responsive complexes are reviewed.
The New group from the University of Sydney, has recentlydeveloped a ligand system which responds as a ratiometric sensorof the mitochondrial copper pool [43]. The design of the ligand isbased on two organic fluorophores, an indolinium and a coumarintethered by an alkyl linkage. The indolinium fluorophore experi-ences quenching in the presence of copper while the coumarin flu-orescence remains constant. This is achieved by conjugation of ahemicyanine functionality that coordinates labile Cu(I) ionsquenching the 600 nm signal. Intracellular copper ion concentra-tion is closely associated with cellular redox status, and is impli-cated in many metabolic disorders. Their studies have revealedthough the application of this probe that cisplatin disrupts theaccumulation of copper within the mitochondria in treated cells.This probe demonstrates how sensing within the mitochondrialenvironment particularly looking at metal ion concentration canreveal aspects of the mechanisms underpinning the therapeuticactivity of metal ion complexes such as cisplatin.
In association with the aforementioned ER lipid droplet stain-ing, neutral cyclometallated Ir(III) complexes referred to in Sec-tion 2.2, several cationic Ir(III) complexes have been investigatedthat label the mitochondria. One example is, complex 6 (Fig. 7)shown in H9c2 cardiomyoblasts with mitochondrial co-stainingby Mitrotracker�Red [40]. Each of the cationic complexes withinthe series exhibited selective accumulation within the mitochon-dria, with the exception of one complex that localised to the ERand lipid droplets. It is suggested that the cationic charge of thecomplexes anchor them within the mitochondrial membrane, pre-sumably due to the transmembrane potential, and as such this maybe a contributing factor to the localisation of these cationic com-plexes. This would be consistent with examples of emissive probescoupled to lipophilic polyaryl cationic phosphonium salts, whichare known to localise within mitochondria [44].
A comparison of the cationic and neutral luminescent Ir(III)complexes developed by Caporale et. al. highlights some key pointsin the assessment of metal complexes for live cell imaging [40].Each of the cationic complexes was notably more lipophilic thantheir neutral counterparts with log D values in the order of 0.6–1.87, however it is not evident that the lipophilicity of the com-plexes had a direct bearing on the efficacy of cellular uptake orlocalisation [18]. The use of ICP-MS was necessary to correlate flu-orescent intensity from the cell (and in some cases account for thetoxicity of a complex) with the total Ir(III) concentration of each ofthese complexes in cells. The large variation in quantum yield andoverall photostability in these structurally related compoundsmeant that this was the only viable method to account for differ-ences in staining. As such the use of ICP-MS should be considereda standard practice as a means of determining quantitative cellularuptake of complexes. For instance, within this study one exceptionwas noted for a cationic complex which seemingly appeared to beless cytotoxic, more akin to the neutral series. In this case the care-ful analysis of the ICP-MS data revealed a low cellular uptake ofthis complex, explaining the minimal toxicity. This further high-

Fig. 7. Mitochondrial staining of cyclometalated cationic Ir(III) complex 6 in live H9c2 cells. Costaining micrographs show mitochondria staining by Ir(III) complex (green)and MitoTracker�Red (red). Merged image shows strong co-localisation of both stains as an orange colour. Scale bar: 10 mm [40]. Copyright � 2017, image adapted andreprinted by permission of John Wiley & Sons, Inc.
Fig. 8. Structures of metal ion complexes that target the mitochondria and elicit cytotoxic effects [47,49,50,52].
240 T.A. Gillam et al. / Coordination Chemistry Reviews 375 (2018) 234–255
lights that an interplay of various uptake and trafficking mecha-nisms may determine the internalisation and localisation of metalion complexes.
Each of the cationic complexes exhibit an elevated cytotoxicityin comparison to their neutral counterparts; however, the toxicityof these complexes is an unfortunate trade off attributed to theirlocalisation within the mitochondria. Internalisation of cationiccomplexes within the mitochondria may disrupt membrane poten-tial, effecting critical energy production pathways resulting in tox-icity. The membrane potential of the mitochondria (negativeinside) is essential to the energy production capabilities of theorganelle [45]. Membrane potential is maintained by efflux of pro-tons out of the mitochondria, creating a charge differential and
proton gradient, which affects the influx of cations, such as Ca(II)[45]. Influx of cationic metal ion complexes may depolarise themitochondrial membrane inciting mitochondrial malfunction andas a result, disruption of cellular energy production. For live cellimaging and tracking applications the toxicity is unwanted; how-ever if complexes can be designed to selectively target diseasedcells, the toxicity can become advantageous for therapeuticapplications.
Emissive metal ion complexes that selectively target and loca-lise in the mitochondria have been a significant focus for the devel-opment of therapeutic compounds. As the mitochondria is a vitalorganelle for cell metabolism and proliferation, it is an excellenttherapeutic target, particularly for cancers where mitochondrial

Fig. 9. Potent anticancer Zn(II) complex and the proposed pathway of action.Reprinted with permission from [60], Copyright � 2016, American ChemicalSociety.
T.A. Gillam et al. / Coordination Chemistry Reviews 375 (2018) 234–255 241
activity is generally elevated [46]. Yang et al. have presented anexcellent example of a therapeutic metal ion complex, namely acationic Re(I) tricarbonyl polypyridyl, with a chloromethylpyridylligand (Fig. 8, complex 7) [47]. Localisation of the complex in themitochondria of A549 cells (a cisplatin resistant human adenocar-cinoma cell line) was achieved due to the charge and highlipophilicity of the complex, which initially facilitated cellularimaging. The inclusion of the chloromethylpyridyl ligand enabledimmobilisation of the complex within the mitochondria throughcovalent reaction with free thiols present on mitochondrial pro-teins. This immobilisation resulted in enhanced cytotoxicity (dueto the prolonged retention of the complex), whereby the complexinduced a series of mitochondrial responses, including inhibitionof respiration, reduced ATP production and an increase in ROS.The authors monitored the changes in these biological pathwaysusing the complex because the phosphorescence of the complexwas oxygen dependent, which could therefore be used to directlymonitor mitochondrial respiration. Importantly, the complexshowed selective action toward cancer cell types.
A series of mitochondrial-targeting cyclometalated Ir(III) com-plexes, with varying degrees of lipophilicity have also been devel-oped as photodynamic, therapeutic agents [48]. Fluorescenceimaging identified localisation of the complexes in the mitochon-dria of HeLa (human cervical cancer derived cell line) and A549cells (human lung carcinoma epithelial derived cell line), with sim-ilar mechanisms of cytotoxicity observed (e.g. elevated ROS,release of apoptosis factors). Importantly, one of the describedcomplexes (with highest lipophilicity and cytotoxicity) could beused to track mitochondrial damage in real time (Fig. 8, complex8). Due to the long-lived triplet excited state of the Ir(III) com-plexes, the photosensitised generation of singlet oxygen (1O2)was possible, which increased the cytotoxicity of the complexesby up to three orders of magnitude. An extension of this workhas involved the conjugation of a coumarin appendage to Ir(III)complexes (Fig. 8, complex 9), with mitochondria targeting, cyto-toxic action and real time image tracking of cell fate again possible[49]. The inclusion of the coumarin was shown to enhance the pho-tophysical properties of the complexes and to increase the cytotox-icity of the complexes. Recent work has extended the therapeuticapplication of these cyclometalated Ir(III) complexes (Fig. 8, com-plex 10) to include inducing and tracking of mitophagy [50]. Defec-tive mitophagy capacity has been associated with conditionsincluding Parkinson’s and cardiovascular disease as well as cancer[51]. In this study, Ir(III) complexes with an ancillary dimethyl-biimidazole ligand were demonstrated to localise in the mitochon-dria of different cancer cell lines. Stimulating previously men-tioned cytotoxicity mechanisms, the phosphorescent complexescould then be used to image the damaged mitochondria and trackmitophagy by time-lapse fluorescence microscopy.
A different therapeutic strategy has been developed by Tsoet al., who synthesised Ir(III) polypyridine complexes with a poly-ethylene glycol (PEG) moiety attached through a photocleavablenitroveratryl linker (Fig. 8, complex 11) [52]. Kept in the dark,PEG appended complexes showed low cytotoxicity, due to the abil-ity of the PEG to minimise interactions with cellular proteins andorganelles. The biocompatibility of the complexes in the dark iselevated by increasing PEG chain length, with cell viability follow-ing the order of PEG (10 kDa) > PEG (5 kDa) > triethylene glycol(TEG). Irradiation at 365 nm triggered the release of the PEG chainand a significant increase in cytotoxicity was observed, even for ashort irradiation time of 5 min (cell viability decreased from 85%to 13%). The intrinsic luminescence of the iridium complexesallowed fluorescence microscopy imaging of the exposed HeLacells, which indicated localisation in the mitochondria. The mech-anism of cytotoxic action was not explored in detail, only to ruleout significant ROS generation as a cause. Whether localisation in
the mitochondria occurred pre- or post-365 nm irradiation (andrelease of the PEG chain) was not determined.
The diverse mechanistic functions of the mitochondria in dis-ease states, let alone in normative cellular function are as yet underexplored. The mitochondria are adaptive in their response to cellu-lar stress and starvation, in part this is why they are so critical tocancer cell growth and survival strategies [53]. As such the devel-opment of metal ion complexes which probe the function of mito-chondria in particular disease states may help address some of thegaps in knowledge that are still present. This will not only inform agreater understanding of the diverse functions of mitochondria,but also can lead to the development of novel therapeutics, whichtarget the tumourigenic capabilities of this organelle.
2.4. Investigation of lysosomal function with metal ion complexes
Lysosomes fulfil many functions within the cell and play a crit-ical role in the transduction of metabolic signaling pathways [53].Abnormality in the arrangement and function of lysosomes can bean indicator of malignancy and therefore this organelle presents asa promising target to monitor metabolic conditions [54]. The lyso-some is closely associated with the process of autophagy, a seriesof events that occur to recycle cellular machinery, enacted whenthe cell is under metabolic stress [55]. Metabolic stress is a markerof tumourigenesis, and thus lysosomal abnormality is associatedwith aggressive malignancy [55]. Complexes that label the lyso-some or report key reactive species within the lysosomal environ-ment are capable of revealing insights into the mechanismsunderpinning various cancers, immune/inflammatory disordersand other metabolic diseases [56,57]. Recent work in the develop-ment of metal ion complexes as lysosomal imaging agents has pre-dominantly focussed on complexes that enable detection oflysosomal ROS [58,59]. Lysosomal ROS sensitive complexes are dis-cussed in Section 4.
A recent study out of the University of Sydney and the Univer-sity of Queensland, by Stacy et al. has demonstrated the mecha-nism by which metal ion complexes that localise to the lysosome

Fig. 10. Structure of the DNA intercalative MRI contrast agent developed byRendina [30].
242 T.A. Gillam et al. / Coordination Chemistry Reviews 375 (2018) 234–255
can be used as therapeutics, while also shedding light on the pre-viously unknown mechanisms of certain anticancer drugs [60]. Inthese studies, a series of zinc(II)–thiosemicarbazone complexeswere synthesised as fluorescent analogues to the potent anticancerdrugs di-2-pyridylketone thiosemicarbazone and 2-acetylpyridinethiosemicarbazone. The complexes were incubated with severaltumour cell types, where the Zn(II) complexes were more toxicthan the non-complexed ligands. Importantly, fluorescence imag-ing revealed that these complexes localise in the lysosomes in allcell types, a mechanistic aspect that could not be directly con-firmed when using non-fluorescent analogues. Further mechanisticstudies, again made possible by the fluorescence properties of thecomplexes, also revealed that the Zn(II) complex could transmetal-late with intracellular Cu(II) ions to form redox active Cu(I) com-plexes (Fig. 9). Subsequent generation of ROS induced lysosomalmembrane permeabilisation, leading to cell death.
2.5. Imaging and therapeutic activity of metal ion complexes withinthe nucleus
The nucleus is the double membrane bound organelle that isthe site of DNA synthesis, protein regulation, and governs manycellular functions by responding dynamically to cellular physiolog-ical and metabolic demands. Altered nuclear structure or beha-viour is an indicator of metabolic changes, in fact an enlargementof the nucleus, changes in shape and chromosomal arrangementare key indicators for cancer diagnosis [61]. As such, imagingagents that enable visualisation of the nucleus or sub-nuclearstructures are beneficial to the study and diagnosis of cancers.
The Rendina group of The University of have labelled thenucleus of A549 human lung carcinoma cells using a Pt(II)–Gd(III) diad complex [30]. The complex is structured with a Gd(III)–diethylenetriamine-pentaacetic acid ligand and is conjugated to aPt(II) 2,20:60,200-tertpyridine complex (Fig. 10, complex 12). ICP-MS analysis revealed that the accumulation of this complex wasone order of magnitude higher in tumour cells, indicating atumour-selective uptake capability, similar to that of other nuclearstaining Pt(II) complexes [62]. This is a particularly useful factor forthe development of MRI contrast agents. The planar polypyridylligands of the Pt(II) centres enable interaction with DNA withinthe nucleus by means of intercalative binding, whilst the Gd(III)metal centre allows this complex to operate as an MRI contrastagent [30]. X-ray fluorescence confirmed the localisation of thecomplex to regions of high zinc and phosphorous density, consis-tent with nuclear staining. The molar ratio of Pt:Gd was also noted
to be 2:1 in vivo supporting that the complex remains stable withinthe cellular environment.
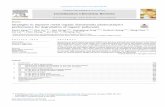
There are some recently described examples of metal ion com-plexes, such as Pt(II), and Ru(II) complexes which permit stainingof the nucleus of live cells with low-cytotoxicity [62,63]. However,many metal ion complexes that localise within the nucleus exhibita higher level of cytotoxicity, often as a result of their ligands inter-calating with DNA or binding with RNA. However, this cytotoxicityhas resulted in positive outcomes, namely the development ofnovel anti-cancer drugs as well as antimicrobial agents [64,65].Therapeutic metal ion complexes with imaging and antibacterialapplications have arisen from the collaborative efforts of Dr.Richard Keene from The University of Adelaide and Prof. Grant Col-lins of the University of New South Wales, where emissive com-plexes have been used to visualise eukaryotic cells and killbacteria. Amongst this work, dinuclear ruthenium complexes havebeen shown to disrupt cytoplasmic and mitochondrial membranesin eukaryotes, yet they exhibit enhanced toxicity toward bacteria[66]. As such these complexes present as promising anti-microbial agents. Previous investigations into oligonuclear poly-pyridyl Ru(II) complexes by Keene and Collins have indicated theirantimicrobial efficacy, while demonstrating 100-fold lower cyto-toxicity to eukaryotic cells [67]. Further studies of cellular uptake,localisation and potential for selective cytotoxicity to cancer cellssupport the effectiveness of this class of complexes as therapeuticagents [68]. The efficacy of platinum based anti-cancer drugs suchas cisplatin promoted the potential application of metal ion com-plexes as therapeutic agents [69]. Keene and Collins have alsodeveloped di-, tri- and tetra-nuclear polypyridyl Ru(II) complexes(Fig. 11, complexes 13, 14, 15 respectively) as anti-cancer agents,which were incubated with cancerous and non-cancerous celllines. Of this series, the tetra-nuclear complex 15, displayed thehighest cytotoxicity, followed by tri- then di-nuclear species, andall complexes showed elevated cytotoxicity within the cancer cellline and displayed significantly higher toxicity again, to bacteria.Localisation in the nucleus and in particular, the RNA rich nucleo-lus of the eukaryotic cells could be visualized by confocal micro-scopy. This series of compounds present as interesting candidatesfor antimicrobial and cancer therapy applications.
Metal ion complexes which localise to specific organelles, notonly enable high resolution visualisation of cellular morphology,they often provide critical information as to metabolic behaviourand environment within the organelle. Organelles fulfil manydynamic, interactive functions within the cell, many of these func-tions have not yet been fully characterised. The application ofimaging technologies afforded by the development of metal ioncomplexes shall undoubtedly contribute to the advancement inunderstanding the intricate behaviour of subcellular structures.With this technology biologists will be able to identify abnormali-ties in cellular function and underpin the diverse mechanisms atplay in many significant diseases.
3. Metal ion complexes with conjugated targeting andfunctional motifs
Metal ion complexes that selectively localise to specific orga-nelles are useful to visualise disease states and associated cellularfunctions; however, to explore cellular mechanisms in greaterdetail a different type of complex is required. The design and syn-thesis of metal ion complexes with specific biomolecule or biorel-evant appendages has gained considerable research interest [70].Complexes with conjugated peptide, protein, metabolite or carbo-hydrate structures have been developed, with the goal of incorpo-rating the biolabelled probe into complex cellular pathways. Suchprobes have been used to directly label and track specific receptor
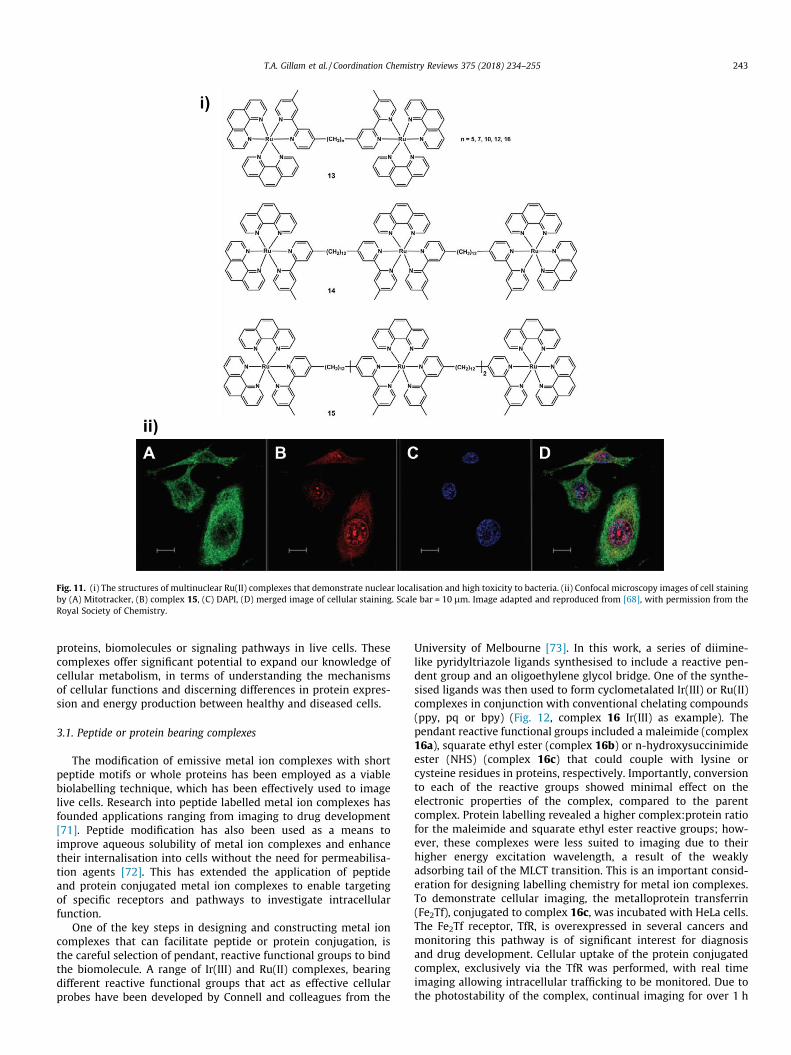
Fig. 11. (i) The structures of multinuclear Ru(II) complexes that demonstrate nuclear localisation and high toxicity to bacteria. (ii) Confocal microscopy images of cell stainingby (A) Mitotracker, (B) complex 15, (C) DAPI, (D) merged image of cellular staining. Scale bar = 10 lm. Image adapted and reproduced from [68], with permission from theRoyal Society of Chemistry.
T.A. Gillam et al. / Coordination Chemistry Reviews 375 (2018) 234–255 243
proteins, biomolecules or signaling pathways in live cells. Thesecomplexes offer significant potential to expand our knowledge ofcellular metabolism, in terms of understanding the mechanismsof cellular functions and discerning differences in protein expres-sion and energy production between healthy and diseased cells.
3.1. Peptide or protein bearing complexes
The modification of emissive metal ion complexes with shortpeptide motifs or whole proteins has been employed as a viablebiolabelling technique, which has been effectively used to imagelive cells. Research into peptide labelled metal ion complexes hasfounded applications ranging from imaging to drug development[71]. Peptide modification has also been used as a means toimprove aqueous solubility of metal ion complexes and enhancetheir internalisation into cells without the need for permeabilisa-tion agents [72]. This has extended the application of peptideand protein conjugated metal ion complexes to enable targetingof specific receptors and pathways to investigate intracellularfunction.
One of the key steps in designing and constructing metal ioncomplexes that can facilitate peptide or protein conjugation, isthe careful selection of pendant, reactive functional groups to bindthe biomolecule. A range of Ir(III) and Ru(II) complexes, bearingdifferent reactive functional groups that act as effective cellularprobes have been developed by Connell and colleagues from the
University of Melbourne [73]. In this work, a series of diimine-like pyridyltriazole ligands synthesised to include a reactive pen-dent group and an oligoethylene glycol bridge. One of the synthe-sised ligands was then used to form cyclometalated Ir(III) or Ru(II)complexes in conjunction with conventional chelating compounds(ppy, pq or bpy) (Fig. 12, complex 16 Ir(III) as example). Thependant reactive functional groups included a maleimide (complex16a), squarate ethyl ester (complex 16b) or n-hydroxysuccinimideester (NHS) (complex 16c) that could couple with lysine orcysteine residues in proteins, respectively. Importantly, conversionto each of the reactive groups showed minimal effect on theelectronic properties of the complex, compared to the parentcomplex. Protein labelling revealed a higher complex:protein ratiofor the maleimide and squarate ethyl ester reactive groups; how-ever, these complexes were less suited to imaging due to theirhigher energy excitation wavelength, a result of the weaklyadsorbing tail of the MLCT transition. This is an important consid-eration for designing labelling chemistry for metal ion complexes.To demonstrate cellular imaging, the metalloprotein transferrin(Fe2Tf), conjugated to complex 16c, was incubated with HeLa cells.The Fe2Tf receptor, TfR, is overexpressed in several cancers andmonitoring this pathway is of significant interest for diagnosisand drug development. Cellular uptake of the protein conjugatedcomplex, exclusively via the TfR was performed, with real timeimaging allowing intracellular trafficking to be monitored. Due tothe photostability of the complex, continual imaging for over 1 h

Fig. 12. Ir(III) complex 16 has been synthesised to enable different bioconjugation chemistry of maleimide (16c), squarate ethyl ester (16b) and NHS ester (16c) [73]. Eu(III)complex 17 with pendent NHS ester has enabled antibody labelling [74].
Fig. 13. (i) Structure of alkyne modified Re(I) complex 18 and subsequent peptide clicked complex 19. (ii) Confocal images show the difference in HeLa cell uptake of thecomplex with and without conjugated HIV-1 Tat peptide. (A) Untreated cells, (B) complex 18 at 100 lM, (C) complex 19 at 20 lM. Image adapted and reprinted withpermission from [75], Copyright � 2014, American Chemical Society.
244 T.A. Gillam et al. / Coordination Chemistry Reviews 375 (2018) 234–255

Fig. 15. Structure of Eu(III)–DTPA C3a conjugated complex 22. Two peptidefragments were individually incorporated via peptide synthesis to complete theC3a protein [77].
T.A. Gillam et al. / Coordination Chemistry Reviews 375 (2018) 234–255 245
was possible, while in comparison Fe2Tf labelled with an organicfluorophore showed a rapid and substantial decrease in emissionintensity. Real time imaging showed the intracellular movementof the complex (dictated by the Fe2Tf), to the early endosomes,prior to release of the iron, following the expected action of thenative protein.
Research out of Macquarie University has shown that covalentattachment of an emissive Eu(III) metal complex to an antibodyusing similar (NHS ester) chemistry, as a practical method to fluo-rescently label antibodies [74]. In this example, a tetradentate b-diketonate Eu complex incorporating a tetraethylene glycol spacerwith a pendent NHS ester to allow antibody conjugation, was syn-thesised (Fig. 12, complex 17). The synthesised complex has supe-rior photophysical properties (2.5 times enhancement inluminescence) to previous generation complexes and the conju-gated antibody was shown to outperform antibody labelled withFITC (�7 times increase in signal-to-noise ratio (SNR)). Time-gated luminescence (TGL) imaging of prostate cancer cells wasdemonstrated using both direct and indirect immunostaining tech-niques, with the complex conjugated to the primary antibody,streptavidin or secondary antibody respectively. The use of themetal ion complex allowed TGL imaging, greatly reducing the noisefrom autofluorescence and improving the SNR.
The ability of a conjugated peptide sequence to enhance the cel-lular uptake of a luminescent Re(I) tricarbonyl polypyridine com-plex has been demonstrated in a collaboration betweenresearchers at Monash University and two Swiss institutes [75].Here, a myristoylated analogue of the HIV-1 Tat peptide was con-jugated to the metal ion complex via click coupling to an alkynemodified pyridine ligand (Fig. 13, complex 18, 19). Such a lipidatedpeptide has been shown to enhance cell permeability, which wasindeed observed (Fig. 13), with an increased uptake of the complexby HeLa cells. The study noted an increase in the logD of the pep-tide conjugated complex from the parent (�0.36 to 0.86), whichcould also be associated with the enhanced cellular uptake. How-ever, it should be mentioned that quantitative intracellular con-centration of the complexes was not evaluated (e.g. by ICP-MS).The observed increased intracellular concentration resulted in anIC50 value of 13 lM, comparable to cisplatin. The parent com-plex showed minimal cell uptake and hence lower cytotoxicity
Fig. 14. Structures of Tb(III) complexes 20 and 21 that are prepared by the click conjugat
(IC50 = 29.9 lM). This strategy presents as an effective method toincrease the cytotoxicity of metal ion complexes, in the contextof treating metabolic diseases such as cancer. This study did notinvestigate selectivity of the complex to healthy or diseased cells,which must be resolved to develop effective therapeutics.
O’Malley and collaborators from Monash University, AustralianNational University and the University of Zurich have also usedclick chemistry to prepare macrocyclic Tb(III) complexes that canfacilitate protein conjugation [76]. The development of complexesthat facilitate highly controllable conjugation chemistry, such asclick, are particularly useful in cases where orientation or con-trolled labelling (1:1 complex:biomolecule ratio) is required. Thisis often unachievable when using NHS ester or maleimide conjuga-tion chemistries. In this case, the clicked lanthanoid complexeswere demonstrated to have increased quantum yield (Fig. 14, com-plex 20), attributed to the lack of water in the inner hydrationsphere, or an increased brightness (Fig. 14, complex 21) due toan increase in the molar absorptivity. The bacterial protein, Escher-ichia coli aspartate/glutamate binding protein, was synthesised tocontain an azide modified, unnatural amino acid, that enabled itto be conjugated to complex 20 and 21. This technique may addversatility to the range and orientation of protein labelling; how-ever, for live cell imaging it may be difficult to take advantage ofthese complexes, due to the high energy excitation light (�300nm) that is required. Incorporation of unnatural amino acids andproduction of proteins by this method is also not readily accessibleand may limit this approach. In the context of studying metabolic
ion of azide bearing compounds to the parent, alkyne functionalised complexes [76].

Fig. 16. Structures of the folic acid (complex 23a) and pteroic acid (complex 23b) labelled Ln(III) metal ion complexes for targeting cell surface receptors [21]. The c, a andbinding sites of the folic acid are identified along with the sole binding site for the pteroic acid.
246 T.A. Gillam et al. / Coordination Chemistry Reviews 375 (2018) 234–255
disease, the incorporation of unnatural amino acids may also alterthe active sites or interactions of proteins within cells. Care mustbe taken to ensure protein interactions studied by this methodare indeed occurring naturally.
In a different approach to synthesise probes for studying pro-tein interactions, Dantas de Araujo et al. from the University ofQueensland developed a Eu(III)–diethylenetriaminepentaaceticacid (DTPA) complex with conjugated human complement ana-phylatoxin (C3a) protein [77]. The C3a receptor is expressed onvarious cell types and is involved in metabolism, inflammationand immunity; however, the interaction of C3a to this receptor isnot well understood [78]. This study described the multistep syn-thesis of the complex, including coupling of a 22 amino acid pep-tide sequence at the N-terminus to the DTPA through amidebond formation and subsequent addition of further peptide frag-ments to complete the 77 residue C3a (Fig. 15, complex 22). DTPAwas selected due to its compatibility with the automated peptidesynthesis process and the sequence was prepared in fragments toensure accurate protein folding. The synthesised protein–Eu(III)complex was shown to behave with similar activity to the nativeC3a and could be used to stimulate C3aR mediated pathways inHEK293 cells. Time-resolved fluorescence was used to study theprotein to receptor interaction in detail and highlight the potentialapplication of such a biolabelled metal ion complex for interrogat-ing biological functions. This method is useful in the case of shortpeptide sequences or smaller proteins that may be produced viaautomated peptide synthesis; however, it is not compatible wheninvestigating larger or quaternary structured proteins.
Peptide and protein labelled metal ion complexes offer a signif-icant range of possibilities for investigating interactions, pathwaysand cellular mechanisms in real time. A range of metal ion com-plexes with different conjugation strategies have been presentedin the recent research, each with advantages and drawbacks. Con-ventional conjugation chemistry such as NHS ester (amide bondformation) is still widely used due to its well understood reactionconditions, from previous bioconjugation research. While this isuseful to advance the field, it is likely that the use of click andbioorthogonal click conjugation will become more commonplace.The intelligent incorporation of the formed trirazole into metalion complexes should enhance the photophysical properties ofthese probes and extend the versatility for in depth investigationof metabolic disease at the cellular level.
3.2. Biolabelling with metabolites
Emissive metal ion complexes have also been appended withother biorelevant species in an attempt to target specific cell recep-tors. The conjugation of folic and pteroic acid to lanthanoid(III)complexes (Fig. 16) has been demonstrated by our research groupat the University of South Australia, where cellular uptake based onthe appended acid species and linker chemistry was investigated[21]. Targeting the folate receptor provides an attractive pathwayfor labelling certain cancer cells due to the overexpression of thisglycoprotein [79]. This investigation in part aimed to determinewhether synthetically simpler fragments of metabolites could beused for targeting or whether the entire compound must be used.There were distinct photophysical differences between complexesbearing folic or pteroic acid. Pteroic acid conjugated complexes dis-played pH sensitive emission, while the emission of folic acid com-plexes was insensitive to pH variations. This was attributed to anincreased separation between the protonated pteridine ring ofthe folic acid and the metal centre, with changes in pH unable toaffect the sensitization process of these complexes. This is signifi-cant for labelling cancer cells, as pH is known to vary in diseasedtissue and cells [80]. Emission from a pH insensitive probe willtherefore correlate to concentration and not the local environment.The length of the linker (represented in Fig. 16, complex 23, wheren = aliphatic chain length of 1 or 5) was shown to have minimaleffect on cellular uptake; however, longer linkers produced adverseeffects on the photophysical properties of the complex due toincreased separation between the metal ion centre and the conju-gated aromatic residue. The study revealed an enhanced uptake offolic acid (complex 23a), over pteroic acid (complex 23b) appendedcomplexes. Significantly, folic acid complexes with shorter linkersshowed higher levels of internalisation. Increased uptake for aprobe of this nature contrasts with previous studies and may sug-gest a more simplistic synthesis strategy is favourable. The conju-gation of folic acid to a Eu(III) complex has been further exploredby Quici et al. who investigated the effect of binding the folic acidat different locations [81]. The uptake of the prepared complexesinto cells expressing high, medium, low and negative levels offolate receptors was studied. A clear trend was observed regardingthe binding site of the folic acid to the complex, with conjugationvia the c carboxyl providing the highest uptake, followed by conju-gation at the a carboxyl and finally through click conjugation via

Fig. 17. Sugar free (24) and glucose (25) and galactose (26) appended cyclomet-alated Ir(III) complexes, synthesised to investigate GLUT mediated cellular uptakeprocesses [85].
T.A. Gillam et al. / Coordination Chemistry Reviews 375 (2018) 234–255 247
the nitrogen indicated in the folic acid structure (Fig. 16). Compet-itive uptake assays (using high expression folate receptor cells)between folic acid and folic acid-complex (c position), showed cellinternalisation was governed by interaction with the folate recep-tor. No uptake was observed in the folate receptor negative cellsand the complex exhibited minimal cytotoxicity. This work alsoinvestigated the fate of the complex after internalisation, by recov-ering the folic acid-complex (c position) 24 h post internalisation.HPLC revealed that the complex remained intact and no hydrolysisof the amide bond had occurred. This is an important finding andmay guide attachment chemistry depending on the required appli-cation of the complex.
3.3. Carbohydrate appended complexes
Carbohydrate appended metal ion complexes are some of theearliest examples of biolabelled metal ion imaging agents and havebeen extensively studied by the likes of Lo and Gottschaldt [82,83].Our research group is also active in this space, with one such exam-ple of research aimed at lanthanoid metal ion complex synthesisfor targeting the mannose receptor [84]. Metal ion complexes with
pendant carbohydrate groups are attractive as cellular tracking andimaging probes, due to the biological dependency on carbohydratespecies for energy production, signaling and synthesis. The abilityto track and visualise changes in the uptake, localisation andbreakdown of carbohydrates could afford significant insight intothe metabolic state of live cells.
A series of cyclometalated Ir(III) complexes appended witheither glucose or galactose species have been synthesised by Lawet al. and their cellular uptake investigated [85]. These complexesentered HeLa cells to some extent, with a mechanism of internali-sation that was difficult to define. Of the complexes investigated,the highest internalisation was observed for a sugar free complex(Fig. 17, complex 24), which was attributed to its higher logP(2.59) than the sugar appended complexes. Reasons to accuratelydescribe the amount of internalised complexes are still underinvestigation; however, this raises an important issue as to whichmechanism governs cellular internalisation and what are theappropriate ligands to enable specific uptake (based solely on thebiolabel interaction). Uptake via the glucose transporter (GLUT) 1and 4 pathways was extensively investigated using complex 25and 26 (Fig. 17), with higher internalisation via this mechanismobserved for the D-glucose appended complex (25). It was observedthat the uptake of this complex could be competitively inhibited byaddition of free D-glucose and 2-deoxy-D-glucose, while uptakewas increased upon stimulation with insulin or 17b-estradiol.These results indicate that the glucose appended complex hasthe potential to monitor sugar uptake and metabolic activity incells; however, non-specific internalisation of other sugar modifiedcomplexes through passive and active pathways remains a signifi-cant issue. Deciphering the differences in uptake between com-plexes bearing different sugars and with different ligandsremains the biggest challenge.
Palmioli et al. has investigated the cellular uptake of dinuclearRe(I) complexes, with different mono and tri glyco-conjugates[86]. Time lapse fluorescence imaging indicated an efficient uptakeof specific glyco-conjugates into HeLa cells, with most rapid uptakeobserved for the more hydrophilic complex (Fig. 18, complex 27).The internalisation mechanism was not determined in this study;however, an interesting result was the absence of fluorescencefor the tri glyco-conjugated complex (Fig. 18, complex 28). Theauthors noted the poor photophysical properties of this complex,which they attributed to the formation of micellular structuresbased on the amphiphilic structure of complex 28. Such an eventwould bring the Re(I) centers into close proximity resulting in tri-plet–triplet annihilation processes and radiation-free decay; thiswas supported by a significant non-radiative decay constant (66� 105 s�1).
Due to the increased energy requirements and overexpressionof certain sugar transporters, cancer cells have been a significantfocus for detection and therapeutic applications of carbohydratemodified metal ion complexes. Fructose has been used to modifya Re(I) polypyridine complex, which showed enhanced uptake bybreast cancer cells compared to both non-cancerous and non-transformed cells [87]. The enhanced uptake was attributed to anoverexpression of the fructose transporter GLUT5 in the breast can-cer cells, with the complex also indicating high cytotoxicity (IC50 =2 lm) upon irradiation. However, a study by Pröhl et al. showed nosignificant cytotoxicity effect by a Ru(I) complex with a chelatingfructose conjugated curcumin ligand [88]. One key difference inthis study was that the chelating ligand was modified with twofructose moieties, suggesting a subtle interplay between complexstructure, internalisation and cytotoxicity. A follow up studydemonstrated negligible difference in uptake and cytotoxicity tobreast cancer cells, between mono-conjugated sucrose and fruc-tose Ru(I) complexes [89]. No evidence for transport via GLUT5pathway was presented, however they did not perform a specific

Fig. 18. Sugar appended metal ion complexes, synthesised to target specific membrane sugar binding and transport proteins [86,90].
248 T.A. Gillam et al. / Coordination Chemistry Reviews 375 (2018) 234–255
free fructose competitive uptake assay. A more comprehensiveinvestigation of internalisation mechanism(s) for this cell line, cov-ering a broad range of sugar appended complexes is warranted. Inthe case of prospective cancer therapy agents, quantification of cel-lular uptake by ICP-MS along with full physiochemical characteri-sation of the complexes must be performed. In the cases whereICP-MS quantification was performed, it was a sugar free complexthat displayed the highest cell uptake, which has been attributed toa higher lipophilicity [87,89].
An anti-proliferative, thioglucose appended compound hasbeen demonstrated byWenzel et al. in the form of a Ru(II) polypyr-idine–Au(I) fragment, binuclear complex [90]. Increased toxicity(compared to cisplatin) to cancer cells was demonstrated bythioglucose tetraacetate modified complexes (Fig. 18, complex 29and 30) with cellular uptake determined to occur via active pro-cesses. Interestingly, uptake via GLUT1 was ruled out by competi-tive studies with free glucose. Fluorescence microscopy showeddifferent localisation of two thioglucose complexes, in the nuclei(Fig. 18, complex 29) and cytosol (Fig. 18, complex 30) respectivelyand DNA studies suggested a different action of toxicity to conven-tional (cisplatin) cancer therapy metal ion complexes. One hypoth-esis offered for the increased toxicity was the improved uptake ofthese complexes by cancer cells; however, no ICP-MS data wasprovided to quantify the intracellular concentration. Unfortu-nately, no data was presented to show selective uptake of the com-plexes by cancer cells only and the exact mechanism of uptake andaction remains unclear.
Without question, carbohydrate appended metal ion complexeshave an important role for investigating cellular metabolism andfor new drug discovery. The known overexpression of carbohy-drate receptors on cancerous cells presents as a highly viablemeans to selectively target cells in the diseased state, for diagnosisand treatment. The focus of future research into carbohydrateappended metal ion complexes should therefore be directed
towards achieving cell internalisation exclusively via specificreceptors. As is apparent in the current research, this is yet to berealised, with inherent complex chemistry also governing cellularuptake. Achieving this may involve a combination of carefullydesigned chemistry, whereby only the conjugated biocompounddirects uptake, or to design assay protocols that force uptake tooccur through this mechanism. For example, controlling theenergy-dependent uptake (by using inhibiting factors or lowertemperature), or by stimulating sugar uptake. The effect that linkerchemistry has on cellular uptake has been discussed in the review,as has the effect of lipophilicity, both of which can be readilyaltered in an attempt to find more selective internalisationpathways.
3.4. Therapeutic compounds conjugated to metal ion complexes
Metal ion complexes with conjugated or coordinating therapeu-tic compounds have recently been investigated as a viable meansto deliver drugs that otherwise display poor pharmacokinetic andpharmacodynamic properties. In these examples, therapeuticagents are linked to a metal ion complex via labile chemistry orthrough coordination with the metal centre, whereby externalstimuli can result in the release of the therapeutic compound. Thistherapeutic release has been achieved by numerous means, includ-ing local environmental stimuli, examples of which will be dis-cussed in the following section. Another release mechanism islight-activated drug delivery, commonly referred to as photo-dynamic therapy (PDT).
The Renfrew group of the University of Sydney has taken advan-tage of PDT to develop novel therapeutic metal ion complexes thatdisplay significantly increased cytotoxicity to cancer cells, over thenon-conjugated drugs. In one example, a Ru(II) phenanthrolinecomplex has been prepared that incorporates the imidazole baseddrug econazole [91]. Due to the presence of the imidazole, this

Fig. 19. Ru(II) complexes with econazole (31, 32) and 6-mercaptoipurine (33) coordinated anticancer drugs [91,93].
T.A. Gillam et al. / Coordination Chemistry Reviews 375 (2018) 234–255 249
compound can chelate directly to the Ru(II) metal centre, withmono and bi drug bearing complexes developed (Fig. 19, complex31, 32). Aqueous stability and photophysical investigations of thecomplexes indicated that complex 32 had favourable propertiesas a therapeutic complex. This is due to water coordination tothe metal centre of complex 31 and loss of emission in aeratedwater. Light-activated release of the econazole from complex 32was achieved by irradiation using 520 nm green light, with oneof the two bound drugs released from the complex. This agreeswith previous studies, where release of a second ligand requiresmuch longer irradiation, or does not occur [92]. Cellular uptakewas achieved in multiple cancer cell lines, with the complex dis-playing cytoplasm localisation. Non-irradiated complexes showedmoderate to low cytotoxicity, however upon irradiation and drugrelease, a significant increase in cytotoxicity was observed. A fur-ther beneficial aspect of this complex, was the loss of fluorescenceupon light-activated drug release, allowing a visual monitoring ofthe process. Unfortunately, no investigation against a non-cancercell line has been performed at this stage.
The Renfrew group has extended the application of the Ru(II)complex by incorporation of the anticancer drug 6-mercaptopurine (6-MP), which is currently used to treat leukemia[93]. In this case, the drug again directly coordinates to the metalcentre in conjunction with other chelating ligands. The effect ofligand chemistry on photo-release of 6-MP was explored in moredetail, where it was found that ancillary ligands had minimal influ-ence on the photo-release of monodentate coordinated 6-MP(Fig. 19, complex 33). Photo-release of bidentate (complex 34)coordinated 6-MP however was significantly influenced by theproperties of the other coordinating ligands [93]. The effects of thisinfluence ranged from a far slower release of the drug, to ligand-solvent substitution, to no evidence of ligand release on irradiation.The careful selection of coordinating ligands has been marked as apotential means to control stability and toxicity of the complexes.The complexes were incubated with MCF-7 cells (human breastcancer derived cell line) and upon irradiation, cells that were incu-bated with complex 33 demonstrated the most significant increasein cytotoxicity. Importantly, in the dark this complex is 20 timesless cytotoxicity than free 6-MP and offers a safer means to deliverthe drug.
The development of metal ion complexes for PDT based on Co(III) have also been demonstrated by Renfrew et al. where varyinglevels of cytotoxicity could be achieved based on the wavelength ofthe irradiating light [94]. In this example, curcumin has been coor-dinated to Co(III) in conjunction with the tris(2-pyridylmethyl)amine (TPA) ligand. Blue light (470 nm) irradiation resulted inreduction of the Co(III) metal centre to Co(II) and subsequentrelease of curcumin, while extended irradiation at this wavelengthcaused photodegradation of the curcumin. Irradiation with green
light (520 nm) caused the curcumin to be released from the com-plex, but did not induce any further degradation to the compound.This difference in response to light activation translated to differ-ences in the cytotoxicity observed in MCF-7 breast cancer cells thathad been incubated with the complexes.
The combination of metal ion complexes coordinated to anti-cancer drugs and PDT presents as a versatile new platform forthe development of safer therapeutics. The ability to focus activat-ing light to specific tissues or regions of the body somewhatnegates the need for specifically selective complexes, howeverthe continuation of this research may result in the developmentof highly targeted and potent anticancer agents.
4. Responsive complexes
One of the rapidly expanding classes of metal ion complexesthat are available in the toolkit for live cell imaging are complexesthat can report on the presence, absence or concentration of speci-fic bio-analytes of interest. Metal ion complexes have long beeninvestigated for their chemo- and bio-sensing abilities, but onlyrecently has this sensing transitioned to live cell applications[95]. It is of significant interest to monitor the presence of certainbiologically active species, in real time, in the intracellular environ-ment, especially in the context of understanding metabolism. Aconsiderable volume of research into metal ion complexes forsensing applications has been around the detection of ROS; how-ever, the scope of analytes has broadened to include small bio-analytes including particularly bio-relevant cations and anions.Researchers have now begun to understand the value of monitor-ing these species in real time, in order to understand the mecha-nisms of various diseases and the significance that certainmetabolites have in disease progression.
4.1. Detecting pathological reactive oxygen and nitrogen species
ROS fulfil crucial functions in many energy production path-ways and secretory mechanisms. However; as previously indicatedthe misrelation or irregular production of ROS produces a plethoraof pathological implications. As such a developing subset of metalion complexes have been emerging as cellular probes of sensingselective ROS and reactive nitrogen species (RNS) [96,97].
Singlet oxygen (1O2) is a toxic ROS that forms the basis of pho-todynamic therapy (PDT) and is considered to be a marker for cellhealth [98–100]. However, this key analyte is difficult to detect dueto its highly reactive nature and thus a brief lifetime (3.5 ls in H2O)[100]. Recently a carbostyril sensitized Eu(III) DO3A complexdeveloped in a collaborative project between the Plush andVoelcker groups of the University of South Australia, has been

Fig. 20. ‘‘Turn-on” Eu(III) complexes 34 and 35 for the visual detection of HOCl[97,102].
Fig. 21. b-Diketonate–Eu(III) complexes 36 and 37 that act as turn ‘‘off” probes forthe detection of HOCl. Complex 36 displays mitochondria localisation, whilecomplex 37 localizes to the lysosome [58].
250 T.A. Gillam et al. / Coordination Chemistry Reviews 375 (2018) 234–255
incorporated into a colourimetric chemosensor for 1O2 [100].Incorporating the Eu(III) complex enabled the sensitive detectionof 1O2 in aqueous media by an ‘‘off–on” response. The complex isfurnished with an anthracene moiety that behaves as a 1O2 trap,resulting in a colourimetric change in emission. The sensing capac-ity of this complex that functions at a neutral pH presents an excel-lent basis for the further development of in vivo lanthanoid basedsensors of 1O2. This same complex has also been attached withinan optically active porous silicon surface through a secondaryamine present in the complex [101]. The development of this func-tionalised 1O2 responsive surface represents an application of ROS
responsive complexes that may form the basis for a point of care‘dipstick’ like diagnostic platform.
Hypochlorous acid (HOCl) has become a ROS of particular inter-est as it is essential for normal immune system function and isderegulated in inflammatory disorders [59,97]. Lanthanoid com-plexes of Tb(III) and Eu(III) baring a [40-(4-amino-3-nitrophenoxy)methylene-2,20:60,200-tertpyridine-6,600-diyl]bis(methylenenitrilo)tetrakis(acetate) ligand have been developed by the Yuangroup from Dalian University of Technology (China), which confera specificity for the detection HOCl over .OH radicals [97]. Thesecomplexes are weakly luminescent due to the presence of an ami-nophenyl PET quencher attached through an acetoxylmethyl ester,which is selectively cleaved by HOCl, imparting an enhancement ofluminescent signal (Fig. 20, complex 34). Further functionalisationof this ligand system has been made by inclusion of a PET quench-ing dinitrophenyl moiety attached via a highly selective HOClcleavable imine linkage (Fig. 20, complex 35) [102]. The resultingcomplex could be used for in vivo imaging. This improved designimparts a further enhancement in HOCl selectivity whilst alsoimproving sensitivity. Complex 35 is non-luminescent due to moreefficient PET quenching, as compared to the weak luminescence ofcomplex 34. Development of this complex has enabled the quanti-tative determination of HOCl in aqueous media and the complexhas been imaged in RAW 264.7 macrophages (murine derivedmacrophage cell line) and Daphnia magna [102].
Although complex 35 provided excellent responses to thedetection of HOCl in live cells, it lacks localisation to particularorganelles that are associated with pathological ROS, such as themitochondria or lysosome. Yuan and colleagues have recentlydeveloped HOCl responsive Eu(III) complexes based on a b-diketonate ligand system [58]. Targeting of the mitochondria andlysosome was achieved by coupling triphenylphosphonium ormorpholine ancillary groups, respectively, onto the aromatic ligandstructure through the formation of a sulfonamide bond (Fig. 21,complex 36 and 37, respectively) [58]. Incorporation of these tar-geting features onto b-diketonate–Eu(III) complexes have allowedfor the visualisation of HOCl with high organelle localization speci-ficity within lysosomes and mitochondria in live cells, exhibiting aHOCl specific ‘‘turn-off” response [58]. This ‘‘turn-off” responseresults from the selective reaction of the diketonate with HOCl, dis-assembling the coordination environment of the sensitizer, by oxi-dation of the coordinating carbonyl groups to carboxylic acids,quenching luminescence. These probes were imaged in HepG2(human hepatoma derived cell line) and RAW 264.7 cell lines aswell as the animal model of Daphnia magna.
Though this work exemplifies the benefits of employing target-ing motifs, realistically a ‘‘turn-on” response is more desirable inapplication. As such, the previously mentioned terpyridine tetrakis(acetate) ligand system that achieves a ‘‘turn-on” response is amore promising lead for the development of responsive HOCl com-plexes. The triphenyl phosphonium targeting system was thereforeapplied to the coordinating appendages of the terpyridine tetrakis(acetate) ligand system, whilst employing a p-nitrophenylquencher in the same way as previously employed for PET quench-ers in this ligand system, by means of an alkyl sulphide bond [44].This mitochondrial targeted complex was highly specific in its’response to HOCl over other ROS, highly emissive, water solubleand permitted high resolution time-resolved imaging of HOCl witha ‘‘turn-on” response, when imaged within the mitochondria of livecells (RAW 264.7 macrophages) and zebrafish [44]. This exempli-fies how revision of a ligand system can result in high analytespecificity, and desirable organelle targeting, with a beneficial‘‘turn-on” response.
Endosomal–lysosomal localised metal ion complexes open awindow of opportunity for biologists to better observe and under-stand the interaction of bioactive species and the precise

Fig. 22. Ir(III) sensor of lysosomal HOCl used to image liver damage in a murine model, and to label the liver of zebrafish. (i) Mouse liver tissue has been imaged following aninduced ischemia (I/R panels) revealing elevated detection of HOCl (high levels of HOCl show as orange in overlayed panels) in comparison to control. (ii) Bright-field (A),luminescence (B), and merged image (C) of a zebrafish following incubation with 100 mm the Ir(III) ferrocene functionalised complex developed by Yuan et al. Image adaptedand reprinted with permission from [59], Copyright � 2017, Elsevier.
Fig. 23. (i) 68Ga complex 38. (ii) PET/CT imaging of tissue hypoxia using complex 33 in (A) CA IX-knockdown and (B) CA IX-expressing shCAIX tumours models in mice.Complex 35 shows strong signal in the bladder and kidneys also, as it is cleared renally without being metabolised. Image adapted and reprinted with permission from [105]Copyright � 2016, American Chemical Society.
T.A. Gillam et al. / Coordination Chemistry Reviews 375 (2018) 234–255 251
involvement of the lysosome in diseases such as cancer. As indi-cated endosomal–lysosomal localisation can be achieved by theincorporation of a targeting group or alternatively by accessingendocytic uptake mechanisms. For example, a Ru(II) complex fur-nished with a cleavable ferrocene quencher (PET disruption ofMLCT emission), has been developed as a diagnostic probe forselectively sensing HOCl in the endosomes-lysosomes of livemacrophages [58]. Where endosomal–lysosomal accumulationwas attributed to the mechanism of uptake occurring mainlythrough caveolae-mediated endocytosis. This complex has demon-strated high biocompatibility and has been imaged in live cells, aswell as in animal models (Daphnia magna and in zebrafish).
A further example of HOCl sensing, by incorporation of a selec-tively cleavable ferrocene quencher, is that of an Ir(III) complexthat has been used to image liver injury in a murine and zebrafishmodel by detection of HOCl [59]. This work stemmed from collab-oration between Dalian University of Technology (China), the KTHRoyal Institute of Technology (Sweden), Woolloongabba hospital
and The University of Queensland (Australia). Imaging with thiscomplex demonstrates its value when applied for whole tissue/organism imaging (Fig. 21). Colocalisation experiments in HepG2cells confirmed the localisation of this complex to the mitochon-dria, confirming the detection was of mitochondrial HOCl, whichis elevated in response to liver injury [59]. This ability to visualiseliver damage could have beneficial applications in the assessmentand study of metabolic diseases that are associated with abnormalHOCl production, such as lung, liver and kidney disease, as well asmyocardial infarction, neurodegenerative disease and cancer [59](Fig. 22).
Hypoxia imaging is a valuable tool for the detection of cancer-ous tissue, and provides useful insight to the design and develop-ment of medicinal interventions [103]. The Donnelly group fromUniversity of Melbourne have made recent advances in the devel-opment of tricarbonyl Tc(I) and Re(I) based hypoxia luminescentand radiolabeled imaging agents [104]. Within the series of com-plexes are Re(I) complexes baring p-nitrophenyl pyridylhydrazone

Fig. 24. Hypoxia responsive cobalt complexes that undergo ligand release in the hypoxic microenvironment of tumour spheroids [109–111].
Fig. 25. A turn ‘‘on” Eu(III) complex 43, for the visual detection of ascorbicacid [113].
252 T.A. Gillam et al. / Coordination Chemistry Reviews 375 (2018) 234–255
functionalised ligands that have demonstrated an elevated uptakewithin hypoxic SH-(SY5Y) (human derived metastatic neuroblas-toma cell line) cells compared to a normoxic control. A nitroimida-zole functionalised 99mTc(I) tricarbonyl complex has proven to bestable in human serum and has been successfully used to imagea human renal cell carcinoma murine xenoplant. One complex inparticular displays tumour specific localization, highlighting thepotential for complexes within this class as tools for clinical diag-nostic SPECT/CT imaging of tumour hypoxia.
The Poulsen group of Griffith University have developed a PETradiotracer for hypoxic tumours based on a sulphonamide func-tionalised 68Ga-DOTA complex (Fig. 23, complex 38) [105]. Thiscomplex is capable of discriminating oxygen levels in tissue, whilebinding to carbonic anhydrase IX (CA IX) exclusively in hypoxiccells. The expression of CA IX is elevated in hypoxic tumours, andit is considered to be a prime histological marker of tissue hypoxia[106–108]. The sulphonamide of the ancillary ligand binds to zincwithin the active sight of CA IX [105]. This complex can be seenlabelling a hypoxic tumour in murine model (Fig. 23), and repre-sents a prospective tool for CA IX positive tumour prognosis.
The Hambley group of the University of Sydney have developedCo(III) complexes that are able to deliver the small fluorescent cou-marin compound, as a model cytotoxin, to the hypoxic core oftumour spheroids [109]. Spheroids are aggregates of tumour cellsthat are highly resistant to permeation by anticancer therapeutics.One complex in a series of compounds developed in this work,Fig. 24 compound 39, has been found to accumulate in hypoxicregions within tumour spheroids (considered to be highlymalignant and difficult to treat). In the hypoxic environment, the
complex is believed to be activated, triggering the release of thecoumarin, by either a reduction or ligand exchange response tohypoxia. The precise mechanism is yet to be elucidated. This exam-ple demonstrates howmetal ion complexes may act as chaperones,escorting a drug to previously inaccessible regions of solid tumourspheroids. Further, careful design of the metal ion complex has alsoenabled targeting tissue hypoxia, as a means for drug release inresponse to local environmental conditions.

T.A. Gillam et al. / Coordination Chemistry Reviews 375 (2018) 234–255 253
Further work out of the University of Sydney has demonstratedhow hypoxic reduction of Co(III)TPA to Co(II)TPA complexes(Fig. 24, complex 40 (as example)) may be monitored within thetumour microenvironment using MRI [110]. The bioreduction ofdiamagnetic Co(III) (MRI-silent) to the detectible paramagneticCo(II) (MRI-active) permits visualisation of the complex at thehypoxic tissue region. This study highlights the potential of Co(III)TPA complexes as candidates for the development of anticancerprodrug therapies.
To demonstrate the use of these complexes as anticancer pro-drugs curcumin, an anticancer drug was coordinated to two Co(III)TPA complexes [111]. Each of these complexes release cur-cumin within the hypoxic tumour microenvironment, via reduc-tion of the complex to Co(II). However, it was noted that changesto the ancillary ligand structure of the complexes has a profoundeffect on the uptake and distribution of the complexes withintumour spheroids. Substitution of coordinated pyridyl ligands,complex 41, for a primary amino group, complex 42, resulted inreduced penetrance into the spheroid. As such it was evident thatthe TPA ligand structure was more suited to the development ofprodrug complexes capable of fully permeating the tumour spher-oid, thus this ligand structure is a better candidate for the develop-ment of prodrugs.
4.2. Biomolecular recognition
Several complexes have been designed to recognise keymetabolically relevant biomolecular species with high specificityin live cells and tissues [112–114]. Detection of these species withresponsive metal ion complexes is generating insights into theaction of particular biomolecules responsible for cell health, andvariations of these biomolecules that may signal metabolicalterations.
Vitamin C (ascorbic acid) has been identified as an essentialnutrient, with roles in metabolic function, with discrepancies inascorbic acid abundance being associated with the progression ofmetabolic disease states such as scurvy [115] and cancer[116,117]. Currently, there are limited tools for the monitoring ordetection of ascorbic acid in living systems. Although there areelectrochemical means for its detection, they commonly sufferfrom interference from other non-specific biomolecules [118]. Flu-orescent detection methods have been developed but are limitedto the detection of ascorbic acid in aqueous media, with low sensi-tivity, and not in vivo. As such, a lanthanoid(III) complex has beendeveloped as a highly selective ascorbic acid sensor based on anovel ligand platform [113]. The ligand (Fig. 25, complex 43) fea-
Fig. 26. Ru(II) complex 45, enabling the visual detection of MGO via a ‘‘turn-on”mechanism [114].
tures a stable free radical, 2,6,6-tetramethyl-1-oxy-4-aminopiperidone ancillary group [113]. The presence of the free radical impartsa quenching effect on the complex: however, in the presence ofascorbic acid the oxygen free radical is reduced selectively(Fig. 25, complex 44). The now protonated complex exhibits a dras-tically enhanced luminescent emission and has been used to imageascorbic acid in real time without background interference in livecells and Daphnia magna using time-gated imaging.
Production of the reactive alpha-ketoaldehyde, methoxyglyoxal(MGO) has been associated with metabolic disorders of the centralnervous system, including Alzheimer’s disease, and other neurode-generative disorders [119,120]. Ru(II) and Ir(III) complexes havebeen developed as ‘‘turn-on” probes for the detection of MGOin vivo [114]. These complexes utilize a 5,6-diamino-phenanthroline ligand that covalently binds MGO, disrupting theelectron transfer of the phenanthroline ligand ensuring efficienttriplet state emission. The Ru(II) variant (Fig. 26, complex 45) ofthese complexes and the ligand’s interaction with MGO (complex46) make these complexes highly biocompatible and have beenused to image MGO with high spatial resolution in RAW cellsand in Daphnia magna, demonstrating their utility as effectivebiosensors.
5. Conclusion and future perspectives
The development of versatile metal ion complexes as imagingagents and therapeutics provide a ‘‘bright light on the horizon”with regard to tackling the burden of metabolic disease on socio-logical and global healthcare systems. Metal ion complexes areuniquely positioned, offering advantageous cross-platform imag-ing capacity, which is highly amenable to the development ofinvestigative and therapeutic compounds. The progression ofmetal ion complexes to biological applications provides an excitingprospect, with current research positioned at the nexus of appliedchemistry and functional biology.
A wealth of knowledge surrounding ligand design has resultedin complexes with specific localisation profiles, metabolite sensingand bio-recognition capabilities [31]. Recent studies have madeexcellent progress in exploring the effects of charge and ligandmodification on cellular uptake and cytotoxicity, elucidating fac-tors that influence the biocompatibility of complexes [40].Although the influence of lipophilicity, charge, ligand specific tar-geting chemistry, and bio-molecular conjugation have been widelyexplored, there remains a degree of ambiguity in the mechanismsof action for metal ion complexes. As such, a more standardisedapproach to characterising complexes and their behaviour in vivoshould be adopted. Accurate quantitation of cellular uptake isrequired in conjugation with peripheral assays that probe themechanisms of cellular internalisation, transport and accumula-tion. Fortunately, metal ion complexes permit a multitude of ana-lytical techniques that can answer these questions. For instance,confocal microscopy can reveal useful information as to the local-isation profile of complexes, but not all complexes are optical emit-ters. In terms of quantifying cellular internalisation, highsensitivity ICP-MS should be used for this analysis. Furtheringthe investigation by detection of complexes in correspondence toother analytes by X-ray fluorescence, FTIR or Raman, for example,provides spatial information as to the localisation of complexesand also reveals aspects of their effect on other analytes withinthe cellular environment. The use of inhibitory and stimulateduptake experiments, which have largely been applied in the evalu-ation of sugar conjugates, is also a necessary measure to determinewhether a bio-conjugated motif is influencing cellular uptake andlocalisation behaviour or if the intrinsic properties of the complexoverride the localisation characteristics of the conjugated motif.

254 T.A. Gillam et al. / Coordination Chemistry Reviews 375 (2018) 234–255
Cytotoxicity of metal ion complexes is a double edged sword inthis field; although undesirable in live cellular imaging, cell deathis a favourable outcome in cancer therapy (so long as it is specificto diseased cells). Many metal ion complexes have demonstratedelevated cytotoxicity compared to cisplatin in diseased systems.Some have even shown considerable promise as antimicrobialagents, with a 100 fold elevation in toxicity to prokaryotic overeukaryotic cells [69]. Perhaps the most advantageous complexesare those that can induce a cytotoxic effect following externalstimulation. Photodynamic therapies are a particularly promisingavenue of metal ion complex application, allowing the selectiveactivation of targeted cell death.
The scope and versatility of metal ion complexes is being rea-lised and a vast amount of new knowledge has been gained fromthe use of metal ion complexes in biological assays. Informationregarding cellular mechanisms, sub-cellular structure and actionsof therapeutics has all been learned using metal ion complexes.This review acknowledges the significant recent advancements inthis field, particularly those made by key Australian researchgroups. The exceptional work of these researchers will undoubt-edly contribute to the glowing future of metal ion complexes andtheir role in the study and treatment of disease.
Acknowledgments
TAG was funded by an Australian Government Research Train-ing Program (RTP) scholarship. MS was funded by an AusIndustry‘‘Researchers in Business” grant. CAB was funded by a FutureIndustry Accelerator Research and Development Voucher Scheme.JLM was funded by a NHMRC Career Development Fellowship(APP1066916). The University of South Australia is kindlyacknowledged for financial support.
References
[1] F.P. Dwyer, E.C. Gyarfas, W.P. Rogers, J.H. Koch, Nature 170 (1952) 190–191.[2] F.P. Dwyer, H.A. Goodwin, E.C. Gyarfas, Aust. J. Chem. 16 (1963) 544–548.[3] C.A. Bader, T. Shandala, E.A. Carter, A. Ivask, T. Guinan, S.M. Hickey, M.V.
Werrett, P.J. Wright, P.V. Simpson, S. Stagni, N.H. Voelcker, P.A. Lay, M. Massi,S.E. Plush, D.A. Brooks, PLoS ONE 11 (2016) 1–24.
[4] E.J. New, A. Congreve, D. Parker, Chem. Sci. 1 (2010) 111–118.[5] J.L. Wedding, H.H. Harris, C.A. Bader, S.E. Plush, R. Mak, M. Massi, D.A. Brooks,
B. Lai, S. Vogt, M.V. Werrett, P.V. Simpson, B.W. Skelton, S. Stagni, Metallomics9 (2017) 382–390.
[6] K.Y. Zhang, H.-W. Liu, M.-C. Tang, A.W.-T. Choi, N. Zhu, X.-G. Wei, K.-C. Lau, K.K.-W. Lo, Inorg. Chem. 54 (2015) 6582–6593.
[7] Y. Chen, R. Guan, C. Zhang, J. Huang, L. Ji, H. Chao, Coord. Chem. Rev. 310(2016) 16–40.
[8] S. Mendis, World Health Organisation: Global status report onnoncommunicable diseases, 2014.
[9] Y. Wang, R. Song, H. Feng, K. Guo, Q. Meng, H. Chi, R. Zhang, Z. Zhang, Sensors16 (2016).
[10] C.A. Bader, R.D. Brooks, Y.S. Ng, A. Sorvina, M.V. Werrett, P.J. Wright, A.G.Anwer, D.A. Brooks, S. Stagni, S. Muzzioli, M. Silberstein, B.W. Skelton, E.M.Goldys, S.E. Plush, T. Shandala, M. Massi, RSC Adv. 4 (2014) 16345–16351.
[11] M.P. Coogan, V. Fernandez-Moreira, Chem. Commun. 50 (2014) 384–399.[12] B.-Z. Zhu, X.-J. Chao, C.-H. Huang, Y. Li, Chem. Sci. 7 (2016) 4016–4023.[13] C.A. Bader, A. Sorvina, P.V. Simpson, P.J. Wright, S. Stagni, S.E. Plush, M. Massi,
D.A. Brooks, FEBS Lett. 590 (2016) 3051–3060.[14] M.V. Werrett, P.J. Wright, P.V. Simpson, P. Raiteri, B.W. Skelton, S. Stagni, A.G.
Buckley, P.J. Rigby, M. Massi, Dalton Trans. 44 (2015) 20636–20647.[15] X. Tian, Q. Zhang, M. Zhang, K. Uvdal, Q. Wang, J. Chen, W. Du, B. Huang, J. Wu,
Y. Tian, Chem. Sci. 8 (2017) 142–149.[16] M.-W. Louie, H.-W. Liu, M.H.-C. Lam, T.-C. Lau, K.K.-W. Lo, Organometallics 28
(2009) 4297–4307.[17] F.L. Thorp-Greenwood, R.G. Balasingham, M.P. Coogan, J. Organomet. Chem.
714 (2012) 12–21.[18] V. Fernandez-Moreira, F.L. Thorp-Greenwood, M.P. Coogan, Chem. Commun.
46 (2010) 186–202.[19] C.A. Bader, E.A. Carter, A. Safitri, P.V. Simpson, P. Wright, S. Stagni, M. Massi, P.
A. Lay, D.A. Brooks, S.E. Plush, Mol. BioSyst. 12 (2016) 2064–2068.[20] C.P. Montgomery, B.S. Murray, E.J. New, R. Pal, D. Parker, Acc. Chem. Res. 42
(2009) 925–937.[21] Z. Du, G.N. Borlace, R.D. Brooks, R.N. Butler, D.A. Brooks, S.E. Plush, Inorg.
Chim. Acta 410 (2014) 11–19.
[22] L. Xu, Y.-Y. Liu, L.-M. Chen, Y.-Y. Xie, J.-X. Liang, H. Chao, J. Inorg. Biochem. 159(2016) 82–88.
[23] J.A.L. Figueroa, C.A. Stiner, T.L. Radzyukevich, J.A. Heiny, Sci. Rep. 6 (2016)20551.
[24] D. Parker, Aust. J. Chem. 64 (2011) 239–243.[25] E. Baggaley, J.A. Weinstein, J.A.G. Williams, Coord. Chem. Rev. 256 (2012)
1762–1785.[26] I. Martinic, S.V. Eliseeva, S. Petoud, J. Lumin. 189 (2017) 19–43.[27] J. He, C.S. Bonnet, S.V. Eliseeva, S. Lacerda, T. Chauvin, P. Retailleau, F.
Szeremeta, B. Badet, S. Petoud, É. Tóth, P. Durand, J. Am. Chem. Soc. 138(2016) 2913–2916.
[28] J.-C.G. Bünzli, S.V. Eliseeva, Basics of lanthanide photophysics, in: P.Hänninen, H. Härmä (Eds.), Lanthanide Luminescence: Photophysical,Analytical and Biological Aspects, Springer, Berlin, Heidelberg, 2011, pp. 1–45.
[29] M.C. Heffern, L.M. Matosziuk, T.J. Meade, Chem. Rev. 114 (2014) 4496–4539.[30] E.L. Crossley, J.B. Aitken, S. Vogt, H.H. Harris, L.M. Rendina, Angew. Chem. Int.
Ed. 49 (2010) 1231–1233.[31] C.A. Puckett, R.J. Ernst, J.K. Barton, Dalton Trans. 39 (2010) 1159–1170.[32] C. Huang, C. Freter, Int. J. Mol. Sci. 16 (2015) 926–949.[33] A. Goel, A. Sharma, M. Kathuria, A. Bhattacharjee, A. Verma, P.R. Mishra, A.
Nazir, K. Mitra, Org. Lett. 16 (2014) 756–759.[34] X. Wu, G. Daniels, P. Lee, M.E. Monaco, Am. J. Clin. Exp. Urol. 2 (2014) 111–
120.[35] P.V. Simpson, I. Casari, S. Paternoster, B.W. Skelton, M. Falasca, M. Massi,
Chem. – A Eur. J. 23 (2017) 6518–6521.[36] H. Malhi, R.J. Kaufman, J. Hepatol. 54 (2011) 795–809.[37] J.R. Cubillos-Ruiz, S.E. Bettigole, L.H. Glimcher, Cell 168 (2017) 692–706.[38] P. Fagone, S. Jackowski, J. Lipid Res. 50 (2009) S311–S316.[39] R. Cao, J. Jia, X. Ma, M. Zhou, H. Fei, J. Med. Chem. 56 (2013) 3636–3644.[40] C. Caporale, C.A. Bader, A. Sorvina, K.D.M. MaGee, B.W. Skelton, T.A. Gillam, P.
J. Wright, P. Raiteri, S. Stagni, J.L. Morrison, S.E. Plush, D.A. Brooks, M. Massi,Chem. – A Eur. J. 23 (2017) 15666–15679.
[41] M.R. Duchen, Diabetes 53 (2004) S96.[42] S. Vyas, E. Zaganjor, M.C. Haigis, Cell 166 (2016) 555–566.[43] C. Shen, J.L. Kolanowski, C.M.N. Tran, A. Kaur, M.C. Akerfeldt, M.S. Rahme, T.
W. Hambley, E.J. New, Metallomics 8 (2016) 915–919.[44] X. Liu, Z. Tang, B. Song, H. Ma, J. Yuan, J. Mater. Chem. B 5 (2017) 2849–2855.[45] L.D. Zorova, V.A. Popkov, E.Y. Plotnikov, D.N. Silachev, I.B. Pevzner, S.S.
Jankauskas, V.A. Babenko, S.D. Zorov, A.V. Balakireva, M. Juhaszova, S.J.Sollott, D.B. Zorov, Anal. Biochem. (2017).
[46] S.E. Weinberg, N.S. Chandel, Nat. Chem. Biol. 11 (2015) 9–15.[47] J. Yang, J.-X. Zhao, Q. Cao, L. Hao, D. Zhou, Z. Gan, L.-N. Ji, Z.-W. Mao, ACS Appl.
Mater. Interfaces 9 (2017) 13900–13912.[48] Y. Li, C.-P. Tan, W. Zhang, L. He, L.-N. Ji, Z.-W. Mao, Biomaterials 39 (2015) 95–
104.[49] R.-R. Ye, C.-P. Tan, L.-N. Ji, Z.-W. Mao, Dalton Trans. 45 (2016) 13042–13051.[50] M.-H. Chen, F.-X. Wang, J.-J. Cao, C.-P. Tan, L.-N. Ji, Z.-W. Mao, ACS Appl.
Mater. Interfaces 9 (2017) 13304–13314.[51] M. Redmann, M. Dodson, M. Boyer-Guittaut, V. Darley-Usmar, J. Zhang, Int. J.
Biochem. Cell Biol. 53 (2014) 127–133.[52] K.K.-S. Tso, K.-K. Leung, H.-W. Liu, K.K.-W. Lo, Chem. Commun. 52 (2016)
4557–4560.[53] M. Boland, A. Chourasia, K. Macleod, Front. Oncol. 3 (2013).[54] H.-J. Yu, Z.-F. Hao, H.-L. Peng, R.-H. Rao, M. Sun, R. Alana, W.C. Ran, H. Chao, L.
Yu, Sens. Actuators, B 252 (2017) 313–321.[55] R.M. Perera, S. Stoykova, B.N. Nicolay, K.N. Ross, J. Fitamant, M. Boukhali, J.
Lengrand, V. Deshpande, M.K. Selig, C.R. Ferrone, J. Settleman, G.Stephanopoulos, N.J. Dyson, R. Zoncu, S. Ramaswamy, W. Haas, N. Bardeesy,Nature 524 (2015) 361–365.
[56] C. Fennelly, R.K. Amaravadi, Lysosomal biology in cancer, in: K. Öllinger, H.Appelqvist (Eds.), Lysosomes: Methods and Protocols, Springer, New York,New York, NY, 2017, pp. 293–308.
[57] S. Hämälistö, M. Jäättelä, Curr. Opin. Cell Biol. 39 (2016) 69–76.[58] H. Ma, B. Song, Y. Wang, C. Liu, X. Wang, J. Yuan, Dyes Pigm. 140 (2017) 407–
416.[59] F. Zhang, X. Liang, W. Zhang, Y.-L. Wang, H. Wang, Y.H. Mohammed, B. Song,
R. Zhang, J. Yuan, Biosens. Bioelectron. 87 (2017) 1005–1011.[60] A.E. Stacy, D. Palanimuthu, P.V. Bernhardt, D.S. Kalinowski, P.J. Jansson, D.R.
Richardson, J. Med. Chem. 59 (2016) 4965–4984.[61] M. Saito, H. Saitoh, Biosci. Biotechnol. Biochem. 76 (2012) 1777–1780.[62] S.S. Pasha, P. Das, N.P. Rath, D. Bandyopadhyay, N.R. Jana, I.R. Laskar, Inorg.
Chem. Commun. 67 (2016) 107–111.[63] Z.-P. Zeng, Q. Wu, F.-Y. Sun, K.-D. Zheng, W.-J. Mei, Inorg. Chem. 55 (2016)
5710–5718.[64] X. Li, K. Heimann, F. Li, J.M. Warner, F. Richard Keene, J. Grant Collins, Dalton
Trans. 45 (2016) 4017–4029.[65] B.J. Pages, D.L. Ang, E.P. Wright, J.R. Aldrich-Wright, Dalton Trans. 44 (2015)
3505–3526.[66] X. Li, K. Heimann, X.T. Dinh, F.R. Keene, J.G. Collins, Mol. BioSyst. 12 (2016)
3032–3045.[67] F. Li, Y. Mulyana, M. Feterl, J.M. Warner, J.G. Collins, F.R. Keene, Dalton Trans.
40 (2011) 5032–5038.[68] X. Li, A.K. Gorle, T.D. Ainsworth, K. Heimann, C.E. Woodward, J. Grant Collins,
F. Richard Keene, Dalton Trans. 44 (2015) 3594–3603.[69] F. Li, J.G. Collins, F.R. Keene, Chem. Soc. Rev. 44 (2015) 2529–2542.

T.A. Gillam et al. / Coordination Chemistry Reviews 375 (2018) 234–255 255
[70] K.K.-W. Lo, Acc. Chem. Res. 48 (2015) 2985–2995.[71] M. Soler, L. Feliu, M. Planas, X. Ribas, M. Costas, Dalton Trans. 45 (2016)
12970–12982.[72] U. Neugebauer, Y. Pellegrin, M. Devocelle, R.J. Forster, W. Signac, N. Moran, T.
E. Keyes, Chem. Commun. (2008) 5307–5309.[73] T.U. Connell, J.L. James, A.R. White, P.S. Donnelly, Chem. – A Eur. J 21 (2015)
14146–14155.[74] N. Sayyadi, I. Justiniano, R.E. Connally, R. Zhang, B. Shi, L. Kautto, A.V. Everest-
Dass, J. Yuan, B.J. Walsh, D. Jin, R.D. Willows, J.A. Piper, N.H. Packer, Anal.Chem. 88 (2016) 9564–9571.
[75] A. Leonidova, V. Pierroz, L.A. Adams, N. Barlow, S. Ferrari, B. Graham, G.Gasser, ACS Med. Chem. Lett. 5 (2014) 809–814.
[76] W.I. O’Malley, E.H. Abdelkader, M.L. Aulsebrook, R. Rubbiani, C.-T. Loh, M.R.Grace, L. Spiccia, G. Gasser, G. Otting, K.L. Tuck, B. Graham, Inorg. Chem. 55(2016) 1674–1682.
[77] A. Dantas de Araujo, C. Wu, K.-C. Wu, R.C. Reid, T. Durek, J. Lim, D.P. Fairlie,Bioconjug. Chem. 28 (2017) 1669–1676.
[78] A. Klos, A.J. Tenner, K.-O. Johswich, R.R. Ager, E.S. Reis, J. Köhl, Mol. Immunol.46 (2009) 2753–2766.
[79] M.D.A. Salazar, M. Ratnam, Cancer Metastasis Rev. 26 (2007) 141–152.[80] L.E. Gerweck, K. Seetharaman, Cancer Res. 56 (1996) 1194.[81] S. Quici, A. Casoni, F. Foschi, L. Armelao, G. Bottaro, R. Seraglia, C. Bolzati, N.
Salvarese, D. Carpanese, A. Rosato, J. Med. Chem. 58 (2015) 2003–2014.[82] M.-W. Louie, H.-W. Liu, M.H.-C. Lam, Y.-W. Lam, K.K.-W. Lo, Chem. – A Eur. J.
17 (2011) 8304–8308.[83] M. Gottschaldt, U.S. Schubert, Chem. – A Eur. J. 15 (2009) 1548–1557.[84] R. Brooks, Z. Du, G. Borlace, D. Brooks, S. Plush, J. Inorg. Chem. 2013 (2013) 8.[85] W.H.-T. Law, L.C.-C. Lee, M.-W. Louie, H.-W. Liu, T.W.-H. Ang, K.K.-W. Lo,
Inorg. Chem. 52 (2013) 13029–13041.[86] A. Palmioli, A. Aliprandi, D. Septiadi, M. Mauro, A. Bernardi, L. De Cola, M.
Panigati, Org. Biomol. Chem. 15 (2017) 1686–1699.[87] K. Yin, K. Zhang, Ka-Shun Tso, M.-W. Louie, H.-W. Liu, K. Kam-Wing Lo,
Organometallics 32 (2013) 5098–5102.[88] M. Pröhl, T. Bus, J.A. Czaplewska, A. Traeger, M. Deicke, H. Weiss, W.
Weigand, U.S. Schubert, M. Gottschaldt, Eur. J. Inorg. Chem. 2016 (2016)5197–5204.
[89] M. Pröhl, P.D. Moser, J.A. Czaplewska, P. Hoffmann, T. Bus, A. Traeger, H.Goerls, U.S. Schubert, M. Gottschaldt, Carbohydr. Res. 446–447 (2017) 19–27.
[90] M. Wenzel, A. de Almeida, E. Bigaeva, P. Kavanagh, M. Picquet, P. Le Gendre, E.Bodio, A. Casini, Inorg. Chem. 55 (2016) 2544–2557.
[91] N. Karaoun, A.K. Renfrew, Chem. Commun. 51 (2015) 14038–14041.[92] L. Zayat, C. Calero, P. Alborés, L. Baraldo, R. Etchenique, J. Am. Chem. Soc. 125
(2003) 882–883.[93] H. Chan, J.B. Ghrayche, J. Wei, A.K. Renfrew, Eur. J. Inorg. Chem. 2017 (2017)
1679–1686.[94] A.K. Renfrew, N.S. Bryce, T. Hambley, Chem. – A Eur. J. 21 (2015) 15224–
15234.[95] D.-L. Ma, V.P.-Y. Ma, D.S.-H. Chan, K.-H. Leung, H.-Z. He, C.-H. Leung, Coord.
Chem. Rev. 256 (2012) 3087–3113.
[96] J. Wu, Y. Yang, L. Zhang, H. Wang, M. Yang, J. Yuan, J. Mater. Chem. B 5 (2017)2322–2329.
[97] Y. Xiao, Z. Ye, G. Wang, J. Yuan, Inorg. Chem. 51 (2012) 2940–2946.[98] M.C. DeRosa, R.J. Crutchley, Coord. Chem. Rev. 233 (2002) 351–371.[99] P.R. Ogilby, Chem. Soc. Rev. 39 (2010) 3181–3209.[100] S.N.A. Jenie, S.M. Hickey, Z. Du, D. Sebben, D.A. Brooks, N.H. Voelcker, S.E.
Plush, Inorg. Chim. Acta 462 (2017) 236–240.[101] S.N.A. Jenie, S.E. Plush, N.H. Voelcker, Langmuir 33 (2017) 8606–8613.[102] L. Xiangli, G. Lianying, S. Bo, T. Zhixin, Y. Jingli, Method. Appl. Fluoresc. 5
(2017) 014009.[103] K. Chia, I.N. Fleming, P.J. Blower, Nucl. Med. Commun. 33 (2012) 217–222.[104] A.J. North, D.J. Hayne, C. Schieber, K. Price, A.R. White, P.J. Crouch, A.
Rigopoulos, G.J. O’Keefe, H. Tochon-Danguy, A.M. Scott, J.M. White, U.Ackermann, P.S. Donnelly, Inorg. Chem. 54 (2015) 9594–9610.
[105] D. Sneddon, R. Niemans, M. Bauwens, A. Yaromina, S.J.A. van Kuijk, N.G.Lieuwes, R. Biemans, I. Pooters, P.A. Pellegrini, N.A. Lengkeek, I. Greguric, K.F.Tonissen, C.T. Supuran, P. Lambin, L. Dubois, S.-A. Poulsen, J. Med. Chem. 59(2016) 6431–6443.
[106] S. Hussain, D.H. Palmer, R. Ganesan, L. Hiller, J. Gregory, P.G. Murray, J.Pastorek, L. Young, N. James, Oncol. Rep. 11 (2004) 1005–1010.
[107] D. Hedley, M. Pintilie, J. Woo, A. Morrison, D. Birle, A. Fyles, M. Milosevic, R.Hill, Clin. Cancer Res. 9 (2003) 5666.
[108] A.L. Harris, Nat. Rev. Cancer 2 (2002) 38–47.[109] N. Yamamoto, A.K. Renfrew, B.J. Kim, N.S. Bryce, T.W. Hambley, J. Med. Chem.
55 (1021) (2012) 11013–11020.[110] E.S. O’Neill, A. Kaur, D.P. Bishop, D. Shishmarev, P.W. Kuchel, S.M. Grieve, G.A.
Figtree, A.K. Renfrew, P.D. Bonnitcha, E.J. New, Inorg. Chem. 56 (2017) 9860–9868.
[111] A. Glenister, C.K.J. Chen, E.M. Tondl, D. Paterson, T.W. Hambley, A.K. Renfrew,Metallomics 9 (2017) 699–705.
[112] L. Zhang, A.F. Martins, Y. Mai, P. Zhao, A.M. Funk, M.V. Clavijo Jordan, S.Zhang, W. Chen, Y. Wu, A.D. Sherry, Chem. – A Eur. J. 23 (2017) 1752–1756.
[113] B. Song, Z. Ye, Y. Yang, H. Ma, X. Zheng, D. Jin, J. Yuan, Sci. Rep. 5 (2015)14194.
[114] W. Zhang, F. Zhang, Y.-L. Wang, B. Song, R. Zhang, J. Yuan, Inorg. Chem. 56(2017) 1309–1318.
[115] Y. Kondo, Y. Inai, Y. Sato, S. Handa, S. Kubo, K. Shimokado, S. Goto, M.Nishikimi, N. Maruyama, A. Ishigami, Proc. Natl. Acad. Sci. 103 (2006) 5723–5728.
[116] C. Maramag, M. Menon, K.C. Balaji, P.G. Reddy, S. Laxmanan, Prostate 32(1997) 188–195.
[117] M.E. Wright, G. Andreotti, J. Lissowska, M. Yeager, W. Zatonski, S.J. Chanock,W.-H. Chow, L. Hou, Eur. J. Cancer 45 (2009) 1824–1830.
[118] G.-H. Wu, Y.-F. Wu, X.-W. Liu, M.-C. Rong, X.-M. Chen, X. Chen, Anal. Chim.Acta 745 (2012) 33–37.
[119] H. Vicente Miranda, O.M.A. El-Agnaf, T.F. Outeiro, Mov. Disord. 31 (2016)782–790.
[120] L.M. Oliveira, A. Lages, R.A. Gomes, H. Neves, C. Família, A.V. Coelho, A.Quintas, BMC Biochem. 12 (2011) 41.